

Abstract
Central carbon metabolism (CCM), including glycolysis, tricarboxylic acid cycle and the pentose phosphate pathway, is the most fundamental metabolic process in the activities of living organisms that maintains normal cellular growth. CCM has been widely used in microbial metabolic engineering in recent years due to its unique regulatory role in cellular metabolism. Using yeast and Escherichia coli as the representative organisms, we summarized the metabolic engineering strategies on the optimization of CCM in eukaryotic and prokaryotic microbial chassis, such as the introduction of heterologous CCM metabolic pathways and the optimization of key enzymes or regulatory factors, to lay the groundwork for the future use of CCM optimization in metabolic engineering. Furthermore, the bottlenecks in the application of CCM optimization in metabolic engineering and future application prospects are summarized.
Keywords: Central carbon metabolism, Metabolic engineering, Microbial chassis, Metabolic pathways, Yeast, Escherichia coli
Background
Since its inception, metabolic engineering has rapidly promoted the biosynthesis of natural products [1, 2]. In the pharmaceutical industry, various high-value pharmaceutical ingredients, including artemisinin, ginsenosides, opioids and vinblastine, have been synthesized using microorganisms [3–6]. The growing environmental awareness of the general public and the scarcity of fossil fuels have gradually revealed the advantages of green biomanufacturing of bulk chemicals [7, 8]. The rise in global oil prices has also heightened interest in biofuels [9, 10].
Metabolic engineering employs genetic engineering techniques to increase the yield of target products by modifying metabolic pathways within cells. The most common methods in metabolic engineering include manipulation of promoter and copy number of target enzymes [11, 12], transcription factor regulation [13], fusion protein construction [14], protein scaffold assembly [15], organelle compartmentalization [16], and dynamic regulatory engineering [17]. The integration of these approaches enables complex and sophisticated metabolic pathway optimization of chassis cells to develop desired optimization strategies such as increasing the metabolic flux of target product-related pathways [18], blocking or attenuating other target product-consuming pathways [19], increasing the catalytic rate of rate-limiting steps [20], and introducing heterologous metabolic pathways [21]. However, a wide range of engineering modifications can have varying degrees of negative impact on the overall metabolism of the chassis strain, causing an imbalance in the metabolic flux of the chassis strain, inhibiting its physiological activity, and ultimately affecting production performance [22]. Because the optimization of the metabolic pathways where the target products are located or adjacent has been relatively well established, further optimization of these pathways has a limited effect on yield improvement. Therefore, recent studies have focused on the global regulation of metabolic flux, looking for breakthroughs in the most fundamental metabolic pathway, central carbon metabolism (CCM), which includes glycolysis, the tricarboxylic acid cycle (TCA cycle), and the pentose phosphate pathway (PPP).
CCM is a major source of energy for cell growth and development and provides precursors for other metabolic activities. Modification of the CCM, which is upstream of biological metabolic activities, often results in the rearrangement of the global metabolic flux of the cell and has a high potential for metabolic engineering applications. On the one hand, optimization of CCM can increase the precursors supply for the targeted compounds. For example, the introduction of the heterologous phosphoketolase phosphotransacetylase (PHK) metabolic pathway can increase the rate of acetyl coenzyme A (acetyl-CoA) synthesis and trigger CCM (glycolysis and the PPP) rearrangement in Saccharomyces cerevisiae, thereby increasing the production of the target product farnesene by 25% [23]. On the other, the manipulation of CCM often causes the rebalance of the availability of energy and the redox cofactors, such as ATP (adenosine triphosphate), NADPH (nicotinamide adenine dinucleotide phosphate) and NADH (nicotinamide adenine dinucleotide), to promote the output of final products by improving the corresponding catalytic steps involved in the biosynthesis pathways. The introduction of the Deinococcus radiodurans response regulator DR1558 into E. coli improves the expression efficiency of the genes related to CCM, and induces the excess generation of NADPH from PPP and supplies the cofactor requirements during PHB biosynthesis [24].
In recent years, metabolic engineering strategies on the optimization of CCM has produced remarkable results in the biosynthesis of many natural products. However, the current status of the application of this optimization strategy has not yet been systematically discussed. In this work, representative chassis strains of yeast and E. coli were selected to summarize the application and potential of CCM in metabolic engineering.
Application of CCM optimization in yeast
Introduction of heterologous metabolic pathways
The introduction of a heterologous CCM metabolic pathway has been shown to be an effective method for regulating CCM in host cells. The introduction of a heterologous CCM that is not found in Saccharomyces species could improve the carbon flux between different pathways of CCM, and promote the biosynthesis of target compounds (Fig. 1).
Fig. 1.
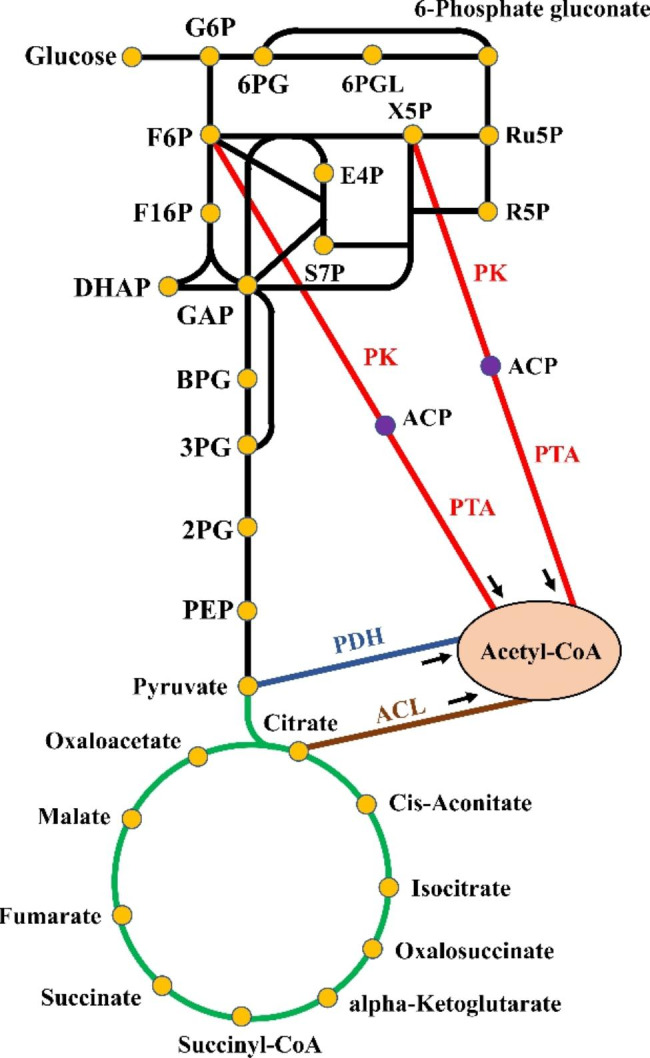
The optimization of CCM by introduction of heterologous pathways to improve the carbon flux in S. cerevisiae. The black line represented glycolysis and pentose phosphate pathway, and the green line represented the tricarboxylic acid cycle. The red, blue and brown lines represented the implemented PHK, PDH and ACL pathways. The orange dot represented the products in CCM, while the purple dot represented ACP. G6P, Glucose 6-phosphate; F6P, Fructose-6-phosphate; F1,6P, Fructose-1,6-bisphosphate; GAP, Glyceraldehyde 3-phosphate; DHAP, Dihydroxyacetone phosphate; BPG, 1,3-Bisphosphoglycerate; 3PG, 3-Phosphoglycerate; 2PG, 2-Phosphoglycerate; PEP, Phosphoenolpyruvate; 6PG, 6-Phosphogluconate; 6PGL, 6-Phosphate glucono-1,5-lactone; X5P, Xylulose 5-phophate; Ru5P, Ribulose 5-phophate; R5P, Ribose 5-phosphate; E4P, Erythrose 4-phosphate; S7P, Sedoheptulose 7-phophate; ACP, Acetyl-phosphate; Acetyl-CoA, Acetyl coenzyme A; PK, Phosphoketolase; PTA, Phosphotransacetylase; PDH, Pyruvate dehydrogenase; ACL, ATP: citrate lyase
PHK pathway
In the CCM of yeast, glucose-6-phosphate (G6P) can generate either fructose-6-phosphate (F6P) in glycolysis or ribulose-5-phosphate (Ru5P) and xylulose 5-phophate (X5P) into the PPP pathway. The introduction of PHK in S. cerevisiae catalyzes the direct production of acetyl-phosphate (ACP) from F6P and X5P to acetyl-CoA via a transacetylation reaction [25]. The only enzymes in the PHK pathway are phosphoketolase (PK) and phosphotransacetylase (PTA). Due to its simple constitution, it is widely used in metabolic engineering.
The PHK pathway facilitates the direct synthesis of acetyl-CoA and the biosynthesis of lipid compounds using acetyl-CoA as a precursor. The knockout of phosphofructokinase (PFK) in Yarrowia lipolytica blocked the metabolic flux of G6P in glycolysis and caused the redox imbalance with excess NADPH production. Then the introduction of the PHK pathway resulted in the accumulation of acetyl-CoA and correction of the redox imbalance by providing a route towards the NADPH-oxidizing lipid synthesis pathway, and a 19% increase in total lipid production was derived in the host strain [26]. In the TCA cycle, citrate can be catalyzed by ATP:citrate lyase (ACL) to produce acetyl-CoA [27]. The mouse-derived ACL and PHK pathway were used to optimize the CCM in fatty acid biosynthesis in Pichia pastoris. Coupled with the subsequent overexpression of NADPH-generating enzymes in TCA cycle and PPP, the engineered strain produced 23.4 g/L of free fatty acids and 2.0 g/L of fatty alcohols [28]. Overexpression of alcohol dehydrogenase 2 (Adh2), acetaldehyde dehydrogenase 6 (Ald6), and exogenous acetyl-CoA synthetase (ACS) variant acsSEL641P following the introduction of the PHK pathway, which provides acetyl-CoA and NADPH, in S. cerevisiae inhibited the synthesis of ethanol and yielded 5100 ± 509 g/CDW (cell dry weight) of fatty acid ethyl esters [29].
The introduction of the PHK pathway also addresses the issue of insufficient erythrose-4-phosphate (E4P) synthesis in S. cerevisiae. The PHK pathway catalyzes the conversion of F6P to acetyl-CoA, which decreases the consumption of metabolic flux in glycolysis while indirectly increasing the metabolic flux in the PPP and promoting E4P accumulation, which provides a large number of precursors for the synthesis of aromatic compounds. The introduction of the PHK pathway in S. cerevisiae shifts the glycolytic flux to E4P synthesis, avoiding the loss of metabolic flux at multiple steps upstream of the glycolysis and PPP. The subsequent promoter optimization and dynamic regulation resulted in a yield of 12.5 g/L for p-hydroxycinnamic acid and a maximum yield on glucose of 154.9 mg/g [30]. The heterologous PHK pathway can increase tyrosol production in the host strain by 135-fold by rearranging the glycolysis and the PPP. Fed batch fermentation using glucose as a carbon source resulted in a total tyrosol and salidroside production of over 10 g/L [31]. Therefore, exogenous introduction of the PHK pathway is an effective strategy for the synthesis of aromatic compounds and derivatives downstream of the PPP pathway, especially in the absence of the available precursor E4P. However, the introduction of the PHK pathway had no significant effect on the biosynthesis of other aromatic compounds. The expression of the PHK pathway did not significantly increase the yield of 2-phenylethanol (2-PE), which may be attributed to the excess carbon flux from pyruvate synthesis that the PHK pathway competes for [32]. Hence, when multiple CCM pathways are regulated in parallel, the introduction of the PHK pathway may be subjected to unknown interference.
The regulatory strategy of the PHK pathway in chassis strain has also been applied to the biosynthesis of other classes of compounds. Protopanaxadiol (PPD), an active triterpene compound, serves as a precursor of high-value ginsenosides. The introduction of the PHK pathway and multicopy integration of endogenous transaldolase 1 (Tal1) and transketolase 1 (Tkl1) in S. cerevisiae increased the PPD yield to 152.37 mg/L [33]. In addition, the introduction of the PHK pathway in S. cerevisiae increased 3-hydroxypropionic acid (3-HP) production by 41.9% and decreased glycerol production by 48.1%. In addition, reducing the expression of phosphoglucose isomerase and overexpressing acetyl-CoA carboxylase (Acc1) and malonyl-CoA reductase (MCR) promoted the metabolic flux to the PPP, yielding 864.5 mg/L of 3-HP (24 times the yield of the initial strain) [34]. Furthermore, after being introduced into S. cerevisiae strain containing the ethanol degradation pathway, the Aspergillus nidulans-derived PHK pathway could be used for the synthesis of polyhydroxybutyrate (PHB) with a yield of 56.4 mg/g in ethanol medium [25].
Other heterologous pathways
Other pathways, in addition to the PHK pathway, can modulate the CCM of the yeast chassis. ACL from A. nidulans increased the mevalonate yield to 2-fold by directly converting citric acid to acetyl-CoA in the TCA cycle of S. cerevisiae [35]. But this strategy is unsuitable for large-scale industrial production due to the limitation of the citric acid flux of the ACL substrate. Besides, the pyruvate dehydrogenase (PDH) pathway of E. coli can directly convert pyruvate from the glycolytic pathway to acetyl-CoA. The absence of ATP consumption in this process conserves more energy for other CCM reactions. After NADP+-dependent modifications in S. cerevisiae, the introduction of PDH pathway resulted in a 2-fold increase in acetyl-CoA [36]. Despite the inferiority of the regulatory effect of the above pathways on CCM compared to the PHK pathway, there may be synergistic effects between these pathways and the PHK pathway. A-ALD in E. coli can not only catalyze the conversion of acetaldehyde to acetyl-CoA, but also promotes the accumulation of large amounts of redox cofactor NADH in the cytoplasm, which would balance the relationship between NADPH consumption and NADH generation in yeast cells [37, 38]. Then the combined use of the A-ALD and PHK pathways in S. cerevisiae resulted in the accumulation of large amounts of acetyl-CoA. Optimization on this basis yielded 279.0 ± 13.0 mg/L of β-amyrin [38].
Optimization of CCM by key enzymes or regulatory factors in yeast
Optimization of key enzymes
The modification of key enzymes in each CCM pathway can rearrange the metabolic flux and facilitate the synthesis of target products. Pyruvate decarboxylase (PDC) initiates the catalysis of pyruvate (a product of glycolysis) to ethanol in S. cerevisiae. The knockdown of the PDC gene effectively shifts the metabolic flux from the ethanol synthesis pathway to the pyruvate-related pathway in the CCM, reducing consumption of the CCM flux by the ethanol synthesis pathway. Pyruvate accumulation was also confirmed by deletion mutants of pdc1 and pdc5 in S. cerevisiae [39]. A yield of 81.0 g/L of 2,3-butanediol was attained by reconstituting the 2,3-butanediol biosynthetic pathway in PDC knockout strains and performing batch fermentation with high concentrations of glucose as a carbon source [40]. In S. cerevisiae, knocking out hexokinase 2 (HXK2) and glucokinase 1 (GLK1) in glycolysis and using tetracycline transactivator protein (tTA) to control hexokinase 1 (HXK1) transcription can shift the metabolic flux from glycolysis to the gluconate synthesis pathway, promoting efficient gluconate biosynthesis, with the final strain showing a 50-fold increase in gluconate production compared to the control strain [41]. Overexpression of glucose-6-phosphate dehydrogenase (Zwf), glucose-6-phosphate isomerase (Pgi), and Pfk1 in P. pastoris, which effectively inhibits the carbon flux of glycolysis, can promote inositol biosynthesis, with inositol production reaching 30.71 g/L [42].
The above reports were optimized for only a few (1–3) key enzymes in CCM. In the metabolic engineering of some target compounds, large-scale optimization of enzymes in CCM has also been performed. The knockdown of 15 relevant CCM enzyme genes in S. cerevisiae and the introduction of 2-pyrone synthase (2-PS) in Gerbera hybrida promote the decarboxylation/condensation reaction of acetyl-CoA and malonyl coenzyme A (malonyl-CoA) to produce triacetic acid lactone (TAL). This process ultimately leads to a 37-fold increase in TAL yield to 2.2 g/L and a 50-fold increase on glucose yield to 0.13 g/g [43].
Optimization of key regulatory factors
In yeast, acetyl-CoA is primarily derived from CCM. The partitioned distribution and insufficient amount of acetyl-CoA limit the ability to synthesize the target product. In Crabtree-negative strain Komagataella phaffii, an ethanol-inducible and constitutive transcriptional regulatory signaling amplifier designed with the transcription activation region of the transcription factor MIT1 can increase ethanol-inducible expression capacity by nearly 20-fold. In addition, this process enables host cells to produce acetyl-CoA independently of CCM using ethanol as a fermentation substrate as well as an acetyl-CoA precursor and inducer. Direct production of acetyl-CoA from ethanol in the cytoplasm via a three-step catalytic process and construction of a biosynthetic pathway increased the yield of the cholesterol-lowering drug simvastatin intermediate, monacolin J, to 3.2 g/L [44].
Ric1 is a transcriptional repressor of multiple genes in the aromatic amino acid biosynthetic pathway in S. cerevisiae [45]. Decreasing Ric1 expression in S. cerevisiae and overexpressing ribose-5-phosphate ketol-isomerase (Rki1) in the PPP resulted in a 7-fold increase in the yield of shikimic acid (SA) to 2.5 g/L. The total yield of muconic acid and the intermediate product protocatechuic acid in the strain developed on this basis was 2.7 g/L [46]. A summary of the applications of CCM optimization in yeast were provided in Table 1.
Table 1.
Optimization of CCM in eukaryotic chassis
| Host | Manipulation | CCM involved | Products | References |
|---|---|---|---|---|
| S. cerevisiae | Introduction of PHK pathway | Glycolysis and PPP | Farnesene | [23] |
| S. cerevisiae | Introduction of PHK pathway | Glycolysis and PPP | Polyhydroxybutyrate | [25] |
| S. cerevisiae | Introduction of PHK pathway | Glycolysis and PPP | p-Hydroxycinnamic acid | [30] |
| S. cerevisiae | Introduction of PHK pathway and A-ALD | Glycolysis and PPP | β-Amyrin | [38] |
| S. cerevisiae |
Introduction of PHK pathway, overexpression of ADH2, ALD6 and acsSEL641P |
Glycolysis and PPP | Fatty acid ethyl esters | [29] |
| S. cerevisiae | Introduction of PHK pathway, multi-copy integration of Tal1 and Tkl1 | Glycolysis and PPP | Protopanaxadiol | [33] |
| S. cerevisiae | Introduction of PK pathway | Glycolysis and PPP | Tyrosol and salidroside | [31] |
| S. cerevisiae | Overexpression of a modified PDH pathway | Glycolysis | Acetyl-CoA | [36] |
| S. cerevisiae | PDC1 deletion | Glycolysis | Pyruvate and lactate | [39] |
| S. cerevisiae | PDC1 deletion | Glycolysis | 2,3-Butanediol | [40] |
| S. cerevisiae | Deletion of HXK2 and GLK1, tTA-controlled expression of HXK1 | Glycolysis | Gluconate | [41] |
| S. cerevisiae | Introduction of PHK pathway, downregulation of phosphoglucose isomerase, and overexpression Acc1 and Mcr | PPP | 3-Hydroxypropionic acid | [34] |
| S. cerevisiae | Repression of Ric1 and overexpression of Rki1 | PPP | Shikimic acid, muconic acid and protocatechuic acid | [46] |
| S. cerevisiae | Introduction of ACL from A. nidulans | TCA cycle | Mevalonate | [35] |
| S. cerevisiae | Knockout of 15 genes of CCM | Glycolysis, PPP and TCA cycle | Triacetic acid lactone | [43] |
| P. pastoris | Overexpression of Zwf, Pgi and Pfk1 | Glycolysis | Inositol | [42] |
| P. pastoris | Control CCM by a synthetic constitutive transcriptional signal amplification device | Glycolysis | Monacolin J | [44] |
| P. pastoris | Introduction of PHK pathway and the ACL from M. musculus | Glycolysis and TCA cycle | Free fatty acids and fatty alcohol | [28] |
| Y. lipolytica | Introduction of PHK pathway and PFK deletion | Glycolysis | Lipid | [26] |
| C. synechocystis | Expression of extra copies of PEPC | TCA cycle | Ethylene | [47] |
| C. synechocystis | Expression of extra copies of PEPC and elevated culture temperature | Glycolysis and TCA cycle | Succinate | [48] |
Optimization of CCM in other eukaryotic chassis
In cyanobacterium Synechocystis sp., the lower carbon flux of the TCA cycle limits the biosynthesis of target products. Increased copy number of the phosphoenolpyruvate carboxylase (PEPC) gene and increased carbon flux into the TCA cycle significantly increased the production of ethylene to 10.5 µg/mL/OD/day), a 1.64-fold increase compared to the original strain [47]. Synechocystis sp. is also an excellent chassis strain for succinate production. Overexpression of PEPC increased succinate yields to 162.3 mg/L after increasing temperature to decrease glycolytic carbon flux and increase TCA cycle carbon flux [48].
Application of CCM optimization in E. coli
Rearrangement of CCM metabolic flux through the PTS system
When E. coli uses glucose as a carbon source, the sugar phosphotransferase system (PTS) transfers the phosphate group from phosphoenolpyruvate (PEP) to glucose to produce G6P and pyruvate. PTS is the primary PEP consumption pathway for E. coli growth [49]. PEP is a key intermediate that links the three major CCM pathways and regulates the expression of some CCM genes [50]. Therefore, regulation of CCM by the PTS system (primarily by deletion of the PTS system) is a widely used CCM regulation strategy in E. coli metabolic engineering.
The deletion of the PTS system in E. coli decreased its carbon consumption, and inhibiting alanine: H+ symporter activity attenuated cell growth inhibition. These optimizations increased β-alanine production to 4.36 g/L [51]. The deletion of the PTS system and knockdown of the transcription factor TyrR, which has a repressive effect on the aromatic amino acid synthesis pathway, significantly increased L-tyrosine production [52, 53]. Further metabolic engineering optimization resulted in melanin and L-dopa yields of 3.22 g/L and 25.53 g/L, respectively [53, 54]. The knockdown of pyruvate kinase (PYK), PEPC, and malic enzymes along with deletion of the PTS system can increase the metabolic flux of the TCA cycle, resulting in a final yield of 5.89 mmol g/DCW (dry cell weight)/h of succinate in the optimized strain [55]. The replacement of the PTS system in E. coli with the galactose transport system resulted in a substantial accumulation of PEP. Further use of PEPC to reroute the metabolic flux of glycolysis to the TCA cycle resulted in a fumaric acid yield of 1.53 g/g dry cell weight [56]. Displacement of the PTS system of E. coli and overexpression of PK increased the precursors of aromatic amino acid synthesis pathway, PEP and E4P. The resulting engineered strain produced 41.7 g/L of tryptophan after fermentation in a 5 L bioreactor [57].
Optimization of CCM by key enzymes or regulatory factors in E. coli
Optimization of key enzymes
In E. coli, PGI and ZWF regulate the metabolic flux into the glycolytic pathway and PPP, respectively. Therefore, CCM optimization by regulation of PGI and ZWF is common in E. coli metabolic engineering. Simultaneous knockdown of PGI and ZWF can increase metabolic flux to the methylerythritol 4-phosphate (MEP) pathway, resulting in the accumulation of isoprenoids and their derivatives and facilitating terpenoid biosynthesis. In high-lycopene production strains constructed using this strategy, lycopene yields of 6.85–7.55 mg/g DCW were achieved [58, 59]. PGI knockdown can increase the metabolic flux to the PPP. Furthermore, ACS overexpression and other optimization methods increased the production of riboflavin to 585.2 mg/L [60]. ZWF knockdown increased the metabolic flux to glycolysis, and further optimization resulted in β-carotene production of 266.4 mg/L [61]. Triosephosphate isomerase (TPIA) is an enzyme that converts dihydroxyacetone phosphate (DHAP) to glyceraldehyde-3-phosphate (GAP). TPIA and ZWF knockdown shift metabolic flux toward pyruvate synthesis, and further optimization can increase 3-HP production by 4.4-fold [62].
CRISPR/Cas9 technology for large-scale gene editing of CCM pathways was used for the optimization of CCM. The use of CRISPR/Cas9 technology to silence a dozen enzymes in the glycolytic and TCA cycle pathways could redirect metabolic flux to the malonyl-CoA synthesis pathway that subsequently increases (2 S)-naringenin production to 421.6 mg/L, a 7.4-fold increase compared to the control strain [63]. Based on CRISPR silencing technology, high-throughput screening of enzymes in the CCM and related pathways that affect the yield of the target product and editing of all genes encoding these enzymes can significantly increase the yield of the target product. This CCM optimiz is currently used in the biosynthesis of (2 S)-pinocembrin and medium-chain fatty acids [64, 65].
CCM can also be optimized by regulating the synthesis of key CCM intermediates such as acetyl-CoA and pyruvate. Acetyl-CoA enters the TCA cycle via citrate generation, a process that depletes acetyl-CoA. Overexpression of TPIA and fructose-bisphosphate aldolase (FBAA) decreases acetyl-CoA flux to the TCA cycle, resulting in a 3-fold increase in PHB concentration in E. coli [66]. In addition, the introduction of an efficient citrate synthase (CS) mutant could also reduce the consumption of acetyl-CoA by the TCA cycle, shifting the metabolic flux to acetate synthesis, which yields 0.24 g/L on glucose in E. coli [67]. Similarly, carbon flux from acetyl-CoA can be transferred to pyruvate by creating the pyruvate dehydrogenase complex mutant. This mutant was overexpressed in E. coli, and by knocking down lactate dehydrogenase (LDHA) and pyruvate oxidase (POXB), it was able to accumulate 17.1 g/L of pyruvate in fermentation [68]. A summary of the applications of CCM with modulated enzymes in E. coli was provided in Fig. 2.
Fig. 2.

The schematic diagram of the CCM with modulated enzymes in E. coli. The black line represented glycolysis and pentose phosphate pathway, and the green line represented the tricarboxylic acid cycle. The red line represented the glucose PTS system and the blue line represented the pathway catalyzed by modulated enzymes. The orange dot represented the products in CCM, while the purple dot represented the products in glucose PTS system. G6P, Glucose 6-phosphate; F6P, Fructose-6-phosphate; F1,6P, Fructose-1,6-bisphosphate; GAP, Glyceraldehyde 3-phosphate; DHAP, Dihydroxyacetone phosphate; 13DPG, 3-Phospho-D-glyceroyl phosphate; 3PG, 3-Phosphoglycerate; 2PG, 2-Phosphoglycerate; PEP, Phosphoenolpyruvate; 6PG15L, 6-Phospho D-glucono-1,5-lactone; X5P, Xylulose 5-phophate; Ru5P, Ribulose 5-phophate; R5P, Ribose 5-phosphate; E4P, Erythrose 4-phosphate; S7P, Sedoheptulose 7-phophate; EI, Phosphotransferases enzyme I; Hpr, Histidine phosphate carrier protein; EIIAB, Phosphotransferases enzyme II A and B; EIIC, Phosphotransferases enzyme II C; PGI, Glucose-6-phosphate isomerase; ZWF, Glucose-6-phosphate dehydrogenase; PYK, Pyruvate kinase; PEPC, Phosphoenolpyruvate carboxylase; TPIA, Triosephosphate isomerase; FBAA, Fructose-bisphosphate aldolase; CS, Citrate synthase; LDHA, Lactate dehydrogenase; POXB, Pyruvate oxidase
Optimization of key regulatory factors
The overexpression of the D. radiodurans response regulator DR1558 in E. coli increased its tolerance to oxidative stress. Moreover, the upregulated expression level of genes involved in CCM and increased accumulation of NADPH form PPP induced by this regulator resulted in PHB production of 5.31 g/L [24].
Other optimization methods
CCM can also be regulated by the switch system. A temperature-sensitive switch system can divide the E. coli fermentation process into growth and production phases. The system regulates the expression of pyruvate carboxylase and oxaloacetic decarboxylase at different temperatures, rebalancing the carbon flux between pyruvate and oxaloacetate. The introduction of this switch system resulted in a record L-threonine yield of 124.03% [70]. Another metabolic switch inhibits the metabolic flux from glycolysis to the TCA cycle and promotes a significant accumulation of acetyl-CoA, yielding 54.2 ± 1.5 mM of the target compound 3-HP with productivity of 32.1 ± 1.3% [71].
CCM optimization in Corynebacterium glutamicum
C. glutamicum is a chassis-engineered strain used for the biosynthesis of polyphenols, where an insufficient supply of acetyl-CoA is an important limiting factor for increasing polyphenol product yield. A moderate reduction of carbon flux in the TCA cycle can effectively decrease the consumption of acetyl-CoA. A 90% reduction in the catalytic activity of CS in the TCA cycle could lead to a 10-fold increase the yield of the target compound naringenin to 19 mg/L [72].
PEP is a key precursor in various biosynthetic pathways. Therefore, reducing PEP depletion can significantly increase the yield of related target products. The knockdown of PEPC in C. glutamicum resulted in a 9.3% increase in the (3R)-acetoin yield to 11.96 g/L [73]. The same strategy elevated isopropanol yield by 1.42-fold [74]. Another common method for decreasing carbon flux consumption by PEP is to optimize the PTS transport system. Knocking out the PTS system and introducing inositol transporter proteins lacking the IolR regulators restored glucose uptake and increased L-serine production to 26.40 g/L [75]. The introduction of mutant inositol transporter proteins in the retained PTS system did not increase hydroxybenzoic acid (HBA) production, but it did shorten the incubation time to achieve the maximum yield. Further suppression of CS expression significantly increased the production rate and yield of hydroxybenzoic acids, reaching 3.1 g/L within 48 h [76].
CCM optimization in Bacillus licheniformis
Because of its robustness and rapid growth, B. licheniformis is widely used as a chassis strain for the synthesis of various biochemicals. Optimization of B. licheniformis CCM can effectively increase the yield of poly-γ-glutamic acid (γ-PGA). Overexpression of PDH and CS directed more carbon fluxes to the TCA cycle, resulting in γ-PGA yields of 34.93% and 11.14%, respectively. Furthermore, knockdown of pyruvate formate-lyase reduced bypass depletion and increased γ-PGA yield by 30.70% [77]. In 2-PE biosynthesis, knockdown of PYK significantly increased PEP supply capacity and allowed for a 79% increase in 2-PE production (0.50 g/L). Deleting the PTS system and replacing it with a transporter system that does not consume PEP could increase the yield of 2-PE [78].
CCM optimization in other prokaryotic chassis
Pseudomonas putida has a substantial advantage in the production of some toxic products due to its high tolerance to organic solvents. The introduction of a promoter-optimized rhamnolipid synthesis pathway in P. putida could direct the metabolic flux of glycolysis and acetyl-CoA to rhamnolipid synthesis, leading to a one-fold increase in rhamnolipid production to 3 g/L [79]. Regulation of CS and ACC gene expression in P. putida using CRISPR/Cas9 technology resulted in an 8-fold increase in acetyl-CoA production [80]. The development of pyruvate-responsive genetic pathways in B. subtilis enables autonomous dynamic control of CCM, resulting in 527 mg/L glucaric acid production, a 154% increase compared to the control strain [81]. The knockdown of the glucose-6-phosphate dehydrogenase gene in Actinosynnema pretiosum reduced the carbon flux in PPP and resulted in a 3-fold increase in the production of ansamitocins [82]. The use of PDC in the development of a CCM metabolic flux control valve device in Zymomonas mobilis resulted in the efficient synthesis of lactate and isobutanol, with lactate and isobutanol production reaching 70% and 65% of the theoretical maximum, respectively [83]. The knockdown of the fructose PTS system in Mannheimia succiniciproducens attenuates the inhibition of carbon catabolism and increases the availability of pyruvic acid. Ultimately, succinate production could be increased to 69.2 g/L by reconstituting the succinate biosynthetic pathway [84].
After knocking out the transcriptional regulator of glyceraldehyde-3-phosphate dehydrogenase and phosphoenolpyruvate carboxykinase CcpNGtg in Geobacillus thermoglucosidasius, the CCM metabolic flux was redirected to the riboflavin biosynthetic pathway, resulting in a 1.51-fold increase in riboflavin production from 171.6 mg/L to 260.3 mg/L [85]. Overexpression of Sp13016, a transcriptional regulator of the glycolytic pathway in Saccharopolyspora pogona, increased the metabolic flux in glycolysis, and decreased the metabolic flux in the TCA cycle and PPP, and resulted in the accumulation of pyruvate and acetyl-CoA, and subsequently increased butenyl-spinosyn production [86]. The development of sensor-assisted transcriptional regulation engineering utilizing the transcriptional regulator QscR in Methylobacterium extorquens increased acetyl-CoA production by approximately 7%, with mevalonate production reaching 2.67 g/L [87]. The applications of CCM optimization in prokaryotic chassis were listed in Table 2.
Table 2.
Optimization of CCM in prokaryotic chassis
| Host | Manipulation | CCM involved | Products | References |
|---|---|---|---|---|
| E. coli | Deletion of PTS system and TyrR repressor | Glycolysis | L-Tyrosine | [52] |
| E. coli | Deletion of PTS system and TyrR repressor | Glycolysis | Melanin | [53] |
| E. coli | Deletion of PTS system and TyrR repressor | Glycolysis | L-DOPA | [54] |
| E. coli | Deletion of PTS system and repression of the activity of Alanine: H+ symporter | Glycolysis | β-alanine | [51] |
| E. coli | Deletion of ZWF and TPIA | Glycolysis | 3-Hydroxypropionic acid | [62] |
| E. coli | Construction of variants of the pyruvate dehydrogenase complex and deletion of LDHA and POXB | Glycolysis | Pyruvate | [68] |
| E. coli | Regulation of carbon distribution by a thermal switch system | Glycolysis | L-Threonine | [70] |
| E. coli | Design and construction of non-oxidative glycolysis pathway | Glycolysis | Acetyl-CoA | [69] |
| E. coli | Silence of a dozen or more CCM enzymes by CRISPR system | Glycolysis and TCA cycle | (2 S)-Naringenin | [63] |
| E. coli | High-throughput screening of CCM key enzymes and fine-tuning of coding genes by CRISPR silencing system | Glycolysis and TCA cycle | (2 S)-Pinocembrin | [64] |
| E. coli | High-throughput screening of CCM key enzymes and fine-tuning of coding genes by CRISPR silencing system | Glycolysis and TCA cycle | Medium chain fatty acids | [65] |
| E. coli | Expression of response regulator DR1558 from D. radiodurans | Glycolysis and TCA cycle | Poly‑3‑hydroxybutyrate | [24] |
| E. coli | Introduction of metabolic toggle switch | Glycolysis and TCA cycle | 3-Hydroxypropionic acid | [71] |
| E. coli | Replacement of PTS system with galactose translocation system | Glycolysis and TCA cycle | Fumaric acid | [56] |
| E. coli | Substitution of PTS system with independent glucose transport system and overexpression of PK | Glycolysis and PPP | L-tryptophan | [57] |
| E. coli | ZWF deletion | Glycolysis and PPP | Lycopene | [59] |
| E. coli | ZWF deletion | Glycolysis and PPP | β-Carotene | [61] |
| E. coli | Overexpression of TPIA and FBAA | TCA cycle | Poly‑3‑hydroxybutyrate | [66] |
| E. coli | Introduction of the efficient citrate synthase variant | TCA cycle | Acetate | [67] |
| E. coli | Deletion of PTS system, PYK, PEPC and Malic enzyme | TCA cycle | Succinate | [55] |
| E. coli | Deletion of PGI and overexpression of ACS | PPP | Riboflavin | [60] |
| C. glutamicum | PEPC deletion | Glycolysis | (3R)-Acetoin | [73] |
| C. glutamicum | PEPC deletion | Glycolysis | Isopropanol | [74] |
| C. glutamicum | Deletion of PTS system and depression of myo-inositol catabolism repressor IolR | Glycolysis | L-Serine | [75] |
| C. glutamicum | Introduction of myo-inositol/proton symporter variant and downregulate of Cs | Glycolysis | Hydroxybenzoic acids | [76] |
| C. glutamicum | Reduction of CS catalysis activity | TCA cycle | Naringenin | [72] |
| B. licheniformis | Deletion of PTS system and PYK | Glycolysis | 2-phenylethanol | [78] |
| B. licheniformis | Overexpression of PDH and Cs and deletion of pyruvate formate-lyase gene | TCA cycle | Poly-γ-glutamic acid | [77] |
| Z. mobilis | Ectopic expression of PDC to construct a CCM control-valve | Glycolysis | Lactate and isobutanol | [83] |
| M. succiniciproducens | Deletion of fructose PTS system | Glycolysis | Succinic acid | [84] |
| G. thermoglucosidasius | Deletion of transcriptional regulator ccpNGtg | Glycolysis and PPP | Riboflavin | [85] |
| B. subtilis | Establishment of a pyruvate-responsive genetic circuit | Glycolysis and TCA cycle | Glucaric acid | [81] |
| P. putida | Introduction of genes for rhamnolipid synthesis | Glycolysis and TCA cycle | Rhamnolipid | [79] |
| S. pogona | Overexpression of the transcriptional regulator Sp13016 | Glycolysis, PPP and TCA cycle | Butenyl-spinosyn | [86] |
| A. pretiosum | ZWF deletion | PPP | Ansamitocins | [82] |
| M. extorquens | Construction of a sensor by transcriptional regulator QscR | PPP and TCA cycle | Acetyl-CoA and Mevalonate | [87] |
Conclusions
The CCM-based optimization strategy can rearrange the metabolic flux in various microbial host strains to increase the supply of precursors in the biosynthetic pathway of target compounds, thereby improving the substrate conversion rate. Therefore, it is critical to maximize the production potential of microbial cell factories and improve compound production efficiency. This strategy is currently being employed to increase the yield of target compounds through metabolic engineering optimization in various chassis strains. Notably, in eukaryotic hosts, optimization of all the three CCM pathways could be used for the biosynthesis of acetyl-CoA derived compounds (e.g., terpenoids and fatty acids derivatives). In the meantime, the optimization of glycolysis and TCA cycles can also be available for the production of organic acids (e.g., pyruvate and succinate), and PPP optimization is used for the metabolic engineering of shikimic acid and aromatic amino acid derivatives (e.g., tyrosol and p-hydroxycinnamic acid). While in prokaryotic hosts, manipulation of glycolysis and PPP is used for the production of a wide range of compounds, including different amino acids (e.g., L-tyrosine and L-threonine) and acetyl-CoA derived chemicals (e.g., terpenes). And the optimization TCA cycle is available for the biosynthesis of organic acids and flavonoids (e.g., naringenin).
However, the regulatory mechanism of CCM is not yet clear due to a large number of genes included, resulting in insufficient CCM optimization approaches. CCM optimization strategies at this stage typically focus on a few functionally defined and repeatedly validated methods, limiting the wide application of CCM optimization in metabolic engineering. These issues can be addressed in the future in the following ways:
To deepen our understanding of CCM regulation network based on species and genetic diversity. As the regulation of CCM is sophisticated, the exploration of regulation mechanisms of CCM from the microbial hosts and other species will be helpful in developing more manipulation methods for CCM optimization.
To develop more transcriptional regulatory factors and gene expression regulatory switches. These regulatory elements can be used to optimize multiple genes and promote the bottleneck reaction(s) of rate-limiting enzyme(s) involved in CCM.
To reduce or eliminate precursor/product consuming pathways related to CCM by balancing metabolic flux or regulating metabolic pathway genes dynamically using key enzymes at metabolic pathway intersections.
Author contributions
Z.W. and X.L. searched and collected the literatures, M.L. and M.M. helped to collect literatures; T.A. and G.W. wrote the original manuscript; D.L. and G.W. edited the draft; Q.Z. reviewed the manuscript. All authors read and approved the manuscript.
Funding
This work was supported by Shandong Provincial Natural Science Foundation (No. ZR2021QC097 to Tianyue An), Introduction and Cultivation Project for Young Creative Talents of Higher Education of Shandong Province (to Guoli Wang), and Yantai School Land Integration Development Project in 2021 (2021XDRHXMPT15 to Defang Li).
Data availability
Not applicable.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Footnotes
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Zhenke Wu and Xiqin Liang contributed equally to this work.
Contributor Information
Defang Li, Email: lidefang@163.com.
Tianyue An, Email: antianyue2007@126.com.
Guoli Wang, Email: trwangli@163.com.
References
- 1.Bailey JE. Toward a science of metabolic engineering. Science. 1991;252:1668–75. doi: 10.1126/science.2047876. [DOI] [PubMed] [Google Scholar]
- 2.Stephanopoulos G, Vallino JJ. Network rigidity and metabolic engineering in metabolite overproduction. Science. 1991;252:1675–81. doi: 10.1126/science.1904627. [DOI] [PubMed] [Google Scholar]
- 3.Ro DK, Paradise EM, Ouellet M, Fisher KJ, Newman KL, Ndungu JM, Ho KA, Eachus RA, Ham TS, Kirby J, et al. Production of the antimalarial drug precursor artemisinic acid in engineered yeast. Nature. 2006;440:940–3. doi: 10.1038/nature04640. [DOI] [PubMed] [Google Scholar]
- 4.Jiang F, Zhou C, Li Y, Deng H, Gong T, Chen J, Chen T, Yang J, Zhu P. Metabolic engineering of yeasts for green and sustainable production of bioactive ginsenosides F2 and 3β, 20S-Di-O-Glc-DM. Acta Pharm Sin B. 2022;12:3167–76. doi: 10.1016/j.apsb.2022.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Galanie S, Thodey K, Trenchard IJ, Filsinger Interrante M, Smolke CD. Complete biosynthesis of opioids in yeast. Science. 2015;349:1095–100. doi: 10.1126/science.aac9373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Zhang J, Hansen LG, Gudich O, Viehrig K, Lassen LMM, Schrübbers L, Adhikari KB, Rubaszka P, Carrasquer-Alvarez E, Chen L, et al. A microbial supply chain for production of the anti-cancer drug vinblastine. Nature. 2022;609:341–7. doi: 10.1038/s41586-022-05157-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lian J, Chao R, Zhao H. Metabolic engineering of a Saccharomyces cerevisiae strain capable of simultaneously utilizing glucose and galactose to produce enantiopure (2R,3R)-butanediol. Metab Eng. 2014;23:92–9. doi: 10.1016/j.ymben.2014.02.003. [DOI] [PubMed] [Google Scholar]
- 8.Zhu X, Tan Z, Xu H, Chen J, Tang J, Zhang X. Metabolic evolution of two reducing equivalent-conserving pathways for high-yield succinate production in Escherichia coli. Metab Eng. 2014;24:87–96. doi: 10.1016/j.ymben.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 9.Zheng Y, Liu Q, Li L, Qin W, Yang J, Zhang H, Jiang X, Cheng T, Liu W, Xu X, Xian M. Metabolic engineering of Escherichia coli for high-specificity production of isoprenol and prenol as next generation of biofuels. Biotechnol Biofuels. 2013;6:57. doi: 10.1186/1754-6834-6-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Choi YJ, Lee SY. Microbial production of short-chain alkanes. Nature. 2013;502:571–4. doi: 10.1038/nature12536. [DOI] [PubMed] [Google Scholar]
- 11.Shen R, Yin J, Ye JW, Xiang RJ, Ning ZY, Huang WZ, Chen GQ. Promoter engineering for enhanced P (3HB-co-4HB) production by Halomonas bluephagenesis. ACS Synth Biol. 2018;7:1897–906. doi: 10.1021/acssynbio.8b00102. [DOI] [PubMed] [Google Scholar]
- 12.Lian J, Jin R, Zhao H. Construction of plasmids with tunable copy numbers in Saccharomyces cerevisiae and their applications in pathway optimization and multiplex genome integration. Biotechnol Bioeng. 2016;113:2462–73. doi: 10.1002/bit.26004. [DOI] [PubMed] [Google Scholar]
- 13.Wang T, Tague N, Whelan SA, Dunlop MJ. Programmable gene regulation for metabolic engineering using decoy transcription factor binding sites. Nucleic Acids Res. 2021;49:1163–72. doi: 10.1093/nar/gkaa1234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhang Y, Li SZ, Li J, Pan X, Cahoon RE, Jaworski JG, Wang X, Jez JM, Chen F, Yu O. Using unnatural protein fusions to engineer resveratrol biosynthesis in yeast and mammalian cells. J Am Chem Soc. 2006;128:13030–1. doi: 10.1021/ja0622094. [DOI] [PubMed] [Google Scholar]
- 15.Liu CL, Dong HG, Xue K, Sun L, Yang Y, Liu X, Li Y, Bai Z, Tan TW. Metabolic engineering mevalonate pathway mediated by RNA scaffolds for mevalonate and isopreneproduction in Escherichia coli. ACS Synth Biol. 2022;11:3305–17. doi: 10.1021/acssynbio.2c00226. [DOI] [PubMed] [Google Scholar]
- 16.Grewal PS, Samson JA, Baker JJ, Choi B, Dueber JE. Peroxisome compartmentalization of a toxic enzyme improves alkaloid production. Nat Chem Biol. 2021;17:96–103. doi: 10.1038/s41589-020-00668-4. [DOI] [PubMed] [Google Scholar]
- 17.Zhou P, Xie W, Yao Z, Zhu Y, Ye L, Yu H. Development of a temperature-responsive yeast cell factory using engineered Gal4 as a protein switch. Biotechnol Bioeng. 2018;115:1321–30. doi: 10.1002/bit.26544. [DOI] [PubMed] [Google Scholar]
- 18.Yim H, Haselbeck R, Niu W, Pujol-Baxley C, Burgard A, Boldt J, Khandurina J, Trawick JD, Osterhout RE, Stephen R, et al. Metabolic engineering of Escherichia coli for direct production of 1,4-butanediol. Nat Chem Biol. 2011;7:445–52. doi: 10.1038/nchembio.580. [DOI] [PubMed] [Google Scholar]
- 19.Raab AM, Gebhardt G, Bolotina N, Weuster-Botz D, Lang C. Metabolic engineering of Saccharomyces cerevisiae for the biotechnological production of succinic acid. Metab Eng. 2010;12:518–25. doi: 10.1016/j.ymben.2010.08.005. [DOI] [PubMed] [Google Scholar]
- 20.Liu Q, Liu Y, Li G, Savolainen O, Chen Y, Nielsen J. De novo biosynthesis of bioactive isoflavonoids by engineered yeast cell factories. Nat Commun. 2021;12:6085. doi: 10.1038/s41467-021-26361-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lu X, Liu Y, Yang Y, Wang S, Wang Q, Wang X, Yan Z, Cheng J, Liu C, Yang X, et al. Constructing a synthetic pathway for acetyl-coenzyme A from one-carbon through enzyme design. Nat Commun. 2019;10:1378. doi: 10.1038/s41467-019-09095-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Meng W, Zhang L, Cao M, Zhang Y, Zhang Y, Li P, Kang Z, Guo S, Xu P, Ma C, et al. 2,3-Butanediol synthesis from glucose supplies NADH for elimination of toxic acetate produced during overflow metabolism. Cell Discov. 2021;7:43. doi: 10.1038/s41421-021-00273-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Meadows AL, Hawkins KM, Tsegaye Y, Antipov E, Kim Y, Raetz L, Dahl RH, Tai A, Mahatdejkul-Meadows T, Xu L, et al. Rewriting yeast central carbon metabolism for industrial isoprenoid production. Nature. 2016;537:694–7. doi: 10.1038/nature19769. [DOI] [PubMed] [Google Scholar]
- 24.Park SH, Kim GB, Kim HU, Park SJ, Choi JI. Enhanced production of poly-3hydroxybutyrate (PHB) by expression of response regulator DR1558 in recombinant Escherichia coli. Int J Biol Macromol. 2019;131:29–35. doi: 10.1016/j.ijbiomac.2019.03.044. [DOI] [PubMed] [Google Scholar]
- 25.Kocharin K, Siewers V, Nielsen J. Improved polyhydroxybutyrate production by Saccharomyces cerevisiae through the use of the phosphoketolase pathway. Biotechnol Bioeng. 2013;110:2216–24. doi: 10.1002/bit.24888. [DOI] [PubMed] [Google Scholar]
- 26.Kamineni A, Consiglio AL, MacEwen K, Chen S, Chifamba G, Shaw AJ, Tsakraklides V. Increasing lipid yield in Yarrowia lipolytica through phosphoketolase and phosphotransacetylase expression in a phosphofructokinase deletion strain. Biotechnol Biofuels. 2021;14:113. doi: 10.1186/s13068-021-01962-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Zhou YJ, Buijs NA, Zhu Z, Qin J, Siewers V, Nielsen J. Production of fatty acid-derived oleochemicals and biofuels by synthetic yeast cell factories. Nat Commun. 2016;7:11709. doi: 10.1038/ncomms11709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Cai P, Wu X, Deng J, Gao L, Shen Y, Yao L, Zhou YJ. Methanol biotransformation toward high-level production of fatty acid derivatives by engineering the industrial yeast Pichia pastoris. Proc Natl Acad Sci U S A. 2022;119:e2201711119. doi: 10.1073/pnas.2201711119. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.de Jong BW, Shi S, Siewers V, Nielsen J. Improved production of fatty acid ethyl esters in Saccharomyces cerevisiae through up-regulation of the ethanol degradation pathway and expression of the heterologous phosphoketolase pathway. Microb Cell Fact. 2014;13:39. doi: 10.1186/1475-2859-13-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Liu Q, Yu T, Li X, Chen Y, Campbell K, Nielsen J, Chen Y. Rewiring carbon metabolism in yeast for high level production of aromatic chemicals. Nat Commun. 2019;10:4976. doi: 10.1038/s41467-019-12961-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Guo W, Huang Q, Feng Y, Tan T, Niu S, Hou S, Chen Z, Du ZQ, Shen Y, Fang X. Rewiring central carbon metabolism for tyrosol and salidroside production in Saccharomyces cerevisiae. Biotechnol Bioeng. 2020;117:2410–19. doi: 10.1002/bit.27370. [DOI] [PubMed] [Google Scholar]
- 32.Hassing EJ, de Groot PA, Marquenie VR, Pronk JT, Daran JG. Connecting central carbon and aromatic amino acid metabolisms to improve de novo 2-phenylethanol production in Saccharomyces cerevisiae. Metab Eng. 2019;56:165–80. doi: 10.1016/j.ymben.2019.09.011. [DOI] [PubMed] [Google Scholar]
- 33.Gao X, Caiyin Q, Zhao F, Wu Y, Lu W. Engineering Saccharomyces cerevisiae for enhanced production of protopanaxadiol with cofermentation of glucose and xylose. J Agric Food Chem. 2018;66:12009–16. doi: 10.1021/acs.jafc.8b04916. [DOI] [PubMed] [Google Scholar]
- 34.Qin N, Li L, Ji X, Li X, Zhang Y, Larsson C, Chen Y, Nielsen J, Liu Z. Rewiring central carbon metabolism ensures increased provision of acetyl-CoA and NADPH required for 3-OH-propionic acid production. ACS Synth Biol. 2020;9:3236–44. doi: 10.1021/acssynbio.0c00264. [DOI] [PubMed] [Google Scholar]
- 35.Rodriguez S, Denby CM, Van Vu T, Baidoo EE, Wang G, Keasling JD. ATP citrate lyase mediated cytosolic acetyl-CoA biosynthesis increases mevalonate production in Saccharomyces cerevisiae. Microb Cell Fact. 2016;15:48. doi: 10.1186/s12934-016-0447-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Cardenas J, Da Silva NA. Engineering cofactor and transport mechanisms in Saccharomyces cerevisiae for enhanced acetyl-CoA and polyketide biosynthesis. Metab Eng. 2016;36:80–9. doi: 10.1016/j.ymben.2016.02.009. [DOI] [PubMed] [Google Scholar]
- 37.Song JY, Park JS, Kang CD, Cho HY, Yang D, Lee S, Cho KM. Introduction of a bacterial acetyl-CoA synthesis pathway improves lactic acid production in Saccharomyces cerevisiae. Metab Eng. 2016;35:38–45. doi: 10.1016/j.ymben.2015.09.006. [DOI] [PubMed] [Google Scholar]
- 38.Liu H, Fan J, Wang C, Li C, Zhou X. Enhanced β-amyrin synthesis in Saccharomyces cerevisiae by coupling an optimal acetyl-CoA supply pathway. J Agric Food Chem. 2019;67:3723–32. doi: 10.1021/acs.jafc.9b00653. [DOI] [PubMed] [Google Scholar]
- 39.Nagamori E, Shimizu K, Fujita H, Tokuhiro K, Ishida N, Takahashi H. Metabolic flux analysis of genetically engineered Saccharomyces cerevisiae that produces lactate under micro-aerobic conditions. Bioprocess Biosyst Eng. 2013;36:1261–5. doi: 10.1007/s00449-012-0870-6. [DOI] [PubMed] [Google Scholar]
- 40.Ishii J, Morita K, Ida K, Kato H, Kinoshita S, Hataya S, Shimizu H, Kondo A, Matsuda F. A pyruvate carbon flux tugging strategy for increasing 2,3-butanediol production and reducing ethanol subgeneration in the yeast Saccharomyces cerevisiae. Biotechnol Biofuels. 2018;11:180. doi: 10.1186/s13068-018-1176-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tan SZ, Manchester S, Prather KL. Controlling central carbon metabolism for improved pathway yields in Saccharomyces cerevisiae. ACS Synth Biol. 2016;5:116–24. doi: 10.1021/acssynbio.5b00164. [DOI] [PubMed] [Google Scholar]
- 42.Zhang Q, Wang X, Luo H, Wang Y, Wang Y, Tu T, Qin X, Su X, Huang H, Yao B, Bai Y, Zhang J. Metabolic engineering of Pichia pastoris for myo-inositol production by dynamic regulation of central metabolism. Microb Cell Fact. 2022;21:112. doi: 10.1186/s12934-022-01837-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Cardenas J, Da Silva NA. Metabolic engineering of Saccharomyces cerevisiae for the production of triacetic acid lactone. Metab Eng. 2014;25:194–203. doi: 10.1016/j.ymben.2014.07.008. [DOI] [PubMed] [Google Scholar]
- 44.Liu Y, Bai C, Liu Q, Xu Q, Qian Z, Peng Q, Yu J, Xu M, Zhou X, Zhang Y, et al. Engineered ethanol-driven biosynthetic system for improving production of acetyl-CoA derived drugs in Crabtree-negative yeast. Metab Eng. 2019;54:275–84. doi: 10.1016/j.ymben.2019.05.001. [DOI] [PubMed] [Google Scholar]
- 45.Bensen ES, Yeung BG, Payne GS. Ric1p and the Ypt6p GTPase function in a common pathway required for localization of trans-golgi network membrane proteins. Mol Biol Cell. 2001;12:13–26. doi: 10.1091/mbc.12.1.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Suástegui M, Yu Ng C, Chowdhury A, Sun W, Cao M, House E, Maranas CD, Shao Z. Multilevel engineering of the upstream module of aromatic amino acid biosynthesis in Saccharomyces cerevisiae for high production of polymer and drug precursors. Metab Eng. 2017;42:134–44. doi: 10.1016/j.ymben.2017.06.008. [DOI] [PubMed] [Google Scholar]
- 47.Durall C, Lindberg P, Yu J, Lindblad P. Increased ethylene production by overexpressing phosphoenolpyruvate carboxylase in the cyanobacterium Synechocystis PCC 6803. Biotechnol Biofuels. 2020;13:16. doi: 10.1186/s13068-020-1653-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hasunuma T, Matsuda M, Kato Y, Vavricka CJ, Kondo A. Temperature enhanced succinate production concurrent with increased central metabolism turnover in the cyanobacterium Synechocystis sp. PCC 6803. Metab Eng. 2018;48:109–20. doi: 10.1016/j.ymben.2018.05.013. [DOI] [PubMed] [Google Scholar]
- 49.Deutscher J, Francke C, Postma PW. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria. Microbiol Mol Biol Rev. 2006;70:939–1031. doi: 10.1128/MMBR.00024-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sauer U, Eikmanns BJ. The PEP-pyruvate-oxaloacetate node as the switch point for carbon flux distribution in bacteria. FEMS Microbiol Rev. 2005;29:765–94. doi: 10.1016/j.femsre.2004.11.002. [DOI] [PubMed] [Google Scholar]
- 51.Li B, Zhang B, Wang P, Cai X, Chen YY, Yang YF, Liu ZQ, Zheng YG. Rerouting fluxes of the central carbon metabolism and relieving mechanism-based inactivation of L-aspartate-α-decarboxylase for fermentative production of β-alanine in Escherichia coli. ACS Synth Biol. 2022;11:1908–18. doi: 10.1021/acssynbio.2c00055. [DOI] [PubMed] [Google Scholar]
- 52.Lütke-Eversloh T, Stephanopoulos G. L-tyrosine production by deregulated strains of Escherichia coli. Appl Microbiol Biotechnol. 2007;75:103–10. doi: 10.1007/s00253-006-0792-9. [DOI] [PubMed] [Google Scholar]
- 53.Chávez-Béjar MI, Balderas-Hernandez VE, Gutiérrez-Alejandre A, Martinez A, Bolívar F, Gosset G. Metabolic engineering of Escherichia coli to optimize melanin synthesis from glucose. Microb Cell Fact. 2013;12:108. doi: 10.1186/1475-2859-12-108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fordjour E, Adipah FK, Zhou S, Du G, Zhou J. Metabolic engineering of Escherichia coli BL21 (DE3) for de novo production of L-DOPA from D-glucose. Microb Cell Fact. 2019;18:74. doi: 10.1186/s12934-019-1122-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Zhao Y, Wang CS, Li FF, Liu ZN, Zhao GR. Targeted optimization of central carbon metabolism for engineering succinate production in Escherichia coli. BMC Biotechnol. 2016;16:52. doi: 10.1186/s12896-016-0284-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Liu H, Song R, Liang Y, Zhang T, Deng L, Wang F, Tan T. Genetic manipulation of Escherichia coli central carbon metabolism for efficient production of fumaric acid. Bioresour Technol. 2018;270:96–102. doi: 10.1016/j.biortech.2018.08.024. [DOI] [PubMed] [Google Scholar]
- 57.Xiong B, Zhu Y, Tian D, Jiang S, Fan X, Ma Q, Wu H, Xie X. Flux redistribution of central carbon metabolism for efficient production of L-tryptophan in Escherichia coli. Biotechnol Bioeng. 2021;118:1393–404. doi: 10.1002/bit.27665. [DOI] [PubMed] [Google Scholar]
- 58.Misawa N, Nakagawa M, Kobayashi K, Yamano S, Izawa Y, Nakamura K, Harashima K. Elucidation of the Erwinia uredovora carotenoid biosynthetic pathway by functional analysis of gene products expressed in Escherichia coli. J Bacteriol. 1990;172:6704–12. doi: 10.1128/jb.172.12.6704-6712.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Zhou Y, Nambou K, Wei L, Cao J, Imanaka T, Hua Q. Lycopene production in recombinant strains of Escherichia coli is improved by knockout of the central carbon metabolism gene coding for glucose-6-phosphate dehydrogenase. Biotechnol Lett. 2013;35:2137–45. doi: 10.1007/s10529-013-1317-0. [DOI] [PubMed] [Google Scholar]
- 60.Lin Z, Xu Z, Li Y, Wang Z, Chen T, Zhao X. Metabolic engineering of Escherichia coli for the production of riboflavin. Microb Cell Fact. 2014;13:104. doi: 10.1186/s12934-014-0104-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Wu Y, Yan P, Li Y, Liu X, Wang Z, Chen T, Zhao X. Enhancing β-carotene production in Escherichia coli by perturbing central carbon metabolism and improving the NADPH supply. Front Bioeng Biotechnol. 2020;8:585. doi: 10.3389/fbioe.2020.00585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Tokuyama K, Ohno S, Yoshikawa K, Hirasawa T, Tanaka S, Furusawa C, Shimizu H. Increased 3-hydroxypropionic acid production from glycerol, by modification of central metabolism in Escherichia coli. Microb Cell Fact. 2014;13:64. doi: 10.1186/1475-2859-13-64. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Wu J, Du G, Chen J, Zhou J. Enhancing flavonoid production by systematically tuning the central metabolic pathways based on a CRISPR interference system in Escherichia coli. Sci Rep. 2015;5:13477. doi: 10.1038/srep13477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Wu J, Zhang X, Zhou J, Dong M. Efficient biosynthesis of (2S)-pinocembrin from d-glucose by integrating engineering central metabolic pathways with a pH-shift control strategy. Bioresour Technol. 2016;218:999–1007. doi: 10.1016/j.biortech.2016.07.066. [DOI] [PubMed] [Google Scholar]
- 65.Wu J, Wang Z, Duan X, Zhou P, Liu P, Pang Z, Wang Y, Wang X, Li W, Dong M. Construction of artificial micro-aerobic metabolism for energy-and carbon-efficient synthesis of medium chain fatty acids in Escherichia coli. Metab Eng. 2019;53:1–13. doi: 10.1016/j.ymben.2019.01.006. [DOI] [PubMed] [Google Scholar]
- 66.Lee SH, Kang KH, Kim EY, Chae TU, Oh YH, Hong SH, Song BK, Jegals J, Park SJ, Lee SY. Metabolic engineering of Escherichia coli for enhanced biosynthesis of poly(3-hydroxybutyrate) based on proteome analysis. Biotechnol Lett. 2013;35:1631–7. doi: 10.1007/s10529-013-1246-y. [DOI] [PubMed] [Google Scholar]
- 67.Tovilla-Coutiño DB, Momany C, Eiteman MA. Engineered citrate synthase alters acetate accumulation in Escherichia coli. Metab Eng. 2020;61:171–80. doi: 10.1016/j.ymben.2020.06.006. [DOI] [PubMed] [Google Scholar]
- 68.Moxley WC, Eiteman MA. Pyruvate production by Escherichia coli by use of pyruvate dehydrogenase variants. Appl Environ Microbiol. 2021;87:e0048721. doi: 10.1128/AEM.00487-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Bogorad IW, Lin TS, Liao JC. Synthetic non-oxidative glycolysis enables complete carbon conservation. Nature. 2013;502:693–7. doi: 10.1038/nature12575. [DOI] [PubMed] [Google Scholar]
- 70.Fang Y, Wang J, Ma W, Yang J, Zhang H, Zhao L, Chen S, Zhang S, Hu X, Li Y, et al. Rebalancing microbial carbon distribution for L-threonine maximization using a thermal switch system. Metab Eng. 2020;61:33–46. doi: 10.1016/j.ymben.2020.01.009. [DOI] [PubMed] [Google Scholar]
- 71.Tsuruno K, Honjo H, Hanai T. Enhancement of 3-hydroxypropionic acid production from glycerol by using a metabolic toggle switch. Microb Cell Fact. 2015;14:155. doi: 10.1186/s12934-015-0342-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Milke L, Ferreira P, Kallscheuer N, Braga A, Vogt M, Kappelmann J, Oliveira J, Silva AR, Rocha I, Bott M, Noack S, Faria N, Marienhagen J. Modulation of the central carbon metabolism of Corynebacterium glutamicum improves malonyl-CoA availability and increases plant polyphenol synthesis. Biotechnol Bioeng. 2019;116:1380–91. doi: 10.1002/bit.26939. [DOI] [PubMed] [Google Scholar]
- 73.Lu L, Mao Y, Kou M, Cui Z, Jin B, Chang Z, Wang Z, Ma H, Chen T. Engineering central pathways for industrial-level (3R)-acetoin biosynthesis in Corynebacterium glutamicum. Microb Cell Fact. 2020;19:102. doi: 10.1186/s12934-020-01363-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Ko YJ, Cha J, Jeong WY, Lee ME, Cho BH, Nisha B, Jeong HJ, Park SE, Han SO. Bio-isopropanol production in Corynebacterium glutamicum: metabolic redesign of synthetic bypasses and two-stage fermentation with gas stripping. Bioresour Technol. 2022;354:127171. doi: 10.1016/j.biortech.2022.127171. [DOI] [PubMed] [Google Scholar]
- 75.Zhang X, Lai L, Xu G, Zhang X, Shi J, Koffas MAG, Xu Z. Rewiring the central metabolic pathway for high-yield L-serine production in Corynebacterium glutamicum by using glucose. Biotechnol J. 2019;14:e1800497. doi: 10.1002/biot.201800497. [DOI] [PubMed] [Google Scholar]
- 76.Kallscheuer N, Marienhagen J. Corynebacterium glutamicum as platform for the production of hydroxybenzoic acids. Microb Cell Fact. 2018;17:70. doi: 10.1186/s12934-018-0923-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Li B, Cai D, Chen S. Metabolic Engineering of central carbon metabolism of Bacillus licheniformis for enhanced production of poly-γ-glutamic acid. Appl Biochem Biotechnol. 2021;193:3540–52. doi: 10.1007/s12010-021-03619-4. [DOI] [PubMed] [Google Scholar]
- 78.Zhan Y, Shi J, Xiao Y, Zhou F, Wang H, Xu H, Li Z, Yang S, Cai D, Chen S. Multilevel metabolic engineering of Bacillus licheniformis for de novo biosynthesis of 2-phenylethanol. Metab Eng. 2022;70:43–54. doi: 10.1016/j.ymben.2022.01.007. [DOI] [PubMed] [Google Scholar]
- 79.Tiso T, Sabelhaus P, Behrens B, Wittgens A, Rosenau F, Hayen H, Blank LM. Creating metabolic demand as an engineering strategy in Pseudomonas putida - rhamnolipid synthesis as an example. Metab Eng Commun. 2016;3:234–44. doi: 10.1016/j.meteno.2016.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Kozaeva E, Volkova S, Matos MRA, Mezzina MP, Wulff T, Volke DC, Nielsen LK, Nikel PI. Model-guided dynamic control of essential metabolic nodes boosts acetyl-coenzyme A-dependent bioproduction in rewired Pseudomonas putida. Metab Eng. 2021;67:373–86. doi: 10.1016/j.ymben.2021.07.014. [DOI] [PubMed] [Google Scholar]
- 81.Xu X, Li X, Liu Y, Zhu Y, Li J, Du G, Chen J, Ledesma-Amaro R, Liu L. Pyruvate-responsive genetic circuits for dynamic control of central metabolism. Nat Chem Biol. 2020;16:1261–68. doi: 10.1038/s41589-020-0637-3. [DOI] [PubMed] [Google Scholar]
- 82.Fan Y, Hu F, Wei L, Bai L, Hua Q. Effects of modulation of pentose-phosphate pathway on biosynthesis of ansamitocins in Actinosynnema pretiosum. J Biotechnol. 2016;230:3–10. doi: 10.1016/j.jbiotec.2016.05.010. [DOI] [PubMed] [Google Scholar]
- 83.Liu Y, Ghosh IN, Martien J, Zhang Y, Amador-Noguez D, Landick R. Regulated redirection of central carbon flux enhances anaerobic production of bioproducts in Zymomonas mobilis. Metab Eng. 2020;61:261–74. doi: 10.1016/j.ymben.2020.06.005. [DOI] [PubMed] [Google Scholar]
- 84.Lee JW, Yi J, Kim TY, Choi S, Ahn JH, Song H, Lee MH, Lee SY. Homo-succinic acid production by metabolically engineered Mannheimia succiniciproducens. Metab Eng. 2016;38:409–17. doi: 10.1016/j.ymben.2016.10.004. [DOI] [PubMed] [Google Scholar]
- 85.Yang Z, Sun Q, Tan G, Zhang Q, Wang Z, Li C, Qi F, Wang W, Zhang L, Li Z. Engineering thermophilic Geobacillus thermoglucosidasius for riboflavin production. Microb Biotechnol. 2021;14:363–73. doi: 10.1111/1751-7915.13543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Tang J, He H, Li Y, Liu Z, Xia Z, Cao L, Zhu Z, Shuai L, Liu Y, Wan Q, et al. Comparative proteomics reveals the effect of the transcriptional regulator Sp13016 on butenyl-spinosyn biosynthesis in Saccharopolyspora pogona. J Agric Food Chem. 2021;69:12554–65. doi: 10.1021/acs.jafc.1c03654. [DOI] [PubMed] [Google Scholar]
- 87.Liang WF, Cui LY, Cui JY, Yu KW, Yang S, Wang TM, Guan CG, Zhang C, Xing XH. Biosensor-assisted transcriptional regulator engineering for Methylobacterium extorquens AM1 to improve mevalonate synthesis by increasing the acetyl-CoA supply. Metab Eng. 2017;39:159–68. doi: 10.1016/j.ymben.2016.11.010. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.