

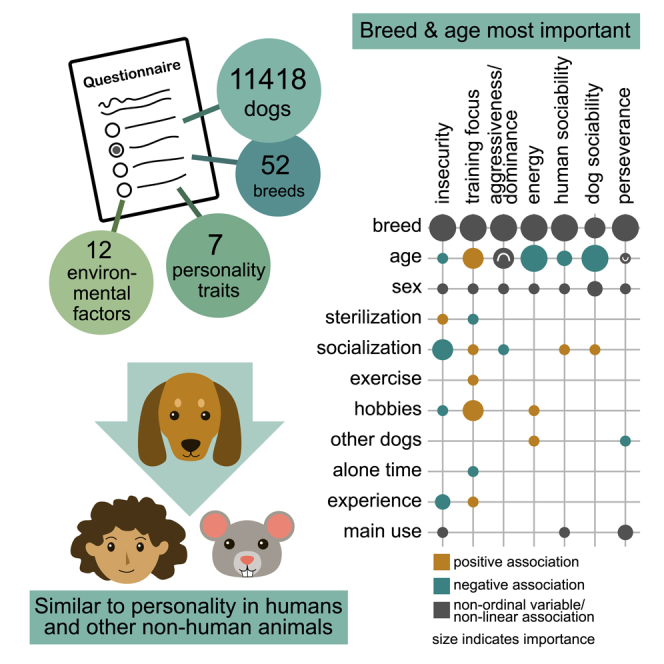
Summary
Personality in pets and other domesticated animals is important for their well-being and it can also influence human-animal relationships. Genetic and environmental factors influencing unwanted behavior in dogs are somewhat well known, but the factors influencing dog personality remain understudied. Here we examined environmental and demographic factors associated with seven broad personality traits in a survey of over 11,000 dogs. We utilized linear models and extensive model validation to examine the factors that have the most significant influences on personality and calculated effect sizes to assess the importance of these variables. Breed and age had the strongest associations with dog personality traits. Some environmental factors, especially puppyhood socialization, were also associated with personality. All factors had small effect sizes, highlighting that a lot of variation in personality remains unexplained. Our results indicate that personality traits are complex and strikingly similar in dogs, humans, and other nonhuman animals.
Subject areas: Canine behavior, Zoology, Behavioral neuroscience
Graphical abstract
Highlights
-
•
Breed was the most important variable and they showed large personality differences
-
•
Age strongly associated with aggressiveness/dominance, energy, and dog sociability
-
•
Puppyhood socialization was the most important environmental variable
-
•
The results highlight the similarity of personality in humans and nonhuman animals
Canine behavior; Zoology; Behavioral neuroscience
Introduction
Animal personality is often defined as behavioral differences between individuals that are relatively stable in time and across contexts.1,2 Personality is highly important for welfare and well-being, as it is a predictor of mortality and physical and mental health in both humans and nonhuman animals.3,4,5,6,7,8 In companion animals, such as pet dogs, personality can also influence the pet-owner relationship.9,10,11 Undesirable behavior can even lead to relinquishment to a shelter or euthanasia.12,13,14 As personality is shared by at least most of the animal kingdom,15,16,17 the study of dog personality can deepen the understanding of personality throughout species. Furthermore, the study of dog personality traits may improve the well-being of both the pet and its owner.
Personality traits are highly genetic, as heritability estimates in humans and across the animal kingdom often range between 40 and 50%.18,19,20,21 However, some variance in personality is explained by the environmental component as well.2,22,23,24,25 The influence of early life environment on personality seems to be especially pronounced. For example, early life adversity is known to heighten fear-related traits and anxiety in humans and rodents.26,27,28,29 Even though personality traits are relatively stable after ontogeny, they also change with age. In humans, conscientiousness and agreeableness often increase whereas neuroticism and extraversion decrease with age.30,31,32
Dog personality has interested researchers for a long time and previous studies have examined the structure of personality, which in many studies consists of traits such as fearfulness/insecurity, energy/activity, training focus, aggressiveness, and sociability.33,34,35,36,37,38,39,40,41 Despite this knowledge on personality structure, the factors influencing dog personality are still mostly undiscovered. Previous studies have indicated that, for example, breeds differ in personalities36,42,43,44,45 and that dogs show similar age differences in personality as humans.10,36,46 Nevertheless, one recent study challenged these breed differences by showing that breed explains only a small part of behavioral variation.47
Here, we utilized a reliable and validated dog personality survey35 and a large dataset of more than eleven thousand pet dogs to examine the demographic and environmental factors associated with seven dog personality traits. We fit general and generalized linear models to assess the independent influence of variables and performed extensive model validation to include only variables with a large effect in the models.
Results
We used a dog personality questionnaire and previously reduced 63 adjective statements into seven dog personality traits: insecurity, training focus, aggressiveness/dominance, energy, human sociability, dog sociability, and perseverance. In this present study, we fit general and generalized linear models to study the environmental and demographic factors associated with these dog personality traits. The possible explanatory variables included the dog’s sex, age, and breed, and 12 variables related to the dog’s past or present environment. We performed stringent model selection and validation to include only the most important variables in the models and also calculated the absolute and relative effect sizes of variables. Final models of all seven traits included all demographic variables (dog’s age, sex, and breed) and one to six environmental variables (Table 1).
Table 1.
Association of explanatory variables with 7 personality traits in the generalized linear models
| Variable | Insecurity |
Training focus |
Energy |
Perseverance |
DF | ||||
|---|---|---|---|---|---|---|---|---|---|
| F | p value | F | p value | F | p value | F | p value | ||
| Sex | 48.90 | 4.18e-11 | 44.48 | 3.46e-10 | 23.81 | 6.37e-06 | 2.24 | 0.206 | 1 |
| Age | 0.00 | 0.981 | 145.84 | 5.28e-15 | 1426.80 | 5.28e-15 | 62.69 | 5.73e-14 | 1 |
| Age2 | 16.36 | 2.16e-04 | 36.31 | 1.69e-08 | 42.47 | 8.96e-10 | 1 | ||
| Breed/breed group | 9.00 | 5.28e-15 | 6.37 | 5.28e-15 | 28.45 | 5.28e-15 | 11.89 | 5.28e-15 | 51 |
| Level of socialization | 58.45 | 5.28e-15 | 26.09 | 5.28e-15 | 4 | ||||
| Sterilization status | 106.86 | 5.28e-15 | 52.27 | 8.41e-12 | 1 | ||||
| Dog experience | 63.25 | 5.28e-15 | 24.75 | 1.37e-14 | 3 | ||||
| Main reason for ownership/use of the dog | 25.79 | 9.19e-11 | 59.49 | 5.28e-15 | 2 | ||||
| Frequency of hobbies | 12.92 | 1.95e-09 | 40.09 | 5.28e-15 | 23.29 | 5.28e-15 | 4 | ||
| Daily exercise | 19.86 | 1.24e-11 | 3 | ||||||
| Average time spent alone | 12.19 | 7.13e-09 | 4 | ||||||
| Other dogs in the household when acquired | 45.30 | 2.35e-10 | 69.23 | 5.28e-15 | 1 | ||||
| Variable |
Aggressiveness/dominance |
Human sociability |
Dog sociability |
DF | |||
|---|---|---|---|---|---|---|---|
| F | p value | F | p value | F | p value | ||
| Sex | 125.41 | 5.28e-15 | 7.79 | 0.013 | 194.46 | 5.28e-15 | 1 |
| Age | 448.82 | 5.28e-15 | 145.58 | 5.28e-15 | 1141.59 | 5.28e-15 | 1 |
| Age2 | 257.57 | 5.28e-15 | 315.55 | 5.28e-15 | 1 | ||
| Breed/breed group | 19.90 | 5.28e-15 | 12.20 | 5.28e-15 | 11.35 | 5.28e-15 | 51 |
| Level of socialization | 16.46 | 3.20e-12 | 12.85 | 2.22e-09 | 9.95 | 3.68e-07 | 4 |
| Main reason for ownership/use of the dog | 37.94 | 5.28e-15 | 2 | ||||
All p values are controlled for false discovery rate. Significant p values are indicated in bold. N = 11 418. F = F-test statistic, DF = degrees of freedom, age2 = age as a quadratic explanatory variable. Supplementary items.
Study cohort
Our study cohort included 11,418 dogs. The age of the dogs varied between 0.18 years and 17.13 years, with a mean of 5.18 years (SD = 3.44 years). Of these dogs, 53% were female and most dogs (76%) were intact. The dogs belonged to 52 breeds or breed groups, the most prevalent breed being the Finnish Lapponian Dog (480 dogs). For ranges, means, and standard deviations of the personality traits, see Table S11. A detailed description of the study cohort can be found in Table S12.
Effect sizes
Age was the most important variable based on relative importance in models of insecurity, training focus, energy, and dog sociability, whereas breed/breed group was the most important variable in models of aggressiveness/dominance, human sociability, and perseverance (Figures 1C, 1D, and S1). In all traits except dog sociability, sex was the least important variable.
Figure 1.

Breeds differ in personality traits and some personality traits are also associated with age and social environment
(A) Breed differences expressed as a heatmap, where colored circles indicate the difference between breed mean of a trait and population mean of the trait. Size of the circle indicates the magnitude of difference (between the breed mean factor score and population mean factor score) and color indicates the direction (blue = lower than average dog, red = higher than average dog). Small gray circles indicate that the breed mean score did not differ from trait mean (95% confidence limits including the mean).
(B) Distributions of the highest and lowest scoring breeds in human sociability. Lines indicate breed mean scores.
(C) Scaled variance-based variable importance (relative effect size) and Cohen’s f2 (absolute effect size) in insecurity.
(D) Scaled variance-based variable importance (relative effect size) and Cohen’s f2 (absolute effect size) in energy.
(E) Association of age with dog sociability. Gray area indicates 95% confidence limits.
(F) Association of the level of socialization experiences with insecurity. Error bars indicate 95% confidence limits. N = 11 418.
Cohen’s f2 estimates ranged between <0.001 and 0.128. Based on Cohen’s suggested cut-offs (small = 0.02, medium = 0.15, large = 0.35), all effects sizes were small or less than small.48 When comparing estimates between variables, breed/breed group obtained the highest value in all traits except dog sociability, in which age had the highest Cohen’s f2 (Figures 1C, 1D, and S1). Environmental variables, sterilization, and sex did not reach the cut-off for small effect sizes, except that socialization reached this cut-off in insecurity (f2 = 0.021).
Sex, age, and sterilization status
Sex was significantly associated with all personality traits except perseverance (Table 1, Figures S2A–S2G). However, the difference between female and male dogs was relatively small in many traits. The largest difference between sexes was discovered in dog sociability (z = 13.94, df = 1, p = 1.14e-41). Female dogs had higher mean scores in insecurity, training focus, and human sociability, and male dogs had higher mean scores in energy, aggressiveness/dominance, and dog sociability. For all pairwise contrasts, see Supplementary Information.
Age was significantly associated with all personality traits except insecurity, which, however, included a significant quadratic effect (Table 1, Figures 1E and S3). The model fit of age was improved by including the quadratic age effect also in training focus, perseverance, aggressiveness/dominance, and dog sociability. Mean score of training focus increased with age, whereas the mean scores of insecurity, energy, dog sociability, and human sociability decreased with age. Aggressiveness/dominance showed a nonlinear effect, with the highest mean score at 9 years of age. In contrast, the mean score in perseverance was lowest in 8-year-old dogs and highest in younger and older dogs.
Sterilization status was included in the insecurity and training focus models (Table 1, Figures S2H–S2I). Compared to intact dogs, neutered dogs had a higher mean score in insecurity (z = 10.21, df = 1, p = 1.14e-22) and a lower mean score in training focus (z = 7.23, df = 1, p = 7.93e-12).
Breed and breed group differences
Breed group was highly associated with all personality traits (Table 1) and breeds differed in all traits (Figures 1A, 1B, 2, and 3). The highest mean insecurity score was observed in Shetland Sheepdog and lowest in Bull-type terriers (difference: z = 8.69, df = 1, p = 1.11e-16). The highest mean training focus score was observed in the “other breed” group and the lowest in Bull-type terriers (difference: z = 6.47, df = 1, p = 1.16e-9). The highest mean energy score was detected in Parson-type terriers and the lowest in Livestock guardian dogs (difference: z = 15.69, df = 1, p = 1.55e-52). The highest mean perseverance score was obtained by Miniature Pinscher and the lowest by Golden Retriever (difference: z = 10.77, df = 1, p = 3.91e-25). In aggressiveness/dominance, there was also a significant difference between the highest-scoring breed group, fighting dogs, and the lowest-scoring breed, Golden Retriever (z = 13.27, df = 1, p = 8.51e-38). The highest mean human sociability score was discovered in Bull-type terriers and the lowest in Livestock guardian dogs (difference: z = 12.46, df = 1, p = 2.16e-33). Finally, in dog sociability, the highest mean score was obtained by Finnish Lapponian Dog and the lowest by Border Collie (difference: z = 13.80, df = 1, p = 7.41e-41). For all pairwise contrasts, see supplemental information.
Figure 2.

Association of breed group with personality traits insecurity, training focus, aggressiveness/dominance, and energy
(A–D) Association of the dog’s breed/breed group with (A) insecurity, (B) training focus, (C) aggressiveness/dominance, and (D) energy in the general and generalized linear models. Error bars indicate 95% confidence limits. N = 11 418.
Figure 3.

Association of breed group with personality traits human sociability, dog sociability, and perseverance
(A–C) Association of the dog’s breed/breed group with (A) human sociability, (B) dog sociability, and (C) perseverance in the linear models. Error bars indicate 95% confidence limits. N = 11 418.
When comparing distributions of trait scores in breeds with highest and lowest mean scores, these distributions mostly overlap, with both breeds including individuals in both ends of the distribution (Figures 1B and S4). However, in many traits, the majority of individuals resided in different parts of the distribution, but this divergence between breeds was highly dependent of the trait. Aggressiveness/dominance, energy, human sociability, and perseverance seemed to have the larger differences between breeds that had the highest and lowest scores in these traits, whereas insecurity, training focus, and dog sociability showed smaller differences.
Socialization
Level of socialization experiences in puppyhood (between 7 weeks and 4 months of age) was included in the models of insecurity, training focus, aggressiveness/dominance, human sociability, and dog sociability (Table 1, Figures 1F and 4A–4E). Higher level of socialization was associated with lower insecurity and aggressiveness/dominance and higher training focus, human sociability, and dog sociability. The largest difference between the poorest socialization level (Q1) and the highest socialization level (Q4) was observed for insecurity (z = 13.02, df = 1, p = 2.05e-36). Most pairwise comparisons between socialization levels reached significance in all traits, and the largest differences were always observed between the poorest socialization level and the highest socialization level (training focus: z = 9.40, df = 1, p = 2.43e-19; aggressiveness/dominance: z = 7.33, df = 1, p = 3.82e-12; human sociability: z = 6.42, df = 1, p = 1.56e-9; dog sociability: z = 5.39, df = 1, p = 5.26e-7). For all pairwise contrasts, see supplemental information.
Figure 4.

Association of puppyhood socialization and main reason for acquiring with personality traits
(A–H) Association of the level of socialization experiences between 7 weeks and 4 months of age with (A) insecurity, (B) training focus, (C) aggressiveness/dominance, (D) human sociability, (E) dog sociability, and association of main reason/use with (F) insecurity, (G) human sociability, (H) perseverance in the general and generalized linear models. Error bars indicate 95% confidence limits. N = 11 418.
Other environmental factors
The main reason for ownership/use of the dog was included in the final models of insecurity, perseverance, and human sociability (Table 1, Figures 4F–4H). The highest mean insecurity score was discovered in pet dogs and the lowest in dogs acquired for hobbies or work (z = 6.90, df = 1, p = 7.25e-11). Pet dogs were also more insecure than dogs considered family members (z = 2.66, df = 1, p = 0.018), which were, in turn, more insecure than hobby/working dogs (z = 6.16, df = 1, p = 7.71e-9). The highest mean perseverance score was obtained by hobby/working dogs and the next highest in dogs considered family members (difference: z = 7.95, df = 1, p = 3.94e-14). The lowest score was obtained by pet dogs (hobby/working dogs vs. pet dogs: z = 10.79, df = 1, p = 3.30e-25; family member dogs vs. pet dogs: z = 5.68, df = 1, p = 1.09e-7). Mean human sociability was higher in family member dogs than in pet dogs (z = 5.86, df = 1, p = 4.13e-8) and hobby/working dogs (z = 7.51, df = 1, p = 1.12e-12). There was no difference in human sociability between pet and hobby/working dogs (z = 1.12, df = 1, p = 0.358).
The frequency of hobbies was included as an explanatory variable for insecurity, training focus, and energy (Table 1, Figures S5A–S5C). The highest mean insecurity score was obtained by dogs that never participated in hobbies and the lowest by dogs that participated in hobbies infrequently (z = 5.64, df = 1, p = 1.40e-7). Mean training focus was highest in dogs participating in hobbies actively and lowest in dogs never participating in hobbies (z = 11.82, df = 1, p = 4.21e-30), and these groups obtained also the highest and lowest mean energy score (z = 8.68, df = 1, p = 1.24e-16). For all pairwise contrasts, see Supplementary Information.
Dog experience was included in the models of insecurity and training focus (Table 1, Figures S6A and S6B). The highest mean insecurity score was obtained by owners’ first dogs and the lowest by dogs that were owners’ fifth dogs or more (z = 12.55, df = 1, p = 7.25e-34). In contrast, the highest training focus score was observed in dogs that were owners’ fifth dogs or more and lowest in second dogs (z = 7.79, df = 1, p = 1.41e-13). For all pairwise contrasts, see Supplementary Information.
The variable “other dogs in the household when acquired” was included in the final models of energy and perseverance (Table 1, Figures S6C and S6D). Dogs that had dog company when they entered the household were more energetic (z = 6.73, df = 1, p = 2.25e-10) but less persevering (z = 8.32, df = 1, p = 2.36e-15) than dogs that were only dogs at that time.
Daily exercise and average time spent alone were included in the final model of training focus (Table 1, Figures S5D and S5E). Dogs exercising over 3 h per day on average had the highest mean training focus, whereas dogs exercising less than 1 h per day had the lowest mean score (z = 7.18, df = 1, p = 1.15e-11). Furthermore, dogs that spent, on average, less than 1 h alone during a typical workday obtained the highest mean training focus, whereas dogs spending over 8 h alone obtained the lowest score (z = 5.33, df = 1, p = 7.06e-7). For all pairwise contrasts, see Supplementary Information.
Discussion
Although many studies have examined the associations of demographic and environmental factors with canine behavior problems and undesirable behavior, less is known about factors influencing dog personality. We utilized our validated dog personality questionnaire35 to examine the association of environmental and demographic variables with seven personality traits: insecurity, training focus, energy, aggressiveness/dominance, human sociability, dog sociability, and perseverance. We discovered that dog breeds notably differed in all personality traits and that age was also highly associated with personality. Some environmental factors, such as the level of socialization in puppyhood, frequency of hobbies, and the main reason for dog ownership were also associated with personality traits. However, their effect sizes were small, indicating a limited association between environment and personality.
Breeds and breed groups differed in personality trait mean scores. Many breeds and breed groups we included have not been studied before, but our results were mostly in line with previous studies. Breed differences in training focus,36,42,43,45,49 insecurity,10,42,43,44,45,46,50 energy,36,42,44,45 aggressiveness/dominance,44,45,51,52 dog sociability,43 and human sociability42,44,45 highly paralleled previous studies. Furthermore, to our knowledge, breed differences in perseverance have not previously been reported. In a validation study of the Monash Canine Personality Questionnaire,36 breed differences in motivation (closely resembling our perseverance) were not detected. Breeds that reached the highest mean score in perseverance (Miniature Pinscher, followed by Asian primitive breeds, Parson-type terriers, Dachshunds, and fighting dogs) are often described as stubborn.53 Furthermore, a previous study discovered that perseverance was weakly associated with fear-aggression,54 and these persevering breeds also scored high in aggressiveness/dominance. In contrast, breeds that scored low in perseverance (including Golden Retriever, Shetland Sheepdog, and Smooth Collie) are all known to be adaptable and compliant.53
We compared the distributions of breeds whose mean scores most differed and showed that even though the distributions mostly overlapped, breeds showed differences in where the majority of individuals resided on the trait score distribution. Therefore, a dog’s breed is not a predictor of its personality, but the probability of showing certain personality traits differs between breeds. Breeds of other domesticated animals also differ in behavior and personality.55,56,57,58,59,60 In conclusion, breed differences in personality seem to be quite stable throughout studies and populations, but many uncommon breeds have not been adequately studied to reliably describe their typical personalities. These breed differences suggest that genetic differences could underlie these differences in behavior between breeds.
Other demographic variables, sex, age, and sterilization status were also associated with personality. Cross-sectional age differences were comparable to breed differences. In contrast, associations of sex and sterilization status were smaller. As discovered in previous studies, the highest mean energy level was found in young dogs, whereas age was nonlinearly related to aggressiveness/dominance, with the highest mean aggressiveness in 8–10-year-old dogs.10,36,46 Similarly to our study, one previous study discovered that dog sociability was highly negatively associated with age.39 We discovered that insecurity was negatively related to age, contrary to some previous studies.39,61 We also noted that training focus was positively associated with age. Some previous studies, including one longitudinal study, agree with our results,36,62 whereas some studies have discovered the opposite association.10,39 These age differences in dogs show parallels to humans and other nonhuman animals.6,7,31,32,63,64,65,66 This could mean that personality traits share a similar genetic and evolutionary background leading to similar development through life or that similar environmental factors shape personality in both humans and other animals. Female dogs were more focused on training and insecure than male dogs, whereas males were more aggressive/dominant, dog sociable, and energetic, agreeing with previous studies.39,44,46,61 Neutered dogs were more insecure, as found in some earlier studies42,61 and less focused on training than intact dogs. Neutering may cause unwanted changes in behavior or, on the other hand, more insecure and less trainable dogs may be more likely to be neutered. Similarly to age, these sex differences parallel those of humans and other nonhuman animals,7,59,67,68,69 possibly indicating a common effect of sex hormones on personality and behavior.
The level of socialization experiences in puppyhood was the most important environmental variable, based on visual inspection of variable level differences and relative and absolute effect sizes. Poorly socialized dogs had higher mean scores in insecurity and aggressiveness/dominance and lower scores in training focus, dog sociability, and human sociability than well-socialized dogs. Dogs obtained after the socialization period, at more than 4 months of age, were similar to poorly socialized dogs in all traits. Of these dogs, 44% were rescues or obtained from a previous owner. Therefore, the results indicate that dogs obtained as rescues or from a previous owner likely have more undesirable characteristics in their personalities, as has been discovered previously.46,70 Early life experiences seem to associate with fearful and aggressive behavior in dogs,50,71,72,73,74,75 but also trainability.75 Overall, early life experiences are important for subsequent personality, behavior, and mental health in dogs, humans, and other animals.26,28,76,77,78,79,80 However, it is also possible that owners willing to socialize their puppies choose the puppy and its breeder more carefully.
Dogs acquired for hobbies and dogs practicing actively were less insecure and more focused on training, energetic, and persevering, as was discovered in earlier studies.10,39,46,50,72,81,82,83 Boldness, training focus, high energy level, and, to some extent, perseverance are important characteristics for hobby dogs that likely make them easier to train and more successful in competitions. Therefore, owners who actively participate in hobbies may select puppies with a certain personality and stop participating with the dog if it proves unsuitable for the hobby. However, it is also possible that participating in hobbies could improve behavioral characteristics.84 Hobbies likely provide environmental enrichment, which has been shown to influence behavior in dogs and other animals as well.84,85,86
Surprisingly, we discovered differences between dogs regarded as pets or family members: pet dogs scored higher in insecurity and lower in human sociability and perseverance than dogs considered family members. Similarly, one previous study found that non-family member dogs were less dog sociable than family member dogs.39 This phenomenon may be related to attachment to the dog, as previous studies have indicated that owners of well-behaving dogs are more satisfied with and attached to them9,10,11 and owners perceiving their dogs as pets had a higher probability of unsuccessful ownership.87 It is also possible that attitude toward dogs, in general, influences the selection of the dog, the owner's perception of the dog’s behavior or their care of the dog.
Other environmental variables, dog experience, living in a multi-dog household when acquired, daily exercise, and average time spent alone during a typical workday showed significant but small associations with personality traits. These parallel previous studies (dog experience,72,83 rearing with conspecifics,88,89 daily exercise, and time spent alone83). However, the reason for these small associations is not known. Due to the parallels of these variables with previous studies and their small importance, we will not speculate on these associations further.
Breed/breed group and age were the most important variables, both relatively and absolutely. Despite their importance, Cohen’s f2 values were small, agreeing with a recent study of behavioral differences between breeds.47 However, effect size estimates in individual differences are commonly small and should only be compared with other studies, not with arbitrary cut-offs.90 Indeed, studies have suggested that correlation cut-offs should be lowered to small = 0.10, medium = 0.20, and large = 0.30.90,91 These cut-points would correspond to Cohen’s f2 estimates of 0.02, 0.05, and 0.10, respectively.48 If following these suggested estimates, the breed would have a large effect size in energy and a medium effect size in aggressiveness/dominance, human sociability, dog sociability, and perseverance. The effect size of the breed would remain small in insecurity and training focus, possibly highlighting a more complex background in these traits. As personality traits are strikingly complex, it is not expected that any variables would explain a large portion of the variance in these traits. Therefore, the variables included in the models, especially breed has an important association with personality traits, and therefore, despite obtaining highly similar results to Morrill et al.,47 we argue that dog breeds do differ.
In conclusion, we discovered many interesting associations between demographic and environmental variables and seven personality traits. The associations of breed and age of the dog with personality traits were more extensive than environmental factors. This was hardly surprising, as the environmental variables mostly considered the current environment of largely adult individuals. This was also paralleled in effect size estimates, which were much higher for breed and age than for environmental variables. Significant breed differences in personality traits indicate that personality is influenced by the individual’s genetic background. They also suggest that different behavioral characteristics are preferred and selected for in different dog breeds. Overall, our results show that personality traits are complex and are likely influenced by the genetic background of the individual and life experiences. This is reflected in the small effect sizes of all explanatory variables. Humans as breeders have a large impact on the personality and well-being of dogs, but our results indicate that individual owners can also influence the development of personality by socializing puppies carefully. Finally, our results show that associations of age, sex, and environment with dog personality resemble those in humans and other nonhuman animals. This similarity indicates that either patterns of personality development are evolutionary very old or due to parallel evolution in dogs and humans. Thus, our findings improve the wider understanding of factors that influence animal and human personalities.
Limitations of the study
Our study has limitations. First, this study is cross-sectional, and therefore causal relationships between explanatory variables and personality traits cannot be inferred. Second, the data is a convenience sample collected online and young, enthusiastic dog owners may be more likely to answer. Third, it is possible that owners of, for example, different breeds perceive their dogs differently. However, the survey used in the study was shown to have good validity and reliability.35 Fourth, to preserve the large sample size and variability in the background of dogs, we created an “obtained after the socialization period” socialization category instead of performing listwise deletion, which could create additional bias. Fifth, we could not collect many variables that would likely be important for the development of personality, including, for example, maternal stress during gestation, quality of maternal care, and socialization experiences before weaning. Future longitudinal studies would be needed to examine the effect of environment on personality.
STAR★Methods
Key resources table
| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Deposited data | ||
| Data used in this study | Figshare | https://doi.org/10.6084/m9.figshare.14479152.v1 |
| Software and algorithms | ||
| Code used in this study | Supplementary material | Data S1 |
| R 4.2.1. | R Core Team | https://cran.r-project.org/ |
Resource availability
Lead contact
Further information and requests for resources should be directed to and will be fulfilled by the lead contact, Hannes Lohi (hannes.lohi@helsinki.fi).
Materials availability
This study did not generate new unique materials.
Experimental model and subject details
Dog owners participated in the study by answering an extensive online behavior questionnaire of their dogs. We did not collect the dog owners’ age and gender identity. Range, mean, and standard deviation of dogs’ age can be found in Table S11 and sex distribution in Table S12. Dog owners provided the information of their dog’s sex, age, breed, environment, and personality. The study was approved by the University of Helsinki Viikki Campus Research Ethics Committee (11.2.2019). Informed consent was obtained from all participants.
Method details
Questionnaire
We utilized our dog personality and unwanted behavior questionnaire, which was previously validated with hypothesis testing. The internal consistency, test-retest reliability, and inter-rater reliability of this questionnaire were also assessed.35 The questionnaire included a personality section, several unwanted behavior sections, a health section, and a background section. A detailed description of the questionnaire and its sections is available as Supplementary Material of our previous study.35
The personality section of the questionnaire included 63 adjective-based statements and the owners indicated how strongly they agreed or disagreed with the statements using a 5-point Likert scale. We previously used factor analysis to reduce these statements into 7 personality traits (factors): insecurity, energy, training focus, aggressiveness/dominance, human sociability, dog sociability, and perseverance.35 We calculated factor scores for each individual and used these as explanatory variables.
Subjects
We previously collected a dataset that included questionnaire responses on 15 371 dogs.35 From this sample, we excluded dogs whose owners did not answer the personality section (2506 dogs). During quality control we also noted one dog whose answers were clearly false (same answering option selected for every question) and excluded it. Furthermore, we also excluded dogs whose owners did not answer the background section (1445 dogs). Finally, we excluded one dog that was a considerable outlier in three out of seven analyses.
The final dataset included 11 418 dogs in 315 breeds and breed variants. Many breeds had only a few responses and thus, we grouped these breeds based on their genetic relationships,92 purpose and use of the breed, and known behavioral similarities. Breeds that were not studied by Parker and others92 were grouped based on purpose and behavioral similarities. For example, our study included more scenthound breeds than Parker et al.,92 which we included in the “scenthounds” breed group. Breeds with an adequate number of responses were handled individually. This grouping resulted in 32 breed groups, 19 individual breeds, and mixed breed dogs (Table S1).
Variables
We included the scores of the seven personality traits as response variables. For the possible explanatory variables, we included the dog’s age, sex, and breed, as well as 12 variables related to the environment: sterilization status, main reason for ownership/use of the dog, where was the dog obtained from, level of socialization, living place, other dogs in the household when the particular dog was acquired, other dogs in the household currently, daily exercise, frequency of hobbies, average time spent alone, life experience of dogs, and dog experience. Level of socialization and frequency of hobbies were variables combined from multiple questions and they are thus described in detail below. All variables are described in Table S2.
Socialization included 7 questions related to experiences between 7 weeks and 4 months of age. These socialization questions were only answered by owners who obtained their dogs before 4 months of age. In these questions, owners stated how often their dog met unfamiliar men, women, children, and dogs, visited city center, and traveled by car and by public transport. The response options were: never, rarely (1–4 times during puppyhood), sometimes (twice a month-twice a week), often (twice a week-once a day), and very often (several times a day). We used principal components analysis (PCA) to reduce these frequencies into fewer components. We first assessed the sampling adequacy with the Kaiser-Meyer-Olkin test using the package psych.93 Then we performed the PCA using a polychoric correlation matrix and without rotation. The screen test and Velicer’s minimum average partial test both suggested one component (Table S3) and we extracted the component scores for individuals with the “Thurstone” estimation method. Finally, many dogs came to the owner after 4 months of age and their owners did not, therefore, answer this question. Thus, we turned this continuous variable into categorical by dividing these factor scores into quartiles and including a fifth category labeled “Obtained after the socialization period”.
The dog owners also indicated how often they engaged in activities other than walking (dog hobbies) with the dog both at home and outside of the home, for example, with an instructor. In frequency outside of home, the options were “never”, “a few times a year”, “1–3 times a month”, “1–2 times a week”, “3–4 times a week”, and “almost every day”. The question asking about the frequency at home had these same options and also included “several times a day” as an option. We divided these answers into three levels: active, infrequent, and never/no hobby. Active included frequency of hobbies of 1–2 times a week or higher. Infrequent included hobby frequencies a few times a year and 1–3 times a month. Never/no hobby included the answering option never. Furthermore, as these variables correlated with each other, we combined them to a single “frequency of hobbies” variable. Dogs that belonged to the category “active” in both variables were placed in the “active” category. Dogs that belonged to the category active only at home were placed in an “active only at home” category. Dogs that participated actively in hobbies only outside of the home were included in an “active only outside” category. Dogs that belonged to the “infrequent” category in either variable were placed in an “infrequent” category in frequency of hobbies. Finally, dogs that belonged to “never/no hobby” category in both variables were placed to the category “never/no hobby” in frequency of hobbies (Table S2).
We evaluated interactions between sex and sterilization, daily exercise and frequency of hobbies, dog’s age and sex, living place and daily exercise, life experience of dogs and dog experience, main reason/use of the dog and frequency of hobbies, other dogs currently and average time spent alone, and between other dogs currently and other dogs when the dog was acquired.
Quantification and statistical analysis
We fitted general and generalized linear models to examine the association of explanatory variables with personality traits, which were used as response variables. Before selecting the best explanatory variables, we fitted general linear models with age, sex, and breed as explanatory variables and investigated the fit of the normal distribution by plotting residuals with packages rcompanion94 and boot95 in R. If normal distribution provided poor fit, we fitted a generalized linear model. The traits insecurity and aggressiveness/dominance had a better model fit with the gamma distribution.
We used an in-house model selection loop in R96 and performed forward stepwise AIC model selection to select explanatory variables. Dog’s age, sex, and breed were included in the starting model in all traits. Besides the possible explanatory variables described above, we also included possible interactions between variables, such as between age and sex, and between sex and sterilization status. These interactions are listed in Tables S4–S10.
AIC model selection tends to overfit models. Thus, we performed extensive model validation to only include relevant explanatory variables in the models. First we performed repeated train-test validation by randomly splitting the data into train (50% of data) and test (50% of data) datasets five times with the package caret97 in R. Thus, this resulted in ten datasets with 50% of the data. We used the aforementioned model selection procedure for all of these datasets. Next, we performed random subsampling validation by splitting the data randomly into 10 equal-sized datasets (10% of data) and performing model selection for these datasets as well. Then we fitted boosted regression models to evaluate the relative importance of variables with the package gbm98 in R.
After the AIC model selection of the validation datasets, we had 20 models for each response variable in total: 10 models in repeated train-test validation datasets and 10 models in random subsampling validation datasets. We calculated the number of times each explanatory variable was included in the chosen models for both validation types. In the final model for each response variable, we decided to include the explanatory variables which were 1) included in all (100%) of the repeated train-test validation dataset models and 2) included in at least 50% of the random subsampling models. The results of model validation are found in Tables S4–S10.
Model fit and extraction of results
We fitted general and generalized linear models using the selected explanatory variables (described above). We then assessed model fit carefully. Firstly, we again plotted the residuals with packages rcompanion94 and boot95 to ensure a good choice of distribution (gamma for insecurity and aggressiveness/dominance, normal distribution for other traits) and to evaluate the presence of heteroscedasticity. Secondly, we inspected outliers with packages broom,99 dplyr,100 and ggplot2.101 At this stage, we detected one outlier, excluded it (as explained in the section “Subjects”) and refitted the models. Thirdly, we checked the linearity assumption of the continuous variable age by fitting generalized additive models with the package gam102 and if this assumption was not met, included both linear and quadratic variables (age and age2) in the general/generalized linear model. Finally, we examined multicollinearity with the variance inflation factor using the package car.103
After examining the fit of the selected models, we extracted the results. Firstly, we conducted analysis of variance (ANOVA) with the package car103 to obtain the overall effects of the explanatory variables. Secondly, to obtain the adjusted means, confidence limits and pairwise comparisons of the levels of categorical explanatory variables, we extracted the estimated marginal means using the package emmeans.104 Finally, we calculated the adjusted means and confidence limits of the continuous explanatory variable age with the package effects.103,105 Due to many pairwise comparisons, all p values were controlled for the false discovery rate (FDR). We set the significance cut-off at p < 0.05.
To compare trait scores between the most extreme breeds, we plotted the trait score distributions of breeds with lowest and highest mean scores in each trait using package ggplot2.101
Effect size
We examined the relative effect size of variables by computing variance-based variable importance using a feature importance ranking measure approach with the package vip.106 Variable importance was scaled so that the most important variable was given the score of 100.
We also examined absolute effect sizes by calculating partial Cohen’s f2 with the package effectsize.107 We followed Cohen’s suggestions48 and defined cut-offs as small = 0.02, medium = 0.15, and large = 0.35.
We conducted all analyses in R version 4.1.2.108
Acknowledgments
We are grateful to the dog owners who participated in the study. We thank Petsofi Ltd. for developing and providing the survey platform. H.L. is a member of HiLIFE. M.S., S.M., E.H., J. E.N., and H.L. are members of Helsinki One Health. This research was funded by the Academy of Finland (grant no. 308887) and the Jane and Aatos Erkko Foundation.
Author contributions
Conceptualization: M.S., S.M., and H.L.; software: J.E.N.; validation: M.S.; formal analysis: M.S.; investigation: M.S., S.M., E.H., S.S., and J.P.;, data curation: M.S., S.M., J.E.N., E.H., and S.S.; writing - original draft: M.S.; writing - review & editing: all authors; visualization: M.S.; supervision: H.L.; project administration: H.L.; funding acquisition: H.L.
Declaration of interests
H.L. was a co-founder and E.H. an employee of Petsofi Ltd., which provided the survey platform for collecting data. The authors declare no other competing interests.
Published: April 19, 2023
Footnotes
Supplemental information can be found online at https://doi.org/10.1016/j.isci.2023.106691.
Contributor Information
Milla Salonen, Email: milla.salonen@utu.fi.
Hannes Lohi, Email: hannes.lohi@helsinki.fi.
Supplemental information
Data and code availability
-
•
Questionnaire data have been deposited at Figshare and are publicly available as of the date of publication. DOI is listed in the key resources table.
-
•
All original code is available in this paper’s supplemental information.
-
•
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.
References
- 1.Carere C., Maestripieri D. In: Animal Personalities: Behavior, Physiology, and Evolution. Carere C., Maestripieri D., editors. University of Chicago Press; 2013. Animal personalities: who cares and why? pp. 1–10. [Google Scholar]
- 2.Stamps J., Groothuis T.G.G. The development of animal personality: relevance, concepts and perspectives. Biol. Rev. 2010;85:301–325. doi: 10.1111/j.1469-185X.2009.00103.x. [DOI] [PubMed] [Google Scholar]
- 3.Strickhouser J.E., Zell E., Krizan Z. Does personality predict health and well-being? A metasynthesis. Health Psychol. 2017;36:797–810. doi: 10.1037/hea0000475. [DOI] [PubMed] [Google Scholar]
- 4.Anglim J., Horwood S., Smillie L.D., Marrero R.J., Wood J.K. Predicting psychological and subjective well-being from personality: a meta-analysis. Psychol. Bull. 2020;146:279–323. doi: 10.1037/BUL0000226. [DOI] [PubMed] [Google Scholar]
- 5.Kotov R., Gamez W., Schmidt F., Watson D. Linking “Big” personality traits to anxiety, depressive, and substance use disorders: a meta-analysis. Psychol. Bull. 2010;136:768–821. doi: 10.1037/a0020327. [DOI] [PubMed] [Google Scholar]
- 6.Weiss A., Inoue-Murayama M., Hong K.W., Inoue E., Udono T., Ochiai T., Matsuzawa T., Hirata S., King J.E. Assessing chimpanzee personality and subjective well-being in Japan. Am. J. Primatol. 2009;71:283–292. doi: 10.1002/ajp.20649. [DOI] [PubMed] [Google Scholar]
- 7.Altschul D.M., Hopkins W.D., Herrelko E.S., Inoue-Murayama M., Matsuzawa T., King J.E., Ross S.R., Weiss A. Personality links with lifespan in chimpanzees. Elife. 2018;7:e33781. doi: 10.7554/ELIFE.33781. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Smith B.R., Blumstein D.T. Fitness consequences of personality: a meta-analysis. Behav. Ecol. 2008;19:448–455. doi: 10.1093/beheco/arm144. [DOI] [Google Scholar]
- 9.Cavanaugh L.A., Leonard H.A., Scammon D.L. A tail of two personalities: how canine companions shape relationships and well-being. J. Bus. Res. 2008;61:469–479. doi: 10.1016/j.jbusres.2007.07.024. [DOI] [Google Scholar]
- 10.Chopik W.J., Weaver J.R. Old dog, new tricks: age differences in dog personality traits, associations with human personality traits, and links to important outcomes. J. Res. Pers. 2019;79:94–108. doi: 10.1016/j.jrp.2019.01.005. [DOI] [Google Scholar]
- 11.Herwijnen I.R.v., van der Borg J.A.M., Naguib M., Beerda B. Dog ownership satisfaction determinants in the owner-dog relationship and the dog’s behaviour. PLoS One. 2018;13:e0204592. doi: 10.1371/journal.pone.0204592. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kwan J.Y., Bain M.J. Owner attachment and problem behaviors related to relinquishment and training techniques of dogs. J. Appl. Anim. Welfare Sci. 2013;16:168–183. doi: 10.1080/10888705.2013.768923. [DOI] [PubMed] [Google Scholar]
- 13.O’Neill D.G., Church D.B., McGreevy P.D., Thomson P.C., Brodbelt D.C. Longevity and mortality of owned dogs in England. Vet. J. 2013;198:638–643. doi: 10.1016/J.TVJL.2013.09.020. [DOI] [PubMed] [Google Scholar]
- 14.New J.C., Salman M.D., King M., Scarlett J.M., Kass P.H., Hutchison J.M. Characteristics of shelter-relinquished animals and their owners compared with animals and their owners in U.S. pet-owning households. J. Appl. Anim. Welfare Sci. 2000;3:179–201. doi: 10.1207/S15327604JAWS0303_1. [DOI] [Google Scholar]
- 15.Gosling S.D. Personality in non-human animals. Soc. Personal. Psychol. Compass. 2008;2:985–1001. doi: 10.1111/j.1751-9004.2008.00087.x. [DOI] [Google Scholar]
- 16.Gosling S.D. From mice to men: what can we learn about personality from animal research? Psychol. Bull. 2001;127:45–86. doi: 10.1037/0033-2909.127.1.45. [DOI] [PubMed] [Google Scholar]
- 17.Mather J.A., Logue D.M. In: Animal Personalities: Behavior, Physiology, and Evolution. Carere C., Maestripieri D., editors. University of Chicago Press; 2013. The bold and the spineless: invertebrate personalities; pp. 12–35. [DOI] [Google Scholar]
- 18.Polderman T.J.C., Benyamin B., De Leeuw C.A., Sullivan P.F., Van Bochoven A., Visscher P.M., Posthuma D. Meta-analysis of the heritability of human traits based on fifty years of twin studies. Nat. Genet. 2015;47:702–709. doi: 10.1038/ng.3285. [DOI] [PubMed] [Google Scholar]
- 19.Røysamb E., Nes R.B., Czajkowski N.O., Vassend O. Genetics, personality and wellbeing. A twin study of traits, facets and life satisfaction. Sci. Rep. 2018;8:12298. doi: 10.1038/s41598-018-29881-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dochtermann N.A., Schwab T., Sih A. The contribution of additive genetic variation to personality variation: heritability of personality. Proc. Biol. Sci. 2015;282:20142201. doi: 10.1098/rspb.2014.2201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Vukasović T., Bratko D. Heritability of personality: a meta-analysis of behavior genetic studies. Psychol. Bull. 2015;141:769–785. doi: 10.1037/bul0000017. [DOI] [PubMed] [Google Scholar]
- 22.Bleidorn W., Hopwood C.J., Lucas R.E. Life events and personality trait change. J. Pers. 2018;86:83–96. doi: 10.1111/jopy.12286. [DOI] [PubMed] [Google Scholar]
- 23.Kandler C., Waaktaar T., Mõttus R., Riemann R., Torgersen S. Unravelling the interplay between genetic and environmental contributions in the unfolding of personality differences from early adolescence to young adulthood. Eur. J. Pers. 2019;33:221–244. doi: 10.1002/PER.2189. [DOI] [Google Scholar]
- 24.Bouchard T.J., Loehlin J.C. Genes, evolution, and personality. Behav. Genet. 2001;31:243–273. doi: 10.1023/A:1012294324713. [DOI] [PubMed] [Google Scholar]
- 25.Shelton D.S., Martins E.P. In: APA handbook of comparative psychology: Basic concepts, methods, neural substrate, and behavior. Call J., Burghardt G.M., Pepperberg I.M., Snowdon C.T., Zentall T., editors. American Psychological Association; 2017. Behavioral variation, adaptation, and evolution; pp. 219–236. [DOI] [Google Scholar]
- 26.Loman M.M., Gunnar M.R., Early Experience, Stress, and Neurobehavioral Development Center Early experience and the development of stress reactivity and regulation in children. Neurosci. Biobehav. Rev. 2010;34:867–876. doi: 10.1016/j.neubiorev.2009.05.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shonkoff J.P., Garner A.S., Committee on Psychosocial Aspects of Child and Family Health. Committee on Early Childhood Adoption and Dependent Care. Section on Developmental and Behavioral Pediatrics. McGuinn L., Pascoe J., Wood D.L., High P.C., Donoghue E., et al. The lifelong effects of early childhood adversity and toxic stress. Pediatrics. 2012;129:e232–e246. doi: 10.1542/peds.2011-2663. [DOI] [PubMed] [Google Scholar]
- 28.Maestripieri D., Carroll K.A. Child abuse and neglect: usefulness of the animal data. Psychol. Bull. 1998;123:211–223. doi: 10.1037/0033-2909.123.3.211. [DOI] [PubMed] [Google Scholar]
- 29.Kikusui T., Mori Y. Behavioural and neurochemical consequences of early weaning in rodents. J. Neuroendocrinol. 2009;21:427–431. doi: 10.1111/j.1365-2826.2009.01837.x. [DOI] [PubMed] [Google Scholar]
- 30.Roberts B.W., Mroczek D. Personality trait change in adulthood. Curr. Dir. Psychol. Sci. 2008;17:31–35. doi: 10.1111/j.1467-8721.2008.00543.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bleidorn W., Schwaba T., Zheng A., Hopwood C.J., Sosa S.S., Roberts B.W., Briley D.A. Personality stability and change: a meta-analysis of longitudinal studies. Psychol. Bull. 2022;148:588–619. doi: 10.1037/BUL0000365. [DOI] [PubMed] [Google Scholar]
- 32.Roberts B.W., Walton K.E., Viechtbauer W. Patterns of mean-level change in personality traits across the life course: a meta-analysis of longitudinal studies. Psychol. Bull. 2006;132:1–25. doi: 10.1037/0033-2909.132.1.1. [DOI] [PubMed] [Google Scholar]
- 33.Jones A.C. 2008. Development and Validation of a Dog Personality Questionnaire. PhD Diss. [Google Scholar]
- 34.Jones A.C., Gosling S.D. Temperament and personality in dogs (Canis familiaris): a review and evaluation of past research. Appl. Anim. Behav. Sci. 2005;95:1–53. doi: 10.1016/j.applanim.2005.04.008. [DOI] [Google Scholar]
- 35.Salonen M., Mikkola S., Hakanen E., Sulkama S., Puurunen J., Lohi H. Reliability and validity of a dog personality and unwanted behavior survey. Animals. 2021;11:1234. doi: 10.3390/ani11051234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Ley J.M., Bennett P.C., Coleman G.J. A refinement and validation of the Monash canine personality questionnaire (MCPQ) Appl. Anim. Behav. Sci. 2009;116:220–227. doi: 10.1016/j.applanim.2008.09.009. [DOI] [Google Scholar]
- 37.Mirkó E., Kubinyi E., Gácsi M., Miklósi Á. Preliminary analysis of an adjective-based dog personality questionnaire developed to measure some aspects of personality in the domestic dog (Canis familiaris) Appl. Anim. Behav. Sci. 2012;138:88–98. doi: 10.1016/j.applanim.2012.02.016. [DOI] [Google Scholar]
- 38.Ley J., Bennett P., Coleman G. Personality dimensions that emerge in companion canines. Appl. Anim. Behav. Sci. 2008;110:305–317. doi: 10.1016/j.applanim.2007.04.016. [DOI] [Google Scholar]
- 39.Kubinyi E., Turcsán B., Miklósi A. Dog and owner demographic characteristics and dog personality trait associations. Behav. Process. 2009;81:392–401. doi: 10.1016/j.beproc.2009.04.004. [DOI] [PubMed] [Google Scholar]
- 40.Turcsán B., Wallis L., Virányi Z., Range F., Müller C.A., Huber L., Riemer S. Personality traits in companion dogs—results from the VIDOPET. PLoS One. 2018;13:e0195448. doi: 10.1371/journal.pone.0195448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Svartberg K., Forkman B. Personality traits in the domestic dog (Canis familiaris) Appl. Anim. Behav. Sci. 2002;79:133–155. doi: 10.1016/S0168-1591(02)00121-1. [DOI] [Google Scholar]
- 42.Tonoike A., Nagasawa M., Mogi K., Serpell J.A., Ohtsuki H., Kikusui T. Comparison of owner-reported behavioral characteristics among genetically clustered breeds of dog (Canis familiaris) Sci. Rep. 2015;5:17710. doi: 10.1038/srep17710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Turcsán B., Kubinyi E., Miklósi Á. Trainability and boldness traits differ between dog breed clusters based on conventional breed categories and genetic relatedness. Appl. Anim. Behav. Sci. 2011;132:61–70. doi: 10.1016/J.APPLANIM.2011.03.006. [DOI] [Google Scholar]
- 44.Eken Asp H., Fikse W.F., Nilsson K., Strandberg E. Breed differences in everyday behaviour of dogs. Appl. Anim. Behav. Sci. 2015;169:69–77. doi: 10.1016/j.applanim.2015.04.010. [DOI] [Google Scholar]
- 45.Serpell J.A., Duffy D.L. In: Domestic Dog Cognition and Behavior. Horowitz A., editor. Springer; 2014. Dog breeds and their behavior; pp. 31–57. [DOI] [Google Scholar]
- 46.Wallis L.J., Szabó D., Kubinyi E. Cross-sectional age differences in canine personality traits; influence of breed, sex, previous trauma, and dog obedience tasks. Front. Vet. Sci. 2019;6:493. doi: 10.3389/fvets.2019.00493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Morrill K., Hekman J., Li X., McClure J., Logan B., Goodman L., Gao M., Dong Y., Alonso M., Carmichael E., et al. Ancestry-inclusive dog genomics challenges popular breed stereotypes. Science. 2022;376:eabk0639. doi: 10.1126/SCIENCE.ABK0639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cohen J. 2nd edition. Lawrence Erlbaum Associates; 1988. Statistical Power Analysis for the Behavioral Sciences. [DOI] [Google Scholar]
- 49.Serpell J.A., Hsu Y.A. Effects of breed, sex, and neuter status on trainability in dogs. Anthrozoös. 2005;18:196–207. doi: 10.2752/089279305785594135. [DOI] [Google Scholar]
- 50.Puurunen J., Hakanen E., Salonen M.K., Mikkola S., Sulkama S., Araujo C., Lohi H. Inadequate socialisation, inactivity, and urban living environment are associated with social fearfulness in pet dogs. Sci. Rep. 2020;10:3527. doi: 10.1038/s41598-020-60546-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Duffy D.L., Hsu Y., Serpell J.A. Breed differences in canine aggression. Appl. Anim. Behav. Sci. 2008;114:441–460. doi: 10.1016/j.applanim.2008.04.006. [DOI] [Google Scholar]
- 52.Hsu Y., Sun L. Factors associated with aggressive responses in pet dogs. Appl. Anim. Behav. Sci. 2010;123:108–123. doi: 10.1016/j.applanim.2010.01.013. [DOI] [Google Scholar]
- 53.American Kennel Club. Dog Breeds. American Kennel Club 2022. https://www.akc.org/dog-breeds/.
- 54.Salonen M., Mikkola S., Hakanen E., Sulkama S., Puurunen J., Lohi H. Personality traits associate with behavioral problems in pet dogs. Transl. Psychiatry. 2022;12:78. doi: 10.1038/s41398-022-01841-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Salonen M., Vapalahti K., Tiira K., Mäki-Tanila A., Lohi H. Breed differences of heritable behaviour traits in cats. Sci. Rep. 2019;9:7949. doi: 10.1038/s41598-019-44324-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Mikkola S., Salonen M., Hakanen E., Sulkama S., Lohi H. Reliability and validity of seven feline behavior and personality traits. Animals. 2021;11:1991. doi: 10.3390/ANI11071991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Sackman J.E., Houpt K.A. Equine personality: association with breed, use, and husbandry factors. J. Equine Vet. Sci. 2019;72:47–55. doi: 10.1016/J.JEVS.2018.10.018. [DOI] [PubMed] [Google Scholar]
- 58.Lloyd A.S., Martin J.E., Bornett-Gauci H.L.I., Wilkinson R.G. Horse personality: variation between breeds. Appl. Anim. Behav. Sci. 2008;112:369–383. doi: 10.1016/J.APPLANIM.2007.08.010. [DOI] [Google Scholar]
- 59.Takeuchi Y., Mori Y. Behavioral profiles of feline breeds in Japan. J. Vet. Med. Sci. 2009;71:1053–1057. doi: 10.1292/jvms.71.1053. [DOI] [PubMed] [Google Scholar]
- 60.Wilhelmy J., Serpell J., Brown D., Siracusa C. Behavioral associations with breed, coat type, and eye color in single-breed cats. J. Vet. Behav. 2016;13:80–87. doi: 10.1016/J.JVEB.2016.03.009. [DOI] [Google Scholar]
- 61.Starling M.J., Branson N., Thomson P.C., McGreevy P.D. “Boldness” in the domestic dog differs among breeds and breed groups. Behav. Process. 2013;97:53–62. doi: 10.1016/j.beproc.2013.04.008. [DOI] [PubMed] [Google Scholar]
- 62.Riemer S., Müller C., Virányi Z., Huber L., Range F. Individual and group level trajectories of behavioural development in Border collies. Appl. Anim. Behav. Sci. 2016;180:78–86. doi: 10.1016/J.APPLANIM.2016.04.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Zablocki-Thomas P.B., Herrel A., Hardy I., Rabardel L., Perret M., Aujard F., Pouydebat E. Personality and performance are affected by age and early life parameters in a small primate. Ecol. Evol. 2018;8:4598–4605. doi: 10.1002/ECE3.3833. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bennett P.C., Rutter N.J., Woodhead J.K., Howell T.J. Assessment of domestic cat personality, as perceived by 416 owners, suggests six dimensions. Behav. Process. 2017;141:273–283. doi: 10.1016/j.beproc.2017.02.020. [DOI] [PubMed] [Google Scholar]
- 65.Gartner M.C., Powell D.M., Weiss A. Personality structure in the domestic cat (Felis silvestris catus), Scottish wildcat (Felis silvestris grampia), clouded leopard (Neofelis nebulosa), snow leopard (Panthera uncia), and African lion (Panthera leo): a comparative study. J. Comp. Psychol. 2014;128:414–426. doi: 10.1037/a0037104. [DOI] [PubMed] [Google Scholar]
- 66.Yamada R., Kuze-Arata S., Kiyokawa Y., Takeuchi Y. Prevalence of 17 feline behavioral problems and relevant factors of each behavior in Japan. J. Vet. Med. Sci. 2020;82:272–278. doi: 10.1292/jvms.19-0519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Hart B.L., Hart L.A. Purdue University Press; 2013. Your Ideal Cat: Insights into Breed and Gender Differences in Cat Behavior. [Google Scholar]
- 68.Allik J., Realo A. In: The Oxford Handbook of the Five Factor Model. Widiger T.A., editor. Oxford University Press; 2017. Universal and specific in the five factor model of personality; pp. 173–190. [DOI] [Google Scholar]
- 69.Lippa R.A. Gender differences in personality and interests: when, where, and why? Soc. Personal. Psychol. Compass. 2010;4:1098–1110. doi: 10.1111/J.1751-9004.2010.00320.X. [DOI] [Google Scholar]
- 70.Lord L.K., Reider L., Herron M.E., Graszak K. Health and behavior problems in dogs and cats one week and one month after adoption from animal shelters. J. Am. Vet. Med. Assoc. 2008;233:1715–1722. doi: 10.2460/JAVMA.233.11.1715. [DOI] [PubMed] [Google Scholar]
- 71.Howell T.J., King T., Bennett P.C. Puppy parties and beyond: the role of early age socialization practices on adult dog behavior. Vet. Med. 2015;6:143–153. doi: 10.2147/vmrr.s62081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Hakanen E., Mikkola S., Salonen M., Puurunen J., Sulkama S., Araujo C., Lohi H. Active and social life associates with lower non-social fear in pet dogs. Sci. Rep. 2020;10:13774. doi: 10.1038/s41598-020-70722-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Tiira K., Lohi H. Early life experiences and exercise associate with canine anxieties. PLoS One. 2015;10:e0141907. doi: 10.1371/journal.pone.0141907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Dietz L., Arnold A.-M.K., Goerlich-Jansson V.C., Vinke C.M. The importance of early life experiences for the development of behavioural disorders in domestic dogs. Beyond Behav. 2018;155:83–114. doi: 10.1163/1568539X-00003486. [DOI] [Google Scholar]
- 75.González-Martínez Á., Martínez M.F., Rosado B., Luño I., Santamarina G., Suárez M.L., Camino F., de la Cruz L.F., Diéguez F.J. Association between puppy classes and adulthood behavior of the dog. J. Vet. Behav. 2019;32:36–41. doi: 10.1016/J.JVEB.2019.04.011. [DOI] [Google Scholar]
- 76.Freedman D.G., King J.A., Elliot O. Critical period in the social development of dogs. Science. 1961;133:1016–1017. doi: 10.1126/science.133.3457.1016. [DOI] [PubMed] [Google Scholar]
- 77.Latham N.R., Mason G.J. Maternal deprivation and the development of stereotypic behaviour. Appl. Anim. Behav. Sci. 2008;110:84–108. doi: 10.1016/j.applanim.2007.03.026. [DOI] [Google Scholar]
- 78.Kikusui T., Takeuchi Y., Mori Y. Early weaning induces anxiety and aggression in adult mice. Physiol. Behav. 2004;81:37–42. doi: 10.1016/j.physbeh.2003.12.016. [DOI] [PubMed] [Google Scholar]
- 79.Würbel H., Stauffacher M. Physical condition at weaning affects exploratory behaviour and stereotypy development in laboratory mice. Behav. Process. 1998;43:61–69. doi: 10.1016/S0376-6357(97)00086-7. [DOI] [PubMed] [Google Scholar]
- 80.Lähdepuro A., Savolainen K., Lahti-Pulkkinen M., Eriksson J.G., Lahti J., Tuovinen S., Kajantie E., Pesonen A.-K., Heinonen K., Räikkönen K. The impact of early life stress on anxiety symptoms in late adulthood. Sci. Rep. 2019;9:4395. doi: 10.1038/s41598-019-40698-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lit L., Schweitzer J.B., Iosif A.-M., Oberbauer A.M. Owner reports of attention, activity, and impulsivity in dogs: a replication study. Behav. Brain Funct. 2010;6:1. doi: 10.1186/1744-9081-6-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Vas J., Topál J., Péch É., Miklósi Á. Measuring attention deficit and activity in dogs: a new application and validation of a human ADHD questionnaire. Appl. Anim. Behav. Sci. 2007;103:105–117. doi: 10.1016/j.applanim.2006.03.017. [DOI] [Google Scholar]
- 83.Sulkama S., Puurunen J., Salonen M., Mikkola S., Hakanen E., Araujo C., Lohi H. Canine hyperactivity, impulsivity, and inattention share similar demographic risk factors and behavioural comorbidities with human ADHD. Transl. Psychiatry. 2021;11:501. doi: 10.1038/s41398-021-01626-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Wells D.L. A review of environmental enrichment for kennelled dogs, Canis familiaris. Appl. Anim. Behav. Sci. 2004;85:307–317. doi: 10.1016/J.APPLANIM.2003.11.005. [DOI] [Google Scholar]
- 85.Gottlieb D.H., Capitanio J.P., Mccowan B. Risk factors for stereotypic behavior and self-biting in rhesus macaques (Macaca mulatta): animal’s history, current environment, and personality. Am. J. Primatol. 2013;75:995–1008. doi: 10.1002/ajp.22161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Vandeleest J.J., McCowan B., Capitanio J.P. Early rearing interacts with temperament and housing to influence the risk for motor stereotypy in rhesus monkeys (Macaca mulatta) Appl. Anim. Behav. Sci. 2011;132:81–89. doi: 10.1016/j.applanim.2011.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Weng H.Y., Kass P.H., Hart L.A., Chomel B.B. Risk factors for unsuccessful dog ownership: an epidemiologic study in Taiwan. Prev. Vet. Med. 2006;77:82–95. doi: 10.1016/j.prevetmed.2006.06.004. [DOI] [PubMed] [Google Scholar]
- 88.Serpell J.A., Duffy D.L. Aspects of juvenile and adolescent environment predict aggression and fear in 12-month-old guide dogs. Front. Vet. Sci. 2016;3:49. doi: 10.3389/fvets.2016.00049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Harvey N.D., Craigon P.J., Blythe S.A., England G.C., Asher L. Social rearing environment influences dog behavioral development. J. Vet. Behav. 2016;16:13–21. doi: 10.1016/j.jveb.2016.03.004. [DOI] [Google Scholar]
- 90.Funder D.C., Ozer D.J. Evaluating effect size in psychological research: sense and nonsense. Adv. Methods Pract. Psychol. Sci. 2019;2:156–168. doi: 10.1177/2515245919847202. [DOI] [Google Scholar]
- 91.Gignac G.E., Szodorai E.T. Effect size guidelines for individual differences researchers. Pers. Indiv. Differ. 2016;102:74–78. doi: 10.1016/J.PAID.2016.06.069. [DOI] [Google Scholar]
- 92.Parker H.G., Dreger D.L., Rimbault M., Davis B.W., Mullen A.B., Carpintero-Ramirez G., Ostrander E.A. Genomic analyses reveal the influence of geographic origin, migration, and hybridization on modern dog breed development. Cell Rep. 2017;19:697–708. doi: 10.1016/j.celrep.2017.03.079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Revelle W.R. Psych: procedures for personality and psychological research. 2018. https://cran.r-project.org/package=psych
- 94.Mangiafico S. 2021. Rcompanion: Functions to Support Extension Education Program Evaluation. R package version 2.4.1. [Google Scholar]
- 95.Canty A., Ripley B. 2021. Boot: Bootstrap R (S-Plus) Functions. R Package Version 1.3-28. [Google Scholar]
- 96.Niskanen J.E., Salonen M., Puurunen J. airGLMs. 2021. https://www.github.com/JNisk/airGLMs
- 97.Kuhn M. 2021. Caret: Classification and Regression Training. R Package Version 6.0-88. [Google Scholar]
- 98.Greenwell B., Boehmke B., Cunningham J., Developers G. 2020. Gbm: generalized boosted regression models. R Packag. version 2.1.8. [Google Scholar]
- 99.Robinson D., Hayes A. Broom: convert statistical analysis objects into tidy tibbles. 2019. https://cran.r-project.org/package=broom
- 100.Wickham H., François R., Henry L., Müller K. Dplyr: a grammar of data manipulation. 2019. https://cran.r-project.org/package=dplyr
- 101.Wickham H. Springer-Verlag New York; 2016. ggplot2: Elegant Graphics for Data Analysis. [DOI] [Google Scholar]
- 102.Hastie T. gam: generalized additive models. 2019. https://cran.r-project.org/package=gam [DOI] [PubMed]
- 103.Fox J., Weisberg S. SAGE Publications; 2019. An R Companion to Applied Regression Third. [Google Scholar]
- 104.Lenth R. emmeans: estimated marginal means, aka least-squares means. 2019. https://cran.r-project.org/package=emmeans
- 105.Fox J. Effect displays in R for generalised linear models. J. Stat. Software. 2003;8:1–27. doi: 10.18637/jss.v008.i15. [DOI] [Google Scholar]
- 106.Greenwell B., Boehmke B. Variable importance plots - an introduction to the vip package. Rom. Jahrb. 2020;12:343–366. doi: 10.32614/RJ-2020-013. [DOI] [Google Scholar]
- 107.Ben-Shachar M., Lüdecke D., Makowski D. Effectsize: estimation of effect size indices and standardized parameters. J. Open Source Softw. 2020;5:2815. doi: 10.21105/JOSS.02815. [DOI] [Google Scholar]
- 108.R Core Team . 2021. R: A Language and Environment for Statistical Computing. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
-
•
Questionnaire data have been deposited at Figshare and are publicly available as of the date of publication. DOI is listed in the key resources table.
-
•
All original code is available in this paper’s supplemental information.
-
•
Any additional information required to reanalyze the data reported in this paper is available from the lead contact upon request.