

Abstract
p300 acts as a transcription coactivator and an acetyltransferase that plays an important role in tumourigenesis and progression. In previous studies, it has been confirmed that p300 is an important regulator in regulating the evolution of malignant tumours and it also has extensive functions. From the perspective of non-posttranslational modification, it has been proven that p300 can participate in regulating many pathophysiological processes, such as activating oncogene transcription, promoting tumour cell growth, inducing apoptosis, regulating immune function and affecting embryo development. In recent years, p300 has been found to act as an acetyltransferase that catalyses a variety of protein modification types, such as acetylation, propanylation, butyylation, 2-hydroxyisobutyration, and lactylation. Under the catalysis of this acetyltransferase, it plays its crucial tumourigenic driving role in many malignant tumours. Therefore, the function of p300 acetyltransferase has gradually become a research hotspot. From a posttranslational modification perspective, p300 is involved in the activation of multiple transcription factors and additional processes that promote malignant biological behaviours, such as tumour cell proliferation, migration, and invasion, as well as tumour cell apoptosis, drug resistance, and metabolism. Inhibitors of p300 have been developed and are expected to become novel anticancer drugs for several malignancies. We review the characteristics of the p300 protein and its functional role in tumour from the posttranslational modification perspective, as well as the current status of p300-related inhibitor research, with a view to gaining a comprehensive understanding of p300.
Keywords: p300, acetyltransferase, tumours, modification, drug resistance, inhibitor
1. Introduction
Since it was discovered that p300 plays a role as an auxiliary transcription activator, many researchers have begun to focus on p300, specifically its role in tumours. p300 can function as a cancer-promoting factor to regulate various biological functions and the malignant progression of various cancers, including lung cancer, stomach cancer, colorectal carcinoma, oesophageal squamous cell carcinoma (ESCC), pancreatic cancer and prostate cancer [1,2,3,4,5,6,7]. In addition, p300 can play an antitumour role in epidermal tumours, osteosarcoma, myelodysplastic syndrome (MDS)-associated leukaemia and human papilloma virus (HPV)-positive head and neck squamous cell carcinoma (HNSCC) [8,9,10,11], and tumours with high EP300 mutation rates have enhanced antitumour immunity [12]. p300 also participates in the regulation of some nontumour diseases and processes, including viral infections [13], chronic constriction injury (CCI) [14], and some cardiovascular diseases and processes (hypertension [15], heart development and heart ageing [16]).
p300 has extensive functions, and it is a common hub of many important biological pathways [17]. For example, p300 can participate in the regulation of many signalling pathways related to p53 [18], the oestrogen regulatory pathway [19], the ATR-CHK1 axis [20] and the cAMP pathway [21]. p300 can also bind to other proteins, such as HIF-α [22], β-catenin [23], p53 [24], TR3 [25], Myb [26] and Smad2/Smad3 [27], and play integral roles in complex physiological and pathological processes. Moreover, p300 expression is modulated by other molecules, such as RAR and ATRA [28]. There is a new global focus on the acetyltransferase activity of p300, and it has been recognised that p300 plays a crucial role in the malignant development of tumours by mediating the development of different types of modifications of histones and nonhistone proteins; these functions have presented new opportunities to expand the knowledge on p300 functions.
2. The Structure of the p300 Protein
p300 was first discovered in 1986 and is a protein with a molecular weight of 300 kDa. p300 is also the nuclear binding target of the adenovirus E1A cancer protein. It has high structural homology with CBP, with an approximately 70% homologous sequence. CBP is an important protein that can bind to cAMP reaction element binding protein (CREB). p300 and CBP have similar but different functions; for example, p300 is mainly involved in transcription and mediates posttranslational modification, while CBP mainly plays a role in cell transformation. The p300 structural domain mainly includes the N-terminal nuclear receptor interaction domain (NRID), transcription adapter zinc finger 1 (TAZ1) and the kinase-induced CREB interaction (KIX) domain [29,30]. The HAT domain is very important for the formation of the transcription complex. Furthermore, it also contains a bromine domain (BD): this domain is the key to the auxiliary transcription function and is involved in the interaction of acetylated modified proteins and the regulation of different physiological and pathological processes; it is also involved in the interaction of the RING domain with the plant homologous domain (PHD), which leads to interactions with the ZZ-type zinc finger domain, TAZ2 domain and IBiD domain [31,32,33], as shown in Figure 1.
Figure 1.

Schematic of the basic structure of p300, which contains the N-terminal nuclear receptor interaction domain (NRID), transcriptional junction zinc finger 1 (TAZ1), kinase-induced CREB interaction domains (KIX), bromine domain (BD), RING domain, plant homology domain (PHD), HAT domain, Z-type zinc finger domain (ZZ), TAZ2 domain and IBiD domain.
3. p300 and Posttranslational Protein Modification
Posttranslational modification of proteins, a form of posttranslational protein processing, involves covalent binding of a small molecular group or a small protein to a specific position in the amino acid side chain of a protein or enzymatic digestion of a protein that causes a break in the covalent bonds of its main chain [34]. The types of posttranslational protein modifications include oxidative modification [35], phosphorylation [36], ubiquitination [37], methylation [38], formylation [39], acetylation [40], propionylation [41], malonylation [42], butyrylation [43], glutarylation [44], succinylation [45], crotonylation [46], and lactylation [47]. Posttranslational protein modifications are associated with different physiological states and the onset of multiple diseases. The functions of posttranslational modifications are diverse and can include the regulation of transcriptional processes, protein–protein interactions, protein localisation and malignant tumour progression.
In particular, p300 can participate in the regulation of posttranslational protein modification by acting as an acetyltransferase and catalyses different types of specific protein modifications, including acetylation, propionylation, butyrylation, n-butyrylation, isobutyrylation, β-hydroxybutyrylation, 2-hydroxyisobutyrylation, crotonylation, isonicotinylation, and lactylation [43,47,48,49,50,51,52,53,54,55,56] (Table 1). p300 itself can also undergo autoacetylation, propionylation, and butyrylation [57,58], and it plays an indispensable role in a variety of physiological and pathological processes, one of which is regulating the malignant progression of tumours. The following figure shows the experimental schematic diagram of mass spectrometry detecting a protein’s posttranslation modification (Figure 2).
Table 1.
Summary of common posttranslational modification types, modification sites, regulatory factors, enrichment modes, and mass shifts in mass spectrometry.
| PTM | Modification Sites | Regulatory Factors | Enrichment Modes | Mass Shif (Da) |
|---|---|---|---|---|
| phosphorylation | serine, threonine, tyrosine | protein kinase; protein phosphatase | titanium dioxide | 80 |
| methylation | lysine, arginine | PKMTs, PRMTs | lysine methylation antibody, arginine methylation antibody | 14, 28, 42 |
| oxidative modification | methionine, tryptophan | ROS | oxPTMs antibody | 4, 16, 32 |
| ubiquitination | lysine | Ubiquitin activator E1, ubiquitin binding enzyme E2 and ubiquitin ligase E3 | lysine ubiquitination antibody | >1000, 114 after trypsinisation |
| acetylation | lysine | p300; SIRT1, SIRT3, SIRT6 | lysine acetylated antibody | 42 |
| propionylation | lysine | p300, NATs (GCN5, PCAF), MYSTs (MOF, MOZ, HBO1); SIRT1-3 | lysine propionylation antibody | 56 |
| butyrylation | lysine | p300, HBO1; SIRT1-3 | lysine butyrylation antibody | 70 |
| malonylation | lysine | SIRT5 | lysine malonylation antibody | 86 |
| glutarylation | lysine | p300, GNATs (GCN5); | lysine glutarylation antibody | 114 |
| SIRT5, SIRT7 | ||||
| hydroxybutyrylation | lysine | p300 | lysine hydroxybutyrylation antibody | 86 |
| 2-hydroxyisobutyrylation | lysine | p300, MYSTs (ESa1p, Tip60); | lysine 2-hydroxyisobutyrylation antibody | 86 |
| HDAC1-3, Rpd3p, CobB | ||||
| succinylation | lysine | GNATs (GCN5, HAT1), CPT1A, KGDHC; SIRT5, SIRT7 | lysine succinylation antibody | 100 |
| crotonylation | lysine | p300; HDAC-3, SIRT1-3 | lysine crotonylation antibody | 68 |
| lactylation | lysine | p300; HDAC1-3, SIRT1-3 | lysine lactylation antibody | 72 |
Figure 2.

Diagram of experiments to detect posttranslational modifications: preparation of purified protein samples, digestion of proteins into peptides, enrichment of specific PTM peptides using appropriate methods, and HPLC/MS/MS analysis of specific enriched PTM peptides.
4. p300 and Tumours
4.1. p300 Can Regulate Transcription
Transcription is the process of transforming genetic information from DNA to RNA. As the first step of protein biosynthesis, transcription is the synthesis step of mRNA and non-coding RNA (tRNA, rRNA, etc.). p300 can regulate transcription, acting as a coactivator of transcription through two mechanisms. First, p300 regulates transcription factor activity, binds enhancers and promotes gene expression by exerting its own transcriptional functions or by cooperating with other molecules. Secondly, p300 regulates transcription factors by promoting modifications that affect the transcription process. The second mechanism has been the focus of recent studies.
p300 can function as an acetyltransferase to modify certain proteins and then participate in the transcription process. For instance, in gastric cancer and melanoma, p300 affects the proliferation, cell cycle and ageing of tumour cells to some extent by acetylating histones to promote transcription of the corresponding genes [59,60], and a similar finding was found in diffuse large B-cell lymphoma (DLBCL) [61]. p300 also regulates transcriptional processes in prostate cancer; on the one hand, p300 recognise phosphorylated AR, which promotes subsequent AR acetylation at K609, thereby facilitating transcription [62]. On the other hand, p300 can promote the malignant progression of prostate cancer by increasing the levels of H3K18ac, H3K27ac and H4ac in the transcription start site (TSS) and antioxidant response element (ARE) regions, which in turn affects the transcriptional level [63]. In breast cancer, the p300 inhibitor A485 can decrease the transcript levels of highly expressed genes by reducing the H3K27ac level in specific genes, such as ER [64]. It has also been demonstrated that DOT1L interacts with the p300/c-Myc complex to enhance EMT-induced stemness properties by recognising promoters such as those of ZEB1 and ZEB2; this promotes DNA methylation and histone acetylation, enhances epithelial−mesenchymal transition (EMT) regulators, and accelerates the malignant progression of breast cancer [65]. Regarding esophageal adenocarcinoma, p300 acetylates K188 and K189 of Maml1, recruiting NACK into the Notch1 ternary complex, which in turn leads to transcription initiation [66]. The interaction of p300 with MRTF-A was verified in MCF-7 cells, and p300 was found to be recruited to the promoter of MRTF-A target genes to acetylate histones and thereby reconfigure chromatin structure [67]. In addition, p300 can play a role in tumour immune regulation. p300 enhances myeloid-derived suppressor cell (MDSC) immunosuppressive function by regulating C/EBPβ acetylation and Arg-1 transcription [68].
Studies on the effect of p300 inhibitors on transcriptional regulation have found that treatment of multiple myeloma cells with A485 preferentially enables the deacetylation of enhancer progenitor-associated histones at the transcriptional level [69]. Similar findings were observed in human colorectal carcinomas [6]. The p300 inhibitor A485 inhibits acetylation of H3 at the CD274 (encoding PD-L1) promoter site and blocks CD274 transcription, providing a mechanistic basis for the combination of A485 and anti-PD-L1 antibodies in the treatment of prostate cancer [70].Treatment of hepatocellular carcinoma (HCC) with the p300 inhibitor B029-2 inhibits the transcription of six metabolic genes, PSPH, PSAT1, ALDH18A1, ATIC, TALDO1 and DTYMK by blocking the binding of H3K18ac and H3K27Ac to the relevant promoters [71]. In macrophages and induced pluripotent stem cells (iPSCs), p300 has been shown to regulate histone lactylation, and p300 is known to catalyse YTHDF2 promoter lactylation in ocular melanoma cells. HDAC1-3 and SIRT1-3 were shown to be demulsification in other studies [72,73] (Table 2). Figure 3 shows a schematic representation of transcription-related pathways regulated by p300.
Table 2.
Summary of the role of p300-mediated posttranslational modification in tumours biological function.
| Biological Function | Tumour Type | Name of Protein | Modification Type | Effects on Tumours | Regulatory Factors | References |
|---|---|---|---|---|---|---|
| transcriptional control | prostate cancer | AR | Acetylation | Promoting AR acetylation at AR K609 site and promoting transcription process | p300 | [62] |
| transcriptional control | breast cancer | Histone3 | Acetylation | Enhance the level of H3K27ac in ER to enhance the transcription level of highly expressed genes | p300 | [64] |
| transcriptional control | esophageal adenocarcinoma | Maml1 | Acetylation | Promote the acetylation of K188 and K189 of Maml1 and promote the transcription process | p300 | [66] |
| transcriptional control | hepatocellular carcinoma | Histone3 | Acetylation | p300 inhibitor B029-2 can block the binding of H3K18ac and H3K27Ac with related promoters and hinder the transcription process | p300 | [71] |
| transcriptional control | ocular melanoma | Promoter of YTHDF2 | Lactylation | p300 catalyses histone lactate and promotes its transcription | p300 | [73] |
| proliferation, migration and invasion | colorectal cancer | MTA2 | Acetylation | Promotes colorectal cancer cell migration and invasion | p300 | [74] |
| proliferation, migration and invasion | liver cancer | Histone3, PGK1 | Acetylation | Promotes proliferation and enhances metastatic capacity of HCC cells | p300 | [71,75] |
| proliferation, migration and invasion | nasopharyngeal carcinoma | Smad2 and Smad3 | Acetylation | Induces EMT and promotes CNE-2 invasion and migration in human nasopharyngeal carcinoma cells | p300 | [76] |
| proliferation, migration and invasion | prostate cancer | AR | Acetylation | Stabilises AR and promotes the proliferation of prostate cancer cells | p300 | [77,78] |
| proliferation, migration and invasion | cervical cancer | HNRNPA1 | Crotonylation | Promotes HeLa cell proliferation, migration, and invasion | p300 | [79] |
| proliferation, migration and invasion | breast cancer | MRTF-A | Acetylation | Increases the viability and promotes the migration of breast cancer cells | p300 | [67] |
| proliferation, migration and invasion | osteosarcoma | JHDM1A | Acetylation | Acetylation of JHDMIA can inhibit the proliferation and invasion of osteosarcoma HOS cells | p300 | [11] |
| proliferation, migration and invasion | T-cell acute lymphoblastic leukaemia | Notch3 | Acetylation | Promotes proliferation of AML cell lines | p300, HDAC1 | [80,81,82] |
| cell apoptosis | pancreatic cancer | Histone | Acetylation | After being treated with C646, it can inhibit the acetylation of H3K27, and then regulate the apoptosis of pancreatic cancer cells. | p300 | [83] |
| cell apoptosis | gastric cancer | Promoter of RASSF2A | Acetylation | Participate in the acetylation of RASSF2A promoter, and then induce apoptosis of gastric cancer cells | p300 | [59] |
| drug resistance formation | breast cancer | FOXO3 | Acetylation | Lapatinib is a chemotherapy drug for breast cancer. p300 mediates the acetylation of FOXO3 and enhances its sensitivity to Lapa-tinib | p300 | [84] |
| drug resistance formation | glioblastoma | Histone3 | Acetylation | CPI-1612 can inhibit the deposition of H3K27Ac in GBM cells, which provides a basis for the sensitisation of TMZ | p300 | [85] |
| drug resistance formation | prostate Cancer | JMJD1A | Acetylation | The acetylation level of JMJD1A increased in drug-resistant prostate cancer cell lines | p300 | [4] |
| drug resistance formation | colorectal cancer | Histone3/4 | Acetylation | p300 expression profile was found to be related to the resistance of colorectal cancer cells to 5-FU | p300 | [86] |
| cellular metabolism | prostate Cancer | Promoter of FASN | Acetylation | Acetylating H3 in FASN gene promoter to regulate lipid metabolism | p300 | [87] |
| cellular metabolism | liver cancer | SREBP-1c | Acetylation | The stability of SREBP-1c is dynamically controlled by p300 acetylation and SIRT1 deacetylation, which is an important mechanism of lipid regulation in hepatocellular carcinoma cells | p300, SIRT1 | [88] |
| cellular metabolism | carcinoma of colon | ENO1 | 2- hydroxyisobutyration | Catalysed the acylation of ENO1 2- hydroxyisobutyrate, and then enhanced its glycolytic ability and lactic acid excretion | p300 | [52] |
Figure 3.

Schematic representation of transcription-related pathways regulated by p300. (A) The p300 gene recognises phosphorylated AR in prostate cancer and promotes AR acetylation in K609, thereby promoting transcription. (B) A485, an inhibitor of p300, can reduce the level of transcription of highly expressed genes by reducing the level of H3K27ac in particular genes (such as ER). (C) In esophageal adenocarcinoma, p300 acetylates K188 and K189 of Maml1, and recruits NACK to the ternary complex of Notch1, resulting in the initiation of transcription. (D) p300 can catalyse the lactate of YTHDF2 promoter in ocular melanoma cells and then regulate the transcription process. (Remarks: The grey arrow is the related process in which p300 inhibitors participate in regulation, and the dotted line is the process in which modification types are removed).
4.2. p300 Regulates Tumour Cell Proliferation, Migration, and Invasion
p300, a tumour-promoting protein, plays an essential role in malignant progression in most solid tumours, and its role has been confirmed in prostate cancer, colorectal cancer, liver cancer, and other diseases. Many years of research have shown that p300 can be involved in biological processes that regulate tumour cell proliferation, migration and invasion, Tumour cell proliferation, migration, and invasion are three important features of the biological behaviour of malignant tumour development. Of these, tumour cell proliferation is the basis of tumour cell growth, development, inheritance and reproduction, and cells all proliferate by division. Tumour cell migration, also known as tumour cell crawling, is a common phenomenon in the pathological process of tumour metastasis. Cellular invasion is the process of expansion and proliferation of malignant tumour cells from their origin to surrounding normal tissue along the interstitial space. Indicating that tumour cells breach the basement membrane. and a clear understanding of the p300 mechanism of action provides a solid basis for further clarification of the role of p300. Furthermore, the focus of research on p300 over the years has shifted, and most studies now focus on the role of p300 as an acetylase.
p300 possesses acetyltransferase activity and plays a procancer role in both solid and haematological cancers. In many solid tumours, such as breast cancer [89], hepatocellular carcinoma [90], oesophageal cancer [91], and cutaneous squamous cell carcinomas [92], increased p300 expression is associated with an aggressive phenotype and a low survival rate; furthermore, histochemistry analysis of a variety of primary tumour tissues has revealed that lower overall levels of H3K18ac are associated with lower survival and a higher risk of recurrence in prostate cancer, renal cancer, lung cancer, pancreatic cancer and breast cancer [93,94,95,96,97]. Wang et al. showed that p300 can acetylate PHF5A, promote cell proliferation, and affect the prognosis of colorectal cancer [98]. In studies of colorectal cancer development, p300 was found to act not only through overexpression of the Wnt signalling pathway but also through interaction with and acetylation of MTA2 to promote migration and invasion [74,99]. HAN et al. suggested that cervical cancer is highly malignant in part because p300 promotes the proliferation, migration, and invasion of HeLa cells by mediating lysine crotonylation and increasing HNRNPA1 expression [79]. p300 can promote the invasion and migration of human nasopharyngeal carcinoma cells (CNE-2 cells), possibly by acetylating Smad2 and Smad3 through the TGF-β signalling pathway and inducing EMT [76]. AR is acetylated, and its stability is regulated by p300, which promotes prostate cancer cell proliferation [77]. In MCF-7 cell experiments, overexpression of p300 and MRTF-A was found to catalyse MRTF-A acetylation and to increase the transcript levels of migration-related genes such as MYL9, MYH9 and CYR61, which in turn increased breast cancer cell viability and promoted cell migration [67]. However, p300 can exert a tumour suppressor effect; for example, in regulating osteosarcoma development, p300 can acetylate JHDMIA K409 and can inhibit the proliferation and invasion of HOS osteosarcoma cells [11].
Regarding haematological tumours, the p300 inhibitor C646 was found to cause apoptosis and slow the proliferation of AML1-ETO-positive acute myeloid leukaemia (AML) cells. Treatment with C464 delayed the growth and induced apoptosis of AML1-ETO-positive AML cell lines and primary parental cells, and the study found that acetylation of AML1-ETO by p300 may be responsible [100,101]. In T-cell acute lymphoblastic leukaemia, p300 and HDAC1 were found to acetylate and deacetylate the K1692 and K1731 positions of Notch3, which plays a key role in T-cell proliferation and the overall malignant progression of T-cell leukaemia [81]. In human leukaemia cell lines, it was also demonstrated that the CREBBP/EP300 bromodomain is essential for regulating the GATA1/MYC regulatory axis during proliferation, and administration of CREBBP/EP300 bromodomain inhibitors was very effective in reducing H3K27ac levels and thus hindering cancer cell proliferation [102].
In successive studies exploring the therapeutic feasibility of p300 inhibitors, it was found that A485, a p300-specific inhibitor, significantly inhibited the growth of ER+ breast cancer cells, which may be related to the down-regulation of enhancer H3K27ac, which inhibits the expression of ER target gene [64]. The antitumour activity of A485 in growth hormone pituitary adenoma was also associated with reduced levels of H3K18ac and H3K27ac [103]. Treatment of colon cancer cells with the isothiazolone-based agent PCAF and the p300 histone acetyltransferase (HAT) co-inhibitors CCT077791 and CCT077792 inhibited histone acetylation and thus prevented colon cancer cell proliferation [104,105]. Treatment of hepatocellular carcinoma cells with the highly potent p300 inhibitor B029-2 significantly reduced the levels of H3K18ac and H3K27ac and significantly reduced the proliferation and metastatic capacity of hepatocellular carcinoma cells [71]. The reason why the combination of curcumin and anti-PD-1 therapy is more effective than anti-PD-1 therapy alone is that curcumin can reduce p300-induced histone acetylation in the promoter region of TGF-β1 because it inhibits p300 expression, thereby activating immune cell function and reducing immune escape [106]. Triple-negative breast cancer cells are sensitive to the p300 inhibitor L002, and treatment with L002 can significantly inhibit the growth of cancer cells. In animal experiments, it was also confirmed that L002 could significantly inhibit tumour growth in vivo, and histochemistry experiments showed that the level of H4ac was significantly decreased in the group that received L002. It was further speculated that L002 might function by decreasing the level of histone acetylation [107]. In summary, p300 is a potential target for the treatment of tumours because it can regulate biological processes such as tumour cell growth, migration and invasion (Table 2), and these processes are presented in Table 2.
4.3. p300 Regulates Tumour Cell Apoptosis
Tumour cell apoptosis is an ordered or procedural manner of cell death, and it is an active death process of tumour cells under the control of specific genes. Apoptotic cells will ultimately be processed by phagocytes. In the case of cancer cells, the interruption of the apoptosis process signifies the development and spread of cancer. Apoptosis significantly affects the fate of tumour cells, and in general, p300 directly and indirectly regulates tumour cell apoptosis and affects tumour progression. p300 affects the following regulatory signalling molecules during the complex pathogenesis of the development of different tumours: c-MYC [108], c-Met, cyclin D1, Bcl2 [2], TRAIL [109], RAR and ATRA [28], Wnt/β-catenin [74], API5 [110], p53 [111], etc. p300 acts as an acetyltransferase, catalysing the acetylation of histones and nonhistone proteins, and plays an integral role in regulating tumour cell apoptosis. Ono et al. performed pancreatic cancer cell-related experiments and found that the expression of apoptosis-related proteins such as cleaved caspases 3, 8, and 9 and PARP was increased after p300 interference, and acetylation of H3K27 was also inhibited by C646 treatment, suggesting that p300 may regulate apoptosis of pancreatic cancer cells via its HAT activity [83]. Liu et al. found that p300 is involved in the acetylation of H3 and H4 on the RASSF2A promoter and regulates RASSF2A expression, which in turn induces the apoptosis of gastric cancer cells [59]. In studies by Fu and others, C646 treatment led to the apoptosis of AML1-ETO-positive AML cell lines and primary parental cells, while normal haematopoietic stem cells were not affected [100,112]. Because p300 can mediate histone acetylation, it is speculated that targeting p300 to regulate histone acetylation may be an important new therapeutic option for AML treatment [101]. Interestingly, p300 can also catalyse the modification of some nonhistone proteins; for example, in the presence of hypoxia or DNA damage, p300 can acetylate p53, which in turn regulates apoptosis [113,114] (Table 2). Figure 4 shows a schematic representation of the pathways involved in the regulation of apoptosis by p300.
Figure 4.

Schematic representation of pathways involved in the regulation of apoptosis by p300. (A) In pancreatic cancer studies, C646 treatment was followed by interference with p300 followed by inhibition of acetylation of H3K27, leading to increased expression of apoptosis-related proteins such as cleaved caspase 3, 8 and 9 and promoting cell apoptosis. (B) C646 treatment leads to apoptosis in AML1-ETO positive AML cell line and primary parental cells, which may be associated with p300-mediated histone acetylation. (C) p300 is involved in the acetylation of the Histones H3 and H4 on RASSF2A promoter and regulates the expression of RASSF2A, thus inducing apoptosis of gastric cancer cells. (D) p300 acetylates p53 and regulates apoptosis in the presence of hypoxia or DNA damage (Remarks: The grey arrow is the related process in which p300 inhibitors participate in regulation, and the dotted line is the process in which modification types are removed).
4.4. p300 Regulates the Formation of Tumour Drug Resistance
Drug resistance, leading to a decrease or lack of efficacy of the drug towards the pathogen, generally refers to the decline or even disappearance of the pathogen’s susceptibility to the drug following repeated drug contact. The emergence of drug resistance is a significant reason for the ineffectiveness of tumour treatment, and it is a problem that needs to be understood and solved. As a regulator closely related to the degree of tumour malignancy, p300 is also involved in the development of drug resistance. Lapatinib is a chemotherapeutic drug for breast cancer, and p300 mediates FOXO3 acetylation and enhances sensitivity to lapatinib [84]. It has also been demonstrated in relevant experiments in cisplatin-resistant bladder cancer cells that cisplatin resistance may be acquired by p300 catalysing foxo3a acetylation[115]. Ono et al. found that HAT inhibition by C646 increased the cytotoxic effect of gemcitabine on pancreatic cancer cell lines at 96 h. The researchers also confirmed that acetylation of H3K27 was inhibited and speculated that the development of gemcitabine resistance in pancreatic cancer was prevented, at least in part, by a HAT-dependent mechanism [83]. In addition, many other related studies have found that p300 inhibition enhances the sensitivity of drug-resistant tumour cells to chemotherapeutic agents by modulating HAT activity. For example, Mladek et al. found that CPI-1612 can effectively block RBBP4/p300 HAT activity by inhibiting the deposition of H3K27Ac in GBM cells, providing a new idea for the sensitisation of glioma to temozolomide (TMZ) [85]. In the study by Huang et al., p300 interference decreased the H3K27ac level, while a strain resistant to EZH2 inhibitors became sensitive. Subsequently, MLL1, which inhibits the binding of p300 to CBP and reduces the level of H3K27ac, was also assessed, providing research directions that could facilitate a more comprehensive understanding of p300 function and the development of tumour therapy [116].
In a study by Xu et al., p300 was found to promote acetylation of JMJD1A K421. The level of JMJD1A acetylation was increased in drug-resistant prostate cancer cell lines, and subsequent experiments also demonstrated that p300 inhibitors significantly inhibited the proliferation of resistant prostate cancer cell lines and increased the sensitivity of these tumour cells to AR antagonists [4]. Additionally, the chemotherapeutic drug 5-FU was found to reduce the ability of p300 to bind to chromatin; 5-FU induced H3 and H4 deacetylation and thus their degradation via lysosomes in a study by Du et al. The p300 expression profile was found to correlate with the resistance of colorectal cancer cells to 5-FU [86]. In response to DNA damage, p300 promotes DNA replication and repair through acetylation of H3 and/or H4 and induces chemotherapy and radiation therapy resistance in a variety of tumour types [117] (Table 2). Figure 5 shows a schematic of the modulation of resistance by p300.
Figure 5.

Schematic of the modulation of resistance by p300. (A) p300 promoted the acetylation of JMJD1A k421, and the acetylation level of JMJD1A increased in drug-resistant prostate cancer cell lines. (B) p300 mediates the acetylation of FOXO3 and enhances the sensitivity to lapatinib in the treatment of breast cancer. (C) C646 inhibits the acetylation of H3K27 to a degree that inhibits gemcitabine resistance in the treatment of pancreatic cancer. (D) After treatment with p300 inhibitor CPI-1612, the activity of RBBP4/p300 HAT was effectively blocked, which made the treatment of glioblastoma sensitised to temozolomide (TMZ). (Remarks: The grey arrow is the related process in which p300 inhibitors participate in regulation, and the dotted line is the process in which modification types are removed).
4.5. p300 Regulates Tumour Metabolic Processes
p300 is a typical acetyltransferase and transcriptional coactivator that not only regulates transcription, apoptosis, and protein localisation, proliferation, migration, and invasion but also participates in metabolic processes. As we all know, tumour cell metabolism, means that tumour cells will acquire unique metabolic preferences depending on their tissue of origin, the degree of genetic changes, and the degree of interaction with hormones and systemic metabolites. That is, tumour occurrence is dependent on the reprogramming of cellular metabolism. p300 is involved in the regulation of tumour cell metabolism primarily as follows, for example, p300 can regulate glucose metabolism and fat metabolism. According to early studies on p300, it has a unique role in the control of metabolic processes as an acetyltransferase.
In studies related to prostate cancer, p300 was found to have a tumour-promoting effect and to regulate the expression of FASN, a regulator of lipid metabolism, by acetylating H3 in the FASN gene promoter. Therefore, p300 may be involved in the regulation of lipid metabolism [87]. Early studies of cellular lipid metabolism revealed that the stability of SREBP-1c is dynamically controlled by p300 acetylation and SIRT1 deacetylation, the latter being a mechanism specific to HepG2 cells [88]. A485, which regulates FOXO1 deacetylation and degradation via a proteasome-dependent pathway, has been shown to inhibit hepatic lipogenesis and glycoprotein production [118]. p300 and HDAC1 acetylate and deacetylate the catalytic subunit of adenosine monophosphate-activated protein kinase (AMPK); these proteins were found to enhance the interactions of the upstream kinase LKB1 when AMPK is deacetylated and lead to phosphorylation and activation of AMPK, resulting in lipolysis in human hepatocytes [107].
p300 can increase the H3K18Ac and H3K27Ac levels in the promoter regions of the metabolism-related genes PSAT1, ATIC and TALDO1 in Huh-7 cells, which in turn promotes glycolysis [71]. Similar studies have also demonstrated that B029-2, which regulates the acetylation of H3 K18 and K27, reduces the glycolytic capacity of hepatocellular carcinoma cells; thus, it has been explored as a potential p300-targeting drug for cancer treatment [71]. In addition to acetylation of nonhistone proteins, p300 catalyses 2-hydroxyisobutylation and lactylation of nonhistone proteins. Recent studies have also shown that p300 regulates glycolysis and lactate excretion by mediating the 2-hydroxyisobutyl conversion of ENO1 to induce acetylase activity, thereby regulating colon cancer metabolism [52]. Treatment with C646 also inhibits the metabolism of HepG2 (or Huh7) hepatocellular carcinoma cells via this mechanism [119]. p300 is also involved in the metabolic processes of immune cells. p300 helps macrophages take up extracellular lactate via monocarboxylic acid transporters (MCTs) and catalyses lactylation of HMGB1 in macrophages. C646 inhibits p300 acetylase activity and inhibits lactate-induced lactylation of HMGB1, two regulatory mechanisms by which p300 participates in macrophage metabolism [120]. In addition, deletion of the p300 gene to detect its effect on metabolic processes in serum induced changes in metabolites such as glutamate/glutamine and choline glycerate, and this finding was verified in HCT116 cells [121] (Table 2). Figure 6 shows a schematic diagram of the mechanisms of p300 in tumour metabolic regulation.
Figure 6.

Schematic diagram of the mechanisms of p300 in tumour metabolism regulation. p300 catalyses many different types of posttranslational modifications, such as acetylation, β-hydroxyisobutyrylation and lactylation, in tumour cells to modulate glycometabolism and lipid metabolism. (Remarks: The grey arrow is the related process in which p300 inhibitors participate in regulation, and the dotted line is the process in which modification types are removed).
5. p300 Inhibitors
In many malignancies, p300 is considered a tumour promoter, a prospective target for tumour therapy, and a biomarker of tumour prognosis; as such, the development and application of p300 inhibitors is a priority.
Several p300-related inhibitors have been discovered over many years of scientific research and clinical exploration. For example, C646 not only controls the c-Met/Akt pathway and cyclin D1 expression but also induces the apoptosis of gastric cancer cells [2]. One study also found that C646 promotes the degradation of JMJD1A, which enhances the sensitivity of enzalutamide-resistant cells to C646, thus improving prostate cancer prognosis [4]. In addition, C646 enhances antitumour activity in colorectal cancer by blocking TRIB3 acetylation and promoting TRIB3 degradation [122]. C646 can also be used in the treatment of haematological tumours, and studies suggest that C646 may be a promising drug for widespread use in the treatment of AML [123,124]. Another inhibitor, CCS1477, inhibits prostate cancer cell proliferation and reduces the expression of AR and c-MYC regulatory genes, making it a promising novel prostate cancer treatment [78]. A high-throughput screen identified the histone acetylation compound L002 as a highly effective inhibitor of p300 in lymphoma cells and triple-negative breast cancer, making it a promising cancer treatment [107]. Both PU139 and PU141 are p300 inhibitors that significantly inhibit neuroblastoma growth and are effective against neuroblastoma [125]. Virtual ligand screening identified the p300 inhibitors anacardic acid [126], curcumin [127,128,129], chrysanthemum and isothiazole [130]. LTK-13 and LTK-14 are selective p300 inhibitors that inhibit the acetylation of p53 [131]. Other p300 inhibitors, such as RTK1 [132], LTK19 [131], and garcinol [131], have been discovered and employed in cancer therapy.
A485 is the most well-known selective p300 inhibitor, and its mechanism of action has been relatively clear for many years. It regulates tumourigenesis and progression by regulating transcription levels, thus affecting biological behaviour and immune function. p300 was also found to play a role in a variety of tumours and cell lines, such as prostate cancer, melanoma and various haematological tumours (multiple myeloma cell lines, AML cell lines and non-Hodgkin lymphoma cell lines) [70,133]. IIn other studies, four alkaloid p300 inhibitors were assessed, and their antitumour effects were validated in subsequent clinical studies [134]. In addition, some p300-related inhibitors, such as cyclopentenone prostaglandin, Y08197, CBP30, and B029-2, have been identified [71,135,136,137]. So far, two p300 inhibitors have been identified in clinical trials, including CCS1477 and FT-7051. Among them, CCS1477 has used in a phase I/II clinical trial in patients with metastatic castration prostate cancer. The other is FT-7051, which has started a phase one clinical trial in patients with metastatic castration resistant prostate cancer.
With the development of experiments related to p300 inhibitors, it is found that the curative effect of p300 inhibitors in the treatment of tumours is not a very ideal one. This is largely due to the fact that limited curative effect of p300 inhibitor monotherapy, thus, the majority of workers in scientific research try their best to explore a better treatment regime. Of these, combination therapy with inhibitors of p300 has an excellent therapeutic outlook. What is most notable is that some studies have begun to explore the therapeutic effect of the p300 inhibitor in combination with the PD-L1 antibody/anti-PD-L1 antibody. For example, in the treatment of colorectal cancer, the antitumour effect of C646 combined with PD-L1 antibody is obviously enhanced [122]. A485 combined with anti-PD-L1 antibody significantly inhibited androgen independent metastatic tumour growth in the setting of prostate cancer [70]. Based on these experiments, we can find that the p300 inhibitor in combination with the PD-L1 antibody/anti-PD-L1 antibody has a better therapeutic outlook in the treatment of various tumours. On the side, it can also be seen that the relationship between p300 and PDL1 and its related mechanism is worthy of investigation. Indeed, in some studies, there are also some explorations on its mechanism. For example, in Liu et al.’s study, p300 inhibitors can inhibit p300 from blocking the expression of PD-L1 induced by intrinsic and IFN-γ, and significantly improve the therapeutic effect of PD-L1 blocking on PCa.
p300 has been shown to play a key role in IFN-γ induced PD-L1 expression, and could be a good target for overcoming adaptive resistance induced by the PD-1/PD-L1 pathway [70]. In Gao et al.’ s research, it was confirmed that the acetylation and deacetylation of k263 site of PD-L1 were dynamically regulated by p300 and HDAC2, and modification of acetylation may promote PD-L1 translocation into the nucleus, and are involved in the regulation of IFN, NF-kB, MHC I, and other immune response gene expression by binding to DNA, thereby promoting tumours immune evasion [138]. there is not so much research on the relationship between p300 and PDL1 and its biological mechanism is relatively scarce, therefore, this will also be a research idea worthy of future study. We summarise some inhibitors of p300 and their main functions in Table 3.
Table 3.
Summary of representative p300 inhibitors and their main functions.
| Name | Structure | Main Function | Clinical Trial Landscape | References |
|---|---|---|---|---|
| C646 |
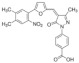
|
C646 regulates tumour cell drug resistance in cancers such as prostate cancer. C646 can enhance the antitumour activity in colorectal cancer by blocking the acetylation of TRIB3 and promoting the degradation of TRIB3. C646 can block the tumour cell cycle, induce tumour cell apoptosis, and inhibit tumour cell proliferation and growth in lung cancer and melanoma | N/A | [108,122,130] |
| A485 |
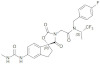
|
A485 regulates antitumour activity in a variety of solid tumours such as melanoma and prostate cancer and in haematological tumours (such as multiple myeloma, acute myeloid leukaemia, and non-Hodgkin lymphoma). Combination of an anti-PD-L1 antibody with A485 can further enhance its antitumour efficacy | N/A | [70,124,133,139] |
| CCS1477 |
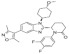
|
CCS1477 can exert anticancer effects in prostate cancer by decreasing AR expression and regulating C-MYC gene expression | Phase I/IIa clinical trial for patients with advanced solid tumours | [78] |
| B029-2 |
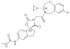
|
B029-2 can inhibit glycolysis, induce tumour cell cycle arrest, and inhibit tumour cell proliferation, thus delaying the malignant progression of liver cancer | N/A | [71] |
| B026 |
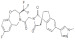
|
B026 can significantly inhibit the expression of MYC in leukaemia cell lines and lymphoma cell lines and significantly decrease the level of H3K27Ac, and it has better effects and stability than A485. In addition, B026 can inhibit the proliferation of prostate cancer Cells | N/A | [140] |
| CCT077791 |
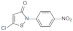
|
CCT077791 treatment for 24 h inhibited colon cancer cell proliferation by reducing the H3 and H4 acetylation levels in HCT116 and HT29 cells | N/A | [105] |
| CBP30 |
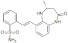
|
CBP30 can impair Treg differentiation and inhibit their function, which in turn enhances antitumor immune function and exerts antitumour effects. CBP30 also inhibits human Th17 responses | N/A | [137,141] |
6. Conclusions and Future Perspectives
The currently known functions of p300 are very broad; it participates in both cancer and nontumour diseases. As detailed in the above introduction to the roles of p300 in cancer, p300 plays a major role in posttranslational modification, which affects not only transcriptional regulation but also tumour cell proliferation, migration, invasion, and apoptosis, as well as the development of drug resistance and metabolic regulatory processes.
A large number of drugs targeting p300 have been studied in cell and animal experiments, but only two p300 inhibitors, such as CCS1477 and FT-7051, have been verified in clinical trials, which also reveals that it is extremely urgent for us to explore the clinical application value of more p300 inhibitors. In addition, in the experiment of p300 inhibitor, there are relatively few related studies to elaborate the treatment scheme of p300 inhibitor to maximise its efficacy, but novel explorations of p300 inhibitor treatment regimens remain. For example, the p300 inhibitor in combination with an anti-PD-L1 antibody is applied to experimental animals with tumours, greatly enhancing the anticancer efficacy of p300 inhibitor monotherapies.
Therefore, in future research, when discussing the mechanism and function of p300, researchers may focus on the following two aspects: on the one hand, at the level of posttranslational modification, we will continue to explore the relevant mechanism of catalytic action of p300 acetyltransferase; and on the other hand, we will focus on the existing posttranslational modification of p300 as a brand new target in order to develop more inhibitors of p300 and to further investigate a more scientific and efficient combination therapy scheme of p300 inhibitors and other anticancer drugs for clinical transformation.
Author Contributions
Z.C. and W.H. conceived the review framework for this article, W.H. and Q.Z. designed the article content and wrote the manuscript, Z.C. provided article polishing support, W.H., Q.Z., K.W., Y.Z. and Q.M. provided suggestions and content modification and polishing. All authors have read and agreed to the published version of the manuscript.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Funding Statement
We acknowledge the support from National Natural Science Foundation of China 82173017, Natural Science Basis Research Plan in Shaanxi Province of China (Program No. 2021JQ-337) and the Scientific and Technological Innovation Major Base of Guangxi (No.2022-36-Z05), Guangxi Key Research and Development Project (No. AB20117001), Guangxi science and technology bases and talent special project (No. AD17129062).
Footnotes
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
References
- 1.Gao Y., Geng J., Hong X., Qi J., Teng Y., Yang Y., Qu D., Chen G. Expression of p300 and CBP is associated with poor prognosis in small cell lung cancer. Int. J. Clin. Exp. Pathol. 2014;7:760–767. [PMC free article] [PubMed] [Google Scholar]
- 2.Wang Y.M., Gu M.L., Meng F.S., Jiao W.R., Zhou X.X., Yao H.P., Ji F. Histone acetyltransferase p300/CBP inhibitor C646 blocks the survival and invasion pathways of gastric cancer cell lines. Int. J. Oncol. 2017;51:1860–1868. doi: 10.3892/ijo.2017.4176. [DOI] [PubMed] [Google Scholar]
- 3.Li M., Song S.W., Ge Y., Jin J.Y., Li X.Y., Tan X.D. The Ras-ERK signaling pathway regulates acetylated activating transcription factor 2 via p300 in pancreatic cancer cells. Ann. Transl. Med. 2020;8:1234. doi: 10.21037/atm-20-5880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Xu S., Fan L., Jeon H.Y., Zhang F., Cui X., Mickle M.B., Peng G., Hussain A., Fazli L., Gleave M.E., et al. p300-Mediated Acetylation of Histone Demethylase JMJD1A Prevents Its Degradation by Ubiquitin Ligase STUB1 and Enhances Its Activity in Prostate Cancer. Cancer Res. 2020;80:3074–3087. doi: 10.1158/0008-5472.CAN-20-0233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bi Y., Kong P., Zhang L., Cui H., Xu X., Chang F., Yan T., Li J., Cheng C., Song B., et al. EP300 as an oncogene correlates with poor prognosis in esophageal squamous carcinoma. J. Cancer. 2019;10:5413–5426. doi: 10.7150/jca.34261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ishihama K., Yamakawa M., Semba S., Takeda H., Kawata S., Kimura S., Kimura W. Expression of HDAC1 and CBP/p300 in human colorectal carcinomas. J. Clin. Pathol. 2007;60:1205–1210. doi: 10.1136/jcp.2005.029165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hou X., Li Y., Luo R.Z., Fu J.H., He J.H., Zhang L.J., Yang H.X. High expression of the transcriptional co-activator p300 predicts poor survival in resectable non-small cell lung cancers. Eur. J. Surg. Oncol. 2012;38:523–530. doi: 10.1016/j.ejso.2012.02.180. [DOI] [PubMed] [Google Scholar]
- 8.Cheng G., Liu F., Asai T., Lai F., Man N., Xu H., Chen S., Greenblatt S., Hamard P.J., Ando K., et al. Loss of p300 accelerates MDS-associated leukemogenesis. Leukemia. 2017;31:1382–1390. doi: 10.1038/leu.2016.347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Xie X., Piao L., Bullock B.N., Smith A., Su T., Zhang M., Teknos T.N., Arora P.S., Pan Q. Targeting HPV16 E6-p300 interaction reactivates p53 and inhibits the tumorigenicity of HPV-positive head and neck squamous cell carcinoma. Oncogene. 2014;33:1037–1046. doi: 10.1038/onc.2013.25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ichise T., Yoshida N., Ichise H. CBP/p300 antagonises EGFR-Ras-Erk signalling and suppresses increased Ras-Erk signalling-induced tumour formation in mice. J. Pathol. 2019;249:39–51. doi: 10.1002/path.5279. [DOI] [PubMed] [Google Scholar]
- 11.Wang Y., Sun B., Zhang Q., Dong H., Zhang J. p300 Acetylates JHDM1A to inhibit osteosarcoma carcinogenesis. Artif. Cells Nanomed. Biotechnol. 2019;47:2891–2899. doi: 10.1080/21691401.2019.1638790. [DOI] [PubMed] [Google Scholar]
- 12.Zhu G., Pei L., Li Y., Gou X. EP300 mutation is associated with tumor mutation burden and promotes antitumor immunity in bladder cancer patients. Aging. 2020;12:2132–2141. doi: 10.18632/aging.102728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ferrari R., Gou D., Jawdekar G., Johnson S.A., Nava M., Su T., Yousef A.F., Zemke N.R., Pellegrini M., Kurdistani S.K., et al. Adenovirus small E1A employs the lysine acetylases p300/CBP and tumor suppressor Rb to repress select host genes and promote productive virus infection. Cell Host Microbe. 2014;16:663–676. doi: 10.1016/j.chom.2014.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Zhu X.Y., Huang C.S., Li Q., Chang R.m., Song Z.b., Zou W.y., Guo Q.L. P300 exerts an epigenetic role in chronic neuropathic pain through its acetyltransferase activity in rats following chronic constriction injury (CCI) Mol. Pain. 2012;8:1. doi: 10.1186/1744-8069-8-84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rai R., Sun T., Ramirez V., Lux E., Eren M., Vaughan D.E., Ghosh A.K. Acetyltransferase p300 inhibitor reverses hypertension-induced cardiac fibrosis. J. Cell Mol. Med. 2019;23:3026–3031. doi: 10.1111/jcmm.14162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Ghosh A.K. p300 in Cardiac Development and Accelerated Cardiac Aging. Aging Dis. 2020;11:916–926. doi: 10.14336/AD.2020.0401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Goodman R.H., Smolik S. CBP/p300 in cell growth, transformation, and development. Genes. Dev. 2000;14:1553–1577. doi: 10.1101/gad.14.13.1553. [DOI] [PubMed] [Google Scholar]
- 18.Grossman S.R. p300/CBP/p53 interaction and regulation of the p53 response. Eur. J. Biochem. 2001;268:2773–2778. doi: 10.1046/j.1432-1327.2001.02226.x. [DOI] [PubMed] [Google Scholar]
- 19.Lakshmanan M.D., Shaheer K. Endocrine disrupting chemicals may deregulate DNA repair through estrogen receptor mediated seizing of CBP/p300 acetylase. J. Endocrinol. Investig. 2020;43:1189–1196. doi: 10.1007/s40618-020-01241-5. [DOI] [PubMed] [Google Scholar]
- 20.Ogiwara H., Kohno T. CBP and p300 histone acetyltransferases contribute to homologous recombination by transcriptionally activating the BRCA1 and RAD51 genes. PLoS ONE. 2012;7:e52810. doi: 10.1371/journal.pone.0052810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Raisner R., Kharbanda S., Jin L., Jeng E., Chan E., Merchant M., Haverty P.M., Bainer R., Cheung T., Arnott D., et al. Enhancer Activity Requires CBP/P300 Bromodomain-Dependent Histone H3K27 Acetylation. Cell Rep. 2018;24:1722–1729. doi: 10.1016/j.celrep.2018.07.041. [DOI] [PubMed] [Google Scholar]
- 22.Jayatunga M.K., Thompson S., McKee T.C., Chan M.C., Reece K.M., Hardy A.P., Sekirnik R., Seden P.T., Cook K.M., McMahon J.B., et al. Inhibition of the HIF1alpha-p300 interaction by quinone- and indandione-mediated ejection of structural Zn(II) Eur. J. Med. Chem. 2015;94:509–516. doi: 10.1016/j.ejmech.2014.06.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sun Y., Kolligs F.T., Hottiger M.O., Mosavin R., Fearon E.R., Nabel G.J. Regulation of β-catenin transformation by the p300 transcriptional coactivator. Proc. Natl. Acad. Sci. USA. 2000;97:12613–12618. doi: 10.1073/pnas.220158597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Dyson H.J., Wright P.E. Role of Intrinsic Protein Disorder in the Function and Interactions of the Transcriptional Coactivators CREB-binding Protein (CBP) and p300. J. Biol. Chem. 2016;291:6714–6722. doi: 10.1074/jbc.R115.692020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Li G.D., Fang J.X., Chen H.Z., Luo J., Zheng Z.H., Shen Y.M., Wu Q. Negative regulation of transcription coactivator p300 by orphan receptor TR3. Nucleic. Acids Res. 2007;35:7348–7359. doi: 10.1093/nar/gkm870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Joy S.T., Henley M.J., De Salle S.N., Beyersdorf M.S., Vock I.W., Huldin A.J.L., Mapp A.K. A Dual-Site Inhibitor of CBP/p300 KIX is a Selective and Effective Modulator of Myb. J. Am. Chem. Soc. 2021;143:15056–15062. doi: 10.1021/jacs.1c04432. [DOI] [PubMed] [Google Scholar]
- 27.Nishihara A., Hanai J.I., Okamoto N., Yanagisawa J., Kato S., Miyazono K., Kawabata M. Role of p300, a transcriptional coactivator, in signalling of TGF-β. Genes Cells. 1998;3:613–623. doi: 10.1046/j.1365-2443.1998.00217.x. [DOI] [PubMed] [Google Scholar]
- 28.Dietze E.C., Caldwell L.E., Marcom K., Collins S.J., Yee L., Swisshelm K., Hobbs K.B., Bean G.R., Seewaldt V.L. Retinoids and retinoic acid receptors regulate growth arrest and apoptosis in human mammary epithelial cells and modulate expression of CBP/p300. Microsc. Res. Tech. 2002;59:23–40. doi: 10.1002/jemt.10174. [DOI] [PubMed] [Google Scholar]
- 29.Wang F., Marshall C.B., Ikura M. Transcriptional/epigenetic regulator CBP/p300 in tumorigenesis: Structural and functional versatility in target recognition. Cell. Mol. Life Sci. 2013;70:3989–4008. doi: 10.1007/s00018-012-1254-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Chan H.M., La Thangue N.B. p300/CBP proteins: HATs for transcriptional bridges and scaffolds. J. Cell Sci. 2001;114:2363–2373. doi: 10.1242/jcs.114.13.2363. [DOI] [PubMed] [Google Scholar]
- 31.Delvecchio M., Gaucher J., Aguilar-Gurrieri C., Ortega E., Panne D. Structure of the p300 catalytic core and implications for chromatin targeting and HAT regulation. Nat. Struct Mol. Biol. 2013;20:1040–1046. doi: 10.1038/nsmb.2642. [DOI] [PubMed] [Google Scholar]
- 32.Park S., Stanfield R.L., Martinez-Yamout M.A., Dyson H.J., Wilson I.A., Wright P.E. Role of the CBP catalytic core in intramolecular SUMOylation and control of histone H3 acetylation. Proc. Natl. Acad. Sci. USA. 2017;114:E5335–E5342. doi: 10.1073/pnas.1703105114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Liu X., Wang L., Zhao K., Thompson P.R., Hwang Y., Marmorstein R., Cole P.A. The structural basis of protein acetylation by the p300/CBP transcriptional coactivator. Nature. 2008;451:846–850. doi: 10.1038/nature06546. [DOI] [PubMed] [Google Scholar]
- 34.Walsh C.T., Garneau-Tsodikova S., Gatto G.J., Jr. Protein posttranslational modifications: The chemistry of proteome diversifications. Angew. Chem. Int. Ed. Engl. 2005;44:7342–7372. doi: 10.1002/anie.200501023. [DOI] [PubMed] [Google Scholar]
- 35.Perdivara I., Deterding L.J., Przybylski M., Tomer K.B. Mass spectrometric identification of oxidative modifications of tryptophan residues in proteins: Chemical artifact or post-translational modification? J. Am. Soc. Mass Spectrom. 2010;21:1114–1117. doi: 10.1016/j.jasms.2010.02.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Humphrey S.J., James D.E., Mann M. Protein Phosphorylation: A Major Switch Mechanism for Metabolic Regulation. Trends Endocrinol. Metab. 2015;26:676–687. doi: 10.1016/j.tem.2015.09.013. [DOI] [PubMed] [Google Scholar]
- 37.Kaiser P., Wohlschlegel J. Identification of ubiquitination sites and determination of ubiquitin-chain architectures by mass spectrometry. Methods Enzym. 2005;399:266–277. doi: 10.1016/S0076-6879(05)99018-6. [DOI] [PubMed] [Google Scholar]
- 38.Hashimoto H., Vertino P.M., Cheng X. Molecular coupling of DNA methylation and histone methylation. Epigenomics. 2010;2:657–669. doi: 10.2217/epi.10.44. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Jiang T., Zhou X., Taghizadeh K., Dong M., Dedon P.C. N-formylation of lysine in histone proteins as a secondary modification arising from oxidative DNA damage. Proc. Natl. Acad. Sci. USA. 2007;104:60–65. doi: 10.1073/pnas.0606775103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Turner B.M. Histone acetylation and an epigenetic code. BioEssays. 2000;22:836–845. doi: 10.1002/1521-1878(200009)22:9<836::AID-BIES9>3.0.CO;2-X. [DOI] [PubMed] [Google Scholar]
- 41.Liu B., Lin Y., Darwanto A., Song X., Xu G., Zhang K. Identification and characterization of propionylation at histone H3 lysine 23 in mammalian cells. J. Biol. Chem. 2009;284:32288–32295. doi: 10.1074/jbc.M109.045856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Peng C., Lu Z., Xie Z., Cheng Z., Chen Y., Tan M., Luo H., Zhang Y., He W., Yang K., et al. The first identification of lysine malonylation substrates and its regulatory enzyme. Mol. Cell Proteom. 2011;10:M111.012658. doi: 10.1074/mcp.M111.012658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chen Y., Sprung R., Tang Y., Ball H., Sangras B., Kim S.C., Falck J.R., Peng J., Gu W., Zhao Y. Lysine propionylation and butyrylation are novel post-translational modifications in histones. Mol. Cell Proteom. 2007;6:812–819. doi: 10.1074/mcp.M700021-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tan M., Peng C., Anderson K.A., Chhoy P., Xie Z., Dai L., Park J., Chen Y., Huang H., Zhang Y., et al. Lysine glutarylation is a protein posttranslational modification regulated by SIRT5. Cell Metab. 2014;19:605–617. doi: 10.1016/j.cmet.2014.03.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zhang Z., Tan M., Xie Z., Dai L., Chen Y., Zhao Y. Identification of lysine succinylation as a new post-translational modification. Nat. Chem. Biol. 2011;7:58–63. doi: 10.1038/nchembio.495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tan M., Luo H., Lee S., Jin F., Yang J.S., Montellier E., Buchou T., Cheng Z., Rousseaux S., Rajagopal N., et al. Identification of 67 histone marks and histone lysine crotonylation as a new type of histone modification. Cell. 2011;146:1016–1028. doi: 10.1016/j.cell.2011.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Zhang D., Tang Z., Huang H., Zhou G., Cui C., Weng Y., Liu W., Kim S., Lee S., Perez-Neut M., et al. Metabolic regulation of gene expression by histone lactylation. Nature. 2019;574:575–580. doi: 10.1038/s41586-019-1678-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cheng Z., Tang Y., Chen Y., Kim S., Liu H., Li S.S., Gu W., Zhao Y. Molecular characterization of propionyllysines in non-histone proteins. Mol. Cell Proteom. 2009;8:45–52. doi: 10.1074/mcp.M800224-MCP200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Sabari B.R., Tang Z., Huang H., Yong-Gonzalez V., Molina H., Kong H.E., Dai L., Shimada M., Cross J.R., Zhao Y., et al. Intracellular crotonyl-CoA stimulates transcription through p300-catalyzed histone crotonylation. Mol. Cell. 2015;58:203–215. doi: 10.1016/j.molcel.2015.02.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang T., Chen K., Yao W., Zheng R., He Q., Xia J., Li J., Shao Y., Zhang L., Huang L., et al. Acetylation of lactate dehydrogenase B drives NAFLD progression by impairing lactate clearance. J. Hepatol. 2021;74:1038–1052. doi: 10.1016/j.jhep.2020.11.028. [DOI] [PubMed] [Google Scholar]
- 51.Lv L., Xu Y.P., Zhao D., Li F.L., Wang W., Sasaki N., Jiang Y., Zhou X., Li T.T., Guan K.L., et al. Mitogenic and oncogenic stimulation of K433 acetylation promotes PKM2 protein kinase activity and nuclear localization. Mol. Cell. 2013;52:340–352. doi: 10.1016/j.molcel.2013.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Huang H., Tang S., Ji M., Tang Z., Shimada M., Liu X., Qi S., Locasale J.W., Roeder R.G., Zhao Y., et al. p300-Mediated Lysine 2-Hydroxyisobutyrylation Regulates Glycolysis. Mol. Cell. 2018;70:663–678.e6. doi: 10.1016/j.molcel.2018.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Zhu Z., Han Z., Halabelian L., Yang X., Ding J., Zhang N., Ngo L., Song J., Zeng H., He M., et al. Identification of lysine isobutyrylation as a new histone modification mark. Nucleic. Acids. Res. 2021;49:177–189. doi: 10.1093/nar/gkaa1176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Jiang Y., Li Y., Liu C., Zhang L., Lv D., Weng Y., Cheng Z., Chen X., Zhan J., Zhang H. Isonicotinylation is a histone mark induced by the anti-tuberculosis first-line drug isoniazid. Nat. Commun. 2021;12:5548. doi: 10.1038/s41467-021-25867-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Huang H., Zhang D., Weng Y., Delaney K., Tang Z., Yan C., Qi S., Peng C., Cole P.A., Roeder R.G., et al. The regulatory enzymes and protein substrates for the lysine β-hydroxybutyrylation pathway. Sci. Adv. 2021;7:eabe2771. doi: 10.1126/sciadv.abe2771. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Kaczmarska Z., Ortega E., Goudarzi A., Huang H., Kim S., Marquez J.A., Zhao Y., Khochbin S., Panne D. Structure of p300 in complex with acyl-CoA variants. Nat. Chem. Biol. 2017;13:21–29. doi: 10.1038/nchembio.2217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Thomas S.P., Denu J.M. Short-chain fatty acids activate acetyltransferase p300. eLife. 2021;10:1–23. doi: 10.7554/eLife.72171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Zhao L.J., Loewenstein P.M., Green M. The adenovirus E1A oncoprotein N-terminal transcriptional repression domain enhances p300 autoacetylation and inhibits histone H3 Lys18 acetylation. Genes Cancer. 2015;6:30–37. doi: 10.18632/genesandcancer.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Liu C., Pan Y., Wang X., Lu J., Huang B., Li X. Activation of RASSF2A by p300 induces late apoptosis through histone hyperacetylation. Cell Biol. Int. 2010;34:1133–1139. doi: 10.1042/CBI20090388. [DOI] [PubMed] [Google Scholar]
- 60.Bandyopadhyay D., Okan N.A., Bales E., Nascimento L., Cole P.A., Medrano E.E. Down-regulation of p300/CBP histone acetyltransferase activates a senescence checkpoint in human melanocytes. Cancer Res. 2002;62:6231–6239. [PubMed] [Google Scholar]
- 61.Haery L., Lugo-Picó J.G., Henry R.A., Andrews A.J., Gilmore T.D. Histone acetyltransferase-deficient p300 mutants in diffuse large B cell lymphoma have altered transcriptional regulatory activities and are required for optimal cell growth. Mol. Cancer. 2014;13:1–13. doi: 10.1186/1476-4598-13-29. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sawant M., Mahajan K., Renganathan A., Weimholt C., Luo J., Kukshal V., Jez J.M., Jeon M.S., Zhang B., Li T., et al. Chronologically modified androgen receptor in recurrent castration-resistant prostate cancer and its therapeutic targeting. Sci. Transl. Med. 2022;14:eabg4132. doi: 10.1126/scitranslmed.abg4132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ianculescu I., Wu D.Y., Siegmund K.D., Stallcup M.R. Selective roles for cAMP response element-binding protein binding protein and p300 protein as coregulators for androgen-regulated gene expression in advanced prostate cancer cells. J. Biol. Chem. 2012;287:4000–4013. doi: 10.1074/jbc.M111.300194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Waddell A., Mahmud I., Ding H., Huo Z., Liao D. Pharmacological Inhibition of CBP/p300 Blocks Estrogen Receptor Alpha (ERalpha) Function through Suppressing Enhancer H3K27 Acetylation in Luminal Breast Cancer. Cancers. 2021;13:2799. doi: 10.3390/cancers13112799. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Cho M.H., Park J.H., Choi H.J., Park M.K., Won H.Y., Park Y.J., Lee C.H., Oh S.H., Song Y.S., Kim H.S., et al. DOT1L cooperates with the c-Myc-p300 complex to epigenetically derepress CDH1 transcription factors in breast cancer progression. Nat. Commun. 2015;6:7821. doi: 10.1038/ncomms8821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Jin K., Zhou W., Han X., Wang Z., Li B., Jeffries S., Tao W., Robbins D.J., Capobianco A.J. Acetylation of Mastermind-like 1 by p300 Drives the Recruitment of NACK to Initiate Notch-Dependent Transcription. Cancer Res. 2017;77:4228–4237. doi: 10.1158/0008-5472.CAN-16-3156. [DOI] [PubMed] [Google Scholar]
- 67.He H., Wang D., Yao H., Wei Z., Lai Y., Hu J., Liu X., Wang Y., Zhou H., Wang N., et al. Transcriptional factors p300 and MRTF-A synergistically enhance the expression of migration-related genes in MCF-7 breast cancer cells. Biochem. Biophys. Res. Commun. 2015;467:813–820. doi: 10.1016/j.bbrc.2015.10.060. [DOI] [PubMed] [Google Scholar]
- 68.Wang W., Chen Y., Du R., Xia X., Zhang Y., Guo H., Ma J., Tian J., Wang S. C/EBPbeta enhances immunosuppression activity of myeloid-derived suppressor cells by a P300-mediated acetylation modification. Inflamm. Res. 2022;71:1547–1557. doi: 10.1007/s00011-022-01639-2. [DOI] [PubMed] [Google Scholar]
- 69.Hogg S.J., Motorna O., Cluse L.A., Johanson T.M., Coughlan H.D., Raviram R., Myers R.M., Costacurta M., Todorovski I., Pijpers L., et al. Targeting histone acetylation dynamics and oncogenic transcription by catalytic P300/CBP inhibition. Mol. Cell. 2021;81:2183–2200.e2113. doi: 10.1016/j.molcel.2021.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Liu J., He D., Cheng L., Huang C., Zhang Y., Rao X., Kong Y., Li C., Zhang Z., Liu J., et al. p300/CBP inhibition enhances the efficacy of programmed death-ligand 1 blockade treatment in prostate cancer. Oncogene. 2020;39:3939–3951. doi: 10.1038/s41388-020-1270-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Cai L.Y., Chen S.J., Xiao S.H., Sun Q.J., Ding C.H., Zheng B.N., Zhu X.Y., Liu S.Q., Yang F., Yang Y.X., et al. Targeting p300/CBP Attenuates Hepatocellular Carcinoma Progression through Epigenetic Regulation of Metabolism. Cancer Res. 2021;81:860–872. doi: 10.1158/0008-5472.CAN-20-1323. [DOI] [PubMed] [Google Scholar]
- 72.Moreno-Yruela C., Zhang D., Wei W., Bæk M., Liu W., Gao J., Danková D., Nielsen A.L., Bolding J.E., Yang L., et al. Class I histone deacetylases (HDAC1–3) are histone lysine delactylases. Sci. Adv. 2022;8:1–15. doi: 10.1126/sciadv.abi6696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Li X., Yang Y., Zhang B., Lin X., Fu X., An Y., Zou Y., Wang J.X., Wang Z., Yu T. Lactate metabolism in human health and disease. Signal. Transduct. Target. Ther. 2022;7:305. doi: 10.1038/s41392-022-01151-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhou J., Zhan S., Tan W., Cheng R., Gong H., Zhu Q. P300 binds to and acetylates MTA2 to promote colorectal cancer cells growth. Biochem. Biophys. Res. Commun. 2014;444:387–390. doi: 10.1016/j.bbrc.2014.01.062. [DOI] [PubMed] [Google Scholar]
- 75.Hu H., Zhu W., Qin J., Chen M., Gong L., Li L., Liu X., Tao Y., Yin H., Zhou H., et al. Acetylation of PGK1 promotes liver cancer cell proliferation and tumorigenesis. Hepatology. 2017;65:515–528. doi: 10.1002/hep.28887. [DOI] [PubMed] [Google Scholar]
- 76.Liao Z.W., Zhao L., Cai M.Y., Xi M., He L.R., Yu F., Zhou T.C., Liu M.Z. P300 promotes migration, invasion and epithelial-mesenchymal transition in a nasopharyngeal carcinoma cell line. Oncol. Lett. 2017;13:763–769. doi: 10.3892/ol.2016.5491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Zhong J., Ding L., Bohrer L.R., Pan Y., Liu P., Zhang J., Sebo T.J., Karnes R.J., Tindall D.J., van Deursen J., et al. p300 acetyltransferase regulates androgen receptor degradation and PTEN-deficient prostate tumorigenesis. Cancer Res. 2014;74:1870–1880. doi: 10.1158/0008-5472.CAN-13-2485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Welti J., Sharp A., Brooks N., Yuan W., McNair C., Chand S.N., Pal A., Figueiredo I., Riisnaes R., Gurel B., et al. Targeting the p300/CBP Axis in Lethal Prostate Cancer. Cancer Discov. 2021;11:1118–1137. doi: 10.1158/2159-8290.CD-20-0751. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Han X., Xiang X., Yang H., Zhang H., Liang S., Wei J., Yu J. p300-Catalyzed Lysine Crotonylation Promotes the Proliferation, Invasion, and Migration of HeLa Cells via Heterogeneous Nuclear Ribonucleoprotein A1. Anal. Cell. Pathol. 2020;2020:5632342. doi: 10.1155/2020/5632342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Kitabayashi I., Aikawa Y., Yokoyama A., Hosoda F., Nagai M., Kakazu N., Abe T., Ohki M. Fusion of MOZ and p300 histone acetyltransferases in acute monocytic leukemia with a t(8;22)(p11;q13) chromosome translocation. Leukemia. 2001;15:89–94. doi: 10.1038/sj.leu.2401983. [DOI] [PubMed] [Google Scholar]
- 81.Palermo R., Checquolo S., Giovenco A., Grazioli P., Kumar V., Campese A.F., Giorgi A., Napolitano M., Canettieri G., Ferrara G., et al. Acetylation controls Notch3 stability and function in T-cell leukemia. Oncogene. 2012;31:3807–3817. doi: 10.1038/onc.2011.533. [DOI] [PubMed] [Google Scholar]
- 82.Tottone L., Zhdanovskaya N., Carmona Pestana A., Zampieri M., Simeoni F., Lazzari S., Ruocco V., Pelullo M., Caiafa P., Felli M.P., et al. Histone Modifications Drive Aberrant Notch3 Expression/Activity and Growth in T-ALL. Front. Oncol. 2019;9:198. doi: 10.3389/fonc.2019.00198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Ono H., Basson M.D., Ito H. P300 inhibition enhances gemcitabine-induced apoptosis of pancreatic cancer. Oncotarget. 2016;7:51301–51310. doi: 10.18632/oncotarget.10117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Mahmud Z., Gomes A.R., Lee H.J., Aimjongjun S., Jiramongkol Y., Yao S., Zona S., Alasiri G., Gong G., Yague E., et al. EP300 and SIRT1/6 Co-Regulate Lapatinib Sensitivity Via Modulating FOXO3-Acetylation and Activity in Breast Cancer. Cancers. 2019;11:1067. doi: 10.3390/cancers11081067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Mladek A.C., Yan H., Tian S., Decker P.A., Burgenske D.M., Bakken K., Hu Z., He L., Connors M.A., Carlson B.L., et al. RBBP4-p300 axis modulates expression of genes essential for cell survival and is a potential target for therapy in glioblastoma. Neuro. Oncol. 2022;24:1261–1272. doi: 10.1093/neuonc/noac051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Du C., Huang D., Peng Y., Yao Y., Zhao Y., Yang Y., Wang H., Cao L., Zhu W.G., Gu J. 5-Fluorouracil targets histone acetyltransferases p300/CBP in the treatment of colorectal cancer. Cancer Lett. 2017;400:183–193. doi: 10.1016/j.canlet.2017.04.033. [DOI] [PubMed] [Google Scholar]
- 87.Gang X., Yang Y., Zhong J., Jiang K., Pan Y., Jeffrey Karnes R., Zhang J., Xu W., Wang G., Huang H. P300 acetyltransferase regulates fatty acid synthase expression, lipid metabolism and prostate cancer growth. Oncotarget. 2016;7:15135–15149. doi: 10.18632/oncotarget.7715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Ponugoti B., Kim D.H., Xiao Z., Smith Z., Miao J., Zang M., Wu S.Y., Chiang C.M., Veenstra T.D., Kemper J.K. SIRT1 deacetylates and inhibits SREBP-1C activity in regulation of hepatic lipid metabolism. J. Biol. Chem. 2010;285:33959–33970. doi: 10.1074/jbc.M110.122978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Xiao X.S., Cai M.Y., Chen J.W., Guan X.Y., Kung H.F., Zeng Y.X., Xie D. High Expression of p300 in Human Breast Cancer Correlates with Tumor Recurrence and Predicts Adverse Prognosis. Chin. J. Cancer Res. 2011;23:201–207. doi: 10.1007/s11670-011-0201-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Yokomizo C., Yamaguchi K., Itoh Y., Nishimura T., Umemura A., Minami M., Yasui K., Mitsuyoshi H., Fujii H., Tochiki N., et al. High expression of p300 in HCC predicts shortened overall survival in association with enhanced epithelial mesenchymal transition of HCC cells. Cancer Lett. 2011;310:140–147. doi: 10.1016/j.canlet.2011.06.030. [DOI] [PubMed] [Google Scholar]
- 91.Li Y., Yang H.-X., Luo R.-Z., Zhang Y., Li M., Wang X., Jia W.-H. High Expression of p300 Has an Unfavorable Impact on Survival in Resectable Esophageal Squamous Cell Carcinoma. Ann. Thorac. Surg. 2011;91:1531–1538. doi: 10.1016/j.athoracsur.2010.12.012. [DOI] [PubMed] [Google Scholar]
- 92.Chen M.K., Cai M.Y., Luo R.Z., Tian X., Liao Q.M., Zhang X.Y., Han J.D. Overexpression of p300 correlates with poor prognosis in patients with cutaneous squamous cell carcinoma. Br. J. Derm. 2015;172:111–119. doi: 10.1111/bjd.13226. [DOI] [PubMed] [Google Scholar]
- 93.Attar N., Kurdistani S.K. Exploitation of EP300 and CREBBP Lysine Acetyltransferases by Cancer. Cold. Spring Harb. Perspect. Med. 2017;7:a026534. doi: 10.1101/cshperspect.a026534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Manuyakorn A., Paulus R., Farrell J., Dawson N.A., Tze S., Cheung-Lau G., Hines O.J., Reber H., Seligson D.B., Horvath S., et al. Cellular histone modification patterns predict prognosis and treatment response in resectable pancreatic adenocarcinoma: Results from RTOG 9704. J. Clin. Oncol. 2010;28:1358–1365. doi: 10.1200/JCO.2009.24.5639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Seligson D.B., Horvath S., Shi T., Yu H., Tze S., Grunstein M., Kurdistani S.K. Global histone modification patterns predict risk of prostate cancer recurrence. Nature. 2005;435:1262–1266. doi: 10.1038/nature03672. [DOI] [PubMed] [Google Scholar]
- 96.Mosashvilli D., Kahl P., Mertens C., Holzapfel S., Rogenhofer S., Hauser S., Buttner R., Von Ruecker A., Muller S.C., Ellinger J. Global histone acetylation levels: Prognostic relevance in patients with renal cell carcinoma. Cancer Sci. 2010;101:2664–2669. doi: 10.1111/j.1349-7006.2010.01717.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Seligson D.B., Horvath S., McBrian M.A., Mah V., Yu H., Tze S., Wang Q., Chia D., Goodglick L., Kurdistani S.K. Global levels of histone modifications predict prognosis in different cancers. Am. J. Pathol. 2009;174:1619–1628. doi: 10.2353/ajpath.2009.080874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Wang Z., Yang X., Liu C., Li X., Zhang B., Wang B., Zhang Y., Song C., Zhang T., Liu M., et al. Acetylation of PHF5A Modulates Stress Responses and Colorectal Carcinogenesis through Alternative Splicing-Mediated Upregulation of KDM3A. Mol. Cell. 2019;74:1250–1263.e1256. doi: 10.1016/j.molcel.2019.04.009. [DOI] [PubMed] [Google Scholar]
- 99.Lazarova D.L., Wong T., Chiaro C., Drago E., Bordonaro M. p300 Influences Butyrate-Mediated WNT Hyperactivation In Colorectal Cancer Cells. J. Cancer. 2013;4:491–501. doi: 10.7150/jca.6582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Gao X.N., Lin J., Ning Q.Y., Gao L., Yao Y.S., Zhou J.H., Li Y.H., Wang L.L., Yu L. A histone acetyltransferase p300 inhibitor C646 induces cell cycle arrest and apoptosis selectively in AML1-ETO-positive AML cells. PLoS ONE. 2013;8:e55481. doi: 10.1371/journal.pone.0055481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Dutta R., Tiu B., Sakamoto K.M. CBP/p300 acetyltransferase activity in hematologic malignancies. Mol. Genet. Metab. 2016;119:37–43. doi: 10.1016/j.ymgme.2016.06.013. [DOI] [PubMed] [Google Scholar]
- 102.Garcia-Carpizo V., Ruiz-Llorente S., Sarmentero J., Grana-Castro O., Pisano D.G., Barrero M.J. CREBBP/EP300 bromodomains are critical to sustain the GATA1/MYC regulatory axis in proliferation. Epigenetics Chromatin. 2018;11:30. doi: 10.1186/s13072-018-0197-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Ji C., Xu W., Ding H., Chen Z., Shi C., Han J., Yu L., Qiao N., Zhang Y., Cao X., et al. The p300 Inhibitor A-485 Exerts Antitumor Activity in Growth Hormone Pituitary Adenoma. J. Clin. Endocrinol. Metab. 2022;107:e2291–e2300. doi: 10.1210/clinem/dgac128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Chen Q., Yang B., Liu X., Zhang X.D., Zhang L., Liu T. Histone acetyltransferases CBP/p300 in tumorigenesis and CBP/p300 inhibitors as promising novel anticancer agents. Theranostics. 2022;12:4935–4948. doi: 10.7150/thno.73223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Stimson L., Rowlands M.G., Newbatt Y.M., Smith N.F., Raynaud F.I., Rogers P., Bavetsias V., Gorsuch S., Jarman M., Bannister A., et al. Isothiazolones as inhibitors of PCAF and p300 histone acetyltransferase activity. Mol. Cancer Ther. 2005;4:1521–1532. doi: 10.1158/1535-7163.MCT-05-0135. [DOI] [PubMed] [Google Scholar]
- 106.Guo L., Li H., Fan T., Ma Y., Wang L. Synergistic efficacy of curcumin and anti-programmed cell death-1 in hepatocellular carcinoma. Life Sci. 2021;279:119359. doi: 10.1016/j.lfs.2021.119359. [DOI] [PubMed] [Google Scholar]
- 107.Yang H., Pinello C.E., Luo J., Li D., Wang Y., Zhao L.Y., Jahn S.C., Saldanha S.A., Chase P., Planck J., et al. Small-molecule inhibitors of acetyltransferase p300 identified by high-throughput screening are potent anticancer agents. Mol. Cancer Ther. 2013;12:610–620. doi: 10.1158/1535-7163.MCT-12-0930. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Ogiwara H., Sasaki M., Mitachi T., Oike T., Higuchi S., Tominaga Y., Kohno T. Targeting p300 Addiction in CBP-Deficient Cancers Causes Synthetic Lethality by Apoptotic Cell Death due to Abrogation of MYC Expression. Cancer Discov. 2016;6:430–445. doi: 10.1158/2159-8290.CD-15-0754. [DOI] [PubMed] [Google Scholar]
- 109.Zhang B., Chen D., Liu B., Dekker F.J., Quax W.J. A novel histone acetyltransferase inhibitor A485 improves sensitivity of non-small-cell lung carcinoma cells to TRAIL. Biochem. Pharm. 2020;175:113914. doi: 10.1016/j.bcp.2020.113914. [DOI] [PubMed] [Google Scholar]
- 110.Sharma V.K., Lahiri M. Interplay between p300 and HDAC1 regulate acetylation and stability of Api5 to regulate cell proliferation. Sci. Rep. 2021;11:16427. doi: 10.1038/s41598-021-95941-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Avantaggiati M.L., Ogryzko V., Gardner K., Giordano A., Levine A.S., Kelly K. Recruitment of p300/CBP in p53-dependent signal pathways. Cell. 1997;89:1175–1184. doi: 10.1016/S0092-8674(00)80304-9. [DOI] [PubMed] [Google Scholar]
- 112.Fu L., Huang W., Jing Y., Jiang M., Zhao Y., Shi J., Huang S., Xue X., Zhang Q., Tang J., et al. AML1-ETO triggers epigenetic activation of early growth response gene l, inducing apoptosis in t(8;21) acute myeloid leukemia. FEBS J. 2014;281:1123–1131. doi: 10.1111/febs.12673. [DOI] [PubMed] [Google Scholar]
- 113.Janknecht R. The versatile functions of the transcriptional coactivators p300 and CBP and their roles in disease. Histol. Histopathol. 2002;17:657–668. doi: 10.14670/HH-17.657. [DOI] [PubMed] [Google Scholar]
- 114.Gopalakrishna Iyer N., Chin S.F., Ozdag H., Daigo Y., Hu D.E., Cariati M., Brindle K., Aparicio S., Caldas C. p300 regulates p53-dependent apoptosis after DNA damage in colorectal cancer cells by modulation of PUMA/p21 levels. Proc. Natl. Acad. Sci. USA. 2004;101:7386–7391. doi: 10.1073/pnas.0401002101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Shiota M., Yokomizo A., Kashiwagi E., Tada Y., Inokuchi J., Tatsugami K., Kuroiwa K., Uchiumi T., Seki N., Naito S. Foxo3a expression and acetylation regulate cancer cell growth and sensitivity to cisplatin. Cancer Sci. 2010;101:1177–1185. doi: 10.1111/j.1349-7006.2010.01503.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Huang X., Yan J., Zhang M., Wang Y., Chen Y., Fu X., Wei R., Zheng X.L., Liu Z., Zhang X., et al. Targeting Epigenetic Crosstalk as a Therapeutic Strategy for EZH2-Aberrant Solid Tumors. Cell. 2018;175:186–199.e119. doi: 10.1016/j.cell.2018.08.058. [DOI] [PubMed] [Google Scholar]
- 117.Liu Y., Mayo M.W., Nagji A.S., Hall E.H., Shock L.S., Xiao A., Stelow E.B., Jones D.R. BRMS1 suppresses lung cancer metastases through an E3 ligase function on histone acetyltransferase p300. Cancer Res. 2013;73:1308–1317. doi: 10.1158/0008-5472.CAN-12-2489. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Zhou F., Liu Q., Zhang L., Zhu Q., Wang S., Zhu K., Deng R., Liu Y., Yuan G., Wang X., et al. Selective inhibition of CBP/p300 HAT by A-485 results in suppression of lipogenesis and hepatic gluconeogenesis. Cell Death Dis. 2020;11:745. doi: 10.1038/s41419-020-02960-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Yuan Y., Yuan H.F., Geng Y., Zhao L.N., Yun H.L., Wang Y.F., Yang G., Zhang X.D. Aspirin modulates 2-hydroxyisobutyrylation of ENO1K281 to attenuate the glycolysis and proliferation of hepatoma cells. Biochem. Biophys. Res. Commun. 2021;560:172–178. doi: 10.1016/j.bbrc.2021.04.083. [DOI] [PubMed] [Google Scholar]
- 120.Yang K., Fan M., Wang X., Xu J., Wang Y., Tu F., Gill P.S., Ha T., Liu L., Williams D.L., et al. Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell Death Differ. 2022;29:133–146. doi: 10.1038/s41418-021-00841-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Bundy J.G., Iyer N.G., Gentile M.S., Hu D.E., Kettunen M., Maia A.T., Thorne N.P., Brenton J.D., Caldas C., Brindle K.M. Metabolic consequences of p300 gene deletion in human colon cancer cells. Cancer Res. 2006;66:7606–7614. doi: 10.1158/0008-5472.CAN-05-2999. [DOI] [PubMed] [Google Scholar]
- 122.Shang S., Yang Y.W., Chen F., Yu L., Shen S.H., Li K., Cui B., Lv X.X., Zhang C., Yang C., et al. TRIB3 reduces CD8+ T cell infiltration and induces immune evasion by repressing the STAT1-CXCL10 axis in colorectal cancer. Sci. Transl. Med. 2022;14:eabf0992. doi: 10.1126/scitranslmed.abf0992. [DOI] [PubMed] [Google Scholar]
- 123.Giotopoulos G., Chan W.I., Horton S.J., Ruau D., Gallipoli P., Fowler A., Crawley C., Papaemmanuil E., Campbell P.J., Gottgens B., et al. The epigenetic regulators CBP and p300 facilitate leukemogenesis and represent therapeutic targets in acute myeloid leukemia. Oncogene. 2016;35:279–289. doi: 10.1038/onc.2015.92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Lasko L.M., Jakob C.G., Edalji R.P., Qiu W., Montgomery D., Digiammarino E.L., Hansen T.M., Risi R.M., Frey R., Manaves V., et al. Discovery of a selective catalytic p300/CBP inhibitor that targets lineage-specific tumours. Nature. 2017;550:128–132. doi: 10.1038/nature24028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Gajer J.M., Furdas S.D., Grunder A., Gothwal M., Heinicke U., Keller K., Colland F., Fulda S., Pahl H.L., Fichtner I., et al. Histone acetyltransferase inhibitors block neuroblastoma cell growth in vivo. Oncogenesis. 2015;4:e137. doi: 10.1038/oncsis.2014.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Tan J., Chen B., He L., Tang Y., Jiang Z., Yin G., Wang J., Jiang X. Anacardic acid (6-pentadecylsalicylic acid) induces apoptosis of prostate cancer cells through inhibition of androgen receptor and activation of p53 signaling. Chin. J. Cancer. Res. 2012;24:275–283. doi: 10.1007/s11670-012-0264-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Balasubramanyam K., Varier R.A., Altaf M., Swaminathan V., Siddappa N.B., Ranga U., Kundu T.K. Curcumin, a novel p300/CREB-binding protein-specific inhibitor of acetyltransferase, represses the acetylation of histone/nonhistone proteins and histone acetyltransferase-dependent chromatin transcription. J. Biol. Chem. 2004;279:51163–51171. doi: 10.1074/jbc.M409024200. [DOI] [PubMed] [Google Scholar]
- 128.Zheng M., Zhu J., Lu T., Liu L., Sun H., Liu Z., Tian J. p300-mediated histone acetylation is essential for the regulation of GATA4 and MEF2C by BMP2 in H9c2 cells. Cardiovasc. Toxicol. 2013;13:316–322. doi: 10.1007/s12012-013-9212-4. [DOI] [PubMed] [Google Scholar]
- 129.Liu L., Fu Y., Zheng Y., Ma M., Wang C. Curcumin inhibits proteasome activity in triple-negative breast cancer cells through regulating p300/miR-142-3p/PSMB5 axis. Phytomedicine. 2020;78:153312. doi: 10.1016/j.phymed.2020.153312. [DOI] [PubMed] [Google Scholar]
- 130.Bowers E.M., Yan G., Mukherjee C., Orry A., Wang L., Holbert M.A., Crump N.T., Hazzalin C.A., Liszczak G., Yuan H., et al. Virtual ligand screening of the p300/CBP histone acetyltransferase: Identification of a selective small molecule inhibitor. Chem. Biol. 2010;17:471–482. doi: 10.1016/j.chembiol.2010.03.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Mantelingu K., Reddy B.A., Swaminathan V., Kishore A.H., Siddappa N.B., Kumar G.V., Nagashankar G., Natesh N., Roy S., Sadhale P.P., et al. Specific inhibition of p300-HAT alters global gene expression and represses HIV replication. Chem. Biol. 2007;14:645–657. doi: 10.1016/j.chembiol.2007.04.011. [DOI] [PubMed] [Google Scholar]
- 132.Ravindra K.C., Selvi B.R., Arif M., Reddy B.A., Thanuja G.R., Agrawal S., Pradhan S.K., Nagashayana N., Dasgupta D., Kundu T.K. Inhibition of lysine acetyltransferase KAT3B/p300 activity by a naturally occurring hydroxynaphthoquinone, plumbagin. J. Biol. Chem. 2009;284:24453–24464. doi: 10.1074/jbc.M109.023861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Kim E., Zucconi B.E., Wu M., Nocco S.E., Meyers D.J., McGee J.S., Venkatesh S., Cohen D.L., Gonzalez E.C., Ryu B., et al. MITF Expression Predicts Therapeutic Vulnerability to p300 Inhibition in Human Melanoma. Cancer Res. 2019;79:2649–2661. doi: 10.1158/0008-5472.CAN-18-2331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Huang M., Huang J., Zheng Y., Sun Q. Histone acetyltransferase inhibitors: An overview in synthesis, structure-activity relationship and molecular mechanism. Eur. J. Med. Chem. 2019;178:259–286. doi: 10.1016/j.ejmech.2019.05.078. [DOI] [PubMed] [Google Scholar]
- 135.Zou L.J., Xiang Q.P., Xue X.Q., Zhang C., Li C.C., Wang C., Li Q., Wang R., Wu S., Zhou Y.L., et al. Y08197 is a novel and selective CBP/EP300 bromodomain inhibitor for the treatment of prostate cancer. Acta. Pharm. Sin. 2019;40:1436–1447. doi: 10.1038/s41401-019-0237-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Ravindra K.C., Narayan V., Lushington G.H., Peterson B.R., Prabhu K.S. Targeting of histone acetyltransferase p300 by cyclopentenone prostaglandin Delta(12)-PGJ(2) through covalent binding to Cys(1438) Chem. Res. Toxicol. 2012;25:337–347. doi: 10.1021/tx200383c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Ghosh S., Taylor A., Chin M., Huang H.R., Conery A.R., Mertz J.A., Salmeron A., Dakle P.J., Mele D., Cote A., et al. Regulatory T Cell Modulation by CBP/EP300 Bromodomain Inhibition. J. Biol. Chem. 2016;291:13014–13027. doi: 10.1074/jbc.M115.708560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Gao Y., Nihira N.T., Bu X., Chu C., Zhang J., Kolodziejczyk A., Fan Y., Chan N.T., Ma L., Liu J., et al. Acetylation-dependent regulation of PD-L1 nuclear translocation dictates the efficacy of anti-PD-1 immunotherapy. Nat. Cell Biol. 2020;22:1064–1075. doi: 10.1038/s41556-020-0562-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Diesch J., Le Pannerer M.M., Winkler R., Casquero R., Muhar M., van der Garde M., Maher M., Herraez C.M., Bech-Serra J.J., Fellner M., et al. Inhibition of CBP synergizes with the RNA-dependent mechanisms of Azacitidine by limiting protein synthesis. Nat. Commun. 2021;12:6060. doi: 10.1038/s41467-021-26258-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Yang Y., Zhang R., Li Z., Mei L., Wan S., Ding H., Chen Z., Xing J., Feng H., Han J., et al. Discovery of Highly Potent, Selective, and Orally Efficacious p300/CBP Histone Acetyltransferases Inhibitors. J. Med. Chem. 2020;63:1337–1360. doi: 10.1021/acs.jmedchem.9b01721. [DOI] [PubMed] [Google Scholar]
- 141.Hammitzsch A., Tallant C., Fedorov O., O’Mahony A., Brennan P.E., Hay D.A., Martinez F.O., Al-Mossawi M.H., de Wit J., Vecellio M., et al. CBP30, a selective CBP/p300 bromodomain inhibitor, suppresses human Th17 responses. Proc. Natl. Acad. Sci. USA. 2015;112:10768–10773. doi: 10.1073/pnas.1501956112. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
Not applicable.