

Abstract
Clonal hematopoiesis of indeterminate potential (CHIP) describes a widespread expansion of genetically variant hematopoietic cells that increases exponentially with age and is associated with increased risks of cancers, cardiovascular disease, and other maladies. Here we discuss how environmental contexts associated with CHIP, such as old age, infections, chemotherapy, or cigarette smoking, alter tissue microenvironments to facilitate the selection and expansion of specific CHIP mutant clones. Further, we consider major remaining gaps in knowledge, including intrinsic effects, clone size thresholds and factors affecting clonal competition, that will determine future application of this field in transplant and preventive medicine.
Introduction
Hematopoietic stem cells (HSCs) reside in the bone marrow where they divide, differentiate, and self-renew to maintain all blood cells throughout an organism’s life. (Pinho and Frenette, 2019) Over time, HSCs naturally acquire somatic mutations, some of which enable selective advantages over other HSCs in a context-dependent fashion (Figure 1). As a result of this competition, oligoclonality in the bone marrow has been observed, initially based on patterns of X chromosome inactivation and later confirmed in normal individuals harboring TET2 mutations. (Busque et al., 2012) The term clonal hematopoiesis of indeterminate potential (CHIP) was first coined in 2015 by Steensma et al. to define a phenotype in which hematopoietic cells harboring somatic mutations clonally expand in the absence of hematological disease. (Steensma et al., 2015) A number of studies have defined the genetic landscape of CHIP, with most mutations occurring in pre-leukemic driver genes that are recurrently implicated in the pathogenesis of hematological malignancies and disorders.(Bolton et al., 2020, Genovese et al., 2014, Miller and Steensma, 2020, Jan et al., 2017) While CHIP was originally defined by detection of mutations in genes related to hematological neoplasms at a variant allele frequency (VAF) of ≥ 2%, newer methods now enable detection of CHIP at a much lower VAF. (Steensma et al., 2015)
Figure 1: Environmental stressors promote mutation-specific clonal expansion.
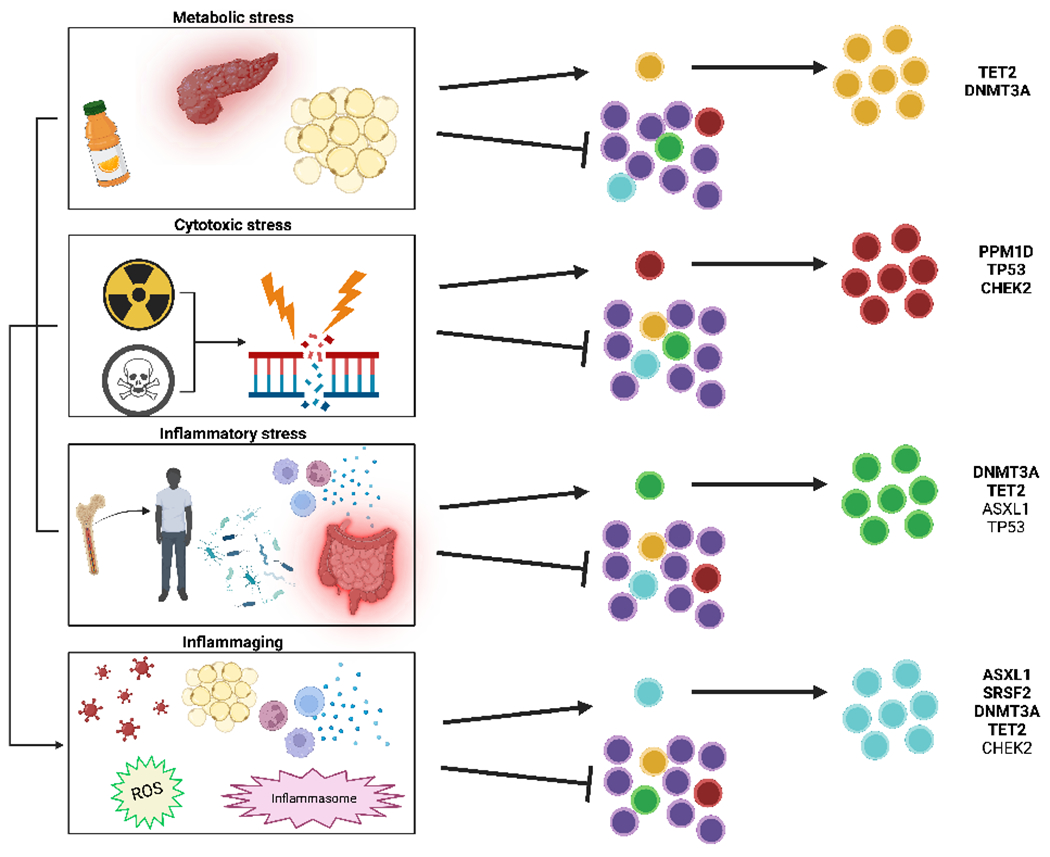
Over an organism’s lifetime, HSCs encounter cellular stresses which may induce somatic mutations in common driver genes. Depending on the context, somatic mutations may confer a selection advantage for certain clones to persist and expand. Environmental effects on clonal competition are mutation specific. Genotoxic stressors such as chemotherapy and radiation provide a selection advantage for clones bearing mutations in p53, PPM1D, and CHEK2, whereas infection and inflammation promote the expansion of clones with mutations in TET2 and DNMT3A. Studies on the metabolic effects of clonal expansion suggest that metabolites, like vitamin C, may impact the function of mutations, such as TET2.
CHIP is found, by conservative measures, in more than 10% of individuals by the eighth decade of life and poses increased risks of adverse outcomes including severe infection, kidney disease, hematological and non-hematological cancers, cardiovascular disease, and all-cause mortality. (Jaiswal et al., 2017, Genovese et al., 2014, Zink et al., 2017, Jaiswal et al., 2014, Kessler et al., 2022, Kar et al., 2022, Bolton et al., 2021, Zekavat et al., 2021a, Dawoud et al., 2020, van Deuren et al., 2021) Despite the increased prevalence of CHIP with age, CHIP mutations appear to be relatively stable over time in the absence of additional stressors. (Midic et al., 2020) This stability implies that extrinsic factors provide opportunities for somatic mutations to undergo “somatic evolution,” facilitating the selection and expansion of clones in specific microenvironments.(Laconi et al., 2020) There is an increasing body of evidence showing that particular stressors can drive expansion of specific CHIP mutant clones (Figure 1). Studies exploring the mechanisms by which specific mutant clones outcompete other HSCs and expand in certain contexts have provided insight into the selective environments that drive their expansion. Understanding the differential fitness landscapes that promote the selective expansion of CHIP variants will enhance our ability to not only manage clonal expansion but also reduce risks associated with CHIP. (King et al., 2020)
CHIP and Disease Risk
CHIP is associated with a widening array of diseases, many of which are age-associated (Table 1). Patients with CHIP have an increased risk of developing hematological malignancies, with the effect size varying according to VAF, clonal complexity, mutation type, and gene(s) mutated.(Desai et al., 2018, Abelson et al., 2018, Zink et al., 2017, Genovese et al., 2014, Jaiswal et al., 2014) Deep sequencing analysis has revealed that the risk of individuals with CHIP developing acute myeloid leukemia (AML) may occur at VAFs as low as 0.005-0.01,(Young et al., 2019, Desai et al., 2018, Abelson et al., 2018) with further increase in AML risk proportionate to VAF.(Abelson et al., 2018, Genovese et al., 2014) Mutations commonly identified in CHIP include those in epigenetic modifiers (TET2, DNMT3A, IDH1, IDH2, and ASXL1), splicing factors (U2AF1, SRSF2), DNA damage repair genes (PPM1D, TP53) and inflammatory mediators (JAK2, STAT3, MYD88), most of which are known drivers of hematological cancers. The risk of future blood cancers and overall decreased survival varies by mutation, with mutations in DNMT3A and TET2 conferring a lesser risk of progression compared to PPM1D, TP53.(Genovese et al., 2014, Abelson et al., 2018, Chou et al., 2011, Gelsi-Boyer et al., 2012, Park et al., 2020, Yuan et al., 2016a, Ley et al., 2010, Jaiswal et al., 2014, Desai et al., 2018)
Table 1:
Suspected CHIP-mediated diseases by driver mutation
DNMT3A and TET2 are the most commonly mutated genes in CHIP. (Buscarlet et al., 2017, Feusier et al., 2021, Genovese et al., 2014, Jaiswal et al., 2014, Xie et al., 2014) DNMT3A mutations are present in at least 20% of AML patients, (Gaidzik et al., 2013, Lauber et al., 2020, Park et al., 2020, Yan et al., 2011, Ley et al., 2010) with roughly 60% of these occurring at the R882 codon. (Gaidzik et al., 2013, Lauber et al., 2020, Ley et al., 2010, Park et al., 2020, Ribeiro et al., 2012) Mutations at this hotspot locus, located within the methyltransferase catalytic domain, have been linked with a higher risk of AML development and poorer outcomes compared to other mutations within the same gene. (Gaidzik et al., 2013, Kumar et al., 2018, Lauber et al., 2020, Ley et al., 2010, Renneville et al., 2012, Ribeiro et al., 2012, Yuan et al., 2016a, Young et al., 2019) Similarly, mutations in TET2, a methylcytosine dioxygenase, occur at high frequencies in myeloid neoplasms (Abdel-Wahab et al., 2009, Delhommeau et al., 2009, Xie et al., 2014), and patients harboring mutations in TET2 tend to have decreased survival and response to therapy. (Wang et al., 2019) DNMT3A and TET2 regulate methylation and demethylation of the genome, respectively, though through different mechanisms.(Izzo et al., 2020) Despite these mechanistic differences, functional studies suggest that mutations in both DNMT3A (Lauber et al., 2020, Park et al., 2020, Russler-Germain et al., 2014, Sandoval et al., 2019, Yan et al., 2011) and TET2 (Lopez-Moyado et al., 2019, Rasmussen et al., 2015, Cimmino et al., 2017) alter the expression of genes involved in proliferation, differentiation, and oncogenesis. These epigenetic changes allow the mutant cells to propagate and persist, enabling the acquisition of other mutations that further drive leukemogenesis. (Bezerra et al., 2020, Celik et al., 2015, Guryanova et al., 2016, Kronke et al., 2013, Loberg et al., 2019, McKerrell et al., 2015, Meyer et al., 2016, Yang et al., 2016, Jan et al., 2017) While the mechanistic roles of DNMT3A and TET2 in malignant hematopoiesis have been thoroughly summarized elsewhere, (Chaudry and Chevassut, 2017, Im et al., 2014, Venugopal et al., 2021, Yang et al., 2015, Bowman and Levine, 2017, Nakajima and Kunimoto, 2014, Feng et al., 2019) it is worth noting that blood cells carrying CHIP mutations may not only increase the risk of AML but also may impact non-hematological tumors by modulating the immune microenvironment.(Kleppe et al., 2015) Indeed, CHIP has been associated with a variety of solid tumors in multiple large cohort studies,(Bolton et al., 2020, Kar et al., 2022, Kessler et al., 2022) strongly suggesting that CHIP may also affect the progression of solid tumors.
In addition to its impact on cancer development and prognosis, CHIP has been noted to affect autologous stem-cell transplant (ASCT) outcomes for lymphoma. Patients with CHIP mutations had higher incidence of therapy-related myeloid neoplasms (TMNs) following ASCT. Furthermore, within this cohort, patients harboring PPM1D mutations had overall lower survival.(Gibson et al., 2017b) An increased prevalence of unexplained cytopenias in patients receiving ASCT of hematopoietic cells harboring CHIP mutations has been reported, suggesting graft dysfunction. (Gibson et al., 2017a) However, others report no adverse outcomes with CHIP post-ASCT, and even improved survival, suggesting that adverse effects may be mutation-specific (Grimm et al., 2019, Heini et al., 2021).
CHIP also has recently been implicated in impacting both risk and response to infections. Patients harboring CHIP were found to have increased risk of severe COVID-19, sepsis and other infections.(Bolton et al., 2021, Dawoud et al., 2020, Jaiswal and Ebert, 2019, Kessler et al., 2022, Zekavat et al., 2021a) These risks may be attributable to impaired function of downstream immune cells, which has been reported for cells bearing CHIP mutations.(Jaiswal and Ebert, 2019, Zekavat et al., 2021a)
One phenomenon of great interest related to CHIP is its link to cardiovascular disease and all cause-mortality.(Jaiswal et al., 2014, Sano et al., 2018c, Libby et al., 2019, Dorsheimer et al., 2019, Cremer et al., 2020, Abegunde et al., 2018, Kar et al., 2022, Kessler et al., 2022) One of the earliest studies found that CHIP carriers had a two-fold risk of coronary heart disease and ischemic stroke (Jaiswal 2014). Subsequent studies in patient cohorts found a four-times greater risk of having early myocardial infarction (MI),(Jaiswal et al., 2017) and congestive heart failure in CHIP patients(Dorsheimer et al., 2019), independent of traditional cardiovascular risk factors. Recently, however, two recent large epidemiological studies using the UK Biobank indicate that the increased risks of cardiovascular disease may be less dramatic (HR 1.1, 1.03-1.17).(Kar et al., 2022, Kessler et al., 2022) Specific mutations, including TET2, DNMT3A and ASXL1 showed an increased incidence of coronary artery disease, coronary artery calcifications, and early MI, with risks of cardiovascular disease that correlate with VAF.(Jaiswal et al., 2014, Bick et al., 2020a, Kar et al., 2022, Jaiswal et al., 2017) Studies using both human samples and mouse models revealed the pro-inflammatory environment generated by the mutant clones drives adverse cardiovascular outcomes.(Jaiswal et al., 2017, Fuster et al., 2017, Sano et al., 2018a, Bick et al., 2020a) However, differing pro-inflammatory phenotypes drive cardiovascular dysfunction and remodeling in CHIP with mutations in PPM1D (Yura et al., 2021), TP53 (Sano et al., 2021), DNMT3A (Abplanalp et al., 2021, Heyde et al., 2021, Sano et al., 2018c) and TET2 (Sano et al., 2018b, Wang et al., 2020), respectively. CHIP also has been linked to COPD,(Buscarlet et al., 2017, Miller et al., 2021a, Zink et al., 2017) diabetes,(Fuster et al., 2020, Jaiswal et al., 2014) psychiatric illnesses,(Zink et al., 2017) early menopause,(Honigberg et al., 2021) osteoporosis,(Kim et al., 2021) liver disease,(Wong et al., 2022) renal dysfunction,(Dawoud et al., 2021) and epigenetic aging.(Robertson et al., 2019) The correlation between these diseases and CHIP could be manifestations of tissue dysfunction due to common causes, like aging, smoking, and inflammation; however, it is possible that CHIP-driven inflammation contributes to the development of these diseases. As we discuss below, inflammation itself is a driver of CHIP, setting up a feed-forward loop of CHIP and disease. Thus, CHIP has important implications for the future management of patients, most immediately in relation to cardiovascular outcomes and precision oncology.(Libby et al., 2019, Miller and Steensma, 2020, Steensma, 2018, Sidlow et al., 2020)
It is worth noting that the factors that drive CHIP may have little to do with the mechanisms by which mutant clones promote disease. Epidemiologic studies will continue to be important to uncover key disease associations. Importantly, understanding what drives CHIP and its health consequences will provide two different avenues for preventive medicine and therapeutic management, respectively. With an eye toward prevention, here we will highlight known associations between environmental stressors and CHIP, including mechanisms of clonal expansion where known.
Chemotherapy and Radiation as Drivers of CHIP
Chemotherapies and radiation destroy malignant cells by inducing DNA damage, impairing DNA replication, damaging DNA repair mechanisms, inhibiting pro-survival signaling pathways, impairing transcription and translation, and evoking metabolic and cellular stress.(Krisl and Doan, 2017) In addition to cancer treatments, chemotherapy and radiation are useful in managing bone marrow diseases,(Lee et al., 2018) transplants,(Krisl and Doan, 2017) immunodeficiencies,(Lum et al., 2019) and autoimmune diseases(2004) where cytotoxic conditioning regimens make space for engraftment, prevent rejection, and dampen an overactive immune system. These reagents universally impair the hematopoietic system either as the primary goal or as an off-target effect of tumor targeting.(Shao et al., 2013) Quiescent HSCs are more protected against the acute stress of chemotherapy and radiation in part via induction of a strong P53-dependent DNA damage response (DDR), which promotes senescence instead of cell death(Mohrin et al., 2010). However, HSCs eventually lose clonogenic activity, undergo cell death, and have decreased fitness, defined as overall ability to persist and expand, in a P53-dependent manner.(Marusyk et al., 2010, Bondar and Medzhitov, 2010) Furthermore, the quiescent nature of HSCs limits DNA repair primarily due to the use of error-prone non-homologous end joining.(Mohrin et al., 2010) Thus, HSC survival comes at the expense of increased risk of mutagenesis and dampened HSC expansion.(Mohrin et al., 2010, Schoedel et al., 2016) Specific pre-existing CHIP mutants are evolutionarily advantaged to expand and persist under the stress of chemotherapy and radiation. Multiple reports have shown patients with a prior history of cancer and cancer treatment are more likely to have certain CHIP mutations (Bolton et al., 2020, Boucai et al., 2018, Coombs et al., 2017, Olszewski et al., 2019, Hsu et al., 2018, Kahn et al., 2018, Wong et al., 2018), with selective clonal expansion dependent on the mechanism of the cytotoxic stressor as described below (see also Table 2).
Table 2:
Extrinsic Mechanisms of Clonal Expansion
| Class | Gene | Mechanism | Description | Model | Reference(s) |
|---|---|---|---|---|---|
| Aging | |||||
|
| |||||
| CHIP | next generation sequencing | Cross sectional study association | human | Jaiswal et al., 2014(Jaiswal et al., 2014), Xie et al., 2014(Xie et al., 2014), Buscarlet et al., 2017(Buscarlet et al., 2017), van Zeventer et al., 2021(van Zeventer et al., 2021) | |
|
|
|||||
| DNMT3A, TET2 | exome sequencing | Association Study between CH risk and longer leukocyte telomere length | human n=200,453 | Kar et al., 2022 (Kar et al., 2022) | |
|
|
|||||
| ASXL1 | Truncated ASXL1 | Truncated ASXL1 expands in murine model of aging via interactions with BAP1 to activate mTORC for survival | mouse | Fujino et al., 2021 (Fujino et al., 2021) | |
|
| |||||
| Chemotherapy/Radiation | |||||
|
| |||||
| CHIP | Radioactive iodine (RAI) | Cross-sectional study showed RAI was linked with increased prevalence of CH | human n= 279 | Boucai et al., 2018 (Boucai et al., 2018) | |
|
|
|||||
| CHIP | Chemotherapy followed by ASCT | Longitudinal association study found MRD-samples treated with chemotherapy-ASCT was linked to CHIP expansion | human n=149 | Eskelund et al., 2020 (Eskelund et al., 2020) | |
|
|
|||||
| PPM1D | Chemotherapy (including topoisomerase I and II inhibitors, taxanes, platinum-based therapies, etoposide cytarabine, doxorubicin), XRT, Radionuclide | Multiple cross-sectional studies, mouse model, and in vitro experiments linked PPM1D to specific cytotoxic stressors | human 10,138 human n=5,649 human n=119 human n=686 human n=1,185 mouse, |
Bolton et al., 2020 (Bolton et al., 2020) Coombs et al., 2017 (Coombs et al., 2017) Wong et al., 2018 (Wong et al., 2018) Swisher et al., 2016 (Swisher et al., 2016) Kim et al., 2019 (Kim et al., 2019) Hsu et al., 2018 (Hsu et al., 2018) Kahn et al., 2018 (Kahn et al., 2018) | |
|
|
|||||
| CHEK2 | Chemotherapy (Topoisomerase II inhibitors, Platinum based therapies) | Cross-sectional study | human n= 10,138 | Bolton et al., 2020 (Bolton et al., 2020) | |
|
|
|||||
| TP53 | Chemotherapy (including platinum base therapies, taxanes, 5-FU), XRT | Multiple cross-sectional studies, and mouse models that show TP53 selective advantage under these cytotoxic stressors | human n= 10,138 human n= 5,649 human n=119 Mouse models |
Bolton et al., 2020 (Bolton et al., 2020) Coombs et al., 2017(Coombs et al., 2017) Wong et al., 2018 (Wong et al., 2018) Bondar et al., 2010(Bondar and Medzhitov, 2010) Marusyk et al., 2010(Marusyk et al., 2010) Wlodarski et al., 1998 (Wlodarski et al., 1998) | |
|
|
|||||
| TP53 | n-ethyl-n-nitrosourea | Treatment of Tp53+/− HSPCs showed competitive advantage | mouse | Wong et al., 2015 (Wong et al., 2015) | |
| TET2 | Chemotherapy (including doxorubicin and cytosine arabinoside) | TET2KOAML cells are less sensitive to treatment; cross sectional study linked to TET2. | AML cell lines; human n=5,649 | Morinishi et al., 2020 (Morinishi et al., 2020) Coombs et al., 2017 (Coombs et al., 2017) | |
| TET2 | Spaceflight (XRT) | Astronauts that experienced space flight had earlier expansion of TET2. | human, n=2 | Mencia-Trinchant et al., 2020 (Mencia-Trinchant et al., 2021) | |
| DNMT3A | Busulfan | Dnmt3a−/− HSCs expand with treatment. | mouse | Chen et al., 2020 (Chen et al., 2020) | |
|
| |||||
| Tobacco/Smoking | |||||
|
| |||||
| ASXL1 | Current and former smoking history | Multiple cross-sectional studies | human n=5,649 n=502,524 n= 200,453 n= 628.388 n= 10,138 |
Coombs et al., 2017 (Coombs et al., 2017) Dawoud et al., 2020(Dawoud et al., 2020) Kar et al., 2022 (Kar et al., 2022) Kessler et al., 2022(Kessler et al., 2022) Bolton et al., 2020 (Bolton et al., 2020) | |
| DNMT3A, TET2 | Current/former tobacco use and smoking | Cross-sectional study | human n=5,649 n=502,524 |
Coombs et al., 2017 (Coombs et al., 2017) Dawoud et al., 2020(Dawoud et al., 2020) | |
| PPM1D | Current/former tobacco use | Cross-sectional study | human n=5,649 | Coombs et al., 2017 (Coombs et al., 2017 | |
| DNMT3A, SRSF2, SR3B1 | Smoking status (current) | Cross-sectional study | human n=200,453 | Kar et al., 2022 (Kar et al., 2022) | |
| CHIP | Smoking history | Cross-sectional study | human n=12,380 | Genovese et al., 2014 (Genovese et al., 2014) | |
|
| |||||
| Inflammation | |||||
|
| |||||
| DNMT3A | Elevated serum MIP1a | Cross sectional study showing increased risk of DNMT3A-CHIP | human n=200,453 | Kar et al., 2022 (Kar et al., 2022) | |
| DNMT3A | Chronic IFNy signaling via M. avium infection | Infection promotes DNMT3A KO expansion due to resistance to inflammation-mediated myeloid differentiation. | mouse | Hormaechea-Agulla et al., 2021 (Hormaechea-Agulla et al., 2021) | |
| TET2 | LPS injection | TET2 clones selectively expanded via lncRNA Morrbid and aTLR4/NK-kB/IL-6/STAT3 pathway | mouse | Cai et al., 2018 (Cai et al., 2018) | |
| TET2 | TNFa signaling | TET2 selectively expanded with direct TNFa signaling | mouse | Abegunde et al., 2018 Abegunde et al., 2018) | |
| TET2 | Disruption of intestinal barrier integrity | Increased microbial translocation and signaling, particularly from Lactobacillus, promotes inflammation and TET2 expansion | mouse | Meisel et al., 2018 (Meisel et al., 2018) | |
| TET2 | broad spectrum antibiotics | Antibiotics slowed the expansion of TET2-deficient clones | mouse | Zeng et al., 2019 (Zeng et al., 2019) | |
| CEBPa | Direct injection of IL-1b | IL-1b promotes KO CEBPa MPP3 clonal expansion, while CEBPa activates wildtype competitors for myeloid expansion and exhaustion. | mouse | Higa et al., 2021 (Higa et al., 2021) | |
| ASXL1 | Tissue editing With Inducible Stem cell Tagging via Recombination (TWISTR) knock-in of CH genes and single cell analysis | Single cell sequencing of knocked-in CHIP homologs zebrafish HSCs demonstrated resistance to inflammation in CH stem cells and promotion of cytokine production in myeloid lineage cells. | zebrafish | Avagyan et al., 2021(Avagyan et al., 2021) | |
| TP53 | poly:IC serial injection | TP53 clones were significantly expanded via IFNy production caused by poly:IC treatment in competitive transplant models | mouse | Rodriguez-Meira et al., 2022 (Rodriguez-Meira et al., 2022) | |
|
| |||||
| Autoimmunity | |||||
|
| |||||
| DNMT3A and PPM1D | Inflammatory environment caused by inflammatory bowel disease | Cross sectional study found higher prevalence of CHIP in patients with ulcerative colitis | human n=187 | Zhang et al., 2019 (Zhang et al., 2019) | |
|
| |||||
| HSCT | |||||
|
| |||||
| DNMT3A | Autologous HSCT | Cross sectional study found DNMT3A associated with HSCT | human n= 35 | Wong et al., 2018 (Wong et al., 2018) | |
| DNMT3A | Murine competitive transplant | DNMT3A null HSCs outcompete wildtype HSCs in murine transplant settings | mouse | Challen et al., 2011(Challen et al., 2011) Jeong et al., 2018(Jeong et al., 2018) | |
| PPM1D | Murine competitive transplant | Truncated PPM1D R451X HSCs fails to outcompete wildtype HSCs in competitive murine transplant settings | mouse | Hsu et al., 2018(Hsu et al., 2018) | |
| TET2 | Serial HSC transplant | Tet2 null HSCs exhaust similarly to wildtype HSCs in serial transplant settings | mouse | Ostrander et al., 2020(Ostrander et al., 2020) | |
| ASXL1 | Murine competitive transplant | Truncated ASXL1 are at a disadvantage in engraftment and cell function in a murine transplant setting | mouse | Fujino et al., 2021(Fujino et al., 2021) | |
| SRSF2 | Murine competitive transplant | Heterozygous Srsf2 P95H HSCs cannot outcompete WT HSCs in engraftment or lineage output in competitive murine transplants | mouse | Kon et al., 2018(Kon et al., 2018) | |
| SF3B1 | Murine competitive transplant | Sf3b1-K700E mutant HSCs are at a disadvantage compared to wild-type HSCs in both young or old recipient mice. | mouse | Mupo et al., 2017(Mupo et al., 2017) | |
| U2AF1 | Murine competitive transplant | U2AF1 chimerism in transplanted mice is significantly reduced in mutant U2AF1 variants: S34F, Q157P, and Q157R. | mouse | Yoshida et al., 2011(Yoshida et al., 2011) | |
|
| |||||
| Metabolic | |||||
|
| |||||
| TET2 | Hyperglycemia | Hyperglycemia drives TET2 myelopoiesis and MPN. | mouse | Cai et al., 2021(Cai et al., 2021) | |
| TET2 | Vitamin C deficiency | Cross-sectional study where lower plasma vitamin C concentrations was associated with a higher frequency of TET2 mutations | human n = 215 | Chen et al., 2021(Chen et al., 2021) | |
| TET2 | Apolipoprotein B | High ApoB was associated with TET2 clonal expansion, whereas alcohol consumption was protective in this cross-sectional study | human n = 200,453 | Kar et al., 2022(Kar et al., 2022) | |
| TET2, DNMT3A, ASXL1 | BMI | High BMI was protective against DNMT3A clonal expansion, but associated with TET2 and ASXL1 CHIP in this cross-sectional study | human n = 628,388 | Kessler et al., 2022(Kessler et al., 2022) | |
| DNMT3A | Fatty bone marrow model | Self-renewal and engraftment advantage was observed in DNMT3A mutant cells and was mediated by IL-6 | mouse | Zioni et al., 2022(Zioni et al., 2021) | |
| CHIP | Unhealthy diet, low LDL, Low HDL, insulin resistance, and hypertension are associated with increased risk of CHIP. | Several cross sectional or longitudinal association studies link CHIP with metabolic syndrome. | human n = 44,111 | Bhattacharya et al., 2021(Bhattacharya et al., 2021), Kar et al., 2022(Kar et al., 2022), van Deuren et al., 2022 (van Deuren et al., 2021) | |
|
| |||||
| Other | |||||
|
| |||||
| CHIP | Down syndrome | Observational study found early signs of CHIP in Down’s syndrome patients | human n=51 | Liggett et al., 2021 (Liggett et al., 2021) | |
| DNTM3A, PPM1D | Growth factor usage, RBC transfusion | Cross sectional study-linked association | human n=5,649 | Coombs et al., 2017 (Coombs et al., 2017) | |
| TET2 | Atherosclerosis | Atherosclerosis accelerates HSC proliferation and expansion of TET2−/− cells | Mathematical modeling, mouse | Heyde et al., 2021 (Heyde et al., 2021) | |
DDR genes
Mutations in DDR genes (TP53, PPM1D, and CHEK2) have the strongest association with prior cancer treatment, and clones with mutations in these genes expand more under the stress of specific cytotoxic therapies.(Bolton et al., 2020, Coombs et al., 2017, Swisher et al., 2016, Wong et al., 2015) Mutations in DDR genes are infrequent in the absence of prior cytotoxic exposure,(Xie et al., 2014, Genovese et al., 2014, Jaiswal et al., 2014, Eskelund et al., 2020) but prevalent in patients with TMN, indicating the selective advantage under cytotoxic stress.(Bolton et al., 2020, Gibson et al., 2017b, Hsu et al., 2018, Lindsley et al., 2017)
Studies have shown that PPM1D mutations often occur in exon 6 and confer resistance to chemotherapeutics, especially platinum-based therapies.(Bolton et al., 2020, Hsu et al., 2018, Kahn et al., 2018, Kim et al., 2019) PPM1D mutations are typically gain of function truncation mutations that stabilize the protein and suppress the DDR, enabling mutant clones to resist apoptosis.(Hsu et al., 2018, Kahn et al., 2018) Similar to the selective advantages observed in large patient cohorts,(Bolton et al., 2020) in vitro studies show that PPM1D mutants are resistant to specific cytotoxic therapies (cisplatin, etoposide, doxorubicin, cytarabine) but not others (5-fluorouracil (5-FU), vincristine).(Hsu et al., 2018, Kahn et al., 2018) These differences are likely explained by these treatments’ differing mechanisms of action. For instance, vincristine impairs mitosis by microtubule interference and inhibits trafficking of DNA repair proteins, including P53, which interacts with PPM1D.(Kahn et al., 2018, Poruchynsky et al., 2015) It is plausible that microtubule interference blocks PPM1D from interacting with downstream DDR mediators, thereby preventing its ability to inhibit apoptosis.(Kahn et al., 2018, Poruchynsky et al., 2015)
Mutations in TP53 are a strong risk factor in the development of TMN.(Abelson et al., 2018, Desai et al., 2018, Gillis et al., 2017, Wong et al., 2015) P53 has multiple functions and may regulate HSC response to cytotoxic stress by regulating apoptosis,(Wu et al., 2005) proliferation,(Bondar and Medzhitov, 2010, Wlodarski et al., 1998, Wu et al., 2005) quiescence and self-renewal.(Chen et al., 2008, Liu et al., 2009) Cytotoxic drivers of TP53 mutant expansion include platinum, radiation, and taxane-based therapies.(Bolton et al., 2020) Furthermore, in vitro studies suggest that 5-FU(Wlodarski et al., 1998) and doxorubicin(Sano et al., 2021) confer a selective advantage for TP53 mutant clones. The expansion of TP53 mutant clones may be dependent on mutation type and may occur via multiple mechanisms, as TP53 mutations have been suggested to cause both gain- and loss-of-function. Interestingly, TP53 mutation has been reported to promote DNA H3K27 trimethylation via enhanced interaction with EZH2, thereby reinforcing HSC-self renewal during radiation stress. (Chen et al., 2019) Whether this mechanism underlies selective CHIP expansion of TP53 mutants during other cytotoxic stresses remains unknown, though it is likely that both DDR impairment and epigenetic changes underlie the selective advantages of this mutant.
Epigenetic modifiers
The role of cytotoxic stress on clones bearing mutations in epigenetic modifiers is less clear. A few small studies have shown clonal expansion in non-leukemic HSC clones harboring mutations in the epigenetic modifiers DNMT3A and TET2.(Wong et al., 2016) Coombs et al showed cytotoxic therapy was associated with TET2,(Coombs et al., 2017) but this was not recapitulated in later large cohort studies.(Bolton et al., 2020) Conversely, mutations in DNMT3A and TET2 are known to confer treatment resistance to chemotherapies in AML, and numerous studies have linked specific DNMT3Amut in AML with poor clinical outcomes.(Lauber et al., 2020, Ley et al., 2010, Ribeiro et al., 2012, Yuan et al., 2016a) For example, DNMT3AR882mut protects against anthracycline-mediated cell death in AML via an impaired chromatin remodeling response that prevents an appropriate DDR.(Guryanova et al., 2016) Restoration of this pathway sensitized DNMT3AR882mut cells to anthracycline.(Guryanova et al., 2016) Conversely, administration of azacytidine, a DNA methyltransferase inhibitor, suppressed Dnmt3AR882mut clones and prolonged survival in AML patients harboring DNMT3AR882mut. (Scheller et al., 2021) In mouse models, DNMT3Anull cells expand following exposure to busulfan, but not 5-FU, highlighting the specificity of certain stressors to drive clonal expansion.(Chen et al., 2020) The effects of chemotherapies on DNMT3Amut clonal selection may also be specific to the type of mutation.(Yuan et al., 2016b)
For TET2, one proposed mechanism driving treatment resistance is that hypermethylation caused by TET2 deletion promotes a quiescent stem-cell like state, desensitizing cells to chemotherapies such as cytosine arabinoside and doxorubicin.(Morinishi et al., 2020) The use of hypomethylating agents to target specific TET2mut may prove beneficial in sensitizing TET2mut cells to chemotherapy.(Stölzel et al., 2021)
The impact of mutations in IDH1/2 and ASXL1 on chemoresistance in AML remain unclear.(Brunner et al., 2019, Molenaar et al., 2018, Ni et al., 2020, Paschka et al., 2010, Paschka et al., 2015) Determining the mechanisms by which chemotherapy interacts with DNMT3A and other epigenetic modifier mutant clones will provide greater insights on how these mutant clones respond to cytotoxic pressures. (Chaudry and Chevassut, 2017, Metzeler et al., 2012, Venugopal et al., 2021, Yang et al., 2015, Yang et al., 2021)
Spliceosome complex proteins
Clones with mutations in spliceosome proteins (SRSF2 SF3B1, and U2AF1) are more likely to persist in AML patients following chemotherapy and are observed in a high proportion of secondary AMLs.(Lachowiez et al., 2021) Still, whether spliceosome mutants have selective advantages with certain chemotherapeutics or whether selection is a consequence of other genetic lesions within the leukemic clones remains unknown.
Finally, the clinical correlation between the selective advantage of non-leukemic epigenetic and spliceosome driver mutations in CHIP under cytotoxic therapy may be limited by the type of cytotoxic therapies used in large cancer patient cohort studies.(Bolton et al., 2020, Coombs et al., 2017) While prior studies in leukemic patients suggest epigenetic modifiers and spliceosome regulators play important roles in response to therapy, how various cytotoxic therapies regulate non-leukemic CHIP clones remains unclear. Further clinical and mechanistic studies are needed to determine whether cytotoxic drivers negatively or positively impact these classes of CHIP mutants. Expanding our understanding of these selective processes could revolutionize personalized medicine, such that treatment regimens are carefully chosen to avoid the selection of certain mutant clones.
Immunotherapies
Current cancer research has focused on the development of immunotherapies to prime, augment, or sustain immune responses to cancer. These therapies pose a greater risk of immune-mediated stress compared to the cytotoxic therapies described above.(Esfahani et al., 2020) Immune checkpoint inhibitors (ICIs) may induce autoimmunity, including immune-mediated destruction of the hematological compartment,(Davis et al., 2019) while chimeric antigen receptor-T cell (CAR T) therapy is known to induce cytokine release syndrome mediated largely by interleukin 1 (IL-1) and IL-6.(Norelli et al., 2018) To date, the impact of immunotherapies on CHIP remains controversial, with some studies indicating that immunotherapies may promote expansion of CHIP clones but others reporting no impact on clonal expansion. (Bolton et al., 2020, Miller et al., 2020)
While there is no consensus about the impact of immunotherapy on CHIP emergence, CHIP mutations are known to affect CAR T cell function. A case study found CAR T cells harboring a TET2 CHIP mutation had enhanced expansion, prolonged activation, increased cytokine production, and memory-like properties,(Fraietta et al., 2018) consistent with a known role for DNA methylation in promoting memory-like enhanced and sustained T cell function;(Akbari et al., 2021, Ladle et al., 2016, Wang et al., 2021) similar results were observed in a pre-clinical mouse model.(Jain et al., 2021) Deletion of DNMT3A in CAR T cells also promotes cell survival, limits exhaustion, and enhances therapeutic efficacy.(Prinzing et al., 2021) One study suggested that CHIP mutations may influence CAR T therapy as patients with CHIP had higher rate of complete response to CAR T therapy, but with increased rates of cytokine release syndrome.(Miller et al., 2021b) Aside from CHIP mutations in CAR T cells themselves, tumor-infiltrating leukocytes harboring CHIP mutations have also been observed in solid tumor samples (Kleppe et al., 2015). Thus, CHIP could impact the efficacy of immunotherapy via non-cell autonomous mechanisms that have not yet been explored.
Smoking
Cigarette smoking induces cytotoxic DNA damage and causes substantial tissue damage and inflammation. Smoking damages the HSC niche and subsequently impairs HSC homing, causes proliferative exhaustion of HSCs, and induces extramedullary hematopoiesis.(Morales-Mantilla et al., 2020, Siggins et al., 2014, Tura-Ceide et al., 2017) Increased risk of CHIP has been linked with a history of smoking,(Bolton et al., 2020, Genovese et al., 2014, Coombs et al., 2017, Kar et al., 2022) as well as smoking-associated pulmonary diseases such as COPD (Buscarlet et al., 2017, Zink et al., 2017) and lung cancer.(Zajkowicz et al., 2015, Coombs et al., 2017, Kessler et al., 2022) Among common CHIP mutations, mutations in ASXL1, have the most significant association with smoking status.(Bolton et al., 2020, Coombs et al., 2017, Kar et al., 2022, Kessler et al., 2022, Dawoud et al., 2020) Mutant ASXL1 regulates DNA transcription by enhancing the ubiquitinase BAP1, resulting in decreased histone ubiquitination and increased AKT/mTor signaling. This results in increased cell cycle progression even in the setting of DNA damage.(Fujino and Kitamura, 2020) Likely, smoking not only contributes to increased mutational burden of ASXL1 mutant clones but also provides a selective advantage for this clone via further induction of DNA and tissue damage and inflammation; further mechanistic studies of the relationship between smoking and ASXL1 mutations are still pending.
Inflammation is a common source of stress that induces the regulation and repair of physiological insults. Inflammation can be triggered by many sources, including old age, infections, cancer, radiation, and underlying pathogenic conditions like atherosclerosis. (Bick et al., 2020b, Gartung et al., 2019, Hansson et al., 2006, Matatall et al., 2016, Pietras, 2017) Inflammation is mediated by various signals, such as pathogen- and damage-associated molecular patterns (PAMPs and DAMPs), as well as pro-inflammatory cytokines including tumor necrosis factor alpha (TNFα), interferon alpha (IFNα), interferon gamma (IFNγ), IL-1β, and IL-6. As reviewed elsewhere, these signals influence HSCs via both direct and indirect mechanisms that impact quiescence, promote differentiation at the expense of self-renewal, and affect cell survival.(King and Goodell, 2011, Baldridge et al., 2010, Essers et al., 2009, Caiado et al., 2021, Pietras, 2017, Hormaechea-Agulla et al., 2021) Long-term consequences of chronic inflammation include impaired HSC self-renewal and subsequent HSC terminal exhaustion.(Matatall et al., 2016)
Several studies in both humans and animal models demonstrate that selected CHIP clones can resist inflammation-induced proliferative stress and gain a selective advantage in inflammatory conditions. Studies in patients with HIV(Bick et al., 2020b, Dharan et al., 2021) and aplastic anemia(Ogawa, 2016, Zhang et al., 2019, Yoshizato et al., 2015) indicate that inflammation can lead to the selection of specific CHIP clones, and can predispose to progression to AML and myelodysplastic syndrome (MDS).(Kristinsson et al., 2011)
Resistance to Inflammation-Mediated Depletion
Loss-of-function TET2 clones have been extensively studied in murine models and these TET2 mutants selectively expand, particularly in the myeloid compartment, in response to pro-inflammatory stimuli TNFα and lipopolysaccharide.(Abegunde et al., 2018, Cai et al., 2018) In a key study by Meisel et al., the presence of Tet2 null hematopoietic cells disrupted intestinal barrier integrity and increased systemic microbial signaling, particularly from Lactobacillus, which was critical in promoting further clonal expansion.(Meisel et al., 2018) Conversely, broad-spectrum antibiotic therapy slowed the expansion of Tet2-deficient clones.(Zeng et al., 2019) Mechanistically, the long noncoding RNA (IncRNA) Morrbid and the TLR4/NF-κB/IL-6/Stat3 pathway may be crucial for Tet2 clonal expansion and persistence.(Cai et al., 2018) Pharmacological inhibition of Shp2 with inhibitor SHP099 or inhibition of Stat3 with E3330 led to a loss of Morrbid hyperactivity in Tet2-deficient hematopoietic stem and progenitor cells (HSPCs), suppressing selective clonal expansion.(Cai et al., 2018) This study highlights the role of IncRNAs in modifying cell-death to inflammation by making them more resistant to inflammation-induced apoptosis.
While DNMT3A mutations are not as extensively studied as TET2, our recent findings shed light on mechanisms by which inflammation can positively select for Dnmt3a KO clones. We found that Dnmt3a KO murine clones outcompete wildtype murine cells in response to chronic IFNγ stimulation caused by Mycobacterium avium infection.(Hormaechea-Agulla et al., 2021) Much like TET2 clones, chronically IFNγ-stimulated Dnmt3a null HSCs are resistant to apoptosis, as caspase 3/7 activity is significantly decreased compared to IFNγ-stimulated wildtype HSCs. Most strikingly, Dnmt3A clones showed dampened differentiation in response to IFNγ compared to wildtype cells. Since we previously showed that excessive IFNγ-stimulated myeloid differentiation is a major mechanism of HSC depletion during chronic infection,(Matatall et al., 2016) reduced IFNγ-dependent differentiation provides a clear selective advantage to Dnmt3a KO clones. Indeed, Dnmt3a KO clones do not upregulate pro-differentiation genes, nor do they display significant myeloid differentiation upon IFNγ stimulation in vitro. Consistent with this phenotype, the promoter regions of pro-differentiation genes were hypermethylated in Dnmt3a KO HSCs from infected mice compared to WT.(Hormaechea-Agulla et al., 2021) Further, a recent study showed that IFNγ-dependent DNMT3Amut clonal expansion was attributable to DNA hypomethylation and altered expression of Txnip and p21, allowing preservation of HSC quiescence.(Zhang et al., 2022) These findings demonstrate that epigenetic changes underlie transcriptional activation of inflammation-driven HSC differentiation and provide a clear mechanism by which Dnmt3a KO clones are selected to expand in inflammatory conditions.(Hormaechea-Agulla et al., 2021, Zhang et al., 2022)
While these studies provide a potential mechanism by which CHIP mutants, such as DNMT3A, expand in the setting of inflammatory stress, some parallel studies of humans and human cells have not mirrored exactly these findings. For instance, the lineage output of naive Dnmt3a KO HSCs in our murine model differed from what has been observed in human DNMT3A R882 mutated HSPCs.(Nam et al., 2022) Single-cell multi-omics of DNMT3A R882 CD34+ HSC populations in multiple myeloma patients revealed a myeloid biased differentiation and megakaryocytic expansion within humans, suggesting that broad transcriptional consequences of DNMT3A mutations affect differentiation even in the absence of a strong exogenous source of inflammation.(Nam et al., 2022) Altered methylation of human DNMT3A R882 mutant clones showed aberrant methylation status, but the differentially methylated regions included MYC as opposed to the ATF transcription factors that we highlighted. Thus, while murine models can shed light on the mechanisms by which DNMT3A mutant clones are positively selected during inflammation, mechanistic studies in humans must be done to confirm relevance.
Depletion of wild-type competitors
Other CHIP clones encode transcriptional alteration(s) that promote positive selection and suppression of competing wild type (WT) clones.(Higa et al., 2021) Our recent study showed that Cebpa KO MPP3 clones expand in response to chronic IL-1ß signaling. Transcriptionally, Cebpa deficiency prevents IL-ß-driven repression of self-renewal genes like Foxo3 and Mycn, which are directly regulated by Cebpa, thereby giving mutant clones a self-renewal advantage over WT cells. Interestingly, Cebpa KO clones can also suppress WT competitors by promoting excessive myeloid differentiation in a competitive transplant model. Indeed, RNA-seq analysis demonstrated that WT MPP3s exposed to chronic IL-1ß and in contact with Cebpa KO clones showed a significant increase in myeloid differentiation genes compared to WT MPP3s stimulated by chronic IL-1ß alone.(Higa et al., 2021)
Avagyan et al 2021 expanded upon the ability of mutant clones to promote differentiation of their competitors. Utilizing a novel technique called Tissue editing With Inducible Stem cell Tagging via Recombination (TWISTR) in a zebrafish model, Avagyan et al created a mosaic zebrafish that expressed 12 mutated CH genes during development. Interestingly, clones bearing frameshift mutations in asxl1, the zebrafish homolog of human ASXL1, demonstrated clonal dominance. Single-cell RNA sequencing of marrow cells showed asxl1 mutant macrophage and neutrophil clones were highly inflammatory, with upregulation of inflammatory cytokines il1β and tnfβ. More strikingly, the asxl1 mutant progenitors showed an inflammation-resistant molecular profile, upregulating genes that modulate the effects of inflammatory stress such as socs3a and nr4a1. These findings suggest that CHIP clones contribute to the generation of inflammatory cytokines known to deplete WT HSCs. At the same time, the clonal mutant progenitor cell types resist the deleterious effects of these inflammatory signals and expand.(Avagyan et al., 2021) It is interesting to note that some CHIP mutations (e.g. in MYD88, STAT3) play a key role in transmitting inflammatory signaling, suggesting a direct route for inflammation resistance. These findings highlight how mutant HSCs can promote their own selection by preserving self-renewal while WT competitors are outcompeted or exhausted through excessive myeloid differentiation.
CHIP as a cause and consequence of inflammation
While CHIP mutants have survival and selective advantages in response to inflammation, CHIP itself also drives inflammation, setting up a feed-forward loop, as noted in the above example of asxl1-mutant zebrafish and reported in association with other mutations. For instance, PPM1D and TET2 mutant clones have been shown to produce immune cell progeny that generate significantly higher than normal levels of the pro-inflammatory cytokine IL-6 upon exposure to chemotherapy or lipopolysaccharide, respectively.(Cai et al., 2018, Jaiswal et al., 2017, Yura et al., 2021) TET2-mutant clones contribute to atherosclerosis and heart disease via increased production of IL-6 by downstream macrophages, and DNMT3A has recently been suggested to contribute to inflammation in the heart by the same mechanism. (Abplanalp et al., 2021, Bick et al., 2020a) As previously noted, TET2 mutations may also contribute to systemic inflammation by causing intestinal barrier compromise. As genetic mosaicism is now recognized to occur widely across tissues,(Hsieh et al., 2020) additional proinflammatory consequences of CHIP mutations are likely to arise.
Human studies support the notion that CHIP can contribute to inflammation-mediated diseases such as severe COVID-19, sepsis, and cardiovascular disease.(Bolton et al., 2021, Dawoud et al., 2020, Jaiswal and Ebert, 2019, Kessler et al., 2022, Zekavat et al., 2021b) While increased IL-6 has been linked to heart disease in animal models and blockade of IL-6 signaling can reduce cardiovascular risk,(Kessler et al., 2022) no empirical evidence has been published suggesting that IL-6 itself is capable of promoting CHIP. Using the presence of the IL6R p.Asp358Ala coding mutation as a proxy to assess the potential for IL-6R blockade in individuals with CHIP, Bick et al., 2020 found that impaired IL-6R signaling reduced cardiovascular disease risk but did not impact CHIP status, particularly DNMT3A and TET2 CHIP. Furthermore, transcriptomic studies have not shown upregulation of canonical inflammatory pathways in CHIP, and the role of IL-6 signaling in CHIP-associated CVD risk remains an open question.(Kessler et al., 2022, Weinstock et al., 2021) Thus, while mechanistic studies in murine models highlight ways that inflammation drives CHIP, the role of inflammation as a driver of CHIP in humans requires further experimental validation. Such a role may be even more complex to identify in humans due to the impact of cell-intrinsic genetic modifiers such as TCL1A on the selective expansion of CHIP clones. (Weinstock et al., 2021)
The extent to which inflammation suppresses competitor cells or changes the niche to allow for favorable expansion of CHIP clones has yet to be fully explored.(Rodriguez-Meira et al., 2022) While we have shown that CHIP clones resist the differentiation and exhaustion effects of inflammation, future studies could highlight more active processes by which CHIP clones suppress their competitors.(Rodriguez-Meira et al., 2022)
Autoimmunity
Autoimmune disease mediated in part by dysregulated cytokine production, including IL-1, IFNγ, and TNFα,(Italiani et al., 2018, Muzes et al., 2012, Lubberts and van den Berg, 2003) generates a chronic inflammatory environment that could facilitate the expansion of selected CHIP clones. Autoimmune diseases like inflammatory bowel disease, systemic lupus erythematosus, and rheumatoid arthritis are well known to carry increased risk of myeloid malignancies.(Bekele and Patnaik, 2020, Boddu and Zeidan, 2019, Ramadan et al., 2012) The risk of hematological malignancy in these diseases has often been attributed to the use of therapies such as cyclophosphamide, methotrexate, and TNFα blockade.(Ramadan et al., 2012) However, increased risk of leukemia has been observed in patients with arthritis with no prior history of treatment, suggesting inflammation itself may have a role in hematological malignancy risk.(Boddu and Zeidan, 2019) One small study found an increased prevalence of CHIP mutations in arthritis patients, with higher VAFs correlated with more severe disease, although this was not replicated in a separate study.(Tariq et al., 2020, Savola et al., 2018) Another small cohort study of inflammatory bowel disease patients found increased VAF for DNMT3A and PPM1D compared to prior cohort studies, which was independent of treatment history. Additionally, serum levels of IFNγ were elevated in patients with DNMT3Amut CH,(Zhang et al., 2019) further implicating this inflammatory cytokine in the selection of DNMT3Amut clones. Indeed, altered epigenetic profiles are known to facilitate immune dysfunction in autoimmune diseases.(Mazzone et al., 2019) Altogether, mutations in epigenetic modifiers could provide a selective advantage for CHIP expansion and subsequent autoimmune disease progression. It is important to note that most of these studies on autoimmune diseases are in small cohorts that are underpowered. Further mechanistic studies are warranted to elucidate the role of autoimmunity and its treatment in CHIP expansion.
Hematopoietic Stem Cell Transplants
HSCT is a last-resort treatment for hematological diseases like AML, MDS, aplastic anemia, and SCD. However, the stress of HSCT promotes loss of HSC proliferative and regenerative capabilities(Flach et al., 2014, Harrison and Astle, 1982) through telomere shortening(Colla et al., 2015, Notaro et al., 1997) and epigenetic changes,(Soraas et al., 2019) decreasing overall HSC fitness. Furthermore, preconditioning for bone marrow transplant via irradiation and cytotoxic regimens can damage the niche(Pinho and Frenette, 2019) and generate an inflammatory environment, further impairing HSPC fitness. Several studies have demonstrated that some CHIP clones have a selective repopulation advantage during transplantation stress.
Both murine and human studies have shown that TET2-deficient HSCs have increased self-renewal and myeloid-biased repopulation expansion after transplantation.(Kunimoto and Nakajima, 2020, Moran-Crusio et al., 2011, Ortmann et al., 2019, Ostrander et al., 2020, Shide et al., 2012) A study by Abegunde et al. showed that TET2 mutant clones avoid HSC suppression by inhibiting TNFα signaling.(Abegunde et al., 2018) However, contradictory studies suggest that TET2null HSCs unexpectantly exhaust at the same rate as control HSCs in serial transplantation.(Ostrander et al., 2020) Furthermore, Yamashita et. al. found that TNFα plays a key role in facilitating HSC survival and myeloid differentiation;(Yamashita and Passegue, 2019) therefore, if TET2 loss of function inhibits TNFα signaling one might expect impaired HSC survival. Overall, the precise mechanism of TET2-mutant clonal expansion after transplantation remains poorly defined and may involve pathways other than TNFα.
Murine studies have shown that Dnmt3a mutant clones have virtually limitless self-renewal and competitive repopulating capacities,(Challen et al., 2011, Kunimoto and Nakajima, 2020) allowing them to outcompete their normal counterparts over time, at least in the context of stem cell transplantation after irradiation preconditioning. Indeed, DNMT3A loss-of-function HSCs can regenerate over at least 12 transplant generations despite the stress of serial transplantations.(Jeong et al., 2018) Allsopp et. al. suggested that DNMT3Anull embryonic stem cells have elongated telomeres that can extend the replicative lifespan of serially transplanted HSCs.(Allsopp et al., 2003, Gonzalo et al., 2006) Resistance to inflammation-mediated decay may also contribute to persistence of Dnmt3a-mutant clones after transplantation, a topic of ongoing exploration.
In contrast to TET2 and DNMT3A, ASXL1, SRSF2, SF3B1, U2AF1, and PPM1D are unable to outcompete wildtype HSPCs in murine transplant settings.(Fujino et al., 2021, Yoshida et al., 2011, Hsu et al., 2018, Kon et al., 2018, Mupo et al., 2017) PPM1D mutant clones lack proliferative advantage and have overall decreased fitness after HSC transplantation, as shown in a series of patients after autologous transplant.(Hsu et al., 2018, Wong et al., 2018) These studies suggest that stabilization of the DDR and resistance to apoptosis conferred by PPM1D mutations confer little to no benefit in the context of the proliferative stress of HSCT.(Hsu et al., 2018, Kahn et al., 2018, Wong et al., 2018)
While gain-of-function mutations in PPM1D were not selected following HSCT, TP53 mutant HSC clones have a competitive expansion advantage under transplant stress.(Chen and Liu, 2019, Chen et al., 2019) Chen et al. demonstrated that a gain-of-function mutation allows TP53 to have enhanced interaction with EZH2, reinforcing H3K27 trimethylation and thereby enhancing the mutant clone’s self-renewal and differentiation capacity.(Chen and Liu, 2019, Chen et al., 2019) Interestingly, TP53 HSC mutant expansion also occurs in a non-irradiated transplant model in absence and, to a greater extent, in the presence of doxorubicin chemotherapy.(Sano et al., 2021) Thus, both mechanisms of altered epigenetics and DDR pathway dysfunction may facilitate selective advantage of TP53 mutant clones due to both transplantation and chemotherapy stress.
CHIP has been observed in patients following autologous (Chitre et al., 2018, Gibson et al., 2017b) and allogeneic stem cell transplants (Frick et al., 2019) and may affect stem cell repopulation and engraftment. Accumulating evidence shows that HSCs harboring CHIP mutations can be found in the donor and can be transplanted to the recipient (Gibson et al., 2017a, Nawas et al., 2021). Recipients who received donor grafts with CHIP mutations experience a more biased myeloid lineage expansion, increasing the risk of the development of adverse hematological outcomes, including therapy-related myeloid neoplasms and cytopenias.(Chitre et al., 2018, Gibson et al., 2017a, McNerney and Le Beau, 2018) Furthermore, Pierola, et al. showed in an in vivo study that CHIP mutant HSC clones lacking DNA damage response systems can survive chemotherapy-induced stress and DNA damage while acquiring secondary mutations, ultimately leading to secondary malignancies. (Alfonso Pierola et al., 2016) Altogether, these discoveries are leading to increased scrutiny of the mutational landscape in HSCT donors.
Aging
Aging is the stressor that is most strongly associated with CH.(Bolton et al., 2020, Genovese et al., 2014, Jaiswal et al., 2014, Xie et al., 2014) Sequencing studies in the elderly reveal that clonal hematopoiesis is most prevalent in those aged >70. While many CH clones can emerge, the most prevalent age-associated clones are DNMT3A, TET2, ASXL1, SF3B1, and SRSF2.(Buscarlet et al., 2017, Genovese et al., 2014, McKerrell et al., 2015, Jaiswal et al., 2014, Xie et al., 2014)
While aging can lead to the expansion of various CHIP clones, it is noteworthy that the expansion of these clones is not directly correlated with CHIP-associated clinical outcomes. A study by Zeventer et al., 2021 investigated CHIP in individuals over 80 years old and found no association between cardiovascular morbidity and the presence of CHIP. There were, however, associations between CHIP and deaths due to hematological malignancies (hazard ratio 1.48, CI 95%).(van Zeventer et al., 2021) Of note, the most prevalent clones in this study were DNMT3A and TET2 mutants, both of which are associated with cardiovascular diseases.(Jaiswal et al., 2017) These opposing findings indicate that aging can lead to the expansion of clones but that the major impacts on health occur before a threshold age, suggesting that the age CHIP onset may be the most relevant factor.
Mechanistically, aging can be considered a multi-factorial stressor that leads to various effects on the hematopoietic compartment, as explored in several reviews.(Jaiswal and Ebert, 2019, Lee et al., 2019, Libby et al., 2019) The mechanisms underlying age-associated clonal expansion likely overlap with inflammatory stimuli discussed above. Indeed, there is a baseline increase in production of pro-inflammatory cytokines such IFNγ, TNFα, and IL-6(Chung et al., 2006, Li et al., 2011, Sanada et al., 2018) over time, frequently called “inflammaging,” which could promote the expansion of DNMT3A(Hormaechea-Agulla et al., 2021) and TET2(Abegunde et al., 2018, Meisel et al., 2018) CHIP clones. In addition, aged HSCs have increased histone and DNA methylation, resulting in activation of self-renewal signatures and silencing of differentiation programs.(Sun et al., 2014, Nachun et al., 2021, Beerman, 2017) As a result, aged HSCs display defective homing to the bone marrow niche, impaired lymphocytic differentiation, and increased myeloid and platelet lineage priming.(Franceschi and Campisi, 2014, Kovtonyuk et al., 2016, Lee et al., 2019) This overproduction of myeloid cells may contribute to another feed forward loop of increased inflammation and CHIP during aging.(Franceschi et al., 2000)
Interestingly, Kar et al., 2022 recently reported that longer leukocyte telomere length was associated with increased DNMT3A- and TET2-mutant CHIP.(Kar et al., 2022) Long telomeres may be yet another mechanism by which these clones persist despite inflammation and replicative stress, but this proposed mechanism has not yet been formally demonstrated.
A study by Fujino et al., 2021 showed that C-terminally truncated ASXL1 clones expanded in aged mice. (Fujino et al., 2021) Mechanistically, truncated Asxl1 interacts with BAP1 to deubiquitinate Akt and activate mTORC, leading to the survival and expansion of these aged Asxl1 HSCs. However, Asxl1 clones exhibited impaired engraftment, myeloid-biased hematopoiesis, and significantly decreased homing ability to the bone marrow niche compared to wildtype transplanted HSCs, consistent with the observation that ASXL1 clones do not survive the stresses of HSCT.(Fujino et al., 2021)
Other investigators have used modeling to argue that a constant fitness advantage conferred by the CHIP mutation is sufficient to model the CHIP seen in humans.(Fabre et al., 2021, Watson et al., 2020) In fact, Fabre et al conclude that “many clones grew more rapidly early in life compared with the rate in old age”.(Fabre et al., 2021) In contrast, Monte Carlo modeling has been used to show that cancer-causing mutations have greater positive selection late in life, thus explaining the age-dependence of leukemias and other cancers.(Rozhok and DeGregori, 2019) Additional studies, such as using mouse and non-human primate models, will be required to determine whether CHIP occurs simply as a matter of time or if extrinsic pressures are required.
Metabolic Stress
Metabolism plays a crucial role in maintaining and regulating HSC homeostasis,(Ito et al., 2019)) and there has been increasing interest in uncovering how metabolic factors impact CHIP. In particular, TET2 mutant clones have been known to interact with endocrine and metabolic axes, although the reports of these associations have been mixed. In a type II diabetes mouse model, a TET2 mutant clone was shown to facilitate insulin resistance in mice.(Fuster et al., 2020) In addition, elevated glucose levels have been found to destabilize TET2 in human peripheral blood samples,(Wu et al., 2018) and increased glucose levels drove leukemia formation in mice harboring TET2-mutations.(Cai et al., 2021) Two recent large cohort studies found a higher prevalence of CHIP in people with an unhealthy diet(Bhattacharya et al., 2021) and in postmenopausal women with a high BMI and poor diet quality.(Haring et al., 2021) A longitudinal study of 20 patients showed that clonal expansion was associated with lower high-density-lipoprotein (HDL) cholesterol and insulin resistance but not with BMI, hypertension, hyperglycemia or total cholesterol.(van Deuren et al., 2021) However, UK BioBank data revealed hypertension, low total cholesterol, and low LDL to be associated with CHIP but not obesity or diabetes.(Kar et al., 2022) A separate analysis of UK BioBank data showed that elevated BMI was negatively associated with DNMT3A mutation, but positively associated with TET2 and ASXL1 mutant clones.(Kessler et al., 2022) One confounder that may explain these mixed results is that vitamin C has been shown to regulate HSC homeostasis by activating TET2.(Lee et al., 2020) Lower plasma vitamin C concentrations was associated with a higher frequency of TET2 mutations in an elderly cohort.(Chen et al., 2021) For DNMT3A, murine models showed that a fatty-bone marrow environment can provide a selective advantage for DNMT3A-mut cells which may be mediated by IL-6.(Zioni et al., 2021) Altogether, early studies suggest an important role for metabolic and dietary stresses in the pathogenesis of CHIP.
Other Stresses That Expand CHIP Clones
While we focused in this review on specific DNA-damaging and inflammatory environmental drivers of clonal expansion, CHIP has also been linked with various other potential drivers including psychiatric diseases,(Zink et al., 2017) gout,(Kessler et al., 2022) and Down syndrome.(Liggett et al., 2021, Tong et al.). In individuals with Down syndrome, clonal hematopoiesis is observed much earlier in life with the most common mutations being found in TET2; although notably this CHIP did not reach levels observed in the elderly.(Liggett et al., 2021) Trisomy 21 has been shown to induce an enhanced IFN signature in the hematopoietic compartment, which may facilitate early clonal expansion,(Sullivan et al., 2016) and CHIP in individuals with Down Syndrome is associated with dysregulated immune patterns in peripheral leukocytes.(Liggett et al., 2021) Selective clonal expansion in other diseases like SCD is less clear.(Pincez et al., 2021, Liggett et al., 2022) A recent report suggests that space flight may accelerate clonal expansion,(Mencia-Trinchant et al., 2021) and many more associations and drivers of CHIP are constantly being discovered.
Concluding Remarks
The natural acquisition of mutations throughout life generates a range of genetically distinct HSPCs. Despite the ubiquity of CHIP-associated mutations, most individuals never develop CHIP, suggesting that environmental pressures are critical determinants of clonal expansion. (King et al., 2020) Here we reviewed the contexts in which some genetically variant clones expand over time. We highlight specific examples of imposed external stress such as platinum-based chemotherapies that drive the expansion of clones containing PPM1D mutations, which in turn increase the risk of secondary malignancy. As the field increasingly identifies environmental factors that may provide a selective advantage for certain clones, the clinical implications of these associations and whether they can be modulated to improve human health continue to be explored.
Among genes known to be associated with CHIP, DDR factors such as P53 and PPM1D have been associated with chemotherapies and radiation. Mutations affecting epigenetic modifiers — DNMT3A, TET2, ASXL1 — and JAK2 have been associated with bacterial translocation, infections, and inflammatory conditions, and one would expect that MYD88 and STAT3 mutant clones are similarly selected in the setting of inflammation. Relatively little is known about the environmental conditions that drive expansion of clones containing mutations in splicing factors such as SF3B1, SRSF2, and U2AF1, which are among the top 10 genes mutated in CHIP. These mutations have been reported to drive inflammatory responses, but further work will be required to determine if they follow the pattern of inflammation-driven selection that has been reported for TET2, DNMT3A, ASXL1, and JAK2.
Whereas some variant clones may contribute to heart disease or stroke, others may be neutral in their impact on human health or may be beneficial by enabling long-term hematopoiesis in the elderly. Indeed, despite our tendency towards binary thinking, mutant clones may live in the liminal space between pathogenic and beneficial. For instance, whereas TET2 clones drive increased IL-6-mediated inflammation, they may also enable persistent hematopoiesis in the challenging conditions of advanced age or serve to suppress competing clones with more serious adverse health effects, essentially constituting non-malignant ‘fitness peaks’. While mutant VAF >10% is strongly associated with increased heme malignancy risk, what other factor(s) govern progression to pre-malignant states like clonal cytopenia of undetermined significance (CCUS) and/or leukemia in this setting remain to be clearly identified. For some mutations, downstream consequences of mutations on differentiated cell function may have an outsized effect on health. Further, most mechanistic studies to elucidate environmental drivers of CHIP have been done in mice. While informative, these models have limitations, the most obvious of which is the short lifespan of the mouse relative to humans and the clean (if not sterile) and non-challenging housing conditions. Paired epidemiologic association studies have been a tremendous strength in this respect, but utilization of clinical trials and non-human primate studies will also be important.
While a range of studies highlights how the environment (e.g., obesity, infection, prior exposure to chemotherapy) significantly alters which clones are selected to survive and expand, the extent to which changes in environment can shape clonal selection or modify disease risk remains to be seen. Lessons from the CANTOS trial examining IL-1ß inhibition to reduce cancer risk are a case in point: while greatly reducing lung cancer risk and potentially reducing major adverse cardiovascular events in patients with TET2-mutant CHIP, canakimumab leaves patients vulnerable to potentially deadly infections. (Ridker et al., 2017, Svensson et al., 2022) Outcomes from such trials indicate that endpoints for intervention must be carefully defined. Furthermore, basal inflammatory signaling, such as is mediated by the microbiome,(Josefsdottir et al., 2017) is essential for normal hematopoiesis and must be taken into account when considering anti-inflammatory approaches. Perhaps it is more productive to address the aberrant immunometabolic features of immune cells derived from mutant clones than to reduce the size of the clone itself. Similarly, one can consider more nuanced methods (beyond brute inhibition of inflammation) to modulate the bone marrow microenvironment to reduce the selective advantage of certain CHIP clones. Finally, while an intervention may prevent the expansion of a specific CHIP clone, it also may enhance the emergence of an alternate clone with unknown impact on health.
Evidence that PPM1D mutant clones expand only in the context of certain chemotherapies raises the potential of using personalized medicine approaches in the selection of therapy. One can imagine an array of decision-making algorithms to guide therapeutic approaches to cancer or other conditions while taking a patient’s somatic mutational background into account. Innumerable studies, including an assignment of the relative risk of various mutations and mutant genes, will be needed to make such a personalized approach truly evidence based.
The relative accessibility of blood enabled the discovery of CHIP and its health implications. Recent studies show that clonal expansion also occurs in other tissues and may be a precursor to solid tumors.(Kakiuchi and Ogawa, 2021, Wijewardhane et al., 2021) Non-oncologic health consequences of clonal expansion in other tissues have yet to be defined, and knowledge in this area will undoubtedly expand in the coming years. These future studies will shape our understanding of how systemic therapies or lifestyle modifications impact clonal evolution in non-hematologic tissues.
King and colleagues analyze how environmental contexts associated with clonal hematopoiesis of indeterminate potential (CHIP) alter tissue microenvironments to facilitate the selection and expansion of specific CHIP mutant clones. They also consider major remaining gaps in knowledge that will determine future application of this field in transplant and preventive medicine.
Acknowledgements:
The authors thank Catherine Gillespie for editing the final manuscript, and Margaret Goodell, Daisuke Nakada, and Yun Huang and their lab members for useful discussions. KYK was supported by NIH R35 HL155672, R01 HL136333, R01 HL134880, R01 AI141716, and a grant from the Helis Foundation. TW was supported by NIH R01 HL136333S. JD was supported by NIH R01 AG067584 and R01 AG066544. EMP was supported by NIH R01 DK119394 and the Cleo Meador and George Ryland Scott Endowed Chair in Hematology. MF was supported by the Ruth L. Kirschstein National Research Service Award (NRSA) Individual Predoctoral Fellowship to Promote Diversity in Health-Related Research (F31-Diversity-1F31HL156500) and the American Society of Hematology Minority Hematology Graduate Student Fellowship (MAF).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- 2004. Autoimmune diseases, cyclophosphamide. In: MORELAND LW (ed.) Rheumatology and Immunology Therapy. Berlin, Heidelberg: Springer Berlin Heidelberg. [Google Scholar]
- ABDEL-WAHAB O, MULLALLY A, HEDVAT C, GARCIA-MANERO G, PATEL J, WADLEIGH M, MALINGE S, YAO J, KILPIVAARA O, BHAT R, HUBERMAN K, THOMAS S, DOLGALEV I, HEGUY A, PAIETTA E, LE BEAU MM, BERAN M, TALLMAN MS, EBERT BL, KANTARJIAN HM, STONE RM, GILLILAND DG, CRISPINO JD & LEVINE RL 2009. Genetic characterization of TET1, TET2, and TET3 alterations in myeloid malignancies. Blood, 114, 144–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ABEGUNDE SO, BUCKSTEIN R, WELLS RA & RAUH MJ 2018. An inflammatory environment containing TNFalpha favors Tet2-mutant clonal hematopoiesis. Exp Hematol, 59, 60–65. [DOI] [PubMed] [Google Scholar]
- ABELSON S, COLLORD G, NG SWK, WEISSBROD O, MENDELSON COHEN N, NIEMEYER E, BARDA N, ZUZARTE PC, HEISLER L, SUNDARAVADANAM Y, LUBEN R, HAYAT S, WANG TT, ZHAO Z, CIRLAN I, PUGH TJ, SOAVE D, NG K, LATIMER C, HARDY C, RAINE K, JONES D, HOULT D, BRITTEN A, MCPHERSON JD, JOHANSSON M, MBABAALI F, EAGLES J, MILLER JK, PASTERNACK D, TIMMS L, KRZYZANOWSKI P, AWADALLA P, COSTA R, SEGAL E, BRATMAN SV, BEER P, BEHJATI S, MARTINCORENA I, WANG JCY, BOWLES KM, QUIROS JR, KARAKATSANI A, LA VECCHIA C, TRICHOPOULOU A, SALAMANCA-FERNANDEZ E, HUERTA JM, BARRICARTE A, TRAVIS RC, TUMINO R, MASALA G, BOEING H, PANICO S, KAAKS R, KRAMER A, SIERI S, RIBOLI E, VINEIS P, FOLL M, MCKAY J, POLIDORO S, SALA N, KHAW KT, VERMEULEN R, CAMPBELL PJ, PAPAEMMANUIL E, MINDEN MD, TANAY A, BALICER RD, WAREHAM NJ, GERSTUNG M, DICK JE, BRENNAN P, VASSILIOU GS & SHLUSH LI 2018. Prediction of acute myeloid leukaemia risk in healthy individuals. Nature, 559, 400–404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ABPLANALP WT, CREMER S, JOHN D, HOFFMANN J, SCHUHMACHER B, MERTEN M, RIEGER MA, VASA-NICOTERA M, ZEIHER AM & DIMMELER S 2021. Clonal Hematopoiesis-Driver DNMT3A Mutations Alter Immune Cells in Heart Failure. Circ Res, 128, 216–228. [DOI] [PubMed] [Google Scholar]
- AGRAWAL M, NIROULA A, CUNIN P, MCCONKEY M, KIM PG, WONG WJ, WEEKS LD, LIN AE, MILLER PG, GIBSON CJ, SEKAR A, NEUBERG DS, BICK AG, NATARAJAN P, NIGROVIC PA, RAO DA & EBERT BL 2021. The Association between Clonal Hematopoiesis and Gout. Blood, 138, 595–595.34436531 [Google Scholar]
- AKBARI B, GHAHRI-SAREMI N, SOLTANTOYEH T, HADJATI J, GHASSEMI S & MIRZAEI HR 2021. Epigenetic strategies to boost CAR T cell therapy. Mol Ther, 29, 2640–2659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ALFONSO PIEROLA A, MARCHESINI M, TAKAHASHI K, GAÑÁN-GóMEZ I, FIORINI E, OGOTI Y, IRLES E, MONTALBAN-BRAVO G, SOFIA S, DWYER KC, GARCIA-MANERO G & COLLA S 2016. The Role of Chip-Related DNA Damage Response Dysfunction in Therapy-Related Myeloid Neoplasms. Blood, 128, 958–958. [Google Scholar]
- ALLSOPP RC, MORIN GB, DEPINHO R, HARLEY CB & WEISSMAN IL 2003. Telomerase is required to slow telomere shortening and extend replicative lifespan of HSCs during serial transplantation. Blood, 102, 517–20. [DOI] [PubMed] [Google Scholar]
- AVAGYAN S, HENNINGER JE, MANNHERZ WP, MISTRY M, YOON J, YANG S, WEBER MC, MOORE JL & ZON LI 2021. Resistance to inflammation underlies enhanced fitness in clonal hematopoiesis. Science, 374, 768–772. [DOI] [PubMed] [Google Scholar]
- BALDRIDGE MT, KING KY, BOLES NC, WEKSBERG DC & GOODELL MA 2010. Quiescent haematopoietic stem cells are activated by IFN-gamma in response to chronic infection. Nature, 465, 793–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BEERMAN I 2017. Accumulation of DNA damage in the aged hematopoietic stem cell compartment. Semin Hematol, 54, 12–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BEKELE DI & PATNAIK MM 2020. Autoimmunity, Clonal Hematopoiesis, and Myeloid Neoplasms. Rheum Dis Clin North Am, 46, 429–444. [DOI] [PubMed] [Google Scholar]
- BEZERRA MF, LIMA AS, PIQUE-BORRAS MR, SILVEIRA DR, COELHO-SILVA JL, PEREIRA-MARTINS DA, WEINHAUSER I, FRANCA-NETO PL, QUEK L, CORBY A, OLIVEIRA MM, LIMA MM, DE ASSIS RA, DE MELO CAMPOS P, DUARTE BK, BENDIT I, ROCHA V, REGO EM, TRAINA F, SAAD ST, BELTRAO EI, BEZERRA MA & LUCENA-ARAUJO AR 2020. Co-occurrence of DNMT3A, NPM1, FLT3 mutations identifies a subset of acute myeloid leukemia with adverse prognosis. Blood, 135, 870–875. [DOI] [PubMed] [Google Scholar]
- BHATTACHARYA R, ZEKAVAT SM, HAESSLER J, FORNAGE M, RAFFIELD L, UDDIN MM, BICK AG, NIROULA A, YU B, GIBSON C, GRIFFIN G, MORRISON AC, PSATY BM, LONGSTRETH WT, BIS JC, RICH SS, ROTTER JI, TRACY RP, CORREA A, SESHADRI S, JOHNSON A, COLLINS JM, HAYDEN KM, MADSEN TE, BALLANTYNE CM, JAISWAL S, EBERT BL, KOOPERBERG C, MANSON JE, WHITSEL EA, PROGRAM NT-OFPM, NATARAJAN P & REINER AP 2021. Clonal Hematopoiesis Is Associated With Higher Risk of Stroke. Stroke, STROKEAHA121037388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BICK AG, PIRRUCCELLO JP, GRIFFIN GK, GUPTA N, GABRIEL S, SALEHEEN D, LIBBY P, KATHIRESAN S & NATARAJAN P 2020a. Genetic Interleukin 6 Signaling Deficiency Attenuates Cardiovascular Risk in Clonal Hematopoiesis. Circulation, 141, 124–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BICK AG, POPADIN K, THORBALL CW, UDDIN MM, ZANNI M, YU B, CAVASSINI M, RAUCH A, TARR P, SCHMID P, BERNASCONI E, GUNTHARD H, LIBBY P, BOERWINKLE E, MCLAREN P, BALLANTYNE CM, GRINSPOON SK, NATARAJAN P & FELLAY J 2020b. Increased CHIP Prevalence Amongst People Living with HIV. medRxiv. [Google Scholar]
- BODDU PC & ZEIDAN AM 2019. Myeloid disorders after autoimmune disease. Best Pract Res Clin Haematol, 32, 74–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BOLTON KL, KOH Y, FOOTE MB, IM H, JEE J, SUN CH, SAFONOV A, PTASHKIN R, MOON JH, LEE JY, JUNG J, KANG CK, SONG KH, CHOE PG, PARK WB, KIM HB, OH MD, SONG H, KIM S, PATEL M, DERKACH A, GEDVILAITE E, TKACHUK KA, WILEY BJ, CHAN IC, BRAUNSTEIN LZ, GAO T, PAPAEMMANUIL E, ESTHER BABADY N, PESSIN MS, KAMBOJ M, DIAZ LA JR., LADANYI M, RAUH MJ, NATARAJAN P, MACHIELA MJ, AWADALLA P, JOSEPH V, OFFIT K, NORTON L, BERGER MF, LEVINE RL, KIM ES, KIM NJ & ZEHIR A 2021. Clonal hematopoiesis is associated with risk of severe Covid-19. Nat Commun, 12, 5975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BOLTON KL, PTASHKIN RN, GAO T, BRAUNSTEIN L, DEVLIN SM, KELLY D, PATEL M, BERTHON A, SYED A, YABE M, COOMBS CC, CALTABELLOTTA NM, WALSH M, OFFIT K, STADLER Z, MANDELKER D, SCHULMAN J, PATEL A, PHILIP J, BERNARD E, GUNDEM G, OSSA JEA, LEVINE M, MARTINEZ JSM, FARNOUD N, GLODZIK D, LI S, ROBSON ME, LEE C, PHAROAH PDP, STOPSACK KH, SPITZER B, MANTHA S, FAGIN J, BOUCAI L, GIBSON CJ, EBERT BL, YOUNG AL, DRULEY T, TAKAHASHI K, GILLIS N, BALL M, PADRON E, HYMAN DM, BASELGA J, NORTON L, GARDOS S, KLIMEK VM, SCHER H, BAJORIN D, PARAISO E, BENAYED R, ARCILA ME, LADANYI M, SOLIT DB, BERGER MF, TALLMAN M, GARCIA-CLOSAS M, CHATTERJEE N, DIAZ LA JR., LEVINE RL, MORTON LM, ZEHIR A & PAPAEMMANUIL E 2020. Cancer therapy shapes the fitness landscape of clonal hematopoiesis. Nat Genet, 52, 1219–1226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BONDAR T & MEDZHITOV R 2010. p53-mediated hematopoietic stem and progenitor cell competition. Cell Stem Cell, 6, 309–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BOUCAI L, FALCONE J, UKENA J, COOMBS CC, ZEHIR A, PTASHKIN R, BERGER MF, LEVINE RL & FAGIN JA 2018. Radioactive Iodine-Related Clonal Hematopoiesis in Thyroid Cancer Is Common and Associated With Decreased Survival. J Clin Endocrinol Metab, 103, 4216–4223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BOWMAN RL & LEVINE RL 2017. TET2 in Normal and Malignant Hematopoiesis. Cold Spring Harb Perspect Med, 7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- BRUNNER AM, NEUBERG DS, WANDER SA, SADRZADEH H, BALLEN KK, AMREIN PC, ATTAR E, HOBBS GS, CHEN YB, PERRY A, CONNOLLY C, JOSEPH C, BURKE M, RAMOS A, GALINSKY I, YEN K, YANG H, STRALEY K, AGRESTA S, ADAMIA S, BORGER DR, IAFRATE A, GRAUBERT TA, STONE RM & FATHI AT 2019. Isocitrate dehydrogenase 1 and 2 mutations, 2-hydroxyglutarate levels, and response to standard chemotherapy for patients with newly diagnosed acute myeloid leukemia. Cancer, 125, 541–549. [DOI] [PubMed] [Google Scholar]
- BUSCARLET M, PROVOST S, ZADA YF, BARHDADI A, BOURGOIN V, LEPINE G, MOLLICA L, SZUBER N, DUBE MP & BUSQUE L 2017. DNMT3A and TET2 dominate clonal hematopoiesis and demonstrate benign phenotypes and different genetic predispositions. Blood, 130, 753–762. [DOI] [PubMed] [Google Scholar]
- BUSQUE L, PATEL JP, FIGUEROA ME, VASANTHAKUMAR A, PROVOST S, HAMILOU Z, MOLLICA L, LI J, VIALE A, HEGUY A, HASSIMI M, SOCCI N, BHATT PK, GONEN M, MASON CE, MELNICK A, GODLEY LA, BRENNAN CW, ABDEL-WAHAB O & LEVINE RL 2012. Recurrent somatic TET2 mutations in normal elderly individuals with clonal hematopoiesis. Nat Genet, 44, 1179–81. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CAI Z, KOTZIN JJ, RAMDAS B, CHEN S, NELANUTHALA S, PALAM LR, PANDEY R, MALI RS, LIU Y, KELLEY MR, SANDUSKY G, MOHSENI M, WILLIAMS A, HENAO-MEJIA J & KAPUR R 2018. Inhibition of Inflammatory Signaling in Tet2 Mutant Preleukemic Cells Mitigates Stress-Induced Abnormalities and Clonal Hematopoiesis. Cell Stem Cell, 23, 833–849 e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CAI Z, LU X, ZHANG C, NELANUTHALA S, AGUILERA F, HADLEY A, RAMDAS B, FANG F, NEPHEW K, KOTZIN JJ, WILLIAMS A, HENAO-MEJIA J, HANELINE L & KAPUR R 2021. Hyperglycemia cooperates with Tet2 heterozygosity to induce leukemia driven by proinflammatory cytokine-induced lncRNA Morrbid. J Clin Invest, 131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CAIADO F, PIETRAS EM & MANZ MG 2021. Inflammation as a regulator of hematopoietic stem cell function in disease, aging, and clonal selection. J Exp Med, 218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CELIK H, MALLANEY C, KOTHARI A, OSTRANDER EL, EULTGEN E, MARTENS A, MILLER CA, HUNDAL J, KLCO JM & CHALLEN GA 2015. Enforced differentiation of Dnmt3a-null bone marrow leads to failure with c-Kit mutations driving leukemic transformation. Blood, 125, 619–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHALLEN GA, SUN D, JEONG M, LUO M, JELINEK J, BERG JS, BOCK C, VASANTHAKUMAR A, GU H, XI Y, LIANG S, LU Y, DARLINGTON GJ, MEISSNER A, ISSA JP, GODLEY LA, LI W & GOODELL MA 2011. Dnmt3a is essential for hematopoietic stem cell differentiation. Nat Genet, 44, 23–31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHAUDRY SF & CHEVASSUT TJ 2017. Epigenetic Guardian: A Review of the DNA Methyltransferase DNMT3A in Acute Myeloid Leukaemia and Clonal Haematopoiesis. Biomed Res Int, 2017, 5473197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHEN J, ELLISON FM, KEYVANFAR K, OMOKARO SO, DESIERTO MJ, ECKHAUS MA & YOUNG NS 2008. Enrichment of hematopoietic stem cells with SLAM and LSK markers for the detection of hematopoietic stem cell function in normal and Trp53 null mice. Exp Hematol, 36, 1236–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHEN J, MATATALL KA, FENG X, HORMAECHEA-AGULLA D, MAHARJAN M, YOUNG N & KING KY 2020. Dnmt3a-null hematopoietic stem and progenitor cells expand after busulfan treatment. Exp Hematol, 91, 39–45 e2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHEN J, NIE D, WANG X, WANG L, WANG F, ZHANG Y, CHEN X, CAO P, LI M, MA X, YU T, ZHOU X, TIAN W, ZHANG Y, LI Y, TAN Y, SUN W, FANG J, LIU M, XU W & LIU H 2021. Enriched clonal hematopoiesis in seniors with dietary vitamin C inadequacy. Clin Nutr ESPEN, 46, 179–184. [DOI] [PubMed] [Google Scholar]
- CHEN S & LIU Y 2019. p53 involvement in clonal hematopoiesis of indeterminate potential. Curr Opin Hematol, 26, 235–240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHEN S, WANG Q, YU H, CAPITANO ML, VEMULA S, NABINGER SC, GAO R, YAO C, KOBAYASHI M, GENG Z, FAHEY A, HENLEY D, LIU SZ, BARAJAS S, CAI W, WOLF ER, RAMDAS B, CAI Z, GAO H, LUO N, SUN Y, WONG TN, LINK DC, LIU Y, BOSWELL HS, MAYO LD, HUANG G, KAPUR R, YODER MC, BROXMEYER HE, GAO Z & LIU Y 2019. Mutant p53 drives clonal hematopoiesis through modulating epigenetic pathway. Nat Commun, 10, 5649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHITRE S, STOLZEL F, CUTHILL K, STREETLY M, GRAHAM C, DILL C, MOHAMEDALI A, SMITH A, SCHETELIG J, ALTMANN H, BORNHAUSER M & MUFTI GJ 2018. Clonal hematopoiesis in patients with multiple myeloma undergoing autologous stem cell transplantation. Leukemia, 32, 2020–2024. [DOI] [PubMed] [Google Scholar]
- CHOU WC, CHOU SC, LIU CY, CHEN CY, HOU HA, KUO YY, LEE MC, KO BS, TANG JL, YAO M, TSAY W, WU SJ, HUANG SY, HSU SC, CHEN YC, CHANG YC, KUO YY, KUO KT, LEE FY, LIU MC, LIU CW, TSENG MH, HUANG CF & TIEN HF 2011. TET2 mutation is an unfavorable prognostic factor in acute myeloid leukemia patients with intermediate-risk cytogenetics. Blood, 118, 3803–10. [DOI] [PubMed] [Google Scholar]
- CHUNG HY, SUNG B, JUNG KJ, ZOU Y & YU BP 2006. The molecular inflammatory process in aging. Antioxid Redox Signal, 8, 572–81. [DOI] [PubMed] [Google Scholar]
- CIMMINO L, DOLGALEV I, WANG Y, YOSHIMI A, MARTIN GH, WANG J, NG V, XIA B, WITKOWSKI MT, MITCHELL-FLACK M, GRILLO I, BAKOGIANNI S, NDIAYE-LOBRY D, MARTIN MT, GUILLAMOT M, BANH RS, XU M, FIGUEROA ME, DICKINS RA, ABDEL-WAHAB O, PARK CY, TSIRIGOS A, NEEL BG & AIFANTIS I 2017. Restoration of TET2 Function Blocks Aberrant Self-Renewal and Leukemia Progression. Cell, 170, 1079–1095 e20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- COLLA S, ONG DS, OGOTI Y, MARCHESINI M, MISTRY NA, CLISE-DWYER K, ANG SA, STORTI P, VIALE A, GIULIANI N, RUISAARD K, GANAN GOMEZ I, BRISTOW CA, ESTECIO M, WEKSBERG DC, HO YW, HU B, GENOVESE G, PETTAZZONI P, MULTANI AS, JIANG S, HUA S, RYAN MC, CARUGO A, NEZI L, WEI Y, YANG H, D’ANCA M, ZHANG L, GADDIS S, GONG T, HORNER JW, HEFFERNAN TP, JONES P, COOPER LJ, LIANG H, KANTARJIAN H, WANG YA, CHIN L, BUESO-RAMOS C, GARCIA-MANERO G & DEPINHO RA 2015. Telomere dysfunction drives aberrant hematopoietic differentiation and myelodysplastic syndrome. Cancer Cell, 27, 644–57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- COOMBS CC, ZEHIR A, DEVLIN SM, KISHTAGARI A, SYED A, JONSSON P, HYMAN DM, SOLIT DB, ROBSON ME, BASELGA J, ARCILA ME, LADANYI M, TALLMAN MS, LEVINE RL & BERGER MF 2017. Therapy-Related Clonal Hematopoiesis in Patients with Non-hematologic Cancers Is Common and Associated with Adverse Clinical Outcomes. Cell Stem Cell, 21, 374–382 e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CREMER S, KIRSCHBAUM K, BERKOWITSCH A, JOHN D, KIEFER K, DORSHEIMER L, WAGNER J, RASPER T, ABOU-EL-ARDAT K, ASSMUS B, SERVE H, RIEGER M, DIMMELER S & ZEIHER AM 2020. Multiple Somatic Mutations for Clonal Hematopoiesis Are Associated With Increased Mortality in Patients With Chronic Heart Failure. Circ Genom Precis Med, 13, e003003. [DOI] [PubMed] [Google Scholar]
- DAVIS EJ, SALEM JE, YOUNG A, GREEN JR, FERRELL PB, ANCELL KK, LEBRUN-VIGNES B, MOSLEHI JJ & JOHNSON DB 2019. Hematologic Complications of Immune Checkpoint Inhibitors. Oncologist, 24, 584–588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DAWOUD AAZ, GILBERT RD, TAPPER WJ & CROSS NCP 2021. Clonal myelopoiesis promotes adverse outcomes in chronic kidney disease. Leukemia. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DAWOUD AAZ, TAPPER WJ & CROSS NCP 2020. Clonal myelopoiesis in the UK Biobank cohort: ASXL1 mutations are strongly associated with smoking. Leukemia, 34, 2660–2672. [DOI] [PubMed] [Google Scholar]
- DELHOMMEAU F, DUPONT S, DELLA VALLE V, JAMES C, TRANNOY S, MASSE A, KOSMIDER O, LE COUEDIC JP, ROBERT F, ALBERDI A, LECLUSE Y, PLO I, DREYFUS FJ, MARZAC C, CASADEVALL N, LACOMBE C, ROMANA SP, DESSEN P, SOULIER J, VIGUIE F, FONTENAY M, VAINCHENKER W & BERNARD OA 2009. Mutation in TET2 in myeloid cancers. N Engl J Med, 360, 2289–301. [DOI] [PubMed] [Google Scholar]
- DESAI P, MENCIA-TRINCHANT N, SAVENKOV O, SIMON MS, CHEANG G, LEE S, SAMUEL M, RITCHIE EK, GUZMAN ML, BALLMAN KV, ROBOZ GJ & HASSANE DC 2018. Somatic mutations precede acute myeloid leukemia years before diagnosis. Nat Med, 24, 1015–1023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DHARAN NJ, YEH P, BLOCH M, YEUNG MM, BAKER D, GUINTO J, ROTH N, FTOUNI S, OGNENOVSKA K, SMITH D, HOY JF, WOOLLEY I, PELL C, TEMPLETON DJ, FRASER N, ROSE N, HUTCHINSON J, PETOUMENOS K, DAWSON SJ, POLIZZOTTO MN, DAWSON MA & GROUP AS 2021. HIV is associated with an increased risk of age-related clonal hematopoiesis among older adults. Nat Med, 27, 1006–1011. [DOI] [PubMed] [Google Scholar]
- DORSHEIMER L, ASSMUS B, RASPER T, ORTMANN CA, ECKE A, ABOU-EL-ARDAT K, SCHMID T, BRUNE B, WAGNER S, SERVE H, HOFFMANN J, SEEGER F, DIMMELER S, ZEIHER AM & RIEGER MA 2019. Association of Mutations Contributing to Clonal Hematopoiesis With Prognosis in Chronic Ischemic Heart Failure. JAMA Cardiol, 4, 25–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ESFAHANI K, ROUDAIA L, BUHLAIGA N, DEL RINCON SV, PAPNEJA N & MILLER WH JR. 2020. A review of cancer immunotherapy: from the past, to the present, to the future. Curr Oncol, 27, S87–S97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ESKELUND CW, HUSBY S, FAVERO F, KLAUSEN TW, RODRIGUEZ-GONZALEZ FG, KOLSTAD A, PEDERSEN LB, RÄTY RK, GEISLER CH, JERKEMAN M, WEISCHENFELDT J & GRØNBÆK K 2020. Clonal hematopoiesis evolves from pretreatment clones and stabilizes after end of chemotherapy in patients with MCL. Blood, 135, 2000–2004. [DOI] [PubMed] [Google Scholar]
- ESSERS MA, OFFNER S, BLANCO-BOSE WE, WAIBLER Z, KALINKE U, DUCHOSAL MA & TRUMPP A 2009. IFNalpha activates dormant haematopoietic stem cells in vivo. Nature, 458, 904–8. [DOI] [PubMed] [Google Scholar]
- FABRE MA, ALMEIDA JGD, FIORILLO E, MITCHELL E, DAMASKOU A, RAK J, ORRÙ V, MARONGIU M, VIJAYABASKAR MS, BAXTER J, HARDY C, ABASCAL F, CHAPMAN MS, WILLIAMS N, NANGALIA J, MARTINCORENA I, CAMPBELL PJ, MCKINNEY EF, CUCCA F, GERSTUNG M & VASSILIOU GS 2021. The longitudinal dynamics and natural history of clonal haematopoiesis. bioRxiv, 2021.08.12.455048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FENG Y, LI X, CASSADY K, ZOU Z & ZHANG X 2019. TET2 Function in Hematopoietic Malignancies, Immune Regulation, and DNA Repair. Front Oncol, 9, 210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FEUSIER JE, ARUNACHALAM S, TASHI T, BAKER MJ, VANSANT-WEBB C, FERDIG A, WELM BE, RODRIGUEZ-FLORES JL, OURS C, JORDE LB, PRCHAL JT & MASON CC 2021. Large-Scale Identification of Clonal Hematopoiesis and Mutations Recurrent in Blood Cancers. Blood Cancer Discov, 2, 226–237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FLACH J, BAKKER ST, MOHRIN M, CONROY PC, PIETRAS EM, REYNAUD D, ALVAREZ S, DIOLAITI ME, UGARTE F, FORSBERG EC, LE BEAU MM, STOHR BA, MENDEZ J, MORRISON CG & PASSEGUE E 2014. Replication stress is a potent driver of functional decline in ageing haematopoietic stem cells. Nature, 512, 198–202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FRAIETTA JA, NOBLES CL, SAMMONS MA, LUNDH S, CARTY SA, REICH TJ, COGDILL AP, MORRISSETTE JJD, DENIZIO JE, REDDY S, HWANG Y, GOHIL M, KULIKOVSKAYA I, NAZIMUDDIN F, GUPTA M, CHEN F, EVERETT JK, ALEXANDER KA, LIN-SHIAO E, GEE MH, LIU X, YOUNG RM, AMBROSE D, WANG Y, XU J, JORDAN MS, MARCUCCI KT, LEVINE BL, GARCIA KC, ZHAO Y, KALOS M, PORTER DL, KOHLI RM, LACEY SF, BERGER SL, BUSHMAN FD, JUNE CH & MELENHORST JJ 2018. Disruption of TET2 promotes the therapeutic efficacy of CD19-targeted T cells. Nature, 558, 307–312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FRANCESCHI C, BONAFE M & VALENSIN S 2000. Human immunosenescence: the prevailing of innate immunity, the failing of clonotypic immunity, and the filling of immunological space. Vaccine, 18, 1717–20. [DOI] [PubMed] [Google Scholar]
- FRANCESCHI C & CAMPISI J 2014. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci, 69 Suppl 1, S4–9. [DOI] [PubMed] [Google Scholar]
- FRICK M, CHAN W, ARENDS CM, HABLESREITER R, HALIK A, HEUSER M, MICHONNEAU D, BLAU O, HOYER K, CHRISTEN F, GALAN-SOUSA J, NOERENBERG D, WAIS V, STADLER M, YOSHIDA K, SCHETELIG J, SCHULER E, THOL F, CLAPPIER E, CHRISTOPEIT M, AYUK F, BORNHAUSER M, BLAU IW, OGAWA S, ZEMOJTEL T, GERBITZ A, WAGNER EM, SPRIEWALD BM, SCHREZENMEIER H, KUCHENBAUER F, KOBBE G, WIESNETH M, KOLDEHOFF M, SOCIE G, KROEGER N, BULLINGER L, THIEDE C & DAMM F 2019. Role of Donor Clonal Hematopoiesis in Allogeneic Hematopoietic Stem-Cell Transplantation. J Clin Oncol, 37, 375–385. [DOI] [PubMed] [Google Scholar]
- FUJINO T, GOYAMA S, SUGIURA Y, INOUE D, ASADA S, YAMASAKI S, MATSUMOTO A, YAMAGUCHI K, ISOBE Y, TSUCHIYA A, SHIKATA S, SATO N, MORINAGA H, FUKUYAMA T, TANAKA Y, FUKUSHIMA T, TAKEDA R, YAMAMOTO K, HONDA H, NISHIMURA EK, FURUKAWA Y, SHIBATA T, ABDEL-WAHAB O, SUEMATSU M & KITAMURA T 2021. Mutant ASXL1 induces age-related expansion of phenotypic hematopoietic stem cells through activation of Akt/mTOR pathway. Nat Commun, 12, 1826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FUJINO T & KITAMURA T 2020. ASXL1 mutation in clonal hematopoiesis. Exp Hematol, 83, 74–84. [DOI] [PubMed] [Google Scholar]
- FUSTER JJ, MACLAUCHLAN S, ZURIAGA MA, POLACKAL MN, OSTRIKER AC, CHAKRABORTY R, WU CL, SANO S, MURALIDHARAN S, RIUS C, VUONG J, JACOB S, MURALIDHAR V, ROBERTSON AA, COOPER MA, ANDRES V, HIRSCHI KK, MARTIN KA & WALSH K 2017. Clonal hematopoiesis associated with TET2 deficiency accelerates atherosclerosis development in mice. Science, 355, 842–847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- FUSTER JJ, ZURIAGA MA, ZORITA V, MACLAUCHLAN S, POLACKAL MN, VIANA-HUETE V, FERRER-PEREZ A, MATESANZ N, HERRERO-CERVERA A, SANO S, COOPER MA, GONZALEZ-NAVARRO H & WALSH K 2020. TET2-Loss-of-Function-Driven Clonal Hematopoiesis Exacerbates Experimental Insulin Resistance in Aging and Obesity. Cell Rep, 33, 108326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GAIDZIK VI, SCHLENK RF, PASCHKA P, STOLZLE A, SPATH D, KUENDGEN A, VON LILIENFELD-TOAL M, BRUGGER W, DERIGS HG, KREMERS S, GREIL R, RAGHAVACHAR A, RINGHOFFER M, SALIH HR, WATTAD M, KIRCHEN HG, RUNDE V, HEIL G, PETZER AL, GIRSCHIKOFSKY M, HEUSER M, KAYSER S, GOEHRING G, TELEANU MV, SCHLEGELBERGER B, GANSER A, KRAUTER J, BULLINGER L, DOHNER H & DOHNER K 2013. Clinical impact of DNMT3A mutations in younger adult patients with acute myeloid leukemia: results of the AML Study Group (AMLSG). Blood, 121, 4769–77. [DOI] [PubMed] [Google Scholar]
- GARTUNG A, YANG J, SUKHATME VP, BIELENBERG DR, FERNANDES D, CHANG J, SCHMIDT BA, HWANG SH, ZURAKOWSKI D, HUANG S, KIERAN MW, HAMMOCK BD & PANIGRAHY D 2019. Suppression of chemotherapy-induced cytokine/lipid mediator surge and ovarian cancer by a dual COX-2/sEH inhibitor. Proc Natl Acad Sci U S A, 116, 1698–1703. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GELSI-BOYER V, BRECQUEVILLE M, DEVILLIER R, MURATI A, MOZZICONACCI MJ & BIRNBAUM D 2012. Mutations in ASXL1 are associated with poor prognosis across the spectrum of malignant myeloid diseases. J Hematol Oncol, 5, 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GENOVESE G, KAHLER AK, HANDSAKER RE, LINDBERG J, ROSE SA, BAKHOUM SF, CHAMBERT K, MICK E, NEALE BM, FROMER M, PURCELL SM, SVANTESSON O, LANDEN M, HOGLUND M, LEHMANN S, GABRIEL SB, MORAN JL, LANDER ES, SULLIVAN PF, SKLAR P, GRONBERG H, HULTMAN CM & MCCARROLL SA 2014. Clonal hematopoiesis and blood-cancer risk inferred from blood DNA sequence. N Engl J Med, 371, 2477–87. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GIBSON CJ, KENNEDY JA, NIKIFOROW S, KUO FC, ALYEA EP, HO V, RITZ J, SOIFFER R, ANTIN JH & LINDSLEY RC 2017a. Donor-engrafted CHIP is common among stem cell transplant recipients with unexplained cytopenias. Blood, 130, 91–94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GIBSON CJ, LINDSLEY RC, TCHEKMEDYIAN V, MAR BG, SHI J, JAISWAL S, BOSWORTH A, FRANCISCO L, HE J, BANSAL A, MORGAN EA, LACASCE AS, FREEDMAN AS, FISHER DC, JACOBSEN E, ARMAND P, ALYEA EP, KORETH J, HO V, SOIFFER RJ, ANTIN JH, RITZ J, NIKIFOROW S, FORMAN SJ, MICHOR F, NEUBERG D, BHATIA R, BHATIA S & EBERT BL 2017b. Clonal Hematopoiesis Associated With Adverse Outcomes After Autologous Stem-Cell Transplantation for Lymphoma. J Clin Oncol, 35, 1598–1605. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GILLIS NK, BALL M, ZHANG Q, MA Z, ZHAO Y, YODER SJ, BALASIS ME, MESA TE, SALLMAN DA, LANCET JE, KOMROKJI RS, LIST AF, MCLEOD HL, ALSINA M, BAZ R, SHAIN KH, ROLLISON DE & PADRON E 2017. Clonal haemopoiesis and therapy-related myeloid malignancies in elderly patients: a proof-of-concept, case-control study. Lancet Oncol, 18, 112–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GONZALO S, JACO I, FRAGA MF, CHEN T, LI E, ESTELLER M & BLASCO MA 2006. DNA methyltransferases control telomere length and telomere recombination in mammalian cells. Nat Cell Biol, 8, 416–24. [DOI] [PubMed] [Google Scholar]
- GRIMM J, BILL M, JENTZSCH M, BEINICKE S, HANTSCHEL J, GOLDMANN K, SCHULZ J, CROSS M, FRANKE GN, BEHRE G, VUCINIC V, PONISCH W, LANGE T, NIEDERWIESER D & SCHWIND S 2019. Clinical impact of clonal hematopoiesis in acute myeloid leukemia patients receiving allogeneic transplantation. Bone Marrow Transplant, 54, 1189–1197. [DOI] [PubMed] [Google Scholar]
- GURYANOVA OA, SHANK K, SPITZER B, LUCIANI L, KOCHE RP, GARRETT-BAKELMAN FE, GANZEL C, DURHAM BH, MOHANTY A, HOERMANN G, RIVERA SA, CHRAMIEC AG, PRONIER E, BASTIAN L, KELLER MD, TOVBIN D, LOIZOU E, WEINSTEIN AR, GONZALEZ AR, LIEU YK, ROWE JM, PASTORE F, MCKENNEY AS, KRIVTSOV AV, SPERR WR, CROSS JR, MASON CE, TALLMAN MS, ARCILA ME, ABDEL-WAHAB O, ARMSTRONG SA, KUBICEK S, STABER PB, GONEN M, PAIETTA EM, MELNICK AM, NIMER SD, MUKHERJEE S & LEVINE RL 2016. DNMT3A mutations promote anthracycline resistance in acute myeloid leukemia via impaired nucleosome remodeling. Nat Med, 22, 1488–1495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HAFERLACH T, NAGATA Y, GROSSMANN V, OKUNO Y, BACHER U, NAGAE G, SCHNITTGER S, SANADA M, KON A, ALPERMANN T, YOSHIDA K, ROLLER A, NADARAJAH N, SHIRAISHI Y, SHIOZAWA Y, CHIBA K, TANAKA H, KOEFFLER HP, KLEIN HU, DUGAS M, ABURATANI H, KOHLMANN A, MIYANO S, HAFERLACH C, KERN W & OGAWA S 2014. Landscape of genetic lesions in 944 patients with myelodysplastic syndromes. Leukemia, 28, 241–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HANSSON GK, ROBERTSON AK & SODERBERG-NAUCLER C 2006. Inflammation and atherosclerosis. Annu Rev Pathol, 1, 297–329. [DOI] [PubMed] [Google Scholar]
- HARING B, REINER AP, LIU J, TOBIAS DK, WHITSEL E, BERGER JS, DESAI P, WASSERTHEIL-SMOLLER S, LAMONTE MJ, HAYDEN KM, BICK AG, NATARAJAN P, WEINSTOCK JS, NGUYEN PK, STEFANICK M, SIMON MS, EATON CB, KOOPERBERG C & MANSON JE 2021. Healthy Lifestyle and Clonal Hematopoiesis of Indeterminate Potential: Results From the Women’s Health Initiative. J Am Heart Assoc, 10, e018789. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HARRISON DE & ASTLE CM 1982. Loss of stem cell repopulating ability upon transplantation. Effects of donor age, cell number, and transplantation procedure. J Exp Med, 156, 1767–79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HEINI AD, PORRET N, ZENHAEUSERN R, WINKLER A, BACHER U & PABST T 2021. Clonal Hematopoiesis after Autologous Stem Cell Transplantation Does Not Confer Adverse Prognosis in Patients with AML. Cancers (Basel), 13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HEYDE A, ROHDE D, MCALPINE CS, ZHANG S, HOYER FF, GEROLD JM, CHEEK D, IWAMOTO Y, SCHLOSS MJ, VANDOORNE K, IBORRA-EGEA O, MUNOZ-GUIJOSA C, BAYES-GENIS A, REITER JG, CRAIG M, SWIRSKI FK, NAHRENDORF M, NOWAK MA & NAXEROVA K 2021. Increased stem cell proliferation in atherosclerosis accelerates clonal hematopoiesis. Cell, 184, 1348–1361 e22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HIGA KC, GOODSPEED A, CHAVEZ JS, DE DOMINICI M, DANIS E, ZABEREZHNYY V, RABE JL, TENEN DG, PIETRAS EM & DEGREGORI J 2021. Chronic interleukin-1 exposure triggers selection for Cebpa-knockout multipotent hematopoietic progenitors. J Exp Med, 218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HONIGBERG MC, ZEKAVAT SM, NIROULA A, GRIFFIN GK, BICK AG, PIRRUCCELLO JP, NAKAO T, WHITSEL EA, FARLAND LV, LAURIE C, KOOPERBERG C, MANSON JE, GABRIEL S, LIBBY P, REINER AP, EBERT BL, PROGRAM NT-OFPM & NATARAJAN P 2021. Premature Menopause, Clonal Hematopoiesis, and Coronary Artery Disease in Postmenopausal Women. Circulation, 143, 410–423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HORMAECHEA-AGULLA D, MATATALL KA, LE DT, KAIN B, LONG X, KUS P, JAKSIK R, CHALLEN GA, KIMMEL M & KING KY 2021. Chronic infection drives Dnmt3a-loss-of-function clonal hematopoiesis via IFNgamma signaling. Cell Stem Cell, 28, 1428–1442 e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HSIEH A, MORTON SU, WILLCOX JAL, GORHAM JM, TAI AC, QI H, DEPALMA S, MCKEAN D, GRIFFIN E, MANHEIMER KB, BERNSTEIN D, KIM RW, NEWBURGER JW, PORTER GA JR., SRIVASTAVA D, TRISTANI-FIROUZI M, BRUECKNER M, LIFTON RP, GOLDMUNTZ E, GELB BD, CHUNG WK, SEIDMAN CE, SEIDMAN JG & SHEN Y 2020. EM-mosaic detects mosaic point mutations that contribute to congenital heart disease. Genome Med, 12, 42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- HSU JI, DAYARAM T, TOVY A, DE BRAEKELEER E, JEONG M, WANG F, ZHANG J, HEFFERNAN TP, GERA S, KOVACS JJ, MARSZALEK JR, BRISTOW C, YAN Y, GARCIA-MANERO G, KANTARJIAN H, VASSILIOU G, FUTREAL PA, DONEHOWER LA, TAKAHASHI K & GOODELL MA 2018. PPM1D Mutations Drive Clonal Hematopoiesis in Response to Cytotoxic Chemotherapy. Cell Stem Cell, 23, 700–713 e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- IM AP, SEHGAL AR, CARROLL MP, SMITH BD, TEFFERI A, JOHNSON DE & BOYIADZIS M 2014. DNMT3A and IDH mutations in acute myeloid leukemia and other myeloid malignancies: associations with prognosis and potential treatment strategies. Leukemia, 28, 1774–83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ITALIANI P, MANCA ML, ANGELOTTI F, MELILLO D, PRATESI F, PUXEDDU I, BORASCHI D & MIGLIORINI P 2018. IL-1 family cytokines and soluble receptors in systemic lupus erythematosus. Arthritis Res Ther, 20, 27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ITO K, BONORA M & ITO K 2019. Metabolism as master of hematopoietic stem cell fate. Int J Hematol, 109, 18–27. [DOI] [PMC free article] [PubMed] [Google Scholar]
- IZZO F, LEE SC, PORAN A, CHALIGNE R, GAITI F, GROSS B, MURALI RR, DEOCHAND SD, ANG C, JONES PW, NAM AS, KIM KT, KOTHEN-HILL S, SCHULMAN RC, KI M, LHOUMAUD P, SKOK JA, VINY AD, LEVINE RL, KENIGSBERG E, ABDEL-WAHAB O & LANDAU DA 2020. DNA methylation disruption reshapes the hematopoietic differentiation landscape. Nat Genet, 52, 378–387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JAIN N, ZHAO Z, IYER A, LOPEZ M, FEUCHT J, KOCHE R, YANG J, ZHAN Y & SADELAIN M 2021. Emergence of a hyper-proliferative phenotype in TET2 edited human CAR T cells. Cancer Research, 81, LB153–LB153. [Google Scholar]
- JAISWAL S & EBERT BL 2019. Clonal hematopoiesis in human aging and disease. Science, 366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JAISWAL S, FONTANILLAS P, FLANNICK J, MANNING A, GRAUMAN PV, MAR BG, LINDSLEY RC, MERMEL CH, BURTT N, CHAVEZ A, HIGGINS JM, MOLTCHANOV V, KUO FC, KLUK MJ, HENDERSON B, KINNUNEN L, KOISTINEN HA, LADENVALL C, GETZ G, CORREA A, BANAHAN BF, GABRIEL S, KATHIRESAN S, STRINGHAM HM, MCCARTHY MI, BOEHNKE M, TUOMILEHTO J, HAIMAN C, GROOP L, ATZMON G, WILSON JG, NEUBERG D, ALTSHULER D & EBERT BL 2014. Age-related clonal hematopoiesis associated with adverse outcomes. N Engl J Med, 371, 2488–98. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JAISWAL S, NATARAJAN P, SILVER AJ, GIBSON CJ, BICK AG, SHVARTZ E, MCCONKEY M, GUPTA N, GABRIEL S, ARDISSINO D, BABER U, MEHRAN R, FUSTER V, DANESH J, FROSSARD P, SALEHEEN D, MELANDER O, SUKHOVA GK, NEUBERG D, LIBBY P, KATHIRESAN S & EBERT BL 2017. Clonal Hematopoiesis and Risk of Atherosclerotic Cardiovascular Disease. N Engl J Med, 377, 111–121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JAN M, EBERT BL & JAISWAL S 2017. Clonal hematopoiesis. Semin Hematol, 54, 43–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JANISZEWSKA H, BAK A, SKONIECZKA K, JASKOWIEC A, KIELBINSKI M, JACHALSKA A, CZYZEWSKA M, JAZWIEC B, KULISZKIEWICZ-JANUS M, CZYZ J, KULICZKOWSKI K & HAUS O 2018. Constitutional mutations of the CHEK2 gene are a risk factor for MDS, but not for de novo AML. Leuk Res, 70, 74–78. [DOI] [PubMed] [Google Scholar]
- JEONG M, PARK HJ, CELIK H, OSTRANDER EL, REYES JM, GUZMAN A, RODRIGUEZ B, LEI Y, LEE Y, DING L, GURYANOVA OA, LI W, GOODELL MA & CHALLEN GA 2018. Loss of Dnmt3a Immortalizes Hematopoietic Stem Cells In Vivo. Cell Rep, 23, 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- JOSEFSDOTTIR KS, BALDRIDGE MT, KADMON CS & KING KY 2017. Antibiotics impair murine hematopoiesis by depleting the intestinal microbiota. Blood, 129, 729–739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KAHN JD, MILLER PG, SILVER AJ, SELLAR RS, BHATT S, GIBSON C, MCCONKEY M, ADAMS D, MAR B, MERTINS P, FERESHETIAN S, KRUG K, ZHU H, LETAI A, CARR SA, DOENCH J, JAISWAL S & EBERT BL 2018. PPM1D-truncating mutations confer resistance to chemotherapy and sensitivity to PPM1D inhibition in hematopoietic cells. Blood, 132, 1095–1105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KAKIUCHI N & OGAWA S 2021. Clonal expansion in non-cancer tissues. Nat Rev Cancer, 21, 239–256. [DOI] [PubMed] [Google Scholar]
- KAR SP, QUIROS PM, GU M, JIANG T, LANGDON R, IYER V, BARCENA C, VIJAYABASKAR MS, FABRE MA, CARTER P, BURGESS S & VASSILIOU GS 2022. Genome-wide analyses of 200,453 individuals yields new insights into the causes and consequences of clonal hematopoiesis. medRxiv, 2022.01.06.22268846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KESSLER MD, DAMASK A, O’KEEFFE S, VAN METER M, BANERJEE N, SEMRAU S, LI D, WATANABE K, HOROWITZ J, HOUVRAS Y, GILLIES C, MBATCHOU J, WHITE RR, KOSMICKI JA, LEBLANC MG, JONES M, CENTER RG, COLLABORATION G-RD, GLASS DJ, LOTTA LA, CANTOR MN, ATWAL GS, LOCKE AE, FERREIRA MAR, DEERING R, PAULDING C, SHULDINER AR, THURSTON G, SALERNO W, REID JG, OVERTON JD, MARCHINI J, KANG HM, BARAS A, ABECASIS GR & JORGENSON E 2022. Exome sequencing of 628,388 individuals identifies common and rare variant associations with clonal hematopoiesis phenotypes. medRxiv, 2021.12.29.21268342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KIM B, WON D, LEE ST & CHOI JR 2019. Somatic mosaic truncating mutations of PPM1D in blood can result from expansion of a mutant clone under selective pressure of chemotherapy. PLoS One, 14, e0217521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KIM PG, NIROULA A, SHKOLNIK V, MCCONKEY M, LIN AE, SLABICKI M, KEMP JP, BICK A, GIBSON CJ, GRIFFIN G, SEKAR A, BROOKS DJ, WONG WJ, COHEN DN, UDDIN MM, SHIN WJ, PIRRUCCELLO J, TSAI JM, AGRAWAL M, KIEL DP, BOUXSEIN ML, RICHARDS JB, EVANS DM, WEIN MN, CHARLES JF, JAISWAL S, NATARAJAN P & EBERT BL 2021. Dnmt3a-mutated clonal hematopoiesis promotes osteoporosis. J Exp Med, 218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KIMISHIMA Y, MISAKA T, YOKOKAWA T, WADA K, UEDA K, SUGIMOTO K, MINAKAWA K, NAKAZATO K, ISHIDA T, OSHIMA M, KOIDE S, SHIDE K, SHIMODA K, IWAMA A, IKEDA K & TAKEISHI Y 2021. Clonal hematopoiesis with JAK2V617F promotes pulmonary hypertension with ALK1 upregulation in lung neutrophils. Nat Commun, 12, 6177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KING KY & GOODELL MA 2011. Inflammatory modulation of HSCs: viewing the HSC as a foundation for the immune response. Nat Rev Immunol, 11, 685–92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KING KY, HUANG Y, NAKADA D & GOODELL MA 2020. Environmental influences on clonal hematopoiesis. Exp Hematol, 83, 66–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KLEPPE M, COMEN E, WEN HY, BASTIAN L, BLUM B, RAPAPORT FT, KELLER M, GRANOT Z, SOCCI N, VIALE A, YOU D, BENEZRA R, WEIGELT B, BROGI E, BERGER MF, REIS-FILHO JS, LEVINE RL & NORTON L 2015. Somatic mutations in leukocytes infiltrating primary breast cancers. NPJ Breast Cancer, 1, 15005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KON A, YAMAZAKI S, NANNYA Y, KATAOKA K, OTA Y, NAKAGAWA MM, YOSHIDA K, SHIOZAWA Y, MORITA M, YOSHIZATO T, SANADA M, NAKAYAMA M, KOSEKI H, NAKAUCHI H & OGAWA S 2018. Physiological Srsf2 P95H expression causes impaired hematopoietic stem cell functions and aberrant RNA splicing in mice. Blood, 131, 621–635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KOVTONYUK LV, FRITSCH K, FENG X, MANZ MG & TAKIZAWA H 2016. Inflamm-Aging of Hematopoiesis, Hematopoietic Stem Cells, and the Bone Marrow Microenvironment. Front Immunol, 7, 502. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KRISL JC & DOAN VP 2017. Chemotherapy and Transplantation: The Role of Immunosuppression in Malignancy and a Review of Antineoplastic Agents in Solid Organ Transplant Recipients. Am J Transplant, 17, 1974–1991. [DOI] [PubMed] [Google Scholar]
- KRISTINSSON SY, BJORKHOLM M, HULTCRANTZ M, DEROLF AR, LANDGREN O & GOLDIN LR 2011. Chronic immune stimulation might act as a trigger for the development of acute myeloid leukemia or myelodysplastic syndromes. J Clin Oncol, 29, 2897–903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- KRONKE J, BULLINGER L, TELEANU V, TSCHURTZ F, GAIDZIK VI, KUHN MW, RUCKER FG, HOLZMANN K, PASCHKA P, KAPP-SCHWORER S, SPATH D, KINDLER T, SCHITTENHELM M, KRAUTER J, GANSER A, GOHRING G, SCHLEGELBERGER B, SCHLENK RF, DOHNER H & DOHNER K 2013. Clonal evolution in relapsed NPM1-mutated acute myeloid leukemia. Blood, 122, 100–8. [DOI] [PubMed] [Google Scholar]
- KUMAR D, MEHTA A, PANIGRAHI MK, NATH S & SAIKIA KK 2018. DNMT3A (R882) mutation features and prognostic effect in acute myeloid leukemia in Coexistent with NPM1 and FLT3 mutations. Hematol Oncol Stem Cell Ther, 11, 82–89. [DOI] [PubMed] [Google Scholar]
- KUNIMOTO H & NAKAJIMA H 2020. Clonal hematopoiesis: Molecular basis and clinical relevance. Leuk Res, 98, 106457. [DOI] [PubMed] [Google Scholar]
- LACHOWIEZ CA, LOGHAVI S, FURUDATE K, MONTALBAN-BRAVO G, MAITI A, KADIA T, DAVER N, BORTHAKUR G, PEMMARAJU N, SASAKI K, ALVARADO Y, YILMAZ M, SHORT NJ, CHIEN K, OHANIAN M, PIERCE S, PATEL KP, JABBOUR E, RAVANDI F, KANTARJIAN HM, GARCIA-MANERO G, TAKAHASHI K, KONOPLEVA MY & DINARDO CD 2021. Impact of splicing mutations in acute myeloid leukemia treated with hypomethylating agents combined with venetoclax. Blood Adv, 5, 2173–2183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LACONI E, MARONGIU F & DEGREGORI J 2020. Cancer as a disease of old age: changing mutational and microenvironmental landscapes. Br J Cancer, 122, 943–952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LADLE BH, LI KP, PHILLIPS MJ, PUCSEK AB, HAILE A, POWELL JD, JAFFEE EM, HILDEMAN DA & GAMPER CJ 2016. De novo DNA methylation by DNA methyltransferase 3a controls early effector CD8+ T-cell fate decisions following activation. Proc Natl Acad Sci U S A, 113, 10631–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LAUBER C, CORREIA N, TRUMPP A, RIEGER MA, DOLNIK A, BULLINGER L, ROEDER I & SEIFERT M 2020. Survival differences and associated molecular signatures of DNMT3A-mutant acute myeloid leukemia patients. Sci Rep, 10, 12761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LEE J, YOON SR, CHOI I & JUNG H 2019. Causes and Mechanisms of Hematopoietic Stem Cell Aging. Int J Mol Sci, 20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LEE MKS, DRAGOLJEVIC D, BERTUZZO VEIGA C, WANG N, YVAN-CHARVET L & MURPHY AJ 2020. Interplay between Clonal Hematopoiesis of Indeterminate Potential and Metabolism. Trends Endocrinol Metab, 31, 525–535. [DOI] [PubMed] [Google Scholar]
- LEE SE, PARK SS, JEON YW, YOON JH, CHO BS, EOM KS, KIM YJ, LEE S, MIN CK, KIM HJ, CHO SG, KIM DW & LEE JW 2018. Optimal conditioning regimen for haplo-identical stem cell transplantation in adult patients with acquired severe aplastic anemia: Prospective de-escalation study of TBI and ATG dose. Am J Hematol, 93, 1368–1375. [DOI] [PubMed] [Google Scholar]
- LEY TJ, DING L, WALTER MJ, MCLELLAN MD, LAMPRECHT T, LARSON DE, KANDOTH C, PAYTON JE, BATY J, WELCH J, HARRIS CC, LICHTI CF, TOWNSEND RR, FULTON RS, DOOLING DJ, KOBOLDT DC, SCHMIDT H, ZHANG Q, OSBORNE JR, LIN L, O’LAUGHLIN M, MCMICHAEL JF, DELEHAUNTY KD, MCGRATH SD, FULTON LA, MAGRINI VJ, VICKERY TL, HUNDAL J, COOK LL, CONYERS JJ, SWIFT GW, REED JP, ALLDREDGE PA, WYLIE T, WALKER J, KALICKI J, WATSON MA, HEATH S, SHANNON WD, VARGHESE N, NAGARAJAN R, WESTERVELT P, TOMASSON MH, LINK DC, GRAUBERT TA, DIPERSIO JF, MARDIS ER & WILSON RK 2010. DNMT3A mutations in acute myeloid leukemia. N Engl J Med, 363, 2424–33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LI Z, CAI X, CAI CL, WANG J, ZHANG W, PETERSEN BE, YANG FC & XU M 2011. Deletion of Tet2 in mice leads to dysregulated hematopoietic stem cells and subsequent development of myeloid malignancies. Blood, 118, 4509–18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LIBBY P, SIDLOW R, LIN AE, GUPTA D, JONES LW, MOSLEHI J, ZEIHER A, JAISWAL S, SCHULZ C, BLANKSTEIN R, BOLTON KL, STEENSMA D, LEVINE RL & EBERT BL 2019. Clonal Hematopoiesis: Crossroads of Aging, Cardiovascular Disease, and Cancer: JACC Review Topic of the Week. J Am Coll Cardiol, 74, 567–577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LIGGETT LA, CATO LD, WEINSTOCK JS, ZHANG Y, NOURAIE SM, GLADWIN MT, GARRETT ME, ASHLEY-KOCH A, TELEN MJ, CUSTER B, KELLY S, DINARDO CL, SABINO EC, LOUREIRO P, CARNEIRO-PROIETTI AB, MAXIMO C, CONSORTIUM NT-OFPM, REINER AP, ABECASIS GR, WILLIAMS DA, NATARAJAN P, BICK AG & SANKARAN VG 2022. Clonal hematopoiesis in sickle cell disease. J Clin Invest, 132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LIGGETT LA, GALBRAITH MD, SMITH KP, SULLIVAN KD, GRANRATH RE, ENRIQUEZ-ESTRADA B, KINNING KT, SHAW JR, RACHUBINSKI AL, ESPINOSA JM & DEGREGORI J 2021. Precocious clonal hematopoiesis in Down syndrome is accompanied by immune dysregulation. Blood Adv, 5, 1791–1796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LINDSLEY RC, SABER W, MAR BG, REDD R, WANG T, HAAGENSON MD, GRAUMAN PV, HU ZH, SPELLMAN SR, LEE SJ, VERNERIS MR, HSU K, FLEISCHHAUER K, CUTLER C, ANTIN JH, NEUBERG D & EBERT BL 2017. Prognostic Mutations in Myelodysplastic Syndrome after Stem-Cell Transplantation. N Engl J Med, 376, 536–547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LIU Y, ELF SE, MIYATA Y, SASHIDA G, LIU Y, HUANG G, DI GIANDOMENICO S, LEE JM, DEBLASIO A, MENENDEZ S, ANTIPIN J, REVA B, KOFF A & NIMER SD 2009. p53 regulates hematopoietic stem cell quiescence. Cell Stem Cell, 4, 37–48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LOBERG MA, BELL RK, GOODWIN LO, EUDY E, MILES LA, SANMIGUEL JM, YOUNG K, BERGSTROM DE, LEVINE RL, SCHNEIDER RK & TROWBRIDGE JJ 2019. Sequentially inducible mouse models reveal that Npm1 mutation causes malignant transformation of Dnmt3a-mutant clonal hematopoiesis. Leukemia, 33, 1635–1649. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LOPEZ-MOYADO IF, TSAGARATOU A, YUITA H, SEO H, DELATTE B, HEINZ S, BENNER C & RAO A 2019. Paradoxical association of TET loss of function with genome-wide DNA hypomethylation. Proc Natl Acad Sci U S A, 116, 16933–16942. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LUBBERTS E & VAN DEN BERG WB 2003. Cytokines in the pathogenesis of rheumatoid arthritis and collagen-induced arthritis. Adv Exp Med Biol, 520, 194–202. [DOI] [PubMed] [Google Scholar]
- LUM SH, HOENIG M, GENNERY AR & SLATTER MA 2019. Conditioning Regimens for Hematopoietic Cell Transplantation in Primary Immunodeficiency. Curr Allergy Asthma Rep, 19, 52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- LUNDBERG P, KAROW A, NIENHOLD R, LOOSER R, HAO-SHEN H, NISSEN I, GIRSBERGER S, LEHMANN T, PASSWEG J, STERN M, BEISEL C, KRALOVICS R & SKODA RC 2014. Clonal evolution and clinical correlates of somatic mutations in myeloproliferative neoplasms. Blood, 123, 2220–8. [DOI] [PubMed] [Google Scholar]
- MARUSYK A, PORTER CC, ZABEREZHNYY V & DEGREGORI J 2010. Irradiation selects for p53-deficient hematopoietic progenitors. PLoS Biol, 8, e1000324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MAS-PEIRO S, HOFFMANN J, FICHTLSCHERER S, DORSHEIMER L, RIEGER MA, DIMMELER S, VASA-NICOTERA M & ZEIHER AM 2020. Clonal haematopoiesis in patients with degenerative aortic valve stenosis undergoing transcatheter aortic valve implantation. Eur Heart J, 41, 933–939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MATATALL KA, JEONG M, CHEN S, SUN D, CHEN F, MO Q, KIMMEL M & KING KY 2016. Chronic Infection Depletes Hematopoietic Stem Cells through Stress-Induced Terminal Differentiation. Cell Rep, 17, 2584–2595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MAZZONE R, ZWERGEL C, ARTICO M, TAURONE S, RALLI M, GRECO A & MAI A 2019. The emerging role of epigenetics in human autoimmune disorders. Clin Epigenetics, 11, 34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MCKERRELL T, PARK N, MORENO T, GROVE CS, PONSTINGL H, STEPHENS J, UNDERSTANDING SOCIETY SCIENTIFIC G, CRAWLEY C, CRAIG J, SCOTT MA, HODKINSON C, BAXTER J, RAD R, FORSYTH DR, QUAIL MA, ZEGGINI E, OUWEHAND W, VARELA I & VASSILIOU GS 2015. Leukemia-associated somatic mutations drive distinct patterns of age-related clonal hemopoiesis. Cell Rep, 10, 1239–45. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MCNERNEY ME & LE BEAU MM 2018. The Harmful Consequences of Increased Fitness in Hematopoietic Stem Cells. Cell Stem Cell, 23, 634–635. [DOI] [PubMed] [Google Scholar]
- MEISEL M, HINTERLEITNER R, PACIS A, CHEN L, EARLEY ZM, MAYASSI T, PIERRE JF, ERNEST JD, GALIPEAU HJ, THUILLE N, BOUZIAT R, BUSCARLET M, RINGUS DL, WANG Y, LI Y, DINH V, KIM SM, MCDONALD BD, ZURENSKI MA, MUSCH MW, FURTADO GC, LIRA SA, BAIER G, CHANG EB, EREN AM, WEBER CR, BUSQUE L, GODLEY LA, VERDU EF, BARREIRO LB & JABRI B 2018. Microbial signals drive pre-leukaemic myeloproliferation in a Tet2-deficient host. Nature, 557, 580–584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MENCIA-TRINCHANT N, MACKAY MJ, CHIN C, AFSHINNEKOO E, FOOX J, MEYDAN C, BUTLER D, MOZSARY C, VERNICE NA, DARBY C, SCHATZ MC, BAILEY SM, MELNICK AM, GUZMAN ML, BOLTON K, BRAUNSTEIN LZ, GARRETT-BAKELMAN F, LEVINE RL, HASSANE DC & MASON CE 2021. Clonal hematopoiesis before, during, and after human spaceflight. Cell Rep, 34, 108740. [DOI] [PubMed] [Google Scholar]
- METZELER KH, WALKER A, GEYER S, GARZON R, KLISOVIC RB, BLOOMFIELD CD, BLUM W & MARCUCCI G 2012. DNMT3A mutations and response to the hypomethylating agent decitabine in acute myeloid leukemia. Leukemia, 26, 1106–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MEYER SE, QIN T, MUENCH DE, MASUDA K, VENKATASUBRAMANIAN M, ORR E, SUAREZ L, GORE SD, DELWEL R, PAIETTA E, TALLMAN MS, FERNANDEZ H, MELNICK A, LE BEAU MM, KOGAN S, SALOMONIS N, FIGUEROA ME & GRIMES HL 2016. DNMT3A Haploinsufficiency Transforms FLT3ITD Myeloproliferative Disease into a Rapid, Spontaneous, and Fully Penetrant Acute Myeloid Leukemia. Cancer Discov, 6, 501–15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MIDIC D, RINKE J, PERNER F, MULLER V, HINZE A, PESTER F, LANDSCHULZE J, ERNST J, GRUHN B, MATZIOLIS G, HEIDEL FH, HOCHHAUS A & ERNST T 2020. Prevalence and dynamics of clonal hematopoiesis caused by leukemia-associated mutations in elderly individuals without hematologic disorders. Leukemia, 34, 2198–2205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MILLER P, QIAO D, ROJAS-QUINTERO J, HONIGBERG MC, SPERLING AS, GIBSON CJ, BICK AG, NIROULA A, MCCONKEY ME, SANDOVAL B, MILLER B, SHI W, VISWANATHAN K, LEVENTHAL MJ, WERNER L, MOLL M, CADE B, BARR RG, CORREA A, CUPPLES LA, GHARIB SA, JAIN D, GOGARTEN S, LANGE L, LONDON S, MANICHAIKUL A, O’CONNOR G, OELSNER E, REDLINE S, RICH SS, ROTTER JI, RAMACHANDRAN V, YU B, SHOLL LM, NEUBERG D, JAISWAL S, LEVY B, OWEN C, NATARAJAN P, SILVERMAN EK, VAN GALEN P, TESFAIGZI Y, CHO M & EBERT BL 2021a. Association of Clonal Hematopoiesis with Chronic Obstructive Pulmonary Disease. Blood. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MILLER PG, GIBSON CJ, MEHTA A, SPERLING AS, FREDERICK DT, MANOS MP, MIAO B, HACOHEN N, HODI FS, BOLAND GM & EBERT BL 2020. Fitness Landscape of Clonal Hematopoiesis Under Selective Pressure of Immune Checkpoint Blockade. JCO Precis Oncol, 4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MILLER PG, SPERLING AS, BREA EJ, LEICK MB, FELL GG, JAN M, GOHIL SH, TAI YT, MUNSHI NC, WU CJ, NEUBERG DS, MAUS MV, JACOBSON C, GIBSON CJ & EBERT BL 2021b. Clonal hematopoiesis in patients receiving chimeric antigen receptor T-cell therapy. Blood Adv, 5, 2982–2986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MILLER PG & STEENSMA DP 2020. Implications of Clonal Hematopoiesis for Precision Oncology. JCO Precision Oncology, 639–646. [DOI] [PubMed] [Google Scholar]
- MOHRIN M, BOURKE E, ALEXANDER D, WARR MR, BARRY-HOLSON K, LE BEAU MM, MORRISON CG & PASSEGUE E 2010. Hematopoietic stem cell quiescence promotes error-prone DNA repair and mutagenesis. Cell Stem Cell, 7, 174–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MOLENAAR RJ, RADIVOYEVITCH T, NAGATA Y, KHURSHED M, PRZYCHODZEN B, MAKISHIMA H, XU M, BLEEKER FE, WILMINK JW, CARRAWAY HE, MUKHERJEE S, SEKERES MA, VAN NOORDEN CJF & MACIEJEWSKI JP 2018. IDH1/2 Mutations Sensitize Acute Myeloid Leukemia to PARP Inhibition and This Is Reversed by IDH1/2-Mutant Inhibitors. Clin Cancer Res, 24, 1705–1715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MORALES-MANTILLA DE, HUANG X, ERICE P, PORTER P, ZHANG Y, FIGUEROA M, CHANDRA J, KING KY, KHERADMAND F & RODRIGUEZ A 2020. Cigarette Smoke Exposure in Mice using a Whole-Body Inhalation System. J Vis Exp. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MORAN-CRUSIO K, REAVIE L, SHIH A, ABDEL-WAHAB O, NDIAYE-LOBRY D, LOBRY C, FIGUEROA ME, VASANTHAKUMAR A, PATEL J, ZHAO X, PERNA F, PANDEY S, MADZO J, SONG C, DAI Q, HE C, IBRAHIM S, BERAN M, ZAVADIL J, NIMER SD, MELNICK A, GODLEY LA, AIFANTIS I & LEVINE RL 2011. Tet2 loss leads to increased hematopoietic stem cell self-renewal and myeloid transformation. Cancer Cell, 20, 11–24. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MORINISHI L, KOCHANOWSKI K, LEVINE RL, WU LF & ALTSCHULER SJ 2020. Loss of TET2 Affects Proliferation and Drug Sensitivity through Altered Dynamics of Cell-State Transitions. Cell Syst, 11, 86–94 e5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MUPO A, SEILER M, SATHIASEELAN V, PANCE A, YANG Y, AGRAWAL AA, IORIO F, BAUTISTA R, PACHARNE S, TZELEPIS K, MANES N, WRIGHT P, PAPAEMMANUIL E, KENT DG, CAMPBELL PC, BUONAMICI S, BOLLI N & VASSILIOU GS 2017. Hemopoietic-specific Sf3b1-K700E knock-in mice display the splicing defect seen in human MDS but develop anemia without ring sideroblasts. Leukemia, 31, 720–727. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MUZES G, MOLNAR B, TULASSAY Z & SIPOS F 2012. Changes of the cytokine profile in inflammatory bowel diseases. World J Gastroenterol, 18, 5848–61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NACHUN D, LU AT, BICK AG, NATARAJAN P, WEINSTOCK J, SZETO MD, KATHIRESAN S, ABECASIS G, TAYLOR KD, GUO X, TRACY R, DURDA P, LIU Y, JOHNSON C, RICH SS, VAN DEN BERG D, LAURIE C, BLACKWELL T, PAPANICOLAOU GJ, CORREA A, RAFFIELD LM, JOHNSON AD, MURABITO J, MANSON JE, DESAI P, KOOPERBERG C, ASSIMES TL, LEVY D, ROTTER JI, REINER AP, WHITSEL EA, WILSON JG, HORVATH S, JAISWAL S & CONSORTIUM, N. T.-O. F. P. M. 2021. Clonal hematopoiesis associated with epigenetic aging and clinical outcomes. Aging Cell, 20, e13366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NAKAJIMA H & KUNIMOTO H 2014. TET2 as an epigenetic master regulator for normal and malignant hematopoiesis. Cancer Sci, 105, 1093–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NAM AS, DUSAJ N, IZZO F, MURALI R, MYERS RM, MOUHIEDDINE T, SOTELO J, BENBARCHE S, WAARTS M, GAITI F, TAHRI S, LEVINE R, ABDEL-WAHAB O, GODLEY LA, CHALIGNE R, GHOBRIAL I & LANDAU DA 2022. Single-cell multi-omics of human clonal hematopoiesis reveals that DNMT3A R882 mutations perturb early progenitor states through selective hypomethylation. bioRxiv, 2022.01.14.476225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NANGALIA J, MASSIE CE, BAXTER EJ, NICE FL, GUNDEM G, WEDGE DC, AVEZOV E, LI J, KOLLMANN K, KENT DG, AZIZ A, GODFREY AL, HINTON J, MARTINCORENA I, VAN LOO P, JONES AV, GUGLIELMELLI P, TARPEY P, HARDING HP, FITZPATRICK JD, GOUDIE CT, ORTMANN CA, LOUGHRAN SJ, RAINE K, JONES DR, BUTLER AP, TEAGUE JW, O’MEARA S, MCLAREN S, BIANCHI M, SILBER Y, DIMITROPOULOU D, BLOXHAM D, MUDIE L, MADDISON M, ROBINSON B, KEOHANE C, MACLEAN C, HILL K, ORCHARD K, TAURO S, DU MQ, GREAVES M, BOWEN D, HUNTLY BJP, HARRISON CN, CROSS NCP, RON D, VANNUCCHI AM, PAPAEMMANUIL E, CAMPBELL PJ & GREEN AR 2013. Somatic CALR mutations in myeloproliferative neoplasms with nonmutated JAK2. N Engl J Med, 369, 2391–2405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NAWAS MT, SCHETELIG J, DAMM F, LEVINE RL, PERALES MA, GIRALT SA, VANDENBRINK MR, ARCILA ME, ZEHIR A, PAPAEMMANUIL E, KLUSSMEIER A, SCHMIDT AH, MAIWALD S, BOLTON KL & TAMARI R 2021. The clinical implications of clonal hematopoiesis in hematopoietic cell transplantation. Blood Rev, 46, 100744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NEWELL LF, DUNLAP J, GATTER K, BAGBY GC, PRESS RD, COOK RJ, FLETCHER L, LEONARD JT, LEONG KM, BUBALO JS, OLYAEI A, DELOUGHERY TG, MAZIARZ RT, MAYNARD E, ORLOFF SL & ENESTVEDT CK 2021. Graft-versus-host disease after liver transplantation is associated with bone marrow failure, hemophagocytosis, and DNMT3A mutations. Am J Transplant, 21, 3894–3906. [DOI] [PubMed] [Google Scholar]
- NGUYEN YTM, FUJISAWA M, NGUYEN TB, SUEHARA Y, SAKAMOTO T, MATSUOKA R, ABE Y, FUKUMOTO K, HATTORI K, NOGUCHI M, MATSUBARA D, CHIBA S & SAKATA-YANAGIMOTO M 2021. Tet2 deficiency in immune cells exacerbates tumor progression by increasing angiogenesis in a lung cancer model. Cancer Sci. [DOI] [PMC free article] [PubMed] [Google Scholar]
- NI J, HONG J, LONG Z, LI Q, XIA R & ZENG Q 2020. Mutation profile and prognostic relevance in elderly patients with de novo acute myeloid leukemia treated with decitabine-based chemotherapy. Int J Lab Hematol, 42, 849–857. [DOI] [PubMed] [Google Scholar]
- NORELLI M, CAMISA B, BARBIERA G, FALCONE L, PUREVDORJ A, GENUA M, SANVITO F, PONZONI M, DOGLIONI C, CRISTOFORI P, TRAVERSARI C, BORDIGNON C, CICERI F, OSTUNI R, BONINI C, CASUCCI M & BONDANZA A 2018. Monocyte-derived IL-1 and IL-6 are differentially required for cytokine-release syndrome and neurotoxicity due to CAR T cells. Nat Med, 24, 739–748. [DOI] [PubMed] [Google Scholar]
- NOTARO R, CIMMINO A, TABARINI D, ROTOLI B & LUZZATTO L 1997. In vivo telomere dynamics of human hematopoietic stem cells. Proc Natl Acad Sci U S A, 94, 13782–5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- OGAWA S 2016. Clonal hematopoiesis in acquired aplastic anemia. Blood, 128, 337–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- OLSZEWSKI AJ, CHORZALSKA AD, KIM AS, QUESENBERRY PJ, LOPRESTI ML, FENTON MA, REAGAN JL, BUTERA JN, SAHIN I, HAMEL C, ROBISON J, PETERSEN M & DUBIELECKA PM 2019. Clonal haematopoiesis of indeterminate potential among cancer survivors exposed to myelotoxic chemotherapy. Br J Haematol, 186, e31–e35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ORAN B, CHAMPLIN RE, WANG F, TANAKA T, SALIBA RM, AL-ATRASH G, GARCIA-MANERO G, KANTARJIAN H, CAO K, SHPALL EJ, ALOUSI AM, MEHTA RS, POPAT U, FUTREAL A & TAKAHASHI K 2021. Donor clonal hematopoiesis increases risk of acute graft versus host disease after matched sibling transplantation. Leukemia. [DOI] [PubMed] [Google Scholar]
- ORTMANN CA, DORSHEIMER L, ABOU-EL-ARDAT K, HOFFRICHTER J, ASSMUS B, BONIG H, SCHOLZ A, PFEIFER H, MARTIN H, SCHMID T, BRUNE B, SCHEICH S, STEFFEN B, RIEMANN J, HERMANN S, DUKAT A, BUG G, BRANDTS CH, WAGNER S, SERVE H & RIEGER MA 2019. Functional Dominance of CHIP-Mutated Hematopoietic Stem Cells in Patients Undergoing Autologous Transplantation. Cell Rep, 27, 2022–2028 e3. [DOI] [PubMed] [Google Scholar]
- OSTRANDER EL, KRAMER AC, MALLANEY C, CELIK H, KOH WK, FAIRCHILD J, HAUSSLER E, ZHANG CRC & CHALLEN GA 2020. Divergent Effects of Dnmt3a and Tet2 Mutations on Hematopoietic Progenitor Cell Fitness. Stem Cell Reports, 14, 551–560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PAPAEMMANUIL E, GERSTUNG M, MALCOVATI L, TAURO S, GUNDEM G, VAN LOO P, YOON CJ, ELLIS P, WEDGE DC, PELLAGATTI A, SHLIEN A, GROVES MJ, FORBES SA, RAINE K, HINTON J, MUDIE LJ, MCLAREN S, HARDY C, LATIMER C, DELLA PORTA MG, O’MEARA S, AMBAGLIO I, GALLI A, BUTLER AP, WALLDIN G, TEAGUE JW, QUEK L, STERNBERG A, GAMBACORTI-PASSERINI C, CROSS NC, GREEN AR, BOULTWOOD J, VYAS P, HELLSTROM-LINDBERG E, BOWEN D, CAZZOLA M, STRATTON MR, CAMPBELL PJ & CHRONIC MYELOID DISORDERS WORKING GROUP OF THE INTERNATIONAL CANCER GENOME, C. 2013. Clinical and biological implications of driver mutations in myelodysplastic syndromes. Blood, 122, 3616–27; quiz 3699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PARK DJ, KWON A, CHO BS, KIM HJ, HWANG KA, KIM M & KIM Y 2020. Characteristics of DNMT3A mutations in acute myeloid leukemia. Blood Res, 55, 17–26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PASCHKA P, SCHLENK RF, GAIDZIK VI, HABDANK M, KRONKE J, BULLINGER L, SPATH D, KAYSER S, ZUCKNICK M, GOTZE K, HORST HA, GERMING U, DOHNER H & DOHNER K 2010. IDH1 and IDH2 mutations are frequent genetic alterations in acute myeloid leukemia and confer adverse prognosis in cytogenetically normal acute myeloid leukemia with NPM1 mutation without FLT3 internal tandem duplication. J Clin Oncol, 28, 3636–43. [DOI] [PubMed] [Google Scholar]
- PASCHKA P, SCHLENK RF, GAIDZIK VI, HERZIG JK, AULITZKY T, BULLINGER L, SPATH D, TELEANU V, KUNDGEN A, KOHNE CH, BROSSART P, HELD G, HORST HA, RINGHOFFER M, GOTZE K, NACHBAUR D, KINDLER T, HEUSER M, THOL F, GANSER A, DOHNER H & DOHNER K 2015. ASXL1 mutations in younger adult patients with acute myeloid leukemia: a study by the German-Austrian Acute Myeloid Leukemia Study Group. Haematologica, 100, 324–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PIETRAS EM 2017. Inflammation: a key regulator of hematopoietic stem cell fate in health and disease. Blood, 130, 1693–1698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PINCEZ T, LEE SSK, ILBOUDO Y, PREUSS M, PHAM HUNG D’ALEXANDRY D’ORENGIANI A-L, BARTOLUCCI P, GALACTÉROS F, JOLY P, BAUER DE, LOOS RJF, LINDSLEY RC & LETTRE G 2021. Clonal hematopoiesis in sickle cell disease. Blood, 138, 2148–2152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PINHO S & FRENETTE PS 2019. Haematopoietic stem cell activity and interactions with the niche. Nat Rev Mol Cell Biol, 20, 303–320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PORUCHYNSKY MS, KOMLODI-PASZTOR E, TROSTEL S, WILKERSON J, REGAIRAZ M, POMMIER Y, ZHANG X, KUMAR MAITY T, ROBEY R, BUROTTO M, SACKETT D, GUHA U & FOJO AT 2015. Microtubule-targeting agents augment the toxicity of DNA-damaging agents by disrupting intracellular trafficking of DNA repair proteins. Proc Natl Acad Sci U S A, 112, 1571–6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- PRINZING B, ZEBLEY CC, PETERSEN CT, FAN Y, ANIDO AA, YI Z, NGUYEN P, HOUKE H, BELL M, HAYDAR D, BROWN C, BOI SK, ALLI S, CRAWFORD JC, RIBERDY JM, PARK JJ, ZHOU S, VELASQUEZ MP, DERENZO C, LAZZAROTTO CR, TSAI SQ, VOGEL P, PRUETT-MILLER SM, LANGFITT DM, GOTTSCHALK S, YOUNGBLOOD B & KRENCIUTE G 2021. Deleting DNMT3A in CAR T cells prevents exhaustion and enhances antitumor activity. Sci Transl Med, 13, eabh0272. [DOI] [PMC free article] [PubMed] [Google Scholar]
- RAMADAN SM, FOUAD TM, SUMMA V, HASAN S & LO-COCO F 2012. Acute myeloid leukemia developing in patients with autoimmune diseases. Haematologica, 97, 805–17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- RASMUSSEN KD, JIA G, JOHANSEN JV, PEDERSEN MT, RAPIN N, BAGGER FO, PORSE BT, BERNARD OA, CHRISTENSEN J & HELIN K 2015. Loss of TET2 in hematopoietic cells leads to DNA hypermethylation of active enhancers and induction of leukemogenesis. Genes Dev, 29, 910–22. [DOI] [PMC free article] [PubMed] [Google Scholar]
- RAUCH PJ, SILVER AJ, GOPAKUMAR J, MCCONKEY M, SINHA E, FEFER M, SHVARTZ E, SUKHOVA G, LIBBY P, EBERT BL & JAISWAL S 2018. Loss-of-Function Mutations in Dnmt3a and Tet2 Lead to Accelerated Atherosclerosis and Convergent Macrophage Phenotypes in Mice. Blood, 132, 745. [Google Scholar]
- RENNEVILLE A, BOISSEL N, NIBOUREL O, BERTHON C, HELEVAUT N, GARDIN C, CAYUELA JM, HAYETTE S, REMAN O, CONTENTIN N, BORDESSOULE D, PAUTAS C, BOTTON S, REVEL T, TERRE C, FENAUX P, THOMAS X, CASTAIGNE S, DOMBRET H & PREUDHOMME C 2012. Prognostic significance of DNA methyltransferase 3A mutations in cytogenetically normal acute myeloid leukemia: a study by the Acute Leukemia French Association. Leukemia, 26, 1247–54. [DOI] [PubMed] [Google Scholar]
- RIBEIRO AF, PRATCORONA M, ERPELINCK-VERSCHUEREN C, ROCKOVA V, SANDERS M, ABBAS S, FIGUEROA ME, ZEILEMAKER A, MELNICK A, LOWENBERG B, VALK PJ & DELWEL R 2012. Mutant DNMT3A: a marker of poor prognosis in acute myeloid leukemia. Blood, 119, 5824–31. [DOI] [PubMed] [Google Scholar]
- RIDKER PM, MACFADYEN JG, THUREN T, EVERETT BM, LIBBY P, GLYNN RJ & GROUP CT 2017. Effect of interleukin-1beta inhibition with canakinumab on incident lung cancer in patients with atherosclerosis: exploratory results from a randomised, double-blind, placebo-controlled trial. Lancet, 390, 1833–1842. [DOI] [PubMed] [Google Scholar]
- ROBERTSON NA, HILLARY RF, MCCARTNEY DL, TERRADAS-TERRADAS M, HIGHAM J, SPROUL D, DEARY IJ, KIRSCHNER K, MARIONI RE & CHANDRA T 2019. Age-related clonal haemopoiesis is associated with increased epigenetic age. Curr Biol, 29, R786–R787. [DOI] [PubMed] [Google Scholar]
- RODRIGUEZ-MEIRA A, NORFO R, WEN WX, CHÉDEVILLE AL, RAHMAN H, O’SULLIVAN J, WANG G, LOUKA E, KRETZSCHMAR WW, PATERSON A, BRIERLEY C, MARTIN J-E, DEMEULE C, BASHTON M, SOUSOS N, HAMBLIN A, GUERMOUCHE H, PASQUIER F, MARZAC C, GIRODON F, DRUMMOND M, HARRISON C, PLO I, JACOBSEN SEW, PSAILA B, THONGJUEA S, ANTONY-DEBRÉ I & MEAD AJ 2022. Deciphering TP53 mutant Cancer Evolution with Single-Cell Multi-Omics. bioRxiv, 2022.03.28.485984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ROZHOK A & DEGREGORI J 2019. A generalized theory of age-dependent carcinogenesis. Elife, 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- RUSSLER-GERMAIN DA, SPENCER DH, YOUNG MA, LAMPRECHT TL, MILLER CA, FULTON R, MEYER MR, ERDMANN-GILMORE P, TOWNSEND RR, WILSON RK & LEY TJ 2014. The R882H DNMT3A mutation associated with AML dominantly inhibits wild-type DNMT3A by blocking its ability to form active tetramers. Cancer Cell, 25, 442–54. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SANADA F, TANIYAMA Y, MURATSU J, OTSU R, SHIMIZU H, RAKUGI H & MORISHITA R 2018. Source of Chronic Inflammation in Aging. Front Cardiovasc Med, 5, 12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SANDOVAL JE, HUANG YH, MUISE A, GOODELL MA & REICH NO 2019. Mutations in the DNMT3A DNA methyltransferase in acute myeloid leukemia patients cause both loss and gain of function and differential regulation by protein partners. J Biol Chem, 294, 4898–4910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SANO S, OSHIMA K, WANG Y, KATANASAKA Y, SANO M & WALSH K 2018a. CRISPR-Mediated Gene Editing to Assess the Roles of Tet2 and Dnmt3a in Clonal Hematopoiesis and Cardiovascular Disease. Circ Res, 123, 335–341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SANO S, OSHIMA K, WANG Y, MACLAUCHLAN S, KATANASAKA Y, SANO M, ZURIAGA MA, YOSHIYAMA M, GOUKASSIAN D, COOPER MA, FUSTER JJ & WALSH K 2018b. Tet2-Mediated Clonal Hematopoiesis Accelerates Heart Failure Through a Mechanism Involving the IL-1beta/NLRP3 Inflammasome. J Am Coll Cardiol, 71, 875–886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SANO S, WANG Y, OGAWA H, HORITANI K, SANO M, POLIZIO AH, KOUR A, YURA Y, DOVIAK H & WALSH K 2021. TP53-mediated therapy-related clonal hematopoiesis contributes to doxorubicin-induced cardiomyopathy by augmenting a neutrophil-mediated cytotoxic response. JCI Insight, 6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SANO S, WANG Y & WALSH K 2018c. Clonal Hematopoiesis and Its Impact on Cardiovascular Disease. Circ J, 83, 2–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SANO S, WANG Y, YURA Y, SANO M, OSHIMA K, YANG Y, KATANASAKA Y, MIN KD, MATSUURA S, RAVID K, MOHI G & WALSH K 2019. JAK2 (V617F) -Mediated Clonal Hematopoiesis Accelerates Pathological Remodeling in Murine Heart Failure. JACC Basic Transl Sci, 4, 684–697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SAVOLA P, LUNDGREN S, KERANEN MAI, ALMUSA H, ELLONEN P, LEIRISALO-REPO M, KELKKA T & MUSTJOKI S 2018. Clonal hematopoiesis in patients with rheumatoid arthritis. Blood Cancer J, 8, 69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SCHELLER M, LUDWIG AK, GÖLLNER S, ROHDE C, KRÄMER S, STÄBLE S, JANSSEN M, MÜLLER J-A, HE L, BÄUMER N, ARNOLD C, GERS J, SCHÜNUNG M, THIEDE C, NIEDERWIESER C, NIEDERWIESER D, SERVE H, BERDEL WE, THIEM U, HEMMERLING I, LEUSCHNER F, PLASS C, SCHLESNER M, ZAUGG J, MILSOM MD, TRUMPP A, PABST C, LIPKA DB & MÜLLER-TIDOW C 2021. Hotspot DNMT3A mutations in clonal hematopoiesis and acute myeloid leukemia sensitize cells to azacytidine via viral mimicry response. Nature Cancer, 2, 527–544. [DOI] [PubMed] [Google Scholar]
- SCHOEDEL KB, MORCOS MNF, ZERJATKE T, ROEDER I, GRINENKO T, VOEHRINGER D, GOTHERT JR, WASKOW C, ROERS A & GERBAULET A 2016. The bulk of the hematopoietic stem cell population is dispensable for murine steady-state and stress hematopoiesis. Blood, 128, 2285–2296. [DOI] [PubMed] [Google Scholar]
- SHAO L, WANG Y, CHANG J, LUO Y, MENG A & ZHOU D 2013. Hematopoietic stem cell senescence and cancer therapy-induced long-term bone marrow injury. Transl Cancer Res, 2, 397–411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SHIDE K, KAMEDA T, SHIMODA H, YAMAJI T, ABE H, KAMIUNTEN A, SEKINE M, HIDAKA T, KATAYOSE K, KUBUKI Y, YAMAMOTO S, MIIKE T, IWAKIRI H, HASUIKE S, NAGATA K, MARUTSUKA K, IWAMA A, MATSUDA T, KITANAKA A & SHIMODA K 2012. TET2 is essential for survival and hematopoietic stem cell homeostasis. Leukemia, 26, 2216–23. [DOI] [PubMed] [Google Scholar]
- SIDLOW R, LIN AE, GUPTA D, BOLTON KL, STEENSMA DP, LEVINE RL, EBERT BL & LIBBY P 2020. The Clinical Challenge of Clonal Hematopoiesis, a Newly Recognized Cardiovascular Risk Factor. JAMA Cardiol, 5, 958–961. [DOI] [PubMed] [Google Scholar]
- SIGGINS RW, HOSSAIN F, REHMAN T, MELVAN JN, ZHANG P & WELSH DA 2014. Cigarette Smoke Alters the Hematopoietic Stem Cell Niche. Med Sci (Basel), 2, 37–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SORAAS A, MATSUYAMA M, DE LIMA M, WALD D, BUECHNER J, GEDDE-DAHL T, SORAAS CL, CHEN B, FERRUCCI L, DAHL JA, HORVATH S & MATSUYAMA S 2019. Epigenetic age is a cell-intrinsic property in transplanted human hematopoietic cells. Aging Cell, 18, e12897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- STEENSMA DP 2018. Clinical Implications of Clonal Hematopoiesis. Mayo Clin Proc, 93, 1122–1130. [DOI] [PubMed] [Google Scholar]
- STEENSMA DP, BEJAR R, JAISWAL S, LINDSLEY RC, SEKERES MA, HASSERJIAN RP & EBERT BL 2015. Clonal hematopoiesis of indeterminate potential and its distinction from myelodysplastic syndromes. Blood, 126, 9–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- STÖLZEL F, FORDHAM SE, LIN W-Y, BLAIR H, ELSTOB C, NANDANA D, MOHR B, RUHNKE L, KUNADT D, DILL C, ALLSOP D, PIDDOCK R, SOURA E-N, PARK C, FADLY M, RAHMAN T, ALHARBI A, WOBUS M, ALTMANN H, RÖLLIG C, WAGENFÜHR L, JONES GL, MENNE T, JACKSON GH, MARR HJ, FITZGIBBON J, ONEL K, MEGGENDORFER M, HEIDENREICH O, HAFERLACH T, VILLAR S, ARICETA B, DIAZ RA, PROSPER F, MONTESINOS P, MARTINEZ-LOPEZ J, BORNHÄUSER M & ALLAN JM 2021. Biallelic TET2 mutation sensitizes to 5’-azacitidine in acute myeloid leukemia. medRxiv. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SULLIVAN KD, LEWIS HC, HILL AA, PANDEY A, JACKSON LP, CABRAL JM, SMITH KP, LIGGETT LA, GOMEZ EB, GALBRAITH MD, DEGREGORI J & ESPINOSA JM 2016. Trisomy 21 consistently activates the interferon response. Elife, 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SUN D, LUO M, JEONG M, RODRIGUEZ B, XIA Z, HANNAH R, WANG H, LE T, FAULL KF, CHEN R, GU H, BOCK C, MEISSNER A, GOTTGENS B, DARLINGTON GJ, LI W & GOODELL MA 2014. Epigenomic profiling of young and aged HSCs reveals concerted changes during aging that reinforce self-renewal. Cell Stem Cell, 14, 673–88. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SVENSSON EC, MADAR A, CAMPBELL CD, HE Y, SULTAN M, HEALEY ML, XU H, D’ACO K, FERNANDEZ A, WACHE-MAINIER C, LIBBY P, RIDKER PM, BESTE MT & BASSON CT 2022. TET2-Driven Clonal Hematopoiesis and Response to Canakinumab: An Exploratory Analysis of the CANTOS Randomized Clinical Trial. JAMA Cardiol. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SWISHER EM, HARRELL MI, NORQUIST BM, WALSH T, BRADY M, LEE M, HERSHBERG R, KALLI KR, LANKES H, KONNICK EQ, PRITCHARD CC, MONK BJ, CHAN JK, BURGER R, KAUFMANN SH & BIRRER MJ 2016. Somatic Mosaic Mutations in PPM1D and TP53 in the Blood of Women With Ovarian Carcinoma. JAMA Oncol, 2, 370–2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- TARIQ F, ALOBAIDI B, XAVIER M, MCCORKINDALE M, VELTMAN J, ISAACS J, PRATT A, ANDERSON A & COLLIN M 2020. THU0026 CLONAL HAEMATOPOIESIS ASSOCIATED SOMATIC MUTATIONS IN RHEUMATOID ARTHRITIS. Annals of the Rheumatic Diseases, 79, 226. [Google Scholar]
- TONG RS, WONG WH, O’LEARY DR, JAY KS, GRANGE DK, BERMAN J, HITZLER JK & DRULEY TE 2018. The Clonal Hematopoietic Spectrum of Down Syndrome and ML-DS. Blood, 132, 3839–3839. [Google Scholar]
- TURA-CEIDE O, LOBO B, PAUL T, PUIG-PEY R, COLL-BONFILL N, GARCIA-LUCIO J, SMOLDERS V, BLANCO I, BARBERA JA & PEINADO VI 2017. Cigarette smoke challenges bone marrow mesenchymal stem cell capacities in guinea pig. Respir Res, 18, 50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- VAN DEUREN RC, ANDERSSON-ASSARSSON JC, KRISTENSSON FM, STEEHOUWER M, SJÖHOLM K, SVENSSON P-A, PIETERSE M, GILISSEN C, TAUBE M, JACOBSON P, PERKINS R, BRUNNER HG, NETEA MG, PELTONEN M, CARLSSON B, HOISCHEN A & CARLSSON LMS 2021. Expansion of mutation-driven haematopoietic clones is associated with insulin resistance and low HDL-cholesterol in individuals with obesity. bioRxiv, 2021.05.12.443095. [Google Scholar]
- VAN ZEVENTER IA, SALZBRUNN JB, DE GRAAF AO, VAN DER REIJDEN BA, BOEZEN HM, VONK JM, VAN DER HARST P, SCHURINGA JJ, JANSEN JH & HULS G 2021. Prevalence, predictors, and outcomes of clonal hematopoiesis in individuals aged >/=80 years. Blood Adv, 5, 2115–2122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- VANNUCCHI AM, LASHO TL, GUGLIELMELLI P, BIAMONTE F, PARDANANI A, PEREIRA A, FINKE C, SCORE J, GANGAT N, MANNARELLI C, KETTERLING RP, ROTUNNO G, KNUDSON RA, SUSINI MC, LABORDE RR, SPOLVERINI A, PANCRAZZI A, PIERI L, MANFREDINI R, TAGLIAFICO E, ZINI R, JONES A, ZOI K, REITER A, DUNCOMBE A, PIETRA D, RUMI E, CERVANTES F, BAROSI G, CAZZOLA M, CROSS NC & TEFFERI A 2013. Mutations and prognosis in primary myelofibrosis. Leukemia, 27, 1861–9. [DOI] [PubMed] [Google Scholar]
- VENUGOPAL K, FENG Y, SHABASHVILI D & GURYANOVA OA 2021. Alterations to DNMT3A in Hematologic Malignancies. Cancer Res, 81, 254–263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WANG R, GAO X & YU L 2019. The prognostic impact of tet oncogene family member 2 mutations in patients with acute myeloid leukemia: a systematic-review and meta-analysis. BMC Cancer, 19, 389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WANG W, LIU W, FIDLER T, WANG Y, TANG Y, WOODS B, WELCH C, CAI B, SILVESTREROIG C, AI D, YANG YG, HIDALGO A, SOEHNLEIN O, TABAS I, LEVINE RL, TALL AR & WANG N 2018. Macrophage Inflammation, Erythrophagocytosis, and Accelerated Atherosclerosis in Jak2 (V617F) Mice. Circ Res, 123, e35–e47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WANG Y, SANO S, YURA Y, KE Z, SANO M, OSHIMA K, OGAWA H, HORITANI K, MIN KD, MIURA-YURA E, KOUR A, EVANS MA, ZURIAGA MA, HIRSCHI KK, FUSTER JJ, PIETRAS EM & WALSH K 2020. Tet2-mediated clonal hematopoiesis in nonconditioned mice accelerates age-associated cardiac dysfunction. JCI Insight, 5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WANG Y, TONG C, DAI H, WU Z, HAN X, GUO Y, CHEN D, WEI J, TI D, LIU Z, MEI Q, LI X, DONG L, NIE J, ZHANG Y & HAN W 2021. Low-dose decitabine priming endows CAR T cells with enhanced and persistent antitumour potential via epigenetic reprogramming. Nat Commun, 12, 409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WATSON CJ, PAPULA AL, POON GYP, WONG WH, YOUNG AL, DRULEY TE, FISHER DS & BLUNDELL JR 2020. The evolutionary dynamics and fitness landscape of clonal hematopoiesis. Science, 367, 1449–1454. [DOI] [PubMed] [Google Scholar]
- WEINSTOCK JS, GOPAKUMAR J, BURUGULA BB, UDDIN MM, JAHN N, BELK JA, DANIEL B, LY N, MACK TM, LAURIE CA, BROOME JG, TAYLOR KD, GUO X, SINNER MF, VON FALKENHAUSEN AS, KÄÄB S, SHULDINER AR, O’CONNELL JR, LEWIS JP, BOERWINKLE E, BARNES KC, CHAMI N, KENNY EE, LOOS RJ, FORNAGE M, HOU L, LLOYD-JONES DM, REDLINE S, CADE BE, PSATY BM, BIS JC, BRODY JA, SILVERMAN EK, YUN JH, QIAO D, PALMER ND, FREEDMAN BI, BOWDEN DW, CHO MH, DEMEO DL, VASAN RS, YANEK LR, BECKER LC, KARDIA S, PEYSER PA, HE J, RIENSTRA M, HARST PVD, KAPLAN R, HECKBERT SR, SMITH NL, WIGGINS KL, ARNETT DK, IRVIN MR, TIWARI H, CUTLER MJ, KNIGHT S, MUHLESTEIN JB, CORREA A, RAFFIELD LM, GAO Y, ANDRADE MD, ROTTER JI, RICH SS, TRACY RP, KONKLE BA, JOHNSEN JM, WHEELER MM, SMITH JG, MELANDER O, NILSSON PM, CUSTER BS, DUGGIRALA R, CURRAN JE, BLANGERO J, MCGARVEY S, WILLIAMS LK, XIAO S, YANG M, GU CC, CHEN Y-DI, LEE W-J, MARCUS GM, KANE JP, PULLINGER CR, SHOEMAKER MB, DARBAR D, RODEN D, ALBERT C, KOOPERBERG C, ZHOU Y, MANSON JE, DESAI P, JOHNSON A, MATHIAS R, BLACKWELL TW, ABECASIS GR, SMITH AV, KANG HM, SATPATHY AT, et al. 2021. Clonal hematopoiesis is driven by aberrant activation of TCL1A. bioRxiv, 2021.12.10.471810. [Google Scholar]
- WIJEWARDHANE N, DRESSLER L & CICCARELLI FD 2021. Normal Somatic Mutations in Cancer Transformation. Cancer Cell, 39, 125–129. [DOI] [PubMed] [Google Scholar]
- WLODARSKI P, WASIK M, RATAJCZAK MZ, SEVIGNANI C, HOSER G, KAWIAK J, GEWIRTZ AM, CALABRETTA B & SKORSKI T 1998. Role of p53 in hematopoietic recovery after cytotoxic treatment. Blood, 91, 2998–3006. [PubMed] [Google Scholar]
- WOLACH O, SELLAR RS, MARTINOD K, CHERPOKOVA D, MCCONKEY M, CHAPPELL RJ, SILVER AJ, ADAMS D, CASTELLANO CA, SCHNEIDER RK, PADERA RF, DEANGELO DJ, WADLEIGH M, STEENSMA DP, GALINSKY I, STONE RM, GENOVESE G, MCCARROLL SA, ILIADOU B, HULTMAN C, NEUBERG D, MULLALLY A, WAGNER DD & EBERT BL 2018. Increased neutrophil extracellular trap formation promotes thrombosis in myeloproliferative neoplasms. Sci Transl Med, 10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WONG TN, MILLER CA, JOTTE MRM, BAGEGNI N, BATY JD, SCHMIDT AP, CASHEN AF, DUNCAVAGE EJ, HELTON NM, FIALA M, FULTON RS, HEATH SE, JANKE M, LUBER K, WESTERVELT P, VIJ R, DIPERSIO JF, WELCH JS, GRAUBERT TA, WALTER MJ, LEY TJ & LINK DC 2018. Cellular stressors contribute to the expansion of hematopoietic clones of varying leukemic potential. Nat Commun, 9, 455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WONG TN, MILLER CA, KLCO JM, PETTI A, DEMETER R, HELTON NM, LI T, FULTON RS, HEATH SE, MARDIS ER, WESTERVELT P, DIPERSIO JF, WALTER MJ, WELCH JS, GRAUBERT TA, WILSON RK, LEY TJ & LINK DC 2016. Rapid expansion of preexisting nonleukemic hematopoietic clones frequently follows induction therapy for de novo AML. Blood, 127, 893–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WONG TN, RAMSINGH G, YOUNG AL, MILLER CA, TOUMA W, WELCH JS, LAMPRECHT TL, SHEN D, HUNDAL J, FULTON RS, HEATH S, BATY JD, KLCO JM, DING L, MARDIS ER, WESTERVELT P, DIPERSIO JF, WALTER MJ, GRAUBERT TA, LEY TJ, DRULEY T, LINK DC & WILSON RK 2015. Role of TP53 mutations in the origin and evolution of therapy-related acute myeloid leukaemia. Nature, 518, 552–555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WONG WJ, EMDIN C, BICK A, ZEKAVAT SM, NIROULA A, PIRRUCCELLO J, DICHTEL L, GRIFFIN G, UDDIN MM, GIBSON CJ, KOVALCIK V, LIN AE, MCCONKEY ME, VROMMAN A, SELLAR RS, KIM PG, AGRAWAL M, WEINSTOCK J, LONG MT, YU B, BANERJEE R, NICHOLLS RC, KELLY M, LOH P-R, MCCARROLL S, BOERWINKLE E, VASAN R, JAISWAL S, JOHNSON A, CHUNG RT, COREY K, LEVY D, BALLANTYNE C, GROUP, N. T. H. W., EBERT BL & NATARAJAN P 2022. Clonal hematopoiesis and risk of chronic liver disease. medRxiv, 2022.01.17.22269409. [Google Scholar]
- WU D, HU D, CHEN H, SHI G, FETAHU IS, WU F, RABIDOU K, FANG R, TAN L, XU S, LIU H, ARGUETA C, ZHANG L, MAO F, YAN G, CHEN J, DONG Z, LV R, XU Y, WANG M, YE Y, ZHANG S, DUQUETTE D, GENG S, YIN C, LIAN CG, MURPHY GF, ADLER GK, GARG R, LYNCH L, YANG P, LI Y, LAN F, FAN J, SHI Y & SHI YG 2018. Glucose-regulated phosphorylation of TET2 by AMPK reveals a pathway linking diabetes to cancer. Nature, 559, 637–641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- WU WS, HEINRICHS S, XU D, GARRISON SP, ZAMBETTI GP, ADAMS JM & LOOK AT 2005. Slug antagonizes p53-mediated apoptosis of hematopoietic progenitors by repressing puma. Cell, 123, 641–53. [DOI] [PubMed] [Google Scholar]
- XIE M, LU C, WANG J, MCLELLAN MD, JOHNSON KJ, WENDL MC, MCMICHAEL JF, SCHMIDT HK, YELLAPANTULA V, MILLER CA, OZENBERGER BA, WELCH JS, LINK DC, WALTER MJ, MARDIS ER, DIPERSIO JF, CHEN F, WILSON RK, LEY TJ & DING L 2014. Age-related mutations associated with clonal hematopoietic expansion and malignancies. Nat Med, 20, 1472–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YAMASHITA M & PASSEGUE E 2019. TNF-alpha Coordinates Hematopoietic Stem Cell Survival and Myeloid Regeneration. Cell Stem Cell, 25, 357–372 e7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YAN XJ, XU J, GU ZH, PAN CM, LU G, SHEN Y, SHI JY, ZHU YM, TANG L, ZHANG XW, LIANG WX, MI JQ, SONG HD, LI KQ, CHEN Z & CHEN SJ 2011. Exome sequencing identifies somatic mutations of DNA methyltransferase gene DNMT3A in acute monocytic leukemia. Nat Genet, 43, 309–15. [DOI] [PubMed] [Google Scholar]
- YANG L, RAU R & GOODELL MA 2015. DNMT3A in haematological malignancies. Nat Rev Cancer, 15, 152–65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YANG L, RODRIGUEZ B, MAYLE A, PARK HJ, LIN X, LUO M, JEONG M, CURRY CV, KIM SB, RUAU D, ZHANG X, ZHOU T, ZHOU M, REBEL VI, CHALLEN GA, GOTTGENS B, LEE JS, RAU R, LI W & GOODELL MA 2016. DNMT3A Loss Drives Enhancer Hypomethylation in FLT3-ITD-Associated Leukemias. Cancer Cell, 30, 363–365. [DOI] [PubMed] [Google Scholar]
- YANG Y, DAI Y, YANG X, WU S & WANG Y 2021. DNMT3A Mutation-Induced CDK1 Overexpression Promotes Leukemogenesis by Modulating the Interaction between EZH2 and DNMT3A. Biomolecules, 11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YOKOKAWA T, MISAKA T, KIMISHIMA Y, WADA K, MINAKAWA K, SUGIMOTO K, ISHIDA T, MORISHITA S, KOMATSU N, IKEDA K & TAKEISHI Y 2021. Crucial role of hematopoietic JAK2 V617F in the development of aortic aneurysms. Haematologica, 106, 1910–1922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YOSHIDA K, SANADA M, SHIRAISHI Y, NOWAK D, NAGATA Y, YAMAMOTO R, SATO Y, SATO-OTSUBO A, KON A, NAGASAKI M, CHALKIDIS G, SUZUKI Y, SHIOSAKA M, KAWAHATA R, YAMAGUCHI T, OTSU M, OBARA N, SAKATA-YANAGIMOTO M, ISHIYAMA K, MORI H, NOLTE F, HOFMANN WK, MIYAWAKI S, SUGANO S, HAFERLACH C, KOEFFLER HP, SHIH LY, HAFERLACH T, CHIBA S, NAKAUCHI H, MIYANO S & OGAWA S 2011. Frequent pathway mutations of splicing machinery in myelodysplasia. Nature, 478, 64–9. [DOI] [PubMed] [Google Scholar]
- YOSHIZATO T, DUMITRIU B, HOSOKAWA K, MAKISHIMA H, YOSHIDA K, TOWNSLEY D, SATO-OTSUBO A, SATO Y, LIU D, SUZUKI H, WU CO, SHIRAISHI Y, CLEMENTE MJ, KATAOKA K, SHIOZAWA Y, OKUNO Y, CHIBA K, TANAKA H, NAGATA Y, KATAGIRI T, KON A, SANADA M, SCHEINBERG P, MIYANO S, MACIEJEWSKI JP, NAKAO S, YOUNG NS & OGAWA S 2015. Somatic Mutations and Clonal Hematopoiesis in Aplastic Anemia. N Engl J Med, 373, 35–47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YOUNG AL, TONG RS, BIRMANN BM & DRULEY TE 2019. Clonal hematopoiesis and risk of acute myeloid leukemia. Haematologica, 104, 2410–2417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YU B, ROBERTS MB, RAFFIELD LM, ZEKAVAT SM, NGUYEN NQH, BIGGS ML, BROWN MR, GRIFFIN G, DESAI P, CORREA A, MORRISON AC, SHAH AM, NIROULA A, UDDIN MM, HONIGBERG MC, EBERT BL, PSATY BM, WHITSEL EA, MANSON JE, KOOPERBERG C, BICK AG, BALLANTYNE CM, REINER AP, NATARAJAN P, EATON CB, NATIONAL HEART L & BLOOD INSTITUTE TC 2021. Supplemental Association of Clonal Hematopoiesis With Incident Heart Failure. J Am Coll Cardiol, 78, 42–52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YUAN XQ, PENG L, ZENG WJ, JIANG BY, LI GC & CHEN XP 2016a. DNMT3A R882 Mutations Predict a Poor Prognosis in AML: A Meta-Analysis From 4474 Patients. Medicine (Baltimore), 95, e3519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YUAN XQ, ZHANG DY, YAN H, YANG YL, ZHU KW, CHEN YH, LI X, YIN JY, LI XL, ZENG H & CHEN XP 2016b. Evaluation of DNMT3A genetic polymorphisms as outcome predictors in AML patients. Oncotarget, 7, 60555–60574. [DOI] [PMC free article] [PubMed] [Google Scholar]
- YURA Y, MIURA-YURA E, KATANASAKA Y, MIN KD, CHAVKIN N, POLIZIO AH, OGAWA H, HORITANI K, DOVIAK H, EVANS MA, SANO M, WANG Y, BOROVIAK K, PHILIPPOS G, DOMINGUES AF, VASSILIOU G, SANO S & WALSH K 2021. The Cancer Therapy-Related Clonal Hematopoiesis Driver Gene Ppm1d Promotes Inflammation and Non-Ischemic Heart Failure in Mice. Circ Res, 129, 684–698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZAJKOWICZ A, BUTKIEWICZ D, DROSIK A, GIGLOK M, SUWINSKI R & RUSIN M 2015. Truncating mutations of PPM1D are found in blood DNA samples of lung cancer patients. Br J Cancer, 112, 1114–20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZEKAVAT SM, LIN SH, BICK AG, LIU A, PARUCHURI K, WANG C, UDDIN MM, YE Y, YU Z, LIU X, KAMATANI Y, BHATTACHARYA R, PIRRUCCELLO JP, PAMPANA A, LOH PR, KOHLI P, MCCARROLL SA, KIRYLUK K, NEALE B, IONITA-LAZA I, ENGELS EA, BROWN DW, SMOLLER JW, GREEN R, KARLSON EW, LEBO M, ELLINOR PT, WEISS ST, DALY MJ, BIOBANK JAPAN P, FINNGEN C, TERAO C, ZHAO H, EBERT BL, REILLY MP, GANNA A, MACHIELA MJ, GENOVESE G & NATARAJAN P 2021a. Hematopoietic mosaic chromosomal alterations increase the risk for diverse types of infection. Nat Med, 27, 1012–1024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZEKAVAT SM, VIANA-HUETE V, ZURIAGA MA, UDDIN MM, TRINDER M, PARUCHURI K, MATESANZ N, ZORITA V, FERRER-PÉREZ A, AMORÓS-PÉREZ M, DAMRAUER SM, BALLANTYNE CM, NIROULA A, GIBSON CJ, PIRRUCCELLO J, GRIFFIN G, EBERT BL, LIBBY P, FUSTER V, ZHAO H, NATARAJAN P, BICK AG, FUSTER JJ & KLARIN D 2021b. TP53-mediated clonal hematopoiesis confers increased risk for incident peripheral artery disease. medRxiv. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZENG H, HE H, GUO L, LI J, LEE M, HAN W, GUZMAN AG, ZANG S, ZHOU Y, ZHANG X, GOODELL MA, KING KY, SUN D & HUANG Y 2019. Antibiotic treatment ameliorates Ten-eleven translocation 2 (TET2) loss-of-function associated hematological malignancies. Cancer Lett, 467, 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZHANG CR, OSTRANDER EL, KUKHAR O, MALLANEY C, SUN J, HAUSSLER E, CELIK H, KOH WK, KING KY, GONTARZ P & CHALLEN GA 2022. Txnip Enhances Fitness of Dnmt3a-Mutant Hematopoietic Stem Cells via p21. Blood Cancer Discov, OF1–OF20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZHANG CRC, NIX D, GREGORY M, CIORBA MA, OSTRANDER EL, NEWBERRY RD, SPENCER DH & CHALLEN GA 2019. Inflammatory cytokines promote clonal hematopoiesis with specific mutations in ulcerative colitis patients. Exp Hematol, 80, 36–41 e3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZINK F, STACEY SN, NORDDAHL GL, FRIGGE ML, MAGNUSSON OT, JONSDOTTIR I, THORGEIRSSON TE, SIGURDSSON A, GUDJONSSON SA, GUDMUNDSSON J, JONASSON JG, TRYGGVADOTTIR L, JONSSON T, HELGASON A, GYLFASON A, SULEM P, RAFNAR T, THORSTEINSDOTTIR U, GUDBJARTSSON DF, MASSON G, KONG A & STEFANSSON K 2017. Clonal hematopoiesis, with and without candidate driver mutations, is common in the elderly. Blood, 130, 742–752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ZIONI N, CHAPAL ILANI N, PETROVICH-KOPITMAN E, SAÇMA M, GEIGER H, SCHELLER M, MUELLER-TIDOW C, KAUSHANSKY N & SHLUSH LI 2021. Fatty Bone Marrow Positively Selects Pre-Leukemic HSPCs with a DNMT3A-mutation. Blood, 138, 596–596.34436528 [Google Scholar]