

Abstract
Limbic-predominant age-related TDP-43 encephalopathy neuropathologic change (LATE-NC) is the most prevalent subtype of TDP-43 proteinopathy, affecting up to 1/3rd of aged persons. LATE-NC often co-occurs with hippocampal sclerosis (HS) pathology. It is currently unknown why some individuals with LATE-NC develop HS while others do not, but genetics may play a role. Previous studies found associations between LATE-NC phenotypes and specific genes: TMEM106B, GRN, ABCC9, KCNMB2, and APOE. Data from research participants with genomic and autopsy measures from the National Alzheimer’s Coordinating Center (NACC; n = 631 subjects included) and the Religious Orders Study and Memory and the Rush Aging Project (ROSMAP; n = 780 included) were analyzed in the current study. Our goals were to reevaluate disease-associated genetic variants using newly collected data and to query whether the specific genotype/phenotype associations could provide new insights into disease-driving pathways. Research subjects included in prior LATE/HS genome-wide association studies (GWAS) were excluded. Single nucleotide variants (SNVs) within 10 kb of TMEM106B, GRN, ABCC9, KCNMB2, and APOE were tested for association with HS and LATE-NC, and separately for Alzheimer’s pathologies, i.e. amyloid plaques and neurofibrillary tangles. Significantly associated SNVs were identified. When results were meta-analyzed, TMEM106B, GRN, and APOE had significant gene-based associations with both LATE and HS, whereas ABCC9 had significant associations with HS only. In a sensitivity analysis limited to LATE-NC + cases, ABCC9 variants were again associated with HS. By contrast, the associations of TMEM106B, GRN, and APOE with HS were attenuated when adjusting for TDP-43 proteinopathy, indicating that these genes may be associated primarily with TDP-43 proteinopathy. None of these genes except APOE appeared to be associated with Alzheimer’s-type pathology. In summary, using data not included in prior studies of LATE or HS genomics, we replicated several previously reported gene-based associations and found novel evidence that specific risk alleles can differentially affect LATE-NC and HS.
Supplementary Information
The online version contains supplementary material available at 10.1186/s40478-021-01250-2.
Keywords: Dementia, Proteinopathy, Pleiotropy, Arteriolosclerosis, SNP, Mixed pathology
Introduction
The present study focused on genetic contributions to transactive response DNA binding protein 43 kDa (TDP-43) proteinopathy and hippocampal sclerosis (HS). One or both of these pathologic features are observed in ~ 30% of brains among persons > 80 years at death [42]. The TDP-43 protein serves multiple functions in gene expression regulation at the levels of both transcription and translation [14, 21, 42, 52]. TDP-43 proteinopathy (aberrantly misfolded and mislocalized TDP-43 protein) is strongly associated with cognitive impairment [11, 41]. This pathologic hallmark was discovered in diseases that are now considered to be a clinical-pathologic spectrum that includes amyotrophic lateral sclerosis (ALS) and frontotemporal lobe degeneration with TDP-43 (FTLD-TDP) [50].
HS is a pathologic finding characterized by selective neuronal loss and gliosis of the hippocampal formation [3, 46]. First described as a pathologic phenomenon in epilepsy [19], HS is a descriptive and relatively nonspecific term used in both neuropathologic and neuroradiographic practice. However, in a subset of cases with HS, TDP-43 proteinopathy is also present [3, 11, 35, 46].
Limbic-predominant age-related TDP-43 encephalopathy (LATE) is a prevalent disease entity characterized by TDP-43 proteinopathy, with greatly increased risk for cognitive impairment, in aged populations [42]. LATE is not a subtype of FTLD-TDP because the associated disease(s) is not the frontotemporal dementia (FTD) clinical syndrome; rather, the presence of the neuropathologic changes underlying LATE (LATE-NC) is an amnestic dementia syndrome [40–42, 59]. HS pathology commonly co-occurs with LATE-NC and was the first neuropathologic change associated with the condition [17, 47]. However, some persons with LATE-NC have no HS, segmental/patchy HS, or unilateral HS [25]. It is currently unknown why some individuals with LATE-NC develop HS pathology while others do not, but genetics may help explain these phenomena.
Several genes and single nucleotide variants (SNVs) have been linked with LATE-NC phenotypes [42]. Risk for HS was previously associated with SNVs that are also known FTLD-TDP risk alleles, including rs5848 from the GRN gene on chromosome 17 and rs1990622 near the TMEM106B gene on chromosome 7 [6, 16, 39, 56, 62, 65]. In a genome-wide association study (GWAS), a SNV in the ABCC9 gene (rs704178/rs704180) on chromosome 12 was associated with HS risk [43]. A separate GWAS found that rs9637454, an SNV in the KCNMB2 gene on chromosome 3, was associated with HS risk [8]. Additional evidence exists linking the APOE ε4 allele, a strong risk factor for Alzheimer’s disease (AD), with increased HS and LATE-NC risk [60, 66, 68]. A study analyzing gene-based associations between the GRN, TMEM106B, ABCC9, and KCNMB2 genes and HS found Bonferroni-corrected significant associations for ABCC9 assuming a recessive mode of inheritance (MOI) and nominally significant associations with GRN, TMEM106B, and KCNMB2 [29]. However, a separate study replicated the associations between GRN and TMEM106B SNVs with LATE-NC, but did not find an association between an ABCC9 variant and LATE-NC or HS pathologies [23]. To the best of our knowledge, there has not been a prior study that found genomic associations with LATE-NC but not HS or vice versa.
In the current study, we analyzed genomic data from the Alzheimer’s Disease Genetics Consortium (ADGC) along with clinical and pathological data from the National Alzheimer’s Coordinating Center (NACC) and the Rush University Religious Orders Study and Memory and Aging Project (ROSMAP) to investigate the associations between prior identified putative risk genes – KCNMB2, TMEM106B, ABCC9, GRN, and APOE – and LATE-NC. While only analyzing participants not included in our prior studies [29, 43], we sought to test whether or not previously reported LATE-NC risk genes can be replicated for LATE-NC neuropathologic phenotypes (specifically, TDP-43 proteinopathy and HS) while also testing for the presence of novel risk alleles in those genes.
Material and methods
Study participants
Representative photomicrographs were taken, showing results from research participants with LATE-NC and LATE-NC + HS, in the University of Kentucky AD Research Center Autopsy cohort, using methods as previously described [44].
Phenotypic data from NACC (March 2021 data freeze) were linked with genotype data from the ADGC. Individuals who died at age 65 years or older were included. Similar to other studies using NACC data [27], individuals were excluded from the NACC cohort if at least one of 19 rare brain diseases were diagnosed (See Additional File 1: Supplemental Table 1) or if they were missing any adjustment variables or both endophenotypes under study.
The ROSMAP study has been described in detail elsewhere [36]. Briefly, data were acquired from two well-characterized cohort studies of aging and dementia. The Religious Orders Study (ROS), begun in 1994, and the Rush Memory and Aging Project (MAP), begun in 1997, involve older adults who enrolled without dementia, agreed to annual clinical evaluations and organ donation at death, and signed an Anatomical Gift Act for brain donation. Written informed consent was obtained from participants, and research was carried out in accordance with Institutional Review Board (IRB)-approved protocols. ROSMAP data are available online at the Rush Alzheimer’s Disease Center Resource Sharing Hub (https://www.radc.rush.edu/), as well as on the Accelerating Medicines Partnership-Alzheimer’s Disease (AMP-AD) Knowledge Portal (syn3219045).
For both the NACC and ROSMAP datasets, individuals were excluded from the analyses if they were included in either of two previous studies of HS genomics [29, 43]. In ROSMAP, participants were excluded based on IID if they were included in the Nelson et al. HS GWAS from 2014. In NACC, HS and TDP-43 were defined using variables from the v10 NACC Neuropathology (NP) dataset which were not available for the participants included in the previous studies. Thus, the NACC and ROSMAP participants included in the current study are a true replication cohort for these earlier HS genomics studies.
Neuropathological endophenotype definitions
In the NACC NP dataset, LATE-NC was defined as either present or absent using the “distribution of TDP-43 immunoreactive inclusions” variables indicating if TDP-43 proteinopathy was observed in either the hippocampus (NPTDPC NACC field), entorhinal/inferior temporal cortex (NPTDPD), or neocortex (NPTDPE) in a case lacking overall diagnosis of FTLD-TDP. A LATE-NC case was defined as definitely having TDP-43 in the hippocampus, entorhinal/inferior temporal cortex, or neocortex. LATE-NC was considered unknown if TDP-43 data were unavailable in all three regions. HS was defined as either present or absent based on the “hippocampal sclerosis of CA1 and/or subiculum” (NPHIPSCL) variable using the “unilateral,” “bilateral,” and “present but laterality not assessed” response categories.
In the ROSMAP data set, LATE-NC was defined dichotomously using the “TDP-43 stage” (tdp_st4) variable and collapsing the 2nd and 3rd stages in cases lacking FTLD-TDP. HS was defined dichotomously by the “hippocampal sclerosis was rated as definitely present with CA1 region affected” response category of the “definite presence of typical hippocampal sclerosis” (hspath_typ) variable.
Quality control of genotype data
For NACC participants, genomic data from the ADGC imputed using the Haplotype Reference Consortium (ADGC-HRC) were used [38]. The genetic data for ROSMAP were also imputed using the HRC and the methods have been described in detail elsewhere [18]. Standard GWAS quality control (QC) procedures were performed separately on the ADGC and ROSMAP genotype data using PLINK1.9 [37, 54]. SNVs were excluded if they were missing in more than 5% of samples, if they had a minor allele frequency less than 1%, or if they had Hardy–Weinberg Equilibrium (HWE) p-values < 1 × 10–6 among AD controls. Individuals were excluded if they were missing more than 5% of genotypes. Two individuals were considered related if they had an identity by descent measure of at least 0.25, which indicates that they are second-degree relatives. For related pairs, the individual with the lowest call rate was excluded.
NACC and ROSMAP genotype data were separately merged with 1000 Genomes Project Phase 3 data. Principal components (PCs) were calculated for the merged data sets using the “pca” procedure in PLINK1.9, and the first two PCs were plotted. The ADGC-HRC and ROSMAP individuals with first and second PCs that overlapped with those of the 1000 Genomes individuals of known European ancestry were identified and all other individuals were excluded from the analysis.
Variant-level associations
All statistical analyses were conducted in R programming language [55], version 4.0.4. Associations between each endophenotype and each SNV were conducted separately in the NACC and ROSMAP datasets using binary logistic regression models assuming each of the three most common MOI: additive, dominant, and recessive. SNVs were excluded from the analyses if they were multiallelic or if there were fewer than 15 minor alleles present across all participants. All regression models were fit using the glm function in R assuming a binomial distribution and a logit link function and were adjusted for age at death, sex, ADGC data selection round (for NACC data) or ROS/MAP study (for ROSMAP data), and the first three genetic PCs. Odds ratios (OR) were calculated for each SNV. Since some endophenotypes were only available in a subset of participants, PCs were calculated separately for each endophenotype. NACC and ROSMAP SNV-level results were meta-analyzed using a fixed-effect, inverse-variance meta-analysis via the metagen function from the meta R package, version 4.18-0 [7]. For targeted analyses of previously reported SNVs, an additive MOI was assumed unless there existed previous evidence of association with another MOI. Additionally, LATE-NC-by-SNV interaction terms were tests for models of HS and were removed if they failed to reach statistical significance (p < 0.05). Plots of cohort-specific and meta-analyzed SNV-level p-values were created using LocusZoom Standalone, version 1.4 (https://genome.sph.umich.edu/wiki/LocusZoom_Standalone) [53], and the ggplot2 R package, version 3.3.3 [67]. Linkage disequilibrium estimates were computed using LDlink with the CEU population (https://ldlink.nci.nih.gov/) [34]. Jaccard similarity coefficients were used to estimate the similarity between binary variables and were calculated by dividing the size of their intersection by the size of their union via the clusteval R package, version 0.1 [58].
Gene-based associations
Gene boundaries for KCNMB2, TMEM106B, ABCC9, GRN, and APOE were defined based on their canonical transcripts using the Genome Reference Consortium Human Build 37 (GRCh37/hg19) gene range list from PLINK (https://www.cog-genomics.org/plink/1.9/resources). All genes were flanked by an additional 10 kb to include potential regulatory regions. See Additional File 1: Supplemental Table 2 for the positions used to define the gene boundaries.
For each gene, endophenotype, and MOI, all SNV-level p-values were combined using the aggregated Cauchy association test (ACAT) [33]. All ACAT analyses were run using R functions provided by the authors (https://github.com/yaowuliu/ACAT). Equal weights were assumed for all SNVs in the ACAT analyses and statistical significance was defined as a p-value < 0.05.
SNV prioritization and follow-up analyses
Prioritized SNVs were identified using a Bonferroni-corrected threshold for significance that accounts for the effective number of independent tests in a given genetic region. The effective number of independent tests in a region was calculated for each endophenotype using the method of Gao et al. [20]. Briefly, Pearson’s correlation coefficient was calculated for all pairs of SNVs and these coefficients were placed in a square matrix. The eigenvalues of the matrix were then computed and ordered from largest to smallest and the effective number of independent tests was defined to be the smallest number of ordered eigenvalues that account for 99.5% of the sum of all eigenvalues. The Bonferroni-corrected threshold for identifying prioritized SNVs in a given genetic region was defined as 0.05 divided by the largest estimated number of independent tests in the region.
Prioritized SNVs were investigated for expression quantitative trait loci (eQTL) associations using the Genotype-Tissue Expression (GTEx) Project’s V8 public data [15], the BRAINEAC Brain eQTL Almanac (http://braineac.org/) [57], and Functional Annotation of Human Long Noncoding RNAs via Molecular Mapping (FANTOM5) database (data accessed via: https://www.ebi.ac.uk/gxa/experiments/E-MTAB-3358/Results). Prioritized SNVs were also investigated for associations with other molecular mechanisms using the INFERring the molecular mechanisms of NOncoding genetic variants (INFERNO) software assuming a threshold on r2 of 0.5 and a threshold on LD block size of 500 kb (http://inferno.lisanwanglab.org/index.php) [4].
Sensitivity analyses
Additional analyses tested if the study’s results were dependent upon a priori analytic approaches. All gene-based analyses were also conducted assuming 0 kb and 25 kb of flanking around each gene. All top SNVs were tested for associations with AD-related neuropathologies, to see if there were indications that the HS and LATE-NC associations were being driven by AD. Additionally, since TDP-43 in the amygdala was not included in the dichotomous LATE-NC definition, all top SNVs were also tested for associations with LATE-NC Stage 1 (vs. LATE-NC Stage 0) to determine if any amygdala-specific associations were missed in the primary analyses.
Results
The phenotypes of interest in the current study are autopsy-confirmed LATE-NC and HS. Specific examples of those pathologies are depicted in Fig. 1. Some brains have LATE-NC without HS (Fig. 1b). However, individuals with LATE-NC are at increased risk of having comorbid HS (Fig. 1c).
Fig. 1.
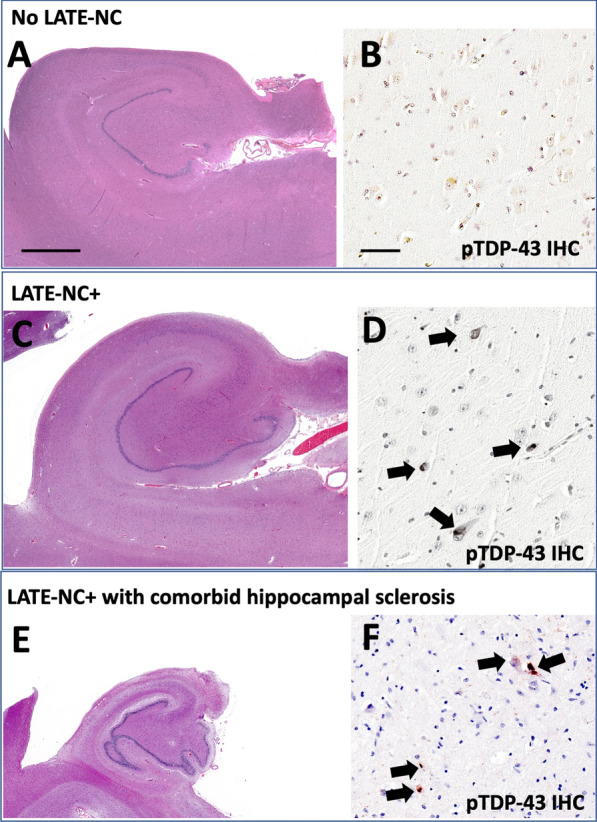
Photomicrographs of human hippocampi depict the main neuropathologic endophenotypes analyzed in the current study. Hippocampal sclerosis (HS) is evaluated with H&E stain (panels A, C, E), whereas LATE-NC is operationalized with phospho-TDP-43 immunohistochemistry (IHC; panels B, D, and F). All photomicrographs depict mid-level hippocampal sections dissected in the coronal plane. Panels A and B show stained brain sections from a woman (APOE e3/e4) who died at age 83; autopsy revealed neither LATE-NC nor HS. Panels C and D are from a man (APOE e3/e4) who died at age 93 with LATE-NC Stage 2. Panels E and F are from a woman (APOE e3/e3) who died at age 95 with LATE-NC Stage 2 and comorbid HS. Note the relatively atrophic hippocampal profile in Panel E in comparison to a or c (same scale bar); the HS + profile in panel E also demonstrates parenchymal rarefaction which can be appreciated even at low magnification. Phospho-TDP-43 immunoreactive intraneuronal inclusions are highlighted with arrows in panels D and F. The representative photomicrographs were from research participants of the University of Kentucky AD Research Center. Scale bar = 2 mm in A, C, and E, 75 microns in A, D, and F
The participants included and excluded, and the reasons for exclusion, are shown in Fig. 2. In the ROSMAP data set, a total of n = 795 individuals had available data for at least one of the endophenotypes along with GWAS data and were not included in earlier studies of HS [29, 43]. In the NACC data set, n = 633 individuals had available data for at least one of the endophenotypes along with GWAS data and were not included in the earlier studies of HS [29, 43]. While not all FTLD subtypes were explicitly excluded among NACC participants, no FTLD cases were included in the final sample likely due inclusion criteria applied by the ADGC during genotyping. Table 1 shows a summary of individual characteristics and endophenotypes for both NACC and ROSMAP participants. ROSMAP participants tended to be older at death (p < 0.001), were more likely to be female (p < 0.001), and were less likely to be an HS case (p = 0.007) than NACC participants. HS was less prevalent than LATE-NC in both cohorts (NACC: HS 14.1%, LATE-NC 29.4%; ROSMAP: HS 9.4%, LATE-NC 33.2%). In ROSMAP, both HS cases and LATE-NC cases tended to be older at death (both p < 0.001) and were less likely to be male (p = 0.054 and p < 0.001, respectively) than their respective controls. There were no identified statistically significant differences in basic demographic characteristics between HS and/or LATE-NC cases and their respective controls in NACC. Additional participant characteristics stratified by combined LATE-NC and HS case status are included in Additional File 1: Supplemental Tables 4 and 5.
Fig. 2.

Included and excluded research participants, along with criteria for exclusion. A flowchart summarizing inclusions and exclusions for National Alzheimer's Coordinating Center (NACC) and Religious Orders Study and Rush Memory and Aging Project (ROSMAP) participants
Table 1.
Participant characteristics stratified by endophenotype status
| NACC | ROSMAP | |||||
|---|---|---|---|---|---|---|
| Number of Participants (%) | Age at Death, Mean (SD) | Female, N (%) | Number of Participants (%) | Age at Death, Mean (SD) | Female, N (%) | |
| HS | ||||||
| Overall | N = 631 | 85.9 (8.3) | 319 (50.6) | N = 780 | 88.7 (7.2) | 525 (67.3) |
| No | 542 (85.9) | 85.9 (8.4) | 270 (49.8) | 707 (90.6) | 88.3 (7.2) | 468 (66.2) |
| Yes | 89 (14.1) | 86.0 (7.5) | 49 (55.1) | 73 (9.4) | 92.0 (6.4) | 57 (78.1) |
| LATE-NC | ||||||
| Overall | N = 512 | 85.1 (7.9) | 207 (50.2) | N = 747 | 89.1 (7.1) | 506 (67.7) |
| No | 291 (70.6) | 84.9 (8.1) | 138 (47.4) | 499 (66.8) | 87.9 (7.3) | 315 (63.1) |
| Yes | 121 (29.4) | 85.4 (7.3) | 66 (57.0) | 248 (33.2) | 91.5 (6.1) | 191 (77.0) |
Participant characteristics stratified by hippocampal sclerosis (HS) and limbic-predominant age-related TDP-43 encephalopathy neuropathological changes (LATE-NC) case status. NACC = National Alzheimer's Coordinating Center; ROSMAP = Religious Orders Study and Rush Memory and Aging Project; SD = standard deviation; HS = hippocampal sclerosis; LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological changes
Persons with HS tended to also have LATE-NC and the reverse was also true among individuals in both datasets (Jaccard coefficients of 0.589 and 0.575 in NACC and ROSMAP, respectively); see Fig. 3. Of the 732 ROSMAP participants with available case data for both LATE-NC and HS, 93% of HS cases were also LATE-NC cases. Of the 410 NACC participants with available case data for both LATE-NC and HS, 73% of HS cases were also LATE-NC cases.
Fig. 3.
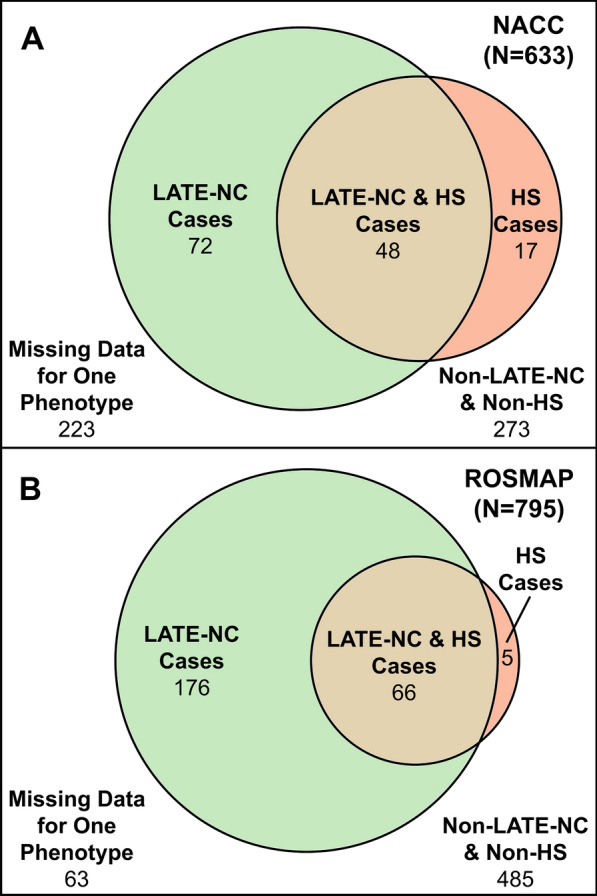
Venn diagrams of the overlap between endophenotypes across studies. Venn diagrams of the overlap between limbic-predominant age-related TDP-43 encephalopathy neuropathological change (LATE-NC) and hippocampal sclerosis (HS) cases in A. National Alzheimer's Coordinating Center (NACC) and B. Religious Orders Study and Rush Memory and Aging Project (ROSMAP). LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological change; HS = hippocampal sclerosis; NACC = National Alzheimer's Coordinating Center; ROSMAP = Religious Orders Study and Rush Memory and Aging Project
Across the KCNMB2, TMEM106B, ABCC9, GRN, and APOE genes, each flanked by 10 kb, a total of 1,580 SNVs passed QC in NACC while 1,532 SNVs passed QC in ROSMAP. A total of 1,438 SNVs were shared between NACC and ROSMAP and were included in the meta-analysis (Additional File 1: Supplemental Table 2).
Gene-based associations
The adjusted meta-analyzed, SNV-level results were combined within genes via ACAT to obtain gene-based p-values. At the gene level, TMEM106B and APOE were significantly associated with both HS and LATE-NC while ABCC9 and GRN were significantly associated with HS only (Table 2). Neither HS nor LATE-NC were significantly associated with KCNMB2. The meta-analyzed gene-based results were largely similar to when they were conducted separately in the NACC and ROSMAP datasets. Additionally, these results were largely unchanged when 0 kb and 25 kb of flanking were added to each gene.
Table 2.
Gene-based results for risk genes
| Chr | Gene | Endophenotype | MOI | ||
|---|---|---|---|---|---|
| Additive | Dominant | Recessive | |||
| 3 | KCNMB2 | HS | 0.718 | 0.632 | 0.478 |
| LATE-NC | 0.980 | 0.995 | 0.473 | ||
| 7 | TMEM106B | HS | 0.006 | 0.052 | 0.005 |
| LATE-NC | < 0.001 | 0.004 | < 0.001 | ||
| 12 | ABCC9 | HS | 0.036 | 0.072 | 0.006 |
| LATE-NC | 0.901 | 0.440 | 0.912 | ||
| 17 | GRN | HS | 0.004 | 0.348 | 0.003 |
| LATE-NC | 0.164 | 0.628 | 0.069 | ||
| 19 | APOE | HS | 0.014 | 0.017 | 0.333 |
| LATE-NC | < 0.001 | < 0.001 | 0.064 | ||
Aggregated Cauchy association test (ACAT) gene-based p-values for hippocampal sclerosis (HS) and limbic-predominant age-related TDP-43 encephalopathy neuropathological changes (LATE-NC). Each gene is flanked by 10 kb. All SNV-level analyses were adjusted for sex, age at death, cohort/study, and the first three genetic principal components and meta-analyzed across National Alzheimer's Coordinating Center (NACC) and Religious Orders Study and Rush Memory and Aging Project (ROSMAP) participants. Chr. = chromosome; HS = hippocampal sclerosis; LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological changes; MOI = mode of inheritance.
Prioritized SNVs and follow-up analyses
The effective number of independent tests for TMEM106B ± 10 kb was estimated to be 25, GRN ± 10 kb was estimated to be 16, KCNMB2 ± 10 kb was estimated to be 104, APOE ± 10 kb was estimated to be 14, and ABCC9 ± 10 kb was estimated to be 71. The Bonferroni-corrected thresholds for a genetic region was calculated by dividing 0.05 by the corresponding estimated effective number of independent tests in the region.
One hundred and ten SNVs in the TMEM106B ± 10 kb locus had adjusted meta-analytic associations with HS or LATE-NC less than the Bonferroni-corrected threshold (Fig. 4a). At the TMEM106B ± 10 kb locus, rs7781670 had the smallest adjusted meta-analytic p-value for LATE-NC assuming an additive MOI (p = 2.97 × 10–5). rs7781670 also met the Bonferroni-corrected threshold for the TMEM106B ± 10 kb locus for HS when assuming a recessive MOI (p = 1.63 × 10–3) and was a significant eQTL in GTEx for TMEM106B in the cerebellum (p = 4.7 × 10–7) and the cortex (p = 2.6 × 10–5). In INFERNO, these prioritized SNVs were associated with both eQTLs and Roadmap enhancers in blood, connective, and epithelial tissues and just Roadmap enhancers in brain, heart, immune organ, liver, and skeletal tissues, among others.
Fig. 4.

Variant-level results for TMEM106B and GRN. Adjusted, meta-analytic, single nucleotide variant (SNV)-level p-values for hippocampal sclerosis (HS) and limbic-predominant age-related TDP-43 encephalopathy neuropathological change (LATE-NC) across A. TMEM106B ± 10 kb and B. GRN ± 10 kb. All analyses were adjusted for sex, age at death, cohort/study, and the first three genetic principal components. Horizontal dashed lines represent the Bonferroni-corrected thresholds for significance that account for the number of independent tests in each genomic region. A diamond represents the SNV with the smallest p-value. The previously identified TMEM106B SNV (Rutherford et al. [62]) is labeled and identified with an arrow. MOI = mode of inheritance; LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological change; HS = hippocampal sclerosis
Fourteen SNVs in the GRN ± 10 kb locus had adjusted meta-analytic associations with HS or LATE-NC less than the Bonferroni-corrected threshold (Fig. 4b). rs5848 had the smallest adjusted meta-analytic p-value in the GRN ± 10 kb locus and met the Bonferroni-corrected threshold for HS (additive MOI p = 2.16 × 10–4; recessive MOI p = 1.91 × 10–4). rs5848 also had the smallest adjusted meta-analytic p-value for LATE-NC in the GRN ± 10 kb locus, but it did not meet the Bonferroni-corrected threshold. In GTEx, rs5848 was a significant eQTL for GRN expression in numerous tissues including thyroid (p = 2.2 × 10–16), caudate (p = 2.0 × 10–12), cortex (p = 2.0 × 10–9), and frontal cortex (p = 4.4 × 10–9). In INFERNO, these prioritized SNVs were associated with both eQTLs and Roadmap enhancers in adipose, connective, endocrine, heart, and nervous tissues, with just eQTLs in blood vessel tissue, and with just Roadmap enhancers in brain, blood, immune organ, liver, and skeletal muscle tissues, among others.
No SNVs in the KCNMB2 ± 10 kb locus had adjusted meta-analytic associations with HS or LATE-NC that met the Bonferroni-corrected threshold (Additional File 1: Supplemental Fig. 1).
The APOE ± 10 kb locus was strongly associated with LATE-NC. Four SNVs (rs429358, rs769449, rs10414043, and rs7256200), all in high linkage disequilibrium with one another (all r2 > 0.95), had adjusted meta-analytic associations with LATE-NC that met the Bonferroni-corrected threshold assuming an additive MOI (all p-values ≤ 2.56 × 10–8) (Fig. 5). While none of the APOE SNVs were associated with APOE expression levels in the evaluated data sets, rs769449 and rs10414043 were significant sQTLs in GTEx for TOMM40 in cerebellar hemisphere tissue (p = 4.0 × 10–10 and p = 1.4 × 10–5, respectively). In INFERNO, these prioritized SNVs were associated with both Roadmap and FANTOM5 enhancers in adipose, blood, brain, connective, epithelial, liver, nervous, skeletal muscle, smooth muscle, and stem cell tissues and with just Roadmap enhancers in endocrine, heart, and immune organ tissues, among others.
Fig. 5.

Variant-level results for APOE. Adjusted, meta-analytic, single nucleotide variant (SNV)-level p-values for hippocampal sclerosis (HS) and limbic-predominant age-related TDP-43 encephalopathy neuropathological change (LATE-NC) across APOE ± 10 kb. All analyses were adjusted for sex, age at death, cohort/study, and the first three genetic principal components. The horizontal dashed line represents the Bonferroni-corrected threshold for significance that accounts for the number of independent tests in the APOE ± 10 kb region. A diamond represents the SNV with the smallest p-value. MOI = mode of inheritance; LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological change; HS = hippocampal sclerosis
The ABCC9 ± 10 kb locus was most strongly associated with HS and contained 13 SNVs with adjusted meta-analytic p-values for HS less than the Bonferroni-corrected threshold (Fig. 6). rs1914361 had the smallest adjusted meta-analytic p-value with HS assuming a recessive MOI (p = 1.70 × 10–4). In prior studies with cohorts of research subjects that did not overlap with the current study, the ABCC9/HS association was strongest for the recessive MOI models [29, 43, 48]. All other SNVs that also met the Bonferroni-corrected threshold when assuming a recessive MOI were in high linkage disequilibrium with rs1914361 (all r2 > 0.75). rs1914361 was a significant eQTL in the GTEx data set for the expression of ABCC9 in several tissues, including brain (nucleus accumbens, caudate, cortex, and putamen) and artery tissues (tibial and aorta) (Fig. 7a). Notably, rs1914361 minor alleles were positively correlated with ABCC9 expression in brain tissues (Fig. 7b) and negatively correlated with ABCC9 expression in artery tissues (Fig. 7c). Furthermore, relative to rs704178, a previously identified ABCC9 HS SNV, rs1914361 had a similarly strong association with ABCC9 gene expression in GTEx (rs704178: p = 4.00 × 10–13; rs1914361: p = 7.10 × 10–12) and a stronger association with ABCC9 gene expression in BRAINEAC (rs704178: p = 6.80 × 10–4; rs1914361: p = 2.10 × 10–7) (Table 3). In INFERNO, these prioritized SNVs were associated with Roadmap enhancers in adipose, blood vessel, connective, heart, live, skeletal muscle, and smooth muscle tissues, among others.
Fig. 6.

Variant-level results for ABCC9. Adjusted, meta-analytic, single nucleotide variant (SNV)-level p-values for hippocampal sclerosis (HS) and limbic-predominant age-related TDP-43 encephalopathy neuropathological change (LATE-NC) across ABCC9 ± 10 kb assuming a recessive mode of inheritance (MOI). A recessive MOI was assumed for ABCC9 since it has consistently been the MOI with the strongest HS association for ABCC9 [43, 48, 29]. All analyses were for sex, age at death, cohort/study, and the first three genetic principal components. The horizontal dashed line represents the Bonferroni-corrected threshold for significance that accounts for the number of independent tests in the ABCC9 ± 10 kb region. A diamond represents the SNV with the smallest p-value. The previously identified ABCC9 SNV [43] is labeled and identified with an arrow. MOI = mode of inheritance; LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological change; HS = hippocampal sclerosis
Fig. 7.

Expression quantitative trait loci (eQTL) analyses for rs1914361 and ABCC9 across tissue types. Expression quantitative trait loci (eQTL) analyses for rs1914361 and ABCC9 gene expression across human tissues in the Genotype-Tissue Expression (GTEx) database. a Multi-tissue eQTL plot of rs1914361 and ABCC9 gene expression; b ABCC9 normalized gene expression stratified by rs1914361 minor alleles in the nucleus acumbens region of the brain; and c ABCC9 normalized gene expression stratified by rs1914361 minor alleles in the aorta region of the artery. GTEx = Genotype-Tissue Expression; NES = normalize effect size; eQTL = expression quantitative trait loci
Table 3.
Most significant expression quantitative trait loci (eQTL) p-values for ABCC9 in BRAINEAC and GTEx databases
| Gene | SNV | Most Significant eQTL P-value | |
|---|---|---|---|
| BRAINEAC | GTEx | ||
| ABCC9 | rs704178 | 6.80E-04 | 4.00E-13 |
| rs1914361 | 2.10E-07 | 7.10E-12 | |
eQTL = expression quantitative trait loci; GTEx = Genotype-Tissue Expression; SNV = single-nucleotide variant
SNV-level regression analyses
In their respective regression models, the GRN SNV rs5848 (p = 0.010), the APOE SNV rs769449 (p < 0.001), and APOE ε4 carrier status (p < 0.001) all had nominally significant adjusted meta-analytic associations with LATE-NC and the TMEM106B SNV rs7781670 had a borderline-significant adjusted meta-analytic association with LATE-NC (p = 0.057) (Table 4). All odds ratio estimates were consistent across NACC and ROSMAP with the exception of the ABCC9 SNV rs1914361 when assuming a recessive MOI (NACC: OR = 0.98; ROSMAP: OR = 1.40). Notably, the odds ratio estimates for LATE-NC were very similar between the APOE SNV rs769449 (meta-analytic OR = 1.95) and APOE ε4 carrier status (meta-analytic OR = 2.05), which likely reflects the fact that rs769449 minor allele counts are strongly correlated with APOE ε4 counts (NACC: r2 = 0.746; ROSMAP: r2 = 0.712). In a sensitivity analysis with smaller sample sizes, the TMEM106B SNV rs1990622, but none of the other tested SNVs, had nominally significant adjusted associations with TDP-43 in the amygdala (LATE-NC Stage 1 vs. LATE-NC Stage 0); see Additional File 1: Supplemental Table 6.
Table 4.
Adjusted limbic predominant age-related TDP-43 encephalopathy neuropathological changes (LATE-NC) odds ratios for risk variants
| Gene | MOI | SNV | Effect Allele | NACC | ROSMAP | Meta-Analysis | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| OR | P-value | OR | P-value | OR | 95% CI | P-value | ||||
| TMEM106B | Additive | rs1990622 | A | 1.39 | 0.051 | 1.08 | 0.484 | 1.16 | (0.97, 1.39) | 0.099 |
| TMEM106B | Additive | rs7781670 | C | 1.47 | 0.024 | 1.09 | 0.415 | 1.19 | (1.00, 1.43) | 0.057 |
| GRN | Additive | rs5848 | T | 1.40 | 0.042 | 1.23 | 0.089 | 1.29 | (1.06, 1.56) | 0.010 |
| ABCC9 | Additive | rs1914361 | G | 1.16 | 0.354 | 1.16 | 0.171 | 1.16 | (0.97, 1.39) | 0.098 |
| ABCC9 | Recessive | rs1914361 | G | 0.98 | 0.933 | 1.40 | 0.077 | 1.25 | (0.92, 1.71) | 0.151 |
| ABCC9 | Additive | rs704178 | G | 0.95 | 0.764 | 1.07 | 0.536 | 1.03 | (0.86, 1.24) | 0.732 |
| ABCC9 | Recessive | rs704178 | G | 0.80 | 0.433 | 1.16 | 0.394 | 1.05 | (0.78, 1.40) | 0.759 |
| APOE | Additive | rs769449 | A | 1.70 | 0.004 | 2.22 | < 0.001 | 1.95 | (1.51, 2.52) | < 0.001 |
| APOE | N/A | e4 Carrier | N/A | 1.88 | 0.010 | 2.16 | < 0.001 | 2.05 | (1.54, 2.74) | < 0.001 |
Adjusted effects of single nucleotide variants (SNV) on limbic-predominant age-related TDP-43 encephalopathy neuropathological change (LATE-NC). All models adjust for sex, age at death, first three principal components and cohort/study. For rs1990622, rs7781670, and rs704178, the effect alleles are the risk-associated alleles and not the minor alleles. NACC = National Alzheimer's Coordinating Center; ROSMAP = Religious Orders Study and Rush Memory and Aging Project; MOI = mode of inheritance; SNV = single-nucleotide variant; OR = odds ratio; CI = confidence interval.
No LATE-NC-by-SNV interactions were significant in the adjusted HS models, so the interaction terms were removed. The TMEM106B SNVs (rs1990622 and rs7781670), the GRN SNV (rs5848), one of the ABCC9 SNVs (rs1914361), the APOE SNV (rs769449), and APOE ε4 carrier status all had nominally significant, adjusted meta-analytic associations with HS (Table 5). When these models were adjusted for LATE-NC, all models had nominally significant adjusted meta-analytic associations with HS with the exception of the APOE SNV (rs769449) and APOE ε4 carrier status (Table 5, Fig. 8), suggesting that the association between APOE status and HS is related to a more direct interaction between APOE and LATE-NC (i.e., TDP-43 proteinopathy). By contrast, the association between HS and the ABCC9 SNV rs704178 becomes nominally significant with larger odds ratio estimates when adjusted for LATE-NC.
Table 5.
Hippocampal sclerosis (HS) odds ratios for risk variants with and without adjustment for LATE-NC status
| Gene | MOI | SNV | Effect Allele | LATE-NC Adjusted | NACC | ROSMAP | Meta-Analysis | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| OR | P-value | OR | P-value | OR | 95% CI | P-value | |||||
| TMEM106B | Additive | rs1990622 | A | Yes | 1.23 | 0.396 | 1.61 | 0.017 | 1.44 | (1.07, 1.95) | 0.017 |
| No | 1.43 | 0.044 | 1.55 | 0.019 | 1.49 | (1.15, 1.91) | 0.002 | ||||
| TMEM106B | Additive | rs7781670 | C | Yes | 1.20 | 0.456 | 1.53 | 0.029 | 1.39 | (1.03, 1.88) | 0.030 |
| No | 1.47 | 0.034 | 1.50 | 0.028 | 1.48 | (1.15, 1.91) | 0.002 | ||||
| GRN | Additive | rs5848 | T | Yes | 1.37 | 0.168 | 1.37 | 0.123 | 1.37 | (1.02, 1.84) | 0.039 |
| No | 1.67 | 0.004 | 1.43 | 0.057 | 1.56 | (1.21, 2.00) | < 0.001 | ||||
| ABCC9 | Additive | rs1914361 | G | Yes | 1.92 | 0.005 | 1.31 | 0.152 | 1.52 | (1.14, 2.03) | 0.004 |
| No | 1.64 | 0.004 | 1.35 | 0.092 | 1.49 | (1.17, 1.90) | 0.001 | ||||
| ABCC9 | Recessive | rs1914361 | G | Yes | 3.87 | < 0.001 | 1.58 | 0.124 | 2.23 | (1.42, 3.51) | < 0.001 |
| No | 2.69 | < 0.001 | 1.64 | 0.075 | 2.12 | (1.45, 3.09) | < 0.001 | ||||
| ABCC9 | Additive | rs704178 | G | Yes | 1.51 | 0.079 | 1.52 | 0.034 | 1.52 | (1.13, 2.04) | 0.006 |
| No | 1.09 | 0.618 | 1.42 | 0.059 | 1.23 | (0.96, 1.57) | 0.099 | ||||
| ABCC9 | Recessive | rs704178 | G | Yes | 1.77 | 0.121 | 1.48 | 0.171 | 1.58 | (1.02, 2.47) | 0.042 |
| No | 1.33 | 0.292 | 1.43 | 0.180 | 1.38 | (0.95, 2.01) | 0.090 | ||||
| APOE | Additive | rs769449 | A | Yes | 1.15 | 0.589 | 1.54 | 0.094 | 1.33 | (0.93, 1.90) | 0.118 |
| No | 1.30 | 0.188 | 2.02 | 0.004 | 1.54 | (1.14, 2.09) | 0.005 | ||||
| APOE | N/A | e4 Carrier | N/A | Yes | 1.74 | 0.114 | 1.34 | 0.318 | 1.49 | (0.96, 2.31) | 0.075 |
| No | 1.79 | 0.024 | 1.91 | 0.018 | 1.84 | (1.28, 2.66) | 0.001 | ||||
Adjusted effects of single nucleotide variants (SNV) on hippocampal sclerosis (HS). A separate regression model was fit for each variant, mode of inheritance (MOI), and limbic-predominant age-related TDP-43 encephalopathy neuropathological change (LATE-NC) adjustment. All models also adjust for sex, age at death, first three principal components and cohort/study. For rs1990622, rs7781670, and rs704178, the effect alleles are the risk-associated alleles and not the minor alleles. NACC = National Alzheimer's Coordinating Center; ROSMAP = Religious Orders Study and Rush Memory and Aging Project; MOI = mode of inheritance; SNV = single-nucleotide variant; LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological change; OR = odds ratio; CI = confidence interval.
Fig. 8.

Adjusted odds ratios for hippocampal sclerosis (HS) across variants. Adjusted odds ratio estimates and 95% confidence intervals for genetic single nucleotide variants (SNV) and APOE ε4 carrier status from separate regression models of hippocampal sclerosis (HS) fit using data from the National Alzheimer's Coordinating Center (NACC), the Religious Orders Study and Rush Memory and Aging Project (ROSMAP), and the meta-analysis of NACC and ROSMAP. All regression models were adjusted for sex, age at death, cohort/study, and the first three genetic principal components. Regression models were also adjusted for limbic-predominant age-related TDP-43 encephalopathy neuropathological change (LATE-NC) case status by the including LATE-NC status as an additional predictor variable and these odds ratio estimates are represented by triangles. For each variant, the effect allele is defined as the HS risk-causing allele (HA odds ratio estimates > 1.0), and not necessarily the minor allele. An additive mode of inheritance (MOI) is assumed for all variants except for rs704178 where a dominant MOI was assumed (since a recessive MOI resulted in a significant protective effect for HS). HS = hippocampal sclerosis; LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological change; NACC = National Alzheimer's Coordinating Center; ROSMAP = Religious Orders Study and Rush Memory and Aging Project
An issue raised by the ABCC9/HS association results was whether this correlation was driven by cases lacking LATE-NC, i.e. the minority of cases with HS pathology that lacked TDP-43 proteinopathy. A separate sensitivity analysis was performed that excluded the cases with HS pathology that lacked LATE-NC. Results are shown in Additional File 1: Supplemental Table 3, which may be compared with Table 5. The odds ratio estimates for the association between ABCC9 risk variants and HS pathology was essentially unchanged by removing the LATE-NC-HS + cases.
Additional sensitivity analyses were performed testing if the top LATE/HS-related SNVs were associated with ADNC, i.e. Braak NFT stages or CERAD neuritic plaque densities. In these analyses, only the APOE SNV (rs769449) and APOE ε4 carrier status were found to be associated with ADNC (Additional File 1: Supplemental Tables 7 and 8). These results suggest that the associations between HS and LATE-NC and the non-APOE SNVs were likely independent of ADNC.
Discussion
Using large genetic data sets with complementary autopsy-derived data, we demonstrated that the neuropathological endophenotypes of LATE-NC and HS showed replication for associations with a number of previously identified risk genes. The strong association between TMEM106B and TDP-43 proteinopathy—including LATE-NC Stage 1—was once again replicated. Interestingly, ABCC9 was not associated with LATE-NC but was associated with HS pathology. Our study adds to the growing body of literature on the overlapping genetics of HS and LATE-NC while also highlighting several genetic loci unique to each disease entity.
We replicated significant gene-based associations between HS and the TMEM106B, ABCC9, GRN, and APOE genes along with the rs7781670 (TMEM106B) and rs5848 (GRN) SNVs. Furthermore, we identified novel SNV-level associations between LATE-NC and rs7781670 and rs769449. The association of LATE-NC and rs7781670 is intriguing since it was also recently associated with clinical AD in a large AD GWAS [9]. We found no evidence to support the hypothesis that KCNMB2 is a risk gene for either LATE-NC or HS pathologies. However, we note that the sample size of the present study was suitable to detect only relatively large genotype/phenotype associations.
There is an emerging consensus that mixed pathologies are highly prevalent in elderly populations, and there are complex relationships between genotypes and the downstream pathologies. The finding that variants in GRN, TMEM106B, and APOE genes are associated with several neuropathological endophenotypes fits in with recent studies looking at genetic pleiotropy in neurological conditions [12]. Pleiotropic effects have been observed among AD-related neuropathological changes like neuritic plaques, neurofibrillary tangles, and cerebral amyloid angiopathy [13] as well as between LATE-NC and FTLD-TDP [42].
Since the large majority of HS cases were also LATE-NC cases in the current study (Fig. 3), it was striking that some risk genes and SNVs were found to only be associated with HS and not LATE-NC—and vice versa, when statistical models were applied. We did identify several genes that are associated with both neuropathologic endophenotypes. Specifically, the TMEM106B, GRN, and APOE SNVs appear to predispose individuals to LATE-NC (Fig. 10a). Our data indicate that the associations between HS and SNVs in the TMEM106B, GRN, and ABCC9 genes remain statistically significant in a model that adjusts for the presence of LATE-NC (Table 5). However, the impact of TMEM106B and GRN on HS appeared to be attenuated in a statistical model that included TDP-43 proteinopathy, suggesting that their impact on HS may be mediated by their role in LATE-NC. How these genetic SNVs can impact HS secondarily or independently of LATE-NC is not currently known.
Fig. 10.

Diagrams depicting potential causal relationships between the genes under study with positive findings. Diagrams depicting potential causal relationships between the genes under study with positive findings (TMEM106B, ABCC9, GRN, and APOE) and TDP-43 proteinopathy/limbic-predominant age-related TDP-43 encephalopathy (LATE), hippocampal sclerosis (HS), and Alzheimer’s disease (AD). a The candidate genes and their corresponding colors in the diagrams, b a diagram of the current study’s prima facie results, and c a diagram showing hypothetical mechanistic pathways that are compatible with the findings of the current study, including how AD neuropathologic changes (often linked to the APOE risk allele) may fit in with the current study’s results. LATE = limbic-predominant age-related TDP-43 encephalopathy; HS = hippocampal sclerosis; AD = Alzheimer’s disease
While several ABCC9 SNVs have been found to be associated with HS, including rs704178 and rs704180, this is the first study to report an association between the ABCC9 SNV rs1914361 and HS. Notably, rs1914361 was found to be associated with HS in two of the three included cohorts of the original HS GWAS [43], but it was not included in the downstream analyses since its association with HS wasn’t nominally significant in all three cohorts (data not published). It is important to note that prior studies involved completely different sets of included participants (no overlap, as verified with computational methods) but the “direction” of the effect in all cohorts studied was the same. Since rs1914361 was found to also be significantly associated with the expression of ABCC9 (Table 3) and is not in strong linkage disequilibrium with rs704178 (r2 = 0.176), the two loci may represent independent ABCC9 HS risk SNVs.
We also identified divergent patterns in the tissue-level gene expressions of ABCC9 and its homologous gene, ABCC8 (Fig. 9). The proteins encoded by both of these genes function to help regulate the “KATP” potassium channels, which serve as molecular sensors helping to match metabolic needs with cellular reactivity [45, 51]. Multiple lines of evidence link ABCC9 with blood vessels in normal and disease states. In the present study, ABCC9 appeared to be relatively highly expressed in vascular and smooth muscle tissues (Fig. 9), and the correlative impact of the ABCC9/HS risk allele differed in blood vessels in comparison to brain tissue (Fig. 7). Further, the risk-related allele was associated with lower expression of ABCC9 in blood vessels (Fig. 7). ABCC9 has previously been shown to be a marker of vascular mural cells (e.g., pericytes and smooth muscle cells)[2, 5, 10, 30, 64] and the protein product has been implicated in modulating blood flow [45, 51, 61]. In terms of highly penetrant genetic variants, ABCC9 toxic gain-of-function mutations are linked to Cantu Syndrome, a complex phenotype that includes tortuous cerebral blood vessel patterns [31, 32]. ABCC9 loss-of-function mutations cause ABCC9-related Intellectual disability Myopathy Syndrome (AIMS), another complex condition that includes intellectual disability with white matter hyperintensities detected by MRI, even in teenagers [63]. Thus, ABCC9 dysregulation may partly underlie the observation (i.e., may help to explain the phenomenon) that arteriolosclerosis is more severe in brains with LATE/HS than non-LATE/HS brains [1, 22, 24, 28, 49].
Fig. 9.

ABCC8 and ABCC9 gene expression across tissue types. ABCC8 and ABCC9 gene expression in various human tissues in the a Genotype-Tissue Expression (GTEx) and b Functional Annotation of Human Long Noncoding RNAs via Molecular Mapping (FANTOM5) databases. In GTEx, central nervous system (CNS) tissues included Brodmann (1909) area 24, Brodmann (1909) area 9, C1 segment of cervical spinal cord, amygdala, caudate nucleus, cerebellar hemisphere, cerebellum, cerebral cortex, hippocampus proper, hypothalamus, nucleus accumbens, pituitary gland, and substantia nigra; vascular/smooth muscle tissues included aorta, atrium auricular region, coronary artery, tibial artery, endocervix, esophagus muscularis mucosa, urinary bladder, and uterus; and other tissues included all other tissue types. In FANTOM5, CNS tissues included amygdala, brain, caudate nucleus, cerebellum, diencephalon, dorsal thalamus, globus pallidus, hippocampal formation, locus ceruleus, medulla oblongata, middle frontal gyrus, middle temporal gyrus, occipital cortex, occipital lobe, olfactory apparatus, parietal lobe, pituitary gland, putamen, spinal cord, and substantia nigra; vascular/smooth muscle tissue included artery, heart, heart left ventricle, left cardiac atrium, mitral valve, smooth muscle, tricuspid valve, and uterus; and other tissues included all other tissue types. GTEx = Genotype-Tissue Expression; FANTOM5 = Functional Annotation of Human Long Noncoding RNAs via Molecular Mapping; TPM = transcripts per million; CNS = central nervous system
The current study adds to a growing body of literature suggesting that LATE-NC is a potential precursor to HS [42]. It is yet to be seen how exactly the APOE gene and AD-type changes interact with other pathologies, but one hypothesis is that APOE and AD predispose an individual to LATE-NC, which then drives an individual towards severe LATE-NC and HS (Fig. 10). It has been found that TDP-43 proteinopathy localizes to tangle-like structures in many cases with ADNC [26]. Further autopsy-based studies with larger sample sizes are needed.
There are both limitations and strengths to the present study. Because of the characteristics of the sample (largely Caucasian, drawn from a number of different research centers), the degree to which findings are generalizable is unknown, especially with respect to individuals of other ancestries. While this work aims to replicate previous associations, there are many models considered which can inflate false positive rates. Additionally, it can be difficult to show that the associations identified in the current study are independent of ADNC, but several sensitivity analyses provide evidence that at least the non-APOE associations are likely independent of ADNC. Further follow-up studies are needed to investigate the significant associations between APOE and LATE-NC, though even if null this association would still highlight the strong associations that exist between AD and other neurodegenerative diseases, which is interesting in itself. We also note that all the included subjects had high-quality neuropathologic workup for TDP-43 proteinopathy and HS, and all the ADGC subjects were autopsied during 2014 and later. These study design elements constitute strengths of the current study.
Supplementary Information
Additional file 1. Supplemental Table 1 for a summary of the rare conditions excluded from the NACC sample; these conditions are extremely rare among ROSMAP participants. ROSMAP participants included in the Nelson et al. 2014 hippocampal sclerosis (HS) genome wise association study (GWAS) were explicitly excluded from the current study; NACC participants were only included in the current study if version 10 NACC neuropathology (NP) data were available, which were not collected until after 2014. LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological change; HS = hippocampal sclerosis; NACC = National Alzheimer's Coordinating Center; ROSMAP = Religious Orders Study and Rush Memory and Aging Project; GWAS = genome wide association study.
Acknowledgements
We are profoundly grateful to research participants, clinicians, and colleague researchers that helped donate, curate, and analyze the data. The NACC database is funded by NIA/NIH Grant U01 AG016976. NACC data are contributed by the NIA-funded ADCs: P30 AG019610 (PI Eric Reiman, MD), P30 AG013846 (PI Neil Kowall, MD), P30 AG062428-01 (PI James Leverenz, MD) P50 AG008702 (PI Scott Small, MD), P50 AG025688 (PI Allan Levey, MD, PhD), P50 AG047266 (PI Todd Golde, MD, PhD), P30 AG010133 (PI Andrew Saykin, PsyD), P50 AG005146 (PI Marilyn Albert, PhD), P30 AG062421-01 (PI Bradley Hyman, MD, PhD), P30 AG062422-01 (PI Ronald Petersen, MD, PhD), P50 AG005138 (PI Mary Sano, PhD), P30 AG008051 (PI Thomas Wisniewski, MD), P30 AG013854 (PI Robert Vassar, PhD), P30 AG008017 (PI Jeffrey Kaye, MD), P30 AG010161 (PI David Bennett, MD), P50 AG047366 (PI Victor Henderson, MD, MS), P30 AG010129 (PI Charles DeCarli, MD), P50 AG016573 (PI Frank LaFerla, PhD), P30 AG062429-01(PI James Brewer, MD, PhD), P50 AG023501 (PI Bruce Miller, MD), P30 AG035982 (PI Russell Swerdlow, MD), P30 AG028383 (PI Linda Van Eldik, PhD), P30 AG053760 (PI Henry Paulson, MD, PhD), P30 AG010124 (PI John Trojanowski, MD, PhD), P50 AG005133 (PI Oscar Lopez, MD), P50 AG005142 (PI Helena Chui, MD), P30 AG012300 (PI Roger Rosenberg, MD), P30 AG049638 (PI Suzanne Craft, PhD), P50 AG005136 (PI Thomas Grabowski, MD), P30 AG062715-01 (PI Sanjay Asthana, MD, FRCP), P50 AG005681 (PI John Morris, MD), P50 AG047270 (PI Stephen Strittmatter, MD, PhD). The Genotype-Tissue Expression (GTEx) Project was supported by the Common Fund of the Office of the Director of the National Institutes of Health, and by NCI, NHGRI, NHLBI, NIDA, NIMH, and NINDS. The data used for the analyses described in this manuscript were obtained from the GTEx Portal (https://www.gtexportal.org/home/datasets) on 9/17/2020. The National Institutes of Health, National Institute on Aging (NIH-NIA) supported this work through the following grants: ADGC, U01 AG032984, RC2 AG036528; Samples from the National Cell Repository for Alzheimer’s Disease (NCRAD), which receives government support under a cooperative agreement grant (U24 AG21886) awarded by the National Institute on Aging (NIA), were used in this study. We thank contributors who collected samples used in this study, as well as patients and their families, whose help and participation made this work possible; Data for this study were prepared, archived, and distributed by the National Institute on Aging Alzheimer’s Disease Data Storage Site (NIAGADS) at the University of Pennsylvania (U24-AG041689-01); NACC, U01 AG016976; NIA LOAD, U24 AG026395, R01AG041797; Banner Sun Health Research Institute P30 AG019610; Boston University, P30 AG013846, U01 AG10483, R01 CA129769, R01 MH080295, R01 AG017173, R01 AG025259, R01AG33193; Columbia University, P50 AG008702, R37 AG015473; Duke University, P30 AG028377, AG05128; Emory University, AG025688; Group Health Research Institute, UO1 AG006781, UO1 HG004610, UO1 HG006375; Indiana University, P30 AG10133; Johns Hopkins University, P50 AG005146, R01 AG020688; Massachusetts General Hospital, P50 AG005134; Mayo Clinic, P50 AG016574; Mount Sinai School of Medicine, P50 AG005138, P01 AG002219; New York University, P30 AG08051, UL1 RR029893, 5R01AG012101, 5R01AG022374, 5R01AG013616, 1RC2AG036502, 1R01AG035137; Northwestern University, P30 AG013854; Oregon Health & Science University, P30 AG008017, R01 AG026916; Rush University, P30 AG010161, R01 AG019085, R01 AG15819, R01 AG17917, R01 AG30146; TGen, R01 NS059873; University of Alabama at Birmingham, P50 AG016582; University of Arizona, R01 AG031581; University of California, Davis, P30 AG010129; University of California, Irvine, P50 AG016573; University of California, Los Angeles, P50 AG016570; University of California, San Diego, P50 AG005131; University of California, San Francisco, P50 AG023501, P01 AG019724; University of Kentucky, P30 AG028383, AG05144; University of Michigan, P50 AG008671; University of Pennsylvania, P30 AG010124; University of Pittsburgh, P50 AG005133, AG030653, AG041718, AG07562, AG02365; University of Southern California, P50 AG005142; University of Texas Southwestern, P30 AG012300; University of Miami, R01 AG027944, AG010491, AG027944, AG021547, AG019757; University of Washington, P50 AG005136; University of Wisconsin, P50 AG033514; Vanderbilt University, R01 AG019085; and Washington University, P50 AG005681, P01 AG03991. The Kathleen Price Bryan Brain Bank at Duke University Medical Center is funded by NINDS grant # NS39764, NIMH MH60451 and by Glaxo Smith Kline. Genotyping of the TGEN2 cohort was supported by Kronos Science. The TGen series was also funded by NIA grant AG041232 to AJM and MJH, The Banner Alzheimer’s Foundation, The Johnnie B. Byrd Sr. Alzheimer’s Institute, the Medical Research Council, and the state of Arizona and also includes samples from the following sites: Newcastle Brain Tissue Resource (funding via the Medical Research Council, local NHS trusts and Newcastle University), MRC London Brain Bank for Neurodegenerative Diseases (funding via the Medical Research Council),South West Dementia Brain Bank (funding via numerous sources including the Higher Education Funding Council for England (HEFCE), Alzheimer’s Research Trust (ART), BRACE as well as North Bristol NHS Trust Research and Innovation Department and DeNDRoN), The Netherlands Brain Bank (funding via numerous sources including Stichting MS Research, Brain Net Europe, Hersenstichting Nederland Breinbrekend Werk, International Parkinson Fonds, Internationale Stiching Alzheimer Onderzoek), Institut de Neuropatologia, Servei Anatomia Patologica, Universitat de Barcelona. ADNI data collection and sharing was funded by the National Institutes of Health Grant U01 AG024904 and Department of Defense award number W81XWH-12-2-0012. ADNI is funded by the National Institute on Aging, the National Institute of Biomedical Imaging and Bioengineering, and through generous contributions from the following: AbbVie, Alzheimer’s Association; Alzheimer’s Drug Discovery Foundation; Araclon Biotech; BioClinica, Inc.; Biogen; Bristol-Myers Squibb Company; CereSpir, Inc.; Eisai Inc.; Elan Pharmaceuticals, Inc.; Eli Lilly and Company; EuroImmun; F. Hoffmann-La Roche Ltd and its affiliated company Genentech, Inc.; Fujirebio; GE Healthcare; IXICO Ltd.; Janssen Alzheimer Immunotherapy Research & Development, LLC.; Johnson & Johnson Pharmaceutical Research & Development LLC.; Lumosity; Lundbeck; Merck & Co., Inc.; Meso Scale Diagnostics, LLC.; NeuroRx Research; Neurotrack Technologies; Novartis Pharmaceuticals Corporation; Pfizer Inc.; Piramal Imaging; Servier; Takeda Pharmaceutical Company; and Transition Therapeutics. The Canadian Institutes of Health Research is providing funds to support ADNI clinical sites in Canada. Private sector contributions are facilitated by the Foundation for the National Institutes of Health (www.fnih.org). The grantee organization is the Northern California Institute for Research and Education, and the study is coordinated by the Alzheimer's Disease Cooperative Study at the University of California, San Diego. ADNI data are disseminated by the Laboratory for Neuro Imaging at the University of Southern California. We thank Drs. D. Stephen Snyder and Marilyn Miller from NIA who are ex-officio ADGC members. Support was also from the Alzheimer’s Association (LAF, IIRG-08-89720; MP-V, IIRG-05-14147) and the US Department of Veterans Affairs Administration, Office of Research and Development, Biomedical Laboratory Research Program. P.S.G.-H. is supported by Wellcome Trust, Howard Hughes Medical Institute, and the Canadian Institute of Health Research.
Abbreviations
- ACAT
Aggregated Cauchy association test
- AD
Alzheimer’s disease
- ADGC
Alzheimer’s disease genetics consortium
- ADNC
Alzheimer’s disease neuropathological changes
- ADRD
Alzheimer’s disease and related dementias
- ALS
Amyotrophic lateral sclerosis
- AMP-AD
Accelerating Medicines Partnership-Alzheimer’s Disease
- eQTL
Expression quantitative trait loci
- TLD
Frontotemporal lobar degeneration
- FTLD-TDP
Frontotemporal lobe degeneration with TDP-43
- GTEx
Genotype-Tissue Expression
- GWAS
Genome wide association study
- HS
Hippocampal sclerosis
- HWE
Hardy–Weinberg equilibrium
- IRB
Institutional Review
- LATE
Limbic-predominant age-related TDP-43 encephalopathy
- LATE-NC
Limbic-predominant age-related TDP-43 encephalopathy neuropathological changes
- MOI
Mode of inheritance
- NACC
National Alzheimer’s Coordinating Center
- NP
Neuropathology
- OR
Odds ratio
- PC
Principal component
- QC
Quality control
- ROSMAP
Religious Orders Study and the Rush Memory and Aging Project
- SNV
Single-nucleotide variant
- TDP-43
TAR-DNA binding protein 43
Authors' contributions
AJD conceptualized the study, analyzed data, wrote the first draft manuscript, generated graphics, and co-authored the manuscript. PTN conceptualized the study, analyzed data, generated graphics, and co-authored the manuscript. YK and LMPS created analytic pipelines used in the analysis and co-authored the manuscript. KLB, SM, and JSKK imputed the genomic data used in the analysis and co-authored the manuscript. MAT coauthored the manuscript and reviewed the text before submission. MDC coauthored the manuscript and reviewed the text before submission. TJH coauthored the manuscript and reviewed the text before submission. JAS coauthored the manuscript and reviewed the text before submission. ADGC critically reviewed the text before submission. DWF conceptualized the study, helped generate framework and details of for data analyses, and co-authored the manuscript. All authors read and approved the final manuscript.
Funding
This work was supported by the National Institutes of Health [R56-AG057191, R01-AG057187, P30-AG028383, R01-AG059716, K01-AG049164].
Availability of data and materials
NACC data are available upon request via NACC’s website (https://naccdata.org/). ROSMAP data are available online at the Rush Alzheimer’s Disease Center Resource Sharing Hub (https://www.radc.rush.edu/), as well as on the Accelerating Medicines Partnership-Alzheimer’s Disease (AMP-AD) Knowledge Portal (syn3219045). ADGC data are available upon request via the NIAGADS website (https://www.niagads.org/user/login?destination=data/request/new_request/).
Declarations
Ethics approval and consent to participate
All study data were provided, along with data use agreements, from anonymized central data repositories, and each study had IRB approval from the Institutional Review Board (IRB) at the local institution where the research volunteer was recruited for participation. Each study participant signed a consent form for inclusion in the study (allowing her or his data to be stored as a resource in the central data repository), and the data use agreements are compatible with those consent forms.
Consent for publication
Not applicable.
Competing interests
Timothy Hohman is on the Scientific Advisory Board for Vivid Genomics. No other authors have conflicts of interest to report.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Adam J. Dugan, Email: adam.dugan@uky.edu
Peter T. Nelson, Email: pnels2@uky.edu
Yuriko Katsumata, Email: katsumata.yuriko@uky.edu.
Lincoln M. P. Shade, Email: lincoln.shade@uky.edu
Kevin L. Boehme, Email: kevinlboehme@gmail.com
Merilee A. Teylan, Email: mateylan@uw.edu
Matthew D. Cykowski, Email: mdcykowski@houstonmethodist.org
Shubhabrata Mukherjee, Email: smukherj@uw.edu.
John S. K. Kauwe, Email: kauwe@byu.edu
Timothy J. Hohman, Email: timothy.j.hohman@vumc.org
Julie A. Schneider, Email: julie_a_schneider@rush.edu
David W. Fardo, Email: david.fardo@uky.edu
References
- 1.Agrawal S, Yu L, Kapasi A, James BD, Arfanakis K, Barnes LL, et al. Limbic-predominant age-related TDP-43 encephalopathy neuropathologic change and microvascular pathologies in community-dwelling older persons. Brain Pathol. 2021;31:e12939. doi: 10.1111/bpa.12939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Al Feteisi H, Al-Majdoub ZM, Achour B, Couto N, Rostami-Hodjegan A, Barber J. Identification and quantification of blood-brain barrier transporters in isolated rat brain microvessels. J Neurochem. 2018;146:670–685. doi: 10.1111/jnc.14446. [DOI] [PubMed] [Google Scholar]
- 3.Amador-Ortiz C, Lin WL, Ahmed Z, Personett D, Davies P, Duara R, et al. TDP-43 immunoreactivity in hippocampal sclerosis and Alzheimer's disease. Ann Neurol. 2007;61:435–445. doi: 10.1002/ana.21154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Amlie-Wolf A, Tang M, Mlynarski EE, Kuksa PP, Valladares O, Katanic Z, et al. INFERNO: inferring the molecular mechanisms of noncoding genetic variants. Nucl Acids Res. 2018;46:8740–8753. doi: 10.1093/nar/gky686. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ando K, Wang W, Peng D, Chiba A, Lagendijk AK, Barske L, et al. Peri-arterial specification of vascular mural cells from naïve mesenchyme requires Notch signaling. Development. 2019 doi: 10.1242/dev.165589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Aoki N, Murray ME, Ogaki K, Fujioka S, Rutherford NJ, Rademakers R, et al. Hippocampal sclerosis in Lewy body disease is a TDP-43 proteinopathy similar to FTLD-TDP Type A. Acta Neuropathol. 2015;129:53–64. doi: 10.1007/s00401-014-1358-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Balduzzi S, Rücker G, Schwarzer G. How to perform a meta-analysis with R: a practical tutorial. Evidence Based Mental Health. 2019;22:153–160. doi: 10.1136/ebmental-2019-300117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Beecham GW, Hamilton K, Naj AC, Martin ER, Huentelman M, Myers AJ, et al. Genome-wide association meta-analysis of neuropathologic features of Alzheimer's disease and related dementias. PLoS Genet. 2014;10:e1004606. doi: 10.1371/journal.pgen.1004606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Bellenguez C, Küçükali F, Jansen I, Andrade V, Morenau-Grau S, Amin N, et al (2020) Large meta-analysis of genome-wide association studies expands knowledge of the genetic etiology of Alzheimer’s disease and highlights potential translational opportunities. medRxiv: 2020.2010.2001.20200659. 10.1101/2020.10.01.20200659
- 10.Bondjers C, He L, Takemoto M, Norlin J, Asker N, Hellström M, et al. Microarray analysis of blood microvessels from PDGF-B and PDGF-Rbeta mutant mice identifies novel markers for brain pericytes. FASEB J. 2006;20:1703–1705. doi: 10.1096/fj.05-4944fje. [DOI] [PubMed] [Google Scholar]
- 11.Brenowitz WD, Monsell SE, Schmitt FA, Kukull WA, Nelson PT. Hippocampal sclerosis of aging is a key Alzheimer's disease mimic: clinical-pathologic correlations and comparisons with both alzheimer's disease and non-tauopathic frontotemporal lobar degeneration. J Alzheimer's Dis JAD. 2014;39:691–702. doi: 10.3233/jad-131880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Chornenkyy Y, Fardo DW, Nelson PT. Tau and TDP-43 proteinopathies: kindred pathologic cascades and genetic pleiotropy. Lab Invest. 2019;99:993–1007. doi: 10.1038/s41374-019-0196-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chung J, Zhang X, Allen M, Wang X, Ma Y, Beecham G, et al. Genome-wide pleiotropy analysis of neuropathological traits related to Alzheimer’s disease. Alzheimers Res Ther. 2018;10:22. doi: 10.1186/s13195-018-0349-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cohen TJ, Lee VM, Trojanowski JQ. TDP-43 functions and pathogenic mechanisms implicated in TDP-43 proteinopathies. Trends Mol Med. 2011;17:659–667. doi: 10.1016/j.molmed.2011.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Consortium GT The Genotype-Tissue Expression (GTEx) project. Nat Genet. 2013;45:580–585. doi: 10.1038/ng.2653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dickson DW, Baker M, Rademakers R. Common variant in GRN is a genetic risk factor for hippocampal sclerosis in the elderly. Neurodegener Dis. 2010;7:170–174. doi: 10.1159/000289231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Dickson DW, Davies P, Bevona C, Van Hoeven KH, Factor SM, Grober E, et al. Hippocampal sclerosis: a common pathological feature of dementia in very old (> or = 80 years of age) humans. Acta Neuropathol. 1994;88:212–221. doi: 10.1007/bf00293396. [DOI] [PubMed] [Google Scholar]
- 18.Dumitrescu L, Mahoney ER, Mukherjee S, Lee ML, Bush WS, Engelman CD, et al. Genetic variants and functional pathways associated with resilience to Alzheimer’s disease. Brain. 2020;143:2561–2575. doi: 10.1093/brain/awaa209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Eadie MJ. Epilepsy, Ammon's horn sclerosis, and Camille Bouchet. J Hist Neurosci. 2017;26:231–237. doi: 10.1080/0964704x.2016.1224141. [DOI] [PubMed] [Google Scholar]
- 20.Gao X, Starmer J, Martin ER. A multiple testing correction method for genetic association studies using correlated single nucleotide polymorphisms. Genet Epidemiol. 2008;32:361–369. doi: 10.1002/gepi.20310. [DOI] [PubMed] [Google Scholar]
- 21.Guo L, Shorter J. Biology and Pathobiology of TDP-43 and Emergent Therapeutic Strategies. Cold Spring Harb Perspect Med. 2017 doi: 10.1101/cshperspect.a024554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Harrison WT, Lusk JB, Liu B, Ervin JF, Johnson KG, Green CL, et al. Limbic-predominant age-related TDP-43 encephalopathy neuropathological change (LATE-NC) is independently associated with dementia and strongly associated with arteriolosclerosis in the oldest-old. Acta Neuropathol. 2021 doi: 10.1007/s00401-021-02360-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Hokkanen SRK, Kero M, Kaivola K, Hunter S, Keage HAD, Kiviharju A, et al. Putative risk alleles for LATE-NC with hippocampal sclerosis in population- Brain Pathol. 2020;30:364–372. doi: 10.1111/bpa.12773. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ighodaro ET, Abner EL, Fardo DW, Lin AL, Katsumata Y, Schmitt FA, et al. Risk factors and global cognitive status related to brain arteriolosclerosis in elderly individuals. J Cereb Blood Flow Metab. 2017;37:201–216. doi: 10.1177/0271678x15621574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ighodaro ET, Jicha GA, Schmitt FA, Neltner JH, Abner EL, Kryscio RJ, et al. Hippocampal sclerosis of aging can be segmental: two cases and review of the literature. J Neuropathol Exp Neurol. 2015;74:642–652. doi: 10.1097/nen.0000000000000204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Josephs KA, Murray ME, Tosakulwong N, Weigand SD, Serie AM, Perkerson RB, et al. Pathological, imaging and genetic characteristics support the existence of distinct TDP-43 types in non-FTLD brains. Acta Neuropathol. 2019;137:227–238. doi: 10.1007/s00401-018-1951-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Katsumata Y, Abner EL, Karanth S, Teylan MA, Mock CN, Cykowski MD, et al. Distinct clinicopathologic clusters of persons with TDP-43 proteinopathy. Acta Neuropathol. 2020 doi: 10.1007/s00401-020-02211-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Katsumata Y, Fardo DW, Kukull WA, Nelson PT. Dichotomous scoring of TDP-43 proteinopathy from specific brain regions in 27 academic research centers: associations with Alzheimer's disease and cerebrovascular disease pathologies. Acta Neuropathol Commun. 2018;6:142. doi: 10.1186/s40478-018-0641-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Katsumata Y, Nelson PT, Ellingson SR, Fardo DW. Gene-based association study of genes linked to hippocampal sclerosis of aging neuropathology: GRN, TMEM106B, ABCC9, and KCNMB2. Neurobiol Aging. 2017;53:193.e117–193.e125. doi: 10.1016/j.neurobiolaging.2017.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kirkwood PM, Gibson DA, Smith JR, Wilson-Kanamori JR, Kelepouri O, Esnal-Zufiaurre A, et al. Single-cell RNA sequencing redefines the mesenchymal cell landscape of mouse endometrium. FASEB J. 2021;35:e21285. doi: 10.1096/fj.202002123R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Kisilevsky E, Kohly RP, Margolin EA. Dilated and tortuous retinal vessels as a sign of Cantu syndrome. Ophthalmic Genet. 2019;40:453–454. doi: 10.1080/13816810.2019.1666415. [DOI] [PubMed] [Google Scholar]
- 32.Leon Guerrero CR, Pathak S, Grange DK, Singh GK, Nichols CG, Lee JM, et al. Neurologic and neuroimaging manifestations of Cantú syndrome: a case series. Neurology. 2016;87:270–276. doi: 10.1212/wnl.0000000000002861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu Y, Chen S, Li Z, Morrison AC, Boerwinkle E, Lin X. ACAT: a fast and powerful p value combination method for rare-variant analysis in sequencing studies. Am J Hum Genet. 2019;104:410–421. doi: 10.1016/j.ajhg.2019.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Machiela MJ, Chanock SJ. LDlink: a web-based application for exploring population-specific haplotype structure and linking correlated alleles of possible functional variants. Bioinformatics. 2015;31:3555–3557. doi: 10.1093/bioinformatics/btv402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Mackenzie IR, Rademakers R. The role of transactive response DNA-binding protein-43 in amyotrophic lateral sclerosis and frontotemporal dementia. Curr Opin Neurol. 2008;21:693–700. doi: 10.1097/WCO.0b013e3283168d1d. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Mahoney ER, Dumitrescu L, Moore AM, Cambronero FE, De Jager PL, Koran MEI, et al. Brain expression of the vascular endothelial growth factor gene family in cognitive aging and alzheimer’s disease. Mol Psychiatry. 2019 doi: 10.1038/s41380-019-0458-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Marees AT, de Kluiver H, Stringer S, Vorspan F, Curis E, Marie-Claire C, et al. A tutorial on conducting genome-wide association studies: Quality control and statistical analysis. Int J Methods Psychiatr Res. 2018;27:e1608–e1608. doi: 10.1002/mpr.1608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.McCarthy S, Das S, Kretzschmar W, Delaneau O, Wood AR, Teumer A, et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat Genet. 2016;48:1279–1283. doi: 10.1038/ng.3643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Murray ME, Cannon A, Graff-Radford NR, Liesinger AM, Rutherford NJ, Ross OA, et al. Differential clinicopathologic and genetic features of late-onset amnestic dementias. Acta Neuropathol. 2014;128:411–421. doi: 10.1007/s00401-014-1302-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Nag S, Yu L, Capuano AW, Wilson RS, Leurgans SE, Bennett DA, et al. Hippocampal sclerosis and TDP-43 pathology in aging and Alzheimer disease. Ann Neurol. 2015;77:942–952. doi: 10.1002/ana.24388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Nelson PT, Abner EL, Schmitt FA, Kryscio RJ, Jicha GA, Smith CD, et al. Modeling the association between 43 different clinical and pathological variables and the severity of cognitive impairment in a large autopsy cohort of elderly persons. Brain Pathol. 2010;20:66–79. doi: 10.1111/j.1750-3639.2008.00244.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Nelson PT, Dickson DW, Trojanowski JQ, Jack CR, Boyle PA, Arfanakis K, et al. Limbic-predominant age-related TDP-43 encephalopathy (LATE): consensus working group report. Brain. 2019;142:1503–1527. doi: 10.1093/brain/awz099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Nelson PT, Estus S, Abner EL, Parikh I, Malik M, Neltner JH, et al. ABCC9 gene polymorphism is associated with hippocampal sclerosis of aging pathology. Acta Neuropathol. 2014;127:825–843. doi: 10.1007/s00401-014-1282-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nelson PT, Gal Z, Wang WX, Niedowicz DM, Artiushin SC, Wycoff S, et al. TDP-43 proteinopathy in aging: Associations with risk-associated gene variants and with brain parenchymal thyroid hormone levels. Neurobiol Dis. 2019;125:67–76. doi: 10.1016/j.nbd.2019.01.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Nelson PT, Jicha GA, Wang WX, Ighodaro E, Artiushin S, Nichols CG, et al. ABCC9/SUR2 in the brain: Implications for hippocampal sclerosis of aging and a potential therapeutic target. Ageing Res Rev. 2015;24:111–125. doi: 10.1016/j.arr.2015.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Nelson PT, Schmitt FA, Lin Y, Abner EL, Jicha GA, Patel E, et al. Hippocampal sclerosis in advanced age: clinical and pathological features. Brain. 2011;134:1506–1518. doi: 10.1093/brain/awr053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Nelson PT, Smith CD, Abner EL, Wilfred BJ, Wang W-X, Neltner JH, et al. Hippocampal sclerosis of aging, a prevalent and high-morbidity brain disease. Acta Neuropathol. 2013;126:161–177. doi: 10.1007/s00401-013-1154-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Nelson PT, Wang W-X, Partch AB, Monsell SE, Valladares O, Ellingson SR, et al. Reassessment of risk genotypes (GRN, TMEM106B, and ABCC9 variants) associated with hippocampal sclerosis of aging pathology. J Neuropathol Exp Neurol. 2015;74:75–84. doi: 10.1097/NEN.0000000000000151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Neltner JH, Abner EL, Baker S, Schmitt FA, Kryscio RJ, Jicha GA, et al. Arteriolosclerosis that affects multiple brain regions is linked to hippocampal sclerosis of ageing. Brain. 2014;137:255–267. doi: 10.1093/brain/awt318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Neumann M, Sampathu DM, Kwong LK, Truax AC, Micsenyi MC, Chou TT, et al. Ubiquitinated TDP-43 in frontotemporal lobar degeneration and amyotrophic lateral sclerosis. Science. 2006;314:130–133. doi: 10.1126/science.1134108. [DOI] [PubMed] [Google Scholar]
- 51.Nichols CG. KATP channels as molecular sensors of cellular metabolism. Nature. 2006;440:470–476. doi: 10.1038/nature04711. [DOI] [PubMed] [Google Scholar]
- 52.Ou SH, Wu F, Harrich D, García-Martínez LF, Gaynor RB. Cloning and characterization of a novel cellular protein, TDP-43, that binds to human immunodeficiency virus type 1 TAR DNA sequence motifs. J Virol. 1995;69:3584–3596. doi: 10.1128/jvi.69.6.3584-3596.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Pruim RJ, Welch RP, Sanna S, Teslovich TM, Chines PS, Gliedt TP, et al. LocusZoom: regional visualization of genome-wide association scan results. Bioinformatics. 2010;26:2336–2337. doi: 10.1093/bioinformatics/btq419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MAR, Bender D, et al. PLINK: a tool set for whole-genome association and population-based linkage analyses. Am J Hum Genet. 2007;81:559–575. doi: 10.1086/519795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.R Core Team (2021) R: a language and environment for statistical computing. 4.0.4 edn. R Foundation for Statistical Computing, City
- 56.Rademakers R, Eriksen JL, Baker M, Robinson T, Ahmed Z, Lincoln SJ, et al. Common variation in the miR-659 binding-site of GRN is a major risk factor for TDP43-positive frontotemporal dementia. Hum Mol Genet. 2008;17:3631–3642. doi: 10.1093/hmg/ddn257. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ramasamy A, Trabzuni D, Guelfi S, Varghese V, Smith C, Walker R, et al. Genetic variability in the regulation of gene expression in ten regions of the human brain. Nat Neurosci. 2014;17:1418–1428. doi: 10.1038/nn.3801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ramey JA (2012) clusteval: Evaluation of Clustering Algorithms. 0.1 edn, City
- 59.Robinson AC, Davidson YS, Horan MA, Pendleton N, Mann DMA. Pathological correlates of cognitive impairment in the university of manchester longitudinal study of cognition in normal healthy old age. J Alzheimer's Dis. 2018;64:483–496. doi: 10.3233/JAD-180171. [DOI] [PubMed] [Google Scholar]
- 60.Robinson JL, Lee EB, Xie SX, Rennert L, Suh E, Bredenberg C, et al. Neurodegenerative disease concomitant proteinopathies are prevalent, age-related and APOE4-associated. Brain. 2018;141:2181–2193. doi: 10.1093/brain/awy146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Rocha MP, Campos MO, Mattos JD, Mansur DE, Rocha HNM, Secher NH, et al. K(ATP) channels modulate cerebral blood flow and oxygen delivery during isocapnic hypoxia in humans. J Physiol. 2020;598:3343–3356. doi: 10.1113/jp279751. [DOI] [PubMed] [Google Scholar]
- 62.Rutherford NJ, Carrasquillo MM, Li M, Bisceglio G, Menke J, Josephs KA, et al. TMEM106B risk variant is implicated in the pathologic presentation of Alzheimer disease. Neurology. 2012;79:717–718. doi: 10.1212/WNL.0b013e318264e3ac. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Smeland MF, McClenaghan C, Roessler HI, Savelberg S, Hansen GÅM, Hjellnes H, et al. ABCC9-related Intellectual disability Myopathy Syndrome is a K(ATP) channelopathy with loss-of-function mutations in ABCC9. Nat Commun. 2019;10:4457. doi: 10.1038/s41467-019-12428-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Song HW, Foreman KL, Gastfriend BD, Kuo JS, Palecek SP, Shusta EV. Transcriptomic comparison of human and mouse brain microvessels. Sci Rep. 2020;10:12358. doi: 10.1038/s41598-020-69096-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Van Langenhove T, van der Zee J, Van Broeckhoven C. The molecular basis of the frontotemporal lobar degeneration-amyotrophic lateral sclerosis spectrum. Ann Med. 2012;44:817–828. doi: 10.3109/07853890.2012.665471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Wennberg AM, Tosakulwong N, Lesnick TG, Murray ME, Whitwell JL, Liesinger AM, et al. Association of Apolipoprotein E ε4 with transactive response DNA-binding protein 43. JAMA Neurol. 2018;75:1347–1354. doi: 10.1001/jamaneurol.2018.3139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Wickham H. ggplot2: Elegant Graphics for Data Analysis. New York, City: Springer-Verlag; 2016. [Google Scholar]
- 68.Yang HS, Yu L, White CC, Chibnik LB, Chhatwal JP, Sperling RA, et al. Evaluation of TDP-43 proteinopathy and hippocampal sclerosis in relation to APOE ε4 haplotype status: a community-based cohort study. Lancet Neurol. 2018;17:773–781. doi: 10.1016/s1474-4422(18)30251-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Additional file 1. Supplemental Table 1 for a summary of the rare conditions excluded from the NACC sample; these conditions are extremely rare among ROSMAP participants. ROSMAP participants included in the Nelson et al. 2014 hippocampal sclerosis (HS) genome wise association study (GWAS) were explicitly excluded from the current study; NACC participants were only included in the current study if version 10 NACC neuropathology (NP) data were available, which were not collected until after 2014. LATE-NC = limbic-predominant age-related TDP-43 encephalopathy neuropathological change; HS = hippocampal sclerosis; NACC = National Alzheimer's Coordinating Center; ROSMAP = Religious Orders Study and Rush Memory and Aging Project; GWAS = genome wide association study.
Data Availability Statement
NACC data are available upon request via NACC’s website (https://naccdata.org/). ROSMAP data are available online at the Rush Alzheimer’s Disease Center Resource Sharing Hub (https://www.radc.rush.edu/), as well as on the Accelerating Medicines Partnership-Alzheimer’s Disease (AMP-AD) Knowledge Portal (syn3219045). ADGC data are available upon request via the NIAGADS website (https://www.niagads.org/user/login?destination=data/request/new_request/).