

Abstract
Aims
The aim of this study was to test the hypothesis that different conformations of misfolded α-synuclein (α-syn) are present in Parkinson’s disease (PD) brain.
Methods
Using two previously characterized conformations of α-syn fibrils, we generated new conformation-selective, α-syn monoclonal antibodies (mAbs). We then interrogated multiple brain regions in a well-characterized autopsy cohort of PD patients (n = 49) with these mAbs, Syn7015 and Syn9029.
Results
Syn7015 detects Lewy bodies (LBs) and Lewy neurites (LNs) formed by pathological α-syn in all brain regions tested, and is particularly sensitive to LNs and small Lewy dots, inclusions believed to form early in the disease. Further, we observed co-localization between Syn7015 and an early marker of α-syn pathology formation, phospho-Ser129-α-syn, and a lack of extensive co-localization with markers of more mature pathology. In comparison, Syn9029 detects Lewy pathology in all regions examined, but indicates significantly fewer LNs than Syn7015. In addition, co-localization of Syn9029 with later markers of α-syn pathology maturation (ubiquitin and P62) suggests that the pathology detected by Syn9029 is older. Semi-quantitative scoring of both LN and LB pathology in nine brain regions further established this trend, with Syn7015 LN scores consistently higher than Syn9029 LN scores.
Conclusions
Our data indicate that different conformations of α-syn pathology are present in PD brain and correspond to different stages of maturity for Lewy pathology. Regional analysis of Syn7015 and Syn9029 immunostaining also provides support for the Braak hypothesis that α-syn pathology advances through the brain.
Keywords: amyloid, conformation selective antibodies, Lewy pathology maturation, Parkinson’s disease
Introduction
Protein aggregates are the key neuropathological hallmarks in a variety of neurodegenerative diseases [1]. In Parkinson’s disease (PD), α-synuclein (α-syn), encoded by the gene SNCA, forms inclusions in axons and neuronal perikarya, referred to as Lewy neurites (LNs) and Lewy bodies (LBs), respectively [2–3]. SNCA duplication, triplication, and point mutations are linked to autosomal dominant PD and directly implicate misfolded α-syn as a causative agent in disease [3–5]. LN and LB α-syn pathology, referred to collectively as Lewy pathology (LP), shows a progressive complexity of morphological changes and post-translational modifications including phosphorylation, ubiquitination, and co-localization with autophagy adaptor protein P62 suggesting that α-syn aggregates mature over time [6–12].
α-Syn pathology appears to develop in a stereotypical, spatiotemporal pattern in the brain, with LN formation preceding LB formation [13–16], which has also been recapitulated in primary neuron culture models [17]. Spreading of pathological α-syn has been proposed as a potential mechanism for the progression of PD [18–20], and this hypothesis has been supported by the observation that embryonic mesencephalic neurons stably grafted into the basal ganglia of PD patients acquire a significant burden of pathological α-syn inclusions after ten years of implantation [21,22]. Transport and cell-to-cell transmission of α-syn aggregates has also been observed in primary neurons [23–25]. Additionally, neuron-based and animal models using in vitro generated α-syn preformed fibrils (PFFs) have recapitulated aspects of PD–like pathology and behavioral phenotypes [17, 26–29]. Importantly, in non-transgenic animals, stereotaxic injection of α-syn PFFs into the striatum led to the development of PD-like LP in interconnected neuronal populations followed by the death of substantia nigra pars compacta (SNPc) dopaminergic neurons and a PD-like motor phenotype. Thus, misfolded α-syn fibrils initiate and propagate pathology in cell and animal models.
We and others have shown that α-syn fibrils can adopt different conformations with distinct biophysical properties in cell-free systems and exhibit discrete phenotypes in cell-based and animal models [19, 20, 30–33]. We have previously generated unique α-syn PFF conformers (designated strains A and B) from recombinant human α-syn by repetitive self-seeded fibrillization in vitro [31]. Biochemical characterization of strain A and B PFFs suggests that the fibril cores compact differently, as partial digestion by proteinase K showed that the N and C termini of strain B fibrils are more susceptible to degradation than those of strain A fibrils. These altered conformations display distinct phenotypes in primary neuron cultures and in vivo animal model studies. For example, strain A and B PFFs can be distinguished by the extent of α-syn pathology induced (A>B) as well as the appearance of insoluble, tau inclusions induced by strain B, but not strain A. These stark phenotypic differences we observed in our model systems led us to ask if misfolded α-syn in LP in PD cases also adopt different conformations.
In our previous studies, we used strain B PFFs as an immunogen and generated a mAb, Syn9029 that is selective for the conformation of strain B fibrils [31]. Here, we used strain A PFFs as an immunogen and generated novel α-syn mAbs selective for the conformation of strain A fibrils, including Syn7015. Using these conformation-selective mAbs, we then investigated the heterogeneity of α-syn LP in the postmortem brains of patients with PD. Specifically, we systematically categorized the morphology of α-syn pathology and patterns of distribution in brainstem, limbic, and neocortical brain regions detected by our new mAbs. Our data support the existence of different α-syn conformations in PD LP that correspond to different maturation stages.
Materials & Methods
Recombinant α-syn purification and in vitro fibrillization
Protein constructs, 21–140 α-syn, 32–140 α-syn, 49–140 α-syn, 58–140 α-syn, 1–120 α-syn, and 1–130 α-syn, were generated through PCR site-directed mutagenesis with the PRK172/human wild-type α-syn plasmid as the template, expressed in transformed Eschericha coli BL21-(DE3)-RIL cells and purified as previously described [26, 27, 34, 35]. Fibrillization, characterization, and storage of the resultant fibrils was performed as previously described [31] and is depicted schematically in Figure 1a.
Figure 1.
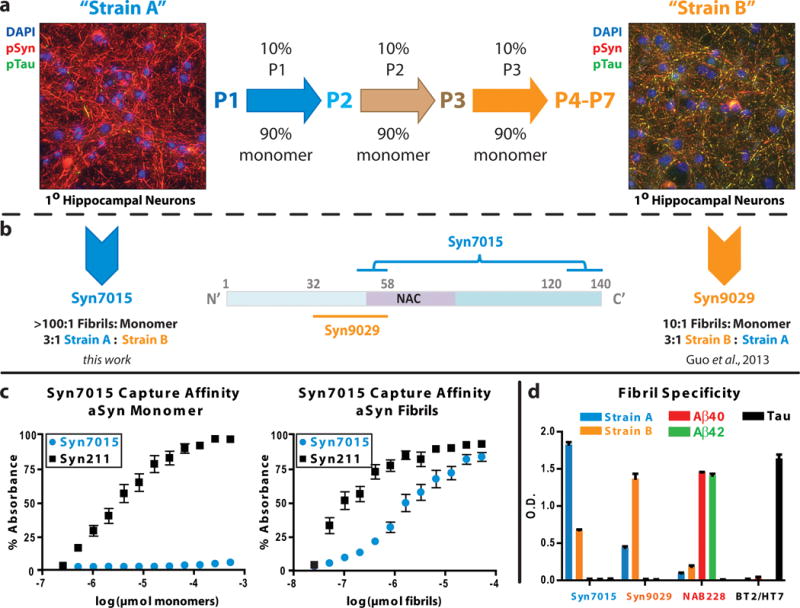
Generation of α-syn conforamtion selective mAbs. (a) De novo strain A (P1) PFFs induce extensive insoluble phospho-Ser129-α-syn (pSyn) positive aggregates (red) in wild-type mouse hippocampal neurons. Serially passaging (center) using 10% PFFs from the prior passage to seed monomeric α-syn generated strain B (P4-P7) α-syn fibrils, which induce both insoluble pSyn (red) and phosphorylated tau or pTau (green) in primary neuronal cultures (right). The aggregates are predominantly found in neuronal process where pSyn and pTau aggregates frequently co-localize (yellow). (b) Syn7015 recognizes a discontinuous epitope with binding elements in the extreme C-terminus and near amino acid 50 whereas Syn9029 recognizes a sequence between amino acid residues 32 and 58. The brackets connecting the two stretches of α-syn are intended to signify the discontinuous epitope recognized by Syn7015. (c) Syn7015 is highly selective for strain A fibrils over α-syn monomer when used as a capture antibody in sandwich ELISA, as compared to the control mAb, Syn211. (d) Comparison of the fibril binding affinity by direct ELISA of Syn7015 and Syn9029 for strain A and strain B α-syn fibrils, Aβ40 and Aβ42 fibrils, and tau fibrils. NAB228 and BT2/HT7 were used as Aβ and tau control mAbs.
Primary neuron cultures, fibril transduction, and immunocytochemistry
Primary neuron cultures were prepared from embryonic day 15–17 embryos of CD1 mice, transduced with either strain A or strain B PFFs, cultured for 18 days and then fixed and stained as previously described [17, 31].
α-Syn ELISA
A 384 well Nunc™ Maxisorp™ plate was coated overnight at 4°C with 50 ng of antibody, Syn7015 or Syn211 [36] and blocked using Block Ace™ solution (AbD Serotec, BUF209, BioRad, Hercules, CA, USA). PFFs were sonicated (10 min., 30 s on, 30 s off, 10°C, high setting, Diagenode Biorupter™, Denville, NJ, USA) and serial double dilutions of α-syn monomer and fibrils were made beginning at 256 and 25.6 ug/mL, respectively. Fibril concentrations were based on the monomer concentration used during assembly. Protein solutions were added to each well and incubated overnight at 4°C. Next, rabbit α-syn mAb MJF-R1 (1:3K; Abcam, Cambride, MA, USA) was added for 4 h at 37°C, followed by goat-anti-rabbit-IgG-HRP (1:10K; Cell Signaling Technology, Danvers, MA, USA) for 1 hour at room temperature. Antibody binding was detected using 1-Step Ultra TMB-ELISA™ Substrate Solution (Thermo Fisher Scientific, Waltham, MA, USA) and a Spectramax M5™ (Molecular Devices, Sunnyvale, CA, USA) plate reader according to the manufacturer’s instructions. Plates were washed in an automatic plate washer between each incubation step. Experiments were performed in triplicate and data are graphed in Figure 1c.
Selectivity for misfolded α-syn was further tested via competition ELISA based on the protocol in [37]. Optimal results were obtained using 5 ng of Syn7015, 20 ng Syn211, and 80 ng of Syn9029 and summarized in Figure S2e–f.
α-Syn Antigen Preparation
Strain A PFFs were generated as described previously [31] (Figure 1a). In PBS, 36.5 μM (0.5 mg/mL) strain A PFFs were sonicated and treated with 0.01% v/v gluteraldehyde for 30 minutes to achieve minimal cross-linking. The reaction was stopped with 1 M Tris, 1 M sodium chloride, pH 7.4. Cross-linking was optimized to eliminate the α-syn monomer band when samples were evaluated by SDS-PAGE (Figure S1a), without disrupting the epitopes of a panel of known α-syn antibodies by dot blot (2 μg protein/dot; Figure S1b) or the ability of the PFFs to seed aggregation of monomeric α-syn.
Anti-α-Syn Antibody Generation and Selection
Animal care and all procedures performed were conducted in accordance with the NIH Guide for the Care and Use of Experimental Animals and approved by the University of Pennsylvania Institutional Animal Care and Use Committee. Murine mAbs were raised against sonicated, cross-linked α-syn stain A PFFs as previously described [31, 36, 38, 39]. Hybridoma supernatants were screened by direct ELISA. Supernatants with selectivity for aggregated forms of α-syn were further evaluated by immunohistochemistry (IHC) and positive hybridomas were expanded and sub-cloned at least twice. Subcloned mAbs were tested as reporters in a direct ELISA for selectivity for Strain A and B α-syn PFFs, amyloid-β fibrils, and tau fibrils (Figures 1d and S2).
Epitope Mapping
Approximately 250 ng/lane of full-length mouse wild-type α-syn, full length human wild-type α-syn, 21–140 α-syn, 32–140 α-syn, 49–140 α-syn, 58–140 α-syn, 1–120 α-syn, and 1–130 α-syn were run via SDS-PAGE on a 5–20% polyacrylamide gradient gel, transferred to a nitrocellulose membrane and probed with Syn7015 (1:1K) and a polyclonal control Ab (HuA) [36]. Representative blots and quantification are shown (Figure S2a–c). Because Syn9029 works poorly via Western blot, its approximate epitope was ascertained via direct ELISA as previously described [31]. Epitope mapping experiments were run in triplicate (Figure S2d) and summarized schematically in Figure 1b (center).
Studied Subjects
Donor brains subjects following neuropathological evaluation were selected from the brain bank at the Center for Neurodegenerative Disease Research at the University of Pennsylvania [39] based on previously published criteria [40] based on previously published criteria [41–43]. Informed consent for autopsy was obtained for all patients from their next of kin and this study was approved by the University of Pennsylvania Institutional Review Board. Forty-nine cases with diagnosed PD according to the U.K. Brain Bank Criteria [44] were evaluated immunohistochemically. Age of disease onset, age at death, and years of education were abstracted from the medical record by an expert neurologist in movement disorders (RSA).
Semi-quantitative IHC
Immunohistochemistry was performed as described previously [40]. Regions examined were a section of the medulla with dorsal motor nucleus of vagus, a midbrain cross-section containing the SNPc and coronal sections of the amygdala including the parahippocampal cortex, the hippocampus at the level of the lateral geniculate nucleus including CA2/3, the entorhinal cortex, the anterior cingulate at the level of the striatum, the superior temporal gyrus from the same coronal section as the hippocampus, and the middle frontal gyrus obtained from the coronal section anterior to the striatum. Each region was assessed for the level of Alzheimer’s disease (AD) neuropathological change [43] and for Braak PD stage [13], according to established criteria as reviewed elsewhere [40].
Syn7015 and Syn9029 mAbs were characterized in ethanol- and formalin-fixed tissue with and without two common antigen retrieval methods (Figure S3). In ethanol-fixed tissue there was no significant gain or alteration of signal by antigen retrieval. Therefore, ethanol-fixed tissue from each PD case was examined using Syn7015 (1:100K), Syn9029 (1:9K) and our standard diagnostic mAb Syn303 [38] (1:60K; Cat# 824301, BioLegend, San Diego, CA, USA) without any antigen retrieval. We examined the brain region described for LBs using semi-quantitative criteria as previously described [40]. LNs; including thick neurites, threads, and dots [6, 14, 16], were scored according to the following inclusion density: 1–2 per region = “rare”; 2–4 per 10× field = 1 (mild); 5–9 = 2 (moderate), and 10+ = 3 (severe). Antibody reproducibility was verified via IHC and immunofluorescence on 8–10 sections from a variety of brain regions across five different brains. LP was quantified separately by investigators blinded to all clinical data and agreement between reviewers was assessed by Spearman correlation and found to be 0.7202 (Syn303 LN), 0.7875 (Syn303 LB), 0.7414 (Syn7015 LN), 0.6511 (Syn7015 LB), 0.7578 (Syn9029 LN) and 0.7974 (Syn9029 LB), all Ps < 0.0001.
Double-labeling Immunofluorescence
Double labeling immunofluorescence of Syn7015 with Syn9029 and of both mAbs Syn7015 and Syn9029 with 81A (phospho-Ser129; Cat# 825701, BioLegend), anti-ubiquitin (Cat# MAB1510, EMD Millipore, Darmstadt, Germany), and anti-P62 (Cat# 8878-M01, Abnova, Neihu District, Taipei City, Taiwan) in PD tissue was conducted as previously described [31, 39] using Alexa Fluor 488- and 594-conjugated secondary Abs (Thermo Fisher Scientific, Waltham, MA, USA). Sections were treated for autofluorescence with Sudan black solution and cover-slipped with Vecta-shield-DAPI mounting medium (Vector Laboratories, Burlingame, CA, USA). For co-labeling experiments, mouse mAbs Syn7015 and Syn9029 were targeted with IgG-specific secondary Abs (Alexa-594 IgG2b and Alexa-488 IgG3, respectively).
Statistical Analyses
Fisher’s exact test or Wilcoxon-Mann-Whitney test was used to compare categorical or ordinal variables between clinical groups. Two-sample t-test or one-way ANOVA was similarly used to compare linear clinical variables.
For comparisons of semi-quantitative α-syn pathology scores between antibodies, we applied Kruskal-Wallis test, followed by post hoc Wilcoxon signed rank sum test to determine significant antibody pair differences. Each brain region was analyzed separately and P-values were corrected for multiple comparisons. Because LB and LN scores are not directly comparable to one another, each pathology score was analyzed separately. Statistical calculations were performed using Stata/IC (13.1, College Station, TX, USA). Figures were prepared with Adobe Illustrator™ (CS2, San Jose, CA, USA) and GraphPad Prism (6.0, La Jolla, CA, USA).
Results
Generation and characterization of anti-α-syn conformation-selective mAbs
When α-syn monomer is aggregated de novo to form PFFs, the first passage (P1) fibrils efficiently recruit endogenous α-syn to misfold and accumulate as phosphorylated pathological aggregates when added to wild-type mouse hippocampal neuron cultures (Figure 1a, left) [17]. Subsequent fibrillization initiated with 5–10% of P1 PFFs as seeds generated second passage (P2) fibrils, and, after several iterations of this sequential seeding paradigm (Figure 1a, center), a new conformation of PFFs emerged at passages 4–7 (P4-P7) that simultaneously induced both α-syn and tau pathology (Figure 1a, right). We hereafter refer to P1 PFFs as strain A and P4-P7 PFFs as strain B [31].
Using strain B PFFs as an immunogen, we generated a mAb, Syn9029, that binds preferentially to strain B fibrils (~3:1 B:A, Figure 1b, right, 1d) [31]. To generate mAbs selective for the strain A fibrils, we immunized BALB/c mice with strain A PFFs and produced a number of high affinity mAbs. One mAb, Syn7015 (Figure 1b, left), had excellent selectivity for fibrillar α-syn over monomer (Figure 1c). Epitope mapping of Syn7015 revealed a complex, discontinuous epitope with the C-terminal sequence near amino acid 130 being essential for binding and the more N-terminal sequence near amino acid 49–58 enhancing mAb binding (Figures 1b, center; and S2a–c). In comparison, Syn9029 had an N-terminal epitope between amino acid 32–58 that is partially masked by the C-terminal sequence beyond amino acid 120 (Figures 1b, center; and S2d). Syn7015 binds strain A PFFs in preference to strain B PFFs (~3:1 A:B; Figure 1d) and both mAbs are selective for α-syn in preference to Aβ40, Aβ42, or tau (Figure 1d). Preference for α-syn fibrils for these new mAbs was further examined by competition ELISA (Figure S2e–f). A large excess of monomeric α-syn was required to inhibit binding of Syn7015 to strain A PFFs (EC50 = 37.2 nM) as compared to Syn211 (EC50 = 444.7 pM). Similarly, a large excess of α-syn monomer was required to inhibit Syn9029 binding to strain B PFFs (EC50 = 868.5 nM) as compared to Syn211 (EC50 = 1.7 nM).
Performance of conformation-selective α-syn mAbs by IHC in human brain tissue
We next sought to characterize these new mAbs in human brain samples from normal controls and PD patients. For comparison, adjacent sections were stained with Syn7015, Syn9029, and the previously described α-syn mAb, Syn303 [38]. We did not observe Syn7015 or Syn9029 immunoreactivity in healthy control brains. In contrast, Syn303 detected normal monomeric, synaptic α-syn present in the neuropil (Figure S4). In brain regions believed to be affected early in the disease course, each mAb displayed a strong preference for pathological accumulations of α-syn (Figure 2) and both Syn7015 and Syn9029 detected prominent LB (Figure 2, black arrowheads) and LN pathology, the latter subdivided into thick LNs (Figure 2, black double-arrows), threads (Figure 2, red arrowheads), and dots (Figure 2, red double-arrows). Large LBs (black arrowheads) and thick LNs (black double-arrows) were prominent in the dorsal motor nucleus of the vagus (Figure 2a–c) and substantia nigra (Figure 2d–f). In the amygdala, in addition to LBs and thick LNs, pathological threads (red arrowheads) and dots (red double-arrows) accumulated in large quantities (Figure 2g–i). Though all three mAbs were able to detect the various types of LP, we noticed that Syn7015 stained more LN pathology than the other mAbs in the substantia nigra and amygdala.
Figure 2.

Subcortical Lewy body (LB) and Lewy neurite (LN) pathology as detected by α-syn conformation-selective antibodies. Syn303, Syn7015 and Syn9029 each have a strong preference for pathological α-syn in (a–c) the dorsal motor nucleus of the medulla (d–f) the substantia nigra pars compacta, and (g–i) the central nucleus of the amygdala. Images are taken from serial sections. Bar = 100 μm. Representative LBs (black arrowheads), thick LNs (black double-arrows), thread LNs (red arrowheads), and dot LN pathology (red double-arrows) are shown.
In hippocampal and cortical regions of a small cohort of PD patients, our conformation-selective mAbs showed more divergent staining properties (Figure 3). While Syn7015 and Syn303 detected relatively similar degrees of LBs and LNs in the entorhinal cortex (Figure 3a,b), Syn9029 labeled only a subset of the pathological α-syn, mostly LBs, in this region (Figure 3c). LP in the hippocampus CA2 region primarily takes the form of a plexus of LNs readily seen by Syn303 (Figure 3d), but only a small fraction of this pathology was detected by Syn9029 (Figure 3f). In contrast, Syn7015 detected LNs in the CA2 in greater abundance than Syn303, particularly threads (red arrowheads) and dots (red double-arrows; Figure 3f). In the anterior cingulate gyrus, Syn303 and Syn9029 readily detected LBs and rare LNs (Figure 3g, i), whereas Syn7015 stained both LBs and an abundance of LNs not labeled by the other mAbs (Figure 3h, red arrow heads and red double arrows).
Figure 3.

Cortical Lewy body (LB) and Lewy neurite (LN) pathology as detected by α-syn conformation-selective antibodies. (a–c) Syn7015 reveals significantly more LNs than Syn303 or Syn9029 in the hippocampus CA2. (d–f) In the entorhinal cortex on the same section, LBs and LNs are readily detected by Syn303 and Syn7015, while Syn9029 stains LBs, but not LNs. (g–i) Anterior cingulate gyrus is frequently filled with LNs, including thread and dot LNs, as indicated by staining with Syn7015 in addition to the severe pathology already detected by Syn303. Syn9029 pathology is limited to a subset of the LBs and LNs. Images are taken from serial sections. Bar = 100 μm. Representative LBs (black arrowheads), thick LNs (black double-arrows), thread LNs (red arrowheads), and dot LN pathology (red double-arrows) are shown.
Because these findings suggested that (i) Syn7015 labeled a population of LNs not detected by the other mAbs in addition to the LP detected by all three mAbs and (ii)) Syn9029 preferentially recognized LBs, we performed a dilution series with all three antibodies on adjacent sections of anterior cingulate cortex to rule out the effect of concentration on the LP detected (Figure S5). Syn7015 retained the ability to detect both LBs and LNs at progressively lower antibody concentrations (Figure S5 e vs k). Syn9029 showed a strong preference for staining LBs even at the highest concentration tested (Figure S5c), and both Syn303 and Syn9029 detected LBs in preference to LNs as the concentration of mAb was reduced (Figure S5 d vs j and f vs l). Activity against pathological α-syn in tissue could be eliminated by pre-adsorption of Syn7015 or Syn9029 with recombinant α-syn PFFs (Figure S6a, g). Consistent with the selectivity of these antibodies, Syn7015 was more efficiently neutralized by strain A PFFs and Syn9029 preferentially blocked by strain B PFFs (Figure S6a, g). Additionally, other strain A selective mAbs were able to stain LBs and extensive amounts of LNs (e.g. Syn7020 and Syn7068 in Figure S7). To extend these observations, we performed double-labeling immunofluorescence in the substantia nigra, entorhinal cortex, hippocampus CA, and anterior cingulate, and found that Syn7015 detects abundant, small LN pathology that is not labeled by Syn9029 (Figure S8, white arrows). Conversely, the majority of thicker LNs and LBs showed good co-localization between the two mAbs (Figure S8, white arrowheads). Collectively, these data suggest that Syn7015 detects a form of pathological α-syn common throughout all types of LP, including the smallest and therefore presumably earliest LNs. Conversely, Syn9029 appears to identify a conformation of aggregated α-syn primarily found in thicker LNs and LBs, which are traditionally thought of as being more mature inclusions.
Conformational change during maturation of pathological α-syn inclusions in PD
Based on our initial observations of the properties of Syn9029 and Syn7015, we hypothesized that Syn7015 detects a conformation of pathological α-syn that is present in the earliest α-syn pathology, whereas Syn9029 detects a more mature species of LP. To test this hypothesis, we looked for co-localization of LP and other markers of morphological maturation of α-syn pathology (e.g. pSyn, ubiquitin, and P62) [7–9, 12]. All Syn7015-positive pathology co-localized with pSyn, including the thread and dot type LNs that did not co-localize with Syn303 (Figure 4a). Syn9029 also completely co-localized with pSyn (Figure 4b). Ubiquitin co-localized with both Syn7015 (Figure 4c) and Syn9029 (Figure 4d) in LBs, but only rarely in LNs. P62 only weakly co-localized with Syn7015 labeled LBs (Figure 4e), but often showed extensive co-localization with LBs stained by Syn9029 (Figure 4f). These results are summarized in Figure 5a and suggest that α-syn confirmation changes as the morphology of LP matures from small, Syn7015 and pSyn positive dots and threads into larger LNs and LBs that remain positive not only for pSyn, and Syn7015, but are also positive for Syn9029, ubiquitin, and P62 (Figure 5b).
Figure 4.

Specific post-translational markers of Lewy pathology maturity variably co-localize with α-syn conformation-selective antibodies. pSyn pathology co-localized with (a) Syn7015 positive thread and dot LNs and LBs and (b) Syn9029 LNs and LBs. Ubiquitination is limited to thick LNs and LBs and rarely co-localizes with (c) Syn7015 thread or dot LN pathology, while frequently co-localizing with (d) Syn9029 pathology. P62 is seen in a subset of α-syn LB pathology, a subset that is rarely (e) Syn7015 positive, but is (f) Syn9029 positive. Bar = 100 μm
Figure 5.

Conformation-selective antibodies highlight α-syn pathology in early and late stages of Lewy pathology (LP) maturation. (a) Conformational mAbs Syn7015 and Syn9029 see pathological α-syn inclusions at different maturational stages in LP evolution or formation, based on immunofluoresence observations in PD brain. (b) Proposed LP maturation model, in which pSyn pathology is first readily detected by Syn7015 in longitudinally transversely sectioned axonal processes. Subsequently, a portion of these α-syn inclusions undergo a conformational compaction into denser structures including thick Lewy neurites and Lewy bodies, in neuronal perikarya that become detectable by Syn9029, as well as by antibodies to ubiquitin and P62.
Conformation-selective antibodies detect progressive pathological α-syn maturation in PD brains
Because Syn7015 and Syn9029 could distinguish α-syn inclusions of different maturity, we hypothesized that these antibodies would demonstrate pathological changes in misfolded α-syn conformation within different brain regions and stages of PD. To test this hypothesis, we examined a well-characterized autopsy cohort of 49 PD patients with these conformation-selective α-syn mAbs (Table 1). Forty-one (84%) of the cases were male. Average years of education was 16.4 (SD 2.1) years, average age at disease onset was 63.7 (SD 9.2) years, and average disease duration was 13.6 (SD 5.8) years. The majority (75.5%) of patients had neocortical α-syn pathology (Braak PD stage 5–6; Table 1). Twelve, nine, and five PD cases met criteria for low, intermediate, or high likelihood of AD neuropathological change [43]. The remaining 23 cases did not have concomitant AD neuropathological change.
Table 1.
Demographic and clinical features of all cases studied
| Disease Group | Parkinson‘s disease | Alzheimer’s disease |
|---|---|---|
| N | 49 | 8 |
| Gender, male : female | 41 : 8 | 5 : 3 |
| Education, years | 16.4 (2.1) | 15.3 (3.5) |
| Age of disease onset, years | 63.7 (9.2) | 71.2 (3.9) |
| Age at death, years | 77.4 (7.3) | 80.5 (4.7) |
| Disease duration, years | 13.6 (5.8) | 10.3 (3.3) |
| Braak Parkinson’s disease stagea, n. None |
0 | 8 |
| 3 | 1 | -- |
| 4 | 11 | -- |
| 5 | 14 | -- |
| 6 | 23 | -- |
| Thal Phase, n. None |
23 | 0 |
| 1 / 2 | 9 | 0 |
| 3 | 5 | 2 |
| 4 / 5 | 12 | 6 |
| Braak tau stage, n. | 2 | 0 |
| None | ||
| I / II | 27 | 0 |
| III / IV | 14 | 0 |
| V / VI | 6 | 8 |
All data are mean (SD) unless otherwise specified.
Stages are 0–6. The Alzheimer’s disease cases are co-morbid amygdala-predominant variant of Lewy pathology.
-- = not applicable
We examined nine regions in each PD case for LNs and LBs, using the mAbs Syn303, Syn7015, and Syn9029. Detailed semi-quantitative scores for all cases examined are shown in Table S1. Semi-quantitative LN scores, as determined by each individual antibody, were significantly different in all brain regions examined except for the middle frontal cortex (Table S2). Specifically, when comparing LN pathological severity as determined by each antibody, Syn7015 detected significantly higher burdens of LN, followed by Syn303, followed by Syn9029, in almost all regions. In contrast, LB pathology burden was equally detected by all three antibodies in almost all regions examined (Table S2). In the amygdala, Syn9029 detected less LB pathology than either Syn303 or Syn7015. Frequency plots for LN and LB semi-quantitative scores for the substantia nigra and amygdala are shown in Figure 6a, b. Taken together, these results support our earlier observation that LN pathologies are differentially detected by our new α-syn conformation-selective antibodies (Syn7015>Syn303>Syn9029), whereas LB pathology is detected more equivalently.
Figure 6.

Distribution plots of semi-quantitative scores demonstrate a preferential staining of Lewy neurites (LNs) by Syn7015 that is maintained throughout the disease course. Semi-quantitative Lewy body (LB) and LN scores for Syn303, Syn7015, and Syn9029 in the (a) substantia nigra and (b) amygdala show equivalent distribution for LBs but not LNs. Distribution plots of semi-quantitative LB and LN scores in the anterior cingulate gyrus for patients classified as (c) Braak PD stage 4, (d) Braak PD stage 5 (e) and Braak PD stage 6 suggest the relative patterns of LB and LN staining are preserved throughout disease progression.
Next, we sought to determine if α-syn pathology varied with respect to changes in Braak PD stage. Eleven, 14, and 23 of our cases met criteria for Braak PD stages four, five, and six. When cases were grouped by Braak stage, each antibody detected similar amounts of LB pathology in all regions examined (eg. Figure 6c–e, left). LN scores in the substantia nigra were predominantly severe and did not vary substantially when patients were organized by Braak PD stage (Figure S9a). In contrast, a progressive accumulation of LNs between Braak PD stages 4 and 6, detected most prominently by Syn7015, was observed in the amygdala, parahippocampal cortex, entorhinal cortex, hippocampus CA, and anterior cingulate gyrus (Figure 6c–e, right, Figure S9b–d). A similar trend was also observed between Braak PD stages 5 and 6 in the middle frontal cortex and superior temporal cortex (Figure S9e). This suggests that the pattern of more severe LN pathology seen by Syn7015 as compared to that stained by Syn303 and especially Syn9029 continued as the overall Braak PD stage advanced.
Pathological α-syn LP in amygdala-predominant AD
Although LP is diagnostic of PD in the appropriate clinical context, LP is also a common co-morbidity in AD, with a subset of AD patients having LP confined to the amygdala [45]. We hypothesized that if Syn7015 pathology is less mature than Syn9029 pathology, Syn7015 pathology should be greater than Syn9029 pathology in AD cases with amygdala-predominant LP. To test this hypothesis and determine if our conformation-selective mAbs detected a similar pattern of LP in AD to that observed in PD, we examined the affected brain regions of eight AD cases with amygdala-predominant LP (Table 1). Both Syn303 and Syn7015 identified moderate to severe burdens of LN in the amygdalae of these cases (Figure 7a, b), in contrast to Syn9029 (Figure 7c). Specifically, the LN pathology detected by the three mAbs in the amygdala was significantly different in these cases (χ22 = 12.41, P = 0.002). This difference was attributable to Syn9029, which detected significantly lower amounts of LN pathology than Syn303 (z = 2.47, P = 0.014) or Syn7015 (z = 2.47, P = 0.013). Conversely, LB pathology was equally detected by all three mAbs in the amygdala (χ22 = 5.42, P = 0.067). These findings suggest that, similar to our findings in PD, the conformation of α-syn pathology in LNs could appear before that of the more mature α-syn pathology stained by Syn9029 in amygdala-predominant AD.
Figure 7.

Syn7015 detects additional pathological burden as compared to Syn9029 in Alzheimer’s disease (AD) with amygdala predominant Lewy pathology (LP). In AD cases with amygdala-predominant LP, α-syn Lewy body (LB) and Lewy neurite (LN) pathology is abundant and recognized by (a) Syn303 and (b) Syn7015, but (c) Syn9029 stains only LBs. Bar = 100 μm.
Discussion
Different conformations of in vitro α-syn fibrils, derived through a serial seeding paradigm, have distinct biochemical and biological properties [31]. In order to further define the role of pathological α-syn conformations in PD, here we developed and characterized conformation-selective mAbs for α-syn that display not only a strong preference for aggregated α-syn PFFs over monomer, but also showed modest selectivity for de novo strain A fibrils. The mAb Syn7015 preferentially binds early passages (P1-P2) of α-syn PFFs, whereas our previously reported strain B-selective mAb, Syn9029 shows a predilection for later passages (P4-P7; Figure 1d). We identified the epitope of Syn7015 as discontinuous with a dependence on both the extreme C-terminus and a stretch of amino acids near position 50 of the α-syn sequence. The epitope for Syn9029 was found to be between amino acid 32 and 58, with removal of the extreme C-terminus enhancing mAb binding. These empirically determined epitopes support the conformational differences previously observed for strain A and strain B PFFs, in which the termini of α-syn are more exposed to proteinase K digestion in strain B PFFs [31].
Characterization of our novel mAbs biochemically and in PD tissue demonstrated that Syn7015 and Syn9029 see overlapping populations of misfolded α-syn LP, and that the LP readily identified by Syn7015 appears to include a less mature form than that detected by Syn9029. Syn9029 shows a strong preference for thick LNs and LBs that are ubiquitinated and often co-localize with P62. In contrast, Syn7015 detects a greater population of α-syn pathology than Syn9029, particularly small LNs and Lewy dots. These results are consistent with the de novo strain A PFF antigen used to generate this mAb being in a conformation that is easily accessible. An abundance of LN pathology detected by Syn7015 in the amygdala, parahippocampal cortex, entorhinal cortex, hippocampus CA, anterior cingulate gyrus, middle frontal cortex and superior temporal cortex of patients with more advanced Braak PD stage suggests that this difference persists throughout the disease course. This finding is in agreement with previous work that shows that LNs are the first pathology described in the progression from early to late stage PD [13], which is supported by studies of in vitro models of pathological α-syn templated propagation wherein strain A PFFs are more potent than strain B PFFs [17]. Moreover, this data suggests that the conformation of pathological α-syn found in these small LNs could be a readily transmissible one. This hypothesis remains to be tested in a larger autopsy cohort, due to the relatively low number of patients in each Braak PD stage and in animal studies where disease progression can be more easily followed longitudinally. We observed a similar phenomenon in AD patients with amygdala-predominant LP. This supports a hypothesis that a conformation of α-syn pathology, labeled efficiently by Syn7015, is the predominant form in the earliest stages of deposition in multiple neurodegenerative diseases.
Collectively, our neuropathological observations match well with the maturation seen in the in vitro generation of strain B fibrils by serial passaging from strain A fibrils [31]. Combining findings from proteinase K digestion, epitope mapping, IHC, and biochemical/biophysical studies, our data suggest a model for α-syn conformational evolution in PD (Figure 5). Misfolding of α-syn first forms smaller, loosely packed structures in which the C-terminus is near the central hydrophobic domain in dots and thin LNs. As thicker LNs and LBs are formed within the neuron, a portion of the misfolded α-syn is converted to more densely compacted species with the termini efficiently excluded from interacting with the amyloid core. Our data also suggest this conformational conversion may be facilitated by recruitment of intracellular machinery responsible for recycling/refolding aggregated proteins, including ubiquitin and P62, consistent with previous descriptions of morphological maturation [6–12].
Other conformational mAbs for α-syn have been recently reported by Vaikath et al. that show selectivity for misfolded species over normal monomer in vitro [46]. Their mAbs were found to have discontinuous epitopes that showed some recognition of a C-terminal fragment of α-syn. Furthermore, the authors describe abundant LN staining, including “micro-aggregates”, that are also pSyn positive. These features are consistent with our characterization of Syn7015, suggesting that these antibodies may be recognizing similar conformers. Based on the analysis of binding to species present at different stages of in vitro α-syn fibrillization, the authors concluded that their mAbs are recognizing “late” α-syn aggregates. Our results here suggest that α-syn aggregation in vivo may have several additional stages of maturation as compared to the process in vitro. Further work to develop tools and methods that clarify the roles of different α-syn structures and conformations throughout disease pathogenesis are necessary.
Structural studies of α-syn fibrils to date have focused mainly on the core residues of α-syn as the termini often fail to adopt well-organized structure [47–50]. However, associations of the C-terminus of α-syn monomer to the NAC domain have been observed spectroscopically [51, 52], and supported experimentally by findings that show an increased rate of aggregation in vitro for C-terminal α-syn truncation mutants [53, 54]. Such an interaction in the fibrillar form may explain the increased potency in vitro of strain A PFFs if it destabilizes fibril packing and enables more efficient fibril shearing and/or dissociation of small, highly potent α-syn fragments. If a subset of the fibrils within thicker LNs and LBs compact into the conformation recognized by Syn9029, toxicity of strain A fibrils might be mitigated. This process would explain the reduced potency of strain B PFFs in vitro. Post-translational modifications like ubiquitination or changes in intracellular compartmentalization may also contribute to this transition in vivo. Neuronal cultures expressing endogenous levels of both α-syn and tau suggest that these more mature structures could seed other amyloidogenic proteins [31]. This model of α-syn seeding tau may be particularly relevant to Parkinson Disease where the LP deposits earlier – and in regions unaffected by – tau pathology.
Our study has several limitations. First, we used a semi-quantitative score which might over- or under-estimate pathological burden, although this approach has been validated in inter-site comparisons of neuropathological assessment [42, 43]. More quantitative methods might be able to measure sub-populations of α-syn inclusions, including LNs, similar to the methods developed for neuritic tau pathology [55] or dystrophic neurites made up of TDP-43 [56]. In addition, the distinct staining patterns produced by our novel mAbs are potentially subject to scoring bias, although our observers were blinded to the case history and scores were aggregated for analysis. Furthermore, only autopsy-confirmed PD patients were selected for the bulk of this analysis and only a subset of brain regions were studied, which could lead to sampling bias. These regions are diagnostic for PD and Braak staging, and therefore these regions are justifiable for characterizing novel α-syn mAbs in this exploratory study. Finally, future studies characterizing α-syn conformers isolated from disease brains will provide important insights on the pathogenesis of α-synucleinopathies.
Collectively, our results support the hypothesis that the conformation of α-syn within LP is changing alongside gross morphological maturation. This finding has potentially important implications for therapeutic agents that target α-syn inclusions. Additional study of the pathological species selectively detected by mAbs like Syn7015 and Syn9029 will improve our understanding of the processes underlying PD progression as well as provide novel targets for biomarker assays and disease modifying therapies.
Supplementary Material
Figure S1. Characterization of antigen used for generation of strain A selective mAbs.
Figure S2. Additional characterization of strain-selective mAbs Syn7015 and Syn9029
Figure S3. Immunohistochemistry (IHC) optimization of Syn7015 and Syn9029 in ethanol and formalin fixed tissue.
Figure S4. Immunohistochemistry (IHC) staining of neuropathologically normal tissue with Syn303, Syn7015, and Syn9029.
Figure S5. Immunohistochemistry (IHC) of Syn303, Syn7015 and Syn9029 mAb dilution series in near serial sections of PD cingulate cortex.
Figure S6. Syn7015 and Syn9029 immunostaining can be removed by serial adsorption with strain A and B PFFs.
Figure S7. Immunohistochemistry (IHC) shows that additional 7000 series mAbs, as exemplified by Syn7020 and Syn7068, also prominently stain thread and dot Lewy neurites (LNs).
Figure S8. Double-labeling immunofluorescence of Syn7015 and Syn9029 co-localizes primarily in Lewy bodies (LBs) and thick Lewy neurites (LNs).
Figure S9. Semi-quantitative scores showing that preferential detection of Lewy neurites (LNs) by Syn7015 is maintained throughout the disease course in multiple brain regions.
Table S1. Demographic information and semi-quantitative scores for all patients in this study
Table S2 Comparative ability of α-syn strain selective antibodies to detect Lewy neurites (LNs) and Lewy bodies (LBs) in Parkinson’s disease brain tissues
Acknowledgments
The authors would like to thank Linh Le, Susan Leight, and Kayla Waits for their technical assistance, Eric Lin for his pilot immunohistochemical study, and Drs. Kurt Brunden and Eddie Lee for their helpful discussion. This work was supported by NIH/NIA training grant (T32-AG000255), the NIH/NINDS Morris K. Udall Parkinson’s Disease Center of Excellence (P50 NS053488), the Michael J. Fox Foundation, the Keefer family, the Parkinson Council, and by a Parkinson’s Disease Foundation Summer Student Fellowship (PDF-SFW-1562). RSA is supported by the NIH (K08-NS093127).
Footnotes
Supporting information
Additional Supporting Information may be found in the online version of the article at the publisher’s website:
“This is the peer reviewed version of the following article: “Novel conformation-selective alpha-synuclein antibodies raised against different in vitro fibril forms show distinct patterns of Lewy pathology in Parkinson’s disease” Neuropathol Appl Neurobiol. 2017, which has been published in final form at DOI: 10.1111/nan.12402. This article may be used for non-commercial purposes in accordance with Wiley Terms and Conditions for Self-Archiving.”
Author Contributions
DJC designed the study, performed experiments, analyzed the data and wrote the manuscript; RSA, MG, and DW analyzed data and reviewed the manuscript; JLR, HMB, RMP, DR and AY acquired and analyzed data and reviewed the manuscript; VM-YL supervised and designed the study; VM-YL and JQT analyzed data and were involved in writing of the manuscript.
Conflict of Interest
All authors declare that they have no conflict of interest.
References
- 1.Forman MS, Trojanowski JQ, Lee VMY. Neurodegenerative diseases: a decade of discoveries paves the way for therapeutic breakthroughs. Nat Med. 2004;10:1055–63. doi: 10.1038/nm1113. [DOI] [PubMed] [Google Scholar]
- 2.Braak H, Sandmann-Keil D, Gai W, Braak E. Extensive axonal Lewy neurites in Parkinson’s disease: a novel pathological feature revealed by alpha-synuclein immunocytochemistry. Neurosci Lett. 1999;265:67–9. doi: 10.1016/s0304-3940(99)00208-6. [DOI] [PubMed] [Google Scholar]
- 3.Spillantini MG, Schmidt ML, Lee VMY, Trojanowski JQ, Jakes R, Goedert M. alpha-Synuclein in Lewy bodies. Nature. 1997;388:839–40. doi: 10.1038/42166. [DOI] [PubMed] [Google Scholar]
- 4.Breydo L, Wu JW, Uversky VN. α-Synuclein misfolding and Parkinson’s disease. Biochim Biophys Acta. 2012;1822:261–85. doi: 10.1016/j.bbadis.2011.10.002. [DOI] [PubMed] [Google Scholar]
- 5.Luna E, Luk KC. Bent out of shape: α-Synuclein misfolding and the convergence of pathogenic pathways in Parkinson’s disease. FEBS Lett. 2015;589:3749–59. doi: 10.1016/j.febslet.2015.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Kanazawa T, Adachi E, Orimo S, Nakamura A, Mizusawa H, Uchihara T. Pale neurites, premature α-synuclein aggregates with centripetal extension from axon collaterals. Brain Pathol. 2012;1:67–78. doi: 10.1111/j.1750-3639.2011.00509.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kuusisto E, Parkkinen L, Alafuzoff I. Morphogenesis of Lewy bodies: dissimilar incorporation of alpha-synuclein, ubiquitin, and p62. J Neuropathol Exp Neurol. 2003;62:1241–53. doi: 10.1093/jnen/62.12.1241. [DOI] [PubMed] [Google Scholar]
- 8.Machiya Y, Hara S, Arawaka S, Fukushima S, Sato H, Sakamoto M, Koyama S, Kato T. Phosphorylated alpha-synuclein at Ser-129 is targeted to the proteasome pathway in a ubiquitin-independent manner. J Biol Chem. 2010;285:40732–44. doi: 10.1074/jbc.M110.141952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wales P, Pinho R, Lázaro DF, Outeiro TF. Limelight on alpha-synuclein: pathological and mechanistic implications in neurodegeneration. J Parkinsons Dis. 2013:415–59. doi: 10.3233/JPD-130216. [DOI] [PubMed] [Google Scholar]
- 10.Fujiwara H, Hasegawa M, Dohmae N, Kawashima A, Masliah E, Goldberg MS, Shen J, Takio K, Iwatsubo T. alpha-Synuclein is phosphorylated in synucleinopathy lesions. Nat Cell Biol. 2002;2:160–4. doi: 10.1038/ncb748. [DOI] [PubMed] [Google Scholar]
- 11.Hasegawa M, Fujiwara H, Nonaka T, Wakabayashi K, Takahashi H, Lee VM, Trojanowski JQ, Mann D, Iwatsubo T. Phosphorylated alpha-synuclein is ubiquitinated in alpha-synucleinopathy lesions. J Biol Chem. 2002;50:49071–6. doi: 10.1074/jbc.M208046200. [DOI] [PubMed] [Google Scholar]
- 12.Wakabayashi K, Tanji K, Mori F, Takahashi H. The Lewy body in Parkinson’s disease: molecules implicated in the formation and degradation of alpha-synuclein aggregates. Neuropathology. 2007;5:494–506. doi: 10.1111/j.1440-1789.2007.00803.x. [DOI] [PubMed] [Google Scholar]
- 13.Braak H, Del Tredici K, Rub U, de Vos RAI, Steur ENHJ, Braak E. Staging of brain pathology related to Parkinson’s disease. Neurobiol Aging. 2003;24:197–211. doi: 10.1016/s0197-4580(02)00065-9. [DOI] [PubMed] [Google Scholar]
- 14.Kanazawa T, Uchihara T, Takahashi A, Nakamura A, Orimo S, Mizusawa H. Three-layered structure shared between Lewy bodies and Lewy neurites-three-dimensional reconstruction of tripled-labeled sections. Brain Pathol. 2008;18:415–22. doi: 10.1111/j.1750-3639.2008.00140.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Orimo S, Uchihara T, Nakamura A, Mori F, Kakita A, Wakabayashi K, Takahashi H. Axonal alpha-synuclein aggregates herald centripetal degeneration of cardiac sympathetic nerve in Parkinson’s disease. Brain. 2008;131:642–50. doi: 10.1093/brain/awm302. [DOI] [PubMed] [Google Scholar]
- 16.Saito Y, Kawashima A, Ruberu NN, Fujiwara H, Koyama S, Sawabe M, Arai T, Nagura H, Yamanouchi H, Hasegawa M, Iwatsubo T, Murayama S. Accumulation of phosphorylated alpha-synuclien in aging human brain. J Neuropathol Exp Neurol. 2003;62:644–54. doi: 10.1093/jnen/62.6.644. [DOI] [PubMed] [Google Scholar]
- 17.Volpicelli-Daley LA, Luk KC, Patel TP, Tanik SA, Riddle DM, Stieber A, Meaney DF, Trojanowski JQ, Lee VMY. Exogenous α-synuclein fibrils induce a Lewy body pathology leading to synaptic dysfunction and neuron death. Neuron. 2011;72:57–71. doi: 10.1016/j.neuron.2011.08.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Brettschneider J, Del Tredici K, Lee VM, Trojanowski JQ. Spreading of pathology in neurodegenerative diseases: a focus on human studies. Nat Rev Neurosci. 2015;16:109–20. doi: 10.1038/nrn3887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Guo JL, Lee VMY. Cell-to-cell transmission of pathogenic proteins in neurodegenerative diseases. Nat Med. 2014;20:130–8. doi: 10.1038/nm.3457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jucker M, Walker LC. Self-propogation of pathogenic protein aggregates in neurodegenerative diseases. Nature. 2013;501:45–51. doi: 10.1038/nature12481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kordower JH, Chu Y, Hauser RA, Freeman TB, Olanow CW. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson’s disease. Nat Med. 2008;14:504–6. doi: 10.1038/nm1747. [DOI] [PubMed] [Google Scholar]
- 22.Li JY, Englund E, Holton JL, Soulet D, Hagell P, Lees AJ, Lashley T, Quinn NP, Rehncrona S, Björklund A, Widner H, Revesz T, Lindvall O, Brundin P. Lewy bodies in grafted neurons in subjects with Parkinson’s disease suggest host-to-graft disease propagation. Nat Med. 2008;14:501–3. doi: 10.1038/nm1746. [DOI] [PubMed] [Google Scholar]
- 23.Brahic M, Bousset L, Bieri G, Melki R, Gitler AD. Axonal transport and secretion of fibrillar forms of α-synuclein, Aβ42 peptide and HTTExon 1. Acta Neuropathol. 2016;131:539–48. doi: 10.1007/s00401-016-1538-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Freundt EC, Maynard N, Clancy EK, Roy S, Bousset L, Souriques Y, Covert M, Melki R, Kirkegaard K, Brahic M. Neuron-to-neuron transmission of α-synuclein fibrils through axonal transport. Ann Neurol. 2012;72:517–24. doi: 10.1002/ana.23747. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Volpicelli-Daley LA, Gamble KL, Schultheiss CE, Riddle DM, West AB, Lee VMY. Formation of α-synuclein Lewy neurite-like aggregates in axons impedes the transport of distinct endosomes. Mol Biol Cell. 2014;25:4010–23. doi: 10.1091/mbc.E14-02-0741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Luk KC, Kehm V, Carroll J, Zhang B, O’Brien P, Trojanowski JQ, Lee VMY. Pathological α-synuclein transmission initiates Parkinson-like neurodegeneration in nontransgenic mice. Science. 2012;338:949–53. doi: 10.1126/science.1227157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Luk KC, Song C, O’Brien P, Stieber A, Branch JR, Brunden KR, Trojanowski JQ, Lee VMY. Exogenous alpha-synuclein fibrils seed the formation of Lewy body-like intracellular inclusions in cultured cells. Proc Natl Acad Sci USA. 2009;106:20051–6. doi: 10.1073/pnas.0908005106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Osterberg VR, Spinelli KJ, Weston LJ, Luk KC, Woltjer RL, Unni VK. Progressive aggregation of alpha-synuclein and selective degeneration of Lewy inclusion bearing neurons in a mouse model of Parkinsonism. Cell Rep. 2015;10:1252–60. doi: 10.1016/j.celrep.2015.01.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Paumier KL, Luk KC, Manfredsson FP, Kanaan NM, Lipton JW, Collier TJ, Steece-Collier K, Kemp CJ, Celano S, Schulz E, Sandoval IM, Fleming S, Dirr E, Polinski NK, Trojanowski JQ, Lee VM, Sortwell CE. Intrastriatal injection of pre-formed mouse α-synuclein fibrils into rats triggers α-synuclein pathology and bilateral nigrostriatal degeneration. Neurobiol Dis. 2015;82:185–99. doi: 10.1016/j.nbd.2015.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Bousset L, Pieri L, Ruis-Arlandis G, Gath J, Jensen PH, Habenstein B, Madiona K, Olieric V, Böckmann A, Meier BH, Melki R. Structural and functional characterization of two alpha-synuclein strains. Nat Commun. 2013;4:2575. doi: 10.1038/ncomms3575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Guo JL, Covell DJ, Daniels JP, Iba M, Stieber A, Zhang B, Riddle DM, Kwong LK, Xu Y, Trojanowski JQ, Lee VMY. Distinct α-synuclein strains differentially promote tau inclusions in neurons. Cell. 2013;154:103–17. doi: 10.1016/j.cell.2013.05.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Peelaerts W, Bousset L, Van den Perren A, Moskalyuk A, Pulizzi R, Giugliano M, Van den Haute C, Melki R, Baekelandt V. α-Synuclein strains cause distinct synucleinopathies after local and systemic administration. Nature. 2015;522:340–4. doi: 10.1038/nature14547. [DOI] [PubMed] [Google Scholar]
- 33.Prusiner SB, Woerman AL, Mordes DA, Watts JC, Rampersaud R, Berry DB, Patel S, Oehler A, Lowe JK, Kravitz SN, Geschwind DH, Glidden DV, Halliday GM, Middleton LT, Gentleman SM, Grinberg LT, Giles K. Evidence for α-synuclein prions causing multiple system atrophy in humans with Parkinsonism. Proc Natl Acad Sci USA. 2015;112:E5308–17. doi: 10.1073/pnas.1514475112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Giasson BI, Murray IVJ, Trojanowski JQ, Lee VMY. A hydrophobic stretch of 12 amino acid residues in the middle of alpha-synuclein is essential for filament assembly. J Biol Chem. 2001;276:2380–6. doi: 10.1074/jbc.M008919200. [DOI] [PubMed] [Google Scholar]
- 35.Volpicelli-Daley LA, Luk KC, Lee VMY. Addition of exogenous alpha-synuclein preformed fibrils to primary neuronal cultures to seed recruitment of endogenous alpha-synuclein to Lewy body and Lewy neurite-like aggregates. Nat Protoc. 2014;9:2135–46. doi: 10.1038/nprot.2014.143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Giasson BI, Jakes R, Goedert M, Duda JE, Leight S, Trojanowski JQ, Lee VMY. A panel of epitope-specific antibodies detects protein domains distributed throughout human alpha-synuclein in Lewy bodies of Parkinson’s disease. J Neurosci Res. 2000;59:528–33. doi: 10.1002/(SICI)1097-4547(20000215)59:4<528::AID-JNR8>3.0.CO;2-0. [DOI] [PubMed] [Google Scholar]
- 37.El-agnaf O. Alpha-synuclein antibodies and use thereof. 20140241984 A1 U.S. patent. 2014
- 38.Baba M, Nakajo S, Tu PH, Tomita T, Nakaya K, Lee VMY, Trojanowski JQ, Iwatsubo T. Aggregation of alpha-synuclein in Lewy bodies of sporadic Parkinson’s disease and dementia with Lewy bodies. Am J Pathol. 1998;152:879–84. [PMC free article] [PubMed] [Google Scholar]
- 39.Duda JE, Giasson BI, Mabon ME, Lee VMY, Trojanowski JQ. Novel antibodies to synuclein show abundant striatal pathology in Lewy body diseases. Ann Neurol. 2002;52:205–10. doi: 10.1002/ana.10279. [DOI] [PubMed] [Google Scholar]
- 40.Toledo JB, Van Deerlin VM, Lee EB, Suh E, Baek Y, Robinson JL, Xie SX, McBride J, Wood EM, Schuck T, Irwin DJ, Gross RG, Hurtig H, McCluskey L, Elman L, Karlawish J, Schellenberg G, Chen-Plotkin A, Wolk D, Grossman M, Arnold SE, Shaw LM, Lee VMY, Trojanowski JQ. A platform for discovery: The University of Pennsylvania Integrated Neurodegenerative Disease Biobank. Alzheimers Dement. 2014;10:477–84. doi: 10.1016/j.jalz.2013.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Irwin DJ, White MT, Toledo JB, Xie SX, Robinson JL, Van Deerin V, Lee VMY, Leverenz JB, Montine TJ, Duda JE, Hurtig HI, Trojanowski JQ. Neuropathologic substrates of Parkinson disease dementia. Ann Neurol. 2012;72:587–98. doi: 10.1002/ana.23659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Montine TJ, Monsell SE, Beach TG, Bigio EH, Bu Y, Caims NJ, Frosch M, Henriksen J, Kofler J, Kukull WA, Lee EB, Nelson PT, Schantz AM, Schneider JA, Sonnen JA, Trojanowski JQ, Vinters HV, Zhou XH, Hyman BT. Multisite assessment of the NIA-AA guidelines for the neuropathologic evaluation of Alzheimer’s disease. Alzheimers Dement. 2016;12:164–9. doi: 10.1016/j.jalz.2015.07.492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Trojanowski JQ, Schuck T, Schmidt ML, Lee VMY. Distribution of phosphate-independent MAP2 epitopes revealed with monoclonal antibodies in microwave-denatured human nervous system tissues. J Neurosci Methods. 1989;2:171–80. doi: 10.1016/0165-0270(89)90030-7. [DOI] [PubMed] [Google Scholar]
- 44.Hughes AJ, Daniel SE, Kilford L, Lees AJ. Accuracy of clinical diagnosis of idiopathic Parkinson’s disease: a clinic-pathological study of 100 cases. J Neurol Neurosurg Psychiatry. 1992;55:181–4. doi: 10.1136/jnnp.55.3.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Leverenz JB, Hamilton R, Tsuang DW, Schantz A, Vavrek D, Larson EB, Kukull WA, Lopez O, Galasko D, Masliah E, Kaye J, Woltjer R, Clark C, Trojanowski JQ, Montine TJ. Empiric refinement of the pathologic assessment of Lewy-related pathology in the dementia patient. Brain Pathol. 2008;18:220–4. doi: 10.1111/j.1750-3639.2007.00117.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Vaikath NN, Majbour NK, Paleologou KE, Ardah MT, van Dam E, van de Berg WD, Forrest SL, Parkkinen L, Gai WP, Hattori N, Takanashi M, Lee SJ, Mann DM, Imai Y, Halliday GM, Li JY, El-Agnaf OM. Generation and characterization of novel conformation-specific monoclonal antibodies for alpha-synuclein pathology. Neurobiol Dis. 2015;79:81–99. doi: 10.1016/j.nbd.2015.04.009. [DOI] [PubMed] [Google Scholar]
- 47.Chen M, Margittai M, Chen J, Langen R. Investigation of alpha-synuclein fibril structure by site-directed spin labeling. J Biol Chem. 2007;282:24970–9. doi: 10.1074/jbc.M700368200. [DOI] [PubMed] [Google Scholar]
- 48.Comellas G, Lemkau LR, Nieuwkoop AJ, Kloepper KD, Ladror DT, Ebisu R, Woods WS, Lipton AS, George JM, Rienstra CM. Structured regions of α-synuclein fibrils include the early-onset Parkinson’s disease mutation sites. J Mol Biol. 2011;411:881–95. doi: 10.1016/j.jmb.2011.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Heise H, Hoyer W, Becker S, Andronesi OC, Riedel D, Baldus M. Molecular-level secondary structure, polymorphism, and dynamics of full-length alpha-synuclein fibrils studied by solid-state NMR. Proc Natl Acad Sci USA. 2005;102:15871–6. doi: 10.1073/pnas.0506109102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Tuttle M, Comellas G, Niewkoop A, Covell DJ, Berthold DA, Kloepper KD, Courtney JM, Kim JK, Barclay AM, Kendall A, Wan W, Stubbs G, Schwieters CD, Lee VMY, George JM, Rienstra CM. Solid-state NMR structure of a pathogenic fibril of full-length human α-synuclein. Nat Struct Mol Biol. 2016;23:409–15. doi: 10.1038/nsmb.3194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Allison JR, Rivers RC, Christodoulou JC, Vendruscolo M, Dobson CM. A relationship between the transient structure in the monomeric state and the aggregation propensities of α-synuclein and β-synuclein. Biochemistry. 2014;53:7170–83. doi: 10.1021/bi5009326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Bertoncini CW, Jung YS, Fernandez CO, Hoyer W, Griesinger C, Jovin TM, Zweckstetter M. Release of long-range tertiary interactions potentiates aggregation of natively unstructured alpha-synuclein. Proc Natl Acad Sci USA. 2005;102:1430–5. doi: 10.1073/pnas.0407146102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Murray IV, Giasson BI, Quinn SM, Koppaka V, Axelsen PH, Ischiropoulos H, Trojanowski JQ, Lee VM. Role of the alpha-synuclein carboxy-terminus on fibril formation in vitro. Biochemistry. 2003;42:8530–40. doi: 10.1021/bi027363r. [DOI] [PubMed] [Google Scholar]
- 54.Serpell LC, Berriman J, Jakes R, Goedert M, Crowther RA. Fiber diffraction of synthetic alpha-synuclein filaments shows amyloid-like cross-beta conformation. Proc Natl Acad Sci USA. 2000;97:4897–902. doi: 10.1073/pnas.97.9.4897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mitchell TW, Nissanov J, Han LY, Mufson EJ, Schneider JA, Cochran EJ, Bennett DA, Lee VM, Trojanowski JQ, Arnold SE. Novel method to quantify neuropil threads in brains from elders with or without cognitive impairment. J Histochem Cytochem. 2000;48:1627–38. doi: 10.1177/002215540004801206. [DOI] [PubMed] [Google Scholar]
- 56.Irwin DJ, Byrne MD, McMillan CT, Cooper F, Arnold SE, Lee EB, Van Deerlin VM, Xie SX, Lee VMY, Grossman M, Trojanowski JQ. Semi-automated digital image analysis of Pick’s disease and TDP-43 proteinopathy. J Histochem Cytochem. 2016;64:54–66. doi: 10.1369/0022155415614303. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Figure S1. Characterization of antigen used for generation of strain A selective mAbs.
Figure S2. Additional characterization of strain-selective mAbs Syn7015 and Syn9029
Figure S3. Immunohistochemistry (IHC) optimization of Syn7015 and Syn9029 in ethanol and formalin fixed tissue.
Figure S4. Immunohistochemistry (IHC) staining of neuropathologically normal tissue with Syn303, Syn7015, and Syn9029.
Figure S5. Immunohistochemistry (IHC) of Syn303, Syn7015 and Syn9029 mAb dilution series in near serial sections of PD cingulate cortex.
Figure S6. Syn7015 and Syn9029 immunostaining can be removed by serial adsorption with strain A and B PFFs.
Figure S7. Immunohistochemistry (IHC) shows that additional 7000 series mAbs, as exemplified by Syn7020 and Syn7068, also prominently stain thread and dot Lewy neurites (LNs).
Figure S8. Double-labeling immunofluorescence of Syn7015 and Syn9029 co-localizes primarily in Lewy bodies (LBs) and thick Lewy neurites (LNs).
Figure S9. Semi-quantitative scores showing that preferential detection of Lewy neurites (LNs) by Syn7015 is maintained throughout the disease course in multiple brain regions.
Table S1. Demographic information and semi-quantitative scores for all patients in this study
Table S2 Comparative ability of α-syn strain selective antibodies to detect Lewy neurites (LNs) and Lewy bodies (LBs) in Parkinson’s disease brain tissues