

Abstract
The non-classical role of vitamin D has been investigated in recent decades. One of which is related to its role in skeletal muscle. Satellite cells are skeletal muscle stem cells that play a pivotal role in skeletal muscle growth and regeneration. This systematic review aims to investigate the effect of vitamin D on satellite cells. A systematic search was performed in Scopus, MEDLINE, and Google Scholar. In vivo studies assessing the effect of vitamin D on satellite cells, published in English in the last ten years were included. Thirteen in vivo studies were analyzed in this review. Vitamin D increases the proliferation of satellite cells in the early life period. In acute muscle injury, vitamin D deficiency reduces satellite cells differentiation. However, administering high doses of vitamin D impairs skeletal muscle regeneration. Vitamin D may maintain satellite cell quiescence and prevent spontaneous differentiation in aging. Supplementation of vitamin D ameliorates decreased satellite cells’ function in chronic disease. Overall, evidence suggests that vitamin D affects satellite cells’ function in maintaining skeletal muscle homeostasis. Further research is needed to determine the most appropriate dose of vitamin D supplementation in a specific condition for the optimum satellite cells’ function.
Keywords: vitamin D, satellite cells, skeletal muscle, in vivo
1. Introduction
Vitamin D is a prohormone that has two main inactive isoforms, namely vitamin D2 (ergocalciferol) and vitamin D3 (cholecalciferol) [1,2]. Vitamin D2 is obtained from ultraviolet (UV) irradiation of ergosterol, a steroid that is found in some plants and fungi. Meanwhile, vitamin D3 is obtained mainly from UV irradiation of 7-dehydrocholesterol in the skin [3,4]. In addition, vitamin D3 is also obtained to a small extent from dietary intake such as oily fish, meat, or egg [2,3].
Vitamin D either obtained from UV exposure or food is then hydroxylated in the liver by vitamin D-25-hydroxylase to become 25-hydroxyvitamin D (25(OH)D) or also called calcidiol. 25(OH)D is a stable metabolite in the blood and best reflects exposure and absorption of Vitamin D. Therefore, 25(OH)D is used as an indicator of vitamin D status [1,2]. Furthermore, 25(OH)D needs to be converted by the enzyme 25(OH)D-1-OHase (CYP27B1) in the kidneys to become 1.25-dihydroxyvitamin D (1.25(OH)2D), the active form of vitamin D or also called calcitriol [5].
The classical function of vitamin D that has long been recognized is its role in the regulation of calcium and phosphate homeostasis and bone metabolism [6,7]. In the last few decades, some studies have indicated the non-classical function of vitamin D in various organs including skeletal muscle [8,9].
Satellite cells are skeletal muscle stem cells located between the basal lamina and sarcolemma which play a pivotal role in skeletal muscle growth and regeneration [10,11]. In homeostatic condition, satellite cells typically are in a quiescent state. The satellite cells will be activated when there are stimuli for skeletal muscle regeneration or hypertrophy, such as during an injury or exercise. Activated satellite cells will become myoblasts which proliferate and differentiate further to form new muscle fibers or fuse with preexisting muscle fibers [11,12]. This process is regulated by several myogenic regulatory factors (such as Myf5, MyoD, MRF4, and myogenin) [13,14]. In addition, some activated satellite cells can also undergo self-renewal and return to a quiescent state to replenish the satellite cell population (Figure 1) [15].
Figure 1.
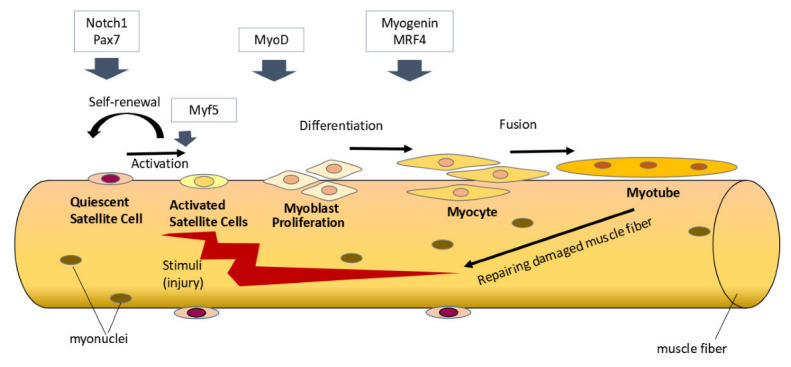
Satellite cells’ activity in skeletal muscle regeneration. Under homeostatic conditions, satellite cells are typically in a quiescent state and express Pax7. Notch signaling plays a role in maintaining the quiescent state of satellite cells. When there are stimuli for skeletal muscle regeneration, various myogenic regulatory factors (Myf5, MyoD, myogenin, and MRF4) regulate satellite cells’ activation, proliferation, and differentiation to form new muscle fibers. Some satellite cells will undergo self-renewal and return to a quiescent state to replenish the satellite cell population. Notch signaling through its regulation of Pax7 plays a role in promoting satellite cell self-renewal.
Several studies have shown that satellite cells express vitamin D receptor (VDR) [16,17,18]. Therefore, vitamin D may play a role in regulating satellite cells’ activity and function. A previous systematic review has discussed the effects of active vitamin D on myogenesis in vitro [19]. However, in vitro models cannot fully mimic the microenvironment of in vivo models [20,21]. Satellite cells’ fate is strongly influenced by their niche. Signals and properties of muscle fibers, basal lamina, as well as microvasculature, and surrounding interstitial cells influence the regulation of satellite cells’ function [22]. When satellite cells are removed from their in vivo niche and cultured in vitro, the satellite cells are activated and committed to proliferation and differentiation, thereby losing their stem cell properties [23]. This systematic review aims to collect the latest evidence from in vivo studies regarding the role of vitamin D on the satellite cells.
2. Methods
2.1. Search Strategy and Selection Process
This systematic review was constructed according to the Preferred Reporting Items for Systematic Reviews Meta-Analysis (PRISMA) guidelines. Relevant articles were identified through the online databases on Scopus, MEDLINE, and Google Scholar using the keywords (“vitamin D” OR “cholecalciferol” OR “calcitriol”) AND (“satellite cell*” OR “skeletal muscle stem cell*”) AND (“in vivo”). The search for articles was also performed manually to ensure getting as many relevant studies as possible. The article search was conducted in July 2022 and repeated in September 2022. The following inclusion and exclusion criteria were used in selecting articles:
Inclusion criteria
Articles are written in English
Published in the last ten years
Intervention with vitamin D supplementation or vitamin D deficiency
Assess the number or function of satellite cells
Exclusion criteria
Review articles
In vitro studies
Vitamin D supplementation in combination with other interventions (drugs/nutrients/exercise)
The study does not state the markers of the satellite cell’s function
The measured outcome of this review is the number of satellite cells characterized by Pax7+ cell quantification or the marker that regulates satellite cells’ activity and function.
2.2. Data Extraction
Data from the included articles were extracted using a standardized form that had been determined and approved by all authors. Any questions and discrepancies were resolved by discussion among the authors. Data extracted included: title, author, year of publication, animal’s species, animal’s age, number of animals per group, route of administration and dosing of vitamin D, and major findings related to satellite cells.
2.3. Risk of Bias Assessment
The risk of bias was evaluated according to the Systematic Review Centre for Laboratory Animal Experimentation (SYRCLE’s) risk of bias tool for animal studies [24,25].
3. Results
3.1. Study Selection
The initial search from Scopus, MEDLINE, and Google Scholar obtained 1421 records. A manual search of the bibliography of the relevant articles yielded four eligible articles. After reducing duplication and screening using titles and abstracts, 48 full-text articles were analyzed further. Thirty-five articles were excluded and a total of thirteen articles were used in this systematic review. The study selection flow chart is shown in Figure 2.
Figure 2.

Study selection flow adapted from Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guideline [26].
3.2. Study Characteristics
From thirteen in vivo studies included, six studies used rats, four studies used mice, two studies used pigs, and one study used chickens as experimental animals. The effect of vitamin D supplementation or restriction was assessed in growing animals, animals with muscle injuries, and aging or diseased animals (Table 1).
Table 1.
Characteristics of selected in vivo studies stratified based on the characteristics of experimental animal models.
| References | Species | Vitamin D form, Dose, Duration | Importance |
|---|---|---|---|
| Animals in the period of prenatal development and postnatal growth | |||
| Hutton et al., 2014 [27] | Male, Ross 708 broiler chicks (n total = 150) |
2240 IU vitamin D3 per kg diet + 2760 IU of 25(OH)D3 per kg diet for 49 days | ↑ mitotically active satellite cells (Pax7+; BrdU+), tended to ↑ density of Pax7+ satellite cells and Myf-5 + satellite cells in pectoralis mayor muscle |
| Zhou et al., 2016 [28] | White gilts (n = 10 per group) | 50 µg/kg vitamin D3 + 50 µg/kg 25(OH)D3 given from mating to weaning | ↑ MyoD1 and ↑ Myogenin mRNA expression in newborn and weaning piglets |
| Oku et al., 2016 [29] | Male Sprague Dawley rats (n = 6 per group) |
Diet with vitamin D restriction for 28 days | ↓ MyoD mRNA expression |
| Thayer et al., 2019 [30] | Sows and their progeny (n total = 69) | 1500 IU/kg vitamin D3; 500 IU/kg vitamin D3 + 25 μg/kg 25(OH)D3; or 1500 IU/kg vitamin D3 + 50 μg/kg 25(OH)D3, during gestation and lactation |
No difference in satellite cells number per fiber in pig’s muscle at birth |
| Reis et al., 2022 [31] | Male and female Wistar Hannover rats (n total = 20) | Maternal vitamin D deficiency | Following 180 days, only in Vitamin D deficiency male adult offspring, there are ↑ local calcitriol, ↑ CYP27B1, ↑ number of satellite cells, ↑ MyoD and ↑myogenin protein expression |
| Animals with muscle injury | |||
| Stratos et al., 2013 [32] | Male Wistar rats (n total = 56) | 332,000 IU/kg body weight vitamin D3 single dose subcutaneous injection after muscle injury | ↑ non myogenic cell proliferation, ↓ apoptosis; Not significant influence the number of satellite cells, ↑ extracellular matrix protein |
| Srikuea et al., 2016 [33] | Male C57BL/6 mice (n = 6 per group) |
1.25(OH)2D3 1 g/kg relative to tibialis anterior muscle wet weight (physiological dose) or 1.25(OH)2D3 1 g/kg relative to mouse body weight (supraphysiological dose), intramuscular injection at day 4–7 post injury | ↑ Vdr expresion in both doses; 1.25(OH)2D3 at a supraphysiological dose: ↓ satellite cell differentiation, delayed regenerative muscle fiber formation, and increased muscular fibrosis |
| Yu et al., 2021 [34] | Male Cyp27b1 knockout (KO) mice | Vitamin D deficiency in Cyp27b1 KO mice | ↓ MyoD, ↓Myf5, ↓MyHC in tibialis anterior muscle of Cyp27b1 KO after injected with BaCl2 |
| Aging or diseased animal | |||
| Faria et al., 2014 [35] | Male old Wistar rats (n = 10 per group) | AIN-93 M maintenance diet or a modified AIN-93 M diet with no vitamin D (to induce vitamin D deficiency) for 9 months | In vitamin D deficient group: ↓ Tibialis anterior weight, ↓ mRNA expression of marker of proliferation (Bmp4, Fgf-2, PCNA) ↓ Notch signaling activity |
| Kinoshita et al., 2015 [36] | Glucocorticoid-treated female Wistar rats (n = 5–9 per group) |
Eldecalcitol (activated vitamin D3 analogue) for 2 or 4 weeks | ↑ Pax7 and ↑ Myogenin mRNA expression during 2 weeks of treatment |
| Tamura et al., 2016 [37] | Diabetic C57BL/6 mice | Vitamin D-deficient diet for 6 weeks | ↓ Pax7 mRNA expression |
| Nakaoka et al., 2019 [38] | Ovariectomized female Sprague–Dawley rats (n = 6 per group) | Vitamin D restriction for 28 days | ↓ Myf-5 and ↓Myogenin mRNA expression |
| Cheung et al., 2020 [39] | C57BL/6 Ctns-/- mice (mouse model of infantile nephropathic cystinosis with Vitamin D insufficiency) | Supplementation with 25(OH)D + 1.25(OH)2D3 (75 μg/kg/ day + 60 ng/kg/day, respectively), for 6 weeks |
Ameliorate the decreased gene expression of Pax7, and MyoD |
VDR: Vitamin D Receptor; BrdU: Bromodeoxyuridine; Pax7: paired box 7; MyoD1: myogenic differentiation 1; Myf5: myogenic factor 5; MyHC: myosin heavy chain; PCNA: proliferating cell nuclear antigen; CYP27B1: Cytochrome P450 Family 27 Subfamily B Member 1; Bmp4: Bone morphogenetic protein 4; Fgf-1: Fibroblast growth factor-2; Igf-1: insulin-like growth factor-1; KO: knockout; mRNA: messenger RNA.
3.3. Risk of Bias Assessment
Table 2 shows the summary of the risk of bias assessment. Seven of thirteen studies described random treatment in the allocation of the subject group [27,28,30,31,33,35,36]. No study has declared a blinding procedure in the intervention and outcome assessment. In the included in vivo studies, several parameters of the risk of bias assessment were not reported. In line with these findings, a previous study reported that the prevalence of reporting the risk of bias in animal studies is still low [40].
Table 2.
Risk of bias summary of in vivo studies.
| References | Selection Bias | Performance Bias | Detection Bias | Attrition Bias | Reporting Bias | Other | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| Hutton et al. [27] |

|
|

|
|
|
|
|
|
|
|
| Zhou et al. [28] |
|
|
|
|
|
|
|
|
|
|
| Oku et al. [29] |
|
|
|
|
|
|
|
|
|
|
| Thayer et al. [30] |
|
|
|
|
|
|
|
|
|
|
| Reis et al. [31] |
|
|
|
|
|
|
|
|
|
|
| Stratos et al. [32] |
|
|
|
|
|
|
|
|
|
|
| Srikuea et al. [33] |
|
|
|
|
|
|
|
|
|
|
| Yu et al. [34] |
|
|
|
|
|
|
|
|
|
|
| Faria et al. [35] |
|
|
|
|
|
|
|
|
|
|
| Kinoshita et al. [36] |
|
|
|
|
|
|
|
|
|
|
| Tamura et al. [37] |
|
|
|
|
|
|
|
|
|
|
| Nakaoka et al. [38] |
|
|
|
|
|
|
|
|
|
|
| Cheung et al. [39] |
|
|
|
|
|
|
|
|
|
|
|
Low risk of bias |
No information/not applicable |
|||||||||
Sequence generation (1), baseline characteristics (2), allocation concealment (3), random housing (4), intervention blinding (5), random outcome assessment (6), outcome blinding (7), incomplete outcome (8), selective outcome reporting (9) and other (10).
3.4. Effect of Vitamin D on Satellite Cells during Prenatal Development and Postnatal Growth
Studies on pigs showed that maternal vitamin D supplementation during pregnancy and lactation improves vitamin D status in newborn and weaning offspring [28,30]. Improved vitamin D status in the pig’s offspring is associated with an increase in the number of primary muscle fibers at birth. However, the number of satellite cells in each muscle fiber was not different from the control [30]. Vitamin D may play a role in muscle growth and development in the early life period not by increasing the number of satellite cells but by increasing the functional capacity of satellite cells to form new muscle fibers. Zhou et al. reported that maternal vitamin D supplementation increased the cross-sectional area of muscle fibers in weaning piglets. In addition, there was also an increase in the MyoD and myogenin mRNA expression, markers of the satellite cell differentiation [28].
A recent study by Reis et al. showed that maternal vitamin D deficiency in rats may affect satellite cell count and skeletal muscle morphology in adult offspring. In that study, maternal vitamin D deficiency selectively increased the number of satellite cells and myogenic regeneration factor in type 2 muscle fibers in adult male offspring. This finding may be due to an increase in local vitamin D production in adult age to compensate for the limited intrauterine vitamin D [31].
Hutton et al. demonstrated that dietary vitamin D supplementation in newly hatched chicks increased mitotically active satellite cells (Pax7+, BrdU+). In addition, the density of Pax7 + satellite cells in fast twitch muscle also tended to increase, but it was not statistically significant (p = 0.07) [27].
In young adult rats, where muscle growth is still occurring but at a slower rate, vitamin D deficiency is associated with decreased MyoD mRNA expression [29].
3.5. Effect of Vitamin D on Satellite Cell during Muscle Injury
Yu et al. used cyp27b1 knockout mice to determine the effect of 1.25(OH)2D deficiency on skeletal muscle regeneration. In the injured muscle of cyp27b1 knockout mice, there was a significant decrease in the number of centrally nucleated fibers and the mRNA expression of Myf5, MyoD, and MyHC [34]. Newly regenerated myofibers are characterized by their centrally located nucleus [41,42]. In that study, decreased centrally nucleated myofiber in cyp27b1 knockout mice indicated that 1.25(OH)2D deficiency was associated with decreased skeletal muscle regeneration. The early stages of skeletal muscle regeneration involve the activation of satellite cells. MyoD and Myf5 are myogenic factors that play a role in the activation of satellite cells [43,44,45]. Thus, decreased skeletal muscle regeneration in 1.25(OH)2D-deficient mice may partly be due to impairment of satellite cell activation.
On the other hand, Srikuea et al. reported that intramuscular injection of 1.25(OH)2D at a supraphysiological dose (1 g/kg mouse body weight) in the injured muscle resulted in decreased satellite cell differentiation and inhibition of skeletal muscle regeneration [33]. Moreover, Stratos et al. showed that, in mice with injured muscle, subcutaneous administration of high doses of vitamin D3 (332,000 IU/kg body weight) had no effect on the number of satellite cells [32].
3.6. Effect of Vitamin D on Satellite Cells during Aging and Diseased States
Faria et al. assessed the effects of vitamin D deficiency in naturally aging rats (24 months old ......rats) [35]. In aging rats with vitamin D deficiency, there was a decrease in the protein expression of the Nocth1 transmembrane fragment and the mRNA expression of Hes1, the target gene of Notch signaling [35]. Notch signaling is one of the key regulators of stem cells in various tissues, including skeletal muscle. In intact skeletal muscle, Notch signaling plays a role in maintaining the quiescent state of skeletal muscle stem cells (satellite cells). Meanwhile, in regenerating muscle, Notch signaling plays a role in self-renewal and expansion of the satellite cell population [46,47]. Vitamin D deficiency in aging rats was also associated with decreased mRNA expression of bone morphogenetic protein 4 (Bmp4) and fibroblast growth factor-2 (Fgf-2), markers that play a role in satellite cell proliferation [35].
Tamura et al. showed that in diabetic mice, vitamin D deficiency augments decreased Pax7 mRNA expression [37]. Pax7 is a transcription factor that plays a critical role in satellite cells’ function. The absence of Pax7 results in the inability of satellite cells to undergo proliferation and self-renewal and lead to a decrease in the population of satellite cells due to precocious differentiation [48]. Pax7 also plays a role in the specification of satellite cells. Deletion of Pax7 also causes alternative stem cell development and leads to hematopoietic or brown adipogenic commitment [49,50].
Cheung et al. used a mouse model of infantile nephropathic cystinosis (Ctsn-/-) which had vitamin D deficiency. In Ctsn-/- mice, there was muscle atrophy and increased adipose tissue browning. Vitamin D repletion increased Pax7, and MyoD mRNA expression and reduced muscle atrophy and adipose tissue browning in these mice [39].
The use of glucocorticoids can induce skeletal muscle atrophy. Kinoshita et al. examined the effect of eldecalcitol (activated vitamin D analog) on rats receiving glucocorticoids. The study showed that eldecalcitol administration for two weeks increased pax7 and myogenin mRNA expression [36].
4. Discussion
In the neonatal period, skeletal muscle undergoes a high growth rate that involves high protein synthesis accompanied by a rapid increase in myonuclei. Satellite cells contribute to the addition of myonuclei to growing muscle fibers in the postnatal period [51]. This process of skeletal muscle growth depends on the proliferation of satellite cells. In 4-day-old rats with a chronically unloaded hindlimb, there was impaired growth of the soleus muscle associated with a decrease in mitotically active satellite cells [52]. The rapid growth of skeletal muscle during early life requires adequate nutrition. Nutrition has a critical influence in providing components for muscle mass synthesis and various signaling involved in the muscle fiber anabolism [51].
Vitamin D is a micronutrient that plays a role in maintaining various body functions throughout life [53]. Srikuea et al. demonstrated that satellite cells are the target cells of vitamin D action and the response of satellite cells to vitamin D varies depending on age. There is a decrease in satellite cell response to vitamin D in aged muscles compared to muscles in the developmental age [33]. These findings support the importance of vitamin D signaling in early life when satellite cell activity is high. Improved vitamin D status is associated with increased proliferation and myogenic capacity of satellite cells in the early weeks of life [27,28]. However, studies do not support the effect of vitamin D signaling on satellite cell number during muscle growth in the early life period [27,30]. A possible explanation is that the high rate of satellite cell proliferation in the early life period plays a role in providing new myonuclei to growing muscles and not increasing the number of satellite cell reserves.
Acute injury involves sudden changes in the form of damage to muscle fibers, infiltration of inflammatory cells, edema, and damage to surrounding tissues. All of these lead to a change in the niche and trigger the activation of satellite cells [54]. Vitamin D signaling appears to influence satellite cells’ function in skeletal muscle regeneration. In mice with vitamin D deficiency, there was a decrease in markers of activation and differentiation of satellite cells [34]. However, high-dose vitamin D supplementation without considering baseline vitamin D status leads to impaired satellite cell differentiation, delayed muscle fiber formation, and fibrosis formation in regenerating muscle [33]. Dosing appears to be a crucial issue when administering vitamin D during muscle regeneration. In vitro studies suggest that the administration of vitamin D supports muscle regeneration in a dose-dependent manner. However, at very high doses, it inhibits muscle formation [55,56,57].
In aging, there are some changes in the satellite cell niche, which causes the satellite cells to lose their quiescence and tend to differentiate prematurely [58]. Exposure of satellite cells in aging mice with serum from young mice can restore the regenerative function of satellite cells [59]. Aging is associated with decreased Notch signaling, a master regulator in maintaining the quiescent state of satellite cells [58]. Decreased Notch signaling in aged rats with vitamin D deficiency suggests that vitamin D may play a role in the regulation of Notch signaling in aging [35]. In in vitro study, Olsson et al. showed that the administration of vitamin D to human-derived myoblasts increased Hes1 mRNA expression, the gene target of Notch signaling [16]. One possible mechanism for vitamin D to regulate Notch signaling is its role in increasing Forkhead box O3 (FOXO3) expression [16]. A previous study demonstrated that FOXO3 is expressed in quiescent satellite cells. FOXO3 modulates Notch signaling by directly increasing Notch receptor expression. The FOXO3-Notch axis is required for satellite cell self-renewal by restoring satellite cell quiescence in regenerating muscle [60].
Vitamin D deficiency in aged rats was also associated with decreased Bmp4 and Fgf2 mRNA expression [35]. BMP signaling plays a role in increasing the satellite cell pool by promoting satellite cell proliferation and preventing precocious differentiation [61]. Stantzou et al. showed that inhibition of BMP signaling decreases the satellite cell pool [62]. Meanwhile, Fgf2 enhances satellite cell proliferation without suppressing differentiation [63]. Faria et al. assumed that the aged rats in their study experienced discrete regeneration episodes due to daily damage, so satellite cell proliferation was needed [35]. In another study, Fgf2 expression increased in aging muscle and triggered satellite cell proliferation and myogenic differentiation in homeostatic conditions. This causes satellite cell depletion and reduced muscle regeneration capacity [64]. Further studies are needed to confirm the role of vitamin D supplementation on Fgf2 in aged rats.
In chronic illness, there may be a chronic injury to the muscle depending on the severity and type of disease. Alterations in energy metabolism, inflammation, or restriction of movement can be factors that cause changes in satellite cells’ activity in diseased states [65,66]. Han et al. reported that an increase in extracellular adenosine (eADO) in diabetic mice decreased the regenerative function of satellite cells [67]. Satellite cells cultured on a high-glucose medium showed decreased proliferation and expression of Pax7, MyoD, and myogenin proteins [68]. In diseased experimental animal models, vitamin D deficiency aggravates the impaired function of satellite cells. Meanwhile, vitamin D supplementation ameliorates the impaired function of these satellite cells. Thus, it is important to pay attention to vitamin D status in various chronic diseases. Further research is needed to determine the dose of vitamin D administration to improve satellite cell function in the setting of chronic disease.
5. Conclusions
In vivo studies support a direct role of vitamin D on satellite cells’ function during muscle growth, injury, aging, or chronic disease. Vitamin D appears to increase satellite cell proliferation in the early life period during rapid muscle growth. Adequate vitamin D status is required to support the satellite cells’ function in skeletal muscle regeneration during acute injury. However, the administration of high doses of vitamin D decreases satellite cell differentiation and delays new muscle fiber formation. Vitamin D deficiency in aging was associated with the decrease in Notch signaling resulting in satellite cells losing their quiescent and differentiating prematurely. Vitamin D supplementation ameliorates the impairment of satellite cell function in chronic disease. Thus, to provide optimal effects on satellite cells’ function, it is necessary to administer vitamin D at a dose according to the physiological needs of each individual. Further research is needed to determine the most appropriate dose and duration of vitamin D supplementation in the various age groups and specific conditions such as in early life, injury, aging, or chronic disease.
Acknowledgments
We would like to thank Budi Setiabudiawan, the head of the Vitamin D Working Group Padjadjaran University, for his support for this work.
Author Contributions
Conceptualization, M.S.A. and N.S.; methodology, H.G.; formal analysis, R.L. and S.S.; investigation, M.S.A. and S.S.; writing—original draft preparation, M.S.A. and H.G.; writing—review and editing, N.S. and R.L.; visualization, N.S. and H.G.; supervision, V.M.T., R.L., N.S. and H.G. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
Grant funding for NS from Universitas Padjadjaran (Riset Percepatan Lektor Kepala), grant number: 1959/UN6.3.1/PT.00/2021.
Footnotes
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Latham C.M., Brightwell C.R., Keeble A.R., Munson B.D., Thomas N.T., Zagzoog A.M., Fry C.S., Fry J.L. Vitamin D Promotes Skeletal Muscle Regeneration and Mitochondrial Health. Front. Physiol. 2021;12:660498. doi: 10.3389/fphys.2021.660498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ran Z., Declan P.N. Vitamin D in health and disease: Current perspectives. Nutr. J. 2010;9:65. doi: 10.1186/1475-2891-9-65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bollen S.E., Atherton P.J. Myogenic, genomic and non-genomic influences of the vitamin D axis in skeletal muscle. Cell Biochem. Funct. 2021;39:48–59. doi: 10.1002/cbf.3595. [DOI] [PubMed] [Google Scholar]
- 4.Lappe J.M. The role of vitamin D in human health: A paradigm shift. Complement. Health Pract. Rev. 2011;16:58–72. doi: 10.1177/1533210110392952. [DOI] [Google Scholar]
- 5.Holick M. Vitamin D Deficiency. N. Engl. J. Med. 2007;357:266–281. doi: 10.1056/NEJMra070553. [DOI] [PubMed] [Google Scholar]
- 6.Dusso A.S., Brown A.J., Slatopolsky E. Vitamin D. Am. J. Physiol.-Ren. Physiol. 2005;289:8–28. doi: 10.1152/ajprenal.00336.2004. [DOI] [PubMed] [Google Scholar]
- 7.DeLuca H.F. The metabolism and functions of vitamin D. Adv Exp Med Biol. 1986;196:361–375. doi: 10.1007/978-1-4684-5101-6_24. [DOI] [PubMed] [Google Scholar]
- 8.Zittermann A., Gummert J.F. Nonclassical vitamin D Actions. Nutrients. 2010;2:408–425. doi: 10.3390/nu2040408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ceglia L. Vitamin D and its role in skeletal muscle. Curr. Opin. Clin. Nutr. Metab. Care. 2009;12:628–633. doi: 10.1097/MCO.0b013e328331c707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Wang Y.X., Rudnicki M.A. Satellite cells, the engines of muscle repair. Nat. Rev. Mol. Cell Biol. 2012;13:127–133. doi: 10.1038/nrm3265. [DOI] [PubMed] [Google Scholar]
- 11.Chen W., Datzkiw D., Rudnicki M.A. Satellite cells in ageing: Use it or lose it. Open Biol. 2020;10:200048. doi: 10.1098/rsob.200048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Motohashi N., Asakura A. Molecular Regulation of Muscle Satellite Cell Self-Renewal. J. Stem. Cell Res. Ther. 2012;S11:e002. doi: 10.4172/2157-7633.S11-e002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Dumont N.A., Wang Y.X., Rudnicki M.A. Intrinsic and extrinsic mechanisms regulating satellite cell function. Development. 2015;142:1572–1581. doi: 10.1242/dev.114223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Marg A., Escobar H., Gloy S., Kufeld M., Zacher J., Spuler A., Birchmeier C., Izsvák Z., Spuler S. Human satellite cells have regenerative capacity and are genetically manipulable. J. Clin. Investig. 2014;124:4257–4265. doi: 10.1172/JCI63992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Almeida C.F., Fernandes S.A., Ribeiro A.F., Jr., Okamoto O.K. Muscle Satellite Cells: Exploring the Basic Biology to Rule Them. Stem Cells Int. 2016;2016:1078686. doi: 10.1155/2016/1078686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Olsson K., Saini A., Strömberg A., Alam S., Lilja M., Rullman E., Gustafsson T. Evidence for Vitamin D receptor expression and direct effects of 1α,25(OH)2D3 in human skeletal muscle precursor cells. Endocrinology. 2016;157:98–111. doi: 10.1210/en.2015-1685. [DOI] [PubMed] [Google Scholar]
- 17.Braga M., Simmons Z., Norris K.C., Ferrini M.G., Artaza J.N. Vitamin D induces myogenic differentiation in skeletal muscle derived stem cells. Endocr. Connect. 2017;6:139–150. doi: 10.1530/EC-17-0008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Srikuea R., Hirunsai M., Charoenphandhu N. Regulation of vitamin D system in skeletal muscle and resident myogenic stem cell during development, maturation, and ageing. Sci. Rep. 2020;10:8239. doi: 10.1038/s41598-020-65067-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Alliband K.H., Kozhevnikova S.V., Parr T., Jethwa P.H., Brameld J.M. In Vitro Effects of Biologically Active Vitamin D on Myogenesis: A Systematic Review. Front. Physiol. 2021;12:736708. doi: 10.3389/fphys.2021.736708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cornelison D.D.W. Context matters: In vivo and in vitro influences on muscle satellite cell activity. J. Cell. Biochem. 2008;105:663–669. doi: 10.1002/jcb.21892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Yin H., Price F., Rudnicki M.A. Satellite cells and the muscle stem cell niche. Physiol. Rev. 2013;93:23–67. doi: 10.1152/physrev.00043.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kuang S., Gillespie M.A., Rudnicki M.A. Niche Regulation of Muscle Satellite Cell Self-Renewal and Differentiation. Cell Stem Cell. 2008;2:22–31. doi: 10.1016/j.stem.2007.12.012. [DOI] [PubMed] [Google Scholar]
- 23.Cosgrove B.D., Sacco A., Gilbert P.M., Blau H.M. A home away from home: Challenges and opportunities in engineering in vitro muscle satellite cell niches. Differentiation. 2009;78:185–194. doi: 10.1016/j.diff.2009.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Hooijmans C.R., Rovers M.M., De Vries R.B.M., Leenaars M., Ritskes-Hoitinga M., Langendam M.W. SYRCLE’s risk of bias tool for animal studies. BMC Med. Res. Methodol. 2014;14:43. doi: 10.1186/1471-2288-14-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Toro M.D., Nowomiejska K., Avitabile T., Rejdak R., Tripodi S., Porta A., Reibaldi M., Figus M., Posarelli C., Fiedorowicz M. Effect of resveratrol on in vitro and in vivo models of diabetic retinophathy: A systematic review. Int. J. Mol. Sci. 2019;20:3503. doi: 10.3390/ijms20143503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Liberati A., Altman D.G., Tetzlaff J., Mulrow C., Gøtzsche P.C., Ioannidis J.P.A., Clarke M., Devereaux P.J., Kleijnen J., Moher D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. PLoS Med. 2009;6:e1000100. doi: 10.1371/journal.pmed.1000100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Hutton K.C., Vaughn M.A., Litta G., Turner B.J., Starkey J.D. Effect of vitamin D status improvement with 25-hydroxycholecalciferol on skeletal muscle growth characteristics and satellite cell activity in broiler chickens. J. Anim. Sci. 2014;92:3291–3299. doi: 10.2527/jas.2013-7193. [DOI] [PubMed] [Google Scholar]
- 28.Zhou H., Chen Y., Lv G., Zhuo Y., Lin Y., Feng B., Fang Z., Che L., Li J., Xu S., et al. Improving maternal vitamin D status promotes prenatal and postnatal skeletal muscle development of pig offspring. [(accessed on 26 September 2022)];Nutrition. 2016 32:1144–1152. doi: 10.1016/j.nut.2016.03.004. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-84969508548&doi=10.1016%2Fj.nut.2016.03.004&partnerID=40&md5=89bfc0901e2b97b308637964014ff6e7. [DOI] [PubMed] [Google Scholar]
- 29.Oku Y., Tanabe R., Nakaoka K., Yamada A., Noda S., Hoshino A., Haraikawa M., Goseki-Sone M. Influences of dietary vitamin D restriction on bone strength, body composition and muscle in rats fed a high-fat diet: Involvement of mRNA expression of MyoD in skeletal muscle. [(accessed on 26 September 2022)];J. Nutr. Biochem. 2016 32:85–90. doi: 10.1016/j.jnutbio.2016.01.013. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-84962343810&doi=10.1016%2Fj.jnutbio.2016.01.013&partnerID=40&md5=1a5a871372bdedafd1caf0e05540d800. [DOI] [PubMed] [Google Scholar]
- 30.Thayer M.T., Nelssen J.L., Langemeier A.J., Morton J.M., Gonzalez J.M., Kruger S.R., Ou Z., Makowski A.J., Bergstrom J.R. The effects of maternal dietary supplementation of cholecalciferol (vitamin D3) and 25(OH)D3 on sow and progeny performance. Transl. Anim. Sci. 2019;3:693–708. doi: 10.1093/tas/txz029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Reis N.G., Assis A.P., Lautherbach N., Gonçalves D.A., Silveira W.A., Morgan H.J.N., Valentim R.R., Almeida L.F., Heck L.C., Zanon N.M., et al. Maternal vitamin D deficiency affects the morphology and function of glycolytic muscle in adult offspring rats. J. Cachexia. Sarcopenia Muscle. 2022;13:2175–2187. doi: 10.1002/jcsm.12986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Stratos I., Li Z., Herlyn P., Rotter R., Behrendt A.K., Mittlmeier T., Vollmar B. Vitamin D increases cellular turnover and functionally restores the skeletal muscle after crush injury in rats. Am. J. Pathol. 2013;182:895–904. doi: 10.1016/j.ajpath.2012.11.006. [DOI] [PubMed] [Google Scholar]
- 33.Srikuea R., Hirunsai M. Effects of intramuscular administration of 1α,25(OH)2D3 during skeletal muscle regeneration on regenerative capacity, muscular fibrosis, and angiogenesis. J. Appl. Physiol. 2016;120:1381–1393. doi: 10.1152/japplphysiol.01018.2015. [DOI] [PubMed] [Google Scholar]
- 34.Yu S., Ren B., Chen H., Goltzman D., Yan J., Miao D. 1,25-Dihydroxyvitamin D deficiency induces sarcopenia by inducing skeletal muscle cell senescence. [(accessed on 26 September 2022)];Am. J. Transl. Res. 2021 13:12638–12649. Available online: http://www.ncbi.nlm.nih.gov/pubmed/34956479%0Ahttp://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC8661220. [PMC free article] [PubMed] [Google Scholar]
- 35.Domingues-Faria C., Chanet A., Salles J., Berry A., Giraudet C., Patrac V., Denis P., Bouton K., Goncalves-Mendes N., Vasson M.-P., et al. Vitamin D deficiency down-regulates Notch pathway contributing to skeletal muscle atrophy in old wistar rats. [(accessed on 26 September 2022)];Nutr. Metab. 2014 11:47. doi: 10.1186/1743-7075-11-47. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-84908096067&doi=10.1186%2F1743-7075-11-47&partnerID=40&md5=1e7f23866adf31b8e2e035f7b8a2e8ee. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kinoshita H., Miyakoshi N., Kasukawa Y., Sakai S., Shiraishi A., Segawa T., Ohuchi K., Fujii M., Sato C., Shimada Y. Effects of eldecalcitol on bone and skeletal muscles in glucocorticoid-treated rats. J. Bone Miner. Metab. 2016;34:171–178. doi: 10.1007/s00774-015-0664-4. [DOI] [PubMed] [Google Scholar]
- 37.Tamura Y., Fujito H., Kawao N., Kaji H. Vitamin D deficiency aggravates diabetes-induced muscle wasting in female mice. [(accessed on 26 September 2022)];Diabetol. Int. 2017 8:52–58. doi: 10.1007/s13340-016-0278-7. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-85013162252&doi=10.1007%2Fs13340-016-0278-7&partnerID=40&md5=e79620fa5e240c9dbb768b3918e7f6dd. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nakaoka K., Yamada A., Noda S., Goseki-Sone M. Influence of dietary vitamin D deficiency on bone strength, body composition, and muscle in ovariectomized rats fed a high-fat diet. Nutrition. 2019;60:87–93. doi: 10.1016/j.nut.2018.09.001. [DOI] [PubMed] [Google Scholar]
- 39.Cheung W.W., Hao S., Wang Z., Ding W., Zheng R., Gonzalez A., Zhan J.Y., Zhou P., Li S., Esparza M.C., et al. Vitamin D repletion ameliorates adipose tissue browning and muscle wasting in infantile nephropathic cystinosis-associated cachexia. J. Cachexia. Sarcopenia Muscle. 2020;11:120–134. doi: 10.1002/jcsm.12497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Macleod M.R., Lawson McLean A., Kyriakopoulou A., Serghiou S., de Wilde A., Sherratt N., Hirst T., Hemblade R., Bahor Z., Nunes-Fonseca C., et al. Risk of Bias in Reports of In Vivo Research: A Focus for Improvement. PLoS biology. 2015;13:e1002273. doi: 10.1371/journal.pbio.1002273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Roman W., Gomes E.R. Nuclear positioning in skeletal muscle. Semin. Cell Dev. Biol. 2018;82:51–56. doi: 10.1016/j.semcdb.2017.11.005. [DOI] [PubMed] [Google Scholar]
- 42.Rodrigues M., Echigoya Y., Maruyama R., Lim K.R.Q., Fukada S.I., Yokota T. Impaired regenerative capacity and lower revertant fibre expansion in dystrophin-deficient mdx muscles on DBA/2 background. Sci. Rep. 2016;6:38371. doi: 10.1038/srep38371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cooper R.N., Tajbakhsh S., Mouly V., Cossu G., Buckingham M., Butler-Browne G.S. In vivo satellite cell activation via Myf5 and MyoD in regenerating mouse skeletal muscle. J. Cell Sci. 1999;112:2895–2901. doi: 10.1242/jcs.112.17.2895. [DOI] [PubMed] [Google Scholar]
- 44.Zammit P.S., Golding J.P., Nagata Y., Hudon V., Partridge T.A., Beauchamp J.R. Muscle satellite cells adopt divergent fates: A mechanism for self-renewal? J. Cell Biol. 2004;166:347–357. doi: 10.1083/jcb.200312007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Cornelison D.D.W., Wold B.J. Single-cell analysis of regulatory gene expression in quiescent and activated mouse skeletal muscle satellite cells. Dev. Biol. 1997;191:270–283. doi: 10.1006/dbio.1997.8721. [DOI] [PubMed] [Google Scholar]
- 46.Gioftsidi S., Relaix F., Mourikis P. The Notch signaling network in muscle stem cells during development, homeostasis, and disease. Skelet. Muscle. 2022;12:9. doi: 10.1186/s13395-022-00293-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Fujimaki S., Ono Y. Notch signaling in the regulation of skeletal muscle stem cells. J. Phys. Fit. Sport. Med. 2018;7:213–219. doi: 10.7600/jpfsm.7.213. [DOI] [Google Scholar]
- 48.Von Maltzahn J., Jones A.E., Parks R.J., Rudnicki M.A. Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc. Natl. Acad. Sci. USA. 2013;110:16474–16479. doi: 10.1073/pnas.1307680110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.An Y., Wang G., Diao Y., Long Y., Fu X., Weng M., Zhou L., Sun K., Cheung T.H., Ip N.Y., et al. A Molecular Switch Regulating Cell Fate Choice between Muscle Progenitor Cells and Brown Adipocytes. Dev. Cell. 2017;41:382–391.e5. doi: 10.1016/j.devcel.2017.04.012. [DOI] [PubMed] [Google Scholar]
- 50.Seale P., Sabourin L.A., Girgis-Gabardo A., Mansouri A., Gruss P., Rudnicki M.A. Pax7 is required for the specification of myogenic satellite cells. Cell. 2000;102:777–786. doi: 10.1016/S0092-8674(00)00066-0. [DOI] [PubMed] [Google Scholar]
- 51.Davis T.A., Fiorotto M.L. Regulation of muscle growth in neonates. Curr. Opin. Clin. Nutr. Metab. Care. 2009;12:78–85. doi: 10.1097/MCO.0b013e32831cef9f. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Kawano F., Takeno Y., Nakai N., Higo Y., Terada M., Ohira T., Nonaka I., Ohira Y. Essential role of satellite cells in the growth of rat soleus muscle fibers. Am. J. Physiol.-Cell Physiol. 2008;295:C458–C467. doi: 10.1152/ajpcell.00497.2007. [DOI] [PubMed] [Google Scholar]
- 53.Bendik I., Friedel A., Roos F.F., Weber P., Eggersdorfer M. Vitamin D: A critical and essential micronutrient for human health. Front. Physiol. 2014;5:248. doi: 10.3389/fphys.2014.00248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Warren G.L., Summan M., Gao X., Chapman R., Hulderman T., Simeonova P.P. Mechanisms of skeletal muscle injury and repair revealed by gene expression studies in mouse models. J. Physiol. 2007;582:825–841. doi: 10.1113/jphysiol.2007.132373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hosoyama T., Iida H., Kawai-Takaishi M., Watanabe K. Vitamin d inhibits myogenic cell fusion and expression of fusogenic genes. Nutrients. 2020;12:2192. doi: 10.3390/nu12082192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ryan K.J.P., Daniel Z.C.T.R., Craggs L.J.L., Parr T., Brameld J.M. Dose-dependent effects of vitamin D on transdifferentiation of skeletal muscle cells to adipose cells. [(accessed on 26 September 2022)];J. Endocrinol. 2013 217:45–58. doi: 10.1530/JOE-12-0234. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-84877107028&doi=10.1530%2FJOE-12-0234&partnerID=40&md5=78beefb1f8149ca108fa2407dcd433d8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Owens D.J., Sharples A.P., Polydorou I., Alwan N., Donovan T., Tang J., Fraser W.D., Cooper R.G., Morton J.P., Stewart C., et al. A systems-based investigation into vitamin D and skeletal muscle repair, regeneration, and hypertrophy. [(accessed on 26 September 2022)];Am. J. Physiol.-Endocrinol. Metab. 2015 309:E1019–E1031. doi: 10.1152/ajpendo.00375.2015. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-84951834947&doi=10.1152%2Fajpendo.00375.2015&partnerID=40&md5=04cafdcd8c67023c6dc1de3e3172bdaa. [DOI] [PubMed] [Google Scholar]
- 58.Parker M.H. The altered fate of aging satellite cells is determined by signaling and epigenetic changes. Front. Genet. 2015;5:59. doi: 10.3389/fgene.2015.00059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Conboy I.M., Conboy M.J., Wagers A.J., Girma E.R., Weissman I.L., Rando T.A. Rejuvenation of aged progenitor cells by exposure to a young systemic environment. [(accessed on 26 September 2022)];Nature. 2005 433:760–764. doi: 10.1038/nature03260. Available online: http://www.nature.com/nature/journal/v433/n7027/abs/nature03260.html. [DOI] [PubMed] [Google Scholar]
- 60.Gopinath S.D., Webb A.E., Brunet A., Rando T.A. FOXO3 promotes quiescence in adult muscle stem cells during the process of self-renewal. Stem Cell Reports. 2014;2:414–426. doi: 10.1016/j.stemcr.2014.02.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ono Y., Calhabeu F., Morgan J.E., Katagiri T., Amthor H., Zammit P.S. BMP signalling permits population expansion by preventing premature myogenic differentiation in muscle satellite cells. Cell Death Differ. 2011;18:222–234. doi: 10.1038/cdd.2010.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Stantzou A., Schirwis E., Swist S., Alonso-Martin S., Polydorou I., Zarrouki F., Mouisel E., Beley C., Julien A., Le Grand F., et al. BMP signaling regulates satellite cell-dependent postnatal muscle growth. Dev. 2017;144:2737–2747. doi: 10.1242/dev.144089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Yablonka-Reuveni Z., Seger R., Rivera A.J. Fibroblast growth factor promotes recruitment of skeletal muscle satellite cells in young and old rats. J. Histochem. Cytochem. 1999;47:23–42. doi: 10.1177/002215549904700104. [DOI] [PubMed] [Google Scholar]
- 64.Chakkalakal J.V., Jones K.M., Basson M.A., Brack A.S. The aged niche disrupts muscle stem cell quiescence. Nature. 2012;490:355–360. doi: 10.1038/nature11438. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Joseph J., Doles J.D. Disease-associated metabolic alterations that impact satellite cells and muscle regeneration: Perspectives and therapeutic outlook. [(accessed on 26 September 2022)];Nutr. Metab. 2021 18:33. doi: 10.1186/s12986-021-00565-0. Available online: https://www.scopus.com/inward/record.uri?eid=2-s2.0-85103197913&doi=10.1186%2Fs12986-021-00565-0&partnerID=40&md5=6ad7f3606e17cf4cc5e78ffcaedb95e1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.McKenna C.F., Fry C.S. Altered satellite cell dynamics accompany skeletal muscle atrophy during chronic illness, disuse, and aging. Curr. Opin. Clin. Nutr. Metab. Care. 2017;20:447–452. doi: 10.1097/MCO.0000000000000409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Han L., Wang G., Zhou S., Situ C., He Z., Li Y., Qiu Y., Huang Y., Xu A., Ong M.T.Y., et al. Muscle satellite cells are impaired in type 2 diabetic mice by elevated extracellular adenosine. Cell Rep. 2022;39:110884. doi: 10.1016/j.celrep.2022.110884. [DOI] [PubMed] [Google Scholar]
- 68.Furuichi Y., Kawabata Y., Aoki M., Mita Y., Fujii N.L., Manabe Y. Excess Glucose Impedes the Proliferation of Skeletal Muscle Satellite Cells Under Adherent Culture Conditions. Front. Cell Dev. Biol. 2021;9:640399. doi: 10.3389/fcell.2021.640399. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.