

Abstract
目的
探讨FEZF1-AS1通过miR-130a-5p/CCND1轴促进非小细胞肺癌(NSCLC)发展的分子机制。
方法
利用TCGA数据库分析FEZF1-AS1在NSCLC中的表达,qRT-PCR检测其在NSCLC癌组织与癌旁组织以及NSCLC细胞系中的表达,并分析其与临床特征的关系。利用数据库预测FEZF1-AS1与hsa-miR-130a-5p的结合位点以及hsa-miR-130a-5p与CCND1的结合位点;应用CCK8、克隆形成实验、划痕实验、Transwell实验检测FEZF1-AS1、hsa-miR-130a-5p对肺癌细胞系增殖、侵袭、迁移能力的影响;双荧光素酶报告实验验证FEZF1-AS1与hsa-miR-130a-5p以及hsa-miR-130a-5p与CCND1存在结合;将H1299、H358分别分为si-NC组、si-FEZF1-AS1组、si-FEZF1-AS1+NC inhibitor组、si-FEZF1-AS1+hsa-miR-130a-5p inhibitor组,应用Western blot检测CCND1的蛋白表达水平,明确FEZF1-AS1/hsa-miR-130a-5p是否通过ceRNA机制调控了CCND1的表达。
结果
FEZF1-AS1在NSCLC中呈高表达,且在NSCLC癌组织中的表达量明显增高(P<0.05),高表达与NSCLC的淋巴结转移有关,在H1299、H358细胞系中的表达量也显著增高(P<0.05);敲减FEZF1-AS1降低了NSCLC细胞的增殖、迁移、侵袭能力(P<0.05);hsa-miR-130a-5p与FEZF1-AS1、CCND1之间存在结合(P<0.05);hsa-miR-130a-5p影响NSCLC细胞的增殖、迁移、侵袭能力(P<0.05);si-FEZF1-AS1降低了CCND1蛋白表达,hsa-miR-130a-5p inhibitor逆转了si-FEZF1-AS对CCND1的敲减作用(P<0.05)。
结论
FEZF1-AS1在NSCLC组织中高表达,且与淋巴结转移有关,促进了NSCLC细胞的增殖、迁移、侵袭,并通过miR-130a-5p/CCND1轴促进了非小细胞肺癌的发展。
Keywords: 非小细胞肺癌, 长链非编码RNA, FEZF1-AS1, miR-130a-5p, CCND1
Abstract
Objective
To explore the molecular mechanism by which FEZF1-AS1 overexpression promotes progression of non-small cell lung cancer (NSCLC) via the miR-130a-5p/CCND1 axis.
Methods
TCGA database was used to analyze FEZF1-AS1 expression levels in NSCLC. FEZF1-AS1 expression was detected by qRT-PCR in clinical specimens of NSCLC tissues and NSCLC cell lines, and its correlation with clinical features of the patients were analyzed. The binding sites of FEZF1-AS1 with hsa-miR-130a-5p and those of hsa-miR-130a-5p with CCND1 were predicted. CCK8 assay, clone formation assay, scratch assay, and Transwell assay were employed to examine the effects of FEZF1-AS1 knockdown and hsa-miR-130a-5p inhibitor on proliferation, invasion, and migration abilities of lung cancer cell lines. Dual luciferase assay was used to verify the binding of FEZF1-AS1 with hsa-miR-130a-5p and the binding of hsa-miR-130a-5p with CCND1. Western blotting was performed to detect the changes in CCND1 protein expression level in H1299 and H358 cells following FEZF1-AS1 knockdown and treatment with hsa-miR-130a-5p inhibitor.
Results
FEZF1-AS1 was highly expressed in NSCLC tissues in close correlation with lymph node metastasis and also in H1299 and H358 cell lines (all P<0.05). FEZF1-AS1 knockdown obviously reduced proliferation, migration, and invasion abilities of NSCLC cells (P<0.05). Dual luciferase assay confirmed the binding of hsa-miR-130a-5p with FEZF1-AS1 and CCND1 (P<0.05), and hsa-miR-130a-5p inhibitor significantly inhibited proliferation, migration, and invasion of NSCLC cells (P<0.05). FEZF1-AS1 knockdown significantly reduced CCND1 protein expression in NSCLC cells, and this effect was strongly inhibited by treatment with hsa-miR-130a-5p inhibitor (P<0.05).
Conclusion
FEZF1-AS1 is highly expressed in NSCLC tissue in close correlation with lymph node metastasis to promote cancer progression through the miR-130a-5p/CCND1 axis.
Keywords: non-small cell lung cancer, lncRNA, FEZF1-AS1, miR-130a-5p, CCND1
肺癌是发病率和死亡率最高的恶性肿瘤,根据国际癌症研究机构2020年发布的全球癌症统计数据显示,中国当年新增肺癌患者约82万人,死亡约71.5万人。只有20%的患者经治疗后能存活过5年,不到7%的患者存活超过10年[1,2]。较差的预后可能是由于对肺癌的发生发展和耐药机制仍不十分清楚。
近年来研究认为非编码RNA(ncRNA)在肿瘤发生发展中起着重要作用并参与耐药[3]。ncRNA不编码蛋白质序列,其中长度超过200nt的ncRNA被称为长链非编码RNA(LncRNA)。LncRNA具有许多功能,如染色质重塑、竞争性内源性RNA(ceRNA)和自然反义转录物等[4],在肿瘤发生的遗传调控、细胞周期、细胞分化、凋亡调控以及生长转移等也发挥重要作用[5]。相关研究还表明LncRNA在对肿瘤病人诊断、靶向治疗耐药及预后判断也极为重要[6-8]。在非小细胞肺癌(NSCLC)中也有研究报道LncRNA可以通过海绵化、PI3K/AKT通路等在其发展中发挥重要作用,并参与表皮细胞生长因子受体酪氨酸激酶酶抑制剂(EGFR-TKIs)的耐药[9,10]。
FEZ家族锌指1反义RNA1(FEZF1-AS1)是一种新的lncRNA,位于7号染色体上,最早由日本科学家通过对21243个全长cDNA测序中发现。FEZF1-AS1在许多癌症中被报道,如结直肠癌[11]、宫颈癌[12]等。周丽亚[13]研究发现FEZF1-AS1在EGFR突变型中晚期肺腺癌表达的阳性率为55.56%(30/54),且表达阳性的患者EGFR-TKI的耐药的发生率高,预后较差。和其他LncRNA一样,FEZF1-AS1也可以通过信号通路和生物过程影响NSCLC细胞的生长、转移和耐药。但目前关于FEZF1-AS1对NSCLC发生的调控机制尚不明确。
为进一步探究FEZF1-AS1在NSCLC中的表达情况,是否通过ceRNA机制调控非小细胞肺癌的增殖、侵袭、迁移等功能促进NSCLC发展,本研究旨在利用生物信息学技术及相关细胞功能学实验,研究FEZF1-AS1在肺癌组织、细胞中的表达水平及影响肺癌细胞生物学功能的具体通路,为FEZF1-AS1将来是否能作为肺癌筛查、诊断及治疗中新的靶点提供依据,也为NSCLC的发生机制和耐药的解决提供新的见解。
1. 材料和方法
1.1. 病理组织标本
收集蚌埠医学院第一附属医院胸外科2020年9月~2022年4月的33例首次确诊的NSCLC患者的手术标本(包括癌组织及癌旁组织),留取的组织标本液氮储存备用。所有肺癌的诊断均经两名病理科医师确诊,收集标本及临床资料均经患者及家属知情同意,且取得蚌埠医学院第一附属医院伦理会审批同意(审批号:BYYFY-2018KY25)。
1.2. 细胞系
非小细胞肺癌细胞系(NCI-H1299、NCI-H358),肺腺癌细胞系(A549、PC-9、NCI-H1975),人支气管上皮细胞系(BEAS-2B)(广州赛库生物技术公司)。所有实验均在蚌埠医科大学第一医院呼吸与危重症学科实验室、呼吸系病临床基础安徽省重点实验室完成。
1.3. 引物设计
FEZF1-AS1、GAPDH引物由上海生工合成,FEZF1-AS1引物序列:F:5'-TTAGGAGGCTTGTTCTGTGT-3',R:5'-GCGCAGGTACTTAAGAAAGA-3';GAPDH引物序列:F:5'-ACAGTCAGCCGCATCTTCT-3',R:5'-GACA AGCTTCCCGTTCTCAG-3'。小干扰RNA及其阴性对照由上海吉玛基因提供,hsa-miR-130a-5p、其余miRNA以及相关阴性对照引物由上海新贝提供,hsa-miR-130a-5p mimics、hsa-miR-130a-5p inhibitor及其阴性对照引物由上海汉恒提供。
1.4. 实验方法
1.4.1. qRT-PCR检测FEZF1-AS1与hsa-miR-130a-5p表达
Trizol(life technologies)法提取NSCLC癌组织及癌旁组织和细胞系中总RNA,逆转录使用逆转录试剂盒(ABclonal),按说明书进行,加入引物,后按qPCR MIX试剂盒(ABclonal)说明书步骤进行扩增,PCR参数按说明书进行设置。扩增后FEZF1-AS1与hsa-miR-130a-5p表达量应用2-ΔΔCt法进行分析。
1.4.2. 细胞培养与传代
于37 ℃ 5%CO2培养箱环境中,应用1640细胞培养基、DMEM细胞培养基(Gibco),进行细胞培养,密度70%~80%后,应用胰酶(Gibco)收取培养细胞进行实验。
1.4.3. 肺癌细胞系进行FEZF1-AS1与hsa-miR-130a-5p转染
将肺癌细胞系在10 cm培养皿中培养至密度70%以上,于6孔板进行铺板,按照siRNA(上海吉玛基因)、hsa-miR-130a-5p inhibitor、hsa-miR-130a-5p mimics(上海汉恒生物)说明书,取2.5 µL Lipofecta mine 2000(Thermo Fisher Scientific)、2.5 µL siRNA、hsa-miR-130a-5p mimics或hsa-miR-130a-5p inhibitor或hsa-miR-130a-5p mimics与Lipofecta mine 2000以及 125 µLOpti-MEM无血清培养基(Gibco)进行转染。
1.4.4. CCK-8细胞增殖实验
将转染后的细胞重悬为20 000 mL的细胞悬液种于96孔板内,放入培养箱,在指定时间将孔板内培养基吸出,将CCK-8试剂(Biosharp)与培养基按比例在加样槽中混合后将孔板放入培养箱孵育,后取出使用酶标仪测量在450 nm处吸光度A,记录结果进行分析(此处取的指定时间为:24、48、72、96 h)。
1.4.5. 细胞克隆形成实验
将转染后的细胞重悬为1000 mL的细胞悬液,取6孔板,在每孔内加入2 mL细胞悬液,放入培养箱培养14 d左右取出,清洗、多聚甲醛进行固定、结晶紫(Solarbio)染色,清洗干净后拍照分析。
1.4.6. 细胞划痕实验
将转染后的细胞,放入培养箱培养,待转染细胞密度达80%以上即可。在6孔板内按三等分划出3条较笔直竖线,吸除板内培养基;倒扣在台面,用记号笔在板底按三等分划出3条较笔直横线,用PBS缓冲液清洗划痕过后的孔2次,每孔加入2 mL 1640基础培养基,采集0 h时划痕图像,并记录采集点,放入培养箱培养24 h或48 h后,采集24 h或48 h时划痕图像,保存并进行结果分析。
1.4.7. Transwell实验
将分装的基质胶放于4 ℃冰箱融化,取出小室,将基质胶(Corning)与1640培养基按1∶8或1∶9进行稀释,每个小室内加入100 µL,放入37 ℃ CO2培养箱孵育1~2 h,待基质胶凝固后,向24孔板内加入含有15%胎牛血清(LONSA SCIENCE SRL)1640培养基800 µL,将转染好的细胞从培养箱中取出,用1 mL 1640完全培养基重悬为200 000/mL的细胞悬液,取200 µL细胞种于小室内;放入37 ℃ CO2培养箱培养24 h后,取出孔板,将24孔板内培养基吸除,在每孔内800 µL 4%多聚甲醛,将小室放于其中,进行细胞固定15 min,清洗小室2次,每孔加入800 µL 1%结晶紫染色15 min;清洗小室1次,将小室倒扣于桌面晾干,拍照记录,分析数据。
1.4.8. 细胞蛋白提取
将培养好的细胞从培养箱中取出,根据细胞量加入相应RIPA强效蛋白裂解液(碧云天),置于冰上裂解30 min;12 000 g 20 min 4 ℃离心,吸取上层清亮液体至EP管内,弃去底部沉淀,向EP管内加入5×蛋白Loading buffer(碧云天),使用恒温金属加热器加热100 ℃ 10 min;制备好的蛋白可立即使用,也可放-80 ℃冰箱储存。
1.4.9. 蛋白浓度测定
在96孔板中分别加入0、1、2、4、8、12、16、20 µL稀释至0.5 mg/ mL的蛋白标准溶液,用标准品稀释液补足到20 µL,制成浓度分别为0、0.025、0.05、0.1、0.2、0.3、0.4、0.5 mg/mL的标准品。加入20 µL待测定蛋白样品在96孔板中,其余空孔加入200 µL BCA工作液,37 ℃ 5% CO2培养箱孵育30 min,将96孔板放入酶标仪,在540 nm处测量出A值,收集数据,根据公式绘制曲线。
1.4.10. Western blotting实验
配制不同浓度上层胶和下层胶,将凝胶(碧云天)放入垂直电泳槽,倒入电泳液,加入样品和预染蛋白Marker(麦客生物),电泳30 min左右,将配制好的快速转膜液放冰箱预冷,剪取适合大小PVDF膜(Merck Millipore),用甲醇激活后,覆盖于胶块表面,转膜20~30 min,将转好的膜放于水平摇床,将快速封闭液倒入清洗盒,于水平摇床上,室温轻晃20~30 min,稀释液配制适量浓度一抗(成都正能生物),放入预冷的抗体盒中,将清洗好的PVDF膜转移至盒中,放4 ℃冰箱过夜,将PVDF膜夹出,放清洗盒内,于水平摇床,用1×PBST清洗,配制适当浓度二抗(成都正能生物),将二抗倒入清洗盒内,于水平摇床室温轻晃1~2 h,用1×PBST清洗5 min,配制ECL发光液(Abbkine),将PVDF膜浸泡在配制好的ECL发光液中30 s,在显影仪内显影,收集结果进行分析。
1.4.11. 荧光素酶报告实验
将转染后的细胞取出,去除培养基,清洗2次,加入适量的1×Lysis Buffer(上海汉恒),反复吹打孔内细胞,于水平摇床上,室温、轻晃孵育15 min,充分裂解,将孔内液体转移至1.5 mL EP管中,4 ℃ 12 000 r/min离心1 min,取上清备用。取20 µL液体,加入100 µL Luciferase Reaction Reagent(上海汉恒),混匀后于酶标仪中检测fLuc信号,然后加入100 µL Luciferase Reaction Reagent II(上海汉恒),混匀后于酶标仪中检测rLuc信号,收集数据进行分析。
1.5. 生物信息学分析
运用Sangerbox数据分析平台(http://past20.sangerbox.com/Index)分析FEZF1-AS1在泛癌中的表达;运用仙桃学术(https://www.xiantao.love/)分析TCGA数据库中FEZF1-AS1在配对与非配对的癌与正常肺组织中表达情况。利用相关数据库预测FEZF1-AS1与hsa-miR-130a-5p的结合位点以及hsa-miR-130a-5p与CCND1的结合位点。
1.6. 统计学分析
采用SPSS 26.0和Graphpad 8.0软件进行统计分析与作图,计数资料以n(%)表示,组间比较采用χ2检验;计量资料以均数±标准差表示,组间比较采用t检验与单因素方差分析;P<0.05表示差异有统计学意义。
2. 结果
2.1. FEZF1-AS1在NSCLC中的表达及与临床特征的关系
利用TCGA数据库发现FEZF1-AS1在泛癌中的表达增高(图1A),在NSCLC中也呈现高表达;在非配对与配对的NSCLC癌组织中的表达量显著高于正常肺组织(P<0.001,图1B)。检测33例NSCLC患者的肺癌组织、癌旁组织及5种肺癌细胞系中FEZF1-AS1的表达情况,结果显示在癌组织的表达量显著高于癌旁组织(P<0.0001,图1C)。根据FEZF1-AS1的表达情况,按照癌/癌旁表达量的中间值3.9作为临界值,将其分为高表达组与低表达组,>3.9为高表达组(16例),≤3.9为低表达组(17例),对其临床特征进行分析。结果显示,在高表达组中,FEZF1-AS1的表达与NSCLC患者的年龄、肿瘤大小、肿瘤TNM分期均无相关性(P>0.05),而与性别、吸烟史及淋巴结转移有关(P<0.05,表1)。在肺癌细胞系中FEZF1-AS1的表达均增高,其中在H1299、H358、PC9细胞的表达显著高于BEAS-2B(P<0.0001,图1D)。因此后续实验选用H1299、H358细胞进行。
图1.
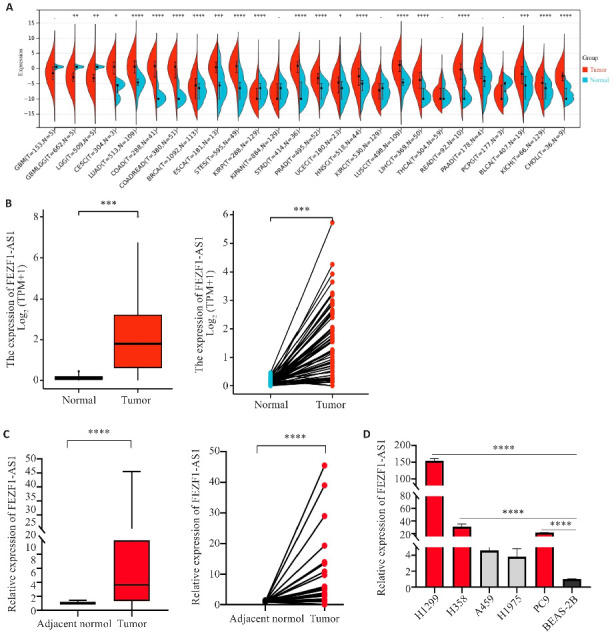
FEZF1-AS1在NSCLC中的表达情况
Fig.1 Expression of FEZF1-AS1 in non-small cell lung cancer (NSCLC) tissues and cell lines. A: Expression of FEZF1-AS1 in pan-cancer in TCGA database. B: Expression of FEZF1-AS1 in unpaired (left) and paired (right) human lung cancer tissues in the TCGA database. C: Expression of FEZF1-AS1 in human NSCLC tissue. D: Expression of FEZF1-AS1 in human lung cancer cell lines. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
表1.
FEZF1-AS1在NSCLC中的表达与临床特征的关系
Tab.1 Relationship between FEZF1-AS1 expression in NSCLC and clinical features of the patients (n)
| Clinical features | Expression of FEZF1-AS1 | χ2 | P | |
|---|---|---|---|---|
| High expression group (n=16) | Low expression group (n=17) | |||
| Age (year) | ||||
| <60 | 5 | 7 | 0.351 | 0.554 |
| ≥60 | 11 | 10 | ||
| Gender | ||||
| Male | 2 | 9 | 6.066 | 0.014 |
| Female | 14 | 8 | ||
| Smoking history | ||||
| Yes | 1 | 6 | 4.160 | 0.041 |
| No | 15 | 11 | ||
| Tumor size (cm) | ||||
| ≤5 | 15 | 16 | 0.002 | 0.965 |
| >5 | 1 | 1 | ||
| Lymph node metastasis | ||||
| Yes | 13 | 6 | 7.127 | 0.008 |
| No | 3 | 11 | ||
| TNM stage | ||||
| Ⅰ-Ⅱ | 13 | 14 | 0.000 | 1.000 |
| Ⅲ-Ⅳ | 3 | 3 | ||
2.2. FEZF1-AS1对非小细胞肺癌的增殖、迁移、侵袭的影响
构建FEZF1-AS1的荧光小干扰RNA(si-FEZF1-AS1),对H1299、H358细胞进行转染(图2A),转染si-FEZF1-AS1后H1299、H358中FEZF1-AS1表达显著降低(P<0.0001,图2B);将H1299、H358分为si-FEZF1-AS1组与si-NC组,两组吸光度在72、96 h有统计学意义差异(P<0.0001,图2C);克隆形成实验结果显示,克隆斑形成数量si-FEZF1-AS1组显著低于si-NC组(P<0.05,图2D)。
图2.
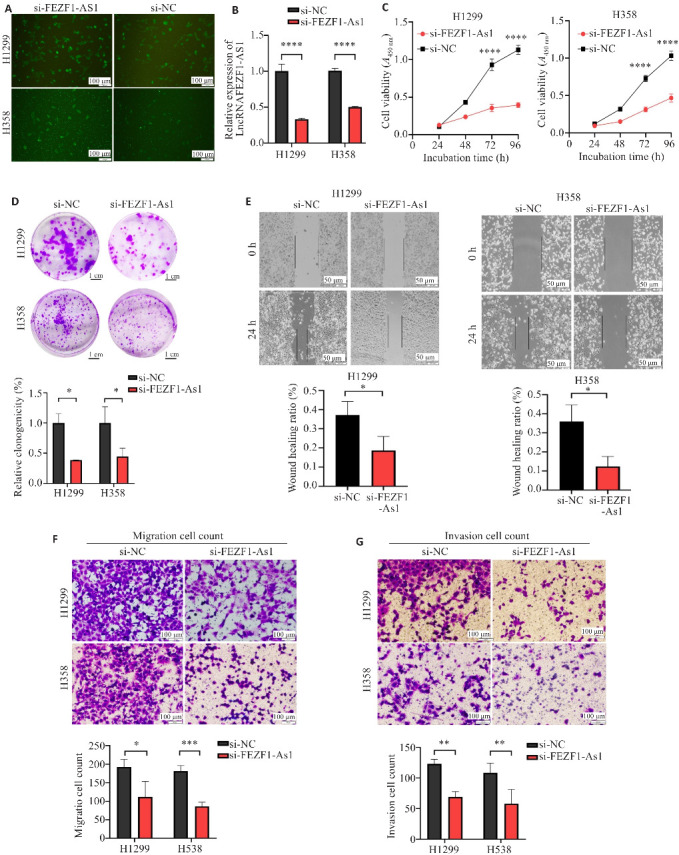
FEZF1-AS1对H1299与H358的增殖、迁移、侵袭功能的影响
Fig.2 Effect of FEZF1-AS1 on proliferation, migration, and invasion of H1299 and H358 cells. A: Fluorescence microscopy of the cells transfected with si-FEZF1-AS1. B: Knockdown efficiency of si-FEZF1-AS1 in H1299 and H358 cells. C: CCK-8 assay of proliferation of H1299 and H358 cells after transfection with si-FEZF1-AS1 and si-NC. D: Formation of H1299 and H358 clones after transfection. E: Healing of H1299 and H358 cell scratches after transfection. F: Transwell migration assay of H1299 and H358 cells after transfection. G: Transwell invasion assay of H1299 and H358 cells after transfection. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
划痕实验结果显示,si-FEZF1-AS1组的划痕愈合率显著低于si-NC组(P<0.05,图2E);si-FEZF1-AS1组的迁移细胞数量显著低于si-NC组(P<0.05,图2F)。Transwell实验结果显示,si-FEZF1-AS1组的侵袭细胞数量显著低于si-NC组(P<0.01,图2G)。
2.3. FEZF1-AS1与hsa-miR-130a-5p存在结合
利用miRbase、targetscan数据库预测发现,FEZF1-AS1与hsa-miR-130a-5p存在3个潜在结合位点(图3A),荧光素酶报告实验结果显示,FEZF1-AS1野生型质粒共转染hsa-miR-130a-5p mimics组的荧光素酶活性显著低于NC mimics组(P<0.001,图3B),而FEZF1-AS1突变型质粒共转染hsa-miR-130a-5p mimics与NC mimics后未出现差异。
图3.
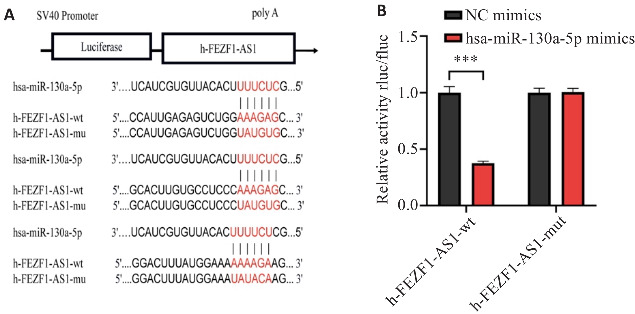
FEZF1-AS1与hsa-miR-130a-5p的结合情况
Fig.3 Binding of FEZF1-AS1 to hsa-miR-130a-5p. A: Prediction of binding sites between FEZF1-AS1 and hsa-miR-130a-5p. B: Dual luciferase reporter assay of the binding between FEZF1-AS1 and hsa-miR-130a-5p. ***P<0.001.
2.4. hsa-miR-130a-5p对非小细胞肺癌的增殖、迁移、侵袭的影响
H1299与H358中si-FEZF1-AS1+hsa-miR-130a-5p inhibitor组的hsa-miR-130a-5p的表达量显著低于si-FEZF1-AS1+NC inhibitor(P<0.0001,图4A)。CKK-8实验结果显示,H1299与H358中si-FEZF1-AS1+hsa-miR-130a-5p inhibitor组的吸光度显著高于si-FEZF1-AS1+NC inhibitor(P<0.001,图4B)。Transwell实验结果显示,si-FEZF1-AS1+hsa-miR-130a-5p inhibitor组的迁移细胞数显著高于si-FEZF1-AS1+NC inhibitor(P<0.05,图4C);si-FEZF1-AS1+hsa-miR-130a-5p inhibitor组的侵袭细胞数显著高于si-FEZF1-AS1+NC inhibitor(P<0.05,图4D)。
图4.
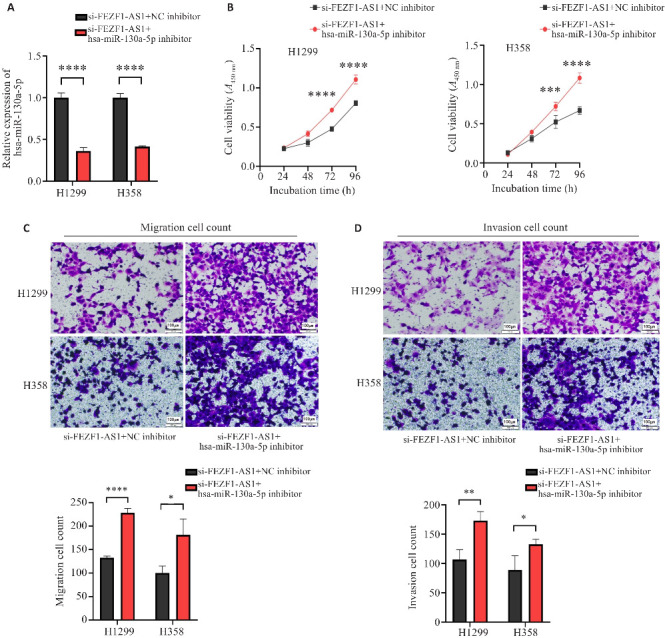
hsa-miR-130a-5p对H1299与H358的增殖、迁移、侵袭功能的影响
Fig.4 Effect of hsa-miR-130a-5p expression level on proliferation, migration, and invasion of H1299 and H358 cells. A: Expression of hsa-miR-130a-5p in the cells after transfection with si-FEZF1-AS1+NC inhibitor and with si-FEZF1-AS1+hsa-miR-130a-5p inhibitor. B: Effect of si-FEZF1-AS1+NC inhibitor and si-FEZF1-AS1+hsa miR-130a-5p inhibitor on cell proliferation. C: Effect of si-FEZF1-AS1+NC inhibitor and si-FEZF1-AS1+hsa miR-130a-5p inhibitor on cell migration. D: Effect of si-FEZF1-AS1+NC inhibitor and si-FEZF1-AS1+hsa-miR-130a-5p inhibitor on cell invasion. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
2.5. hsa-miR-130a-5p与CCND1存在结合
两组细胞转录组测序结果显示,FEZF1-AS1敲减下降的mRNA有4165个(图5A),利用miRDB和ENCORI数据库预测了与hsa-miR-130a-5p结合的靶基因,分别得到940个与1875个靶基因,选取转录组测序中差异度最大的50个mRNA,与之取交集得到了2个mRNA(图5B),通过qRT-PCR发现发现si-FEZF1-AS1组较si-NC组CCND1的表达量显著降低(P<0.0001,图5C、D)。利用miRbase、targetscan数据库对hsa-miR-130a-5p和CCND1的结合位点进行了预测,最终发现了7个结合位点,此处展示3个(图5E),荧光素酶报告实验发现CCND1-3UTR野生型质粒共转染hsa-miR-130a-5p mimics组的荧光素酶活性显著低于NC mimics组(P<0.001,图5F),而CCND1-3UTR突变型质粒共转染hsa-miR-130a-5p mimics与NC mimics后未出现差异。
图5.
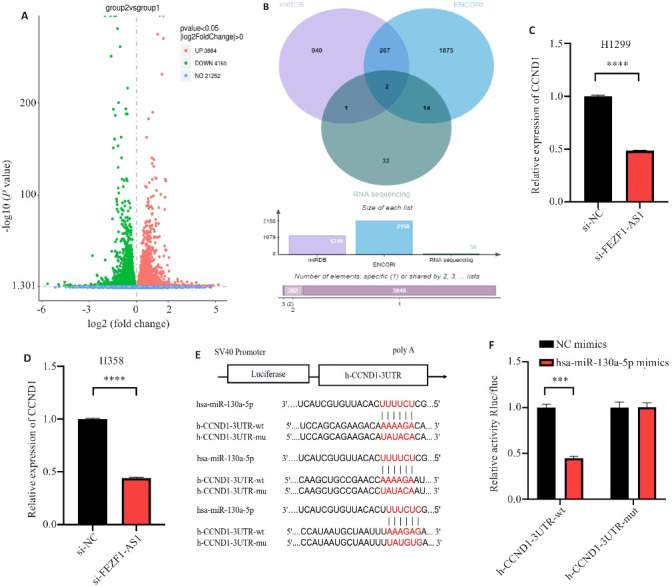
hsa-miR-130a-5p与CCND1的结合情况
Fig.5 Binding status of hsa-miR-130a-5p with CCND1. A: Volcano map of differential gene expression between si-FEZF1-AS1 (group 2) and si-NC groups (group 1) by transcriptome sequencing. B: Database prediction of downstream target proteins and transcriptome sequencing results shown in Venn diagram. C: Knockdown of FEZF1-AS1 and expression of CCND1 in H1299 cells. D: Knockdown of FEZF1-AS1 and expression of CCND1 in H358 cells. E: Prediction of binding sites between hsa-miR-130a-5p and CCND1. F: Dual luciferase reporter assay of binding between hsa-miR-130a-5p and CCND1. ***P<0.001, ****P<0.0001.
2.6. FEZF1-AS1/hsa-miR-130a-5p通过ceRNA机制调控CCND1的表达
Western blot结果显示,si-FEZF1-AS1组较si-NC组CCND1的蛋白表达水平显著降低(P<0.0001,图6A、B),而si-FEZF1-AS1+ hsa-miR-130a-5p inhibitor组相较于si-FEZF1-AS1+NC inhibitor组CCND1的蛋白表达水平显著增高(P<0.05,图6C、D)。
图6.
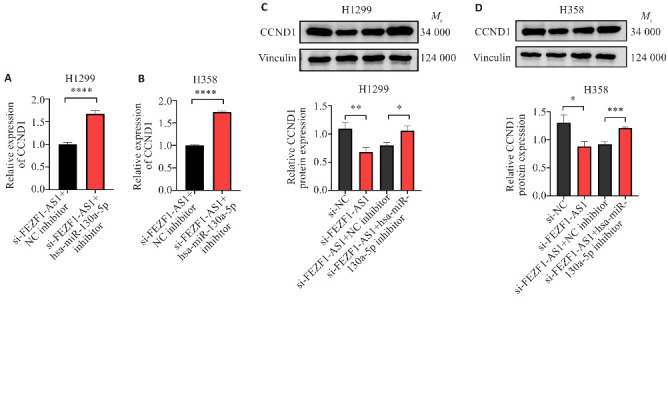
FEZF1-AS1/hsa-miR-130a-5p通过ceRNA机制调控CCND1的表达
Fig.6 FEZF1-AS1/hsa miR-130a-5p regulates CCND1 expression through ceRNA mechanism. A: Expression levels of CCND1 in H1299 cells transfected with si-FEZF1-AS1+NC inhibitor and si-FEZF1-AS1+hsa-miR-130a-5p inhibitor. B: Expression levels of CCND1 in H358 cell transfected with si-FEZF1-AS1+NC inhibitor and si-FEZF1-AS1+hsa-miR-130a-5p inhibitor. C: Protein expression of CCND1 in H1299 cells transfected with si-NC, si-FEZF1-AS1, si-FEZF1-AS1+NC inhibitor and si-FEZF1-AS1+hsa-miR-130a-5p inhibitor. D: Protein expression of CCND1 in H358 cells transfected with si-NC, si-FEZF1-AS1, si-FEZF1-AS1+NC inhibitor and si-FEZF1-AS1+hsa-miR-130a-5p inhibitor. *P<0.05, **P<0.01, ***P<0.001, ****P<0.0001.
3. 讨论
以前对RNA的研究主要集中于编码RNA,但最近ncRNA亦被证明在肿瘤的发生发展中起到了重要的作用,包括LncRNA、CircRNA、miRNA、假基因等,其中LncRNA具备更多的功能,如结合转录因子调节下游基因的表达、产生内源siRNA、与特定蛋白质结合以调节其活性、充当miRNA海绵防止mRNA降解等[14]。已有研究表明,LncRNA对肿瘤增殖、迁移、侵袭、EMT、血管生成、耐药及免疫微环境等均有影响,在肿瘤的发生发展中具有重要作用[15-17]。
FEZF1-AS1作为一种新的LncRNA,在多种肿瘤中均出现差异化表达,如结直肠癌[11]、宫颈癌[12]等,在NSCLC中仅有少量研究。利用生物信息学技术筛查肿瘤潜在的诊疗靶点、结合位点,已成为当代医学重要的分支。本研究利用TCGA数据库分析发现FEZF1-AS1在NSCLC组织中高表达。为了验证数据库的预测,课题组应用qRT-PCR检测33例NSCLC患者的肺癌组织、癌旁组织中FEZF1-AS1的表达情况,结果显示:在癌组织中表达量增高,并且发现FEZF1-AS1的高表达与淋巴结转移有关。这与周丽亚[13]的研究结果不完全相同,但她的研究还显示在表达阳性EGFR突变型中晚期肺腺癌患者耐药的发生率高,预后较差。这与国内其他学者的研究结果相似[12, 18]。究其原因可能是FEZF1-AS1通过促进上皮-间充质转换通路(EMT)、抑制p57抑癌基因、提高磷酸化AKT(pAKT)表达水平等途径促进肿瘤细胞的增殖、侵袭、转移及耐药过程,如EGFR-TKIs耐药[10]。因此明确FEZF1-AS1在NSCLC中的发生机制,对临床预测NSCLC的预后及EGFR-TKIs耐药具有重要价值。
应用qRT-PCR筛选出高表达FEZF1-AS1的NSCLC细胞系H1299、H358,构建si-FEZF1-AS1,通过CCK-8、克隆形成实验、划痕实验、Transwell实验发现:敲低FEZF1-AS1后降低了NSCLC细胞的增殖、迁移和侵袭功能。因此认为FEZF1-AS1促进了非小细胞肺癌的增殖、迁移、侵袭能力。Hui等[19]研究也发现敲低FEZF1-AS1可以下调HMGA2的表达,抑制胃癌细胞的体内致瘤能力,并且调节胃癌细胞的生长。
利用Starbase数据库预测并挑选出与FEZF1-AS1结合的miRNA,发现敲减FEZF1-AS1后hsa-miR-130a-5p的表达量显著增高。细胞功能学实验也证实hsa-miR-130a-5p影响NSCLC细胞的增殖、迁移、侵袭功能。通过荧光素酶报告实验显示:FEZF1-AS1野生型质粒共转染hsa-miR-130a-5p mimics组的荧光素酶活性显著降低,而FEZF1-AS1突变型质粒共转染hsa-miR-130a-5p mimics与NC mimics后未出现差异,这表明FEZF1-AS1与hsa-miR-130a-5p存在结合位点。接着通过CCK-8、Transwell实验发现hsa-miR-130a-5p inhibitor能够逆转si-FEZF1-AS1对肺癌细胞的增殖、迁移和侵袭功能的影响,以上结果表明FEZF1-AS1与hsa-miR-130a-5p存在相互关系。因此认为FEZF1-AS1可以通过hsa-miR-130a-5p调控其在肺癌发生中的作用。
CCND1也被称为细胞周期蛋白D1[20],在正常情况下,CCND1与CDK4或CDK6形成复合物,这对于G1/S细胞周期转换至关重要。它被归类为癌基因,并已被证明可以促进血管生成,肿瘤细胞增殖和对放化疗的耐药性。此外,在没有CDK的情况下,CCND1可以直接与核受体结合,调节细胞增殖、生长和分化等功能[21]。Sun等[22]研究发现USP10可以与CCND1结合并阻止其经历K48连接的多泛素化,从而增加CCND1的稳定性并促进胶质母细胞瘤的增殖和存活。Chen等[23]提出MicroRNA-584通过靶向CCND1抑制胰腺癌中的细胞增殖和侵袭。Hao等[24]认为MicroRNA-195可以通过下调CDC42/CCND1途径从而抑制上皮性卵巢癌中的细胞增殖、迁移和侵袭功能。Liu等[25]证明LY2874455和阿贝西利逆转FGF3/4/19/CCND1扩增介导的NSCLC吉非替尼耐药,以上报道显示CCND1在多种肿瘤中发挥了重要作用。
因此课题组利用miRDB和ENCORI数据库预测了与hsa-miR-130a-5p结合的靶基因,利用miRbase、targetscan数据库预测hsa-miR-130a-5p和CCND1的结合位点,进行荧光素酶报告实验,结果显示:CCND1-3UTR野生型质粒共转染hsa-miR-130a-5p mimics组的荧光素酶活性显著降低,而CCND1-3UTR突变型质粒共转染hsa-miR-130a-5p mimics与NC mimics后未出现差异,这表明CCND1与hsa-miR-130a-5p存在结合,且敲低FEZF1-AS1会使CCND1的表达降低,hsa-miR-130a-5p inhibitor能够逆转FEZF1-AS1对CCND1蛋白水平的敲减作用。因此认为CCND1可以通过ceRNA机制进行调控,且FEZF1-AS1通过miR-130a-5p/CCND1轴促进了NSCLC的发展,此项研究发现尚属首次。Chen等[26]研究发现FEZF1-AS1通过靶向的miR-4497/GBX2轴促进了喉鳞状细胞癌细胞的迁移、侵袭功能。Xie等[27]研究发现FEZF1-AS1通过miR-632/FAM83A轴可以调节体内、外直肠癌细胞增殖、迁移、侵袭。因此认为FEZF1-AS1通过ceRNA调控肿瘤的发生机制及其复杂。
本研究证实了FEZF1–AS1在NSCLC组织中高表达,与NSCLC淋巴结转移有关,促进了NSCLC细胞的增殖、迁移、侵袭,并通过miR-130a-5p/CCND1轴促进了非小细胞肺癌的发展。但本研究仍有不足之处:(1)临床样本较少,且多为早期病例;(2)在NSCLC发生中,FEZF1-AS1及miR-130a-5p/CCND1的上下游是否存在其他的调控机制尚需进一步研究;(3)未来是否能找到有效的作用靶点、克服靶向耐药、能否真正在肺癌的诊断及治疗中产生作用,这还需要我们进一步的探索。
基金资助
国家自然科学基金(82373329);安徽省教育厅自然科学研究重点项目(KJ2018A0997)
Supported by National Natural Science Foundation of China (82373329).
参考文献
- 1. Brody H. Lung cancer[J]. Nature, 2020, 587(7834): S7. DOI: 10.1038/d41586-020-03152-0 [DOI] [PubMed] [Google Scholar]
- 2. Cheung CHY, Juan HF. Quantitative proteomics in lung cancer[J]. J Biomed Sci, 2017, 24(1): 37. DOI: 10.1186/s12929-017-0343-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Li FB, Gu FM, Li Q, et al. ROR1-AS1 knockdown inhibits growth and invasion and promotes apoptosis in NSCLC cells by suppression of the PI3K/Akt/mTOR pathway[J]. J Biochem Mol Toxicol, 2021, 35(5): e22726. DOI: 10.1002/jbt.22726 [DOI] [PubMed] [Google Scholar]
- 4. Fang YW, Fullwood MJ. Roles, functions, and mechanisms of long non-coding RNAs in cancer[J]. Genomics Proteomics Bioinformatics, 2016, 14(1): 42-54. DOI: 10.1016/j.gpb.2015.09.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Barrett T, Troup DB, Wilhite SE, et al. NCBI GEO: archive for functional genomics data sets: 10 years on[J]. Nucleic Acids Res, 2011, 39(Database issue): D1005-10. DOI: 10.1093/nar/gkq1184 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Temraz S, Mukherji D, Alameddine R, et al. Methods of overcoming treatment resistance in colorectal cancer[J]. Crit Rev Oncol Hematol, 2014, 89(2): 217-30. DOI: 10.1016/j.critrevonc.2013.08.015 [DOI] [PubMed] [Google Scholar]
- 7. Zhang MH, Yang Y, Zhao Y, et al. LncRNA DQ786243 expression as a biomarker for assessing prognosis in patients with gastric cancer[J]. Eur Rev Med Pharmacol Sci, 2018, 22(8): 2304-9. [DOI] [PubMed] [Google Scholar]
- 8. Zhang Q, Chen BY, Liu P, et al. XIST promotes gastric cancer (GC) progression through TGF‑β1 via targeting miR-185[J]. J Cell Biochem, 2018, 119(3): 2787-96. DOI: 10.1002/jcb.26447 [DOI] [PubMed] [Google Scholar]
- 9. Yu WJ, Sun ZN, Yang L, et al. lncRNA PTAR promotes NSCLC cell proliferation, migration and invasion by sponging microRNA-101[J]. Mol Med Rep, 2019, 20(5): 4168-74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Chen S, Zhou L, Ran RZ, et al. Circ_0016760 accelerates non-small-cell lung cancer progression through miR-646/AKT3 signaling in vivo and in vitro [J]. Thorac Cancer, 2021, 12(23): 3223-35. DOI: 10.1111/1759-7714.14191 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Bian ZH, Zhang JW, Li M, et al. LncRNA-FEZF1-AS1 promotes tumor proliferation and metastasis in colorectal cancer by regulating PKM2 signaling[J]. Clin Cancer Res, 2018, 24(19): 4808-19. DOI: 10.1158/1078-0432.ccr-17-2967 [DOI] [PubMed] [Google Scholar]
- 12. Liang M, Li YK, Dai TT, et al. lncRNA FEZF1-AS1 regulates biological behaviors of cervical cancer by targeting miRNA-1254[J]. Food Sci Nutr, 2021, 9(9): 4722-37. DOI: 10.1002/fsn3.2315 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. 周丽亚. FEZF1-AS1和AKT3在中晚期肺腺癌中的表达及与EGFR-TKIs疗效关系的研究[D]. 蚌埠: 蚌埠医学院, 2022: 1-2. [Google Scholar]
- 14. Shi CL, Sun L, Song YS. FEZF1-AS1: a novel vital oncogenic lncRNA in multiple human malignancies[J]. Biosci Rep, 2019, 39(6): BSR20191202. DOI: 10.1042/bsr20191202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Li M, Wei L, Liu PY, et al. Lnc-ATG9B-4 aggravates progress of hepatocellular carcinoma through cell proliferation and migration by upregulating CDK5[J]. Exp Biol Med, 2021, 246(2): 177-86. DOI: 10.1177/1535370220963197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16. Luo HL, Luo T, Liu JJ, et al. Macrophage polarization-associated lnc-Ma301 interacts with caprin-1 to inhibit hepatocellular carcinoma metastasis through the Akt/Erk1 pathway[J]. Cancer Cell Int, 2021, 21(1): 422. DOI: 10.1186/s12935-021-02133-1 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Wang J, Tan LR, Yu XT, et al. lncRNA ZNRD1-AS1 promotes malignant lung cell proliferation, migration, and angiogenesis via the miR-942/TNS1 axis and is positively regulated by the m6A reader YTHDC2[J]. Mol Cancer, 2022, 21(1): 229. DOI: 10.1186/s12943-022-01705-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Liu WW, Li Y, Zhang Y, et al. Circulatinglong non-coding RNA FEZF1-AS1 and AFAP1-AS1 serve as potential diagnostic biomarkers for gastric cancer[J]. Pathol Res Pract, 2020, 216(1): 152757. DOI: 10.1016/j.prp.2019.152757 [DOI] [PubMed] [Google Scholar]
- 19. Hui YJ, Yang Y, Li DP, et al. LncRNA FEZF1-AS1 modulates cancer stem cell properties of human gastric cancer through miR-363-3p/HMGA2[J]. Cell Transplant, 2020, 29: 963689720925059. DOI: 10.1177/0963689720925059 [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 20. Zhu DW, Huang J, Liu N, et al. PSMC2/CCND1 axis promotes development of ovarian cancer through regulating cell growth, apoptosis and migration[J]. Cell Death Dis, 2021, 12(8): 730. DOI: 10.1038/s41419-021-03981-5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Wei YJ, Chen W, Li ZH, et al. EIF3H stabilizes CCND1 to promotes intrahepatic cholangiocarcinoma progression via Wnt/β-catenin signaling[J]. FASEB J, 2022, 36(12): e22647. DOI: 10.1096/fj.202200913r [DOI] [PubMed] [Google Scholar]
- 22. Sun T, Xu YJ, Jiang SY, et al. Suppression of the USP10/CCND1 axis induces glioblastoma cell apoptosis[J]. Acta Pharmacol Sin, 2021, 42(8): 1338-46. DOI: 10.1038/s41401-020-00551-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Chen G, Hu M, Qu XS, et al. MicroRNA-584 directly targets CCND1 and inhibits cell proliferation and invasion in pancreatic cancer[J]. Mol Med Rep, 2019, 19(1): 719-26. [DOI] [PubMed] [Google Scholar]
- 24. Hao XY, Jia QQ, Yuan JL, et al. MicroRNA-195 suppresses cell proliferation, migration and invasion in epithelial ovarian carcinoma via inhibition of the CDC42/CCND1 pathway[J]. Int J Mol Med, 2020, 46(5): 1862-72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Liu DC, Liu HG, Gan JD, et al. LY2874455 and abemaciclib reverse FGF3/4/19/CCND1 amplification mediated gefitinib resistance in NSCLC[J]. Front Pharmacol, 2022, 13: 918317. DOI: 10.3389/fphar.2022.918317 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Chen XD, Cheng P, Hu CH. LncRNA FEZF1-AS1 accelerates the migration and invasion of laryngeal squamous cell carcinoma cells through miR-4497 targeting GBX2[J]. Eur Arch Otorhinolaryngol, 2021, 278(5): 1523-35. DOI: 10.1007/s00405-021-06636-5 [DOI] [PubMed] [Google Scholar]
- 27. Xie RJ, Liu CB, Liu LF, et al. Long non-coding RNA FEZF1-AS1 promotes rectal cancer progression by competitively binding miR-632 with FAM83A[J]. Acta Biochim Biophys Sin, 2022, 54(4): 452-62. DOI: 10.3724/abbs.2022022 [DOI] [PMC free article] [PubMed] [Google Scholar]