

Abstract
Abstract concepts can potentially be represented using metaphorical mappings to concrete domains. This view predicts that when linguistic metaphors are processed, they will invoke sensory-motor simulations. Here, I examine evidence from neuroimaging and lesion studies that addresses whether metaphors in language are embodied in this manner. Given the controversy in this area, I first outline some criteria by which the quality of neuroimaging and lesion studies might be evaluated. I then review studies of metaphors in various sensory-motor domains such as action, motion, texture, taste, and time. Idioms are examined next. I also address some neuroimaging studies that can speak to the question of metaphoric conceptual organization without explicit use of linguistic metaphors. Evidence suggests that metaphors are indeed grounded in sensory-motor systems. The case of idioms is less clear, and I suggest that they might be grounded in a qualitatively different manner than metaphors at higher levels of the action hierarchy. For some specific abstract concepts, there is also nonlinguistic neural evidence for metaphoric conceptual organization.
Keywords: metaphors, idioms, embodiment, grounding, abstract concepts
1. Introduction
Any comprehensive theory of the conceptual or semantic system requires an account of abstract concept representation. Abstract concepts can be defined as those that cannot be perceived directly with the five senses (vision, hearing, touch, taste, smell). Concepts such idea or theory as abstract in this sense in that they have no direct sensory correlates, as opposed to concrete concepts such as chair or thunder. While embodiment theories, which suggest that concepts are grounded in action-perception systems (Barsalou 2008; Kiefer and Pulvermuller 2012), apply clearly to concrete concepts, it is not obvious how they can account for abstract concepts.1 Abstract lexical concepts, treated as a single board category, activate mid- and anterior superior temporal sulcus, and inferior frontal gyrus, relative to concrete concepts (Wang et al. 2010). These are multifunctional areas that can be argued to represent embodiment of abstract concepts. However, these areas are also prominently related to executive and phonological processes, and hence the precise nature of grounding is less clear. A number of accounts that seek to explain grounding abstract concepts have been proposed (Kiefer and Harpaintner 2020; Villani et al. 2019; Borghi et al. 2018; Desai, Reilly, and van Dam 2018). These theories point out that abstract concepts do not form a unified category, but rather come in many flavors. Different kinds of abstract concepts can be grounded through their refence to events and situations, meta-cognition, interoception, and emotion.
A different and influential answer to the puzzle of abstract concepts is provided by appealing to analogical mappings between abstract and concrete domains in the form of metaphors. If abstract concepts can be understood in terms of analogical or metaphoric mappings with concrete concepts, which do have sensory-motor features, then abstract concepts can be grounded as well. Figurative language (e.g., metaphors, idioms, proverbs, clichés) is pervasive in everyday life. Pollio et al. (1977) estimated that people use about 6 nonliteral fixed expressions per minute of discourse, including 1.8 new metaphors and 4.08 conventionalized metaphors. This may reflect an underlaying metaphorical structure of the conceptual system. This view was promoted most prominently by Lakoff and colleagues, as Conceptual Metaphor Theory (CMT; Lakoff 1993; Lakoff and Johnson 1980, 1999). A conceptual metaphor is a projection of the basic experiential structure from concrete domains of experience (e.g., action, spatial orientation, movement) to abstract domains (e.g., mental processes, morality, mathematics). For example, the abstract concept of understanding something can be based on the action of grasping an object. This is manifested in the linguistic metaphor grasping an idea, with mappings grasping ⇔ understanding and object ⇔ idea. This view suggests that metaphors are embodied. Abstract ideas are not just described in terms of concrete domains when we use metaphors in language, but are understood in terms of concrete domains.
Several criticisms of CMT and related theories have also arisen (McGlone 2011, 2007; Murphy 1996, 1997). A central question that arises from this proposal is whether metaphor in language necessarily translates to metaphor in conceptual processing. A familiar or conventional metaphor can develop a homonymous sense. A metaphor such as He grasped the idea may be processed similarly to He understood the idea, because grasp has two distinct meanings: one reflecting the action and the other completely abstract, to understand. The embodiment account suggests, instead, that He grasped the idea should be processed similarly to He grasped the handle. The understand sense of grasp is processed through cross-domain mapping with action, and is not abstract.
Here, I review evidence from neuroimaging, brain stimulation, and lesion studies that speaks to the debate on embodiment of metaphoric and idiomatic language. A significant body of behavioral research is exists that has been reviewed elsewhere (Gibbs 2011) and will not be addressed here (for an alternative view, see Casasanto and Gijssels 2015). I begin with a tutorial review of factors to consider when designing and evaluating neuroimaging studies. We will use these criteria to evaluate studies, address some of the criticisms of this neuroimaging work, and attempt to distinguish between good and bad criticisms.
2. Metaphor Embodiment
A number of neuroimaging studies have examined the question of the grounding of metaphoric language, typically by comparing metaphoric sentences to non-metaphoric ones, and examining differential activations in relevant sensory-motor brain areas. A typical comparison is between a sensory-motor metaphor (MET; e.g., The man grasped the idea; She had a rough day) and an abstract or non-sensory-motor control sentence (ABS; The man wanted the item; She had a bad day). Literal action sentences are also often used (LIT2; The man grasped the handle; She had a rough blanket). The key prediction is that sensory-motor areas should be activated for MET > ABS contrast, and they should also be seen for the LIT > ABS contrast. Note that the contrast MET > LIT is not particularly useful with respect to the question of embodiment. This contrast is relevant to the question of how metaphors are processed in general, which is an important but separate issue. The embodiment view predicts a null result in sensory-motor cortex (roughly equal activation for MET and LIT), or somewhat stronger activation for LIT.
Another approach used in several studies is to omit the ABS condition and instead use semantic somatotopy. For example, arm-related and leg-related metaphors can be used as controls for each other by comparing them directly. Here, more activation in an arm region of interest (ROI) is predicted for MET-arm, and more in the leg ROI for MET-leg. Other types of sentences used in studies are idioms (IDI; The country lifted the veil on its nuclear program) and fictive motion sentences (FIC; The road goes through the valley). FIC sentences are motion metaphors that use an inanimate agent that could not literally undergo the described motion. Here, I will adopt this terminology (MET, LIT, ABS, IDI, FIC) to describe various studies for clarity, instead of using the condition names used by the study authors. The notation A > B indicates areas activated more (e.g., those with a higher magnitude of BOLD response in an fMRI study) for condition A relative to condition B.
2.1. How should neuroimaging studies be evaluated?
A number of factors affect the quality and reliability of results, and neuroimaging studies vary widely on some of the dimensions. I briefly review some of the important factors to keep in mind when designing and evaluating studies.
(1). Characteristics of stimulus materials.
Selecting a large number of metaphors and creating appropriate control conditions to study metaphor embodiment is a nontrivial, painstaking task that almost inevitably involves some form of compromise. Most neuroimaging studies have used sentence stimuli containing familiar metaphors. It is common to match conditions on a number of psycholinguistic variables, such as number of words, imageability/concreteness of content words, and word frequency. It is expected that MET and ABS conditions will have lower concreteness than LIT, because of the use of abstract entities (e.g., idea). Another important consideration is the overall naturalness, familiarity, or processing difficulty of the sentences. Ideally, one wants these conditions to have similar level of difficulty. If one condition is more unnatural and difficult (typically MET), it would recruit additional executive resources that can lead to widespread activation, which would be incorrectly attributed as semantic. Many studies use either a rating or behavioral task to estimate and control for one or more of these interrelated variables (by matching conditions, or less ideally, by including it as a covariate in the analysis), which is a strength. Syntactic structure is another variable that ideally should be matched, but this is often difficult while maintaining similar naturalness and other constraints. It is virtually impossible to achieve syntactic match for idioms while maintaining naturalness for a large number of stimuli. Many studies use MET and ABS conditions are that equated on the number of words, but not syntactic structure (e.g., MET: The man fell under her spell; ABS: The merchant was greedy and gluttonous.). Ideally, the ABS conditions should also have a meaning that is very similar to that of the MET condition (e.g., MET: He grasped the idea; ABS: He understood the idea). This is again difficult or impossible to achieve with a large number of stimuli while maintaining naturalness, and hence most studies do not have this feature. One approach is to use only a small number of sentences, which makes close matching on myriad variables more tractable. However, this necessitates repeating the stimuli several times for each participant, because using a large number of subjects, such as 50 or 60, which is commonly done in low-cost behavioral experiments, is usually infeasible in neuroimaging. This results in repetition suppression (reduced BOLD response for repeated stimuli) and a potentially less robust response. Investigators usually strike a compromise between equating the conditions for naturalness, processing difficulty, syntactic structure, and lexical variables such as frequency, at the expense of one of these variables.
Care should also be taken to distinguish between the specific types of figurative language under study. Some studies use a mix of metaphors, idioms, and proverbs, but label this condition ‘metaphor’ or ‘idiom,’ which can potentially be misleading (see Section 4 for further discussion).
With respect to control conditions, some studies use a low-level baseline such as hash marks (‘#####’), false fonts (‘♋♌♍♎’), or fixation (‘+’). This does not control for phonological processing3 and covert speech, which also engage motor and premotor areas. These are precisely the areas of interest for action metaphors, creating a confound. While these sentence > low-level-baseline contrasts can create appealing visualizations by showing large activation clusters in the motor cortex, they can be misleading because they can result from phonology and covert speech rather than semantics. In these cases, critical comparisons are not MET > low-level-baseline, but contrasts that compare sentences with each other, such as MET-arm > MET-leg.
(2). Definition of ROIs.
In order to reduce number of statistical tests, investigators often pick one or more “region(s) of interest” (ROI) based on a hypothesis and restrict the analysis to these. Whole-brain analysis, compared to an ROI approach, has many desirable features. It is theoretically neutral, does not require an anatomical hypothesis, and can reveal expected as well as unexpected or novel results. It eliminates the suspicion that an ROI might have been picked in a post-hoc manner to produce the best results. However, in practice it often tends to be too harsh or stringent, due to the large number of comparisons that require corrections. Relying only on whole-brain analyses would lead to many false negatives. Hence, most authors rely on hypothesis-driven ROIs, which are interrogated for predicted activation patterns. In a roughly decreasing order of desirability, ROIs can be defined in following ways. (1) Definition using a localizer task, and use at individual level. (2) Definition using a localizer task, use at a group level. (3) Regions (typically spherical) around peak coordinates from prior studies. (4) Anatomically defined small and large regions.
Due to anatomical precision at the subject level, (1) is the most desirable option especially if there is significant variability in the location of relevant functional regions, such as motion-sensitive region MT+. However, this option is not used frequently because of the significant added time to run a localizer task long enough to reliably detect a region in single subjects. If limited scanning time is available, spending a significant chunk on the localizer reduces the time available for the main experiment, resulting in a potentially weak result in the task of interest. Hence, most studies use options (2)-(4). It should also be noted that the localizer task and control conditions should be chosen appropriately, so that the resulting ROI is not too general or too restrictive.
Use of large ROIs, and averaging of signal within the ROI, can help reduce noise but is also susceptible to problems with reverse inference (Poldrack 2006). That is, a large region is likely to have subdivisions involved in many functions, and even a small region can be multi-functional. Activation can be observed from non-specific processes that are not well-controlled for, such as attention, decision-making, or phonological and orthographic processing. On the other hand, it is possible to reduce the impact of such non-specific processes by using a carefully designed control condition. Defining well circumscribed ROIs precisely, and in an a priori manner adds to the rigor of the study. Majority of the metaphor studies discussed here have reasonable ROI definitions, coupled with appropriate control conditions, so that reverse inference is not a significant problem in those cases.
(3). Statistical power.
In recent years, there has been increased awareness of the importance of sample size, and potential problems caused by small sample sizes (Turner et al. 2018; Cremers, Wager, and Yarkoni 2017; Button et al. 2013; Pajula and Tohka 2016). An experiment with a small number of subjects is susceptible to not only to false negatives, but also to false positives (Button et al. 2013). A sample size in terms of number of subjects between 20 and 30 is frequently recommended (Desmond and Glover 2002; Murphy and Garavan 2004; Pajula and Tohka 2016) and is also practical. Sample size also depends on size of the effect, number of trials per subject, and anatomical variability of ROIs. Effect size can be expected to be small in case of metaphors compared to a well-matched control condition, while anatomical variability may be high especially higher-order sensory-motor cortex. This is very different from contrasts that use very different conditions or have a sensory difference, such as finger tapping vs. rest, pictures vs. scrambled pictures, pain vs. little or no pain. Number of trials per condition is an equally important variable. Several neuroimaging studies of metaphor don’t perform strongly in this department, using a relatively small sample size of 7-15 subjects and/or a small number of trials per condition.
Now, we turn to studies of metaphors in various sensory-motor domains, discussing first positive and then negative findings. Studies of idioms are discussed next. Table 1 summarizes these studies.
Table 1.
A summary of studies of metaphor and idiom processing.
| Study | # Subj | Conditions (sentences) | Domain | Presentation modality | # items per cond | Definition of ROIs | Task |
|---|---|---|---|---|---|---|---|
| Neuroimaging studies | |||||||
| Romero Lauro et al. (2013) | 24 | Literal, metaphoric, idiomatic, fictive, abstract, upper and lower limb | Action/motion | Visual | 21 | Spherical regions centered on peaks from previous studies | Judge whether a task sentence is congruent with the main sentence using a button press |
| Boulenger et al. (2012) | 18 | Arm- and leg-related literal and idiomatic | Action (arm/leg) | Visual | 38 | Anatomical (hand drawn on average surface) | Answer occasional yes/no questions after catch trials |
| Raposo et al. (2009) | 22 | Arm- and Leg idiomatic | Action (arm/leg) | Auditory | 28 | Finger and foot movements | Word relatedness judgment on 50% the trials |
| Desai et al. (2011) | 22 | Literal, metaphoric, abstract | Action (hand/arm) | Visual | 81 | Whole brain, sequential hand movement | Sentence familiarity judgment after each run |
| Desai et al. (2013) | 27 | Literal, metaphoric, idiomatic, abstract | Action (hand/arm) | Visual | 40 | Whole brain, sequential hand movements | Sensibility judgment on 10% of the trials |
| Lacey et al. 2012 | 7 | Metaphoric and abstract | Texture | Auditory | 54 | Haptic and visual texture perception | Button press after each sentence has been understood |
| Aziz-Zadeh et al. (2006) | 12 | Literal, metaphoric for mouth, hand, foot | Action | Visual | 5 items x 8 repetitions | Videos of mouth, hand, and foot actions | Passive reading; recall after scan |
| Lacey et al. (2017) | 12 | Body part metaphors, body part literal; control literal corresponding to both | Body parts | Auditory | 60 | Visual, motor, and sensory localizers | Positive/neutral/negative valence judgement |
| Saygin et al. (2010) | 136 | Literal motion, fictive motion, static | Motion | Audiovisual (speaking face) | 65 | MT+ localizer (moving vs. stationary dots) | Button press on detecting an anomalous sentence (~11% of trials) |
| Boulenger et al. (2009) | 18 | Literal, idiom for arm and foot | Action | Visual (RSVP) | 38 | Movements of finger or foot | Yes/No question with a button press after catch trials |
| Chen et al. (2008) | 14 | Literal motion, metaphoric motion, non-motive | Motion | Visual | 35 | Anatomically defined temporal and occipital regions | Sensibility judgment after each sentence |
| Wallentin et al. (2005a) | 15 | Literal motion, metaphoric motion, corresponding static | Motion | Auditory | 32 | Anatomically defined | Sensibility judgment after each sentence with a button press |
| Wallentin et al. (2005b) | 18 | Literal, metaphoric, fictive motion, nonsense | Motion | Visual and auditory | 40 | Whole brain | Sensibility judgment after each sentence with a button press |
| Lai & Desai (2016) | 22 | Literal motion, spatial metaphoric motion, temporal metaphoric motion, and three corresponding controls | Spatial motion, temporal motion | Visual | 62 | Spherical ROIs relevant to temporal and spatial processing, centered on coordinates from previous studies | Button press after catch trials (25%) indicating whether a probe word was related to the sentence |
| Citron & Goldberg (2014) | 26 | Metaphor, abstract | Taste | Visual | 37 | Anatomically defined gustatory areas based on WFU PickAtlas | Answer yes/no comprehension questions with a buttonpress on occasional catch trials |
| Brain stimulation/lesion studies | |||||||
| Cacciari et al. (2011) | 8 | Literal, metaphoric, fictive motion, and non-motion | Motion | Visual | 27 | Leg motor cortex localized with TMS | Passive reading, recognition test after |
| Reilly et al. (2019) | 24 | Literal, metaphoric, abstract | Action | Visual | 12-14 per SOA | Hand motor cortex localized with TMS | Sensibility judgment with a left hand button press for each sentence |
| Fernandino et al. (2013) | 20 | Literal, metaphoric, idiomatic, abstract | Action | Visual | 25 | -(Parkinson’s patients) | Sensibility judgment for each sentence |
| Johari et al. (2021) | 23 | Literal, metaphoric, idiomatic, visual | Action | Visual | 20 | HD-tDCS centered on hand motor cortex (group coordinates) | Sensibility judgment for each sentence |
Sixteen subjects were included in the study, but MT+ ROIs could be defined only for 13 subjects.
3. Metaphors
3.1. Action
Desai et al. (2011) used LIT, MET, and ABS sentences that were matched in syntactic structure as well as overall difficulty in a sensibility judgement task. The latter variable is an important potential confound, as more difficult or unfamiliar sentences can activate executive processing areas such as those related to attention and decision making. The metaphors were non-idiomatic but conventional or familiar metaphors, and a meaningfulness judgment task (detect nonsense sentences inserted as catch trials) was used. In a whole-brain analysis, a higher order motor area associated with complex action planning and hand-object interaction in the left anterior inferior parietal lobe (aIPL) was activated not only for LIT > ABS, but also for MET > ABS (Fig. 1a).
Figure 1.
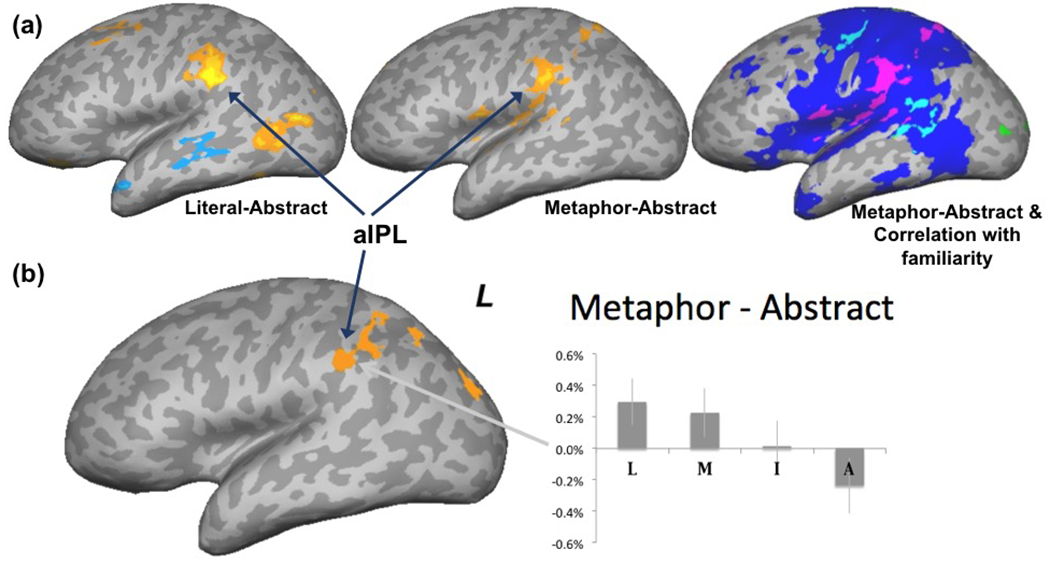
(a) Activations from Desai et al. (2011) for literal and metaphor sentences (yellow-orange) compared to abstract (blue) sentences (left and central panel). In the rightmost panel, blue indicates activation from a motor localizer task, pink is action MET > ABS activation overlapping with the localizer, and cyan shows correlation with metaphor familiarity after factoring out abstract sentence familiarity also overlapping with the localizer. (b) MET-ABS activation from Desai et al. (2013). Graph shows percentage signal change in the aIPL cluster for literal (L), metaphor (M), idiom(I), and abstract (A) conditions. Adapted from Desai et al. (2011) and Desai et al. (2013) respectively.
AIPL has a well-established role in tool use, action planning, and goal-oriented action execution from numerous neuroimaging and lesion studies (Randerath et al. 2017; Vingerhoets 2014; Binkofski, Klann, and Caspers 2016; Jax, Buxbaum, and Moll 2006; Johnson-Frey, Newman-Norlund, and Grafton 2005; Buxbaum et al. 2007; Peeters, Rizzolatti, and Orban 2013) in addition to action semantics in single-word studies (e.g., Kuhnke, Kiefer, and Hartwigsen 2020). It is the central node in a tool use network formed by structural connections to posterior middle temporal and inferior frontal regions (Ramayya, Glasser, and Rilling 2009). The aIPL cluster covered both anterior supramarignal gyrus (aSMG) and the anterior end of the intraparietal sulcus (aIPS). Examining a large number of neuroimaging studies, Orban and Caruana (2014) found that aIPS was activated by observing a tool being grasped, while aSMG was activated by observation of moving the tool to achieve a goal (e.g., picking up an object). Similarly during action execution, they suggested that aIPS is associated with planning the grasp, and aSMG with planning the tool movement to achieve a goal. Ideomotor apraxia is often the result of damage to aIPL (Haaland, Harrington, and Knight 2000; Jax, Buxbaum, and Moll 2006). It results in impairments in imitating actions and gestures, pantomiming, recognizing object-related pantomimes, and planning object-related actions (Varney and Damasio 1987; Buxbaum, Johnson-Frey, and Bartlett-Williams 2005; Goldenberg and Karnath 2006). TMS to aSMG causes a delay in planning goal-oriented actions, but not in responses to arbitrary stimuli (Tunik, Lo, and Adamovich 2008), and TMS to aIPS results in on-line adjustments of grasp (Tunik, Frey, and Grafton 2005). Thus, converging evidence from neuroimaging, lesion, and brain stimulation studies point to a role of aIPL in planning and execution of goal directed actions. This region is recruited in service of comprehension of both literal and metaphoric action language, presumably supporting higher-order simulations of object-related actions.
The results also showed that the response in a primary motor area was inversely related to metaphor sentence familiarity, but not to abstract sentence familiarity, showing a familiarity × condition interaction (Fig 1a, rightmost panel). This suggests a progression whereby primary motor cortex is recruited for processing relatively unfamiliar metaphors, and this reliance reduces as metaphors become more familiar. Note that very familiar (but non-idiomatic) metaphors still recruit the higher order action planning area in the left aIPL, which did not significantly vary with metaphor familiarity.
Desai et al. (2013) used MET, LIT, ABS, and IDI sentences. The sentences were and longer and more varied in their syntactic structure compared to the Desai et al. (2011) study, due to inclusion of idiomatic sentences that were matched in length to other conditions. The MET > ABS comparison yielded similar results to that study, showing activation in the left aIPL in a whole-brain analysis, with additional activation in the intraparietal sulcus and middle occipital gyrus (Fig 1b). AIPL shows a linear trend, LIT > MET > IDI > ABS.
An additional finding in these two studies concerns the differences between the metaphoric and literal sentences. While metaphoric sentences were similar to the literal sentence in the aIPL, they activated an additional area in the mid superior temporal sulcus and middle temporal gyrus (mSTS/mMTG)4. This area was also seen in the ABS > LIT, and can be considered a signature of abstract semantics (whatever its internal format may be). Thus, action metaphors showed similarities to both literal and abstract sentences, and were not identical to literal action sentences.
In an EEG study, Lai et al. (2019) examined the time-course of activation for action metaphors used in the Desai et al. . (2011) study. They found that the frontal concreteness effect was similar for both LIT and MET sentences compared to ABS, with the same time-course. This suggests that action verbs in both metaphoric and literal contexts are interpreted similarly, as a concrete action. The MET > ABS comparison showed additional negativity in posterior electrodes, possibly reflecting additional resources recruited for metaphor interpretation. Thus, action metaphors showed both similarities and differences relative to literal action sentences.
Romero Lauro et al. (2013) found activation in a hand sentence ROI in the precentral gyrus for MET > ABS for upper limb action sentences. The same ROI also showed activation for upper limb LIT > ABS contrast.
3.2. Texture and Body Parts
Lacey et al. (2012) used texture metaphors such as She had a rough day and contrasted them with similar ABS5 sentences (She had a bad day). In functionally defined texture-sensitive ROIs in left OP1, left OP3, and right OP1, greater activation for MET than ABS was found (Fig. 2a). While the small sample size (n=7 subjects) is a reason to be cautions, the results are suggestive.
Figure 2.
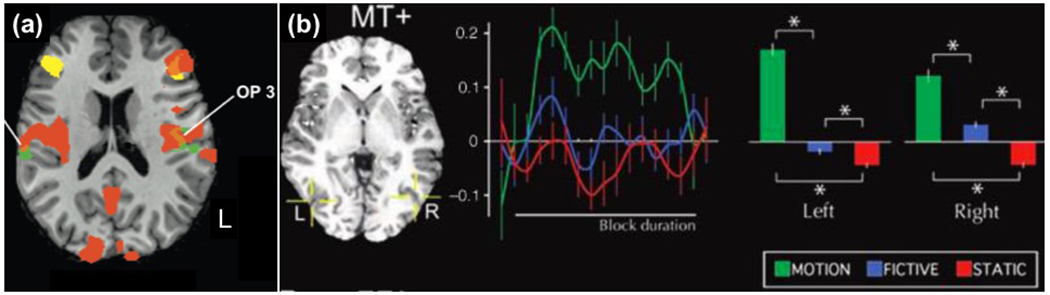
(a) The main result from Lacey et al. (2012) study of texture metaphors. Orange – subject-specific haptic texture localizer; yellow – visual texture localizer; brown/green – textural metaphors and their overlap with haptic localizer in sensory area OP3 in the left hemisphere, in OP1 in the right hemisphere. (b) Results from Saygin et al. (2010) study that used motion, fictive motion, and static sentences. Graphs shows activations in individually localized area MT+. Adapted from Lacey et al. (2012) and Saygin et al. (2010) respectively.
Lacey et al. (Lacey et al. 2017) used body-part-specific metaphors (He is under her thumb), LIT sentences with body parts (She scraped her knuckles), and corresponding control sentences without body part terms (ABS: He is under her control; LIT-control: She scraped her plate). Metaphoric sentences had more words, and had lower naturalness, familiarity, and interpretability than the literal body part sentences, but still contained metaphors that had relatively high familiarity. Potential concerns related to time-on-task effects is alleviated by the fact that metaphor sentences had faster RTs than their control sentences. The interaction contrast (MET > ABS) > (LIT > LIT-control) activated a cluster in the left STG, and partly overlapped with extrastriate body area (EBA) identified in the visual localizer task. However, the motivation behind this contrast not clear, because one expects both metaphoric body part and literal body part sentences to activate the EBA, and not one relative to the other. Contrasts MET > ABS and LIT > LIT-control were also examined separately, and overlap with limb-selective EBA for the former, but not the latter, was found. An additional interaction contrast, (limb metaphor > limb-control) > (face metaphor > face-control) also revealed activation in the EBA, showing that this activation is specific to limb metaphors, and is not a general characteristic of metaphor processing. The main puzzling aspect of the results is the question of why LIT sentences did not activate EBA. If metaphoric sentences are grounded in visual areas that process body parts, surely literal body-part sentences should also be grounded in this manner. At least two explanations can be offered. One, the metaphor control sentences were abstract, while the literal control sentences referred to physical actions and manipulable objects (She scraped her plate; She closed her door; His chair was small). These control sentences may also activate the EBA to some extent due to implied use of body parts, weakening the contrast LIT > LIT-control. Secondly, the small sample size (n=12 subjects) may have reduced power to detect effects.
3.3. Motion
Saygin et al. (2010) used motion-related LIT sentences (I drove from Modesto to Fresno), FIC sentences (The highway runs from Modesto to Fresno) and static ABS sentences (Modesto and Fresno are in California). Motion-sensitive area MT+ localized in each subject using a separate localizer run. They found strong activation in both the left and right MT+ for LIT > ABS. They also found a small but significant activation for FIC > ABS in both of these regions, suggesting that metaphoric motion engages motion-related simulations (Fig. 2b). They observed a gradient, LIT > FIC > ABS, which is reminiscent of the gradient found in Desai et al. 2011 and Desai et al. 2013 studies in aIPL (LIT > MET > IDI > ABS) for action metaphors.
Chen et al. (2008) used motion predicate metaphors (PM; The man fell under her spell), literal motion (LM; The child fell under the slide), and non-motive abstract sentences (NM; The merchant was greedy and gluttonous). MET and LIT sentences were matched on a number of variables including concreteness, imageability, familiarity, naturalness, and plausibility. MET nouns had a higher written frequency than the other two types. The ABS sentences had lower concreteness and imageability, and greater naturalness and plausibility. Behaviorally, there was no difference in RT between the sentences, but METs were less accurate than the other two conditions. MET > ABS comparison reveal activation in pMTG. However, the LIT > ABS comparison did not show any activations for LIT, making the interpretation of pMTG as representing conceptual motion difficult. They suggest a posterior-to-anterior gradient of abstraction in motion processing, where literal sentences activate more posterior regions, and starting around MT+. Gradually more abstracted motion is processed as one moves along the lateral temporal lobe. A relatively small number of participants (n=14) might affect the results.
Wallentin et al. (2005a) used FIC and LIT motion sentences and compared them to static ABS sentences. Motion-related sentences together activated the pMTG compared to static sentences, and there were no differences among LIT and FIC sentences in this region. The critical FIC > ABS contrast was not presented, but the FIC condition activated pMTG relative to meaningless control sentences containing nonwords. Another study by Wallentin et al. (2005b) used LIT, MET, and FIC sentences, but did not contain an ABS condition, and hence is difficult to evaluate.
Lai and Desai (2016) used spatial metaphors (fictive motion) (The trail crawled until the end of the hills) and literal motion sentences (The caterpillar crawled towards the top of the tree), with corresponding static sentences (The trail faced the end of the hills; The caterpillar looked at the top of the tree). This study also included temporal motion sentences, which will be discussed in the next section. Each sentence type was matched to its control on a number of psycholinguistic variables, and on rated comprehension difficulty. In ROIs defined by coordinates from previous studies, the spatial FIC sentences activated the pITG ROI anterior to area MT+, relative to its control sentences. The MT+ ROI did not show a difference in either fictive or literal motion sentences relative to their controls.
3.4. Taste
Citron and Goldberg (2014) used taste metaphors (She looked at him sweetly) contrasted with ABS (She looked at him kindly), and found activation for metaphors in the left gustatory cortex (lateral orbitofrontal cortex, frontal operculum, and anterior insula). This activation was also found for single taste words compared to abstract words, making it unlikely that the activation reflects general metaphoric processing.
3.5. Time
Lai and Desai (2016) used metaphors for temporal duration such as Her seminar stretches across the afternoon, with corresponding static control sentences (Her seminar is in the afternoon) . Activation from these sentences was examined in specific locations identified in a meta-analysis of temporal perception studies (Wiener, Turkeltaub, and Coslett 2010). Coordinates in the left anterior insula, IFG, and the right claustrum was upregulated for temporal metaphors relative to their static controls, but not for spatial metaphors or literal motion sentences relative to their controls. This suggests that portions of the temporal perception network may be used to process temporal motion.
3.6. Brain stimulation studies
Cacciari et al. (2011) used LIT, MET, FIC, and IDI motion sentences compared with ABS sentences in a TMS study. TMS to the left primary leg area resulted in greater motor evoked potentials for LIT, MET, and FIC sentences (but not IDI sentences) than ABS sentences.
Reilly et al. (2019) applied TMS at three time points (150 ms, 300 ms, 450 ms) after the onset of the verb in LIT, MET, and ABS sentences. With hand motor TMS at the 300 ms point, both LIT and MT sentences produced a significant slowing of RT compared to the control occipital TMS in the item analysis. Additionally, these two conditions also differed from ABS at 300 ms. (In the linear mixed effects analysis, the metaphoric condition produced a trending result at p < 0.053.).
HD-tDCS stimulation to hand motor cortex was used by Johari et al. (2021), who found interactions (MET, ABS) × (real, sham stimulation) and (LIT, ABS) × (real, sham) in RT in a meaningfulness judgment task. The action sentences were speeded up with the application of HD-tDCS relative to the visual control sentences, which is consistent with stimulation parameters that are known to induce excitation.
These studies suggest a causal or functional role of motor areas in processing action metaphors as well as literal action language.
3.7. Negative results
In an fMRI study, Aziz-Zadeh et al. (2006) tested LIT and MET sentences. Action ROIs were defined by observation of actions. They found that LIT, but not MET sentences activated the action ROIs.
In Parkinson’s patients and in age-matched controls, Fernandino et al. (2013) tested comprehension of LIT, MET, IDI, and ABS sentences using a meaningfulness judgment task. For the group (patients, controls) × sentence type (MET, ABS) interaction, and did not find effects for response times. This interaction was significant for LIT and IDI sentences. In patients, there was a MET > ABS effect for RTs (p < 0.005), while this difference was not significant in the control group. However, the critical RT interaction did not reach significance.
3.8. Discussion
On a fairly consistent bases, sensory-motor metaphors activate corresponding sensory-motor areas. Action metaphors are the most commonly examined, but metaphors in other domains also show similar patterns. This result is supplemented by brain stimulation studies as well. While not all studies are ideal (for example, several use a small participant and/or stimulus set size, some don’t report the critical MET > ABS contrast but rather discuss MET > LIT), there is sufficient consistency across studies to suggest that metaphors are grounded in sensory-motor system.
The negative result reported by Fernandino et al. (2013) is puzzling, given that in the same study, they found the interaction for both LIT and IDI action sentence RTs. A possible explanation lies in the accuracy, where trending interaction was found (p < 0.08) for metaphors. The MET accuracy was lower than ABS in patients, while it was higher in the control group. This can be a case of speed- accuracy tradeoff (Fernandez et al. 2018; Bogacz et al. 2010), where some of the increase in RT is absorbed by a reduction in accuracy.
On the other hand, for Aziz-Zadeh et al. (2006), two methodological factors may explain the negative result for MET. First is the low number of subjects in the study (n=12), which is about half of the recommended number and reduces the power. Secondly, there were only five different sentences per condition, which were repeated eight times each. Repetition suppression can play a role in reducing the magnitude of the BOLD response.
In case of motion metaphors, it is noteworthy that only one study found activation in the traditional motion perception complex MT+ (Saygin et al. 2010). The other three studies reported activation around this area, in pMTG or pITG. PMTG and pITG are indeed associated with action and motion, and hence the interpretation of this activation as motion-related can be justified. However, pMTG also has a more general role in sentence and verb frame processing. While these factors are accounted for by the control sentences, there may be a lingering doubt whether this activation is truly specific to motion semantics. Importantly, only Saygin et al. (2010) used a subject-specific localizer task to identify MT+. They found significant individual variability in its location, as much as 10 mm on the anterior-posterior axis. This high variability may explain why the other three studies, which used a common anatomical location across subjects, failed to find activations in MT+, and highlights the importance of subject-specific localizers.
Some studies show in higher-order sensory-motor cortex, such as aIPL, for metaphors and literal sentences. Some authors have argued that activation only in primary areas (such as M1 or V1) constitutes valid evidence for embodiment. Activation of higher-order areas does not count, because these areas are multi-functional and modality nonspecific, and hence could represent some other processes. I disagree with this claim for multiple reasons. First, sensory-motor systems have been well-studied and are known to extend well beyond primary regions. It is entirely possible for a process to have a visual component even if it occurs outside V1, and analogously for motor processes to occur outside M1 or auditory processes outside A1. For example, even in the macaque, corticospinal tracts receive only about 35% of the fibers from the primary motor cortex, and the rest of the input is from premotor, parietal, cingulate, and SMA regions (Galea and Darian-Smith 1994). It is not possible to simply declare that these regions are not “real” motor areas, because they are (Rizzolatti and Luppino 2001). A simplistic dichotomy between “modality specific” and “modality nonspecific” areas is mistaken.
The concern for potential confounds is legitimate, as indeed most if not all brain areas are multi-functional, depending on what is meant by “area” and “function.” That is why control conditions are included to account for nonspecific processes such as those related to phonology, orthography, attention, general sentence processing, executive processing, or task performance. Activation for a metaphoric sentence relative to a low-level baseline such rest or fixation could arise from myriad non-specific processes. But if a control condition that accounts for such processes is included, then activations in higher-order sensory-motor cortex can indeed be interpreted as evidence for embodiment. Many studies have taken great pains to carefully select stimulus materials for this reason. It is always possible that a confound was missed by the experimenters. However, the onus then is on the critic to (1) identify the specific confound, and (2) show that the confound could plausibly result in activation in the particular areas that were interpreted as reflecting sensory-motor processes. The best response is to conduct a study in which the putative confound is eliminated, and show that the purported sensory-motor activation disappears. It is not sufficient to simply state that activation in higher-order sensory-motor areas could reflect any process and does not count. Functions of brain areas are not arbitrary, and their activation in neuroimaging studies is also not arbitrary. Much is known about many higher-order areas, which can and should be used to interpret results. While reverse inference can indeed be used in an inappropriate manner (Poldrack 2006), it is not inherently undesirable or a “bad thing,” and can also be used in a valid and informative manner. Several investigators have suggested that conceptual processing involves a hierarchy of areas ranging from primary to higher level sensory-motor areas, to multimodal and heteromodal areas such as angular gyrus (Kiefer and Harpaintner 2020; Fernandino et al. 2016). Context and task demands determine the depth of activation at various levels of this hierarchy in a flexible manner.
While majority of the studies have examined action and motion metaphors, limited evidence exists for other domains as well, such as taste and time. Further examination of these other domains is clearly desirable. For example, a single study has examined temporal metaphors (Lai and Desai, 2016) that used coordinates from a meta-analysis of temporal perception studies to identify areas relevant to time processing. An obvious next step is to use temporal processing tasks as localizers at an individual subject level. Cross-linguistic studies, especially in languages that may use different metaphors than English (e.g., with a different direction of motion in space and time, or with different mappings between pitch and space), is another direction that would be of value.
Metaphoric representation and abstraction
Jamrozik et al. (2016) propose that the metaphoric abstraction process has two characteristics: relational highlighting and sensory-motor shedding. Deeper relational meaning is highlighted, while superficial sensory-motor features are eliminated by the metaphor. For example, in the metaphor Problems galloped into their marriage, surface features such as four legs of a horse being off the ground are eliminated, while a deeper meaning is highlighted. Here, I present a partially different view. I propose that the intuitively appealing dichotomy between “deep” vs. “shallow,” or “core” vs. “surface” features of concepts, is mistaken. There is no essence of a concept, or a core, deep, or real meaning that is independent from the so-called surface features. They are inextricably intertwined so as to make this distinction fundamentally unsound. Metaphors highlight which sensory-motor features and relations are emphasized and can affect levels of detail, but never eliminate them altogether. Relational meaning is certainly important, but relations can exist only between entities. If they are between sensory-motor features, then they are sensory-motor relations. There is no core or deep meaning of gallop that is independent from the supposedly shallow features such as motion. Even additional details of speed and manner of motion are retained in the metaphor. If the metaphor is changed to Problems seeped into their marriage, the slow, gradual, and quiet manner of motion is highlighted. Metaphors work by sensory-motor selection, and not by sensory-motor shedding.
Metaphors were found to have similarities to literal sentences in sensory-motor areas, but also to abstract sentences in the middle temporal lobe. Abstract concepts like understand, when expressed metaphorically as grasp, indeed have sensory-motor bases. However, the metaphoric grasp also has similarities to understand expressed literally. The mid-STS and anterior STG regions, activated by abstract concepts in single-word studies, and by abstract sentences, are also activated by action metaphors (the blue region in the left panel of Fig. 1a shows this activation, which is similar between ABS and MET conditions). This has implications for two models of metaphoric conceptual processing outlined by Murphy (1996). In the strong version, abstract concepts are defined entirely using the metaphors they are used in, and have no independent content. In the weak version, abstract concepts have basic independent representations, but are shaped by metaphoric usage (see also Gibbs 1994a). The results above are more consistent with the weak version. The most common interpretation of temporal activation relates to phonology and phonological neighborhoods. A component of meaning could be represented by activating other concepts, and the temporal lobe system represents the phonological access point to other concepts. This does not necessarily imply that this representation itself is abstract or symbolic. Abstract concepts themselves, and their neighbors accessed through phonology, could be grounded indirectly in sensory-motor systems, or in affective, interoceptive, and meta-cognitive systems (Desai, Reilly, and van Dam 2018; Borghi et al. 2019). The results only show that action metaphors are not identical to literal sentences, and also have similarities to sentences with abstract meaning. A metaphor such as theories are buildings is used when someone suggest that a certain theory has a solid foundation. But people are still aware of differences between theories and buildings. It is possible that the concept of theory has an independent representation that is skeletal or impoverished, and is enhanced and enriched by frequent metaphoric alignment with buildings. This independent representation can be enhanced in different, event contradictory ways (e.g., by saying that a theory is slippery or fluid, which would not be possible for a building). On the other hand, concepts are likely dynamic and flexible, and need not have a fixed or static representation at all. The problem of multiple and contradictory metaphors can be handled by a view in which conceptual representations are created on the fly based on the context (Gibbs 1996). The question then is, what neural representation of theory is activated in a neutral context such as in I like theories? Experiments that can test whether such a representation is a “weighted average” of representations based on different metaphors for theories, or has an independent, non-metaphoric component, would be of value.
Theories are simultaneously more than buildings and less than buildings, and an appeal to additional metaphors is necessary if that is the only tool available. Furthermore, establishing appropriate connections between domains and discarding others seem to require prior understanding of the concepts. Abstract concepts appear to lack a prior structure that can be readily mapped to a concrete domain. The role of metaphoric mapping is precisely to provide that structure, but this assumes a pre-existing concept (presumably based on direct experience), which can be structured. Hence, while it is not obvious that metaphor can provide a complete solution to the problem of abstract concept representation (Dove 2009; Barsalou and Wiemer-Hastings 2005), it may certainly play a role.
4. Idioms
Some of the behavioral and neuroimaging literature on metaphor processing commonly uses the terms ‘metaphor’ and ‘idiom’ interchangeably, and some consider idioms to be equivalent to conventional metaphors. For the purposes of the current article, I distinguish between metaphors and idioms (sometimes also called “dead metaphors”). An idiom is a string of constituents whose meaning is not necessarily derived from that of the constituents. Idiomatic meaning is retrieved from long-term memory. In an idiom such as spill the beans, both spill and beans are essential elements. Drop the beans or spill the marbles do not convey the same meaning. On the other hand, the metaphoric notion ‘grasping as understanding’ can be applied to many cases. Unlike idioms, metaphors – either novel or conventionalized – do not have a unique standardized meaning. A case such as grasp the idea is a familiar or conventional metaphor, but is not an idiom, because one can grasp a theory, grasp the situation, and so on. This productive nature of metaphors that allows for online combination of words distinguishes them from idioms.
Idioms are theoretically important due to their controversial nature in two aspects: metaphoricity and compositionality (Sanford 2014; Vulchanova et al. 2019). Non-compositional models of idiom processing suggest that idioms are stored and retrieved as chunks of constructions, and function as a single lexical item (e.g., Swinney and Cutler 1979; Gibbs 1994b). Compositional models propose that idioms are analyzed compositionally at some level, and single words can have influence on its interpretation (e.g., Cacciari and Tabossi 1988; Hamblin and Gibbs 1999). Hybrid models that take elements of both proposals also exist (Titone and Connine 1999). For our purposes, if the abstract meaning of an idiom is retrieved as a single lexical item, then it should not involve sensory-motor systems in the same way that metaphors do. If idioms are found to be grounded in sensory-motor systems, then compositional or hybrid accounts are supported. Idioms also vary in the degree of their decomposability or transparency, and this can affect the level of compositional analysis. Neuroimaging studies have not addressed this variable systematically, and hence we will not discuss it here but note it as a caveat.
4.1. Positive findings
Boulenger et al. (2009) used arm- and leg-related IDI and LIT action sentences in an fMRI study. There was no ABS condition in this study, and the baseline only contained hash marks matched in length to the sentences. A differential effect for arm- and leg-related idioms was found in corresponding ROIs in the motor strip, showing semantic somatotopy, in a late analysis window that started from 3 s after the offset of the final critical word (habit in Pablo kicked the habit).
In a MEG study, arm- and leg- related idiomatic and literal sentences were used by Boulenger et al. (2012). In an analysis collapsing LIT and IDI sentences, significantly greater activity was found in the arm ROI for arm sentences relative to the leg sentences. Activation of leg sentences in the leg ROI relative to that from arm sentence did not reach significance. In another analysis examining individual time points (as opposed to time 50-ms time windows used in the previous analysis), both arm and leg ROIs showed a body part × ROI interaction.
In Parkinson’s patients, Fernandino et al. (2013) found increased response times for literal and idiomatic action sentences relative to abstract sentences in a an interaction with a control group (IDI, ABS) × (patient, control).
Romero Lauro et al. (2013) found activation in a hand sentence ROI in the precentral gyrus for IDI > ABS for upper limb action sentences. The same ROI also showed activation for upper limb LIT > ABS and MET > ABS contrasts, supporting its role in action semantic processing.
Johari et al. (2021) applied HD-tDCS centered on the hand motor cortex and found interactions (IDI, ABS) × (real, sham stimulation) and (LIT, ABS) × (real, sham stimulation) in RT in a meaningfulness judgment task, with relative facilitation for IDI and LIT sentences.
4.2. Negative findings
Raposo et al. (2009) used arm- and leg-related LIT and IDI sentences in an fMRI study. In the critical analysis, an interaction between arm/leg sentences and arm/leg ROIs was observed for LIT, but not for IDI sentences.
Cacciari et al. (2011) failed to find an effect on motor evoked potentials (MEPs) for IDI > ABS with TMS to leg motor cortex, while finding this effect for LIT and MET sentences.
Desai et al. (2013) found no activation in motor areas such as aIPL for IDI sentences, while finding this activation for LIT and MET sentences relative to ABS.
4.3. Discussion
Given the existence of several positive as well as negative results, the case of idioms warrants closer examination. Studies by Boulenger et al. (2009) and Boulenger et al. (2012) are among the best designed, due to the use of somatotopy. Rather than using an ‘abstract’ condition, where concepts may have indirect associations with actions, use of semantic somatotopy in these studies provides an ideal control, because actions are compared with other actions executed with a different effector. These studies also use individual localizer tasks rather than using group coordinates of other studies. They also have some limitations. For example, Boulenger et al. (2012) report only analyses that collapse literal and idiomatic sentences.
Stimulus materials used in some studies are also less-than-ideal. One issue is that the stimuli appear to be a mix of idioms and metaphors, rather than just idioms. For example, sentences such as She caught the sun and He grasped the idea are metaphors rather than idioms. A second issue is that some of the stimuli, while containing idioms, may not be processed as idioms due to a lack of context. For example, it is not clear that the sentence John scraped the barrel (Boulenger, Shtyrov, and Pulvermuller 2012) fully activates an idiomatic meaning. (Contrast with John could not find anyone qualified for this job and now he is scraping the barrel, which is more likely to be understood in the intended idiomatic sense.) The isolated sentence could be processed more like a literal sentence if presented with minimal context. The general criticism of a lack of context to fully engage an idiomatic sense is applicable to all of the studies. In some studies, this problem is mitigated by changing the context in which action verb is encountered. Desai et al. (2013) and Fernandino et al. (2013) used inanimate agents in idiomatic sentences (The country lifted the veil on its nuclear program) and animate agents in literal sentences (The craftsman lifted the pebble from the ground). The context provided by the noun phrase makes literal interpretation of the action verb implausible for idioms. However, this is still limited context consisting of a single noun phrase. Studies that provide a more extensive context to clearly activate an idiomatic meaning (e.g., The country was very secretive until it lifted the veil on its nuclear program) would be valuable. Such stimuli create additional difficulties in matching conditions on psycholinguistic variables, which have to be addressed (e.g., by presenting a context sentence separately).
4.4. Re-analysis of Desai et al. (2013) study
Given the inconsistency in results, I more closely examined results of the Desai et al. (2013) study that used IDI, MET, LIT, and ABS conditions. While studies typically examine only a few “motor areas,” namely premotor cortex and aIPL, the motor system is extensive and partly hierarchical. Subcortical areas, especially the basal ganglia (BG), play a well-known role in action control. It is possible that action language, including idioms, engages the motor system, but at higher levels in the hierarchy. Some evidence is provided by Fernandino et al. (2013), who reported impairment of action idioms in Parkinson’s patients, where subcortical motor circuits play a central role in action impairment.
Striatum and globus pallidus are central regions associated with action selection, planning, and modulation of ongoing action (Jin, Tecuapetla, and Costa 2014; Graybiel 1998; Markowitz et al. 2018; Kim and Hikosaka 2015). Within the striatum, lateral dorsal striatum/putamen is associated with rigid and automatic form of stimulus bound behavior that is less relevant to action here. In contrast, medial dorsal striatum/caudate is associated with flexible, goal-directed action, and contingencies between actions and consequences (Grahn, Parkinson, and Owen 2008). Lehericy et al. (2006) found activation in caudate and anterior globus pallidus when comparing complex actions to simple actions (that included movements with and without simple temporal sequencing). Using peaks in right and left caudate (±14, 10, 9) and anterior globus pallidus (±18, 2, 2) from Lehericy et al. (2006), I defined spherical ROIs with 5 mm radius. Comparing action idiom to abstract sentences using t-tests, results indicated significantly greater activation in the right caudate (p < 0.002) and left caudate (p < 0.042). Trends in the right and left anterior globus pallidus (p < 0.085 and p < 0.065 respectively) were observed. The right caudate activation was also significant after corrections for multiple comparisons (Benjamini and Hochberg 1995).
These results suggest a role of BG, and specifically caudate, in action idiom processing. Caudate has a role in facilitating appropriate motor programs or action schemas while inhibiting competing ones, and in selection of appropriate action sub-goals (Grahn, Parkinson, and Owen 2008; Jin, Tecuapetla, and Costa 2014), which may be used to ground action idioms. In addition to the well-known direct, indirect, and hyperdirect pathways, additional pathways between BG and cortex have been identified (for reviews, see Milardi et al. 2019; Haber 2016). For example, a direct cortico-pallidal pathway projects mainly to sensorimotor regions, and has a likely role in sensorimotor integration (Cacciola et al. 2019). Projections between caudate and sensory-motor regions, including premotor cortex and pre-SMA, have a role in planning and learning sequential movements (Haber 2016). Connectivity between BG and cerebellum is implicated in motor timing, and in movement disorders such as dystonia (Milardi et al. 2019).
The results suggest a role of BG and associated connections to sensory-motor cortical regions and cerebellum, in action idiom comprehension. A potential role of BG circuits in idiom comprehension is in action selection and planning. Action idioms may rely on selection of action schemas, but this activation may not fully percolate all the way to higher order cortical areas associated with action execution, such as the aIPL. This grounding may be more automatic and low-level, whereby simulations weaken after action planning stage. It is interesting to note that pre-SMA was also activated for IDI > ABS contrast (significant after whole-brain correction), which is tightly connected to the basal ganglia, and also has a role in action selection and initiation (Nachev, Kennard, and Husain 2008; Nachev et al. 2007; Akkal, Dum, and Strick 2007). When stimulation is applied to cortical motor areas, as in Johri et al. (2021), it may percolate through motor circuits to the level of BG through cortico-BG loops, leading to modulation of idiom processing through cortical stimulation. While Johari et al. used a behavioral measure, Cacciari et al. (2011) used MEPs in their TMS study. If idioms are processed primarily at the level of action selection and action plans, this may reduce effects at the level of MEPs in muscles, potentially explaining the negative finding and the difference between the two stimulation studies.
A concern with this interpretation is that while BG have a well-established role in action control, they also have a more general role in executive function (Riva, Taddei, and Bulgheroni 2018; Guo et al. 2018) . Could the observed activation represent non-specific processes related to general task demands or task difficulty? Two arguments can be presented against this possibility. First, idiom sentences in this study were carefully designed to have similar difficulty as abstract sentences. No difference in RT (in a meaningfulness judgment task) was found between IDI and ABS sentences, either in subject analysis or in item analysis. Secondly, MET sentences in this study also had similar (numerically higher, but statistically no-significant) RT to IDI sentences, and hence can be expected to have at least as much executive demands as the IDI condition. No activation in the ROIs was observed for MET > ABS contrast. This makes it unlikely that the activation here represents general sentence processing or decision-making processes, even for non-literal language.
Could the activation of BG represent an inhibition of action simulations rather than their planning, given that BG loops also play a role in action inhibition? This would imply that while LIT and MET sentences involve simulating actions at some level, comprehension of IDI involves the opposite process of action inhibition, and hence they are not grounded in the same manner. This is unlikely, given that brain stimulation results, such as those in Johari et al. (2021), result in identical effects for LIT and IDI sentences. If one type of sentence was understood through activating action simulations, while the other by inhibiting them, stimulation of motor cortex should result in opposite effects (facilitation in one case while interference in the other), which is not the case.
In sum, this result suggests a possibility that action idioms, due to their well-learned nature, are processed at the level of relatively abstract action schemas in BG and pre-SMA, while action metaphors are processed on-line in higher order motor cortex with relatively more detailed simulations. This may reconcile inconsistent positive and negative findings related to action idiom processing. This supports compositional and hybrid accounts of idiom processing that suggest some level of compositional analysis and influence of individual words in an idiom.
5. Metaphoric organization without linguistic metaphors
The studies discussed above pertain to processing of linguistic metaphors. They show that when metaphoric language is processed, it is understood in terms of the source domain. CMT, however, suggests that there is a metaphoric relationship between abstract and concrete domains at the conceptual level, not just at the linguistic level. A mapping between concrete and abstract domains exist independently on language, and metaphors in language are a reflection of this underlaying conceptual relationship. The fact that linguistic metaphors appear to be grounded in sensory-motor processing, as reviewed above, supports this view, but it is not sufficient to establish an underlaying nonlinguistic relationship. Relationships between nonlinguistic domains of knowledge may not necessarily be reflected in linguistic metaphors (Casasanto 2009), and vice versa. While there is behavioral evidence for such nonlinguistic relationship, at least in some circumstances (Casasanto and Bottini 2014), neural evidence is very limited. Here, I describe three studies that speak to this question.
To examine whether visuospatial mechanisms may be used to process even isolated words, Quadflieg et al. (2011) used words associated with up/down location (airplane/carpet), positive/negative valence (beauty/accident), and high/low power (boss/assistant). Using a classifier that was trained to classify the physical position of shapes, they showed that location and valence (but not power) can also be classified using the same classifier without further training, especially using the ROI of left IPS. This suggests that salient features of location and valence can be grounded in spatial processes, even when they are not used explicitly in metaphors. A potential objection is that the IPS is multi-functional area, and in addition to spatial processing, is important to visual attention. These results could reflect differences in attentional demands or time-on-task differences between conditions, and not grounding in spatial processing per se. The fMRI task involved responses only on catch trials, and hence this possibility cannot be definitively ruled out. However, Quadflieg et al. compared response times and accuracy of their stimuli off-line, and found no differences between conditions, mitigating this concern.
Zanolie et al. (2012) examined the relationship between power difference in positions (e.g., boss vs. servant) and the vertical dimension of space, reflecting the conceptual metaphor power is up, in an ERP study. They found that congruent trials (high power associated with upper visual field) elicited a larger N1 than incongruent trials. The N1 can be interpreted as indexing a shift in spatial attention. This result is consistent with the notion that semantics of power are interpreted metaphorically through space, without presence of linguistic metaphors.
Tavares et al. (2015) used a social game to examine dimensions of power (e.g., dominance, hierarchy, competence) and affiliation (e.g., intimacy, trustworthiness, love). Just as power is associated with the vertical dimension of space, affiliation is frequently described as a horizontal dimension of space or distance and is reflected in linguistic metaphors (She is very close to her niece; We are growing apart). In the experiment, subjects developed relationships with other characters in the game by making voluntary choices in terms of power and affiliation, in absence of linguistic metaphors. They found that the hippocampus tracked the social coordinates defined by power and affiliation, when subjects “move” in this social space by making voluntary choices. Hippocampus is crucial for coding spatial and temporal context of events (Eichenbaum and Cohen 2014). Absolute social distance was tracked by posterior cingulate, which is associated with spatial navigation. This suggests that abstract concepts of power and affiliation, and the resulting social relationship, may be represented in a spatiotemporal format even without any explicit use of linguistic metaphors.
6. Conclusions
Abstract concepts can be understood by relating them metaphorically to concrete domains. Evidence from brain imaging, brain stimulation, and lesion studies, on balance, shows that when abstract concepts are expressed as linguistic metaphors to sensory-motor domains, they are understood in terms of those domains. Metaphors are embodied and grounded in sensory-motor domains. Such metaphors are understood not through a polysemous sense that is abstract, but through simulations in action-perception systems at varying levels of detail. Processing of these metaphors is not identical to that of literal concrete language, however, but also retains an abstract component. Abstract concepts may have independent representations that are shaped and enriched by metaphors.
The case of idioms is less clear. Neuroimaging evidence suggests that at least, sensory-motor systems are not likely to be activated in the same way or to the same extent for idioms as they are for metaphors or for literal sentences. Based on preliminary evidence, I suggest that idioms may still be grounded, but in a qualitatively different manner than metaphors. They could be grounded through more schematic action planning, control, and goal circuits that sit higher in the action hierarchy in the basal ganglia.
Finally, there is neural evidence that some abstract concepts and relations, such as power and affiliation, have a metaphoric organization even in the absence of linguistic metaphors. Some components of the abstract conceptual system may indeed have underlying metaphoric basis, at least under certain conditions. Fleshing out the bounds of this organization -- for example, the precise types of abstract concepts that are organized in this way, their flexibility, and the necessary conditions and context -- awaits further investigation.
Figure 3.
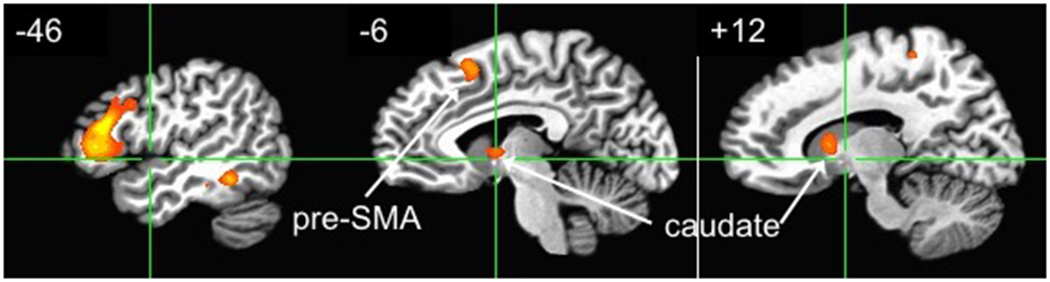
Activations in IDI > ABS contrast in data from Desai et al. (2013), at voxelwise p < 0.005. Frontal and pre-SMA activations are significant in whole-brain analysis, while the right caudate activation is significant in an ROI analysis. Numbers indicate Talairach x-coordinates of the slice (negative is left hemisphere).
Funding:
This work was supported by NIH/NIDCD grants R01DC010783, R56DC010783, and R01DC017162.
Footnotes
This is not meant to imply that other theories, such as the amodal symbol view, do in fact account for abstract concepts. Whether other theories can account either concrete or abstract concepts is indeed not clear, but this is a separate question that will not be addressed here.
Abstract sentences are also “literal” in that they are non-figurative. Here, the term LIT is reserved for non-figurative sentences that denote a concrete sensory-motor event, such as “throwing a ball.” ABS is used for literal sentences that denote events that do not directly entail action/perception, such as “considering my options.”
Many neuroimaging studies present sentence stimuli visually that are read silently. Note that this does not obviate the importance of controlling for phonology. Activation of phonological representations during reading is automatic and rapid (and similarly, orthography is activated to some extent by auditory word presentations).
This activation could be seen only due to the whole-brain analysis used in the study. If the analyses were targeted only to specific action ROIs, this area relevant to abstract semantics and with interesting theoretical implications would not be seen. This provides an example of the potential value of whole-brain analyses.
They used the term ‘literal’ for this condition. Under the current nomenclature, such conditions are termed ABS.
Conflict of Interest: Author Rutvik H. Desai declares that he has no conflict of interest.
Ethical approval: This article does not contain any studies with human participants or animals performed by any of the authors.
References
- Akkal D, Dum RP, and Strick PL. 2007. ‘Supplementary motor area and presupplementary motor area: targets of basal ganglia and cerebellar output’, J Neurosci, 27: 10659–73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aziz-Zadeh L, Wilson SM, Rizzolatti G, and Iacoboni M. 2006. ‘Congruent embodied representations for visually presented actions and linguistic phrases describing actions’, Curr Biol, 16: 1818–23. [DOI] [PubMed] [Google Scholar]
- Barsalou LW 2008. ‘Grounded cognition’, Annu Rev Psychol, 59: 617–45. [DOI] [PubMed] [Google Scholar]
- Barsalou LW, and Wiemer-Hastings K. 2005. ‘Situating abstract concepts.’ in Pecher D and Zwaan RA (eds.), Grounding cognition: The role of perception and action in memory, language, and thought (Cambridge University Press: New York: ). [Google Scholar]
- Benjamini Y, and Hochberg Y. 1995. ‘Controlling the false discovery rate: a practical and powerful approach to multiple testing’, Journal of Royal Statistical Society B, 57: 298–300. [Google Scholar]
- Binkofski Ferdinand Christoph, Klann Juliane, and Caspers Svenja. 2016. ‘On the Neuroanatomy and Functional Role of the Inferior Parietal Lobule and Intraparietal Sulcus.’ in, Neurobiology of Language. [Google Scholar]
- Bogacz R, Wagenmakers EJ, Forstmann BU, and Nieuwenhuis S. 2010. ‘The neural basis of the speed-accuracy tradeoff’, Trends Neurosci, 33: 10–6. [DOI] [PubMed] [Google Scholar]
- Borghi AM, Barca L, Binkofski F, Castelfranchi C, Pezzulo G, and Tummolini L. 2018. ‘Words as social tools: Language, sociality and inner grounding in abstract concepts’, Phys Life Rev. [DOI] [PubMed] [Google Scholar]
- ———. 2019. ‘Words as social tools: Language, sociality and inner grounding in abstract concepts’, Phys Life Rev, 29: 120–53. [DOI] [PubMed] [Google Scholar]
- Boulenger V, Hauk O, and Pulvermuller F. 2009. ‘Grasping ideas with the motor system: semantic somatotopy in idiom comprehension’, Cereb Cortex, 19: 1905–14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boulenger V, Shtyrov Y, and Pulvermuller F. 2012. ‘When do you grasp the idea? MEG evidence for instantaneous idiom understanding’, Neuroimage, 59: 3502–13. [DOI] [PubMed] [Google Scholar]
- Button Katherine S., Ioannidis John P. A., Mokrysz Claire, Brian a. Nosek, Jonathan Flint, Emma S. J. Robinson, and Marcus R. Munafò. 2013. ‘Power failure: why small sample size undermines the reliability of neuroscience’, Nature Reviews Neuroscience, 14. [DOI] [PubMed] [Google Scholar]
- Buxbaum LJ, Kyle K, Grossman M, and Coslett HB. 2007. ‘Left inferior parietal representations for skilled hand-object interactions: evidence from stroke and corticobasal degeneration’, Cortex, 43: 411–23. [DOI] [PubMed] [Google Scholar]
- Buxbaum LJ, Johnson-Frey SH, and Bartlett-Williams M. 2005. ‘Deficient internal models for planning hand–object interactions in apraxia’, Neuropsychologia, 43: 917–29. [DOI] [PubMed] [Google Scholar]
- Cacciari C, Bolognini N, Senna I, Pellicciari MC, Miniussi C, and Papagno C. 2011. ‘Literal, fictive and metaphorical motion sentences preserve the motion component of the verb: a TMS study’, Brain Lang, 119: 149–57. [DOI] [PubMed] [Google Scholar]
- Cacciari C, and Tabossi P. 1988. ‘The comprehension of idioms’, Journal of Memory and Language, 27: 668–83. [Google Scholar]
- Cacciola A, Milardi D, Bertino S, Basile GA, Calamuneri A, Chillemi G, Rizzo G, Anastasi G, and Quartarone A. 2019. ‘Structural connectivity-based topography of the human globus pallidus: Implications for therapeutic targeting in movement disorders’, Mov Disord, 34: 987–96. [DOI] [PubMed] [Google Scholar]
- Casasanto D 2009. ‘When is a lingusitic metaphor a conceptual metaphor.’ in Vyvyan Evans and Stephanie Pourcel (eds.), New Directions in Cognitive Linguistics (John Benjamins: Philadelphia: ). [Google Scholar]
- Casasanto D, and Bottini R. 2014. ‘Spatial language and abstract concepts’, Wiley Interdiscip Rev Cogn Sci, 5: 139–49. [DOI] [PubMed] [Google Scholar]
- Casasanto Daniel, and Gijssels Tom. 2015. ‘What makes a metaphor an embodied metaphor?’, Linguistics Vanguard, 1. [Google Scholar]
- Chen E, Widick P, and Chatterjee A. 2008. ‘Functional-anatomical organization of predicate metaphor processing’, Brain Lang, 107: 194–202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Citron FM, and Goldberg AE. 2014. ‘Metaphorical sentences are more emotionally engaging than their literal counterparts’, J Cogn Neurosci, 26: 2585–95. [DOI] [PubMed] [Google Scholar]
- Cremers HR, Wager TD, and Yarkoni T. 2017. ‘The relation between statistical power and inference in fMRI’, PLoS ONE, 12: e0184923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desai RH, Conant LL, Binder JR, Park H, and Seidenberg MS. 2013. ‘A piece of the action: modulation of sensory-motor regions by action idioms and metaphors’, Neuroimage, 83: 862–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desai RH, Reilly M, and van Dam W. 2018. ‘The multifaceted abstract brain’, Philos Trans R Soc Lond B Biol Sci, 373: 20170122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desai Rutvik H, Jeffrey R Binder, Lisa L Conant, Quintino R Mano, and Mark S Seidenberg. 2011. ‘The Neural Career of Sensory-motor Metaphors.’, Journal of Cognitive Neuroscience, 23: 2376–86. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Desmond JE, and Glover GH. 2002. ‘Estimating sample size in functional MRI (fMRI) neuroimaging studies: statistical power analyses’, J Neurosci Methods, 118: 115–28. [DOI] [PubMed] [Google Scholar]
- Dove G 2009. ‘Beyond perceptual symbols: a call for representational pluralism’, Cognition, 110: 412–31. [DOI] [PubMed] [Google Scholar]
- Eichenbaum H, and Cohen NJ. 2014. ‘Can we reconcile the declarative memory and spatial navigation views on hippocampal function?’, Neuron, 83: 764–70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandez L, Huys R, Issartel J, Azulay JP, and Eusebio A. 2018. ‘Movement Speed-Accuracy Trade-Off in Parkinson’s Disease’, Front Neurol, 9: 897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandino L, Binder JR, Desai RH, Pendl SL, Humphries CJ, Gross WL, Conant LL, and Seidenberg MS. 2016. ‘Concept Representation Reflects Multimodal Abstraction: A Framework for Embodied Semantics’, Cereb Cortex, 26: 2018–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fernandino L, Conant LL, Binder JR, Blindauer K, Hiner B, Spangler K, and Desai RH. 2013. ‘Where is the action? Action sentence processing in Parkinson’s disease’, Neuropsychologia, 51: 1510–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galea MP, and Darian-Smith I. 1994. ‘Multiple corticospinal neuron populations in the macaque monkey are specified by their unique cortical origins, spinal terminations, and connections’, Cereb Cortex, 4: 166–94. [DOI] [PubMed] [Google Scholar]
- Gibbs Jr., Raymond W. 1996. ‘Why many concepts are metaphorical’, Cognition, 61: 309–19. [DOI] [PubMed] [Google Scholar]
- Gibbs Raymond W. 2011. ‘Evaluating Conceptual Metaphor Theory’, Discourse Processes, 48: 529–62. [Google Scholar]
- Gibbs RW. 1994a. ‘Figurative thought and figurative language.’ in Gernsbacher MA (ed.), Handbook of Psycholinguistics (Academic PRess: San Deigo, CA: ). [Google Scholar]
- ———. 1994b. The poetics of mind : figurative thought, language, and understanding (Cambridge University Press: Cambridge [England] ; New York, NY, USA: ). [Google Scholar]
- Goldenberg G, and Karnath HO. 2006. ‘The neural basis of imitation is body part specific’, J Neurosci, 26: 6282–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grahn JA, Parkinson JA, and Owen AM. 2008. ‘The cognitive functions of the caudate nucleus’, Prog Neurobiol, 86: 141–55. [DOI] [PubMed] [Google Scholar]
- Graybiel AM. 1998. ‘The basal ganglia and chunking of action repertoires’, Neurobiology of Learning and Memory, 70: 119–36. [DOI] [PubMed] [Google Scholar]
- Guo Y, Schmitz TW, Mur M, Ferreira CS, and Anderson MC. 2018. ‘A supramodal role of the basal ganglia in memory and motor inhibition: Meta-analytic evidence’, Neuropsychologia, 108: 117–34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haaland KY, Harrington DL, and Knight RT. 2000. ‘Neural representations of skilled movement’, Brain, 123: 2306–13. [DOI] [PubMed] [Google Scholar]
- Haber SN. 2016. ‘Corticostriatal circuitry.’ in Pfaff DW and Volkow N (eds.), Neuroscience in the 21st Century: From Basic to Clinical (Springer: New York, NY: ). [Google Scholar]
- Hamblin JL, and Gibbs RW Jr. 1999. ‘Why you can’t kick the bucket as you slowly die: Verbs in idiom comprehension’, Journal of Psycholinguistic Research, 28: 25–39. [Google Scholar]
- Jamrozik A, McQuire M, Cardillo ER, and Chatterjee A. 2016. ‘Metaphor: Bridging embodiment to abstraction’, Psychon Bull Rev. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jax SA, Buxbaum LJ, and Moll AD. 2006. ‘Deficits in movement planning and intrinsic coordinate control in ideomotor apraxia’, J Cogn Neurosci, 18: 2063–76. [DOI] [PubMed] [Google Scholar]
- Jin X, Tecuapetla F, and Costa RM. 2014. ‘Basal ganglia subcircuits distinctively encode the parsing and concatenation of action sequences’, Nat Neurosci, 17: 423–30. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johari K, Riccardi N, Malyutina S, Modi M, and Desai RH. 2021. ‘HD-tDCS over motor cortex facilitates figurative and literal action sentence processing’, Neuropsychologia, 159: 107955. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnson-Frey SH, Newman-Norlund R, and Grafton ST. 2005. ‘A distributed left hemisphere network active during planning of everyday tool use skills’, Cereb Cortex, 15: 681–95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kiefer M, and Pulvermuller F. 2012. ‘Conceptual representations in mind and brain: theoretical developments, current evidence and future directions’, Cortex, 48: 805–25. [DOI] [PubMed] [Google Scholar]
- Kiefer Markus, and Harpaintner Marcel. 2020. ‘Varieties of abstract concepts and their grounding in perception or action’, Open Psychology, 2: 119–37. [Google Scholar]
- Kim HF, and Hikosaka O. 2015. ‘Parallel basal ganglia circuits for voluntary and automatic behaviour to reach rewards’, Brain, 138: 1776–800. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhnke P, Kiefer M, and Hartwigsen G. 2020. ‘Task-Dependent Recruitment of Modality-Specific and Multimodal Regions during Conceptual Processing’, Cereb Cortex, 30: 3938–59. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lacey S, Stilla R, Deshpande G, Zhao S, Stephens C, McCormick K, Kemmerer D, and Sathian K. 2017. ‘Engagement of the left extrastriate body area during body-part metaphor comprehension’, Brain Lang, 166: 1–18. [DOI] [PubMed] [Google Scholar]
- Lacey S, Stilla R, and Sathian K. 2012. ‘Metaphorically feeling: comprehending textural metaphors activates somatosensory cortex’, Brain Lang, 120: 416–21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai VT, and Desai RH. 2016. ‘The grounding of temporal metaphors’, Cortex, 76: 43–50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai VT, Howerton O, and Desai RH. 2019. ‘Concrete processing of action metaphors: Evidence from ERP’, Brain Res, 1714: 202–09. [DOI] [PubMed] [Google Scholar]
- Lakoff G 1993. ‘The contemporary theory of metaphor.’ in Ortony A (ed.), Metaphor and Thought (Cambridge University Press; ). [Google Scholar]
- Lakoff G, and Mark Johnson. 1980. Metaphors We Live By (University of Chicago Press: Chicago: ). [Google Scholar]
- ———. 1999. Philosophy in the Flesh: The Embodied Mind and its Challenge to Western Thought (Basic Books: New York: ). [Google Scholar]
- Lehericy S, Bardinet E, Tremblay L, Van de Moortele PF, Pochon JB, Dormont D, Kim DS, Yelnik J, and Ugurbil K. 2006. ‘Motor control in basal ganglia circuits using fMRI and brain atlas approaches’, Cereb Cortex, 16: 149–61. [DOI] [PubMed] [Google Scholar]
- Markowitz JE, Gillis WF, Beron CC, Neufeld SQ, Robertson K, Bhagat ND, Peterson RE, Peterson E, Hyun M, Linderman SW, Sabatini BL, and Datta SR. 2018. ‘The Striatum Organizes 3D Behavior via Moment-to-Moment Action Selection’, Cell, 174: 44–58 e17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McGlone Matthew S. 2007. ‘What is the explanatory value of a conceptual metaphor?’, Language & Communication, 27: 109–26. [Google Scholar]
- –––. 2011. ‘Hyperbole, Homunculi, and Hindsight Bias: An Alternative Evaluation of Conceptual Metaphor Theory’, Discourse Processes, 48: 563–74. [Google Scholar]
- Milardi D, Quartarone A, Bramanti A, Anastasi G, Bertino S, Basile GA, Buonasera P, Pilone G, Celeste G, Rizzo G, Bruschetta D, and Cacciola A. 2019. ‘The Cortico-Basal Ganglia-Cerebellar Network: Past, Present and Future Perspectives’, Front Syst Neurosci, 13: 61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy GL 1996. ‘On metaphoric representation’, Cognition, 60: 173–204. [DOI] [PubMed] [Google Scholar]
- –––. 1997. ‘Reasons to doubt the present evidence for metaphoric representation’, Cognition, 62: 99–108. [DOI] [PubMed] [Google Scholar]
- Murphy K, and Garavan H. 2004. ‘An empirical investigation into the number of subjects required for an event-related fMRI study’, Neuroimage, 22: 879–85. [DOI] [PubMed] [Google Scholar]
- Nachev P, Wydell H, O’Neill K, Husain M, and Kennard C. 2007. ‘The role of the pre-supplementary motor area in the control of action’, Neuroimage, 36 Suppl 2: T155–63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nachev Parashkev, Kennard Christopher, and Husain Masud. 2008. ‘Functional role of the supplementary and pre-supplementary motor areas’, Nature reviews. Neuroscience, 9: 856–69. [DOI] [PubMed] [Google Scholar]
- Orban GA, and Caruana F. 2014. ‘The neural basis of human tool use’, Front Psychol, 5: 310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pajula J, and Tohka J. 2016. ‘How Many Is Enough? Effect of Sample Size in Inter-Subject Correlation Analysis of fMRI’, Comput Intell Neurosci, 2016: 2094601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peeters Ronald R, Giacomo Rizzolatti, and Guy a Orban. 2013. ‘Functional properties of the left parietal tool use region.’, Neuroimage, 78: 83–93. [DOI] [PubMed] [Google Scholar]
- Poldrack RA 2006. ‘Can cognitive processes be inferred from neuroimaging data?’, Trends in Cognitive Sciences, 10: 59–63. [DOI] [PubMed] [Google Scholar]
- Pollio Howard R. 1977. Psychology and the poetics of growth : figurative language in psychology, psychotherapy, and education (L. Erlbaum Associates ;distributed by the Halsted Press Division of J. Wiley: Hillsdale, N.J. New York: ). [Google Scholar]
- Quadflieg Susanne, Joset a Etzel, Valeria Gazzola, Christian Keysers, Schubert Thomas W, Waiter Gordon D, and Macrae C Neil. 2011. ‘Puddles, Parties, and Professors: Linking Word Categorization to Neural Patterns of Visuospatial Coding.’, Journal of Cognitive Neuroscience: 2636–49. [DOI] [PubMed] [Google Scholar]
- Ramayya Ashwin G, Matthew F Glasser, and James K Rilling. 2009. ‘A DTI Investigation of Neural Substrates Supporting Tool Use.’, Cerebral cortex (New York, N.Y. : 1991): 507–16. [DOI] [PubMed] [Google Scholar]
- Randerath J, Valyear KF, Philip BA, and Frey SH. 2017. ‘Contributions of the parietal cortex to increased efficiency of planning-based action selection’, Neuropsychologia. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raposo A, Moss HE, Stamatakis EA, and Tyler LK. 2009. ‘Modulation of motor and premotor cortices by actions, action words and action sentences’, Neuropsychologia, 47: 388–96. [DOI] [PubMed] [Google Scholar]
- Reilly M, Howerton O, and Desai RH. 2019. ‘Time-Course of Motor Involvement in Literal and Metaphoric Action Sentence Processing: A TMS Study’, Front Psychol, 10: 371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riva D, Taddei M, and Bulgheroni S. 2018. ‘The neuropsychology of basal ganglia’, Eur J Paediatr Neurol, 22: 321–26. [DOI] [PubMed] [Google Scholar]
- Rizzolatti G, and Luppino G. 2001. ‘The cortical motor system’, Neuron, 31: 889–901. [DOI] [PubMed] [Google Scholar]
- Lauro Romero, Leonor J Giulia Mattavelli, Papagno Costanza, and Tettamanti Marco. 2013. ‘She runs, the road runs, my mind runs, bad blood runs between us: Literal and figurative motion verbs: An fMRI study.’, Neuroimage, 83C: 361–71. [DOI] [PubMed] [Google Scholar]
- Sanford D 2014. ‘Idiom as the intersection of conceptual and syntactic schemas’, Language and Cognition, 6: 492–509. [Google Scholar]
- Saygin Ayse Pinar, Stephen McCullough, Morana Alac, and Emmorey Karen. 2010. ‘Modulation of BOLD Response in Motion-sensitive Lateral Temporal Cortex by Real and Fictive Motion Sentences.’, Journal of Cognitive Neuroscience, 22: 2480–90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swinney D, and Cutler A. 1979. ‘The access and processing of idiomatic expressions’, Journal of Verbal Learning and Verbal Behavior, 18: 523–34. [Google Scholar]
- Tavares RM, Mendelsohn A, Grossman Y, Williams CH, Shapiro M, Trope Y, and Schiller D. 2015. ‘A Map for Social Navigation in the Human Brain’, Neuron, 87: 231–43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Titone DA, and Connine CM. 1999. ‘On the compositional and noncompositional nature of idiomatic expressions’, Journal of Pragmatics, 31: 1655–74. [Google Scholar]
- Tunik E, Frey SH, and Grafton ST. 2005. ‘Virtual lesions of the anterior intraparietal area disrupt goal-dependent on-line adjustments of grasp’, Nat Neurosci, 8: 505–11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tunik E, Lo OY, and Adamovich SV. 2008. ‘Transcranial magnetic stimulation to the frontal operculum and supramarginal gyrus disrupts planning of outcome-based hand-object interactions’, J Neurosci, 28: 14422–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Turner BO, Paul EJ, Miller MB, and Barbey AK. 2018. ‘Small sample sizes reduce the replicability of task-based fMRI studies’, Commun Biol, 1: 62. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varney NR, and Damasio H. 1987. ‘Locus of lesion in impaired pantomime recognition’, Cortex, 23: 699–703. [DOI] [PubMed] [Google Scholar]
- Villani C, Lugli L, Liuzza MT, and Borghi AM. 2019. ‘Varieties of abstract concepts and their multiple dimensions’, Language and Cognition, 11: 403–30. [Google Scholar]
- Vingerhoets G 2014. ‘Contribution of the posterior parietal cortex in reaching, grasping, and using objects and tools’, Front Psychol, 5: 151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vulchanova Mila, Milburn Evelyn, Vulchanov Valentin, and Baggio Giosuè. 2019. ‘Boon or Burden? The Role of Compositional Meaning in Figurative Language Processing and Acquisition’, Journal of Logic, Language and Information, 28: 359–87. [Google Scholar]
- Wallentin M, Lund TE, Ostergaard S, Ostergaard L, and Roepstorff A. 2005. ‘Motion verb sentences activate left posterior middle temporal cortex despite static context’, Neuroreport, 16: 649–52. [DOI] [PubMed] [Google Scholar]
- Wallentin M, Østergaarda S, Lund TE, Østergaard L, and Roepstorff A. 2005. ‘Concrete spatial language: See what I mean?’, Brain and Language, 92: 221–33. [DOI] [PubMed] [Google Scholar]
- Wang Jing, Julie a Conder, David N Blitzer, and Svetlana V Shinkareva. 2010. ‘Neural representation of abstract and concrete concepts: A meta-analysis of neuroimaging studies.’, Human brain mapping, 31: 1459–68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiener Martin, Turkeltaub Peter, and Coslett HB. 2010. ‘The image of time: A voxel-wise meta-analysis’, Neuroimage, 49: 1728–40. [DOI] [PubMed] [Google Scholar]
- Zanolie K, Sv Dantzig I. Boot, Wijnen J, Schubert TW, Giessner SR, and Pecher D. 2012. ‘Mighty metaphors: behavioral and ERP evidence that power shifts attention on a vertical dimension’, Brain Cogn, 78: 50–8. [DOI] [PubMed] [Google Scholar]