


Abstract
DNA topoisomerases are ubiquitous enzymes that govern the topological interconversions of DNA thereby playing a key role in many aspects of nucleic acid metabolism. Recently determined crystal structures of topoisomerase fragments, representing nearly all the known subclasses, have been solved. The type IB enzymes are structurally distinct from other known topoisomerases but are similar to a class of enzymes referred to as tyrosine recombinases. A putative topoisomerase I open reading frame from the kinetoplastid Leishmania donovani was reported which shared a substantial degree of homology with type IB topoisomerases but having a variable C-terminus. Here we present a molecular model of the above parasite gene product, using the human topoisomerase I crystal structure in complex with a 22 bp oligonucleotide as a template. Our studies indicate that the overall structure of the parasite protein is similar to the human enzyme; however, major differences occur in the C-terminal loop, which harbors a serine in place of the usual catalytic tyrosine. Most other structural themes common to type IB topoisomerases, including secondary structural folds, hinged clamps that open and close to bind DNA, nucleophilic attack on the scissile DNA strand and formation of a ternary complex with the topoisomerase I inhibitor camptothecin could be visualized in our homology model. The validity of serine acting as the nucleophile in the case of the parasite protein model was corroborated with our biochemical mapping of the active site with topoisomerase I enzyme purified from L.donovani promastigotes.
INTRODUCTION
DNA topoisomerases represent a family of DNA-processing enzymes that essentially solve the topological problems arising in cells during DNA replication, transcription, recombination, chromatin assembly and chromosome segregation (1–2). They are broadly classified into type I and type II topoisomerases. Type II enzymes are dimeric, cleaving both strands of a duplex and passing another region of intact DNA through this transient double-stranded break (in an ATP-dependent manner) resulting in changes in linking number in steps of two. In contrast, type I enzymes are monomeric, require no additional energy cofactor and they transiently nick a single strand of DNA duplex, allowing for single-step changes in the linking number of circular DNAs. Type I enzymes are further divided into type IA (bacterial) and IB (eukaryotic and vaccinia virus) enzymes. Type IA topoisomerases are able to relax only negatively supercoiled DNA, require magnesium and a single-stranded stretch of DNA for function and become transiently covalently attached to the 5′ end of the nicked single strand. However, topoisomerases IB are able to relax both positively and negatively supercoiled DNA with equal efficiency, do not require a single-stranded region of DNA or metal ions for function and form a transient covalent bond with the 3′ end of the nicked DNA strand. With the structural characterizations of fragments of Escherichia coli topoisomerase I (a type IA enzyme) (3,4), yeast topoisomerase I (type IB enzyme) (5), vaccinia topoisomerase (type IB enzyme) (6), and more recently determined structures of human topoisomerase I in complex with DNA (7–9) and of a catalytic fragment of vaccinia topoisomerase (10), it is clear that the type IA and IB topoisomerases share no structural similarity (11).
The fact that topoisomerases are proven drug targets (12,13) has led to the study of these enzymes from an array of parasitic organisms, so as to delineate common and distinguishing features between the host and parasite enzymes. Work on DNA topoisomerases from the kinetoplastid protozoan parasites (14) has been a major focus of interest in this regard. Recently, the DNA sequence of a topoisomerase I-like gene from the kinetoplastid Leishmania donovani has been reported (15). The deduced amino acid sequence of this gene showed an extensive degree of homology with the central core of other eukaryotic type IB topoisomerases, including several conserved motifs. Only the C-terminus of the parasite protein was markedly dissimilar from the C-terminal domain of type IB topoisomerases that normally contains the active site tyrosine within a conserved SKXXY motif (15). Prediction analysis using NetPhos indicated that within the C-terminal domain of this parasite protein, serine residues and not tyrosine had maximum probability of acting as potential phosphorylation sites for enzyme catalysis.
From our laboratory, we have previously reported ATP-independent, Mg-dependent type I topoisomerase activity from cell extracts of L.donovani promastigotes (16), which relaxes both positive and negative supercoils in topological steps of unity. The enzyme is sensitive to topoisomerase I-specific inhibitors like camptothecin (CPT) and is also a target for potential chemotherapy of anti-leishmanial drugs (17–19). The homogenously purified active enzyme is 67 kDa. The present work reports the mapping of the active site residue of the endogenously purified L.donovani topoisomerase I (LdTOP1) enzyme. The work also includes molecular modeling studies to gain an insight into the tertiary structure of the protein from the deduced amino acid sequence of the LdTOP1-like gene, its binary complex with the substrate DNA duplex and a ternary complex involving the enzyme, DNA and CPT, a potent inhibitor of the enzymatic process. The structure prediction and computational analysis was based on available crystal structures of human topoisomerase I. These studies indicate a common structural and functional theme for the human topoisomerase I and the parasite topoisomerase I-like gene product. So this work lends strong support to the existence of a topoisomerase I protein in the kinetoplastid protozoon L.donovani, which diverges from other eukaryotic type IB topoisomerases in the possession of a variable active site.
MATERIALS AND METHODS
Enzyme, DNA and chemicals
Type I topoisomerase was purified from L.donovani strain D1700 (MHOM/SD/68/SI) promastigotes as previously described by Chakraborty et al. (16). Rat liver topoisomerase I was purified according to a published protocol (20). Plasmid pHOT1 DNA (containing the high-affinity topoisomerase I cleavage site that is derived from the Tetrahymena ribosomal gene repeat, a hexadecameric sequence) was purchased from TopoGEN. The nick translation kit was purchased from Boehringer Mannheim (Roche). DNase I and Sephadex G-25 were from Amersham Pharmacia Biotech. Thin layer chromatography (TLC) was performed on silica gel plates (Macherey-Nagel Polygram SilG).
Biochemical analysis of covalent linkage through catalytic amino acid residue
A 50 µl reaction mixture containing 50 mM PIPES, pH 6.0, 75 mM KCl, 10 mM MgCl2, 0.5 mM DTT, 0.5 mM EDTA, 30 µg/ml BSA, 25 ng of [α-32P]dCTP nick-translated pHOT1 DNA and 80 U purified LdTOP1 enzyme was incubated at 37°C for 30 min. The reaction was terminated by adding NaOH to 0.2 M. DNase I treatment was done at a concentration of 100 µg/ml followed by further addition of 30 U Bal31 nuclease. Reactions were stopped by adding 10 mM EDTA and 0.5% SDS before processing the DNA–protein complexes through two cycles of a Sephadex G-25 spin column and acetone precipitation. Next, proteolysis was carried out with 50 µg/ml of proteinase K at 37°C for 30 min, the digested products vacuum dried and re-suspended in 30 µl of constant boiling with 6 M HCl. After removal of HCl, the dried hydrolysates were resuspended in 10 µl of H2O containing 200 nmol each of phosphoserine, phoshphothreonine and phosphotyrosine. A few microliters of the samples were spotted on the silica gel plate. Phosphoamino acid mix markers were also spotted on the same plate in the same horizontal line with the earlier spotted samples. TLC, ninhydrin staining and autoradiography of the resolved amino acids of the spots were carried out following published procedures (21).
Molecular modeling and structural analysis
Predictions of three-dimensional structures of topoisomerase I, its binary complex with DNA and ternary complex with DNA and CPT were done by knowledge-based homology modeling using InsightII 98.0 of MSI (Biosym Technologies), ABGEN (22) and our in-house package of MODELYN and ANALYN (23) in UNIX as well as in the MS Windows environment. Energy minimization and molecular dynamics were performed with the InsightII 98.0/Discover package using cff91 forcefield on a Silicon GraphicsR OCTANE workstation. Energy minimizations were done with a convergence criterion of 0.001 kcal/mol, using a combination of steepest descent and conjugate gradient methods of 100 steps each; these steps were repeated until satisfactory conformational parameters were obtained. Molecular dynamics simulations were carried out using a time step of 1 fs over a period of 0.1 µs. Distance constraints were applied to the hydrogen bonding of base pairs of the DNA duplex to keep them intact while running minimization and dynamics on the complexes. Secondary structures were predicted from sequences using SOPM and analyzed from structures using MODELYN and RIBBONS. The electrostatic potential surface of the protein model was determined by MOLMOL (24). TREEVIEW software package version 1.5 (25) was used for the construction of a phylogenetic tree, which read PHYLIP style treefiles produced by CLUSTAL W (26).
RESULTS
Active site mapping of the L.donovani topoisomerase I enzyme
A 67 kDa protein with topoisomerase I activity had been homogenously purified from the nuclear extracts of L.donovani promastigotes (16). From its enzymatic characteristics, the protein could be classified as an eukaryotic type I topoisomerase. When the LdTOP1 was incubated with supercoiled pHOT1 DNA followed by termination of reaction with SDS, nicked species of DNA were the main end products of the reaction, which became further enriched in the presence of CPT (19). Analysis of CPT-induced DNA breaks by an assay using end-labeled DNA (27) show that the Leishmania enzyme is covalently linked to the 3′ end of the broken DNA (data not shown). In order to investigate the chemical nature of the covalent linkage formed between the substrate DNA and the LdTOP1, biochemical mapping studies were carried out by labeling the purified enzyme as described in Materials and Methods, positive control being set up with a rat liver topoisomerase I. Acid hydrolysis of the labeled oligonucleotide–protein complexes yielded orthophosphate as the major labeled species, as shown by TLC analysis of the products (Fig. 1). In the case of the LdTOP1, a significant fraction of the radioactivity was found to co-chromatograph with chemically synthesized phosphoserine/phosphothreonine on thin-layer silica gel in the solvent system mentioned in the Materials and Methods. Under similar conditions, for the rat liver topoisomerase I, the radioactive spot was found to co-migrate with standard phosphotyrosine. Alkaline hydrolysis of the labeled protein led to the disappearance of the radioactive signal of the LdTOP1–oligo complex on TLC plates (data not shown), further confirming that the labeled amino acid is either alkali-labile phosphoserine or phosphothreonine. The signal of the mammalian topoisomerase I–oligo complex was found to be resistant to alkaline pH. Thus, the covalent linkage formed between the L.donovani enzyme and DNA was identified as Ser-P/Thr-P and not the usual Tyr-P as observed for other eukaryotic topoisomerase I (28).
Figure 1.
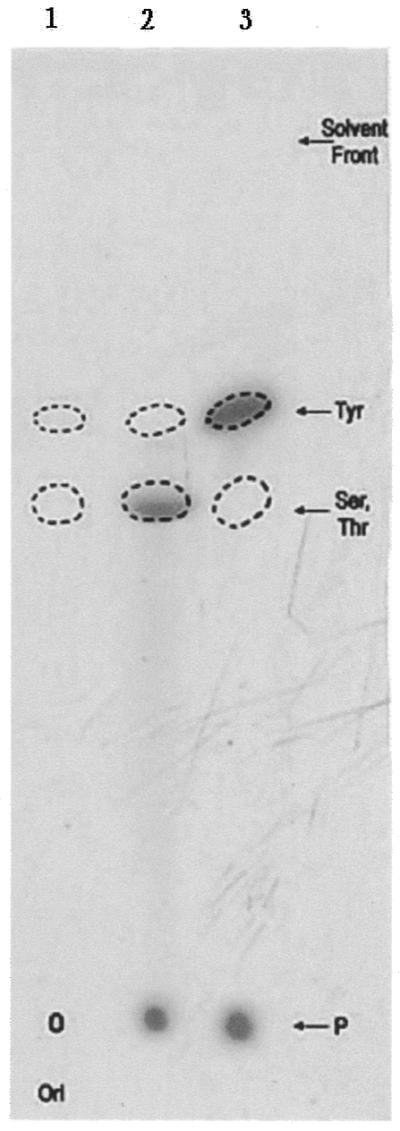
Identification of the phosphoamino acid linkage in the hydrolysate of topoisomerase I–DNA covalent complexes. Lane 1 shows the migration profile of the standard cold phosphoamino acids. Lanes 2 and 3 represent the migration of 32P-labeled acid hydrolysates from rat liver and L.donovani samples, respectively. The positions of the marker compounds (seen by ninhydrin staining) are indicated by dotted outlines. The point of application of samples, i.e. the origin (ori), as well as the solvent front run in the silica gel plate are marked.
Three-dimensional model of putative Leishmania topoisomerase I
Based on combined primary sequence and secondary structure alignments against the known crystal structures of human topoisomerase I (PDB accession nos 1A36, 1A35 and 1A31), a three-dimensional model (PDB accession no. 1JUW) has been presented here for a LdTOP1-like sequence. Like other eukaryotic type I enzymes, the human topoisomerase I is a monomeric protein and it contains 765 amino acids (29). Limited proteolysis studies of human topoisomerase I (30) indicate that it is comprised of four structural domains: a highly charged NH2-terminal domain, a major positively charged core domain, followed by a small linker domain and finally a COOH-terminal domain containing the active site residue. In the crystal structures of engineered catalytically active components of human topoisomerase I, the hydrophilic N-terminal (which is known to be dispensable for activity) had been removed and the protein was in a complex with a 22 bp blunt-ended duplex DNA oligonucleotide. Using these structures as a template, the approximate protein model of LdTOP1-like protein (PDB accession no. 1JUW) was simulated (Fig. 2A) in complex with an identical duplex DNA. The DNA sequence had the preferred consensus sequence of ACTT∧ at positions –4 to –1 in the scissile strand adjacent to the cleavage site (∧), that corresponded to the high affinity eukaryotic topoisomerase I binding site from Tetrahymena rDNA (31). The most reliable homology model generated for the parasite protein comprised 544 residues and showed an RMSD of 0.8 Å over Cα positions for 445 equivalent residues of the human topoisomerase I structure, 1A36, the superposition being depicted in Figure 3. The Leishmania model appeared to be a bi-lobed protein that clamps completely around its DNA substrate. One lobe (comprising two domains resembling the core subdomains I and II of human topoisomerase I) sits on top of the DNA duplex (as oriented in Fig. 2A) just like the ‘cap’ structure referred to in the human protein (32). It is composed of mixed α and β secondary-structural elements and contains two unique ‘nose-cone’ helices that extend 25 Å from the body of the molecule (Fig. 2B). The surfaces of these helices, which are proximal to the DNA, are highly positively charged and are positioned above the DNA major groove, but do not directly interact with the DNA. The second lobe (also comprising two domains; the former resembling the core subdomain III of human topoisomerase I and the latter representing a short variable C-terminal domain) sits below the DNA, is composed of an all α-helical structure except for one three-stranded β-sheet (Fig. 2B) and contains the catalytic residues implicated in the strand cleavage and religation reactions. These two lobes are covalently linked through a continuous helical chain on one side of the DNA molecule labeled the ‘connector’, and touch each other in a small contact region, similar to the ‘lip’ structure in the human topoisomerase I (32). The coiled-coil linker domain extends 50 Å from the body of the second lobe and exposes a highly positively charged face towards the DNA similar to the ‘nose-cone’ helices; however, this domain also fails to make direct interactions with the DNA.
Figure 2.

Ribbon representation of the LdTOP1 model (PDB accession no. 1JUW). (A) The homology model of LdTOP1 in complex with a 22 bp DNA duplex (31) shown in sticks. The LdTOP1 model is colored domain-wise. Core subdomains I, II and III are colored yellow, blue and red, respectively. The linker and C-terminal domain are shown in orange and green, respectively. The part rendered in dark red represents the missing part in the human topoisomerase I crystal structure. (B) The homology model of LdTOP1 depicting its secondary structures: α-helices rendered in sky blue, β-sheets in yellow and random coils in orange. The N- and C-termini of the modeled protein are marked. The length of the nose-cone helices and linker are depicted in angstroms. Also marked are the ‘connector’- and ‘cap’-like structures.
Figure 3.

Superposition of the modeled LdTOP1 (green) on human topoisomerase I (red), both proteins being depicted in Cα trace.
Protein–DNA contacts in the Leishmania protein–DNA model
The model of the Leishmania protein in complex with topoisomerase I–suicide DNA substrate reveals how the parasite enzyme can bind to the DNA and align it for strand cleavage and covalent attachment. In fact, the Leishmania protein is observed to contact the DNA (Fig. 4) mostly via protein to DNA phosphate interactions just like the human topoisomerase I (8,11). Only four protein–DNA contacts derive from two residues, Arg190 and Lys352, which enter the minor groove of the DNA, adjacent to the cleavage site. One of these contacts is base specific, i.e. the hydrogen bond between the side chain NH2 group of Lys352 and the oxygen atom of the favored –1 thymidine base. This lysine residue is conserved at this position in all known sequences of type IB topoisomerases. This interaction explains the observed preference for a thymidine at the –1 position adjacent to the cleavage site, as thymidine is the only base with a hydrogen bond acceptor freely available at this position in the minor groove. The parasite enzyme even exhibits a distinct asymmetry in its contact with the DNA (Fig. 4) like the human topoisomerase I, interacting more frequently with the 5 bp upstream of the cleavage site than it does with an 8 bp region downstream of the cleavage site. The electrostatic potential surface of the Leishmania protein model that plays an important role in its circumferential binding around the negatively charged DNA duplex has been illustrated in Figure 5.
Figure 4.

Schematic representation of the protein–DNA contacts of modeled LdTOP1 with the 22 bp DNA duplex. Major interactions within distances of 3.5 Å are depicted. The phosphate-deoxyribose backbone of the intact strand is shown in blue, and that of the scissile strand is shown in magenta to indicate regions upstream or downstream of the cleavage site. The observed interactions are limited exclusively to protein–phosphate interactions (large gray dots). The –1 and +2 bases are the only bases that are contacted by the protein, as indicated with bold black boxes. Both contacts occur in the minor groove, one between the Lys352 and O2 atom of –1 thymidine and the other between the Arg190 and N3 atom of the guanidine base. Small red dots indicate interactions between Arg190 and the ribose oxygens in positions +2 and +3 on the cleaved strand. The cleavage site, the catalytic triad of Arg314, Arg410 and His453 plus the serine nucleophile (Ser553) contacting the scissile phosphate at the active site are marked.
Figure 5.

Surface representations of the Leishmania protein model (MOLMOL) revealing the prominent groove where DNA duplex can bind. (A) Region spanning core subdomains I and III including the putative hinge of the protein. (B) Part of subdomain III, linker and C-terminal domain. The surface is colored to represent electrostatic potential (–15kbT/e to +15kbT/e) where kbT is the product of the Boltzmann constant and temperature and e is the electronic charge, with negatively and positively charged regions shown in red and blue, respectively.
Putative active site view of the Leishmania protein homology model
The model of the LdTOP1-like gene provides a view of the detailed architecture of the proposed active site of the enzyme (Fig. 6). For eukaryotic type IB topoisomerases the DNA strand cleavage reaction usually proceeds via a nucleophilic attack on the scissile phosphorus atom by the catalytic residue. Interestingly, sequence analysis revealed the absence of the usual SKXXY motif within the C-terminal domain of the parasite topoisomerase I-like open reading frame and a serine (Ser553) aligned with the conserved Tyr723 catalytic residue of the human enzyme. The Ser553 was found to be positioned perfectly for nucleophilic attack and subsequently for covalent attachment to the 3′ end of the broken strand because it is within suitable distance of the O5′-P scissile bond. Other residues near the phosphodiester bond that is cleaved are two arginine and one histidine residue found to be conserved with the human topoisomerase I enzyme in structurally equivalent positions. For the catalytic mechanism of LdTOP1, it seems reasonable to propose that this residue triad contributes to the stabilization of the pentavalently co-ordinated transition state intermediate through hydrogen-bonding; Arg314 and Arg410 lying within hydrogen-bonding distance of one non-bridging oxygen, O1, and His453 lying near the other non-bridging oxygen, O2. In the LdTOP1 model, there is no side-chain atom within a 4 Å radius of the hydroxyl of Ser553 to act as a general base for activating the nucleophilic oxygen. The closest such amino acid is His453, which is unlikely to be involved as a general base since the Nɛ2 atom of His453 is 4.5 Å away from the hydroxyl oxygen of Ser553. Analogously the Nɛ2 atom of His632 is 4.3 Å away from what would be the hydroxyl oxygen of Tyr723 in the crystal structure of the reconstituted human topoisomerase I non-covalent complex. A possible explanation drawn from analogy with the human structure is that the proton of the nucleophile might be abstracted by a hypothetical water molecule as catalysis proceeds. Thus, analysis of the position of participating catalytic residues in the Leishmania protein model is compatible with the human topoisomerase I crystal structure in many aspects but deviating from the host enzyme in its possession of a serine in place of the universal tyrosine. A view of the C-terminal domains of the human topoisomerase I with its catalytic Tyr723 and the Leishmania protein with its putative catalytic Ser553 is presented in Figure 7.
Figure 6.

Detailed architecture of the active site of putative LdTOP1. (A) Structural view of the amino acid residues in the putative active site of the Leishmania protein model. The three-dimensional relationship between the side chains of residues Arg314, Arg410, His453, Lys352 and Ser553 and the +1 and –1 residue belonging to the scissile strand of DNA is shown. The depiction is in solid sticks (carbon, green; nitrogen, blue; oxygen, red; and phosphorus, pink). Hydrogen bonds (dotted lines) and bond distances (in angstroms) are shown. (B) Schematic diagram depicting a possible intermediate stage in the cleavage phase of catalysis by the putative LdTOP1. The active site residues Arg314, Arg410 and His453, from subdomain III as well as the putative catalytic Ser553 of the C-terminal domain, are depicted surrounding the scissile phosphate (+1), which is coordinated in a trigonal bipyramidal fashion.
Figure 7.

Close-up views of the active site pocket of human topoisomerase I (A) and the putative LdTOP1 (B). The proteins are depicted in ribbon representation, coloring being according to amino acid sequence. The positions of the nucleophilic tyrosine (Phe723Tyr) and serine (Ser553) are marked, respectively, in the active sites of human topoisomerase I (A) and the putative LdTOP1 model (B).
Proposed mechanism of topoisomerization by Leishmania-modeled protein
The spatial arrangement of the DNA in the model of LdTOP1, based on the human topoisomearse I co-crystal structure, renders unlikely an enzyme-bridging mechanism for DNA relaxation analogous to what has been proposed for Type IA topoisomerase (Escherichia coli). Stewart et al. (8) proposed a ‘controlled rotation’ mechanism for the relaxation of DNA superhelical tension by type IB topoisomerases from observations concerning the nature of the human topoisomerase I–DNA interactions. Many of these mechanistic features could be visualized in the homology model of LdTOP1–DNA complex. An alternative hypothesis is shown in Figure 8, where it is proposed that once cleavage has occurred, the Leishmania enzyme opens up at the putative hinge region located at the top of the connector in subdomain III to allow rotation and perhaps assumes a conformation approximating that proposed for the DNA-free form of the enzyme. Opening and closing of the protein clamp during DNA binding and release must involve the breaking of the connection between the lips and the lifting of the cap away from the base. In this model, there may be little or no hindrance to DNA rotation, DNA binding being naturally directed by the surface and charge complementation. Either model could accommodate multiple rotation events before religation of the DNA.
Figure 8.

A possible mechanism of topoisomerization by the Leishmania protein. The bilobed protein model (space-filling) is colored blue for the ‘cap’ lobe (subdomains I and II) and yellow for the ‘catalytic’ lobe (subdomain III, linker and the C-terminal domain) the view being down the axis of the bound DNA. The DNA is represented as space-filling and colored according to usual atom colors. (A) The modeled closed form of the Leishmania protein presumably catalyzes cleavage of the scissile strand upon binding the DNA. (B) The parasite protein modeled into an open configuration for DNA binding by allowing upward movement at the putative hinge point.
Proposed mode of binding of CPT to the Leishmania protein–DNA covalent complex
Topoisomerase I is the sole intracellular target for a well-studied class of compounds, the CPTs, which have emerged as potent anticancer drugs and which act by stabilizing the covalent topoisomerase I–DNA complex. CPT inhibition of LdTOP1 has been reported (19). Recently, some modes of binding of CPT to the covalent complex of human topoisomerase I and DNA have been proposed (7,33,34). The model of Redinbo et al. (7) places the CPT molecule into the DNA duplex in a position that partially overlaps with the +1 guanine of the scissile strand; this placement requires that the guanine base be flipped out of the duplex and stacked on the planar CPT. Our results on placing the CPT in an identical manner with the modeled Leishmania protein–DNA covalent complex highlighted that this alkaloid exerts an analogous mode of action on this parasite protein. In this model (Fig. 9), the immutable lactone and 20(S)-hydroxyl moieties of CPT are found to interact with Arg190 and Asp353 and form a stacking interaction with guanine (favored base in the scissile +1 position for CPT action) such that the guanine N-3 is in close proximity to the C-7 of CPT. The amino acid residues whose mutation renders the human topoisomerase I insensitive to CPT could also be identified as important in the Leishmania protein, namely Arg190 and Asp353, equivalent to Arg364 and Asp533 of human topoisomerase I. Only the stacking interaction of CPT with Asn722 of human topoisomerase I was missing in the parasite enzyme. The important feature highlighted in the model of the ternary complex is that the re-entry of the +1 guanine into the DNA duplex is hindered by the stacked CPT and that the 5′-hydroxyl of the dG(+1) is prevented from attacking the enzyme–DNA phosphate bond (phosphoserine in this case) thereby explaining the inhibition of topoI-mediated DNA religation.
Figure 9.

(A) Model of the inhibitor CPT bound to the covalent complex of putative LdTOP1 and DNA. In this model, CPT (space-filling representation with nitrogen atoms in blue, oxygen in red, carbon in green and the A, B and E rings of the molecule indicated) stacks in the DNA duplex in place of the guanine base at the +1 Gua position of the scissile strand, which has been flipped out of the duplex. CPT makes a stacking interaction with this +1 base bringing the C7 atom of the drug in close proximity to the N3 atom of +1 Gua. Two of the CPT-resistant mutations reported in human, represented by Arg190 and Asp353 in the parasite protein model, were found to interact in an analogous manner with the required lactone and 20(S)-hydroxyl moieties of CPT. (B) A schematic representation of the key hydrogen bonds and ring-stacking interactions made between the LdTOP1–DNA covalent complex model and CPT in the proposed CPT-binding mode is shown.
DISCUSSION
With both rat liver and L.donovani DNA topoisomerase I enzymes, formation of the covalent complex with 32P-labeled supercoiled DNA, followed by nucleolytic treatment of the complex, yield a 32P-labeled protein. Analysis of the product after hydrolysis and TLC identified Tyr-P as the labeled amino acid in the case of rat liver topoisomerase I indicating that the covalent bond that forms between DNA and the rat liver enzyme is phosphotyrosine, like topoisomerase enzymes from other species. However, similar analysis of the chemical nature of the labeled LdTOP1–DNA products indicated that the covalent linkage is most likely a phosphoserine/phosphothreonine and not a phosphotyrosine.
Recently, Brocoli et al. (15) reported a sequence from L.donovani that codes for a putative topoisomerase I protein, identical to other topoisomerase I sequences deposited in the GenBank. The predicted protein analysis which uses MAXHOM multiple sequence alignment aligns the query sequence of putative LdTOP1 with different eukaryotic type IB DNA topoisomerase sequences with an identity score of 37% and a similarity score of 49% with the human topoisomerase I. This homology allowed us to build a three-dimensional model of the putative LdTOP1 protein based on the crystal structure of human topoisomerase I (PDB accession no. 1A36). The model predicts that the parasite protein adopts an overall conformation similar to that of the human topoisomerase I, organized into four subdomains. The superposition of the three-dimensional model on the human topoisomerase I structure clearly showed that most of the secondary structural elements are conserved in both the proteins and that they are largely present in the same relative positions. Other interesting features of the Leishmania protein model are the presence of ‘connector’-, ‘cap’- and ‘lip’-like structures similar to that of the human topoisomerase I. The majority of the protein–DNA contacts in our model were found to be equivalent to those in the human topoisomerase I–DNA co-crystal structure, the protein contacting only the central 10 bp of the DNA (–4 to +6). Topoisomerization by the human enzyme could also be visualized in the open and closed modeled conformations of the Leishmania protein, with the hinge-bending movement being controlled by the electrostatic charged surface of the protein.
The predicted active site pocket of the Leishmania protein model is located in a structurally equivalent position to that of the crystallized human topoisomerase I. The four catalytic residues conserved in the human protein including Arg488, Arg590, His632 and Lys532 cluster in the core subdomain III. Mutagenesis studies have indicated that the above two arginine and one histidine residues play an essential role in the nicking closing reaction. The analogous residues in domain III of the Leishmania protein model are Arg314, Arg410, His453 and Lys352. However, the striking feature inferred from analysis of the C-terminal domain of our model is that the reactive tyrosine found in the human topoisomerase I C-terminus was missing in the structurally equivalent active site pocket of the Leishmania protein. Instead, the C-terminal domain of our model was found to harbor a serine residue (Ser553) within a suitable distance of the 5′-OH of the scissile bond in the modeled topoisomerase I–suicide DNA substrate complex. The parasite protein ternary complex model with the CPT inhibitor could account for the interaction with two conserved residues as in human. Interestingly, the stacking interaction with asparagine (Asn722 in human) was absent in the Leishmania model. Ala552 and Ser553 of the parasite protein were found to be occupying positions analogous with Asn722 and Tyr723, respectively, of human topoisomerase I. This unique presence of a serine in our structural model at the position occupied by tyrosine in the human enzyme points towards serine acting as the reactive nucleophile for the parasite enzyme. This observation is strongly supported by identification of Ser-P/Thr-P as the protein–DNA linkage in the covalent complex between LdTOP1 and DNA by biochemical mapping studies.
Broadly, type I topoisomerases and site-specific recombinases are believed to be derived from a common ancestral strand transferase (35). Just as there are two families of site-specific recombinases, one using a tyrosine nucleophile-based DNA catalysis, e.g. tyrosine recombinases like λ integrase (36), while the other performs a serine nucleophile-mediated strand cleavage reaction, e.g. the resolvase-invertase group that includes E.coli γδ resolvase (37); the kinetoplastid protozoan parasite L.donovani possesses a topoisomerase I enzyme with a reactive serine unlike the topoisomerase IB enzymes of other eukaryotic members from yeast to human, that use a catalytic tyrosine.
Since DNA topoisomerase I genes code for ubiquitous proteins, their alignments and comparative analyses can form a strong base for molecular phylogeny. A phylogenetic tree has been constructed (Fig. 10) using the LdTOP1-like sequence and representative topoisomerase I sequences from other eukaryotes including yeast to human. The tree indicates that the kinetoplastid protozoan parasite L.donovani diverged very early from the main line of eukaryotic evolution. The absence of the conserved motif surrounding the active site in the C-terminal domain of LdTOP1 also points to this direction. A possible hypothesis is that this conserved motif in topoisomerase I proteins encompassing the active site tyrosine might be a later acquisition in higher eukaryotes.
Figure 10.

Phylogenetic tree using the amino acid sequences of type I topoisomerases from L.donovani and other organisms. The phylogram is displayed on TREEVIEW that reads PHYLIP format treefiles under the CLUSTAL W program. The evolutionary scale bar is shown on the left, which indicates the relative distance on the tree in arbitrary units.
A future aspect of this work lies in exploiting the differences between the active site domain of host and parasite enzymes by integrating structural information with biochemical experimentation. Such information will provide crucial insights for designing newer drugs with selective toxicity for the parasitic enzyme. This drug screening system would, thereby, form an exciting base for developing antiparasitic chemotherapy.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Professor S. Bhattacharyya, the Director of our Institute, for his interest in this work. Purified rat liver topoisomerase I was kindly provided by A. Roychowdhury. S. Mandal is thanked for the assistance in TLC of samples. This work is supported by a grant (BT/PRO493/MED/09/096/96) from the Department of Biotechnology, Government of India.
PDB accession no. 1JUW
REFERENCES
- 1.Champoux J.J. (1990) Mechanistic aspects of type-I topoisomerases. In Wang,J.C. and Cozzarelli,N.R. (eds), DNA Topology and Its Biological Effects. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 217–242.
- 2.Wang J.C. (1996) DNA topoisomerases. Annu. Rev. Biochem., 65, 635–692. [DOI] [PubMed] [Google Scholar]
- 3.Lima C.D., Wang,J.C. and Mondragon,A. (1994) Three-dimensional structure of the 67 kD N-terminal fragment of E. coli DNA topoisomerase I. Nature, 367, 138–146. [DOI] [PubMed] [Google Scholar]
- 4.Yu L., Zhu,C.X., Tse-Dinh,Y.C. and Fesik,S.W. (1995) Solution structure of the C-terminal single-stranded DNA-binding domain of Escherichia coli topoisomerase I. Biochemistry, 34, 7622–7628. [DOI] [PubMed] [Google Scholar]
- 5.Lue N., Sharma,A. and Mondragon,A. (1995) A 26 kDa yeast DNA topoisomerase I fragment: crystallographic structure and mechanistic implications. Structure, 3, 1315–1322. [DOI] [PubMed] [Google Scholar]
- 6.Sharma A., Hanai,R. and Mondragon,A. (1994) Crystal structure of the N-terminal fragment of vaccinia virus DNA topoisomerase I. Structure, 2, 767–777. [DOI] [PubMed] [Google Scholar]
- 7.Redinbo M.R., Stewart,L., Kuhn,P., Champoux,J.J. and Hol,W.G.J. (1998) Crystal structure of human topoisomerase I in covalent and noncovalent complexes with DNA. Science, 279, 1504–1513. [DOI] [PubMed] [Google Scholar]
- 8.Stewart L., Redinbo,M.R., Qiu,X., Hol,W.G.J. and Champoux,J.J. (1998) A model for the mechanism of human topoisomerase I. Science, 279, 1534–1541. [DOI] [PubMed] [Google Scholar]
- 9.Redinbo M.R., Stewart,L., Champoux,J.J. and Hol,W.G.J. (1999) Structural flexibility in human topoisomerase I revealed in multiple non-isomorphous crystal structures. J. Mol. Biol., 292, 685–696. [DOI] [PubMed] [Google Scholar]
- 10.Cheng C., Kussie,P., Paveltich,N. and Shuman,S. (1998) Conservation of structure and mechanism between eukaryotic topoisomerase I and site-specific recombinases. Cell, 92, 841–850. [DOI] [PubMed] [Google Scholar]
- 11.Redinbo M.R., Champoux,J.J. and Hol,W.G.J. (1999) Structural insights into the function of type IB topoisomerases. Curr. Opin. Stuct. Biol., 9, 29–36. [DOI] [PubMed] [Google Scholar]
- 12.Schneider E., Hsiang,Y.H. and Liu,L.F. (1991) DNA topoisomerases as anticancer drug targets. Adv. Pharmacol., 21, 149–183. [DOI] [PubMed] [Google Scholar]
- 13.Wang J.C. (1994) DNA topoisomerases as targets of therapeutics: an overview. Adv. Pharmacol., 29A, 1–19. [DOI] [PubMed] [Google Scholar]
- 14.Shapiro T.A. (1993) Inhibition of topoisomerases in African trypanosomes. Acta Trop., 54, 251–260. [DOI] [PubMed] [Google Scholar]
- 15.Broccoli S., Marquis J.-F., Papadopoulou,B., Olivier,M. and Drolet,M. (1999) Characterization of a Leishmania donovani gene encoding a protein that closely resembles a type IB topoisomerase. Nucleic Acids Res., 27, 2745–2752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chakraborty A.K., Gupta,A. and Majumder,H.K. (1993) A type 1 DNA topoisomerase from the kinetoplast hemoflagellate Leishmania donovani. Ind. J. Biochem. Biophys., 30, 257. [PubMed] [Google Scholar]
- 17.Ray S., Majumder,H.K., Chakravarty,A. and Mukhopadhyay,S. (1996) Amarogentin, a naturally occurring secoiridoid glycoside and a newly recognised inhibitor of topoisomerase I from Leishmania donovani. J. Nat. Prod., 59, 27–29. [DOI] [PubMed] [Google Scholar]
- 18.Ray S., Sadhukhan,P.K., Mandal,N.B., Mahato,S.B. and Majumder,H.K. (1997) Dual inhibition of DNA topoisomerases of Leishmania donovani by novel indolyl quinolines. Biochem. Biophys. Res. Commun., 230, 171–175. [DOI] [PubMed] [Google Scholar]
- 19.Ray S. Hazra,B., Mittra,B.M., Das,A. and Majumder,H.K. (1998) Diospyrin, a bisnapthoquinone; a novel inhibitor of type I DNA topoisomerase of Leishmania donovani. Mol. Pharmacol., 54, 994–999. [DOI] [PubMed] [Google Scholar]
- 20.Champoux J.J. and McConaughy,B.L. (1976) Purification and characterization of the DNA untwisting enzyme from rat liver. Biochemistry, 15, 4638–4642. [DOI] [PubMed] [Google Scholar]
- 21.Tse Y.-C., Kirkegaard,K. and Wang,J.C. (1980) Covalent bonds between protein and DNA. J. Biol. Chem., 255, 5560–5565. [PubMed] [Google Scholar]
- 22.Mandal C., Kingery,B.D., Anchin,J.M., Subramanium,S. and Linthicum,D.S. (1996) ABGEN: a knowledge based automated approach for antibody structure modeling. Nat. Biotechnol., 14, 323–328. [DOI] [PubMed] [Google Scholar]
- 23.Mandal C. (1998) MODELYN—A Molecular Modelling Program. Version PC-1.0, Indian Copyright No. 9/98.
- 24.Koradi R., Billeter,M. and Wuthrich,K. (1996) MOLMOL: a program for display and analysis of macromolecular structures. J. Mol. Graph., 14, 51–55, 29–32. [DOI] [PubMed] [Google Scholar]
- 25.Thompson J.D., Higgins,D.G. and Gibson,T.J. (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22, 4673–4680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Page R.D.M. (1996) TREEVIEW: an application to display phylogenetic trees on personal computers. Comput. Appl. Biosci., 12, 357–358. [DOI] [PubMed] [Google Scholar]
- 27.Hsiang Y.-H., Hertzberg,R., Hecht,S. and Liu,L.F. (1985) Camptothecin induced protein-linked DNA breaks via mammalian DNA topoisomerase I. J. Biol. Chem., 260, 14873–14878. [PubMed] [Google Scholar]
- 28.Eng W.K., Pandit,S.D. and Sternglanz,R. (1989) Mapping of the active site tyrosine of eukaryotic DNA topoisomerase I. J. Biol. Chem., 264, 13373–13376. [PubMed] [Google Scholar]
- 29.D’Arpa P., Machlin,P.S., Ratrie,H.,III, Rothfield,N.F., Cleveland,D.W. and Earnshaw,W.C. (1988) cDNA cloning of human DNA topoisomerase I: catalytic activity of a 67.7 kDa C-terminal fragment. Proc. Natl Acad. Sci. USA, 85, 2543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Stewart L., Ireton,G.C., Parker,L.H., Madden,K.R. and Champoux,J.J. (1996) Topoisomerase I has a strong binding preference for a conserved hexadecameric sequence in the promoter region of the rRNA gene from Tetrahymena pyriformis. J. Biol. Chem., 271, 7593.8631793 [Google Scholar]
- 31.Andersen A.H., Gocke,E., Bonven,B.J., Neilsen,O.F. and Westergaard,O. (1985) Topoisomerase I has a strong binding preference for a conserved hexadecameric sequence in the promoter region of the rRNA gene from Tetrahymena pyriformis. Nucleic Acids Res., 13, 1543–1557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Champoux J.J. (2001) DNA topoisomerases: structure, function and mechanism Annu. Rev. Biochem., 70, 369–413. [DOI] [PubMed] [Google Scholar]
- 33.Fan Y., Weinstein,J.N., Kohn,K.W., Shi,L.M. and Pommier Y. (1998) Molecular modelling studies of the DNA topoisomerase I ternary cleavable complex with camptothecin. J. Med. Chem., 41, 2216–2226. [DOI] [PubMed] [Google Scholar]
- 34.Kerrigan J.E. and Pilch,D.S. (2001) A structural model for the ternary cleavable complex formed between human topoisomerase I, DNA and camptothecin. Biochemistry, 40, 9792–9798. [DOI] [PubMed] [Google Scholar]
- 35.Cheng C., Kussie,P., Pavletic,N. and Shuman,S. (1998) Conservation of structure and mechanism between eukaryotic topoisomerase I and site-specific recombinases. Cell, 92, 841–850. [DOI] [PubMed] [Google Scholar]
- 36.Kwon H.J., Tirumalai,R., Landy,A. and Ellenberger,T. (1997) Flexibility in DNA recombination: structure of the λ integrase catalytic core. Science, 276, 126–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Yang W. and Steitz,T.A. (1995) Crystal structure of the site-specific recombinase γδ resolvase complexed with a 34 bp cleavage site. Cell, 82, 193–207. [DOI] [PubMed] [Google Scholar]