

Keywords: adjuvant, subolesin, tick, tick-borne disease, vaccine, vaccinomics
Abstract
Ticks represent a major concern for society worldwide. Ticks are also difficult to control, and vaccines represent the most efficacious, safe, economically feasible and environmentally sustainable intervention. The evolution of tick vaccinology has been driven by multiple challenges such as (1) Ticks are difficult to control, (2) Vaccines control tick infestations by reducing ectoparasite fitness and reproduction, (3) Vaccine efficacy against multiple tick species, (4) Impact of tick strain genetic diversity on vaccine efficacy, (5) Antigen combination to improve vaccine efficacy, (6) Vaccine formulations and delivery platforms and (7) Combination of vaccines with transgenesis and paratransgenesis. Tick vaccine antigens evolved from organ protein extracts to recombinant proteins to chimera designed by vaccinomics and quantum vaccinomics. Future directions will advance in these areas together with other novel technologies such as multiomics, AI and Big Data, mRNA vaccines, microbiota-driven probiotics and vaccines, and combination of vaccines with other interventions in collaboration with regions with high incidence of tick infestations and tick-borne diseases for a personalized medicine approach.
Challenge 1: ticks are difficult to control
Ticks and tick-borne pathogens constitute a growing problem with increasing social and economic concern worldwide (e.g. de la Fuente et al., 2023a). Ticks are difficult to control, and traditional control methods are mainly based on the use of chemical acaricides with partial success and drawbacks such as selection of resistant ticks and negative impact on animal health and production and environmental contamination (Agwunobi et al., 2021; Githaka et al., 2022; Gonzaga et al., 2023). A number of reports of establishment of multiacaricides resistant ticks in different parts of the world (Bishop et al., 2023) and growing global public concern of environment pollution due to high use of chemical acaricides has posed serious challenges on continuation the use of conventional methods for tick management.
Under the One Health and sustainability perspective, vaccines are the most effective and safe intervention to reduce tick populations and risks associated with transmitted pathogens (de la Fuente, 2018; reviewed by Estrada-Peña et al., 2022). However, although a number of reports of significant efficacy of other vaccine formulations have been reported (de la Fuente and Kocan, 2003; de la Fuente and Contreras, 2015; Bishop et al., 2023; Parizi et al., 2023), only Bm86/Bm95-based vaccines TickGARD in Australia and Gavac in Cuba were registered and commercialized for the control of Rhipicephalus microplus tick infestations (de la Fuente et al., 2007; Rodríguez-Mallon, 2023). Currently, only Gavac (CIGB, Havana, Cuba; https://www.cigb.edu.cu/en/product/gavac-2/) and Bovimune Ixovac (Lapisa, La Piedad, Michoacán, Mexico; https://lapisa.com/productos/bovimune-ixovac) with Bm86 antigen are still commercially available in some Latin American countries.
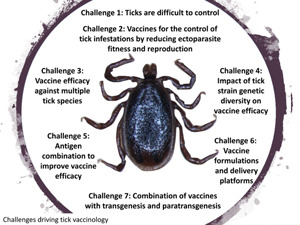
Based on the evolution of vaccinology (Andreano et al., 2019), this review approached the evolution of tick vaccinology to face challenges and advance in the development of new effective anti-tick vaccines and other control interventions (Fig. 1).
Figure 1.

Tick vaccine research in the context of the evolution of vaccinology. Key advances in tick vaccinology are highlighted in red with tick stickers.
Challenge 2: vaccines control tick infestations by reducing ectoparasite fitness and reproduction
The proof-of-concept of anti-tick vaccine was proposed by Allen and Humphreys (1979) using organ specific protein extracts. The first challenge was then approached with the discovery of R. microplus Bm86/Bm95 antigen and the development, registration and commercialization of TickGARD and Gavac vaccines for the control of cattle tick infestations (Willadsen et al., 1988, 1995; Rodríguez et al., 1994; reviewed by de la Fuente and Kocan, 2003; de la Fuente et al., 2007; Rodríguez-Mallon, 2023). The protective mechanism was associated with antibody production in response to vaccine and antibody-antigen interactions in the midgut lumen of ticks feeding on immunized host (Willadsen and Kemp, 1988). This interaction affected tick protein function, which translated into reduction in the number of ticks completing life cycle, weight, oviposition and fertility (de la Fuente and Kocan, 2014). Considering the role of cattle hosts in tick-borne diseases (TBD), these vaccines may not only reduce tick infestations and incidence of TBD in cattle but also in humans and other animal species (Chakraborty et al., 2023). However, due to significant variation in vaccine efficacy reported of 0–100% (de la Fuente and Kocan, 2014; Parizi et al., 2023) against different strains of R. microplus, these vaccines have not been approved in most countries.
Challenge 3: vaccine efficacy against multiple tick species
Despite the advances on anti-tick R. microplus vaccines with Bm86/Bm95 antigens, conserved protective antigens across different tick genera needed to be identified. To address this challenge, Subolesin (SUB; originally named 4D8 and ortholog of Akirin) was discovered by expression library immunization in Ixodes scapularis mouse model (Almazán et al., 2003). The SUB-vaccine protective responses were not only mediated by anti-SUB antibodies entering tick cells by unknown mechanisms and blocking protein translocation to the nucleus to exert its regulatory function, but also through activation of other immune protective mechanisms (de la Fuente et al., 2011, 2021; Merino et al., 2011; Artigas-Jerónimo et al., 2020). The immune response to SUB affects multiple biological processes, which translates in various hosts (e.g. cattle, deer, sheep, dog, rabbit, mouse, chicken) into reduction of fitness and reproduction of different tick species (e.g. Ornithodoros, Ixodes, Haemaphysalis, Amblyomma, Dermacentor, Hyalomma, Rhipicephalus) and other arthropod vectors (e.g. mosquito, sand fly, poultry red mite) and vector-borne pathogens (e.g. Anaplasma, Babesia, Borrelia, Plasmodium) (Artigas-Jerónimo et al., 2018; Parizi et al., 2023) (Table 1). The efficacy and effectiveness of vaccines with SUB antigens have been evaluated not only under pen-controlled conditions (Shakya et al., 2014; Artigas-Jerónimo et al., 2018), but also in field trials (Torina et al., 2014; Mendoza-Martínez et al., 2021). Under field conditions in vaccinated cattle and sheep, the results showed 63% of sheep tick infestations, 8-fold reduction in the per cent of infested cattle, 32–55% reduction in tick weight, reduction in acaricide treatments and in the prevalence of Anaplasma marginale tick-transmitted genotypes (Torina et al., 2014). Recently, SUB vaccine provided a 67% efficacy in cattle infested with R. microplus (Mendoza-Martínez et al., 2021) and 83–90% efficacy in cattle vaccinated with Rhipicephalus appendiculatus SUB and infested with R. appendiculatus, Rhipicephalus decoloratus and Amblyomma variegatum (Kasaija et al., 2020).
Table 1.
Examples of the efficacy of animal immunization with SUB tick protective antigen
| Tick species | Expression system | Host | Dose and delivery | Adjuvant | Efficacy (E) against infestation (%) | Reference |
|---|---|---|---|---|---|---|
| I. scapularis | E. coli | Rabbit | 500 μg 2 doses; S/C | Freund's incomplete adjuvant | 46%, I. scapularis | Almazán et al. (2005) |
| R. microplus | P. pastoris | Chicken | 50 μg 3 doses; S/C | Montanide ISA 50 V | 35.1%, Dermanyssus gallinae gallinae | Harrington et al., 2009 |
| R. microplus | E. coli | Cattle | 100 μg 3 doses; I/M | Montanide ISA 50 V |
R. annulatus larvae, adults, 37–48% |
Almazán et al. (2010) |
| A. americanum | E .coli | Cattle | 100 μm 3 doses; S/C | Montanide ISA 50 V | 55% | de la Fuente et al., (2010) |
| I. scapularis | Vaccinia virus | Mice | 108 pfu given orally | No adjuvant used | 52% | Bensaci et al., 2012 |
| R. microplus | E. coli | White-tailed deer | 100 μg 3 doses; I/M | Montanide ISA 50 V | 83% | Carreón, et al., 2012 |
| R. microplus | P. pastoris | Mice | 25 μg 3 doses; I/M | Montanide ISA 50V2 | I. ricinus larvae, 54% | Moreno-Cid et al., 2013 |
| R. microplus | E. coli | Cattle | 100 μg 3 doses; I/M | Montanide ISA 50 V | 60% | Merino et al., 2013 |
| R. microplus | E. coli | Cattle | 100 μg 2 doses; I/M | Montanide 888 | 44%. | Shakya et al., 2014 |
| R. appendiculatus | E. coli | Cattle | 100 μg 3 doses; I/M | Montanide ISA 50V2 |
R. appendiculatus, 47–90%; A. variegatum, 50–89%; R. decoloratus, 51% |
Kasaija et al., 2020 |
| A. variegatum | E. coli | Cattle | 100 μg 3 doses; I/M | Montanide ISA 50V2 |
R. appendiculatus, 83–86%; A. variegatum, 47–76%; R. decoloratus, 72% |
Kasaija et al., 2020 |
| R. decoloratus | E. coli | Cattle | 100 μg 3 doses; I/M | Montanide ISA 50V2 |
R. appendiculatus 66–89%; A. variegatum 50–89%; R. decoloratus, 51% |
Kasaija et al., 2020 |
| H. longicornis | E.coli | Rabbit | 500 μg 2 doses; I/M | TiterMax Gold | 37.4% | Lee et al., 2020 |
|
R. microplus SUB peptide |
E. coli | Cattle | 100 μg 3 doses; S/C | Montanide ISA 50 | 67% | Mendoza-Martínez et al., 2021 |
| R. microplus | E. coli | Cattle | oral | Montanide ISA 50V2 |
R. decoloratus, 7196%; R. appendiculatus, 87–99% |
Kasaija et al., 2022 |
Abbreviations: S/C, subcutaneous; I/M, intramuscular; pfu, plaque forming units.
Taken together, these results support the efficacy of SUB vaccines against different tick genera and other arthropod vector species. Additionally, other antigens such as p29, Aquaporin, Metalloprotease, Potassium ion channels, Protease inhibitors, Calreticulin, P0, Ferritin 2 and Tropomyosin have shown protection against different tick species (de la Fuente and Kocan, 2003; de la Fuente and Contreras, 2015; Manjunathachar et al., 2019; Abbas et al., 2023; Parizi et al., 2023; de la Fuente et al., 2023b; Nepveu-Traversy et al., 2024).
Challenge 4: impact of tick strain genetic diversity on vaccine efficacy
Even if tick vaccine antigens such as SUB have shown efficacy against multiple tick species, the challenge related to strain genetic diversity and other factors needs to be considered. To face this challenge, a ‘personalized medicine’ approach was proposed considering regional, tick species/strains and host factors.
An example of this approach is the SUB antigen from R. appendiculatus, R decoloratus and A. variegatum, main tick species infesting Bos indicus and crossbred cattle in Uganda (Kasaija et al., 2020). Vaccine formulations with antigens from these tick species were evaluated under controlled pen conditions in both cattle breeds to select R. appendiculatus-derived SUB as the antigen with higher cross-species protection (Kasaija et al., 2020). This vaccine is now under field trial in Uganda (Kabi et al., 2022). Other personalized SUB vaccines have been evaluated against different Indian tick species (Parthasarathi et al., 2023).
These results highlight the importance of personalizing vaccines considering tick, host and livestock farm management factors to improve effectiveness under field conditions.
Challenge 5: antigen combination to improve vaccine efficacy
Antigen combinations have been considered to improve vaccine efficacy and results of experimental trials provided support for this approach (e.g. Vitellin-degrading cysteine endopeptidase (VTDCE), Boophilus yolk pro-cathepsin (BYC) and Glutathione S-transferase (GST-Hl), Parizi et al., 2012; Bm86, SUB and Tropomyosin (TPM), Parthasarathi et al., 2023; Bm86 and P0 peptide, Rodríguez-Mallon et al., 2023) (Table 2). A comparatively higher efficacy was noted when compared with single antigen immunization. However, the main limitation of this approach is that protein-protein physical and immunological interactions may affect protective immune response in vaccinated hosts and thus additional experiments are required to eliminate the possible constraints in developing vaccine formulation using multiple antigens.
Table 2.
Examples of the efficacy of vaccination of animals with SUB combined with other tick/parasite antigens
| Antigen I | Antigen II | Antigen III | Host | Dose and delivery | Adjuvant | Efficacy | Reference |
|---|---|---|---|---|---|---|---|
|
R. microplus BM86 |
H. anatolicum SUB | H.anatolicum tropomyosin (TPM) | Cattle | Each antigen 100 μg 3 doses; I/M; Co-vaccination | Montanide ISA 50V2 | 87.2% and 86.2% against H. anatolicum larvae and adults; 86.7% against R. microplus | Parthasarathi et al., 2023 |
| R. microplus BM86 | SUB peptide of R. microplus | ---- | Cattle | 100 μg 3 doses; S/C; dual vaccine | Montanide ISA 50V2 | 49% | Mendoza-Martínez et al., 2021 |
| R. microplus SUB |
Anaplasma marginale Major surface protein-1 |
---- | Cattle | 120 μg 3 doses; S/C; chimeric vaccine | Montanide ISA 50V2 | 81% | Almazán et al. (2012) |
| R. appendiculatus SUB |
A. variegatum SUB |
R. decoloratus SUB |
Cattle | Cocktail of 100μg each protein prepared 3 doses; S/C; Cocktail vaccine |
Montanide ISA 50V2 |
R. appendiculatus 74–92; A. variegatum 51–69%; R. decoloratus 71% |
Kasaija et al., 2020 |
| R. microplus SUB | heat inactivated Mycobacterium bovis (IV) | ---- | Cattle | 200 μg of SUB mixed with 6 × 106cfu IV in 18 ml PBS for SUB + IV 2 doses; Oral |
IV acts as adjuvant | 65% | Contreras et al. (2019a, 2019b) |
| Subolesin-Akirin chimera (Q38) | ----- | Roe deer | 100 μg 3 dose I/M; chimeric vaccine | Montanide ISA 50V2 | More than 95% against I. ricinus and 46.4% Dermacentor reticulatus larvae | Contreras et al., 2020 | |
| Protective epitopes of I. scapularis SUB | Protective epitopes of A. albopictus Akirin |
||||||
| SUB-MSP1a construct | ---- | Cattle | 100 μg 2 doses; I/M; chimeric vaccine | Montanide ISA 50V2 | 60% | Almazán et al. (2012) | |
| R. microplus SUB |
Anaplasma marginale Major Surface Protein 1a (MSP1a) |
||||||
To approach this limitation, the possibility of combining SUB DNA and protein in a vaccine formulation was considered (Hassan et al., 2020). However, recent research has focused on quantum vaccinomics algorithms for the combination of antigen protective epitopes or immunological quantum (Artigas-Jerónimo et al., 2020; Contreras et al., 2022a, 2022b). As recently proposed (de la Fuente et al., 2023b), in this approach, the prediction, identification and validation of protective epitopes is based on the combination of in vitro, in silico, in music and epitope mapping approaches with systems biology integration of omics datasets, artificial intelligence (AI) and Big Data (Villar et al., 2017; de la Fuente et al., 2018; de la Fuente and Contreras, 2023).
Vaccinomics is based on the integrations of omics dataset for the identification of candidate vaccine protective antigens (Poland et al., 2013; de la Fuente and Merino, 2013; Contreras et al., 2016, 2017, 2019a). The proposal of quantum vaccinomics originated from vaccinomics and the random processes such as immunoglobulin recombination events, direct correlation between atomic coordination and peptide immunogenicity and quantum dynamics of the immune response that has been subjected to optimizing evolution within living organisms supporting quantum immunology (reviewed by de la Fuente and Contreras, 2021). Then, in reference to Albert Einstein quantum of light, immune protective epitopes were proposed as immunological quantum and quantum vaccinomics as the identification and combination of antigen immunological quantum for vaccine development (Artigas-Jerónimo et al., 2020).
Antigens such as Q38 with SUB protective epitopes (Artigas-Jerónimo et al., 2020; de la Fuente et al., 2023b) have shown protection against tick infestations and other arthropod vectors (Merino et al., 2013; Moreno-Cid et al., 2013; Contreras et al., 2020; Letinić et al., 2021) with correlation between SUB-reactive epitopes and vaccine efficacy (Contreras et al., 2022a). The chimeric antigen RmSEI composed of R. microplus Subtilisin inhibitor 7 (RmSI-7), a Trypsin inhibitory like serine protease inhibitor, an interdomain region from the Kunitz inhibitor BmTI-A, and a cysteine-rich AMP-like Microplusin (RmSEI) was designed and showed anti-tick and antimicrobial activities (Costa et al., 2023). This approach can also be used to combine tick with pathogen derived antigens (Shrivastava et al., 2020). Two multiepitopic peptides using amino acid sequences of ferritin-2 (FER2) and tropomyosin (TPM) vitellogenin receptor (VgR) were synthesized and tested against H. anatolicum infestations with more than 80% efficacy (Nandi et al., 2023) (Table 2).
Quantum vaccinomics also considers immune mechanisms mediated by protein post-translational modifications such as carbohydrate alpha-gal (Galα1-3Galβ1-4GlcNAc) present in glycoproteins (Galili, 2021) to address limitations of reductionists methods such as reverse vaccinology (Van Regenmortel, 2018; de la Fuente et al., 2023b). Accordingly, quantum vaccinomics covers some of the proposed top biotechnology trends in 2024 (https://www.startus-insights.com/innovators-guide/top-10-biotech-industry-trends-innovations-in-2021/) including AI, Big Data, gene editing, precision medicine, gene sequencing, biomanufacturing and synthetic biology.
In this way, quantum vaccinomics for protective antigen design considers vaccine efficacy and safety, geographic, environmental and population factors, host-tick-pathogen interactions and derived factors and host immunity for vaccinomics and adversomics.
Challenge 6: vaccine formulations and delivery platforms
Even when protective antigens are identified or designed, formulations and delivery are the key components of vaccine efficacy. Regarding tick control, recent advances in vaccine formulations targeting vector gut microbiota commensal bacteria was found effective (Mateos-Hernández et al., 2020, 2021). Experimental manipulation of the microbiota has been achieved by antibiotic exposure or sterile-rearing conditions of the vector. Anti-microbiota vaccine impacted tick physiology by increasing tick weight during feeding and modulated tick microbiota composition and diversity in a taxon-specific manner. The impact of anti-microbiota vaccines on pathogen development was shown in Plasmodium relictum and the mosquito vector Culex quinquefasciatus (Aželytė et al., 2022), and recently it was reported that perturbations of tick microbiota can impact highly sensitive Borrelia spp. with departure from the modulation induced by the pathogen in the vector microbiota posing a high cost to the spirochete (Wu-Chuang et al., 2021). However, these methods induce global changes in the microbiota and make the depletion of specific bacteria difficult. Recently, anti-microbiota vaccines were proposed as a precise tool for microbiota manipulation (Wu-Chuang et al., 2021; Maitre et al., 2022). Other advances including probiotics and formulations with high alpha-gal content (Cabezas-Cruz and de la Fuente, 2017; Hodžić et al., 2020; Bamgbose et al., 2021) and adjuvants with heat-inactivated alpha-gal-containing bacteria for oral vaccine administration (Contreras et al., 2019b; Kasaija et al., 2022). Oral vaccine formulations combining R. appendiculatus-derived SUB with heat-inactivated mycobacteria resulted in 96% and 99% efficacy against R. decoloratus and R. appendiculatus, respectively (Kasaija et al., 2022).
Tick vaccines have mainly been designed with recombinant antigens, but recent research includes advances in mRNA vaccines (Sajid et al., 2021; Boulanger and Wikel, 2023; Matias et al., 2023). For antigen combination, chimeric antigens on microparticles and mRNA-lipid nanoparticles may be considered for vaccine delivery (Sajid et al., 2021; Matias et al., 2023).
Challenge 7: combination of vaccines with transgenesis and paratransgenesis
Recently, Cas9-mediated gene editing was implemented in ticks by embryo injection and ReMOT Control (Sharma et al., 2022). The CRISPR-Cas molecular machines also provide interventions for paratransgenesis to manipulate tick microbiome and virome composition and function (Ramachandran and Bikard, 2019).
More recently, Frankenbacteriosis was developed for paratransgenic manipulation of tick commensal Sphingomonas bacterium to reduce tick fitness and Anaplasma phagocytophilum pathogen infection (Mazuecos et al., 2023a, 2023b; de la Fuente et al., 2023b).
Transgenesis and paratransgenesis may be combined with anti-tick vaccines and other control interventions including the proposed Suicidalbacteriosis in which tick commensal bacteria are manipulated to produce and secrete antigens protective against ticks and tick-borne pathogens to immunize hosts during blood feeding (de la Fuente et al., 2023b). For example, genetic manipulation of tick microbiome and virome composition and function may produce ticks more susceptible to tick vaccine induced host immune response thus improving vaccine efficacy for the control of tick infestations and vector capacity.
However, application of gene editing technology involves risks since it may produce off target deleterious mutations. A high frequency of off-target effects has been reported in human cells but low in mice and zebrafish (Hwang et al., 2013; Yang et al., 2013). Large genomes may contain identical or homologous DNA sequences to intended target DNA sequence. Gene editing technology may delete these unintended sequences causing mutations which may cause cell death or transformation. Efforts have been made to reduce off-target mutations, but further improvement is required. Another problem is efficient safe delivery of CRISPR-Cas9 into cell types that are hard to transfect. If there is a risk of transferring genes to other species, there is risk of transferring modified sequences. It is difficult to control dispersion of gene driven trait. Moreover, disappearance of whole populations targeted by gene drive may have serious consequences in the ecosystem equilibrium. All these risk factors demand careful assessment of each potential application and need for critical regulatory norms.
Conclusions and future directions
Tick vaccine antigens evolved from organ specific protein extracts to recombinant proteins to vaccinomics algorithms for designing chimeric antigens. Recent advances in tick vaccinology and future directions include discovery of novel protective antigens (de la Fuente and Contreras, 2015; Abbas et al., 2023) including the application of AI and Big Data analytic techniques (de la Fuente et al., 2018), novel vaccine formulations and delivery platforms (Ndawula, 2021; Tabor, 2021; Pereira et al., 2022), mRNA vaccines (Sajid et al., 2021; Matias et al., 2023; Boulanger and Wikel, 2023), vaccinomics and quantum vaccinomics (Poland et al., 2013; de la Fuente and Contreras, 2021, 2023; Contreras et al., 2022b). Other methods include use of formulations with combined protective antigens (Ndawula and Tabor, 2020; Parthasarathi et al., 2021), probiotics and other formulations targeting tick microbiota (Cabezas-Cruz and de la Fuente, 2017; Hodžić et al., 2020; Mateos-Hernández et al., 2020, 2021; Wu-Chuang et al., 2023). To improve vaccine efficacy, post-translational modifications such as alpha-gal have also been considered to improve vaccine efficacy (Hodžić et al., 2020). Moreover, characterization of tick-host-pathogen interactions, immune protective and acaricide-resistance mechanisms (Bhowmick and Han, 2020; Bishop et al., 2023; Waldman et al., 2023), transgenesis and paratransgenesis for the genetic manipulation of commensal bacteria and ticks (Sharma et al., 2022; Mazuecos et al. 2023a; de la Fuente et al., 2023b) and combination of vaccines with other interventions such as natural plant and animal-derived compounds and cultural practices among other interventions (Showler and Saelao, 2022) were considered as possible alternatives. International collaborations with regions with high incidence of tick infestations and TBD (Estrada-Peña and de la Fuente, 2023), personalized medicine approach based on regional, tick species/strains and host-driven variables (Kasaija et al., 2020) are also proposed for sustainable management of the relevant vector.
Acknowledgements
We thank all our collaborators worldwide for their contribution to advance in tick vaccinology.
Data availability statement
All data used in the study is disclosed in the paper and corresponding references.
Authors’ contributions
JF and SG conceived and wrote the article.
Financial support
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Competing interests
None.
Ethical standards
Not applicable.
References
- Abbas MN, Jmel MA, Mekki I, Dijkgraaf I and Kotsyfakis M (2023) Recent advances in tick antigen discovery and anti-tick vaccine development. International Journal of Molecular Sciences 24, 4969. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Agwunobi DO, Yu Z and Liu J (2021) A retrospective review on ixodid tick resistance against synthetic acaricides: implications and perspectives for future resistance prevention and mitigation. Pesticide Biochemistry and Physiology 173, 104776. [DOI] [PubMed] [Google Scholar]
- Allen JR and Humphreys SJ (1979) Immunisation of guinea pigs and cattle against ticks. Nature 280, 491–493. [DOI] [PubMed] [Google Scholar]
- Almazán C, Kocan KM, Bergman DK, Garcia-Garcia JC, Blouin EF and de la Fuente J (2003) Identification of protective antigens for the control of Ixodes scapularis infestations using cDNA expression library immunization. Vaccine 21, 1492–1501. [DOI] [PubMed] [Google Scholar]
- Almazán C, Blas-Machado U, Kocan KM, Yoshioka JH, Blouin EF, Mangold AJ and de la Fuente J (2005) Characterization of three Ixodes scapularis cDNAs protective against tick infestations. Vaccine 23, 4403–4416. [DOI] [PubMed] [Google Scholar]
- Almazán C, Lagunes R, Villar M, Canales M, Rosario-Cruz R, Jongejan F and de la Fuente J (2010) Identification and characterization of Rhipicephalus (Boophilus) microplus candidate protective antigens for the control of cattle tick infestations. Parasitology Research 106, 471–479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almazán C, Moreno-Cantú O, Moreno-Cid JA, Galindo RC, Canales M, Villar M and de la Fuente J (2012) Control of tick infestations in cattle vaccinated with bacterial membranes containing surface-exposed tick protective antigens. Vaccine 30, 265–272. [DOI] [PubMed] [Google Scholar]
- Andreano E, D'Oro U, Rappuoli R and Finco O (2019) Vaccine evolution and its application to fight modern threats. Frontiers in Immunology 10, 1722. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Artigas-Jerónimo S, Villar M, Cabezas-Cruz A, Valdés JJ, Estrada-Peña A, Alberdi P and de la Fuente J (2018) Functional evolution of subolesin/akirin. Frontiers in Physiology 9, 1612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Artigas-Jerónimo S, Comín JJP, Villar M, Contreras M, Alberdi P, Viera IL, Soto L, Cordero R, Valdés JJ, Cabezas-Cruz A, Estrada-Peña A and de la Fuente J (2020) A novel combined scientific and artistic approach for the advanced characterization of interactomes: the akirin/subolesin model. Vaccines 8, 77. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aželytė J, Wu-Chuang A, Žiegytė R, Platonova E, Mateos-Hernandez L, Maye J, Obregon D, Palinauskas V and Cabezas-Cruz A (2022) Anti-microbiota vaccine reduces avian malaria infection within mosquito vectors. Frontiers in Immunology 13, 841835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bamgbose T, Anvikar AR, Alberdi P, Abdullahi IO, Inabo HI, Bello M, Cabezas-Cruz A and de la Fuente J (2021) Functional food for the stimulation of the immune system against malaria. Probiotics and Antimicrobial Proteins 13, 1254–1266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bensaci M, Bhattacharya D, Clark R and Hu LT (2012) Oral vaccination with vaccinia virus expressing the tick antigen subolesin inhibits tick feeding and transmission of Borrelia burgdorferi. Vaccine 30, 6040–6046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhowmick B and Han Q (2020) Understanding tick biology and its implications in anti-tick and transmission blocking vaccines against tick-borne pathogens. Frontiers in Veterinary Science 7, 319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bishop LJ, Stutzer C and Maritz-Olivier C (2023) More than three decades of Bm86: what we know and where to go. Pathogens (Basel, Switzerland) 12, 1071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boulanger N and Wikel S (2023). Vaccines against tick-borne diseases: a big step forward?. Trends in Parasitology 39, 989–990. [DOI] [PubMed] [Google Scholar]
- Cabezas-Cruz A and de la Fuente J (2017). Immunity to α-Gal: toward a single-antigen pan-vaccine to control major infectious diseases. ACS Central Science 3, 1140–1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carreón D, de la Lastra JM, Almazán C, Canales M, Ruiz-Fons F, Boadella M, Moreno-Cid JA, Villar M, Gortázar C, Reglero M, Villarreal R and de la Fuente J (2012) Vaccination with BM86, subolesin and akirin protective antigens for the control of tick infestations in white tailed deer and red deer. Vaccine 30, 273–279. [DOI] [PubMed] [Google Scholar]
- Chakraborty S, Gao S, Allan BF and Smith RL (2023). Effects of cattle on vector-borne disease risk to humans: a systematic review. PLoS Neglected Tropical Diseases 17, e0011152. Advance online publication. 10.1371/journal.pntd.0011152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contreras M, Villar M, Alberdi P and de la Fuente J (2016) Vaccinomics approach to tick vaccine development. Methods in Molecular Biology (Clifton, N.J.) 1404, 275–286. [DOI] [PubMed] [Google Scholar]
- Contreras M, Alberdi P, Fernández De Mera IG, Krull C, Nijhof A, Villar M and de La Fuente J (2017). Vaccinomics approach to the identification of candidate protective antigens for the control of tick vector infestations and Anaplasma phagocytophilum infection. Frontiers in Cellular and Infection Microbiology 7, 360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contreras M, Villar M and de la Fuente J (2019a) A vaccinomics approach for the identification of tick protective antigens for the control of Ixodes ricinus and Dermacentor reticulatus infestations in companion animals. Frontiers in Physiology 10, 977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contreras M, Kasaija PD, Merino O, de la Cruz-Hernandez NI, Gortazar C and de la Fuente J (2019b) Oral vaccination with a formulation combining Rhipicephalus microplus subolesin with heat inactivated Mycobacterium bovis reduces tick infestations in cattle. Frontiers in Cellular and Infection Microbiology 9, 45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contreras M, San José C, Estrada-Peña A, Talavera V, Rayas E, Isabel León C, Luis Núñez J, García Fernández de Mera I and de la Fuente J (2020) Control of tick infestations in wild roe deer (Capreolus capreolus) vaccinated with the Q38 subolesin/akirin chimera. Vaccine 38, 6450–6454. [DOI] [PubMed] [Google Scholar]
- Contreras M, Kasaija PD, Kabi F, Mugerwa S and de la Fuente J (2022a) The correlation between subolesin-reactive epitopes and vaccine efficacy. Vaccines 10, 1327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Contreras M, Artigas-Jerónimo S, Pastor Comín JJ and de la Fuente J (2022b) A quantum vaccinomics approach based on protein-protein interactions. Methods in Molecular Biology 2411, 287–305. [DOI] [PubMed] [Google Scholar]
- Costa GCA, Silva FAA, Manzato VM, Torquato RJS, Gonzalez YG, Parizi LF, da Silva Vaz Junior I and Tanaka AS (2023) A multiepitope chimeric antigen from Rhipicephalus microplus-secreted salivary proteins elicits anti-tick protective antibodies in rabbit. Veterinary Parasitology. 318, 109932. [DOI] [PubMed] [Google Scholar]
- de la Fuente J (2018) Controlling ticks and tick-borne diseases…looking forward. Ticks and Tick-Borne Diseases 9, 1354–1357. [DOI] [PubMed] [Google Scholar]
- de la Fuente J and Contreras M (2015) Tick vaccines: current status and future directions. Expert Review of Vaccines 14, 1367–1376. [DOI] [PubMed] [Google Scholar]
- de la Fuente J and Contreras M (2021) Vaccinomics: a future avenue for vaccine development against emerging pathogens. Expert Review of Vaccines 20, 1561–1569. [DOI] [PubMed] [Google Scholar]
- de la Fuente J and Contreras M (2023) Quantum vaccinomics platforms to advance in vaccinology. Frontiers in Immunology 14, 1172734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Fuente J and Kocan KM (2003) Advances in the identification and characterization of protective antigens for recombinant vaccines against tick infestations. Expert Review of Vaccines 2, 583–593. [DOI] [PubMed] [Google Scholar]
- de la Fuente J and Kocan KM (2014) Development of vaccines for control of tick infestations and interruption of pathogen transmission. In Sonenshine D, Roe M (Eds), Biology of Ticks, 2nd Edn. New York: Oxford University Press, pp. 333–352. [Google Scholar]
- de la Fuente J and Merino O (2013) Vaccinomics, the new road to tick vaccines. Vaccine 31, 5923–5929. [DOI] [PubMed] [Google Scholar]
- de la Fuente J, Almazán C, Canales M, Pérez de la Lastra JM, Kocan KM and Willadsen P (2007) A ten-year review of commercial vaccine performance for control of tick infestations on cattle. Animal Health Research Reviews 8, 23–28. [DOI] [PubMed] [Google Scholar]
- de la Fuente J, Manzano-Roman R, Naranjo V, Kocan KM, Zivkovic Z, Blouin EF, Canales M, Almazán C, Galindo RC, Step DL and Villar M (2010) Identification of protective antigens by RNA interference for control of the lone star tick, Amblyomma americanum. Vaccine 28, 1786–1195. [DOI] [PubMed] [Google Scholar]
- de la Fuente J, Moreno-Cid JA, Canales M, Villar M, de la Lastra JM, Kocan KM, Galindo RC, Almazán C and Blouin EF (2011) Targeting arthropod subolesin/akirin for the development of a universal vaccine for control of vector infestations and pathogen transmission. Veterinary Parasitology 181, 17–22. [DOI] [PubMed] [Google Scholar]
- de la Fuente J, Villar M, Estrada-Peña A and Olivas JA (2018) High throughput discovery and characterization of tick and pathogen vaccine protective antigens using vaccinomics with intelligent big data analytic techniques. Expert Review of Vaccines 17, 569–576. [DOI] [PubMed] [Google Scholar]
- de la Fuente J, Artigas-Jerónimo S and Villar M (2021) Akirin/subolesin regulatory mechanisms at host/tick-pathogen interactions. microLife 3, uqab012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Fuente J, Estrada-Peña A, Rafael M, Almazán C, Bermúdez S, Abdelbaset AE, Kasaija PD, Kabi F, Akande FA, Ajagbe DO, Bamgbose T, Ghosh S, Palavesam A, Hamid PH, Oskam CL, Egan SL, Duarte-Barbosa A, Hekimoğlu O, Szabó MPJ, Labruna MB and Dahal A (2023a) Perception of ticks and tick-borne diseases worldwide. Pathogens (Basel, Switzerland) 12, 1258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de la Fuente J, Mazuecos L and Contreras M (2023b) Innovative approaches for the control of ticks and tick-borne diseases. Ticks and Tick-Borne Diseases 14, 102227. [DOI] [PubMed] [Google Scholar]
- Estrada-Peña A and de la Fuente J (2023) Evolution of tick vaccinology highlights changes in paradigms in this research area. Vaccines,11, 253. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Estrada-Peña A, Mallón AR, Bermúdez S, de la Fuente J, Domingos A, García MPE, Labruna MB, Merino O, Mosqueda J, Nava S, Cruz RL, Szabó M, Tarragona E and Venzal JM (2022) One health approach to identify research needs on Rhipicephalus microplus ticks in the Americas. Pathogens (Basel, Switzerland) 11, 1180. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galili U (2021) Increasing efficacy of enveloped whole-virus vaccines by in situ immune-complexing with the natural anti-gal antibody. Medical Research Archives 9, 2481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Githaka NW, Kanduma EG, Wieland B, Darghouth MA and Bishop RP (2022) Acaricide resistance in livestock ticks infesting cattle in Africa: current status and potential mitigation strategies. Current Research in Parasitology and Vector-Borne Diseases 2, 100090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gonzaga BCF, Barrozo MM, Coutinho AL, Pereira E Sousa LJM, Vale FL, Marreto L, Marchesini P, de Castro Rodrigues D, de Souza EDF, Sabatini GA, Costa-Júnior LM, Ferreira LL, Lopes WDZ and Monteiro C (2023) Essential oils and isolated compounds for tick control: advances beyond the laboratory. Parasites and Vectors, 16, 415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harrington D, Canales M, de la Fuente J, de Luna C, Robinson K, Guy J and Sparagano O (2009) Immunisation with recombinant proteins subolesin and Bm86 for the control of Dermanyssus gallinae in poultry. Vaccine 27, 4056–4063. [DOI] [PubMed] [Google Scholar]
- Hassan IA, Wang Y, Zhou Y, Cao J, Zhang H and Zhou J (2020) Cross protection induced by combined Subolesin-based DNA and protein immunizations against adult Haemaphysalis longicornis. Vaccine 38, 907–915. [DOI] [PubMed] [Google Scholar]
- Hodžić A, Mateos-Hernández L, de la Fuente J and Cabezas-Cruz A (2020) α-Gal-based vaccines: advances, opportunities, and perspectives. Trends in Parasitology 36, 992–1001. [DOI] [PubMed] [Google Scholar]
- Hwang WY, Fu Y, Reyon D, Maeder ML, Tsai SQ, Sander JD, Peterson RT, Yeh JR and Joung JK (2013) Efficient genome editing in zebrafish using a CRISPR-Cas system. Nature Biotechnology 31, 227–229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kabi F, Dhikusooka M, Matovu M, Mugerwa S, Kasaija P, Emudong P, Kirunda H, Contreras M, Gortazar C and de la Fuente J (2022) Monitoring the subolesin vaccine field trial for safer control of cattle ticks amidst increasing acaricide resistance in Uganda. Vaccines 10, 1594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasaija PD, Contreras M, Kabi F, Mugerwa S and de la Fuente J (2020) Vaccination with recombinant subolesin antigens provides cross-tick species protection in Bos indicus and crossbred cattle in Uganda. Vaccines 8, 319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasaija PD, Contreras M, Kabi F, Mugerwa S, Garrido JM, Gortazar C and de la Fuente J (2022) Oral vaccine formulation combining tick subolesin with heat inactivated mycobacteria provides control of cross-species cattle tick infestations. Vaccine 40, 4564–4573. [DOI] [PubMed] [Google Scholar]
- Lee SH, Li J, Moumouni PFA, Okado K, Zheng W, Liu M, Ji S, Kim S, Umemiya-Shirafuji R and Xuan X (2020) Subolesin vaccination inhibits blood feeding and reproduction of Haemaphysalis longicornis in rabbits. Parasites & Vectors 13, 478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Letinić BD, Contreras M, Dahan-Moss Y, Linnekugel I, de la Fuente J and Koekemoer LL (2021) Additional evidence on the efficacy of different akirin vaccines assessed on Anopheles arabiensis (Diptera: Culicidae). Parasites & Vectors 14, 209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maitre A, Wu-Chuang A, Aželytė J, Palinauskas V, Mateos-Hernández L, Obregon D, Hodžić A, Valiente Moro C, Estrada-Peña A, Paoli JC, Falchi A and Cabezas-Cruz A (2022) Vector microbiota manipulation by host antibodies: the forgotten strategy to develop transmission-blocking vaccines. Parasites & Vectors 15, 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manjunathachar HV, Kumar B, Saravanan BC, Choudhary S, Mohanty AK, Nagar G, Chigure G, Ravi Kumar GVPPS, de la Fuente J and Ghosh S (2019) Identification and characterization of vaccine candidates against Hyalomma anatolicum-Vector of Crimean-Congo haemorrhagic fever virus. Transboundary and Emerging Diseases 66, 422–434. [DOI] [PubMed] [Google Scholar]
- Mateos-Hernández L, Obregón D, Maye J, Borneres J, Versille N, de la Fuente J, Estrada-Peña A, Hodžić A, Šimo L and Cabezas-Cruz A (2020) Anti-tick microbiota vaccine impacts Ixodes ricinus performance during feeding. Vaccines 8, 702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mateos-Hernández L, Obregón D, Wu-Chuang A, Maye J, Bornères J, Versillé N, de la Fuente J, Díaz-Sánchez S, Bermúdez-Humarán LG, Torres-Maravilla E, Estrada-Peña A, Hodžić A, Šimo L and Cabezas-Cruz A (2021) Anti-microbiota vaccines modulate the tick microbiome in a taxon-specific manner. Frontiers in Immunology 12, 704621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matias J, Cui Y, Tang X, Sajid A, Arora G, Wu MJ, DePonte K, Muramatsu H, Tam YK, Narasimhan S, Pardi N, Weissman D and Fikrig E (2023) Specific mRNA lipid nanoparticles and acquired resistance to ticks. Vaccine 41, 4996–5002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazuecos L, Alberdi P, Hernández-Jarguín A, Contreras M, Villar M, Cabezas-Cruz A, Simo L, González-García A, Díaz-Sánchez S, Neelakanta G, Bonnet SI, Fikrig E and de la Fuente J (2023a) Frankenbacteriosis targeting interactions between pathogen and symbiont to control infection in the tick vector. iScience 26, 106697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazuecos L, González-García A and de la Fuente J (2023b) Genetic modification, characterization, and co-infection of Franken Sphingomonas and Anaplasma phagocytophilum in tick cells. STAR Protocols 4, 102557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mendoza-Martínez N, Alonso-Díaz MA, Merino O, Fernández-Salas A and Lagunes-Quintanilla R (2021) Protective efficacy of the peptide subolesin antigen against the cattle tick Rhipicephalus microplus under natural infestation. Veterinary Parasitology 299, 109577. [DOI] [PubMed] [Google Scholar]
- Merino O, Almazán C, Canales M, Villar M, Moreno-Cid JA, Estrada-Peña A, Kocan KM and de la Fuente J (2011) Control of Rhipicephalus (Boophilus) microplus infestations by the combination of subolesin vaccination and tick autocidal control after subolesin gene knockdown in ticks fed on cattle. Vaccine 29, 2248–2254. [DOI] [PubMed] [Google Scholar]
- Merino O, Antunes S, Mosqueda J, Moreno-Cid JA, Pérez de la Lastra JM, Rosario-Cruz R, Rodríguez S, Domingos A and de la Fuente J (2013) Vaccination with proteins involved in tick-pathogen interactions reduces vector infestations and pathogen infection. Vaccine 31, 5889–5896. [DOI] [PubMed] [Google Scholar]
- Nandi A, Manisha Solanki V, Tiwari V, Sajjanar B, Sankar M, Saini M, Shrivastava S, Bhure SK and Ghosh S (2023) Protective efficacy of multiple epitope-based vaccine against Hyalomma anatolicum, vector of Theileria annulata and Crimean-Congo Hemorrhagic Fever Virus. Vaccines 11, 881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ndawula C Jr (2021) From bench to field: a guide to formulating and evaluating anti-tick vaccines delving beyond efficacy to effectiveness. Vaccines 9, 1185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ndawula C Jr and Tabor AE (2020) Cocktail anti-tick vaccines: the unforeseen constraints and approaches toward enhanced efficacies. Vaccines 8, 457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nepveu-Traversy ME, Fausther-Bovendo H and Babuadze GG (2024) Human tick-borne diseases and advances in anti-tick vaccine approaches: a comprehensive review. Vaccines 12, 141. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parizi LF, Reck, J Jr., Oldiges DP, Guizzo MG, Seixas A, Logullo C, de Oliveira PL, Termignoni C, Martins JR and Vaz Ida S Jr. (2012) Multi-antigenic vaccine against the cattle tick Rhipicephalus (Boophilus) microplus: a field evaluation. Vaccine 30, 6912–6917. [DOI] [PubMed] [Google Scholar]
- Parizi LF, Githaka NW, Logullo C, Zhou J, Onuma M, Termignoni C and da Silva Vaz I Jr (2023). Universal tick vaccines: candidates and remaining challenges. Animals: an Open Access Journal from MDPI 13, 2031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parthasarathi BC, Kumar B and Ghosh S (2021) Current status and future prospects of multi-antigen tick vaccine. Journal of Vector Borne Diseases 58, 183–192. [DOI] [PubMed] [Google Scholar]
- Parthasarathi BC, Kumar B, Bhure SK, Sharma AK, Manisha, Nagar G, Kumar, S, Nandi, A, Manjunathachar, HV, Chigure, GM, Shakya, M, Sankar, M, Fuente, J and Ghosh S (2023) Co-immunization efficacy of recombinant antigens against Rhipicephalus microplus and Hyalomma anatolicum tick infestations. Pathogens (Basel, Switzerland) 12, 433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pereira DFS, Ribeiro HS, Gonçalves AAM, da Silva AV, Lair DF, de Oliveira DS, Boas DFV, Conrado IDSS, Leite JC, Barata LM, Reis PCC, Mariano RMDS, Santos TAP, Coutinho DCO, Gontijo NF, Araujo RN, Galdino AS, Paes PRO, Melo MM, Nagem RAP, Dutra WO, Silveira-Lemos DD, Rodrigues DS and Giunchetti RC (2022) Rhipicephalus microplus: an overview of vaccine antigens against the cattle tick. Ticks and Tick-Borne Diseases 13, 101828. [DOI] [PubMed] [Google Scholar]
- Poland GA, Kennedy RB, McKinney BA, Ovsyannikova IG, Lambert ND, Jacobson RM and Oberg AL (2013) Vaccinomics, adversomics, and the immune response network theory: individualized vaccinology in the 21st century. Seminars in Immunology 25, 89–103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramachandran G and Bikard D (2019) Editing the microbiome the CRISPR way. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences 374, 20180103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodríguez-Mallon A (2023). The Bm86 discovery: a revolution in the development of anti-tick vaccines. Pathogens (Basel, Switzerland) 12, 231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodríguez-Mallon A, Encinosa Guzmán PE, Bello Y, Domingos A, Antunes S, Kopacek P, Santos AS, Velez R, Perner J, Ledesma Bravo FL, Frantova H, Erhart J, Rodríguez R, Fuentes A, Diago D, Joglar M, Méndez L and Estrada MP (2023) Efficacy of the vaccine candidate based on the P0 peptide against Dermacentor nitens and Ixodes ricinus ticks. Pathogens (Basel, Switzerland) 12, 1365. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodríguez M, Rubiera R, Penichet M, Montesinos R, Cremata J, Falcón V, Sánchez G, Bringas R, Cordovés C, Valdés M, Lleonart R, Herrera L and de la Fuente J (1994) High level expression of the B. microplus Bm86 antigen in the yeast Pichia pastoris forming highly immunogenic particles for cattle. Journal of Biotechnology 33, 135–146. [DOI] [PubMed] [Google Scholar]
- Sajid A, Matias J, Arora G, Kurokawa C, DePonte K, Tang X, Lynn G, Wu MJ, Pal U, Strank NO, Pardi N, Narasimhan S, Weissman D and Fikrig E (2021) mRNA vaccination induces tick resistance and prevents transmission of the Lyme disease agent. Science Translational Medicine 13, eabj9827. [DOI] [PubMed] [Google Scholar]
- Shakya M, Kumar B, Nagar G, de la Fuente J and Ghosh S (2014). Subolesin: a candidate vaccine antigen for the control of cattle tick infestations in Indian situation. Vaccine 32, 3488–3494. [DOI] [PubMed] [Google Scholar]
- Sharma A, Pham MN, Reyes JB, Chana R, Yim WC, Heu CC, Kim D, Chaverra-Rodriguez D, Rasgon JL, Harrell RA, 2nd, Nuss AB and Gulia-Nuss M (2022) Cas9-mediated gene editing in the black-legged tick, Ixodes scapularis, by embryo injection and ReMOT control. iScience, 25, 103781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Showler AT and Saelao P (2022). Integrative alternative tactics for Ixodid control. Insects 13, 302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shrivastava N, Verma A and Dash PK (2020) Identification of functional epitopes of structural proteins and in-silico designing of dual acting multiepitope anti-tick vaccine against emerging Crimean-Congo hemorrhagic fever virus. European Journal of Pharmaceutical Sciences: Official Journal of the European Federation for Pharmaceutical Sciences 151, 105396. [DOI] [PubMed] [Google Scholar]
- Moreno-Cid JA, Pérez de la Lastra JM, Villar M, Jiménez M, Pinal R, Estrada-Peña A, Molina R, Lucientes J, Gortázar C and de la Fuente J and SUB/AKR Vaccine Study Group (2013) Control of multiple arthropod vector infestations with subolesin/akirin vaccines. Vaccine 31, 1187–1196. [DOI] [PubMed] [Google Scholar]
- Tabor AE (2021). A review of Australian tick vaccine research. Vaccines 9, 1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torina A, Moreno-Cid JA, Blanda V, Fernández de Mera IG, de la Lastra JM, Scimeca S, Blanda M, Scariano ME, Briganò S, Disclafani R, Piazza A, Vicente J, Gortázar C, Caracappa S, Lelli RC and de la Fuente J (2014) Control of tick infestations and pathogen prevalence in cattle and sheep farms vaccinated with the recombinant subolesin-major surface protein 1a chimeric antigen. Parasites & Vectors 7, 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Regenmortel MHV (2018) Development of a preventive HIV vaccine requires solving inverse problems which is unattainable by rational vaccine design. Frontiers in Immunology 8, 2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Villar M, Marina A and de la Fuente J (2017) Applying proteomics to tick vaccine development: where are we? Expert Review of Proteomics 14, 211–221. [DOI] [PubMed] [Google Scholar]
- Waldman J, Klafke GM, Tirloni L, Logullo C and da Silva Vaz I Jr (2023) Putative target sites in synganglion for novel ixodid tick control strategies. Ticks and Tick-Borne Diseases 14, 102123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Willadsen P and Kemp DH (1988). Vaccination with ‘concealed’ antigens for tick control. Parasitology Today 4, 196–198. [DOI] [PubMed] [Google Scholar]
- Willadsen P, McKenna RV and Riding GA (1988) Isolation from the cattle tick, Boophilus microplus, of antigenic material capable of eliciting a protective immunological response in the bovine host. International Journal for Parasitology 18, 183–189. [DOI] [PubMed] [Google Scholar]
- Willadsen P, Bird P, Cobon GS and Hungerford J (1995). Commercialisation of a recombinant vaccine against Boophilus microplus. Parasitology 110 Suppl, S43–S50. [DOI] [PubMed] [Google Scholar]
- Wu-Chuang A, Hodžić A, Mateos-Hernández L, Estrada-Peña A, Obregon D and Cabezas-Cruz A (2021) Current debates and advances in tick microbiome research. Current Research in Parasitology & Vector-Borne Diseases 1, 100036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu-Chuang A, Mateos-Hernandez L, Maitre A, Rego ROM, Šíma R, Porcelli S, Rakotobe S, Foucault-Simonin A, Moutailler S, Palinauskas V, Aželytė J, Sǐmo L, Obregon D and Cabezas-Cruz A (2023) Microbiota perturbation by anti-microbiota vaccine reduces the colonization of Borrelia afzelii in Ixodes ricinus. Microbiome 11, 151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang H, Wang H, Shivalila CS, Cheng AW, Shi L and Jaenisch R (2013) One-step generation of mice carrying reporter and conditional alleles by CRISPR/Cas-mediated genome engineering. Cell 154, 1370–1379. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data used in the study is disclosed in the paper and corresponding references.