

Abstract
Multiple nutritional deficiencies (MND) confound studies designed to assess the role of a single nutrient in contributing to the initiation and progression of disease states. Despite the perception of many healthcare practitioners, up to 25% of Americans are deficient in five-or-more essential nutrients. Stress associated with the COVID-19 pandemic further increases the prevalence of deficiency states. Viral infections compete for crucial nutrients with immune cells. Viral replication and proliferation of immunocompetent cells critical to the host response require these essential nutrients, including zinc. Clinical studies have linked levels of more than 22 different dietary components to the likelihood of COVID-19 infection and the severity of the disease. People at higher risk of infection due to MND are also more likely to have long-term sequelae, known as Long COVID.
Keywords: Multiple nutritional deficiencies, Lymphocyte proliferation assay, Myeloperoxidase, Glutathione peroxidase isozyme 4, NADPH oxidase 2
Introduction
Micronutrient depletion is commonly associated with stress, aging, and infectious disease. Nutrients with solid evidence for stress-induced deficiency include magnesium, zinc, iron, calcium, and niacin (Lopresti 2019). An example of the reverse, deficiency-induced stress, includes cobalamin. On average, 41% of Americans have insufficient levels of cobalamin (vitamin B12), as judged by a lymphocyte proliferation assay (LPA) (Bucci 1994). Low maternal plasma B12 and high folate during gestation predict increased HPA-axis stress in the offspring. Higher cortisol responses to stress in children resulting from such pregnancies have been documented during adolescence (median age = 13.6 years) by Krishnaveni et al. (2020). With a higher tendency for stress, these individuals have a greater chance of developing MND. Figure 1 illustrates this interdependence between stress-related deficit in micronutrients and deficiency-induced stress.
Fig. 1.
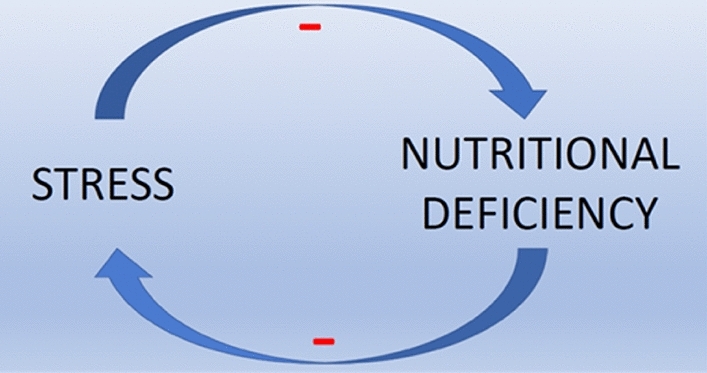
Cyclic nature of stress and nutritional deficiency
What are nutritional deficiencies and how are they determined?
Conventional nutritional science estimates less than 5.7% of people (USA) lack two-or-more essential nutrients (Bird et al. 2017). However, this assessment depends on the criteria employed to define an insufficient level of an essential dietary component. For many nutrients with pleiotropic effects on different organ systems and multiple biochemical pathways, disease in the whole organism manifests in more than one way and at more than one intake level. For example, the dietary requirement for vitamin D to prevent rickets is substantially lower than the requirement to reduce the risk of cancer or for proper immune system functioning (Holick 2020).
Nutritional deficiency based on the LPA
William Shive, at the Clayton Foundation Biochemical Institute (CFBI), developed a robust method (LPA) for assessing individual deficiencies of many micronutrients based on lymphocyte proliferation (Shive et al. 1986). The CFBI at the University of Texas in Austin has a long and distinguished history of nutritional research. Roger John Williams founded the CFBI. He was a nutritional research pioneer who discovered vitamins B5 (pantothenic acid), B6 (pyridoxine), B9 (folate), LA (lipoic acid), and avidin. Shive’s method for rapidly determining deficiency states caused by diet or individual genetic variation finally addresses the concept of ‘biochemical individuality’ initially proposed by Roger J. Williams (1956). LPA determined the incidence of 19 different nutrient disorders in 833 Americans (Bucci 1994). Based on these data, 98% of Americans have at least one problem; 83% have two-or-more insufficiencies, and 25% have five-or-more nutritional deficiencies (Bucci 1994). MND would predict that a single supplement administered to a patient with multiple inadequacies may not correct disease symptoms resulting from the lack of several. Clinical testing of monotherapy is likely to be confounded by other coexisting deficiencies. Correlation of disease with a selected, single inadequacy is expected to have substantial scatter due to the possibility of MND in many (83%) patients.
Association of nutritional disorders with COVID-19 disease
Figure 2 illustrates the relationship between COVID-19 disease, the incidence of nutrient disorders, and the effect of nutritional supplements. The first two columns in this figure include nutrients or conditions (i.e., fructose intolerance and insulin dysfunction) known to be assessed by Shive’s LPA. The incidence of disorders determined by LPA in a group of 833 Americans, expressed as a percentage, is included immediately following the name of the nutrient or condition (Bucci 1994). The third column of Fig. 2 contains nutrients, where LPA has not assessed the deficiency prevalence. Column three includes available data for the frequency of insufficiency determined by other methods. Based on a comprehensive evaluation of peer-reviewed literature in April 2022, those nutrients that had one-or-more credible publications linking them with the risk and severity of COVID-19 are indicated by a green checkmark ( ). Those nutrients that exacerbate the risk of COVID-19 disease or are toxic in excess have a red checkmark (
). Those nutrients that exacerbate the risk of COVID-19 disease or are toxic in excess have a red checkmark ( ). Cobalamin (vitamin B12) is particularly noteworthy, since the incidence of B12 deficiency correlates with COVID-19 disease (Wee 2021), but B12 supplements increase the risk of disease severity (Dalbeni et al. 2021). Most nutrients under consideration, 22 out of 29, have evidence to support the notion that their levels affect COVID-19 disease. Two micronutrients, zinc and selenium, are known to be toxic in excess (Fosmire 1990; Barceloux 1999).
). Cobalamin (vitamin B12) is particularly noteworthy, since the incidence of B12 deficiency correlates with COVID-19 disease (Wee 2021), but B12 supplements increase the risk of disease severity (Dalbeni et al. 2021). Most nutrients under consideration, 22 out of 29, have evidence to support the notion that their levels affect COVID-19 disease. Two micronutrients, zinc and selenium, are known to be toxic in excess (Fosmire 1990; Barceloux 1999).
Fig. 2.

Relationship between nutrient disorders and COVID-19 disease
Literature relating different nutrients with COVID-19 disease: cobalamin/vitamin B12 (Wee 2021; Elham 2021; Galmés et al. 2020; Dalbeni et al. 2021; Clemente-Suárez et al. 2021; van Kempen and Deixler 2021); calcium (Zhou et al. 2020; Osman et al. 2021; Yang et al. 2021; Zeng et al. 2021; El-Kurdi et al. 2020); zinc (Galmés et al. 2020; Dharmalingam et al. 2021; Zeng et al. 2021; Heller et al. 2021; Maares et al. 2022; Notz et al. 2021; Du Laing et al. 2021; Chillon et al. 2022); thiamin/vitamin B1 (Al Sulaiman et al. 2021; Branco de Oliveira et al. 2021); pyridoxine/vitamin B6 (Kumrungsee et al. 2020; Galmés et al. 2020); fructose (Vazirani 2021); magnesium (Jose et al. 2021; Zeng et al. 2021; Trapani et al. 2022; Eskander and Razzaque 2022; van Kempen and Deixler 2021); glucose (Shauly-Aharonov et al. 2021; Wang et al. 2021; Ardestani and Azizi 2021); niacinamide/vitamin B3 (Raines et al. 2020); riboflavin/vitamin B2 (Akasov et al. 2022); oleic acid (Clemente-Suárez et al. 2021); glutamine (Matsuyama et al. 2021; Cengiz et al. 2020; Soliman et al. 2022); inositol (Espinola et al. 2021; Bizzarri et al. 2020; Laganà et al. 2020); choline (Olivari et al. 2020; Freedman et al. 2020; Bizzarri et al. 2020); selenium (Galmés et al. 2020; Im et al. 2020; Dharmalingam et al. 2021; Clemente-Suárez et al. 2021; Fakhrolmobasheri et al. 2022; Khatiwada and Subedi 2021; Schomburg 2022; Heller et al. 2021; Moghaddam et al. 2020; Notz et al. 2021; Du Laing et al. 2021; Zhang et al. 2020; Skesters et al. 2022); cholecalciferol/vitamin D (Galmés et al. 2020; Grant et al. 2020; Im et al. 2020; Clemente-Suárez et al. 2021; van Kempen and Deixler 2021; Weir et al. 2020; Kaya et al. 2021); iron (Galmés et al. 2020; Nai et al. 2021; Habib et al. 2021; Drakesmith et al. 2021; Taneri et al. 2020; Dharmalingam et al. 2021; Zeng et al. 2021; Clemente-Suárez et al. 2021); taurine (Iwegbulem et al. 2022; van Eijk et al. 2022); phosphorus (van Kempen and Deixler 2021; Seers and Davenport 2020); ascorbate/vitamin C (Liu et al. 2020; Hemilä and de Man 2021; Holford et al. 2020; Galmés et al. 2020; Clemente-Suárez et al. 2021; Biancatelli et al. 2020); bioflavonoids (Derosa et al. 2021; Biancatelli et al. 2020; Españo et al. 2021; DI Pierro et al. 2021; Landis et al. 2022); N-acetyl cysteine (Shi and Puyo 2020; Assimakopoulos et al. 2021; Mohanty et al. 2021; De Flora et al. 2020; Zhou et al. 2021; Jorge-Aarón and Rosa-Ester 2020; Wong et al. 2021; Andreou et al. 2020; de Alencar et al. 2021; Kapur et al. 2022); ω-3 fatty acids (Clemente-Suárez et al. 2021; Das 2020; Goc et al. 2021); retinoic acid/vitamin A (Galmés et al. 2020); folate (Galmés et al. 2020; Meisel et al. 2021; Doaei et al. 2021); copper (Galmés et al. 2020; Dharmalingam et al. 2021; Zeng et al. 2021; Andreou et al. 2020); iodine (Fröhlich and Wahl 2021); nickel (Dharmalingam et al. 2021); lithium (Dharmalingam et al. 2021); chromium (Zeng et al. 2021); manganese (Zeng et al. 2021); albumin (El-Kurdi et al. 2020); potassium (Liu et al. 2021; Alfano et al. 2021; Noori et al. 2022); sodium (Gheorghe et al. 2021).
The contribution of zinc and selenium deficiency to morbidity and mortality from COVID-19 disease
There is a clear correlation between zinc or selenium deficiency with morbidity and mortality from COVID-19 infection (Heller et al. 2021; Moghaddam et al. 2020; Maares et al. 2022; Notz et al. 2021; Du Laing et al. 2021). The humoral response, production of antibodies after SARS-CoV-2 vaccination, also correlates with free zinc, but not total serum zinc concentrations (Chillon et al. 2022). Although correlation does not necessarily prove causation, the essential role of zinc in the production of 10% of all proteins (Read et al. 2019) and of selenium in DNA biosynthesis or alkyl-hydroperoxide detoxification (Muller et al. 2007) would argue that there is a mechanistic explanation.
Zinc is a gatekeeper of immune function (Wessels et al. 2017). The knowledge that insufficient dietary zinc produces an elevated risk of infection dates to the early 1960s (Prasad 2001). Evidence suggests zinc deficiency causes cell-mediated immune dysfunction (cytokine storm), cognitive impairment (brain fog), and depression (Prasad 2013; Nowak et al. 2005). Zinc deficiency also is associated with retarded wound healing (Kogan et al. 2017), various types of cancer risk (Skrajnowska and Bobrowska-Korczak 2019), and heart failure (Rosenblum et al. 2020). Much of the pathology associated with suboptimal zinc is also associated with the sequelae of COVID-19, known as Long COVID.
Zinc and COVID-induced loss of smell and taste
Zinc supplements post-infection reduce the duration of anosmia, but not all symptoms resulting from SARS-CoV-2 infection (Abdelmaksoud et al. 2021). A likely explanation for the effect of zinc in shortening the duration of anosmia during COVID-19 disease involves the initial site of infection and the role that zinc plays in olfaction. The nasopharyngeal area is a common site for the first encounter with SARS-CoV-2. Loss of olfaction is an expected early symptom of COVID-19. As SARS-CoV-2 proliferates, it would result in local depletion of zinc, since many proteins (10%) and viral replication both require zinc ions. Localized zinc depletion would deprive olfaction of this essential metal ion. There are two steps in the detection of odorant molecules that require zinc. The first is the cleavage of conjugates that mask odorant detection, and the second is the conjugation of odorant molecules that terminates detection and prevents saturation of the olfactory receptor. The gene for the odorant metabolizing enzyme UDP-glucuronosyltransferase (UGT), elevated during zinc deficiency (tom Dieck et al. 2003), is linked to COVID-induced loss of smell (Shelton et al.2022). The odorant metabolizing enzymes, UGT, and β-glucuronidase are essential for the sense of smell (Neiers et al. 2021). Zinc activates the membrane-associated β-glucuronidase (Yamaguchi et al. 1990). High zinc concentrations can completely inactivate UGT and β-glucuronidase (Schöllhammer et al. 1975). These effects of zinc on β-glucuronidase and UGT can explain the paradoxical association of anosmia with zinc deficiency (Pisano and Hilas 2016) and high-zinc resulting from the intranasal administration of zinc solutions (Alexander and Davidson 2006). Anosmia and dysgeusia are commonly associated with COVID-19 (Zahra et al. 2020) and dietary zinc deficiency (Pisano and Hilas 2016).
Selenium has a dual role in the progression of COVID-19 disease
Keshan disease is a highly fatal disorder first recognized in 1935 and ultimately linked to the consumption of crops grown in low-selenium soils (Chen 2012). Excess dietary selenium is also toxic. The effects of chronic exposure resulting from the consumption of crops grown in high-selenium soils (Lower Cambrian outcrop areas in Southern Shaanxi, China) are documented (Barceloux 1999; Du et al. 2018). Zhang et al. (2020) have pointed out that infections by SARS-CoV-2 and other viruses are more prevalent in those regions of China with low selenium soil content. Selenium deficiency also results in congestive cardiomyopathy, heart failure, cardiomegaly, increased risk of cancer, infection, inflammatory disease, diabetes, retarded wound healing, and infertility, while excess selenium results in brittle hair and brittle, thickened, stratified nails or loss of both hair and nails (Hariharan and Dharmaraj 2020; Fairweather-Tait et al. 2011). In the extreme, selenium deficiency or excess can result in death.
Figure 3 lists three selenium-containing enzymes (in red) essential to life based on gene-knockout experiments in mice (Muller et al. 2007). Glutathione peroxidase isozyme 4 (GPX4) is an enzyme capable of detoxifying alkyl-hydroperoxides, membrane-bound phospholipid-hydroperoxides, and dioxetanes, like those formed during the immune response to pathogens with reactive oxygen species (Belikov et al. 2015; Di Mascio et al. 2019). Thioredoxin reductase is essential for activating ribonucleotide reductase to form the deoxynucleotide precursors for DNA biosynthesis and T-cell proliferation (Sengupta and Holmgren 2014; Muri et al. 2018). Selenium deficiency would have a differential effect on the immune response and SARS-CoV-2 proliferation, since replication of the virus involves RNA biosynthesis (RNA-dependent RNA polymerase) in contrast to lymphocytes and polymorphonuclear neutrophils (PMN), which rely on DNA biosynthesis (Yin et al. 2020). Because of this, low selenium produces a disadvantage to the immune response relative to SARS-CoV-2 replication. Selenium deficiency would also increase the collateral damage to host cells during infection by limiting the host-protective effect of GPX4 (Martinez et al. 2021).
Fig. 3.

Enzymes essential to life; their gene designation; and phenotypic outcome of gene-knockout experiments in mice. Enzymes in red require an essential selenocysteine
The first stage of immune response: proliferation
There are three stages of the immune response to infection by a pathogen, illustrated in Fig. 4. In the first stage, immunocompetent cells that can respond to viral antigens must proliferate to mount an adequate response to invaders that are also increasing in number. The role of many essential nutrients is evident in the first stage of the immune response. Zinc is an integral component of a wide range of proteins (10% overall) necessary for cell replication and is specifically involved in human DNA polymerases and other essential DNA-binding enzymes (Evanics et al. 2003; Petrucco and Percudani 2008). Selenium is necessary for the biosynthesis of deoxynucleotides for DNA replication and the proliferation of immunocompetent cells (Sengupta and Holmgren 2014; Muri et al. 2018). Glutamine is a regulator of DNA replication and cell multiplication and is an essential component of purine and pyrimidine biosynthesis (Zetterberg and Engström 1981; Cory and Cory 2006). Phosphate depletion markedly inhibits DNA synthesis and cell division (Alexander et al. 2022; Engström and Zetterberg 1983; Houillier and Salles 2021). Many other essential nutrients are required in adequate amounts to support the rapid response of the immune system to a pathogen threat. It is not surprising that most of the commonly encountered deficiencies (Fig. 2) have evidence linking them to the development of severe COVID-19 disease. Without an adequate increase in the pathogen-specific and auxiliary cells of the immune system, it will be overwhelmed by a rapidly increasing pathogen.
Fig. 4.

Overall summary of nutrient roles in pathogen response
The second stage of response: innate immunity
The second stage of immune response is the direct destruction of the pathogen by polymorphonuclear neutrophils (PMN) and T lymphocytes. This cytotoxic defense system exposes the pathogen to reactive oxygen species (ROS). Myeloperoxidase (MPO) plays a role in the second stage of the immune response by producing hypochlorous acid/hypochlorite (ClO−, pKA = 7.5), singlet oxygen, and N-chlorotaurine (tau-Cl) (Aratani 2018; Kanofsky 1989; Schuller-Levis and Park 2003). In addition to the production of ROS localized in regions of infection, PMN, in combination with T lymphocytes, use a variety of cytotoxic mechanisms to fight infection, including the release of antimicrobial peptides and the expulsion of their nuclear contents to form ‘extracellular traps’ (Mayadas et al. 2014; Reusch et al. 2021; Witter et al. 2016). Although MPO is not essential to the innate immune response, MPO, PMN, and monocytes play a significant role in the pathology associated with severe infection (Shrivastava et al. 2021; Matsushita et al. 2015; Peyneau et al. 2022). ROS produced by MPO are not an essential component of the host defense response, since genetic deficiency in MPO only renders an individual less capable of clearing pathogenic organisms. However, it does not significantly increase the risk of severe infection unless the individual has diabetes (Nauseef 1988). Severe COVID-19 disease is associated with elevated levels of MPO (Shrivastava et al. 2021). The T cell GPX4 is essential for preventing T cell death by MPO-induced ferroptosis with concomitant loss of effective immune response (Matsushita et al. 2015).
MPO is a heme–iron-containing enzyme implicated in the pathology of chronic inflammation that, in the presence of hydrogen peroxide and chloride ions, will destroy its heme prosthetic group to generate free iron (Maitra et al. 2013). NADPH oxidase 2 (NOX2) requires essential heme–iron, FAD (riboflavin/vitamin B2 derived), and NADPH (niacinamide/vitamin B3 derived) cofactors to produce superoxide (Magnani et al. 2017). Unlike MPO-deficiency (Nauseef 1988), loss of NOX2 results in immunodeficiency and recurrent infections (Noreng et al. 2022). Inability to produce NADPH due to glucose 6-phosphate dehydrogenase (G6PD) deficiency renders an individual prone to infections, presumably due to reduced activity of NOX2, and susceptible to oxidative stress due to reduced activity of glutathione peroxidases (Luzzatto et al. 2020; Mallouh and Abu-Osba 1987). G6PD deficiency also increases the risk of COVID-19 disease (Vick 2020).
Superoxide produced by NOX2 is converted to hydrogen peroxide by superoxide dismutase (Winterbourn et al. 2016). Superoxide dismutase activity in PMN and lymphocytes depends on prosthetic copper and zinc cofactors (Vucić et al. 1997). Human catalase requires a heme–iron and NADPH (niacinamide/vitamin B3 derived) cofactors and detoxifies hydrogen peroxide by its conversion to water and oxygen (Goyal and Basak 2010). Glutathionylation of catalase by NOX2 can result in PMN death (Nagarkoti et al. 2019).
MPO synthesizes ClO− and tau-Cl from hydrogen peroxide, chloride, and taurine (Aratani 2018; Kanofsky 1989; Schuller-Levis and Park 2003). Hydrogen peroxide and alkyl-hydroperoxides can react with ClO− and other oxidants (e.g., metal ions and peroxynitrite) to form singlet oxygen (Miyamoto et al. 2007; Kanofsky 1989). ClO− is a highly reactive oxidant that can degrade the tetrapyrrole rings of hemoglobin, cause vitamin B12 and nitric oxide deficiency, compete with oxygen for hemoglobin binding, and is thought to be responsible for the induction of cytokine storms (Camp et al. 2021; Goud et al. 2021). ClO− reacts rapidly with glutathione to give products that glutathione reductase cannot reduce (Carr and Winterbourn 1997). Tau-Cl is a somewhat less reactive oxidant than ClO− with a longer lifetime and more extended range of action but similar detoxification chemistry by glutathione (Schuller-Levis and Park 2003; Kim and Cha 2014; Carr et al. 2001). Tau-Cl has anti-inflammatory effects by mitigating the inflammatory impact of ClO− (Marcinkiewicz and Kontny 2014; Kim and Cha 2014). Singlet oxygen is the most reactive of the ROS. Despite the limited life of PMN-generated singlet oxygen due to its decomposition by radiative decay (λ = 1270 nm and 1200 nm) it reacts at diffusion-limited rates with various intra- and extra-cellular components to form hydroperoxides and dioxetanes (Hackbarth et al. 2022; Stief 2003; Di Mascio et al. 1990; 2019; Skovsen et al. 2005). Singlet oxygen is also thought to potentiate thrombolysis (Stief 2007). GPX4 (an essential selenium enzyme) is required to prevent death by ferroptosis from the alkyl-peroxides and phospholipid-peroxides formed by singlet oxygen (Stockwell et al. 2020; Yang and Stockwell 2016).
The third stage of response: antibody production
Activated B lymphocytes will differentiate into antibody-producing plasma cells (Nutt et al. 2015). Chillon et al. (2022) have shown that the antibody response to vaccination with SARS-CoV-2 antigens correlates with free zinc but not with total zinc levels (99.999% protein-bound). The average level of free zinc in human serum is about 0.2 μM (0.09–0.42 nM), only 0.0015% of the total zinc, 11.5–15 μM (Chillon et al. 2022; Alker et al. 2019; Maares et al. 2022). The difference between serum free zinc in COVID-19 survivors is, on average, about twice the concentration of free zinc in patients that do not survive infection (0.4 vs. 0.2 nM) (Maares et al. 2022). Males tend to have slightly higher free zinc levels (0.23 vs. 0.21 nM) than females (Chillon et al. 2022; Maares et al. 2019).
IgG antibodies have a weak metal-binding site (copper or zinc) in their Fc region (Glover et al. 2015; Mehta et al. 2021; Yamanaka et al. 2016). Zinc binding to the metal site of IgG antibodies results in protein aggregation and prevents copper-induced degradation (Glover et al. 2015; Mehta et al. 2021). IgG is only one of many zinc-binding proteins in the blood, such as other immunoglobulins (IgM and IgA), albumin, α2-macroglobulin, haptoglobulin, fibrinogen, ceruloplasmin, complement C4, prealbumin, and C-reactive protein (Yamanaka et al. 2016). These proteins, together with other zinc-binding nutrients in the blood, such as the bioflavonoid quercetin, will maintain the ‘free’ zinc levels to facilitate cellular uptake by the ZIP transporters (Singh et al. 2021; Dabbagh-Bazarbachi et al. 2014; Haase and Rink 2014). Free zinc is a potent inhibitor of the SARS-CoV-2 main protease at nanomolar concentrations and is an inhibitor of viral replication (Panchariya et al. 2021). Adverse effects on antibody production at higher free zinc concentrations may explain the observation of maximum response during hypozincemia (Xu et al. 2022). There may be an optimal and dynamic range of free zinc for antibody response to SARS-CoV-2 that may not correspond to other aspects of the immune response.
Summary
Nutritional deficiencies predispose individuals to severe infection by SARS-CoV-2. COVID-19 disease further exacerbates dietary deficiencies. Stress before or after illness also lowers the stores of essential nutrients. Although zinc (33%) and selenium (14%) are prevalent deficiencies, there are more than 22 different nutritional factors (MND) reported to influence infection outcomes. People at higher risk of infection due to MND are also more likely to have long-term sequelae (Long COVID).
Acknowledgements
No grant support was used to prepare this manuscript.
Abbreviations
- MND
Multiple nutritional deficiencies
- LPA
Lymphocyte proliferation assay
- MPO
Myeloperoxidase
- GPX4
Glutathione peroxidase isozyme 4
- NOX2
NADPH oxidase 2
Funding
Not applicable.
Data availability
Enquiries about data availability should be directed to the author.
Declarations
Conflict of interest
The author declares that he has no conflict of interest, financial or otherwise.
Consent for publication
Not applicable.
Footnotes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- Abdelmaksoud AA, Ghweil AA, Hassan MH, Rashad A, Khodeary A, Aref ZF, Sayed MAA, Elsamman MK, Bazeed SES. Olfactory disturbances as presenting manifestation among Egyptian patients with COVID-19: possible role of zinc. Biol Trace Elem Res. 2021;199(11):4101–4108. doi: 10.1007/s12011-020-02546-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akasov RA, Khaydukov EV, Andreyuk DS, Sholina NV, Sheremeta AN, Romanov DV, Kostyuk GP, Panchenko VY, Kovalchuk MV. Riboflavin for COVID-19 adjuvant treatment in patients with mental health disorders: observational study. Front Pharmacol. 2022;13:755745. doi: 10.3389/fphar.2022.755745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Al Sulaiman K, Aljuhani O, Al Dossari M, Alshahrani A, Alharbi A, Algarni R, Al Jeraisy M, Al Harbi S, Al Katheri A, Al Eidan F, Al Bekairy AM, Al Qahtani N, Al Muqrin M, Vishwakarma R, Al Ghamdi G. Evaluation of thiamine as adjunctive therapy in COVID-19 critically ill patients: a two-center propensity score matched study. Crit Care. 2021;25(1):223. doi: 10.1186/s13054-021-03648-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander TH, Davidson TM. Intranasal zinc and anosmia: the zinc-induced anosmia syndrome. Laryngoscope. 2006;116(2):217–220. doi: 10.1097/01.mlg.0000191549.17796.13. [DOI] [PubMed] [Google Scholar]
- Alexander R, Debiec N, Razzaque MS, He P. Inorganic phosphate-induced cytotoxicity. IUBMB Life. 2022;74(1):117–124. doi: 10.1002/iub.2561. [DOI] [PubMed] [Google Scholar]
- Alfano G, Ferrari A, Fontana F, Perrone R, Mori G, Ascione E, Magistroni R, Venturi G, Pederzoli S, Margiotta G, Romeo M, Piccinini F, Franceschi G, Volpi S, Faltoni M, Ciusa G, Bacca E, Tutone M, Raimondi A, Menozzi M, Franceschini E, Cuomo G, Orlando G, Santoro A, Di Gaetano M, Puzzolante C, Carli F, Bedini A, Milic J, Meschiari M, Mussini C, Cappelli G, Guaraldi G. Hypokalemia in patients with COVID-19. Clin Exp Nephrol. 2021;25(4):401–409. doi: 10.1007/s10157-020-01996-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alker W, Schwerdtle T, Schomburg L, Haase H. A Zinpyr-1-based fluorimetric microassay for free zinc in human serum. Int J Mol Sci. 2019;20(16):4006. doi: 10.3390/ijms20164006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Andreou A, Trantza S, Filippou D, Sipsas N, Tsiodras S. COVID-19: The potential role of copper and N-acetylcysteine (NAC) in a combination of candidate antiviral treatments against SARS-CoV-2. In Vivo. 2020;34(3 Suppl):1567–1588. doi: 10.21873/invivo.11946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aratani Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch Biochem Biophys. 2018;640:47–52. doi: 10.1016/j.abb.2018.01.004. [DOI] [PubMed] [Google Scholar]
- Ardestani A, Azizi Z. Targeting glucose metabolism for treatment of COVID-19. Signal Transduct Target Ther. 2021;6(1):112. doi: 10.1038/s41392-021-00532-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Assimakopoulos SF, Aretha D, Komninos D, Dimitropoulou D, Lagadinou M, Leonidou L, Oikonomou I, Mouzaki A, Marangos M. N-acetyl-cysteine reduces the risk for mechanical ventilation and mortality in patients with COVID-19 pneumonia: a two-center retrospective cohort study. Infect Dis (lond) 2021;53(11):847–854. doi: 10.1080/23744235.2021.1945675. [DOI] [PubMed] [Google Scholar]
- Barceloux DG. Selenium. J Toxicol Clin Toxicol. 1999;37(2):145–172. doi: 10.1081/clt-100102417. [DOI] [PubMed] [Google Scholar]
- Belikov AV, Schraven B, Simeoni L. T cells and reactive oxygen species. J Biomed Sci. 2015;22:85. doi: 10.1186/s12929-015-0194-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biancatelli RMLC, Berrill M, Catravas JD, Marik PE. Quercetin and vitamin C: an experimental, synergistic therapy for the prevention and treatment of SARS-CoV-2 related disease (COVID-19) Front Immunol. 2020;11:1451. doi: 10.3389/fimmu.2020.01451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bird, et al. Risk of deficiency in multiple concurrent micronutrients in children and adults in the United States. Nutrients. 2017;9:655. doi: 10.3390/nu9070655. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bizzarri M, Laganà AS, Aragona D, Unfer V. Inositol and pulmonary function. Could myo-inositol treatment downregulate inflammation and cytokine release syndrome in SARS-CoV-2? Eur Rev Med Pharmacol Sci. 2020;24(6):3426–3432. doi: 10.26355/eurrev_202003_20715. [DOI] [PubMed] [Google Scholar]
- Branco de Oliveira MV, Irikura S, Lourenço FHB, Shinsato M, Irikura TCDB, Irikura RB, Albuquerque TVC, Shinsato VN, Orsatti VN, Fontanelli AM, Samegima DAG, Gonçalves MVM, Bernabé DG. Encephalopathy responsive to thiamine in severe COVID-19 patients. Brain Behav Immun Health. 2021;14:100252. doi: 10.1016/j.bbih.2021.100252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bucci LR. A functional analytical technique for monitoring nutrient status and repletion. Part 3: clinical experience. Am Clin Lab. 1994;13:10–11. [PubMed] [Google Scholar]
- Camp OG, Bai D, Gonullu DC, Nayak N, Abu-Soud HM. Melatonin interferes with COVID-19 at several distinct ROS-related steps. J Inorg Biochem. 2021;223:111546. doi: 10.1016/j.jinorgbio.2021.111546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr AC, Winterbourn CC. Oxidation of neutrophil glutathione and protein thiols by myeloperoxidase-derived hypochlorous acid. Biochem J. 1997;327(Pt 1):275–281. doi: 10.1042/bj3270275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carr AC, Hawkins CL, Thomas SR, Stocker R, Frei B. Relative reactivities of N-chloramines and hypochlorous acid with human plasma constituents. Free Radic Biol Med. 2001;30(5):526–536. doi: 10.1016/s0891-5849(00)00495-0. [DOI] [PubMed] [Google Scholar]
- Cengiz M, Uysal BB, Ikitimur H, Ozcan E, Islamoğlu MS, Aktepe E, Yavuzer H, Yavuzer S. Effect of oral L-glutamine supplementation on Covid-19 treatment. Clin Nutr Exp. 2020;33:24–31. doi: 10.1016/j.yclnex.2020.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen J. An original discovery: selenium deficiency and Keshan disease (an endemic heart disease) Asia Pac J Clin Nutr. 2012;21(3):320–326. [PubMed] [Google Scholar]
- Chillon TS, Maares M, Demircan K, Hackler J, Sun Q, Heller RA, Diegmann J, Bachmann M, Moghaddam A, Haase H, Schomburg L. Serum free zinc is associated with vaccination response to SARS-CoV-2. Front Immunol. 2022;13:906551. doi: 10.3389/fimmu.2022.906551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Clemente-Suárez VJ, Ramos-Campo DJ, Mielgo-Ayuso J, Dalamitros AA, Nikolaidis PA, Hormeño-Holgado A, Tornero-Aguilera JF. Nutrition in the actual COVID-19 pandemic. Narrat Rev Nutr. 2021;13(6):1924. doi: 10.3390/nu13061924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cory JG, Cory AH. Critical roles of glutamine as nitrogen donors in purine and pyrimidine nucleotide synthesis: asparaginase treatment in childhood acute lymphoblastic leukemia. In Vivo. 2006;20(5):587–589. [PubMed] [Google Scholar]
- Dabbagh-Bazarbachi H, Clergeaud G, Quesada IM, Ortiz M, O'Sullivan CK, Fernández-Larrea JB. Zinc ionophore activity of quercetin and epigallocatechin-gallate: from Hepa 1–6 cells to a liposome model. J Agric Food Chem. 2014;62(32):8085–8093. doi: 10.1021/jf5014633. [DOI] [PubMed] [Google Scholar]
- Dalbeni A, Bevilacqua M, Teani I, Normelli I, Mazzaferri F, Chiarioni G. Excessive vitamin B12 and poor outcome in COVID-19 pneumonia. Nutr Metab Cardiovasc Dis. 2021;31(3):774–775. doi: 10.1016/j.numecd.2020.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Das UN. Can bioactive lipids inactivate coronavirus (COVID-19)? Arch Med Res. 2020;51(3):282–286. doi: 10.1016/j.arcmed.2020.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Alencar JCG, Moreira CL, Müller AD, Chaves CE, Fukuhara MA, da Silva EA, Miyamoto MFS, Pinto VB, Bueno CG, Neto FL, Gomez LM, Menezes MCS, Marchini JFM, Marino LO, Brandão Neto RA, Souza HP. Double-blind, randomized, placebo-controlled trial with N-acetylcysteine for treatment of severe acute respiratory syndrome caused by Coronavirus disease 2019 (COVID-19) Clin Infect Dis. 2021;72(11):e736–e741. doi: 10.1093/cid/ciaa1443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Flora S, Balansky R, La Maestra S. Rationale for the use of N-acetylcysteine in both prevention and adjuvant therapy of COVID-19. FASEB J. 2020;34(10):13185–13193. doi: 10.1096/fj.202001807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Derosa G, Maffioli P, D'Angelo A, Di Pierro F. A role for quercetin in coronavirus disease 2019 (COVID-19) Phytother Res. 2021;35(3):1230–1236. doi: 10.1002/ptr.6887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dharmalingam K, Birdi A, Tomo S, Sreenivasulu K, Charan J, Yadav D, Purohit P, Sharma P. Trace elements as immunoregulators in SARS-CoV-2 and other viral infections. Indian J Clin Biochem. 2021;36(4):416–426. doi: 10.1007/s12291-021-00961-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Mascio P, Devasagayam TP, Kaiser S, Sies H. Carotenoids, tocopherols and thiols as biological singlet molecular oxygen quenchers. Biochem Soc Trans. 1990;18(6):1054–1056. doi: 10.1042/bst0181054. [DOI] [PubMed] [Google Scholar]
- Di Mascio P, Martinez GR, Miyamoto S, Ronsein GE, Medeiros MHG, Cadet J. Singlet molecular oxygen reactions with nucleic acids, lipids, and proteins. Chem Rev. 2019;119(3):2043–2086. doi: 10.1021/acs.chemrev.8b00554. [DOI] [PubMed] [Google Scholar]
- Doaei S, Gholami S, Rastgoo S, Gholamalizadeh M, Bourbour F, Bagheri SE, Samipoor F, Akbari ME, Shadnoush M, Ghorat F, Mosavi Jarrahi SA, Ashouri Mirsadeghi N, Hajipour A, Joola P, Moslem A, Goodarzi MO. The effect of omega-3 fatty acid supplementation on clinical and biochemical parameters of critically ill patients with COVID-19: a randomized clinical trial. J Transl Med. 2021;19(1):128. doi: 10.1186/s12967-021-02795-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drakesmith H, Pasricha SR, Cabantchik I, Hershko C, Weiss G, Girelli D, Stoffel N, Muckenthaler MU, Nemeth E, Camaschella C, Klenerman P, Zimmermann MB. Vaccine efficacy and iron deficiency: an intertwined pair? Lancet Haematol. 2021;8(9):e666–e669. doi: 10.1016/S2352-3026(21)00201-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Du Y, Luo K, Ni R, Hussain R. Selenium and hazardous elements distribution in plant-soil-water system and human health risk assessment of lower Cambrian, Southern Shaanxi, China. Environ Geochem Health. 2018;40(5):2049–2069. doi: 10.1007/s10653-018-0082-3. [DOI] [PubMed] [Google Scholar]
- Du Laing G, Petrovic M, Lachat C, De Boevre M, Klingenberg GJ, Sun Q, De Saeger S, De Clercq J, Ide L, Vandekerckhove L, Schomburg L. Course and survival of COVID-19 patients with comorbidities in relation to the trace element status at hospital admission. Nutrients. 2021;13(10):3304. doi: 10.3390/nu13103304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elham A. Vitamin B12 deficiency in COVID-19 recovered patients: case report. Int J Pharma Res. 2021;13(1):482–485. [Google Scholar]
- El-Kurdi B, Khatua B, Rood C, Snozek C, Cartin-Ceba R, Singh VP. Mortality from coronavirus disease 2019 increases with unsaturated fat and may be reduced by early calcium and albumin supplementation. Gastroenterology. 2020;159(3):1015–1018.e4. doi: 10.1053/j.gastro.2020.05.057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Engström W, Zetterberg A. Phosphate and the regulation of DNA replication in normal and virus-transformed 3T3 cells. Biochem J. 1983;214(3):695–702. doi: 10.1042/bj2140695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eskander M, Razzaque MS. Can maintaining optimal magnesium balance reduce the disease severity of COVID-19 patients? Front Endocrinol (lausanne) 2022;13:843152. doi: 10.3389/fendo.2022.843152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Españo E, Kim J, Lee K, Kim JK. Phytochemicals for the treatment of COVID-19. J Microbiol. 2021;59(11):959–977. doi: 10.1007/s12275-021-1467-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espinola MSB, Bertelli M, Bizzarri M, Unfer V, Laganà AS, Visconti B, Aragona C. Inositol and vitamin D may naturally protect human reproduction and women undergoing assisted reproduction from covid-19 risk. J Reprod Immunol. 2021;144:103271. doi: 10.1016/j.jri.2021.103271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evanics F, Maurmann L, Yang WW, Bose RN. Nuclear magnetic resonance structures of the zinc finger domain of human DNA polymerase-alpha. Biochim Biophys Acta. 2003;1651(1–2):163–171. doi: 10.1016/s1570-9639(03)00266-8. [DOI] [PubMed] [Google Scholar]
- Fairweather-Tait SJ, Bao Y, Broadley MR, Collings R, Ford D, Hesketh JE, Hurst R. Selenium in human health and disease. Antioxid Redox Signal. 2011;14(7):1337–1383. doi: 10.1089/ars.2010.3275. [DOI] [PubMed] [Google Scholar]
- Fakhrolmobasheri M, Mazaheri-Tehrani S, Kieliszek M, Zeinalian M, Abbasi M, Karimi F, Mozafari AM. COVID-19 and selenium deficiency: a systematic review. Biol Trace Elem Res. 2022;200(9):3945–3956. doi: 10.1007/s12011-021-02997-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fosmire GJ. Zinc toxicity. Am J Clin Nutr. 1990;51(2):225–227. doi: 10.1093/ajcn/51.2.225. [DOI] [PubMed] [Google Scholar]
- Freedman R, Hunter SK, Law AJ, D’Alessandro A, Noonan K, Wyrwa A, Camille Hoffman M. Maternal choline and respiratory coronavirus effects on fetal brain development. J Psychiatr Res. 2020;128:1–4. doi: 10.1016/j.jpsychires.2020.05.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fröhlich E, Wahl R. Physiological role and use of thyroid hormone metabolites - potential utility in COVID-19 patients. Front Endocrinol (lausanne) 2021;12:587518. doi: 10.3389/fendo.2021.587518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galmés S, Serra F, Palou A. Current state of evidence: influence of nutritional and nutrigenetic factors on immunity in the COVID-19 pandemic framework. Nutrients. 2020;12(9):2738. doi: 10.3390/nu12092738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gheorghe G, Ilie M, Bungau S, Stoian AMP, Bacalbasa N, Diaconu CC. Is there a relationship between COVID-19 and hyponatremia? Medicina (kaunas) 2021;57(1):55. doi: 10.3390/medicina57010055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glover ZK, Basa L, Moore B, Laurence JS, Sreedhara A. Metal ion interactions with mAbs: part 1. Mabs. 2015;7(5):901–911. doi: 10.1080/19420862.2015.1062193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goc A, Niedzwiecki A, Rath M. Polyunsaturated ω-3 fatty acids inhibit ACE2-controlled SARS-CoV-2 binding and cellular entry. Sci Rep. 2021;11(1):5207. doi: 10.1038/s41598-021-84850-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goud PT, Bai D, Abu-Soud HM. A multiple-hit hypothesis involving reactive oxygen species and myeloperoxidase explains clinical deterioration and fatality in COVID-19. Int J Biol Sci. 2021;17(1):62–72. doi: 10.7150/ijbs.51811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goyal MM, Basak A. Human catalase: looking for complete identity. Protein Cell. 2010;1(10):888–897. doi: 10.1007/s13238-010-0113-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grant WB, Lahore H, McDonnell SL, Baggerly CA, French CB, Aliano JL, Bhattoa HP. Evidence that Vitamin D supplementation could reduce risk of influenza and COVID-19 infections and deaths. Nutrients. 2020;12(4):988. doi: 10.3390/nu12040988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haase H, Rink L. Zinc signals and immune function. BioFactors. 2014;40(1):27–40. doi: 10.1002/biof.1114. [DOI] [PubMed] [Google Scholar]
- Habib HM, Ibrahim S, Zaim A, Ibrahim WH. The role of iron in the pathogenesis of COVID-19 and possible treatment with lactoferrin and other iron chelators. Biomed Pharmacother. 2021;136:111228. doi: 10.1016/j.biopha.2021.111228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hackbarth S, Islam R, Šubr V, Etrych T, Fang J. Singlet oxygen in vivo: It is all about intensity. J Pers Med. 2022;12(6):891. doi: 10.3390/jpm12060891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hariharan S, Dharmaraj S. Selenium and selenoproteins: it’s role in regulation of inflammation. Inflammopharmacology. 2020;28(3):667–695. doi: 10.1007/s10787-020-00690-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heller RA, Sun Q, Hackler J, Seelig J, Seibert L, Cherkezov A, Minich WB, Seemann P, Diegmann J, Pilz M, Bachmann M, Ranjbar A, Moghaddam A, Schomburg L. Prediction of survival odds in COVID-19 by zinc, age and selenoprotein P as composite biomarker. Redox Biol. 2021;38:101764. doi: 10.1016/j.redox.2020.101764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hemila H, de Man AME. Vitamin C and COVID-19. Front Med (lausanne). 2021;7:559811. doi: 10.3389/fmed.2020.559811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holford P, Carr AC, Jovic TH, Ali SR, Whitaker IS, Marik PE, Smith AD. Vitamin C-an adjunctive therapy for respiratory infection, sepsis and COVID-19. Nutrients. 2020;12(12):3760. doi: 10.3390/nu12123760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holick MF. Sunlight, UV radiation, vitamin D, and skin cancer: how much sunlight do we need? Adv Exp Med Biol. 2020;1268:19–36. doi: 10.1007/978-3-030-46227-7_2. [DOI] [PubMed] [Google Scholar]
- Houillier P, Salles JP. Biochemical assessment of phosphate homeostasis. Arch Pediatr. 2021;28(7):588–593. doi: 10.1016/j.arcped.2021.09.001. [DOI] [PubMed] [Google Scholar]
- Im JH, Je YS, Baek J, Chung MH, Kwon HY, Lee JS. Nutritional status of patients with COVID-19. Int J Infect Dis. 2020;100:390–393. doi: 10.1016/j.ijid.2020.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwegbulem O, Wang J, Pfirrmann RW, Redmond HP. The role of taurine derivatives in the putative therapy of COVID-19-induced inflammation. Ir J Med Sci. 2022;191(1):485–486. doi: 10.1007/s11845-021-02522-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jorge-Aarón RM, Rosa-Ester MP. N-acetylcysteine as a potential treatment for COVID-19. Future Microbiol. 2020;15:959–962. doi: 10.2217/fmb-2020-0074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jose J, Magoon R, Kapoor PM. Magnesium: the neglected cation in COVID-19? J Anaesthesiol Clin Pharmacol. 2021;37(1):141–142. doi: 10.4103/joacp.JOACP_628_20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanofsky JR. Singlet oxygen production by biological systems. Chem Biol Interact. 1989;70(1–2):1–28. doi: 10.1016/0009-2797(89)90059-8. [DOI] [PubMed] [Google Scholar]
- Kapur A, Sharma M, Sageena G. Therapeutic potential of N-acetyl cysteine during COVID-19 epoch. World J Virol. 2022;11(2):104–106. doi: 10.5501/wjv.v11.i2.104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaya MO, Pamukçu E, Yakar B. The role of vitamin D deficiency on COVID-19: a systematic review and meta-analysis of observational studies. Epidemiol Health. 2021;43:e2021074. doi: 10.4178/epih.e2021074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khatiwada S, Subedi A. A mechanistic link between selenium and coronavirus disease 2019 (COVID-19) Curr Nutr Rep. 2021;10(2):125–136. doi: 10.1007/s13668-021-00354-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim C, Cha YN. Taurine chloramine produced from taurine under inflammation provides anti-inflammatory and cytoprotective effects. Amino Acids. 2014;46(1):89–100. doi: 10.1007/s00726-013-1545-6. [DOI] [PubMed] [Google Scholar]
- Kogan S, Sood A, Garnick MS. Zinc and wound healing: a review of zinc physiology and clinical applications. Wounds. 2017;29(4):102–106. [PubMed] [Google Scholar]
- Krishnaven GV, et al. Maternal B12, Folate and homocysteine concentrations and offspring cortisol and cardiovascular responses to stress. J Clin Endocrinol Metab. 2020;105(7):e2591–e2599. doi: 10.1210/clinem/dgz114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kumrungsee T, Zhang P, Chartkul M, Yanaka N, Kato N. Potential role of vitamin B6 in ameliorating the severity of COVID-19 and its complications. Front Nutr. 2020;7:562051. doi: 10.3389/fnut.2020.562051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Laganà AS, Unfer V, Garzon S, Bizzarri M. Role of inositol to improve surfactant functions and reduce IL-6 levels: a potential adjuvant strategy for SARS-CoV-2 pneumonia? Med Hypotheses. 2020;144:110262. doi: 10.1016/j.mehy.2020.110262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landis HE, Getachew B, Tizabi Y. Therapeutic potential of flavonoids and zinc in COVID-19. Medpress Nutr Food Sci. 2022;1(1):202111001. [PMC free article] [PubMed] [Google Scholar]
- Liu F, Zhu Y, Zhang J, Li Y, Peng Z. Intravenous high-dose vitamin C for the treatment of severe COVID-19: study protocol for a multicentre randomised controlled trial. BMJ Open. 2020;10(7):e039519. doi: 10.1136/bmjopen-2020-039519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu S, Zhang L, Weng H, Yang F, Jin H, Fan F, Zheng X, Yang H, Li H, Zhang Y, Li J. Association between average plasma potassium levels and 30-day mortality during hospitalization in patients with COVID-19 in Wuhan. China Int J Med Sci. 2021;18(3):736–743. doi: 10.7150/ijms.50965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lopresti AL. The effects of psychological and environmental stress on micronutrient concentrations in the body: a review of the evidence. Adv Nutr. 2019;11:103–112. doi: 10.1093/advances/nmz082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luzzatto L, Ally M, Notaro R. Glucose-6-phosphate dehydrogenase deficiency. Blood. 2020;136(11):1225–1240. doi: 10.1182/blood.2019000944. [DOI] [PubMed] [Google Scholar]
- Maares M, Hackler J, Haupt A, Heller RA, Bachmann M, Diegmann J, Moghaddam A, Schomburg L, Haase H. Free zinc as a predictive marker for COVID-19 mortality risk. Nutrients. 2022;14(7):1407. doi: 10.3390/nu14071407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Magnani F, Nenci S, Millana Fananas E, Ceccon M, Romero E, Fraaije MW, Mattevi A. Crystal structures and atomic model of NADPH oxidase. Proc Natl Acad Sci U S A. 2017;114(26):6764–6769. doi: 10.1073/pnas.1702293114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maitra D, Shaeib F, Abdulhamid I, Abdulridha RM, Saed GM, Diamond MP, Pennathur S, Abu-Souda HM. Myeloperoxidase acts as a source of free iron during steady-state catalysis by a feedback inhibitory pathway. Free Radic Biol Med. 2013 doi: 10.1016/j.freeradbiomed.2013.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mallouh AA, Abu-Osba YK. Bacterial infections in children with glucose-6-phosphate dehydrogenase deficiency. J Pediatr. 1987;111(6 Pt 1):850–852. doi: 10.1016/s0022-3476(87)80202-0. [DOI] [PubMed] [Google Scholar]
- Marcinkiewicz J, Kontny E. Taurine and inflammatory diseases. Amino Acids. 2014;46(1):7–20. doi: 10.1007/s00726-012-1361-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez SS, Huang Y, Acuna L, Laverde E, Trujillo D, Barbieri MA, Tamargo J, Campa A, Baum MK. Role of selenium in viral infections with a major focus on SARS-CoV-2. Int J Mol Sci. 2021;23(1):280. doi: 10.3390/ijms23010280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsushita M, Freigang S, Schneider C, Conrad M, Bornkamm GW, Kopf M. T cell lipid peroxidation induces ferroptosis and prevents immunity to infection. J Exp Med. 2015;212(4):555–568. doi: 10.1084/jem.20140857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matsuyama T, Yoshinaga SK, Shibue K, Mak TW. Comorbidity-associated glutamine deficiency is a predisposition to severe COVID-19. Cell Death Differ. 2021;28(12):3199–3213. doi: 10.1038/s41418-021-00892-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayadas TN, Cullere X, Lowell CA. The multifaceted functions of neutrophils. Annu Rev Pathol. 2014;9:181–218. doi: 10.1146/annurev-pathol-020712-164023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mehta S, Flores H, Walters B, Sreedhara A. Metal ion interactions with mAbs: Part 2. Zinc-mediated aggregation of IgG1 monoclonal antibodies. Pharm Res. 2021;38(8):1387–1395. doi: 10.1007/s11095-021-03089-7. [DOI] [PubMed] [Google Scholar]
- Meisel E, Efros O, Bleier J, Beit Halevi T, Segal G, Rahav G, Leibowitz A, Grossman E. Folate levels in patients hospitalized with coronavirus disease 2019. Nutrients. 2021;13(3):812. doi: 10.3390/nu13030812. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Miyamoto S, Ronsein GE, Prado FM, Uemi M, Corrêa TC, Toma IN, Bertolucci A, Oliveira MC, Motta FD, Medeiros MH, Mascio PD. Biological hydroperoxides and singlet molecular oxygen generation. IUBMB Life. 2007;59(4–5):322–331. doi: 10.1080/15216540701242508. [DOI] [PubMed] [Google Scholar]
- Moghaddam A, Heller RA, Sun Q, Seelig J, Cherkezov A, Seibert L, Hackler J, Seemann P, Diegmann J, Pilz M, Bachmann M, Minich WB, Schomburg L. Selenium deficiency is associated with mortality. Nutrients. 2020;12(7):2098. doi: 10.3390/nu12072098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mohanty RR, Padhy BM, Das S, Meher BR. Therapeutic potential of N-acetyl cysteine (NAC) in preventing cytokine storm in COVID-19 review of current evidence. Eur Rev Med Pharmacol Sci. 2021;25(6):2802–2807. doi: 10.26355/eurrev_202103_25442. [DOI] [PubMed] [Google Scholar]
- Muller FL, Lustgarten MS, Jang Y, Richardson A, Van Remmen H. Trends in oxidative aging theories. Free Radic Biol Med. 2007;43(4):477–503. doi: 10.1016/j.freeradbiomed.2007.03.034. [DOI] [PubMed] [Google Scholar]
- Muri J, Heer S, Matsushita M, Pohlmeier L, Tortola L, Fuhrer T, Conrad M, Zamboni N, Kisielow J, Kopf M. The thioredoxin-1 system is essential for fueling DNA synthesis during T-cell metabolic reprogramming and proliferation. Nat Commun. 2018;9(1):1851. doi: 10.1038/s41467-018-04274-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagarkoti S, Dubey M, Sadaf S, Awasthi D, Chandra T, Jagavelu K, Kumar S, Dikshit M. Catalase S-glutathionylation by NOX2 and mitochondrial-derived ROS adversely affects mice and human neutrophil survival. Inflammation. 2019;42(6):2286–2296. doi: 10.1007/s10753-019-01093-z. [DOI] [PubMed] [Google Scholar]
- Nai A, Lorè NI, Pagani A, De Lorenzo R, Di Modica S, Saliu F, Cirillo DM, Rovere-Querini P, Manfredi AA, Silvestri L. Hepcidin levels predict Covid-19 severity and mortality in a cohort of hospitalized Italian patients. Am J Hematol. 2021;96(1):E32–E35. doi: 10.1002/ajh.26027. [DOI] [PubMed] [Google Scholar]
- Nauseef WM. Myeloperoxidase deficiency. Hematol Oncol Clin North Am. 1988;2(1):135–158. doi: 10.1016/S0889-8588(18)30634-8. [DOI] [PubMed] [Google Scholar]
- Neiers F, Jarriault D, Menetrier F, Briand L, Heydel JM. The odorant metabolizing enzyme UGT2A1: immunolocalization and impact of the modulation of its activity on the olfactory response. PLoS ONE. 2021;16(3):e0249029. doi: 10.1371/journal.pone.0249029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noori M, Nejadghaderi SA, Sullman MJM, Carson-Chahhoud K, Kolahi AA, Safiri S. Epidemiology, prognosis and management of potassium disorders in covid-19. Rev Med Virol. 2022;32(1):e2262. doi: 10.1002/rmv.2262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noreng S, Ota N, Sun Y, Ho H, Johnson M, Arthur CP, Schneider K, Lehoux I, Davies CW, Mortara K, Wong K, Seshasayee D, Masureel M, Payandeh J, Yi T, Koerber JT. Structure of the core human NADPH oxidase NOX2. Nat Commun. 2022;13(1):6079. doi: 10.1038/s41467-022-33711-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Notz Q, Herrmann J, Schlesinger T, Helmer P, Sudowe S, Sun Q, Hackler J, Roeder D, Lotz C, Meybohm P, Kranke P, Schomburg L, Stoppe C. Clinical significance of micronutrient supplementation in critically ill COVID-19 patients with severe ARDS. Nutrients. 2021;13(6):2113. doi: 10.3390/nu13062113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nowak G, Szewczyk B, Pilc A. Zinc and depression. An update. Pharmacol Rep. 2005;57(6):713–718. [PubMed] [Google Scholar]
- Nutt SL, Hodgkin PD, Tarlinton DM, Corcoran LM. The generation of antibody-secreting plasma cells. Nat Rev Immunol. 2015;15(3):160–171. doi: 10.1038/nri3795. [DOI] [PubMed] [Google Scholar]
- Olivari L, Riccardi N, Rodari P, Angheben A, Artioli P, Salgarello M. COVID-19 pneumonia: increased choline uptake with 18F-choline PET/CT. Eur J Nucl Med Mol Imaging. 2020;47(10):2476–2477. doi: 10.1007/s00259-020-04870-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Osman W, Al Fahdi F, Al Salmi I, Al Khalili H, Gokhale A, Khamis F. Serum calcium and vitamin D levels: correlation with severity of COVID-19 in hospitalized patients in royal hospital, Oman. Int J Infect Dis. 2021;107:153–163. doi: 10.1016/j.ijid.2021.04.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panchariya L, Khan WA, Kuila S, Sonkar K, Sahoo S, Ghoshal A, Kumar A, Verma DK, Hasan A, Khan MA, Jain N, Mohapatra AK, Das S, Thakur JK, Maiti S, Nanda RK, Halder R, Sunil S. Arockiasamy A (2021) Zinc2+ ion inhibits SARS-CoV-2 main protease and viral replication in vitro. Chem Commun (camb) 2021;57(78):10083–10086. doi: 10.1039/d1cc03563k. [DOI] [PubMed] [Google Scholar]
- Petrucco S, Percudani R. Structural recognition of DNA by poly(ADP-ribose)polymerase-like zinc finger families. FEBS J. 2008;275(5):883–893. doi: 10.1111/j.1742-4658.2008.06259.x. [DOI] [PubMed] [Google Scholar]
- Peyneau M, Granger V, Wicky PH, Khelifi-Touhami D, Timsit JF, Lescure FX, Yazdanpanah Y, Tran-Dinh A, Montravers P, Monteiro RC, Chollet-Martin S, Hurtado-Nedelec M, de Chaisemartin L. Innate immune deficiencies are associated with severity and poor prognosis in patients with COVID-19. Sci Rep. 2022;12(1):638. doi: 10.1038/s41598-021-04705-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pierro DIF, Khan A, Bertuccioli A, Maffioli P, Derosa G, Khan S, Khan BA, Nigar R, Ujjan I, Devrajani BR. Quercetin phytosome® as a potential candidate for managing COVID-19. Minerva Gastroenterol (torino). 2021;67(2):190–195. doi: 10.23736/S2724-5985.20.02771-3. [DOI] [PubMed] [Google Scholar]
- Pisano M, Hilas O. Zinc and taste disturbances in older adults: a review of the literature. Consult Pharm. 2016;31(5):267–270. doi: 10.4140/TCP.n.2016.267. [DOI] [PubMed] [Google Scholar]
- Prasad AS. Recognition of zinc-deficiency syndrome. Nutrition. 2001;17(1):67–69. doi: 10.1016/s0899-9007(00)00469-x. [DOI] [PubMed] [Google Scholar]
- Prasad AS. Discovery of human zinc deficiency: its impact on human health and disease. Adv Nutr. 2013;4(2):176–190. doi: 10.3945/an.112.003210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raines NH, Ganatra S, Nissaisorakarn P, Pandit A, Morales A, Asnani A, Sadrolashrafi M, Maheshwari R, Patel R, Bang V, Shreyder K, Brar S, Singh A, Dani SS, Knapp S, Poyan Mehr A, Brown RS, Zeidel ML, Bhargava R, Schlondorff J, Steinman TI, Mukamal KJ, Parikh SM. Niacinamide may be associated with improved outcomes in COVID-19-related acute kidney injury: an observational study. Kidney 360. 2020;2(1):33–41. doi: 10.34067/KID.0006452020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Read SA, Obeid S, Ahlenstiel C, Ahlenstiel G. The role of zinc in antiviral immunity. Adv Nutr. 2019;10(4):696–710. doi: 10.1093/advances/nmz013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reusch N, De Domenico E, Bonaguro L, Schulte-Schrepping J, Baßler K, Schultze JL, Aschenbrenner AC. Neutrophils in COVID-19. Front Immunol. 2021;12:652470. doi: 10.3389/fimmu.2021.652470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenblum H, Wessler JD, Gupta A, Maurer MS, Bikdeli B. Zinc deficiency and heart failure: a systematic review of the current literature. J Card Fail. 2020;26(2):180–189. doi: 10.1016/j.cardfail.2020.01.005. [DOI] [PubMed] [Google Scholar]
- Schöllhammer I, Poll DS, Bickel MH. Liver microsomal beta-glucuronidase and UDP-glucuronyltransferase. Enzyme. 1975;20(5):269–276. doi: 10.1159/000458949. [DOI] [PubMed] [Google Scholar]
- Schomburg L. Selenium deficiency in COVID-19-a possible long-lasting toxic relationship. Nutrients. 2022;14(2):283. doi: 10.3390/nu14020283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuller-Levis GB, Park E. Taurine: new implications for an old amino acid. FEMS Microbiol Lett. 2003;226(2):195–202. doi: 10.1016/S0378-1097(03)00611-6. [DOI] [PubMed] [Google Scholar]
- Seers T, Davenport R. Phosphate metabolism and respiratory alkalosis: a forgotten lesson in COVID-19. Age Ageing. 2020;49(6):927. doi: 10.1093/ageing/afaa176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sengupta R, Holmgren A. Thioredoxin and glutaredoxin-mediated redox regulation of ribonucleotide reductase. World J Biol Chem. 2014;5(1):68–74. doi: 10.4331/wjbc.v5.i1.68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shauly-Aharonov M, Shafrir A, Paltiel O, Calderon-Margalit R, Safadi R, Bicher R, Barenholz-Goultschin O, Stokar J. Both high and low pre-infection glucose levels associated with increased risk for severe COVID-19: new insights from a population-based study. PLoS ONE. 2021;16(7):e02547. doi: 10.1371/journal.pone.0254847. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shelton JF, Shastri AJ, Fletez-Brant K, Aslibekyan S, Auton A. The UGT2A1/UGT2A2 locus is associated with COVID-19-related loss of smell or taste. Nat Genet. 2022;54(2):121–124. doi: 10.1038/s41588-021-00986-w. [DOI] [PubMed] [Google Scholar]
- Shi Z, Puyo CA. N-Acetylcysteine to combat COVID-19: An evidence review. Ther Clin Risk Manag. 2020;16:1047–1055. doi: 10.2147/TCRM.S273700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shive W, Pinkerton F, Humphreys J, Johnson MM, Hamilton WG, Matthews KS. Development of a chemically defined serum- and protein-free medium for growth of human peripheral lymphocytes. Proc Natl Acad Sci USA. 1986;83(1):9–13. doi: 10.1073/pnas.83.1.9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shrivastava S, Chelluboina S, Jedge P, Doke P, Palkar S, Mishra AC, Arankalle VA. Elevated levels of neutrophil activated proteins, alpha-defensins (DEFA1), calprotectin (S100A8/A9) and myeloperoxidase (MPO) are associated with disease severity in COVID-19 patients. Front Cell Infect Microbiol. 2021;11:7512. doi: 10.3389/fcimb.2021.751232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh CK, Chhabra G, Patel A, Chang H, Ahmad N. Dietary phytochemicals in zinc homeostasis: a strategy for prostate cancer management. Nutrients. 2021;13(6):1867. doi: 10.3390/nu13061867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skesters A, Kustovs D, Lece A, Moreino E, Petrosina E, Rainsford KKD. Selenium, selenoprotein P, and oxidative stress levels in SARS-CoV-2 patients during illness and recovery. Inflammopharmacology. 2022;30(2):499–503. doi: 10.1007/s10787-022-00925-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skovsen E, Snyder JW, Lambert JDC, Ogilby PR. Lifetime and diffusion of singlet oxygen in a cell. J Phys Chem B. 2005;109(18):8570–8573. doi: 10.1021/jp051163i. [DOI] [PubMed] [Google Scholar]
- Skrajnowska D, Bobrowska-Korczak B. Role of zinc in immune system and anti-cancer defense mechanisms. Nutrients. 2019;11(10):2273. doi: 10.3390/nu11102273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soliman OM, Thabet AMA, Abudahab GM, Kamel EZ. The impact of glutamine supplementation on the short-term mortality of COVID-19 diseased patients admitted to the ICU: A single-blind randomized clinical trial. Egypt J Anaesth. 2022;38(1):94–100. doi: 10.1080/11101849.2022.2031811. [DOI] [Google Scholar]
- Stief TW. The physiology and pharmacology of singlet oxygen. Med Hypotheses. 2003;60(4):567–572. doi: 10.1016/s0306-9877(03)00026-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stief TW. Singlet oxygen potentiates thrombolysis. Clin Appl Thromb Hemost. 2007;13(3):259–278. doi: 10.1177/1076029607302404. [DOI] [PubMed] [Google Scholar]
- Stockwell BR, Jiang X, Gu W. Emerging mechanisms and disease relevance of ferroptosis. Trends Cell Biol. 2020;30(6):478–490. doi: 10.1016/j.tcb.2020.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taneri PE, Gómez-Ochoa SA, Llanaj E, Raguindin PF, Rojas LZ, Roa-Díaz ZM, Salvador D, Jr, Groothof D, Minder B, Kopp-Heim D, Hautz WE, Eisenga MF, Franco OH, Glisic M, Muka T. Anemia and iron metabolism in COVID-19: a systematic review and meta-analysis. Eur J Epidemiol. 2020;35(8):763–773. doi: 10.1007/s10654-020-00678-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- tom Dieck H, Döring F, Roth HP, Daniel H. Changes in rat hepatic gene expression in response to zinc deficiency as assessed by DNA arrays. J Nutr. 2003;133(4):1004–1010. doi: 10.1093/jn/133.4.1004. [DOI] [PubMed] [Google Scholar]
- Trapani V, Rosanoff A, Baniasadi S, Barbagallo M, Castiglioni S, Guerrero-Romero F, Iotti S, Mazur A, Micke O, Pourdowlat G, Scarpati G, Wolf FI, Maier JA. The relevance of magnesium homeostasis in COVID-19. Eur J Nutr. 2022;61(2):625–636. doi: 10.1007/s00394-021-02704-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Kempen TATG, Deixler E. SARS-CoV-2: influence of phosphate and magnesium, moderated by vitamin D, on energy (ATP) metabolism and on severity of COVID-19. Am J Physiol Endocrinol Metab. 2021;320(1):E2–E6. doi: 10.1152/ajpendo.00474.2020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Eijk LE, Offringa AK, Bernal ME, Bourgonje AR, van Goor H, Hillebrands JL. The disease-modifying role of taurine and its therapeutic potential in coronavirus disease 2019 (COVID-19) Adv Exp Med Biol. 2022;1370:3–21. doi: 10.1007/978-3-030-93337-1_1. [DOI] [PubMed] [Google Scholar]
- Vazirani AA. COVID-19, an incentive to tackle sugar in hospitals and at home. J Endocr Soc. 2021;5(6):bvab037. doi: 10.1210/jendso/bvab037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vick DJ. Glucose-6-phosphate dehydrogenase deficiency and COVID-19 infection. Mayo Clin Proc. 2020;95(8):1803–1804. doi: 10.1016/j.mayocp.2020.05.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vucić M, Gavella M, Bozikov V, Ashcroft SJ, Rocić B. Superoxide dismutase activity in lymphocytes and polymorphonuclear cells of diabetic patients. Eur J Clin Chem Clin Biochem. 1997;35(7):517–521. doi: 10.1515/cclm.1997.35.7.517. [DOI] [PubMed] [Google Scholar]
- Wang W, Shen M, Tao Y, Fairley CK, Zhong Q, Li Z, Chen H, Ong JJ, Zhang D, Zhang K, Xing N, Guo H, Qin E, Guan X, Yang F, Zhang S, Zhang L, He K. Elevated glucose level leads to rapid COVID-19 progression and high fatality. BMC Pulm Med. 2021;21(1):64. doi: 10.1186/s12890-021-01413-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wee AKH. COVID-19’s toll on the elderly and those with diabetes mellitus – is vitamin B12 deficiency an accomplice? Med Hypotheses. 2021;146(2021):11037. doi: 10.1016/j.mehy.2020.110374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weir EK, Thenappan T, Bhargava M, Chen Y. Does vitamin D deficiency increase the severity of COVID-19? Clin Med (lond) 2020;20(4):e107–e108. doi: 10.7861/clinmed.2020-0301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wessels I, Maywald M, Rink L. Zinc as a gatekeeper of immune function. Nutrients. 2017;9(12):1286. doi: 10.3390/nu9121286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams RJ. Biochemical individuality, the basis for the genetotrophic concept. New York: Wiley & Sons Inc; 1956. [Google Scholar]
- Winterbourn CC, Kettle AJ, Hampton MB. Reactive oxygen species and neutrophil function. Annu Rev Biochem. 2016;85:765–792. doi: 10.1146/annurev-biochem-060815-014442. [DOI] [PubMed] [Google Scholar]
- Witter AR, Okunnu BM, Berg RE. The essential role of neutrophils during infection with the intracellular bacterial pathogen Listeria monocytogenes. J Immunol. 2016;197(5):1557–1565. doi: 10.4049/jimmunol.1600599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wong KK, Lee SWH, Kua KP. N-Acetylcysteine as adjuvant therapy for COVID-19 - a perspective on the current state of the evidence. J Inflamm Res. 2021;14:2993–3013. doi: 10.2147/JIR.S306849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu W, Liu Y, Zou X, Luo H, Wu W, Xia J, Chan MTV, Fang S, Shu Y, Wu WKK, Zhang L. Hypozincemia in COVID-19 patients correlates with stronger antibody response. Front Immunol. 2022;12:785599. doi: 10.3389/fimmu.2021.785599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi M, Mori S, Suketa Y. Effects of Ca2+, Zn2+ and Cd2+ on uridine diphosphate-glucuronyltransferase and beta-glucuronidase activities in rat liver microsomes. Chem Pharm Bull (tokyo) 1990;38(1):159–163. doi: 10.1248/cpb.38.159. [DOI] [PubMed] [Google Scholar]
- Yamanaka Y, Matsugano S, Yoshikawa Y, Orino K. Binding analysis of human immunoglobulin G as a zinc-binding protein. Antibodies (basel) 2016;5(2):13. doi: 10.3390/antib5020013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang WS, Stockwell BR. Ferroptosis: death by lipid peroxidation. Trends Cell Biol. 2016;26(3):165–176. doi: 10.1016/j.tcb.2015.10.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang C, Ma X, Wu J, Han J, Zheng Z, Duan H, Liu Q, Wu C, Dong Y, Dong L. Low serum calcium and phosphorus and their clinical performance in detecting COVID-19 patients. J Med Virol. 2021;93(3):1639–1651. doi: 10.1002/jmv.26515. [DOI] [PubMed] [Google Scholar]
- Yin W, Mao C, Luan X, Shen DD, Shen Q, Su H, Wang X, Zhou F, Zhao W, Gao M, Chang S, Xie YC, Tian G, Jiang HW, Tao SC, Shen J, Jiang Y, Jiang H, Xu Y, Zhang S, Zhang Y, Xu HE. Structural basis for inhibition of the RNA-dependent RNA polymerase from SARS-CoV-2 by remdesivir. Science. 2020;368(6498):1499–1504. doi: 10.1126/science.abc1560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zahra SA, Iddawela S, Pillai K, Choudhury RY, Harky A. Can symptoms of anosmia and dysgeusia be diagnostic for COVID-19? Brain Behav. 2020;10(11):e01839. doi: 10.1002/brb3.1839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zeng HL, Yang Q, Yuan P, Wang X, Cheng L. Associations of essential and toxic metals/metalloids in whole blood with both disease severity and mortality in patients with COVID-19. FASEB J. 2021;35(3):e21392. doi: 10.1096/fj.202002346RR. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zetterberg A, Engström W. Glutamine and the regulation of DNA replication and cell multiplication in fibroblasts. J Cell Physiol. 1981;108(3):365–373. doi: 10.1002/jcp.1041080310. [DOI] [PubMed] [Google Scholar]
- Zhang J, Taylor EW, Bennett K, Saad R, Rayman MP. Association between regional selenium status and reported outcome of COVID-19 cases in China. Am J Clin Nutr. 2020;111(6):1297–1299. doi: 10.1093/ajcn/nqaa095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou X, Chen D, Wang L, Zhao Y, Wei L, Chen Z, Yang B. Low serum calcium: a new, important indicator of COVID-19 patients from mild/moderate to severe/critical. Biosci Rep. 2020;40(12):BSR2020690. doi: 10.1042/BSR20202690. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou N, Yang X, Huang A, Chen Z. The potential mechanism of N-acetylcysteine in treating COVID-19. Curr Pharm Biotechnol. 2021;22(12):1584–1590. doi: 10.2174/1389201021999201228212043. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Enquiries about data availability should be directed to the author.