

Abstract
The previous history of contraction and length changes of a muscle influences the size of the stretch reflex and H reflex. Here we ask, is this dependence due to changes in mechanical properties of extrafusal fibres, intrafusal fibres of spindles, or both?
The soleus muscle of human subjects was conditioned using either a voluntary contraction or a contraction evoked by low-strength electrical stimulation, in the range 0–25% of maximum. Following conditioning, reflexes were increased by more than twofold above the no-contraction value by a voluntary contraction of 5% of maximum, or more, but not by electrical stimulation which presumably did not contract the intrafusal fibres of spindles.
When the muscle was conditioned with a contraction at a length shorter than the test length, rather than at the test length, a depressing effect on reflexes was attributed to both the burst of impulses generated in spindles when the muscle was stretched back to the test length and to a reduced stretch sensitivity of muscle spindles.
The experiments demonstrate the importance of keeping the muscle and its spindles in a defined mechanical state when measuring reflexes. They also point to the powerful facilitating influences of conditioning muscle contractions provided they recruit the intrafusal fibres of spindles.
Work from our laboratory has established that the responses of muscle spindles and their spinal reflex action are dependent on the muscle's immediate previous history of contraction and length changes, because muscle exhibits thixotropy (for a review see Proske et al. 1993). To demonstrate extremes of behaviour of spindles and spindle reflexes in relation to muscle history effects, we have devised two forms of conditioning, termed hold-long conditioning and hold-short conditioning. Hold-long conditioning consists of stretching the muscle to a length longer than that at which the test measurements are to be made, then contracting the muscle and holding it at that length for a few seconds before returning it to the test length. Similarly, hold-short conditioning is where the muscle is shortened, contracted and then stretched back to the test length.
The reasoning underlying these forms of conditioning is as follows. As the muscle relaxes following a contraction, a number of stable cross-bridges form between actin and myosin filaments in sarcomeres of muscle fibres (Hill, 1968). The presence of stable cross-bridges gives the muscle a degree of passive stiffness. As a result, when the stiffened muscle fibres are shortened, as occurs after hold-long conditioning, they fall slack. By contrast, slack does not develop if the contraction is carried out at a shorter length and the muscle is then stretched back to the test length, that is, after hold-short conditioning.
We hypothesize that slack is able to form in both extrafusal and intrafusal fibres. The presence of slack in intrafusal fibres has a large effect on the responses of muscle spindles. Their response to locally applied vibration is reduced (Morgan et al. 1991) as is their response to stretch (Proske et al. 1992). That, in turn, leads to depression of the phasic stretch reflex (Gregory et al. 1987) and to facilitation of the H reflex as a result of less post-activation depression from the lower level of spindle background activity (Gregory et al. 1990; Hultborn et al. 1996; Wood et al. 1996).
In all of the animal studies care was taken to make conditioning stimulation strong enough to recruit both α-and γ-motoneurones and the stimulation was given immediately after the conditioning length change had been completed (see Fig. 1). In the human experiments, voluntary contractions were used, and since they were effective, it was assumed that they activated both skeletomotor and fusimotor neurones. This is consistent with the available evidence (Burke et al. 1979). However, it remained uncertain whether the effects on reflexes were the result of changes in mechanical properties of extrafusal fibres, intrafusal fibres, or both. Therefore a central aim of this study was to establish whether fusimotor activation was necessary to reset spindle sensitivity after hold-long conditioning.
Figure 1. Conditioning and testing of human soleus muscle.

The three methods of muscle conditioning are shown, in diagrammatic form, at the top. After stretch of the muscle (hold-long), shortening (hold-short), or no length change (hold-test), a fusimotor-strength contraction is given (filled bar). After the contraction the muscle is kept at that length for several seconds and is then returned to the test length where the test stimulus is given (arrow). The actual conditioning sequences employed in this study are represented by the sample traces shown at the bottom. For each trial the muscle was first pre-conditioned by being stretched, represented by 30 deg ankle rotation in a dorsiflexing direction from the 110 deg test position. At the longer length subjects carried out a 2 s contraction at a strength of 25% MVC. They were then instructed to completely relax while the muscle was held at that length for a further 5 s before being returned to the test length (hold-test) or to a length corresponding to 30 deg plantarflexion from the test position (hold-short). Three seconds after moving to the new position a conditioning contraction was given. The contraction was either voluntary or evoked by percutaneous electrical stimulation. Contraction strengths were adjusted to correspond to 0, 5, 10 and 25% MVC for the voluntary contraction, and 0, 5 and 10% for the electrical stimulation. Contraction strengths were measured as rectified, smoothed EMG (not shown). After the conditioning contraction subjects were again asked to fully relax while the muscle was returned to the test length 5 s later (hold-short) or kept at the test length (hold-test) and then the test stimulus was given to elicit a stretch reflex or H reflex. The top and second traces show ankle torque and soleus EMG, respectively, during pre-conditioning and hold-test conditioning, followed by the test reflex. The third and fourth traces show torque and EMG, respectively, during hold-short conditioning and test reflex. The bottom trace shows the ankle angle. The record of ankle angle is shown as a dashed line for hold-short conditioning and as a continuous line for hold-test conditioning. The reflex EMG in response to the tendon tap is shown on an expanded time scale.
In some recent experiments (Wise et al. 1998), instead of contracting the muscle at a shorter-than-test length, it was simply contracted at the test length (Fig. 1; hold-test). At first sight, hold-short and hold-test conditioning should have left the muscle and its spindles in the same mechanical state, free of any slack. However, there remained some uncertainty about precisely how the hold-short conditioning procedure exerted its effects. Was the movement back to the test length at the end of hold-short conditioning itself sufficient to produce a change in the state of the spindles? If a muscle contraction was necessary, did it have to be strong enough to recruit γ-motoneurones to produce the full effects of muscle conditioning on reflex size, or was a contraction which was purely extrafusal sufficient?
Here we have explored these questions using modified conditioning procedures designed to allow a separation of the three components of conditioning, that is, the movement, the extrafusal contraction and the intrafusal contraction. Contractions assumed to be purely extrafusal, of different strengths, were produced by electrical stimulation of the muscle, while fusimotor recruitment was achieved using voluntary contractions of comparable strengths. Using the stretch reflex as our test, the method allowed us to obtain an estimate of the level of fusimotor activation at various levels of voluntary contraction, in a way that has not been done before.
Our observations suggested that mechanical changes in muscle, after conditioning of the extrafusal fibres only, were insufficient to account for the observed changes in reflex amplitude. In addition, it was found that hold-short and hold-test conditioning did not leave the muscle in precisely the same state. Here we have used both stretch reflex and H reflex testing to explore the reasons underlying this difference.
METHODS
The experiments were performed on eight healthy subjects aged 22–25 years. Subjects all gave informed written consent to the experimental procedures which had been approved by the relevant ethics committee. Subjects were seated in a reclining chair which was adjusted so that the right leg was flexed at the hip by approximately 120 deg and at the knee by approximately 130 deg. The right foot was strapped to a footplate so that the angle formed between the footplate and the lower leg was 110 deg. The footplate could be rotated about an axis co-incident with the axis of the ankle joint and could be fixed in any position, in both the plantar and dorsiflexed directions. A potentiometer aligned with the axis of the ankle joint provided a signal related to joint angle.
The tendon jerk reflex in triceps surae was elicited using a specially constructed tendon hammer. The hammer was designed to deliver blows of equal energy. It consisted of a metal bar that pivoted about its centre. The bar was counterbalanced so that gravitational forces could not influence the speed of the hammer once it had been released by a spring. The bar left the spring before striking the tendon, ensuring that all of the spring's energy was transferred to the hammer. The strength of the tap could be adjusted by altering the tension in the spring.
The triceps surae H reflex was evoked with a 1 ms duration constant current pulse applied though a Ag-AgCl adhesive electrode (3M ‘Red-Dot’ paediatric electrode) positioned in the popliteal fossa. The anode was a rectangular plate (6 cm × 4 cm) strapped above the patella. Conductive gel was applied between the plate and skin. Stimulus strength was adjusted so that the size of the evoked H reflex (unconditioned) was approximately 25–50% of the maximal motor response. Stimulus conditions were adjusted so that a small direct response was visible together with each reflex. This provided a useful control for electrode movement. The recording electrodes used were also Ag-AgCl adhesive electrodes and these were placed over the soleus muscle. Reflexes were measured as the peak-to-peak amplitude of the EMG response, and for each experimental sequence values were averaged over five trials. Reflexes were normalized with respect to the largest reflex recorded in a session, which meant that the calculated averages were always less than one (Wood et al. 1996).
Part of the experiment involved contraction of the triceps surae by electrical stimulation. Distributed stimulation was used to keep stimulus intensity low (Joyce et al. 1969). Usually three moist pad electrodes were strapped over different parts of the calf, but for one subject only two were used, and in one other subject only a single electrode was used. Interleaved trains of pulses (200 μs, 10–32 mA rectangular pulses, 42 pulses per second per channel) were delivered through the stimulating (cathodal) electrodes with respect to an indifferent (anodal) electrode strapped laterally, just below the knee. A stimulus programmer (WPI Pulsemaster A300) rotated through the channels at equal intervals (8 ms where 3 electrodes were used). We found that contractions of 5 and 10% of a maximum voluntary contraction (MVC) could be evoked with stimulus currents below 1.5 times motor threshold, which we felt was safely below the threshold for stimulating fusimotor axons (Gandevia et al. 1986). The electrodes were relatively large discs of 50 mm diameter, chosen to minimize discomfort to the subject. This was considered important, since we wished to avoid central effects that might be evoked by uncomfortable or painful stimulation, and which might therefore influence the size of the test reflex.
Subjects were asked to produce voluntary contractions of different strengths by matching an EMG signal. EMG was used rather than torque because, particularly in the hold-short position, torque levels were too low to be accurately reproducible (Fig. 1). Care was taken to ensure that the subject remained quite relaxed following the conditioning contraction (or no contraction), by monitoring the EMG trace. Trials affected by unintentional contractions were discarded.
The muscle conditioning procedure used in the experiments is shown in Fig. 1. The chosen sequence ensured that for each test measurement the muscle was in a precisely defined state. Conditioning was always begun with a stretch of the muscle, represented by ankle rotation of 30 deg, in a dorsiflexing direction, from the 110 deg test position. At the longer length, subjects were asked to carry out a contraction for about 2 s at a strength equal to 25% MVC, as estimated from the size of the rectified and smoothed EMG signal. Subjects were then asked to relax while the muscle was kept in position for a further 5 s. Stretching of the muscle and contraction was termed ‘pre-conditioning’ (Fig. 1). The idea of the pre-conditioning was to ‘reset’ muscle spindles by producing stable cross-bridges in intrafusal fibres at long lengths so that spindles would finish up slack at the test length. After pre-conditioning the muscle was either shortened back to the test length (30 deg plantar flexion) and contracted a second time, called ‘hold-test’ conditioning, or shortened by a total of 60 deg plantarflexion and contracted, called ‘hold-short’ conditioning. With hold-test conditioning, the test stimulus was given 10 s after the second contraction. With hold-short conditioning the muscle was brought back to the test length 5 s after the contraction and the test stimulus was given 5 s later (Fig. 1).
The second contraction was either a voluntary contraction or an electrically evoked contraction. Four contraction strengths were used: 0, 5, 10 and 25% MVC. Voluntary contractions were measured and displayed to the subject as rectified, smoothed EMG. To help subjects achieve a set level of contraction a visual target was presented showing the required level. Contractions generated by electrical stimulation were graded by altering stimulus strength within a range that was likely to be below fusimotor threshold and was not painful. Stimulus strength was adjusted to produce torque levels equal to those produced by voluntary contractions of 5 and 10% MVC.
Reflex responses were displayed on a Nicolet digital oscilloscope. Data were recorded using an Apple Power Macintosh 7100/66 computer and a MacLab/8s system running Chart software (ADInstruments). Data were analysed using the software package Igor Pro (WaveMetrics, OR, USA).
Statistical analysis was carried out using repeated measures ANOVA, least-significant difference (LSD) analysis and Student's t test. The analysis was carried out using the program SPSS (SPSS Inc., Chicago, IL, USA) run on a Macintosh computer.
RESULTS
The stretch reflex and intrafusal thixotropy
The main result of this study was that after muscle pre-conditioning, no contraction at the test length led to a small reflex but a conditioning voluntary contraction produced a large increase in reflex size. If, instead of a voluntary contraction, the contraction was evoked by percutaneous electrical stimulation, the reflex did not increase significantly. Examples of the effects of voluntary and electrically evoked contractions on the test reflex are shown in Fig. 2.
Figure 2. Sample records of effects on the reflex, after pre-conditioning, of a voluntary contraction of 10% MVC (upper panel) or a 10% contraction evoked by percutaneous electrical stimulation (lower panel).
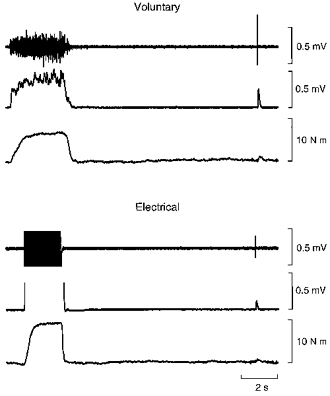
In each panel, the upper trace shows the raw EMG recorded in soleus, the second trace the rectified smoothed EMG record and the bottom trace, torque. Electrical stimulation leads to large artifacts which obscure the EMG record. The reflex after electrical stimulation is smaller than after a voluntary contraction.
A voluntary contraction of 5% MVC more than doubled reflex size and a 10% contraction doubled it again. Interestingly, a 25% contraction produced no further increase (Fig. 3). Electrically evoked contractions at 5 and 10% MVC increased the reflex slightly but non-significantly above the no-contraction value (Fig. 3).
Figure 3. Size of stretch reflex after two forms of muscle conditioning.

Size of stretch reflex (means ± s.e.m.) normalized with respect to its maximal value, plotted against the size of the conditioning contraction. Circles, values obtained when a voluntary conditioning contraction was used; squares, contraction evoked by electrical stimulation. Open symbols, values measured after hold-test conditioning; filled symbols, values measured after hold-short conditioning.
This difference between the effects on reflex size of no contraction or an electrically evoked contraction, when compared with a voluntary contraction, was observed in all subjects. Pooled values for the eight subjects showed a highly significant difference between the two groups (P < 0.01; paired t test).
A repeated measures ANOVA with the factors contraction strength (5, 10 and 25% MVC) and form of conditioning revealed a significance difference between hold-test and hold-short values (P < 0.001). A repeated measures t test showed that there was no significant difference between the 0% values for hold-test versus hold-short. A LSD analysis on the effects of voluntary contractions of different strength after hold-test conditioning showed that the values at 5% were significantly lower than those at 10 and 25% (P < 0.05).
These observations suggest that it is the fusimotor co-activation during a voluntary contraction which is responsible for the observed increase in the reflex after hold-long pre-conditioning.
Two forms of conditioning
An unexpected result emerged when the experiment described above was extended to include hold-short as well as hold-test conditioning.
Our prediction had been that the conditioning contractions during hold-test and hold-short conditioning would both remove any pre-existing slack in intrafusal and extrafusal fibres, so that it was expected that reflexes after the two forms of conditioning would be similar in size. That was clearly not the case for most of the experimental conditions (Fig. 3). When there was no conditioning contraction, reflexes were not significantly different after hold-short and hold-test pre-conditioning (t test; P> 0.05). When a conditioning contraction was present, if it was a voluntary contraction, the reflex after hold-test was significantly larger than that after hold-short (t test; P < 0.05), for all three contraction strengths used. Contractions evoked electrically, and in strength equivalent to 5 and 10% MVC, were ineffective in generating the hold-short-hold-test difference. In fact, hold-test conditioning with electrically evoked contractions led to reflexes which were not significantly different from reflexes after hold-short conditioning, whether this was with an electrically evoked or voluntary contraction, or even when there was no contraction at all. It was concluded that hold-short and hold-test conditioning did not leave spindles and motoneurones in the same state, provided a conditioning voluntary contraction was used. It now remained to explain this result. We decided to explore possible effects at the level of the motoneurones by using an H reflex rather than a stretch reflex.
The H reflex
Here only voluntary contractions of 10% MVC were used to condition the muscle, since the aim was to explore the hold-short-hold-test difference, when it was present.
The first point was to establish whether such a difference was present with the H reflex, using the two forms of conditioning and, indeed, it was (Fig. 4). This meant that whatever mechanism was responsible for the reflexes being small after hold-short conditioning, whether it was of peripheral or central origin, it was not attributable solely to a change in the mechanical state of the spindles since the H reflex showed the effect.
Figure 4. Time course of conditioning effects on stretch reflex and H reflex.

Upper panel: normalized stretch reflexes (means ±s.e.m.) measured after a 10% MVC conditioning contraction plotted against conditioning-test interval. ^, hold-test conditioning; •, hold-short conditioning. Lower panel: H reflexes, elicited using a stimulus strength of 25% of maximal motor response, measured after a 10% MVC conditioning contraction. ^, hold-test conditioning; •, hold-short conditioning.
A number of explanations were considered. One, based on the observations of Hultborn et al. (1996) (see also Táboríková & Sax, 1969; Katz et al. 1977) and supported by some of our own observations (Wood et al. 1996; Wood, 1997), was that the burst of spindle impulses produced on stretching the muscle back to the test length after hold-short conditioning produced a long-lasting depression of motoneurones, called post-activation depression, and that this was responsible for the small reflexes after hold-short conditioning. To test this possibility, the interval between the time of restretch and the test stimulus was increased from the usual 5 s to 10 and 15 s. The rationale was that if the spindle afferent burst during stretch of the muscle was responsible for depression of the motoneurones after hold-short conditioning, test stimuli delivered at the longer intervals should be depressed less and, provided no other mechanisms were operating, the hold-short-hold-test difference should disappear, since it is known that the depression lasts for about 10 s (Hultborn et al. 1996). That was indeed the case for the H reflex, but for the stretch reflex, there was no change with time, and a significant difference persisted for all conditioning-test intervals (Fig. 4). It was concluded that the explanation used to account for changes in the H reflex after the two forms of conditioning could not account for the changes observed with the stretch reflex.
A repeated measures t test showed a significant difference for the hold-short versus hold-test H reflex at the 5 s interval (P < 0.03) but not at the longer intervals. For the stretch reflex there were no significant differences between values at different times, but for both reflexes there was a highly significant effect of conditioning on reflex size (P < 0.001, repeated measures ANOVA).
DISCUSSION
The experiments described in this study have provided a number of important results. First, in confirmation of what was already known, the size of both the stretch reflex and the H reflex are dependent on the previous form of muscle conditioning (for a review see Proske et al. 1993; see also Hagbarth et al. 1995; Hagbarth & Nordin, 1998). Given that both reflexes are used routinely as diagnostic tools and both are notoriously variable in size, it is important to remember that their size may be halved or doubled depending on what has happened to the muscle beforehand.
Intrafusal thixotropy
Perhaps the most important result achieved in this study was that after muscle pre-conditioning the reflex could be facilitated with a voluntary contraction at the test length but not with a contraction of the same strength evoked electrically using a stimulus that was too weak to recruit γ-motoneurones. During voluntary isometric contractions there is good evidence for co-activation of skeletomotor and fusimotor neurones (Vallbo, 1971; Burke et al. 1979; Wilson et al. 1997). Our interpretation of the increase in stretch reflex size produced by the voluntary contraction is that slack in muscle fibres, both extrafusal and intrafusal, produced by pre-conditioning, is taken up by the voluntary contraction and that fusimotor activation is necessary to remove the slack in spindles. The increase in reflex size is not the result of a central facilitation of motoneurones since it is known that H reflexes are depressed following a voluntary contraction (Crone & Nielsen, 1989).
An important finding is that there is some fusimotor activation of spindles during a voluntary contraction of 5% MVC, while at the 10% level, activation is sufficient to maximally facilitate the reflex, that is, reflex amplitude is not increased further by larger voluntary contractions. It is tempting to conclude that the intrafusal slack in spindles produced by the pre-conditioning was fully taken up by the 10% contraction but not by the 5% contraction. A 25% contraction produces no greater fusimotor activation than 10% or, more likely, leaves intrafusal fibres in a mechanical state no different from that after a 10% contraction. In this context, the recent findings by Wilson et al. (1997) showed that, in human tibialis anterior, contraction threshold for spindle acceleration, presumably representing fusimotor threshold, was less than 3.2% MVC. Although we have not systematically studied this point, observations in animal experiments suggest that take-up of slack in intrafusal fibres depends on both the duration and rate of fusimotor stimulation (J. E. Gregory, D. L. Morgan & U. Proske, unpublished observations).
If the contraction was evoked by surface electrical stimulation of the muscle, using distributed stimulation, there was no increase in the size of the reflex. Stimulus strength used was typically less than 1.5 times threshold for a contraction which, with this method of stimulation, is likely to be below threshold for fusimotor neurones (Gandevia et al. 1986). A hold-short movement alone was equally ineffective in increasing reflex size. It had been considered possible that an imposed movement, or internal movement, or lateral compressive forces during an extrafusal contraction, might be effective in removing some of the slack from spindles, but such was not the case. This result confirms our earlier observations on muscle spindles of the anaesthetized cat (Gregory et al. 1986).
Surface electrical stimulation of the muscle will excite some afferent impulses, both in skin and muscle. Muscle afferents will, of course, also be excited by a voluntary contraction. That leaves the question of whether the effects, or lack thereof, of electrical stimulation can be attributed to stimulation of cutaneous afferents. Arguments against an inhibitory action on the reflex by cutaneous afferents include the fact that 5 and 10% contractions produced similar effects and an inhibitory action would not be expected to last for the 8 s between the end of conditioning and the test stimulus. If electrical stimulation produced some inhibition, no stimulation and electrical stimulation would not have been expected to have similar effects on reflexes (Fig. 3).
For the stretch reflex we had also considered the possibility that removal of extrafusal slack would stiffen the muscle and therefore change in some way the characteristics of the mechanical stimulus transmitted to muscle spindles during a tendon tap, compared with the situation when the muscle was slack. While that may be the case, the effect on the reflex was not large enough to be measured in the present experiments. We conclude that muscle history effects on stretch reflexes are dominated by the mechanical state of the intrafusal fibres.
Two forms of conditioning
When hold-long pre-conditioning was followed by no conditioning contraction, or by an electrically evoked contraction, there was no difference in reflex size between hold-short and hold-test conditioning. This means that the movements of the muscle associated with the conditioning procedures, by themselves, do not establish the hold-test- hold-short difference. It requires a voluntary contraction of the muscle to establish this difference.
This leads to the second important result of the study that, provided a conditioning voluntary contraction is given, hold-short and hold-test conditioning are not equivalent in their effects on reflex amplitude (Fig. 4). We believe two factors are responsible for the larger reflexes seen after hold-test than after hold-short conditioning. First, the burst of afferent impulses following restretch to the test length after hold-short conditioning has a depressing effect on reflex amplitude. This may be the main factor operating with the H reflex, where the difference disappeared with longer conditioning-test intervals. However, given that this depression lasts only for about 10 s, it remains to be explained why the difference in stretch reflex size persisted with long conditioning-test intervals (Fig. 4). Certainly any central depressing effect from the burst of impulses after hold-short conditioning should have been well over at 15 s, as was shown by the H reflex. Some insight into possible mechanisms has been provided by recent recordings of spindle afferents in the soleus muscle of the anaesthetized cat (A. K. Wise, J. E. Gregory & U. Proske, unpublished observations). Typically, after hold-test conditioning, resting discharge levels were slightly higher than after hold-short conditioning, suggesting that the mechanical state of spindles was not quite the same. Our earlier work has shown that the size of the stretch reflex in the anaesthetized cat is critically dependent on the interimpulse intervals in the afferent burst evoked by a tendon tap, but not on the number of impulses (Wood et al. 1994). Based on our animal experiments, we propose that the response of human muscle spindles to a tendon tap is greater after hold-test conditioning and that this accounts for the persisting difference in hold-test versus hold-short reflex size at long conditioning-test intervals. By contrast, the short-lasting H reflex difference is due to a central depression. Presumably the tendon jerk is subjected to the same depression but this is overridden by the difference in mechanical state of the spindles.
Although reflex responses were recorded as EMG in soleus, it is very likely that there were similar responses in lateral and medial gastrocnemius as well. Some consideration must also be given to possible influences on the reflex from antagonist muscles. Subjects were instructed to contract agonists only, but antagonist EMG was not routinely monitored to determine the degree of co-contraction, if any, during each contraction. After a movement which introduced slack in antagonists (hold-short conditioning), the level of afferent activity coming from them would be low, so that any reflex effects would be small. After a contraction at the test length any co-contraction of antagonists would have been expected to lead to inhibition, pre- and postsynaptic, of the test muscle, yet the overriding effect seen was a potentiation. We concluded that the effects from antagonists, if any, were small.
To conclude, we have described a number of ‘environmental’ factors which must be taken into account when measuring human stretch reflexes. If the muscle has undergone a brief stretch, this is followed by a period of central depression. Shortening of the muscle after a stretch, brief or long-lasting, is likely to introduce some slack in the intrafusal fibres of muscle spindles. Contraction of intrafusal fibres is required to remove the slack and therefore to sensitize muscle spindles to stretch. Here a voluntary contraction as weak as 10% MVC is sufficient. Finally, any conditioning manoeuvre which leaves spindles in a slightly altered mechanical state is likely to have a large effect on reflex size. All of these factors emphasize that great care must be taken when measuring reflexes and attributing the changes in size to a particular source.
References
- Burke D, Hagbarth K-E, Skuse NF. Voluntary activation of spindle endings in human muscles temporarily paralysed by nerve pressure. The Journal of Physiology. 1979;287:329–336. doi: 10.1113/jphysiol.1979.sp012662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crone C, Nielsen J. Methodological implications of the post-activation depression of the soleus H-reflex in man. Experimental Brain Research. 1989;78:28–32. doi: 10.1007/BF00230683. [DOI] [PubMed] [Google Scholar]
- Gandevia SC, Miller S, Aniss AM, Burke D. Reflex influences on muscle spindle activity in relaxed human leg muscles. Journal of Neurophysiology. 1986;56:159–170. doi: 10.1152/jn.1986.56.1.159. [DOI] [PubMed] [Google Scholar]
- Gregory JE, Mark RF, Morgan DL, Patak A, Polus B, Proske U. Effects of muscle history on the stretch reflex in cat and man. The Journal of Physiology. 1990;424:93–107. doi: 10.1113/jphysiol.1990.sp018057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gregory JE, Morgan DL, Proske U. After-effects in the responses of cat muscle spindles. Journal of Neurophysiology. 1986;56:451–461. doi: 10.1152/jn.1986.56.2.451. [DOI] [PubMed] [Google Scholar]
- Gregory JE, Morgan DL, Proske U. Changes in size of the stretch reflex of cat and man attributed to after-effects in muscle spindles. Journal of Neurophysiology. 1987;58:628–640. doi: 10.1152/jn.1987.58.3.628. [DOI] [PubMed] [Google Scholar]
- Hagbarth K-E, Nordin M. Postural after-contractions in man attributed to muscle spindle thixotropy. The Journal of Physiology. 1998;506:875–883. doi: 10.1111/j.1469-7793.1998.875bv.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagbarth K-E, Nordin M, Bongiovanni LG. After-effects on stiffness and stretch reflexes of human finger flexor muscles attributed to muscle thixotropy. The Journal of Physiology. 1995;482:215–223. doi: 10.1113/jphysiol.1995.sp020511. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hill DK. Tension due to interaction between the sliding filaments in resting striated muscle. The effect of stimulation. The Journal of Physiology. 1968;199:637–684. doi: 10.1113/jphysiol.1968.sp008672. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hultborn H, Illert M, Nielsen J, Paul A, Ballegaard M, Wiese H. On the mechanism of the post-activation depression of the H-reflex in human subjects. Experimental Brain Research. 1996;108:450–462. doi: 10.1007/BF00227268. [DOI] [PubMed] [Google Scholar]
- Joyce GC, Rack PMH, Westbury DR. The mechanical properties of cat soleus muscle during controlled lengthening and shortening movements. The Journal of Physiology. 1969;204:461–474. doi: 10.1113/jphysiol.1969.sp008924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katz R, Morin C, Pierrot-Deseilligny E, Hibino R. Conditioning of H reflex by a preceding subthreshold tendon reflex stimulus. Journal of Neurology, Neurosurgery and Psychiatry. 1977;40:575–580. doi: 10.1136/jnnp.40.6.575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan DL, Proske U, Gregory JE. Responses of primary endings of cat muscle spindles to locally applied vibration. Experimental Brain Research. 1991;87:530–536. doi: 10.1007/BF00227078. [DOI] [PubMed] [Google Scholar]
- Proske U, Morgan DL, Gregory JE. Muscle history dependence of responses to stretch of primary and secondary endings of cat soleus muscle spindles. The Journal of Physiology. 1992;445:81–95. doi: 10.1113/jphysiol.1992.sp018913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Proske U, Morgan DL, Gregory JE. Thixotropy in skeletal muscle and in muscle spindles: A review. Progress in Neurobiology. 1993;41:705–721. doi: 10.1016/0301-0082(93)90032-n. 10.1016/0301-0082(93)90032-N. [DOI] [PubMed] [Google Scholar]
- Táboríková H, Sax DS. Conditioning of H-reflexes by a preceding subthreshold H-reflex stimulus. Brain. 1969;92:203–212. doi: 10.1093/brain/92.1.203. [DOI] [PubMed] [Google Scholar]
- Vallbo ÅB. Muscle spindle response at the onset of isometric voluntary contractions in man. Time difference between fusimotor and skeletomotor effects. The Journal of Physiology. 1971;218:405–431. doi: 10.1113/jphysiol.1971.sp009625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson LR, Gandevia SC, Burke D. Discharge of human muscle spindle afferents innervating ankle dorsiflexors during target isometric contractions. The Journal of Physiology. 1997;504:221–232. doi: 10.1111/j.1469-7793.1997.221bf.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wise AK, Gregory JE, Proske U. Detection of movements of the human forearm during and after co-contractions of muscles acting at the elbow joint. The Journal of Physiology. 1998;508:325–330. doi: 10.1111/j.1469-7793.1998.325br.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood SA. 1997. Factors affecting the size of the phasic stretch reflex. PhD Thesis, Monash University. [Google Scholar]
- Wood SA, Gregory JE, Proske U. The influence of muscle spindle discharge on the human H reflex and the monosynaptic reflex in the cat. The Journal of Physiology. 1996;497:279–290. doi: 10.1113/jphysiol.1996.sp021767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood SA, Proske U, Gregory JE, Morgan DL. Fusimotor activity and the tendon jerk in the anaesthetised cat. Experimental Brain Research. 1994;98:101–109. doi: 10.1007/BF00229114. [DOI] [PubMed] [Google Scholar]