

Abstract
Recent emerging research on intestinal microbiota and its contribution to the central nervous system during health and disease has attracted significant attention. Age-related intestinal microbiota changes initiate brain aging and age-related neurodegenerative disorders. Aging is one of the critical predisposing risk factors for the development of neurodegenerative diseases. Maintaining a healthy gut microbiota is essential for a healthy body and aging, but dysbiosis could initiate many chronic diseases. Understanding the underlying mechanisms of gut microbiota alterations/dysbiosis will help identify biomarkers for aging-related chronic conditions. This review summarizes recent advances in microbiota-neurodegenerative disease research and will enhance our understanding of gut microbiota dysbiosis and its effects on brain aging.
Key Words: brain aging, cognitive decline, dysbiosis, fecal microbiota transplantation, gut-microbiota, neuroinflammation, prebiotics, probiotics
Introduction
“All Diseases Begin in the Gut,” proclaimed by the father of modern medicine, Hippocrates, more than 2000 years ago (Lyon, 2018), still holds its relevance in maintaining optimal health situation and microbiota dysbiosis. Over the past several years, a healthy digestive system’s power and potential in modulating whole body functions have received tremendous attention. Recent studies identified a biochemical link between the brain and the bacteria in the gut, known as the “gut microbiota-brain axis,” the mutualistic and bidirectional communication between the digestive tract and the central nervous system (CNS). The connection between gut and brain works by “bottom-up signaling” (from the digestive system to the brain, also known as the “gut-brain axis”) or “top-down signaling” (from the brain to the gut, also referred to as the “brain-gut axis”). Increasing evidence in the recent few years has highlighted the “bottom-up” and the “ top-down” interplay between the gut microbiota and the brain in many neurological and neurodegenerative diseases (Wang and Wang, 2016).
The gut microbiota has both positive and negative effects on the brain. Under normal body conditions, gut microbiota plays a critical role in many brains’ biological and physiological processes, such as myelination, glial cell activation, and neurogenesis. Additionally, the microbiota can effectively modulate behavior and influence psychological processes such as mood and cognition (Hsiao et al., 2013). The current scientific literature vehemently suggests that the gut microbiome can influence the brain aging process and the initiation and progression of neurodegenerative disorders, making the gut-brain crosstalk a promising and exciting research area in the neuroscience field. Studies have demonstrated that alterations in the gut microbiota composition, also known as dysbiosis, can negatively influence brain functions by initiating neuroinflammation (Fransen et al., 2017). Aging is a complicated process usually accompanied by chronic low-grade neuroinflammation known as inflammaging (Franceschi et al., 2000) along with a decline in cognition and memory (Peters, 2006). Dysbiosis is a disturbance of the commensal homeostasis between the host and the gut microbiota, and the outcome is chronic inflammation and a decreased production of some essential microbiota metabolites such as short-chain fatty acids (SCFAs).
Many studies have underscored the importance of gut microbiota alteration being the root cause of many diseases resulting in severe health implications and co-morbid conditions, such as inflammatory bowel disease, obesity, diabetes, cancer, cardiovascular disease, anxiety, and autism (Blumberg and Powrie, 2012; Cenit et al., 2017). Furthermore, intestinal dysbiosis plays a fundamental role in aging and many age-related neurodegenerative disorders, including Parkinson’s disease (PD) (Gorecki et al., 2019) and Alzheimer’s disease (AD) (Winek et al., 2016). The crucial information related to microbiota and its role in brain aging has emerged only in recent years. However, the evidence on the mechanisms is still unclear, and more studies are needed to determine its role in brain aging and age-related neurodegenerative disorders. Therefore, this review will summarize recent advancements in gut-brain axis dysregulation and related-neurodegenerative diseases. Future therapeutic implications for gut microbiota modulation are also summarized.
The search strategy and selection criteria were limited to articles published in peer-reviewed journals. A literature review of the past two decades (from 2000 to 2021) was conducted by searching the PubMed and Web of Science databases to identify articles reporting the relation between gut microbiota alteration, healthy brain aging, and age-related neuroinflammation and cognitive decline.
Gut Microbiota and Homeostasis
An enormous number of different microbes colonize in the mammalian intestine, and the concentration of these microbiomes increases gradually from the ileum to the colon (Sender et al., 2016). The gut microbiota has both positive and negative effects on the body and the brain, in particular. The host gut microbiota contributes to several biological and physiological processes, including regulating gut motility, preventing foreign pathogen colonization in the gut, releasing neurotransmitters, and producing various antioxidants and metabolites (Bercik et al., 2012; Haseeb Anwar et al., 2019). In normal physiological conditions, the density of the human microbiome is highest in the colon. Four phyla that constitute the bulk of the gut microbiota, namely Firmicutes, Bacteroidetes, Proteobacteria, and Actinobacteria, are the most abundant organisms, approximately 64%, 23%, 8%, and 3% of the population, respectively (Barengolts, 2016). The Firmicutes: Bacteroidetes ratio, the prevalent phyla of the bacterium in the intestine, is an indicator of the standard of metabolic health, and alteration in this ratio represents an index of abnormal conditions (Mariat et al., 2009). Notably, the gut microbiota changes at different developmental stages and aging (Mariat et al., 2009). For example, Proteobacteria predominates in neonate’s guts and considerably reduces from 16% in childhood to 4.6% during adulthood (Shin et al., 2015).
Microbiota populations can be influenced by various stress factors, such as diet, infections, drugs, illness, and aging (O’Mahony et al., 2009; Claesson et al., 2012; Davey et al., 2012). In these abnormal conditions, fecal microbiome biodiversity is decreased and represents underlying features of dysbiosis (Claesson et al., 2011). Gut microbiota dysbiosis occurs when the composition of resident commensal communities changes compared to the normal condition (Petersen and Round, 2014). Studies have shown that gut dysbiosis plays a role in developing and progressing diseases, including; infectious diseases, GI diseases, respiratory diseases, psychological diseases, and autoimmune diseases (Shreiner et al., 2015). Microbiota dysbiosis is also observed in subjects with metabolic and neurological disorders (Blumberg and Powrie, 2012). In addition, current evidence indicates that the impairment in the quantity and composition of gut microbiota affects cognitive functions and synaptic plasticity (Salami, 2021).
Healthy gut microbiota keeps the brain healthy and protects against different types of neurological disorders. Many recent studies investigated the idea that the host microbiota and their microbiome are essential in a healthy brain’s fundamental biological processes. For example, intestinal bacteria maintain microglial cells in a healthy mature steady-state condition during development, which is necessary to prevent neuroinflammatory and neurodegenerative disorders at the later phase of life (Rothhammer et al., 2016; Erny et al., 2017). Microglial cells, resident immune cells in CNS, play an essential role in modulating neurogenesis, maintaining homeostasis and cognition in healthy brains (Graeber and Streit 2010; Williamson et al., 2011). However, microglial overactivation augments neuronal damage, neuroinflammation, and cognitive dysfunction in aging and neurodegenerative diseases. Furthermore, the absence of host-microbiota led to defects in microglia maturation, differentiation, morphology, function, and gene expression profiles in mice harboring no intestinal microbiota and immune activity called germ-free (GF) mice compared with normal mice (Erny et al., 2015). These findings suggest that maternal microbiota contributes to microglia’s early development and has important brain development implications.
Gut Microbiota-Brain Axis
A plethora of evidence has provided new insights into the possible mechanisms and pathways that connect the intestinal microbiota and its metabolites with the brain and their involvement in regulating brain development and function (Hsiao et al., 2013; Erny et al., 2015). Based on the available literature, there are five major possible communication pathways of gut microbiota to influence the brain. These include neural networks, neuroendocrine system, gut microbiota metabolic system, gut immune system, and barriers system pathways (Wang and Wang, 2016), as shown in Figure 1.
Figure 1.
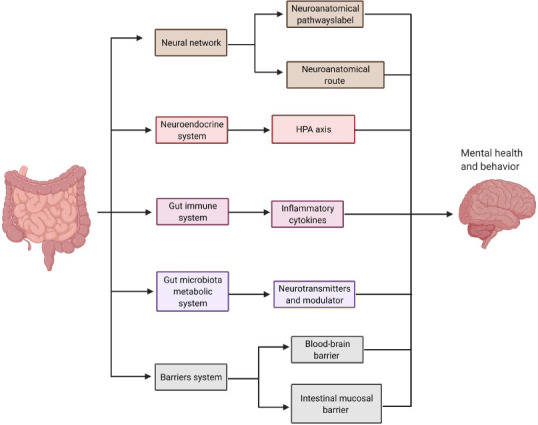
Gut-brain axis.
Five major possible communication pathways of gut microbiota and the brain: the neural network, neuroendocrine system, gut immune system, gut microbiota metabolic system, and barriers system. Created with BioRender.com. HPA: Hypothalamic-pituitary-adrenal.
The gut bacteria can interact with the brain through neuronal networks via two neuroanatomical pathways. The first neuroanatomical pathway directly connects the gut and brain by the autonomic nervous system and vagus nerve in the spinal cord (Bonaz et al., 2017). The second neuroanatomical route is a bidirectional communication through the enteric nervous system in the gut and autonomic nervous system and vagus nerve within the spinal cord (Furness, 2012). Another route of communication between gut bacteria and the brain is through the neuroendocrine system by the hypothalamic-pituitary-adrenal (HPA) axis, which regulates the body’s response to stress (Umesaki et al., 1999; Sudo et al., 2004). Gut microbiota can alter hypothalamic functions by secreting pro-inflammatory mediators that stimulate the release of corticotrophin-releasing hormone, a primary polypeptide hormone regulating the HPA axis. Studies have shown that the lack of gut microbiota or dysregulation of the HPA axis is characterized by increased plasma corticosterone levels which are linked to anxiety and depression-like behaviors (Holsboer 2000; Gosselin and Rivest, 2008). These studies suggest that the gut microbiota maintains CNS homeostasis by modulating the activity of the HPA axis.
Gut bacteria also synthesize some neurotransmitters and neural regulator substances that directly or indirectly influence brain functions (Brenchley and Douek, 2012), such as gamma-aminobutyric acid, 5-hydroxytryptamine, and dopamine. Gut microbiota also releases metabolites such as SCFA and tryptophan, which control brain inflammation. A recent study highlighted the microbial catabolites of dietary tryptophan produced by gut microbiota and released under a specific tryptophan diet-controlled microglia and astrocyte activation by acting through the microglial aryl hydrocarbon receptor (Rothhammer et al., 2018). Tryptophan is one of the essential amino acids supplied by dietary protein and can be catabolized by gut microbiota, producing various indole-derivatives (Roager and Licht, 2018). It has been shown that these tryptophan catabolites have a positive or protective impact on neurological and neuroinflammatory diseases (Roager and Licht 2018; Rothhammer et al., 2018). Another study reported that treatment with SCFAs as a dietary supplement rescues microglial function otherwise impaired in GF animals and can reverse microglia’s immature phenotype (Erny et al., 2015). SCFAs are metabolites produced by the microbiome and derived mainly from the fermentation of non-digestible carbohydrates and the digestion of proteins and peptides (Rios-Covian et al., 2016). It is considered one of the most studied microbiota metabolic products such as acetic, butyric, propionic, valeric, isovaleric, and isobutyric acids (Kotani et al., 2009). Under physiological conditions, SCFAs perform a wide range of physiologic functions, ranging from regulating and activating the immune system to microglial cell homeostasis (Canfora et al., 2015; Levy et al., 2017). These findings further expand our understanding of how gut microbiota impacts microglia in the brain. The gut immune system is another system that influences the brain through innate immune receptors [toll-like receptors (TLRs)] in the digestive system and activates adaptive immune cells (McKernan et al., 2011; Furusawa et al., 2013). TLRs are the first step in inducing cytokines against immune response and can trigger several critical reactions for maintaining gut microbiota homeostasis (McKernan et al., 2011; Hooper et al., 2012). In addition, the microbiota has a direct impact on lymphoid structure development, T cell subsets balance, and epithelial cell function (Hooper et al., 2012). In response to dysbiosis, the gut microbiota triggers the immune system in both the intestine and CNS. Subsequently, the body produces cytokines and other inflammatory mediators that promote systemic inflammation and modulate glial cell activation and function (Cryan et al., 2019).
The barrier system permeability is a unique communication pathway by the intestinal mucosal barrier and blood-brain barrier (BBB). BBB integrity and its development are also dependent on gut microbiota composition, such as Clostridium tyrobutyricum and Bacteroides thetaiotaomicron (Braniste et al., 2014). However, the molecular mechanisms by which gut microbiota regulate the BBB integrity and permeability are still unclear. Maintaining BBB integrity is critical for preserving a homeostatic environment for the normal functioning of the brain. In a recent study, GF mice monocolonized with a beneficial bacterium, Clostridium tyrobutyricum, and Bacteroides thetaiotaomicron, mainly produce a different type of SCFAs a decrease in BBB permeability and up-regulation in the tight junction proteins expression (Braniste et al., 2014). Other studies have shown that environmental factors such as aging, stress, diet modification, and disease could induce intestinal mucosal barrier dysfunction. The mucosal barrier dysfunction facilitates the entry of metabolites, byproducts, and cytokines into the blood circulation and stimulation of TLRs (Brenchley and Douek 2012; Konig et al., 2016), and the outcome is a profound increase in the permeability of BBB for unwanted substances (Rezai-Zadeh et al., 2009).
Animal Models of Aging-Induced Gut Microbiota Modulation
Using rodents as a model system in microbiota-related research is considered a powerful tool that can be utilized under controlled conditions to further understand the gut microbiota brain axis. There are two main animal models that have been used to study the microbiota in normal and pathogenic conditions with aging. In one model, the rodents are treated with antibiotics for several days to induce severe gut microbiota dysbiosis (Frohlich et al., 2016). Antibiotic administration substantially reduces bacterial bioavailability and decreases microbiota diversity (Dudek-Wicher et al., 2018). In addition, antibiotic administration negatively impacts cognition by reducing the expression of cognition-related proteins (brain-derived neurotrophic factor (BDNF) and synaptophysin) in different brain regions (Frohlich et al., 2016). The second model includes raising the rodents in a germ-free environment, called GF rodents (Al-Asmakh and Zadjali 2015; Fransen et al., 2017). The germ-free animal model is an excellent model for studying abnormal brain structure, development, and behavioral dysfunctions (Salami, 2021). Furthermore, other methods have been used to produce a positive or negative intervention in aging research, such as dietary manipulations, treating with probiotics/prebiotics, and fecal microbiota transplantation (FMT) (Drastich et al., 2018). FMT technique is used by transplanting the gut microbiota from aged to young subjects to study whether the shift of gut microbiota in aged subjects can affect cognitive behavior or change the brain structure and functions in the young subjects (Borody et al., 2013).
Initial animal and human studies used fecal analysis to assess the gut microbiota composition and its metabolic products. Numerous advanced techniques, such as meta-omics analysis and next-generation high-throughput sequencings (16S ribosomal RNA (rRNA) microbial profiling), DNA microarrays, metatranscriptomics, metabolomics, and shotgun metagenomics, were utilized to understand the gut microbiota-brain axis and identify microbial communities, composition, and functions (Mariat et al., 2009; Zwielehner et al., 2009; Fraher et al., 2012; Ticinesi et al., 2018; Li et al., 2020). These techniques can also determine the microbial diversity, qualitative and quantitative information of specific bacterial types, and gut microbiota changes during the initiation and progression of the disease states (Fraher et al., 2012).
Gut Microbiota Alterations and the Normal Aging Process
An imbalance of the intestinal homeostasis is linked with the normal aging process and many infectious and neurodegenerative diseases in the elderly (Westfall et al., 2017; Xu et al., 2019). Healthy aging humans tend to have a different composition, stability, and diversity of gut microbiota than young adults (Woodmansey et al., 2004; Mueller et al., 2006; Jeffery et al., 2016). In one clinical study, the microbiota dysbiosis change between the aged (65 years and older) and young (28–46 years old) was analyzed using fecal sampling (Claesson et al., 2011). The study observed that in the elderly, the phylum Bacteroidetes is dominant at 57%, and phylum Firmicutes is dominant at 40%. However, in young subjects, the phylum Bacteroidetes dominated by 41%, and the phylum Firmicutes dominated by 51%. A similar result was observed in other studies showing a decrease in Firmicutes/Bacteroidetes ratio (from 10.9 to 0.6) of the human microbiota between adults (25 to 45 years old) and the elderly (70–90 years old) (Mariat et al., 2009). Another clinical study that recruited 371 people from newborn to centenarians reported a significant difference in 35 genera in aged human gut microbiota by performing 16S RNA sequencing (Xu et al., 2019). Interestingly, beneficial genera (Oxalobacter, Butyrivibrio, and Lactobacillus) were lost, whereas genera (Parvimonas, Butyricimonas, and Anaerotruncus) related to inflammation and disease increased.
Human and rodent gut microbiota share Firmicutes and Bacteroidetes, major microbial phyla (Nguyen et al., 2015; Gopalakrishnan et al., 2021). There were significant differences observed in these phyla ratio levels, communities, and composition. In contrast to humans, the ratio of two main phyla, Firmicutes and Bacteroidetes, was increased in mice and rats with advanced age (18–20 months) to almost nine-fold compared to young (2–6 months) and was accompanied by cognitive impairments and increased anxiety behaviors (Hoffman et al., 2017; Spychala et al., 2018; Li et al., 2020). Similarly, microbiota changes were also investigated by decreasing the bacteroidetes and Tenericutes levels and increasing Firmicutes and Actinobacteria levels, resulting in an increase in the Firmicutes to Bacteroidetes ratio (Kim et al., 2016, 2017).
During aging, microbiota alteration occurs mainly by decreasing microbiota diversity and increasing in pathological bacteria, Proteobacterium, and relatively low proportions of beneficial bacteria, Bifidobacterium. Besides alterations in microbial biodiversity, the production of beneficial microbiota metabolites such as SCFAs is also drastically reduced, which may be linked to the aging-related process (Erny et al., 2015; Chen et al., 2019). The administration of antibiotics in adult mice resulted in decreased SCFAs concentrations and cognitive impairments compared to the control group (Frohlich et al., 2016). These findings indicate that SCFAs change with aging and could play an essential role in the pathophysiology of disease and cognitive decline. Compared to young, aged mice showed a reduction of SCFAs metabolites by 68% acetate and propionate and 80% of the butyrate levels. These reduced levels were in parallel with the cognitive impairments (Spychala et al., 2018). In all these studies, significant changes were observed between the intestine flora and host with aging in both humans and animals, and these changes gradually reach a stage of microbiota dysbiosis. Therefore, understanding the changes in the microbiome with aging may provide potential biomarkers of the initiation of the aging process.
Gut Microbiota Alteration and Age-Related Neuroinflammation
Since chronic low-grade systemic inflammation contributes to age-related diseases, many studies have shown that gut microbiota modulates the inflammaging and could be a key determinant for age-related neuroinflammation and neurodegeneration. The fecal microbiota transplantation was also used to examine the effects of microbiota dysbiosis on the chronic and inflamed immune system with aging (Fransen et al., 2017; Li et al., 2020). It was found that the microbiota triggers innate immunity and produces inflammatory responses mimicking inflammaging with heightened intestinal inflammation and increased circulation of inflammatory cytokines in both serum and brains. Bacteria populating the gut can secrete large amounts of metabolites and byproducts such as double-stranded RNA, lipoproteins, and lipopolysaccharides (LPS). These bacterial components contribute to the signaling pathways involved in the excessive production of pro-inflammatory cytokines such as interleukin (IL)-8 and IL-6 (Biagi et al., 2010). As per published studies, it was observed that aging corresponds to an increase in LPS production level in plasma and brains, a cell membrane byproduct of gram-negative bacteria. At the molecular level, the increase in LPS level is correlated with the increased expression of TLR4, myeloid differential protein-88, and the nuclear translocation of nuclear factor κB in both intestinal and brain tissues (Wu et al., 2021). Another study also showed that bowel inflammation triggers neuroinflammation in rats (Villaran et al., 2010). It was observed that LPS induced ulcerative colitis mediates systemic inflammation leading to exacerbated BBB permeability, induced inflammation in the substantia nigra region of the brain, and dopaminergic loss. They also investigated that non-selective macrophage depletion by an intravenous administration of liposome-encapsulated clodronate reduced the peripheral and neuroinflammatory response. These studies explain that the mucosal translocation of bacterial LPS and LPS-binding proteins into the systemic circulation could also promote the chronic low-grade systemic inflammation mainly found in the older subjects and can initiate CNS inflammation by activation LPS/TLR4 pathway in brain glial cells (Stehle et al., 2012; Ghosh et al., 2015; Kim et al., 2016). In addition, LPS increases the BBB permeability by activating TLR4/IRF-3 signaling pathway in the endothelial cell in the blood vessels and disrupting the intestinal epithelium barrier (Choi et al., 2012). Higher levels of LPS were also found in the hippocampal and superior temporal lobe of the postmortem brain lysates of AD patients (Zhao et al., 2017). In another study, increased bacterial populations in AD patients’ brain tissue were investigated using 16S rRNA sequencing (Emery et al., 2017). Related to these studies, LPS challenged GF mice showed microglial morphological and functional dysregulation and impairments in the release of the pro-inflammatory cytokines and interleukins such as IL-1β, IL-6, and tumor necrosis factor-α (Erny et al., 2015; Kim et al., 2016; Matcovitch-Natan et al., 2016). Accordingly, a significant increase in the pro-inflammatory cytokines and impairments in oxidative stress protein expression was also investigated in young rat serum and brains after transplanting with aged rat microbiota (Li et al., 2020). Based on all these studies, the gut microbiota represented a pro-inflammatory phenotype in aged subjects promoting the idea that age-related neuroinflammation are modulated by chronic peripheral inflammation
Moreover, the substances synthesized by gut microbiota, either as metabolic byproducts or constituents of bacterial structures, can promote protein aggregation in brain tissues in some neurodegenerative disorders. For example, many gut microbiota species are associated with the production of amyloid fibers, such as Escherichia coli (E.coli) and Bacillus subtilis. These amyloid fibers can cross the intestinal and BBB layer, promote amyloid-β protein formation and accumulation in the brain, and enhance AD pathogenesis in the elderly (Kowalski and Mulak, 2019; Li et al., 2019). The findings are supported by studies reporting a reduction in amyloid protein concentration in the serum and brain in the AD transgenic mouse model treated with antibiotics (Minter et al., 2016; Harach et al., 2017). It was hypothesized that the increased levels of stress-related proteins in aging brains could be the possible reason for neuroinflammation and subsequent neurodegenerative diseases (Alsegiani and Shah, 2020). The byproducts of gut microbiota alteration and dysbiosis could alter the expression of various genes and associated synaptic proteins and promote the accumulation of inflammatory proteins in the brain, inducing neuroinflammation, astrocyte activation, neuronal apoptosis, and cognitive decline. These events appear several years before the onset of age-related neurodegeneration disorders, as shown in Figure 2. It is proved that microbiota alteration activity gradually increases with aging, and this imbalance causes glial cells to turn into a reactive state and subsequently induce inflammation in the brain.
Figure 2.
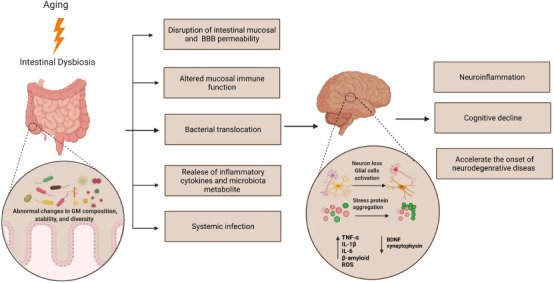
The aging brain starts from the digestive system and subsequently develops into neurodegenerative diseases.
Gut microbiota dysbiosis induces stress protein aggregation in the healthy brain leading to neuroinflammation and brain aging, consequently increasing the risk of developing neurodegenerative disorders. Created with BioRender.com. BDNF: Brain-derived neurotrophic factor; IL: interleukin; ROS: reactive oxygen spcies; TNF-α: tumor necrosis factor alpha.
Gut Microbiota Alteration and Age-Related Cognitive Decline
Many recent studies found that age-related neuroinflammation and cognitive decline are associated with the integrity of microbiota-gut-brain axis function. The gut microbiota alteration with aging amounts to leakiness of the gut epithelial barrier and BBB, loss of the enteric nervous system, altered neurotransmitters, metabolites, and mucosal immune function resulting in excessive pro-inflammatory cytokine release (Biagi et al., 2010; Fransen et al., 2017). So far, there are limited studies available underlying the mechanisms associating intestinal microbiota alterations or dysbiosis with age-related neuroinflammation and cognitive decline in humans. However, few studies reported a low microbiota diversity between healthy elder adults and those with cognitive impairments (Claesson et al., 2011, 2012). A recent preclinical study reported a significant decrease in the anti-inflammatory genus Faecalibacterium in people with subjective cognitive decline compared to the normal healthy group (Sheng et al., 2021). Subjective cognitive decline is the first symptomatic manifestation of preclinical AD (Rabin et al., 2017). It was suggested that the gut microbiota has emerged as an essential player in the development of vascular cognitive impairment in elderly patients with cerebral vascular disease (Li et al., 2018). Vascular cognitive impairment is a cognitive impairment that is attributed to cerebrovascular disease in the elderly such as atherosclerosis, ischemic and hemorrhagic stroke (Li et al., 2018; Iadecola et al., 2019). A summary of human studies comparing the changes in the human gut microbiome composition between living subjects suffering from cognitive symptoms and healthy older subjects is presented in Table 1.
Table 1.
Overview of the main human studies comparing the fecal gut microbiota composition between subjects suffering from cognitive symptoms and healthy older subjects
| Reference | Number and characteristics of subjects | Mean age (yr) | Gut microbiota alteration in cognitive decline patients compared to control healthy older adults |
|---|---|---|---|
| Cattaneo et al., 2017 | 73 patients with dementia | 71 ± 7 (cases) | The microbiota of patients with dementia had higher pro-inflammatory gut microbiota taxon (Escherichia/Shigella) and reduced anti-inflammatory taxon (Eubacterium rectale). |
| 10 controls | 68 ± 8 (controls) | The decrease in the E. rectale population is associated with peripheral systemic inflammation. | |
| Vogt et al., 2017 | 25 patients with dementia | 71 ± 7 (cases) | Subjects with cognitive impairments had decreased Firmicutes and increased Bacteroidetes |
| 25 controls | 69 ± 8 (controls) | Reduced the abundance of 13 anti-inflammatory taxa, including Bifidobacterium, Alistipes, Bilophila, and Clostridium. | |
| Manderino et al., 2017 | 25 with cognitive decline | 68 ± 7 (cases and controls) | Subjects with cognitive impairments had an abundance of 4 bacterial phyla, Bacteroidetes, Firmicutes, Proteobacteria, and Verrucomicrobia. |
| 18 with mild cognitive decline | A strong correlation with cognitive decline was observed in Verrucomicrobia abundance | ||
| Zhuang et al., 2018 | 43 patients with dementia 43 controls | 70 ± 9 (cases and controls) | Subjects with dementia had fecal microbiota composition alteration characterized by an abundance of several species including Bacteroides, Actinobacteria, Ruminococcus, Lachnospiraceae, and Selenomonadalestable |
| Saji et al., 2019 | 34 patients with dementia 94 controls | 75 ± 9 (cases and controls) | Subjects with dementia had a low abundance of Bacteroides, Bifidobacterium and Lactobacillales and high Firmicutes/Bacteroidetes ratio and other bacteria. |
| Sheng et al., 2021 | 67 patients with cognitive decline | 67 ± 6 (cases) | Subjects with dementia had a low abundance of Firmicutes, class Clostridia, Clostridiales, Ruminococcaceae, and Faecalibacterium. |
| 38 controls | 73 ± 8 (controls) | A significant decrease in the abundance of the anti-inflammatory genus Faecalibacterium |
Evidence from animal studies suggests that the alterations in gut microbiota may drive cognitive impairments and behavioral dysregulation in aged mice due to the prolonged exposure of the brain to microbiota metabolites and pro-inflammatory cytokine (Caracciolo et al., 2014; Erny et al., 2015; Leung and Thuret 2015; Manderino et al., 2017). Several recent studies used the FMT technique to investigate the effects of transferring gut microbiota from old to young rodents, focusing on age-related signs of cognitive decline and neuroinflammation. The results suggest that mice receiving an aged microbiota began to behave like older mice by exhibiting impaired motor strength and cognitive behaviors (D’Amato et al., 2020; Li et al., 2020; Wu et al., 2021). Besides cognition impairments, another study observed alteration of one hundred forty proteins involved in synaptic plasticity and neurotransmission in the hippocampus, in addition to downregulation of proteins involved in glucose transport at the BBB by using Label-free quantitative proteomics (D’Amato et al., 2020). All these detected alterations in protein expression in the hippocampus region in young recipient mice contributed to the dysfunction of the normal aging brain process. In addition, a substantial reduction of SCFA-producing bacteria was also reported in young mice receiving fecal transplantation from aged mice.
Moreover, fecal microbiota transplants from aged donor rats significantly decreased the regional homogeneity and the density of dendritic spines. Furthermore, it changed the synaptic structures by reducing the expression of cognition proteins such as BDNF, N-methyl-D-aspartate receptor NR1 subunit, and synaptophysin in the hippocampus and prefrontal cortex regions in the young fecal recipient rats (Li et al., 2020). These studies provide a shred of direct evidence for the contribution of gut microbiota to age-related cognitive decline and in the brain’s physiological function and structure. In contrast to these studies, another study confirmed the increased hippocampal neurogenesis, intestinal growth, and pro-longevity signaling in young GF mice after receiving fecal microbiota from aged mice (Kundu et al., 2019). These findings suggest that gut microbiota transplants from aged subjects could have beneficial protection from neurological and neurodegenerative disorders in young subjects.
Potential Therapeutic Possibilities Targeting Gut Microbiota Dysbiosis to Reduce Age-Related Disorders
As life expectancy began increasing with the turn of this century, the risk of having a neurogenerative disease increased with aging. Having a healthy aging brain and delaying the onset of neurodegenerative disease is a big challenge. So far, the effective pharmacological therapies developed for neurological conditions mainly were based on targeting proteins or genes within the brain. The intestinal microbiome emerged as a promising target for anti-aging interventions based on the emerging appreciation of a gut-microbiota brain axis. Therefore, microbiota alteration could become a promising therapeutic target for neuroinflammation and cognitive decline related to aging and other neurodegenerative diseases.
Fecal microbiota transplantation
Research on the gut-brain axis has identified several promising targets for the efficacy of pharmacological interventions. One possible treatment method involves restoring gut microbiota’s composition and function by introducing fecal contents from healthy young subjects into the aged subject’s gastrointestinal (GI) tract, known as fecal microbiota transplantation (FMT). The FMT method has been used in more than 380 clinical trials to treat many GI target diseases, including constipation, irritable bowel syndrome, and autoimmune diseases (Choi and Cho 2016; Hollingsworth et al., 2021). A recent study reported that the beneficial effects of FMT from 3-months aged mice to 24-month-old mice improve the immune system homeostasis and cognitive functioning, in particular spatial learning and memory (D’Amato et al., 2020). However, the use of FMT is limited due to the unknown long-term efficacy of this method and the reported unexpected side effects. For example, a recently published case study identified a 56-year-old female patient with chronic radiation colitis who developed adhesion ileus after 2 days of the FMT (Harsch and Konturek, 2019). Researchers speculate the adhesive side effect because the transplantation of new bacterial species into the colon could have led to the trapping of a gut segment in a preexisting adhesion. In addition, the possibility of transferring the endotoxins or infectious agents to the recipient subjects could worsen GI complications (De Leon et al., 2013; Schwartz et al., 2013). Besides these factors, sex differences might also play an essential factor since many studies showed that the gut microbiome varies between males and females in human and rodent studies (Dominianni et al., 2015; Borgo et al., 2018). Therefore, further investigation is needed to prove the use of the FMT technique to improve the cognition decline in aged humans. However, isolation of a defined group of bacteria from the fecal sample then transported into the recipient’s intestine would be a safer alternative to the FMT method (Buffie et al., 2015; Wymore Brand et al., 2015).
Probiotics and prebiotics
Supplementation with probiotics and prebiotics is a promising alternative therapeutic option for treating gut alteration in aging. Prebiotics are non-digestible and fermented food components that selectively promote beneficial commensal microbiota growth and activity (Franco-Robles and Lopez, 2015), such as a fiber-rich diet like yogurt, fruits, vegetables, and whole grains. The significant degradation and fermentation products of prebiotic metabolism in the intestine are SCFAs. In contrast, probiotics are supplements that supply beneficial bacteria to GI to modulate gut microbiota composition and function, such as Bifidobacterium lactis (Franco-Robles and Lopez, 2015). In addition, the positive effect of probiotics in healthy cognitive function has been considered recently in many human and animal studies.
Studies have shown that the utilization of prebiotics and probiotics increases the production of SCFA, improves age-related memory alteration, stimulates immune responses, and decreases inflammation and oxidative stress in the brain (Brussow, 2013). Another study suggested that prebiotics and probiotics improve learning and memory in rats by increasing butyrate production in the intestine and simultaneously increasing the BDNF expression and decreasing the pro-inflammatory cytokine concentrations in the hippocampus (Romo-Araiza et al., 2018). Furthermore, it has been found that probiotics can improve CNS dysfunction by increasing both the diversity and intestinal bacteria population (Kwok et al., 2014). One clinical study found that healthy elderly (≥ 65 years) administering a multi-strain probiotic for 12 weeks showed significant improvement in brain function, mental flexibility, alleviation stress, and increased serum BDNF levels (Kim et al., 2021). In addition, a probiotic mixture has also shown an improvement in aged mice memory and reduction in glial activation, and modified microbiota composition in the feces and the brain (Ho et al., 2019; Yang et al., 2020). Also, the single mono-strain probiotic administration improved memory function in young and older adults suffering from mild cognitive impairments (Sanborn et al., 2020; Xiao et al., 2020) and aged mice (Wang et al., 2020). Besides the beneficial effects of probiotic supplementations in cognitive improvement, several issues are necessary to be addressed. The pharmacological effects of the probiotic are strain-specific, which means each probiotic bacterial strain is specific for a unique host type (Musa et al., 2009; Stenman et al., 2020). Therefore, understanding their mode of action in aging research is needed to elucidate the more efficient probiotic strains that can offer health benefits in different individuals. Defining the dose of the probiotics is also needed to achieve the positive effects in preventing diseases and modulating the immune system. However, using probiotics in healthy individuals to modulate the gut microbiota remains highly debated and controversial. In addition, the long-term efficacy of probiotics in the intestine and fecal microbiota is still unknown.
Conclusion and Future Perspective
Even though gut microbiota dysbiosis has been considerably studied in recent years, the underlying causes remain unclear. This review discussed and focused on gut microbial in brain aging neuroinflammation, cognitive decline, and the suggested therapeutic options to reduce dysbiosis and cognitive decline in aging. The discussed studies could pave the way for reinforcing microbiota dysbiosis as a potential biomarker in aging and provide strong evidence for manipulating gut microbiota with probiotic and prebiotic therapy as an essential option to prevent and treat different neurodegenerative diseases. The most critical unmet need in the gut-brain axis is to find the relationship between specific pathological biomarkers and microbiota changes. This has sprouted a plethora of studies aimed at modifying the gut microbiota composition in the elderly that potentially could reduce inflammaging and improve cognitive decline. However, the critical questions to be answered are: does brain aging start from the brain or digestive system? Is microbiota responsible for the incidence of brain aging and pathogenesis? Does peripheral systemic inflammation emerge from gut bacteria? Understanding the role of dysbiosis in aging and altering the gut microbiota may prove helpful in attenuating inflammaging and the associated diseases. Furthermore, identifying the biochemical and mechanical signaling that mediates dysbiosis could help early detection of neuroinflammation and neurodegenerative diseases.
Footnotes
Conflicts of interest: The authors declare no conflicts of interest.
Editor note: ZAS is an Editorial Board member of Neural Regeneration Research. He was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal’s standard procedures, with peer review handled independently of this Editorial Board member and their research groups.
Open peer reviewer: Chuanzhou Li, School of Basic Medical Sciences, Huazhong University of Science and Technology, Hubei Province, China.
Funding: ASA was supported by the College of Pharmacy, Department of Pharmaceutical Chemistry, King Saud University, Riyadh, Saudi Arabia. The study was partly supported by the grants from The American Heart Association, No. 17AIREA33700076/ZAS/2017 and the National Institute of Neurological Disorders and Stroke of the National Institutes of Health, No. R01NS112642 (to ZAS).
P-Reviewers: Li C; C-Editors: Zhao M, Zhao LJ, Li JY; T-Editor: Jia Y.
References
- 1.Al-Asmakh M, Zadjali F. Use of germ-free animal models in microbiota-related research. J Microbiol Biotechnol. 2015;25:1583–1588. doi: 10.4014/jmb.1501.01039. [DOI] [PubMed] [Google Scholar]
- 2.Alsegiani AS, Shah ZA. The role of cofilin in age-related neuroinflammation. Neural Regen Res. 2020;15:1451–159. doi: 10.4103/1673-5374.274330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Barengolts E. Gut microbiota, prebiotics, probiotics, and synbiotics in management of obesity and prediabetes: review of randomized controlled trials. Endocr Pract. 2016;22:1224–1234. doi: 10.4158/EP151157.RA. [DOI] [PubMed] [Google Scholar]
- 4.Bercik P, Collins SM, Verdu EF. Microbes and the gut-brain axis. Neurogastroenterol Motil. 2012;24:405–413. doi: 10.1111/j.1365-2982.2012.01906.x. [DOI] [PubMed] [Google Scholar]
- 5.Biagi E, Nylund L, Candela M, Ostan R, Bucci L, Pini E, Nikkïla J, Monti D, Satokari R, Franceschi C, Brigidi P, De Vos W. Through ageing, and beyond: gut microbiota and inflammatory status in seniors and centenarians. PLoS One. 2010;5:e10667. doi: 10.1371/journal.pone.0010667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Blumberg R, Powrie F. Microbiota, disease, and back to health: a metastable journey. Sci Transl Med. 2012;4:137rv7. doi: 10.1126/scitranslmed.3004184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bonaz B, Sinniger V, Pellissier S. The vagus nerve in the neuro-immune axis: implications in the pathology of the gastrointestinal tract. Front Immunol. 2017;8:1452. doi: 10.3389/fimmu.2017.01452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Borgo F, Garbossa S, Riva A, Severgnini M, Luigiano C, Benetti A, Pontiroli AE, Morace G, Borghi E. Body mass index and sex affect diverse microbial niches within the gut. Front Microbiol. 2018;9:213. doi: 10.3389/fmicb.2018.00213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Borody TJ, Paramsothy S, Agrawal G. Fecal microbiota transplantation: indications, methods, evidence and future directions. Curr Gastroenterol Rep. 2013;15:337. doi: 10.1007/s11894-013-0337-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Braniste V, Al-Asmakh M, Kowal C, Anuar F, Abbaspour A, Tóth M, Korecka A, Bakocevic N, Ng LG, Kundu P, Gulyás B, Halldin C, Hultenby K, Nilsson H, Hebert H, Volpe BT, Diamond B, Pettersson S. The gut microbiota influences blood-brain barrier permeability in mice. Sci Transl Med. 2014;6:263ra158. doi: 10.1126/scitranslmed.3009759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Brenchley JM, Douek DC. Microbial translocation across the GI tract. Annu Rev Immunol. 2012;30:149–173. doi: 10.1146/annurev-immunol-020711-075001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Buffie CG, Bucci V, Stein RR, McKenney PT, Ling L, Gobourne A, No D, Liu H, Kinnebrew M, Viale A, Littmann E, van den Brink MR, Jenq RR, Taur Y, Sander C, Cross JR, Toussaint NC, Xavier JB, Pamer EG. Precision microbiome reconstitution restores bile acid mediated resistance to Clostridium difficile. Nature. 2015;517:205–208. doi: 10.1038/nature13828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Canfora EE, Jocken JW, Blaak EE. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat Rev Endocrinol. 2015;11:577–591. doi: 10.1038/nrendo.2015.128. [DOI] [PubMed] [Google Scholar]
- 14.Caracciolo B, Xu W, Collins S, Fratiglioni L. Cognitive decline, dietary factors and gut-brain interactions. Mech Ageing Dev. 2014;136-137:59–69. doi: 10.1016/j.mad.2013.11.011. [DOI] [PubMed] [Google Scholar]
- 15.Cattaneo A, Cattane N, Galluzzi S, Provasi S, Lopizzo N, Festari C, Ferrari C, Guerra UP, Paghera B, Muscio C, Bianchetti A, Volta GD, Turla M, Cotelli MS, Gennuso M, Prelle A, Zanetti O, Lussignoli G, Mirabile D, Bellandi D, et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol Aging. 2017;49:60–68. doi: 10.1016/j.neurobiolaging.2016.08.019. [DOI] [PubMed] [Google Scholar]
- 16.Cenit MC, Sanz Y, Codoner-Franch P. Influence of gut microbiota on neuropsychiatric disorders. World J Gastroenterol. 2017;23:5486–5498. doi: 10.3748/wjg.v23.i30.5486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Chen R, Xu Y, Wu P, Zhou H, Lasanajak Y, Fang Y, Tang L, Ye L, Li X, Cai Z, Zhao J. Transplantation of fecal microbiota rich in short chain fatty acids and butyric acid treat cerebral ischemic stroke by regulating gut microbiota. Pharmacol Res. 2019;148:104403. doi: 10.1016/j.phrs.2019.104403. [DOI] [PubMed] [Google Scholar]
- 18.Choi HH, Cho YS. Fecal microbiota transplantation: current applications effectiveness, and future perspectives. Clin Endosc. 2016;49:257–265. doi: 10.5946/ce.2015.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Choi JJ, Choi YJ, Chen L, Zhang B, Eum SY, Abreu MT, Toborek M. Lipopolysaccharide potentiates polychlorinated biphenyl-induced disruption of the blood-brain barrier via TLR4/IRF-3 signaling. Toxicology. 2012;302:212–220. doi: 10.1016/j.tox.2012.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Claesson MJ, Cusack S, O'Sullivan O, Greene-Diniz R, de Weerd H, Flannery E, Marchesi JR, Falush D, Dinan T, Fitzgerald G, Stanton C, van Sinderen D, O'Connor M, Harnedy N, O'Connor K, Henry C, O'Mahony D, Fitzgerald AP, Shanahan F, Twomey C, Hill C, Ross RP, O'Toole PW. Composition, variability, and temporal stability of the intestinal microbiota of the elderly. Proc Natl Acad Sci U S A. 2011;108:4586–4591. doi: 10.1073/pnas.1000097107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Claesson MJ, Jeffery IB, Conde S, Power SE, O'Connor EM, Cusack S, Harris HM, Coakley M, Lakshminarayanan B, O'Sullivan O, Fitzgerald GF, Deane J, O'Connor M, Harnedy N, O'Connor K, O'Mahony D, van Sinderen D, Wallace M, Brennan L, Stanton C, et al. Gut microbiota composition correlates with diet and health in the elderly. Nature. 2012;488:178–184. doi: 10.1038/nature11319. [DOI] [PubMed] [Google Scholar]
- 22.Cryan JF, O'Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, Boehme M, Codagnone MG, Cussotto S, Fulling C, Golubeva AV, Guzzetta KE, Jaggar M, Long-Smith CM, Lyte JM, Martin JA, Molinero-Perez A, Moloney G, Morelli E, Morillas E, O'Connor R, et al. The microbiota-gut-brain axis. Physiol Rev. 2019;99:1877–2013. doi: 10.1152/physrev.00018.2018. [DOI] [PubMed] [Google Scholar]
- 23.D'Amato A, Di Cesare Mannelli L, Lucarini E, Man AL, Le Gall G, Branca JJV, Ghelardini C, Amedei A, Bertelli E, Regoli M, Pacini A, Luciani G, Gallina P, Altera A, Narbad A, Gulisano M, Hoyles L, Vauzour D, Nicoletti C. Faecal microbiota transplant from aged donor mice affects spatial learning and memory via modulating hippocampal synaptic plasticity- and neurotransmission-related proteins in young recipients. Microbiome. 2020;8:140. doi: 10.1186/s40168-020-00914-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Davey KJ, O'Mahony SM, Schellekens H, O'Sullivan O, Bienenstock J, Cotter PD, Dinan TG, Cryan JF. Gender-dependent consequences of chronic olanzapine in the rat: effects on body weight, inflammatory, metabolic and microbiota parameters. Psychopharmacology (Berl) 2012;221:155–169. doi: 10.1007/s00213-011-2555-2. [DOI] [PubMed] [Google Scholar]
- 25.De Leon LM, Watson JB, Kelly CR. Transient flare of ulcerative colitis after fecal microbiota transplantation for recurrent Clostridium difficile infection. Clin Gastroenterol Hepatol. 2013;11:1036–1038. doi: 10.1016/j.cgh.2013.04.045. [DOI] [PubMed] [Google Scholar]
- 26.Dominianni C, Sinha R, Goedert JJ, Pei Z, Yang L, Hayes RB, Ahn J. Sex, body mass index, and dietary fiber intake influence the human gut microbiome. PLoS One. 2015;10:e0124599. doi: 10.1371/journal.pone.0124599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Drastich P, Bajer L, Kverka M. Possibilities of therapeutic manipulation of the gut microbiota. Vnitr Lek. 2018;64:665–671. [PubMed] [Google Scholar]
- 28.Dudek-Wicher RK, Junka A, Bartoszewicz M. The influence of antibiotics and dietary components on gut microbiota. Prz Gastroenterol. 2018;13:85–92. doi: 10.5114/pg.2018.76005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Emery DC, Shoemark DK, Batstone TE, Waterfall CM, Coghill JA, Cerajewska TL, Davies M, West NX, Allen SJ. 16S rRNA next generation sequencing analysis shows bacteria in Alzheimer's post-mortem brain. Front Aging Neurosci. 2017;9:195. doi: 10.3389/fnagi.2017.00195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Erny D, Hraběde Angelis AL, Jaitin D, Wieghofer P, Staszewski O, David E, Keren-Shaul H, Mahlakoiv T, Jakobshagen K, Buch T, Schwierzeck V, Utermöhlen O, Chun E, Garrett WS, McCoy KD, Diefenbach A, Staeheli P, Stecher B, Amit I, Prinz M. Host microbiota constantly control maturation and function of microglia in the CNS. Nat Neurosci. 2015;18:965–977. doi: 10.1038/nn.4030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Erny D, Hrabe de Angelis AL, Prinz M. Communicating systems in the body: how microbiota and microglia cooperate. Immunology. 2017;150:7–15. doi: 10.1111/imm.12645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fraher MH, O'Toole PW, Quigley EM. Techniques used to characterize the gut microbiota: a guide for the clinician. Nat Rev Gastroenterol Hepatol. 2012;9:312–322. doi: 10.1038/nrgastro.2012.44. [DOI] [PubMed] [Google Scholar]
- 33.Franceschi C, Bonafè M, Valensin S, Olivieri F, De Luca M, Ottaviani E, De Benedictis G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann N Y Acad Sci. 2000;908:244–254. doi: 10.1111/j.1749-6632.2000.tb06651.x. [DOI] [PubMed] [Google Scholar]
- 34.Franco-Robles E, Lopez MG. Implication of fructans in health: immunomodulatory and antioxidant mechanisms. ScientificWorldJournal. 2015;2015:289267. doi: 10.1155/2015/289267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Fransen F, van Beek AA, Borghuis T, Aidy SE, Hugenholtz F, van der Gaast-de Jongh C, Savelkoul HFJ, De Jonge MI, Boekschoten MV, Smidt H, Faas MM, de Vos P. Aged gut microbiota contributes to systemical inflammaging after transfer to germ-free mice. Front Immunol. 2017;8:1385. doi: 10.3389/fimmu.2017.01385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Fröhlich EE, Farzi A, Mayerhofer R, Reichmann F, Jačan A, Wagner B, Zinser E, Bordag N, Magnes C, Fröhlich E, Kashofer K, Gorkiewicz G, Holzer P. Cognitive impairment by antibiotic-induced gut dysbiosis: Analysis of gut microbiota-brain communication. Brain Behav Immun. 2016;56:140–155. doi: 10.1016/j.bbi.2016.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Furness JB. The enteric nervous system and neurogastroenterology. Nat Rev Gastroenterol Hepatol. 2012;9:286–294. doi: 10.1038/nrgastro.2012.32. [DOI] [PubMed] [Google Scholar]
- 38.Furusawa Y, Obata Y, Fukuda S, Endo TA, Nakato G, Takahashi D, Nakanishi Y, Uetake C, Kato K, Kato T, Takahashi M, Fukuda NN, Murakami S, Miyauchi E, Hino S, Atarashi K, Onawa S, Fujimura Y, Lockett T, Clarke JM, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature. 2013;504:446–450. doi: 10.1038/nature12721. [DOI] [PubMed] [Google Scholar]
- 39.Ghosh S, Lertwattanarak R, Garduño Jde J, Galeana JJ, Li J, Zamarripa F, Lancaster JL, Mohan S, Hussey S, Musi N. Elevated muscle TLR4 expression and metabolic endotoxemia in human aging. J Gerontol A Biol Sci Med Sci. 2015;70:232–246. doi: 10.1093/gerona/glu067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gopalakrishnan V, Dozier EA, Glover MS, Novick S, Ford M, Morehouse C, Warrener P, Caceres C, Hess S, Sellman BR, Cohen TS. Engraftment of bacteria after fecal microbiota transplantation is dependent on both frequency of dosing and duration of preparative antibiotic regimen. Microorganisms. 2021;9:1399. doi: 10.3390/microorganisms9071399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Gorecki AM, Preskey L, Bakeberg MC, Kenna JE, Gildenhuys C, MacDougall G, Dunlop SA, Mastaglia FL, Akkari PA, Koengten F, Anderton RS. Altered gut microbiome in Parkinson's disease and the influence of lipopolysaccharide in a human alpha-synuclein over-expressing mouse model. Front Neurosci. 2019;13:839. doi: 10.3389/fnins.2019.00839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Graeber MB, Streit W. Microglia: biology and pathology. Acta Neuropathol. 2010;119:89–105. doi: 10.1007/s00401-009-0622-0. [DOI] [PubMed] [Google Scholar]
- 43.Harach T, Marungruang N, Duthilleul N, Cheatham V, Mc Coy KD, Frisoni G, Neher JJ, Fåk F, Jucker M, Lasser T, Bolmont T. Reduction of Abeta amyloid pathology in APPPS1 transgenic mice in the absence of gut microbiota. Sci Rep. 2017;7:41802. doi: 10.1038/srep41802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Harsch IA, Konturek PC. Adhesion ileus after fecal microbiota transplantation in long-standing radiation colitis. Case Rep Gastrointest Med. 2019;2019:2543808. doi: 10.1155/2019/2543808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Haseeb A, Shahzad I, Ghulam H, Muhammad NF. IntechOpen. London, UK: 2019. Gut microbiome: a new organ system in body in parasitology and microbiology research. [Google Scholar]
- 46.Ho ST, Hsieh YT, Wang SY, Chen MJ. Improving effect of a probiotic mixture on memory and learning abilities in d-galactose-treated aging mice. J Dairy Sci. 2019;102:1901–1909. doi: 10.3168/jds.2018-15811. [DOI] [PubMed] [Google Scholar]
- 47.Hoffman JD, Parikh I, Green SJ, Chlipala G, Mohney RP, Keaton M, Bauer B, Hartz AMS, Lin AL. Age drives distortion of brain metabolic, vascular and cognitive functions, and the gut microbiome. Front Aging Neurosci. 2017;9:298. doi: 10.3389/fnagi.2017.00298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hollingsworth BA, Cassatt DR, DiCarlo AL, Rios CI, Satyamitra MM, Winters TA, Taliaferro LP. Acute radiation syndrome and the microbiome: impact and review. Front Pharmacol. 2021;12:643283. doi: 10.3389/fphar.2021.643283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science. 2012;336:1268–1273. doi: 10.1126/science.1223490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Hsiao EY, McBride SW, Hsien S, Sharon G, Hyde ER, McCue T, Codelli JA, Chow J, Reisman SE, Petrosino JF, Patterson PH, Mazmanian SK. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell. 2013;155:1451–1463. doi: 10.1016/j.cell.2013.11.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Iadecola C, Duering M, Hachinski V, Joutel A, Pendlebury ST, Schneider JA, Dichgans M. Vascular cognitive impairment and dementia: JACC scientific expert panel. J Am Coll Cardiol. 2019;73:3326–3344. doi: 10.1016/j.jacc.2019.04.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Jeffery IB, Lynch DB, O'Toole PW. Composition and temporal stability of the gut microbiota in older persons. ISME J. 2016;10:170–182. doi: 10.1038/ismej.2015.88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Kim CS, Cha L, Sim M, Jung S, Chun WY, Baik HW, Shin DM. Probiotic supplementation improves cognitive function and mood with changes in gut microbiota in community-dwelling older adults: a randomized, double-blind, placebo-controlled, multicenter trial. J Gerontol A Biol Sci Med Sci. 2021;76:32–40. doi: 10.1093/gerona/glaa090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kim KA, Jeong JJ, Yoo SY, Kim DH. Gut microbiota lipopolysaccharide accelerates inflamm-aging in mice. BMC Microbiol. 2016;16:9. doi: 10.1186/s12866-016-0625-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Kim S, Covington A, Pamer EG. The intestinal microbiota: Antibiotics, colonization resistance, and enteric pathogens. Immunol Rev. 2017;279:90–105. doi: 10.1111/imr.12563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.König J, Wells J, Cani PD, García-Ródenas CL, MacDonald T, Mercenier A, Whyte J, Troost F, Brummer RJ. Human intestinal barrier function in health and disease. Clin Transl Gastroenterol. 2016;7:e196. doi: 10.1038/ctg.2016.54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Kotani A, Miyaguchi Y, Kohama M, Ohtsuka T, Shiratori T, Kusu F. Determination of short-chain fatty acids in rat and human feces by high-performance liquid chromatography with electrochemical detection. Anal Sci. 2009;25:1007–1011. doi: 10.2116/analsci.25.1007. [DOI] [PubMed] [Google Scholar]
- 58.Kwok LY, Wang L, Zhang J, Guo Z, Zhang H. A pilot study on the effect of Lactobacillus casei Zhang on intestinal microbiota parameters in Chinese subjects of different age. Benef Microbes. 2014;5:295–304. doi: 10.3920/BM2013.0047. [DOI] [PubMed] [Google Scholar]
- 59.Leung K, Thuret S. Gut microbiota: a modulator of brain plasticity and cognitive function in ageing. Healthcare (Basel) 2015;3:898–916. doi: 10.3390/healthcare3040898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Levy M, Kolodziejczyk AA, Thaiss CA, Elinav E. Dysbiosis and the immune system. Nat Rev Immunol. 2017;17:219–232. doi: 10.1038/nri.2017.7. [DOI] [PubMed] [Google Scholar]
- 61.Li S, Shao Y, Li K, HuangFu C, Wang W, Liu Z, Cai Z, Zhao B. Vascular cognitive impairment and the gut microbiota. J Alzheimers Dis. 2018;63:1209–1222. doi: 10.3233/JAD-171103. [DOI] [PubMed] [Google Scholar]
- 62.Li Y, Ning L, Yin Y, Wang R, Zhang Z, Hao L, Wang B, Zhao X, Yang X, Yin L, Wu S, Guo D, Zhang C. Age-related shifts in gut microbiota contribute to cognitive decline in aged rats. Aging. 2020;12:7801–7817. doi: 10.18632/aging.103093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Lyon L. 'All disease begins in the gut': was Hippocrates right? Brain. 2018;141:e20. doi: 10.1093/brain/awy017. [DOI] [PubMed] [Google Scholar]
- 64.Manderino L, Carroll I, Azcarate-Peril MA, Rochette A, Heinberg L, Peat C, Steffen K, Mitchell J, Gunstad J. Preliminary evidence for an association between the composition of the gut microbiome and cognitive function in neurologically healthy older adults. J Int Neuropsychol Soc. 2017;23:700–705. doi: 10.1017/S1355617717000492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Mariat D, Firmesse O, Levenez F, Guimarăes V, Sokol H, Doré J, Corthier G, Furet JP. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009;9:123. doi: 10.1186/1471-2180-9-123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Matcovitch-Natan O, Winter DR, Giladi A, Vargas Aguilar S, Spinrad A, Sarrazin S, Ben-Yehuda H, David E, Zelada González F, Perrin P, Keren-Shaul H, Gury M, Lara-Astaiso D, Thaiss CA, Cohen M, Bahar Halpern K, Baruch K, Deczkowska A, Lorenzo-Vivas E, Itzkovitz S, et al. Microglia development follows a stepwise program to regulate brain homeostasis. Science. 2016;353:aad8670. doi: 10.1126/science.aad8670. [DOI] [PubMed] [Google Scholar]
- 67.McKernan DP, Dennison U, Gaszner G, Cryan JF, Dinan TG. Enhanced peripheral toll-like receptor responses in psychosis: further evidence of a pro-inflammatory phenotype. Transl Psychiatry. 2011;1:e36. doi: 10.1038/tp.2011.37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Minter MR, Zhang C, Leone V, Ringus DL, Zhang X, Oyler-Castrillo P, Musch MW, Liao F, Ward JF, Holtzman DM, Chang EB, Tanzi RE, Sisodia SS. Antibiotic-induced perturbations in gut microbial diversity influences neuro-inflammation and amyloidosis in a murine model of Alzheimer's disease. Sci Rep. 2016;6:30028. doi: 10.1038/srep30028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Mueller S, Saunier K, Hanisch C, Norin E, Alm L, Midtvedt T, Cresci A, Silvi S, Orpianesi C, Verdenelli MC, Clavel T, Koebnick C, Zunft HJ, Doré J, Blaut M. Differences in fecal microbiota in different European study populations in relation to age, gender, and country: a cross-sectional study. Appl Environ Microbiol. 2006;72:1027–1033. doi: 10.1128/AEM.72.2.1027-1033.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Musa HH, Wu SL, Zhu CH, Seri HI, Zhu GQ. The potential benefits of probiotics in animal production and health. Journal of Animal and Veterinary Advances. 2009;8:313–321. [Google Scholar]
- 71.Nguyen TL, Vieira-Silva S, Liston A, Raes J. How informative is the mouse for human gut microbiota research? Dis Model Mech. 2015;8:1–16. doi: 10.1242/dmm.017400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.O'Mahony SM, Marchesi JR, Scully P, Codling C, Ceolho AM, Quigley EM, Cryan JF, Dinan TG. Early life stress alters behavior, immunity, and microbiota in rats: implications for irritable bowel syndrome and psychiatric illnesses. Biol Psychiatry. 2009;65:263–267. doi: 10.1016/j.biopsych.2008.06.026. [DOI] [PubMed] [Google Scholar]
- 73.Peters R. Ageing and the brain. Postgrad Med J. 2006;82:84–88. doi: 10.1136/pgmj.2005.036665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Petersen C, Round JL. Defining dysbiosis and its influence on host immunity and disease. Cell Microbiol. 2014;16:1024–1033. doi: 10.1111/cmi.12308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Rabin LA, Smart CM, Amariglio RE. Subjective cognitive decline in preclinical Alzheimer's disease. Annu Rev Clin Psychol. 2017;13:369–396. doi: 10.1146/annurev-clinpsy-032816-045136. [DOI] [PubMed] [Google Scholar]
- 76.Rezai-Zadeh K, Gate D, Town T. CNS infiltration of peripheral immune cells: D-Day for neurodegenerative disease? J Neuroimmune Pharmacol. 2009;4:462–475. doi: 10.1007/s11481-009-9166-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Rios-Covian D, Ruas-Madiedo P, Margolles A, Gueimonde M, de Los Reyes-Gavilan CG, Salazar N. Intestinal short chain fatty acids and their link with diet and human health. Front Microbiol. 2016;7:185. doi: 10.3389/fmicb.2016.00185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018;9:3294. doi: 10.1038/s41467-018-05470-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Romo-Araiza A, Gutiérrez-Salmeán G, Galván EJ, Hernández-Frausto M, Herrera-López G, Romo-Parra H, García-Contreras V, Fernández-Presas AM, Jasso-Chávez R, Borlongan CV, Ibarra A. Probiotics and prebiotics as a therapeutic strategy to improve memory in a model of middle-aged rats. Front Aging Neurosci. 2018;10:416. doi: 10.3389/fnagi.2018.00416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Rothhammer V, Borucki DM, Tjon EC, Takenaka MC, Chao CC, Ardura-Fabregat A, de Lima KA, Gutiérrez-Vázquez C, Hewson P, Staszewski O, Blain M, Healy L, Neziraj T, Borio M, Wheeler M, Dragin LL, Laplaud DA, Antel J, Alvarez JI, Prinz M, et al. Microglial control of astrocytes in response to microbial metabolites. Nature. 2018;557:724–728. doi: 10.1038/s41586-018-0119-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Rothhammer V, Mascanfroni ID, Bunse L, Takenaka MC, Kenison JE, Mayo L, Chao CC, Patel B, Yan R, Blain M, Alvarez JI, Kébir H, Anandasabapathy N, Izquierdo G, Jung S, Obholzer N, Pochet N, Clish CB, Prinz M, Prat A, et al. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat Med. 2016;22:586–597. doi: 10.1038/nm.4106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Saji N, Niida S, Murotani K, Hisada T, Tsuduki T, Sugimoto T, Kimura A, Toba K, Sakurai T. Analysis of the relationship between the gut microbiome and dementia: a cross-sectional study conducted in Japan. Sci Rep. 2019;9:1008. doi: 10.1038/s41598-018-38218-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Salami M. Interplay of good bacteria and central nervous system: cognitive aspects and mechanistic considerations. Front Neurosci. 2021;15:613120. doi: 10.3389/fnins.2021.613120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Sanborn V, Azcarate-Peril MA, Updegraff J, Manderino L, Gunstad J. Randomized clinical trial examining the impact of lactobacillus rhamnosus GG probiotic supplementation on cognitive functioning in middle-aged and older adults. Neuropsychiatr Dis Treat. 2020;16:2765–2777. doi: 10.2147/NDT.S270035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Schwartz M, Gluck M, Koon S. Norovirus gastroenteritis after fecal microbiota transplantation for treatment of Clostridium difficile infection despite asymptomatic donors and lack of sick contacts. Am J Gastroenterol. 2013;108:1367. doi: 10.1038/ajg.2013.164. [DOI] [PubMed] [Google Scholar]
- 86.Sender R, Fuchs S, Milo R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016;14:e1002533. doi: 10.1371/journal.pbio.1002533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Sheng C, Lin L, Lin H, Wang X, Han Y, Liu SL. Altered gut microbiota in adults with subjective cognitive decline: the SILCODE study. J Alzheimers Dis. 2021;82:513–526. doi: 10.3233/JAD-210259. [DOI] [PubMed] [Google Scholar]
- 88.Shin NR, Whon TW, Bae JW. Proteobacteria: microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015;33:496–503. doi: 10.1016/j.tibtech.2015.06.011. [DOI] [PubMed] [Google Scholar]
- 89.Shreiner AB, Kao JY, Young VB. The gut microbiome in health and in disease. Curr Opin Gastroenterol. 2015;31:69–75. doi: 10.1097/MOG.0000000000000139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Spychala MS, Venna VR, Jandzinski M, Doran SJ, Durgan DJ, Ganesh BP, Ajami NJ, Putluri N, Graf J, Bryan RM, McCullough LD. Age-related changes in the gut microbiota influence systemic inflammation and stroke outcome. Ann Neurol. 2018;84:23–36. doi: 10.1002/ana.25250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Stehle JR, Jr, Leng X, Kitzman DW, Nicklas BJ, Kritchevsky SB, High KP. Lipopolysaccharide-binding protein, a surrogate marker of microbial translocation, is associated with physical function in healthy older adults. J Gerontol A Biol Sci Med Sci. 2012;67:1212–1218. doi: 10.1093/gerona/gls178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Stenman LK, Patterson E, Meunier J, Roman FJ, Lehtinen MJ. Strain specific stress-modulating effects of candidate probiotics: A systematic screening in a mouse model of chronic restraint stress. Behav Brain Res. 2020;379:112376. doi: 10.1016/j.bbr.2019.112376. [DOI] [PubMed] [Google Scholar]
- 93.Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu XN, Kubo C, Koga Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J Physiol. 2004;558:263–275. doi: 10.1113/jphysiol.2004.063388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Umesaki Y, Setoyama H, Matsumoto S, Imaoka A, Itoh K. Differential roles of segmented filamentous bacteria and clostridia in development of the intestinal immune system. Infect Immun. 1999;67:3504–3511. doi: 10.1128/iai.67.7.3504-3511.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Villarán RF, Espinosa-Oliva AM, Sarmiento M, De Pablos RM, Argüelles S, Delgado-Cortés MJ, Sobrino V, Van Rooijen N, Venero JL, Herrera AJ, Cano J, Machado A. Ulcerative colitis exacerbates lipopolysaccharide-induced damage to the nigral dopaminergic system: potential risk factor in Parkinson`s disease. J Neurochem. 2010;114:1687–1700. doi: 10.1111/j.1471-4159.2010.06879.x. [DOI] [PubMed] [Google Scholar]
- 96.Vogt NM, Kerby RL, Dill-McFarland KA, Harding SJ, Merluzzi AP, Johnson SC, Carlsson CM, Asthana S, Zetterberg H, Blennow K, Bendlin BB, Rey FE. The impact of intestinal microbiota on bio-medical research: definitions, techniques and physiology of a “new frontier”. Acta Biomed. 2018;89:52–59. doi: 10.23750/abm.v89i9-S.7906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Vogt NM, Kerby RL, Dill-McFarland KA, Harding SJ, Merluzzi AP, Johnson SC, Carlsson CM, Asthana S, Zetterberg H, Blennow K, Bendlin BB, Rey FE. Gut microbiome alterations in Alzheimer's disease. Sci Rep. 2017;7:13537. doi: 10.1038/s41598-017-13601-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Wang HX, Wang YP. Gut microbiota-brain axis. Chin Med J (Engl) 2016;129:2373–2380. doi: 10.4103/0366-6999.190667. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Wang S, Ahmadi S, Nagpal R, Jain S, Mishra SP, Kavanagh K, Zhu X, Wang Z, McClain DA, Kritchevsky SB, Kitzman DW, Yadav H. Lipoteichoic acid from the cell wall of a heat killed Lactobacillus paracasei D3-5 ameliorates aging-related leaky gut, inflammation and improves physical and cognitive functions: from C. elegans to mice. Geroscience. 2020;42:333–352. doi: 10.1007/s11357-019-00137-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Westfall S, Lomis N, Kahouli I, Dia SY, Singh SP, Prakash S. Microbiome, probiotics and neurodegenerative diseases: deciphering the gut brain axis. Cell Mol Life Sci. 2017;74:3769–3787. doi: 10.1007/s00018-017-2550-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Williamson LL, Sholar PW, Mistry RS, Smith SH, Bilbo SD. Microglia and memory: modulation by early-life infection. J Neurosci. 2011;31:15511–15521. doi: 10.1523/JNEUROSCI.3688-11.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Winek K, Dirnagl U, Meisel A. The gut microbiome as therapeutic target in central nervous system diseases: implications for stroke. Neurotherapeutics. 2016;13:762–774. doi: 10.1007/s13311-016-0475-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Woodmansey EJ, McMurdo ME, Macfarlane GT, Macfarlane S. Comparison of compositions and metabolic activities of fecal microbiotas in young adults and in antibiotic-treated and non-antibiotic-treated elderly subjects. Appl Environ Microbiol. 2004;70:6113–6122. doi: 10.1128/AEM.70.10.6113-6122.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Wu ML, Yang XQ, Xue L, Duan W, Du JR. Age-related cognitive decline is associated with microbiota-gut-brain axis disorders and neuroinflammation in mice. Behav Brain Res. 2021;402:113125. doi: 10.1016/j.bbr.2021.113125. [DOI] [PubMed] [Google Scholar]
- 105.Wymore Brand M, Wannemuehler MJ, Phillips GJ, Proctor A, Overstreet AM, Jergens AE, Orcutt RP, Fox JG. The altered schaedler flora: continued applications of a defined murine nicrobial community. ILAR J. 2015;56:169–178. doi: 10.1093/ilar/ilv012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Xiao J, Katsumata N, Bernier F, Ohno K, Yamauchi Y, Odamaki T, Yoshikawa K, Ito K, Kaneko T. Probiotic bifidobacterium breve in iImproving cognitive functions of older adults with suspected mild cognitive impairment: a randomized, double-blind, placebo-controlled trial. J Alzheimers Dis. 2020;77:139–147. doi: 10.3233/JAD-200488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Xu C, Zhu H, Qiu P. Aging progression of human gut microbiota. BMC Microbiol. 2019;19:236. doi: 10.1186/s12866-019-1616-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Yang X, Yu D, Xue L, Li H, Du J. Probiotics modulate the microbiota-gut-brain axis and improve memory deficits in aged SAMP8 mice. Acta Pharm Sin B. 2020;10:475–487. doi: 10.1016/j.apsb.2019.07.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Zhao Y, Jaber V, Lukiw WJ. Secretory products of the human GI tract microbiome and their potential impact on Alzheimer's disease (AD): detection of lipopolysaccharide (LPS) in AD hippocampus. Front Cell Infect Microbiol. 2017;7:318. doi: 10.3389/fcimb.2017.00318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Zhuang ZQ, Shen LL, Li WW, Fu X, Zeng F, Gui L, Lü Y, Cai M, Zhu C, Tan YL, Zheng P, Li HY, Zhu J, Zhou HD, Bu XL, Wang YJ. Gut microbiota is altered in patients with Alzheimer's disease. J Alzheimers Dis. 2018;63:1337–1346. doi: 10.3233/JAD-180176. [DOI] [PubMed] [Google Scholar]
- 111.Zwielehner J, Liszt K, Handschur M, Lassl C, Lapin A, Haslberger AG. Combined PCR-DGGE fingerprinting and quantitative-PCR indicates shifts in fecal population sizes and diversity of Bacteroides, bifidobacteria and Clostridium cluster IV in institutionalized elderly. Exp Gerontol. 2009;44:440–446. doi: 10.1016/j.exger.2009.04.002. [DOI] [PubMed] [Google Scholar]