

Abstract
Messenger RNA (mRNA) holds great potential in developing immunotherapy, protein replacement, and genome editing. In general, mRNA does not have the risk of being incorporated into the host genome and does not need to enter the nucleus for transfection, and it can be expressed even in nondividing cells. Therefore, mRNA‐based therapeutics provide a promising strategy for clinical treatment. However, the efficient and safe delivery of mRNA remains a crucial constraint for the clinical application of mRNA therapeutics. Although the stability and tolerability of mRNA can be enhanced by directly retouching the mRNA structure, there is still an urgent need to improve the delivery of mRNA. Recently, significant progress has been made in nanobiotechnology, providing tools for developing mRNA nanocarriers. Nano‐drug delivery system is directly used for loading, protecting, and releasing mRNA in the biological microenvironment and can be used to stimulate the translation of mRNA to develop effective intervention strategies. In the present review, we summarized the concept of emerging nanomaterials for mRNA delivery and the latest progress in enhancing the function of mRNA, primarily focusing on the role of exosomes in mRNA delivery. Moreover, we outlined its clinical applications so far. Finally, the key obstacles of mRNA nanocarriers are emphasized, and promising strategies to overcome these obstacles are proposed. Collectively, nano‐design materials exert functions for specific mRNA applications, provide new perception for next‐generation nanomaterials, and thus revolution of mRNA technology.
Keywords: Exosomes, Gene editing, Gene therapy, mRNA delivery, mRNA vaccine, Nanocarriers
1. INTRODUCTION
As intermediate tunneling, messenger ribonucleic acid (mRNA) can transport genetic codes from DNA to ribosomes for protein expression, which has emerged as a highly appealing potential to change the vaccination, protein replacement therapy, and other treatments for human diseases. 1 , 2 , 3 , 4 The first study of successful mRNA therapeutics in the preclinical application was published in 1990. 5 In addition, due to the progress of mRNA manufacturing and intercellular delivery strategy, significant development and breakthrough have been made in the mRNA‐based therapy. 6 In addition, mRNA‐based therapeutics exhibit several advantages over traditional functional biomolecules and therapeutic proteins. 7 First, mRNA is endowed with perfect safety under physiological conditions because it is not transferred into the nucleus and does not integrate into the host genome. Therefore, mRNA‐based therapeutics are transient, avoiding the potential risk of insertional mutagenesis. 8 , 9 Second, the initiation of protein translation is promoted once mRNA just reaches the cytoplasm. 10 Third, these theranostic mRNAs can achieve stability and controlled release through mRNA structural modification (such as 5′‐Cap, 5′‐UTR, ORF, 3′‐UTR, and 3′‐poly(A) tail) or nanomaterials delivery, such as lipid‐derived nanoparticles (LNPs), polymer‐based nanoparticles, and hybrid nanoparticle, thus improving the pharmacokinetics of nucleic acid drugs. 11 , 12 , 13 Fourth, it is easy to design and manufacture mRNA using chemical synthesis and enzyme synthesis approaches. 14 Solid‐phase chemical synthesis is generally suitable for the synthesis of short‐chain RNAs (50–100 nt). Moreover, the synthesis yield using this method remains poor and impure with the increase of RNA length. 15 Recently, enzyme synthesis is a popular method based on RNA polymerase (usually T7) and linear DNA to obtain desired mRNA. However, the initial product of mRNA through this method is composed of targeted mRNA, untargeted mRNA, and oligodexynucleotides, which need to be purified for further application. 16
Despite its potential advantages, how to efficiently and stably perform intracellular delivery of mRNA is a significant barrier in the clinical application of mRNA as a therapeutic modality. 14 , 17 , 18 , 19 Additionally, naked mRNA molecules are easily degraded by enzymes and are not easily taken up by the target cells. 20 Therefore, methods and vehicles for mRNA delivery, such as viral vectors, mechanical transfection, and nonviral vectors, have been developed. 11 , 21 , 22 Viral vectors are very efficiency in mRNA transfection and are applied in clinical trials, such as Kymriah for chimeric antigen receptor (CAR) T immunotherapy on lymphoblastic leukemia. 23 , 24 However, viral vectors have disadvantages of potential carcinogenic, high immunogenic, limited gene packaging ability, and low‐volume production. 25 , 26 Mechanical transfection, such as injection of naked mRNA in conventional and self‐amplifying forms, is not widely applied due to their extracellular exonucleases, inefficient cell uptake, unsuccessful endosomal release, or potential cytotoxicity. 27 Nonviral vehicles, including lipid or polymer‐based nanoparticles, exosomes, and ligand‐RNA conjugates, represent safe and efficacious technologies and allow repeated administrations. 28 , 29 Moreover, lipid‐based nanoparticles have been the most extensively used for mRNA‐based therapeutics in preclinical studies and undergoing clinical trials (Table 1). 49 , 50 In addition, some nonviral vehicles possess targeting effects at desired sites in mRNA‐based therapeutics because of their organ‐targeted properties. 50 Furthermore, nonviral nanoparticles are also substantially attractive in mRNA vaccines for virus outbreaks. 51
TABLE 1.
List of nanoparticle mRNA‐based vaccines in clinical trials
| Nanocarrier | Disease | Names | General information | Clinical trials no./references |
|---|---|---|---|---|
| Cancer immunotherapy | ||||
| Liposomes (1,2‐di‐O‐octa decenyl‐3‐trimethyl‐ammonium‐propane (DOTAP), 1,2‐dioleoyl‐sn‐glycero‐3‐phosphoethanolamine (DSPC) | Melanoma | Lipo‐MERIT | Phase I, enrollment, completed, intravenously (IV) administered, elicit of T‐cell responses against malignant melanoma‐associated antigens | NCT02410733 30 |
| Liposomes | Triple‐negative breast cancer | TNBC‐MERIT | Phase I, active, not recruiting | NCT02316457 31 |
| Liposome | Ovarian cancer | W_ova1 vaccine | Phase I, active, not recruiting, IV injection | NCT04163094 |
| LNPs | Colorectal cancer | RO7198457 | Phase I, active, not recruiting, induction of both T‐lymphocyte‐ and memory T‐cell‐dependent immune responses against cancer ‐associated antigens | |
| LNPs | Melanoma | Lipo‐MERIT | Phase I, active, not recruiting, IV injection, elicited T‐cell responses and tumor‐associated antigen | NCT02410733 32 |
| LNPs | KRAS mutant or metastatic cancer | mRNA‐5671/V941 | Phase I/II, completed, intramuscular (IM) injection combination therapy | NCT03948763 |
| LNPs | Metastatic melanoma, epithelial cancer, or gastrointestinal cancer | mRNA‐4650 | Phase I/II, completed, IM injections, intramuscular safety, induced neoantigen‐specific T‐cell response | NCT03480152 32 |
| LNPs | Metastatic HPV16‐related squamous cell carcinoma of head and neck expressing PD‐L1 | BNT113 | Phase I, recruiting | NCT04534205 33 |
| LNPs |
Melanoma Solid tumors |
mRNA‐4157 | Phase II, active, IM Injection, well tolerated | |
| LNPs | Unmodified Melanoma | BNT111 | Phase II, recruiting IV injection | NCT04526899 35 |
| LNPs: | Relapsed lymphoma or malignancies | mRNA‐2416 | Phase I, cessation, intratumoral injection | NCT03323398 |
| LNPs | Solid tumor malignancies or lymphoma | mRNA‐2752 | Phase I, recruiting, intratumoral injection | NCT03739931 |
| Nanocarrier | Disease | Vaccination names | General Information | Clinical trials no./references |
|---|---|---|---|---|
| Prevent viral infection | ||||
| LNPs | Rabies virus | CV7202 (Rabies virus glycoprotein‐mRNA vaccine) | Phase I, completed, IM injection, neutralizing antibody responses; well tolerated | NCT03713086 36 |
| LNPs | Rabies virus | CV7201 (sequence‐modified mRNA of rabies virus glycoprotein) | Phase I, recruited, completed needle injection acceptable tolerability and generated rabies neutralizing antibody responses |
NCT03713086 36 , 37 |
| LNPs | Human papilloma virus | HARE‐40 | Phase I/II, recruiting, intradermal injection | NCT03418480 38 |
| LNPs | Human metapneumovirus and parainfluenza | mRNA‐1653 vaccine (nucleoside‐modified mRNA) | Phase I, completed IM injection | NCT03392389 |
| LNPs | Cytomegalovirus Infection | mRNA‐1443, mRNA‐1647 | Phase I, recruited, completed induce humoral and cellular immune response | |
| LNPs | Chikungunya virus | mRNA‐1388 | Phase I, recruited, completed | NCT03325075 |
| LNPs | Zika virus | mRNA‐1325, mRNA‐1893 | Phase I/II, enrollment, completed well‐tolerated, induced a neutralizing antibody response | |
| LNPs | Influenza viruses H10N8 and H7N9 | mRNA‐1440 vaccine (nucleoside‐modified) | Phase I, enrollment, completed IM Injection reasonable safety, tolerability, stimulate humoral immune response | NCT03076385 40, 41 |
| LNPs | Influenza H7N9 | mRNA‐1851(H10N8 or H7N9 mRNA) | Phase I, completed IM injection well tolerability and immunogenicity | NCT03345043 40 |
| LNPs | Respiratory syncytial virus (RSV), SARS‐CoV‐2 | mRNA‐1345 | Phase III, active, not recruiting IM injection induce protective immunogenicity | |
| LNPs: ionizable lipid, cholesterol, DSPC, PEG2000‐DM | COVID‐19 | mRNA vaccine (full‐length spike protein) | Phase I, completed IM injection induce neutralize antibody against SARS‐CoV‐2 infection | NCT04758962 42 |
| LNPs: ionizable amino lipid, PEGylated lipid, cholesterol, phospholipid |
COVID‐19 SARS‐CoV‐2 |
mRNA vaccine (full‐length prefusion spike protein of SARS‐CoV‐2 | Phase I, terminated IM injection induced significant humoral immune responses | NCT04860258 43 |
| LNPs: ALC‐0315, ALC‐0159, DSPC, cholesterol |
COVID‐19 SARS‐CoV‐2 |
BNT162b2 (spike protein) | Phase III, completed, IM injection | NCT04816669 44 |
| LNPs: PEG2000‐lipid PEG2000‐DMG, DSPC, cholesterol | LUNAR‐COVID‐19 | ARCT‐021 | Phase II, terminated, IM injection | NCT04728347 |
| LNPs: PEG‐lipid, cholesterol, 1,2‐dimyristoyl‐rac‐glycero‐3‐methoxypolyethylene glycol‐2000 (DMG‐PEG), DSPC, ionizable lipid | COVID‐19 | mRNA vaccine (spike protein of SARS‐CoV‐2 | Phase III, recruiting, approved by FDA, IM injection safety and well tolerability protection against SARS‐CoV‐2 infection | NCT04847102 45 |
| LNPs (ionizable lipid DSPC, cholesterol, DMG‐PEG) | SARS‐CoV‐2 |
mRNA‐1273 vaccine (spike protein mRNA vaccine) BNT162 |
Phase III, active, recruiting, approved by FDA, IM injection, 94.1% efficacy in protection against COVID‐19, feasible to storage and delivery |
NCT04860297 46 , 47 , 48 |
| Nanocarrier | Disease | Vaccination names | General information | Clinical trials no./references |
|---|---|---|---|---|
| mRNA for protein replacement | ||||
| LNPs | Ornithine transcarbamylase (OTC) deficiency |
MRT5201 ARCT‐810 (human OTC protein) |
Phase I/II, withdrawn, intravenous (IV) infusion, adequate enzyme replacement for correct OTC deficiency | |
| LNPs | Methylmalonic acidemia (methylmalonyl‐coenzyme A mutase (MUT) deficiency) | mRNA‐3927 | Phase I/II, terminated, IV infusion | NCT03810690 |
| LNPs (lipid, cholesterol, PEG2000, DSPC) | Propionic acidemia | mRNA 3927 (α and β subunits of mitochondrial propionyl‐CoA carboxylase protein) | Phase I/II, recruiting, IV infusion | NCT04159103 |
| LNPs (Poly (β‐amino esters, PEG2000, DOPE, cholesterol) | Cystic fibrosis (CF) | MRT5005 (human CF transmembrane regulator protein) | Phase I/II, recruiting, aerosol inhalation | NCT03375047 |
Here, we reviewed the latest developments in mRNA‐based therapeutics, such as vaccination, genome editing, and protein replacement therapy. Moreover, we also emphasized the advanced delivery platforms for mRNA and their biomedical applications. In addition, we discussed the potential challenges of mRNA‐based therapeutics and provided our future perspectives.
2. mRNA FOR THERAPEUTIC APPLICATIONS
Over the past decades, mRNA‐based therapeutics have been highly promising for treating various human diseases, including cancers, infections, and autoimmune diseases. 52 , 53 , 54 Recently, mRNA‐based vaccines have become a promising alternative in booster immunizations. 1 , 55 In addition, mRNA‐based protein replacement therapy provides a viable alternative and permanent cure for various diseases. 56 , 57 In 1961, as an intermediate hereditary substance, mRNA is first discovered. 58 As early as 1990, scientists have first reported that intramuscular injection of in vitro‐transcribed mRNA into mouse skeletal muscle cells, results in in vivo expression of encoding proteins. Since then, mRNA‐based therapeutics have been exploited in a variety of application. 5 In 2002, the first clinical trial used ex vivo mRNA to transfect dendritic cells (DCs), aiming to stimulate cytotoxic T lymphocyte against cancer. 59 In 2020, mRNA‐based vaccines were approved against COVID‐19 by the U.S Food and Drug Administration (FDA). 60
mRNA‐based therapeutics are composed of two main elements: the mRNA itself and the delivery systems. Most eukaryotic mRNAs have a tripartite structure comprised a 5′ cap structure, a 5′ untranslated region (5′ UTR), the open reading frame (ORF), a 3′ UTR, and a poly (A)‐tail to the 3′ end. The 5′ cap and 3′ poly a tail can protect mRNA from degradation by exonuclease, help bind to ribosomes, and initiate splicing, maturation, and translation. 61 , 62 The 5′ UTR and 3′ UTR are located at the upstream and downstream domains of the mRNA coding region, respectively, and are essential to protect the mRNA from decomposition and degradation. 63 Moreover, the 5′ UTR plays a crucial role in controlling protein translation efficiency due to its binding site for the preinitiation complex initiation of protein translation. 64 In addition, 3′ UTR contains degradation signals of mRNA, and alpha‐globin or beta‐globin 3′ UTR has been developed to improve the stability of heterologous mRNA in cells. 65 The ORF region contains protein‐encoding nucleotide sequences with a start codon and ends with a stop codon. 66 For the production of mRNA, the in vitro transcription (IVT) is performed on a linear pDNA template, and the RNA polymerase of T7 DNA48 synthesizes multiple copies of the RNA transcript. 67 Once the RNA is capped at the 5′ end and purified, it is ready for formulation. Three main types of RNA are currently being developed: self‐amplifying RNA (saRNA), nonreplicating mRNA (nrRNA), and trans‐amplifying mRNA (taRNA) 68 , 69 (Figure 1). In this section, we summarized a detailed overview of mRNA‐based clinical applications, and mRNA deliver system is available in part 3.
FIGURE 1.
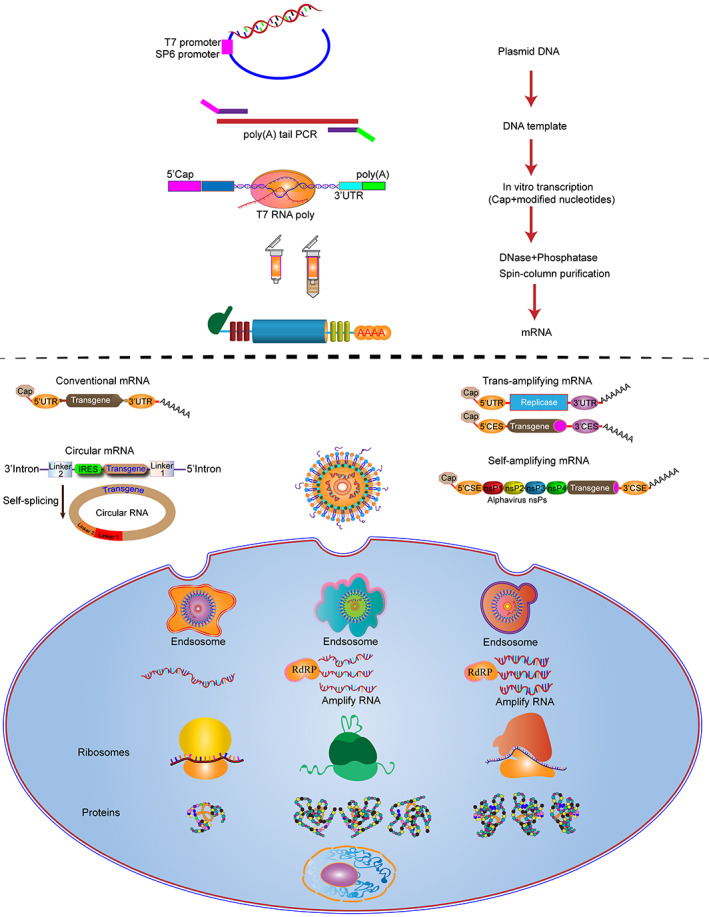
mRNA manufacturing process. In vitro transcribed mRNA encoding the gene of interest has been used to generate conventional mRNA. Use saRNA, which encodes a replicase that self‐amplifies mRNA for positive‐strand RNA viral genomes. The second utilizes a novel bipartite vector system of taRNA, in which the replicase is deleted to form a trans‐replicon. Both protocols improved the half‐life and translation efficiency of mRNA.
2.1. mRNA‐based protein replacement therapy
Over the past few years, evidence has revealed that mRNA‐based therapeutics have gained immense interest and are demonstrating to be viable options in a wide range of diseases, including cancer, 70 infectious disease, 71 and rare genetic diseases. 72 mRNAs can use the human body to translate and encode therapeutic proteins, such as transmembrane, intracellular, intramitochondrial, and secreted proteins. 73 Therefore, mRNA‐based therapeutics can potentially restore the function of a missing or defective protein in diseased cells or create new cellular functions altering the diseased state. 74
In mRNA‐based cancer immunotherapy, synthetic mRNA crosses the host cells through the cell membrane or by endocytosis and are translated to proteins in the cytosol. mRNA‐loaded DC vaccines can present major histocompatibility complexes (MHC) of the antigen‐presenting cells (APCs) to effector cells of the host immune system to induce cytotoxic T lymphocytes and natural killer (NK) cells attacking cancerous cells. 19 Moreover, mRNA‐based anti‐tumor vaccines can help generate an immune‐friendly tumor microenvironment (TME) through activating toll‐like receptors (TLRs) and retinoic acid‐inducible gene I (RIG‐I) and secreting type I interferon, as well as chemokine GRO, MCP‐1, RANTES, and MDC. 75 , 76 In addition, mRNA can encode monoclonal antibodies (mAbs), playing a passive targeted immunotherapy strategy and achieving a more comprehensive oncology therapeutic effect. 77 , 78 Therefore, there are several ongoing clinical trials of mRNA immunotherapies for a variety of cancers.
Cardiovascular diseases (CVDs) lead to more than one‐third of all deaths worldwide, mainly due to ischemic heart disease (IHD). 79 , 80 Recent evidence has demonstrated that mRNA‐based management is promising to improve myocardial regeneration and prevent heart failure. 4 , 81 , 82 The main mechanisms of IHD therapy include inhibition of cardiomyocyte death, promotion of cardiac regeneration, maintenance of coronary plaque, enhancement of angiogenesis, and triggering of cardiac reprogramming. 81 For instance, pyruvate kinase muscle isoenzyme 2 (PMK2) is a critical enzyme in aerobic glycolysis, contributing to cell metabolic reprogramming. 83 Furthermore, PMK2 can induce cardiomyocyte proliferation and is primarily expressed at higher levels in regenerative fetal heart and neonatal cardiomyocyte but not in adult cardiomyocyte. 84 Therefore, in IHD therapeutics, the upregulation of PMK2‐encoding mRNA can re‐invigorate the proliferation of cardiomyocyte, resulting in cardiomyocyte regeneration. 84 Moreover, it has been reported that insulin‐like growth factor‐1 (IGF1) protects cardiomyocytes after ischemia and myocardial infarction (MI). 85 Therefore, promoting the expression of IGF1 in mouse hearts after MI can enhance cell survival and inhibit cell death under hypoxia‐induced apoptosis. 85 , 86 In addition, for MI treatment, overexpression of an angiogenic factor, human vascular endothelial growth factor A (VEGFA)‐encoding mRNA, in mice promotes the differentiation of heart progenitor cells into endothelial cells, improves heart function, and prolongs long‐term survival. 87 Notably, mRNA‐based therapeutics play a vital role in injured hearts repair, angiogenesis, and cardiomyocyte regeneration.
mRNA‐based therapeutics have also become a new pillar for liver disease treatment. Crigle‐Najjar syndrome type 1 (CN1) is a rare metabolic disease characterized by the absence or decreased activity of UDP glucuronosyltransferase family 1 member A1 (UGT1A1), an enzyme required for glucuronidation of unconjugated bilirubin in the liver. 88 The mutation of UGT1A1 gene contributes to severe unconjugated hyperbilirubinemia and consequently develops jaundice and irreversible brain and muscular damage. 89 However, mRNA‐based therapeutics can restore the hepatic expression of UGT1A1 and normalize the levels of bilirubin. 90 Moreover, α‐1 antitrypsin (AAT) deficiency is a recessive disease caused by mutations in the serpin family A member 1 (SERPINA1) gene. 91 As a serine protease inhibitor, AAT neutralizes neutrophil plasminogen in the lung and liver to protect tissues from excessive proteinase activity. 92 , 93 The AAT deficiency can activate the intracellular injury cascade of apoptotic liver cell death, compensate hepatocellular proliferation, and lead to end‐organ injury. 94 Nevertheless, supplement of AAT‐encoding mRNA can significantly produce AAT protein and provide an alternative modality for liver AAT deficiency. 95 In addition, mRNA‐based therapeutics are currently being tested in preclinical and clinical studies for other liver diseases, including thrombotic thrombocytopenic purpura (TTP), 96 glycogen storage disease type 1A (GSD1), 97 acute intermittent porphyria, 98 and factor IX deficiency hemophilia B. 99 Therefore, mRNA‐based therapeutic are optimistic about liver disease treatment through the development of clinical studies.
Studies have shown that mRNA‐based therapy has outstanding advantages in treating neurological diseases. For instance, Kojima et al. have developed an emerging technique to delivery therapeutic catalase mRNA into the cytosol of nerve cell, which cannot only attenuate localized neuroinflammation but also rescue neuroinflammation in the model of Parkinson's disease. Moreover, mRNA‐based therapeutics also hold great potential for treating other diseases, such as leukemia, 74 asthma, 100 Fabry disease, 101 skin disease, 102 , 103 and bone regeneration. 104 , 105
2.2. mRNA vaccines
Vaccines play a pivotal role in the public health and the quality of human life, which not only prevent various infectious diseases but also reduce and eradicate mostly fatal disease, such as tuberculosis, smallpox, ebola, meningitis, and malaria. 106 , 107 Traditionally, vaccines can be roughly subdivided into live attenuated vaccines, inactivated vaccines, subunit vaccines, and toxoid vaccines, which were commonly applied in clinical practice, lastingly provided protection for various diseases. 107 , 108 , 109 Although conventional vaccines can provide long term and durable protection against diseases, they have difficulty in fulfilling pandemic needs as some emerging infectious diseases require more development speed, low‐cost manufacturing, and large‐scale production. mRNA vaccines have emerged as promising alternatives to traditional vaccines due to their advantages of rapid development, high efficacy, and cost‐efficient manufacturing. Furthermore, the mRNA vaccine is more likely to stimulate protective antibody and antigen‐specific T‐cell responses (Figure 2). Notably, the innate immune response is significantly enhanced following secondary immunization. 110 In addition, conventional vaccines are not suitable for cancer vaccine development. In contrast, mRNA vaccines encoded using tumor‐associated antigens (TAAs) can be developed more rapidly with cost‐effective approaches. 111 , 112 , 113
FIGURE 2.
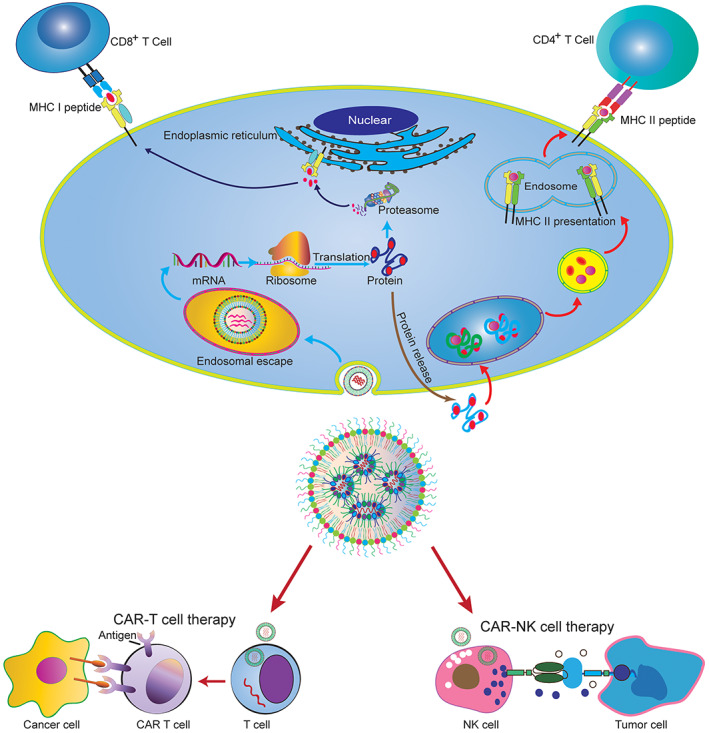
mRNA vaccines and mRNA delivery for CAR T‐cell/CAR NK‐cell engineering. Compared with other types of vaccines, mRNA vaccines have many advantages, including the use of noninfectious elements, reduced production time, and the potential to target multiple diseases. mRNA vaccines can be developed in the laboratory using DNA templates and readily available materials. Therefore, it can be standardized and mass‐produced, and the research speed of vaccines is faster than traditional vaccines. At present, COVID‐19 mRNA vaccines delivered by lipid‐derived nanoparticle (LNP) play a vital role in the fight against coronavirus as the first clinical mRNA vaccine.
Moreover, mRNA vaccines have several advantages over conventional vaccines. 114 , 115 First, mRNA vaccines are scalable due to modified mRNA sequence can satisfy all genetic information requirements to encode all protein. 55 Second, mRNA vaccines are comparatively effective and safe because they only target cytoplasmic delivery, avoiding the risk of genome integration. 116 Third, manufacturing mRNA vaccines on a large scale tends to be industrialized. 116 Therefore, mRNA vaccines have been widely studied in different kinds of diseases.
At present, there are several modes of production of RNA vaccines: template‐directed synthesis, nonreplicating, and self‐amplifying messenger RNA (samRNA) vaccines. 117 Nonreplicating vaccines represent a straightforward approach wherein administered mRNA is directly translated into the cytoplasm of transfected cells to encode the protein antigens of interest. 118 However, samRNA vaccines encode the RNA genome of a single‐stranded RNA virus and contain additional sequences in the coding region for RNA replication. 118 However, both nonreplicating and samRNA vaccines have been proven effective against infectious diseases, and nonreplicating vaccines have strong potential to be used in cancer immunotherapy. 112 , 115 , 118 , 119 So far, most mRNA vaccines have been developed for infectious diseases or cancer immunotherapy. To our knowledge, most ongoing trials or FDA‐approved mRNA therapeutics are based on typical linear mRNA, which can be produced by RNA polymerase‐mediated IVT. However, in contrast to typical linear mRNA, circular mRNA (circRNA) increases mRNA stability and has been found to contribute to robust and durable protein expression. 120 Therefore, the development of circRNA vaccines triggers higher and more durable effective neutralizing antibodies against severe acute respiratory syndrome coronavirus 2 (SARS‐CoV‐2) and shows adequate protection against new SARS‐CoV‐2 variants. 121 In addition, these circRNA vaccines can strongly induce immune responses and are more durable than typical mRNA vaccines. However, the development of circRNA vaccines faces problems, such as efficient cyclization of intramolecular RNA, purification of circRNA, and high cost of IVT production reagents. 122 We hope that advanced technology in circRNA by in vitro or in vivo synthesis can be developed in the coming years to facilitate the development of therapeutic circRNAs.
2.2.1. mRNA vaccines for infectious diseases
mRNA vaccines against infectious diseases, such as influenza, Zika virus, and rabies, are underway in clinical trials. In contrast, the development is relatively slow due to mRNA stability and the lack of an optimized delivery system. However, the coronavirus disease‐2019 (COVID‐19) pandemic has accelerated the development and application of mRNA vaccines. 123 COVID‐19 is a global pandemic caused by SARS‐CoV‐2, which has affected hundreds of millions of people and led to millions of deaths worldwide. 124 , 125 The rapid global spread of COVID‐19 has posed an international health emergency and economic burden as a devastating pandemic of the 21st Century. 126
Two rapid‐response mRNA vaccines (mRNA‐1273 and BNT162b2) against COVID‐19 are licensed by the FDA, which are administered in the general population of many countries and have demonstrated excellent tolerability and high levels of protection against COVID‐19. 127 Moreover, accumulating evidence has confirmed that the effectiveness of the BNT162b2 can achieve a protection rate of more than 90% against COVID‐19. In comparison, mRNA‐1273 has a protection rate of 87.4%, decreasing hospitalization and hospital mortality by 95.8% and 97.9%, respectively. 128 , 129 Unfortunately, the effectiveness and protection of these vaccines are reduced due to SARS‐CoV‐2 variants, including alpha (B.1.1.7 and descendant lineages), beta (B.1.351), gamma (P.1), delta (B.1.617.2 and AY lineages), and omicron (B.1.1.529 and BA lineages). 130 Therefore, further improvement and development of mRNA vaccines are necessary to protect the constantly evolving SARS‐CoV‐2 virus. Fortunately, multiple mRNA vaccines have been designed against acute infectious viruses, such as flaviviruses, influenza viruses, and rabies virus, and chronic infectious viruses, such as hepatitis C virus (HCV) and human immunodeficiency virus (HIV). 131 Moreover, most of these mRNA vaccines have been in the phase of clinical trials. 131
2.2.2. mRNA vaccine for cancer therapy
The clinical application of mRNA‐based tumor vaccine has achieved significant progress over the past decade. 119 , 132 mRNA‐based vaccine is an innovative administration of cancer immunotherapy, and the vaccine plays a vital role in delivering the antigens recognizing by the body's immune cells to induce immune response. 113 , 133 Moreover, mRNA‐based vaccines can code tumor‐specific antigens (TSAs), introduce them into APCs, and synthesize the required antigens, stimulating both humoral and cellular immunity to kill cancer cells. 115 In addition, mRNA‐based vaccines can also encode and express TAAs and TSAs to specifically attack and eliminate cancer cells. 134
To date, some strategies have demonstrated that mRNA‐based vaccines possess the feasibility and tremendous potential to be applied in cancer treatment. For instance, BI1361849, as an mRNA encoding non‐small cell‐associated tumor antigens, can promote the proliferation of functional tumor‐specific CD4 and CD8 cell, killing the cancer cell and improving survival. 135 In addition, DCs‐based mRNA vaccines also provide the necessary modalities to administer the tumor. DCs, as the most efficient APCs, play a vital role in the activation process of helper T and killer T cells through capturing, processing, and presenting antigens to T cells. 136 The DCs‐based mRNA tumor vaccines are produced undergoing multiple procedures: DCs are extracted from the patient's peripheral blood, and the differentiation and maturation are stimulated using the granulocyte‐macrophage colony‐stimulating factor (GM‐CSF) and interleukin‐4 (IL‐4); then mRNA encoding TSAs is transfected; the mRNA‐loaded DCs are finally reinfused into the patients, and the immune system is stimulated to combat cancer cells. 137 For example, the pp65‐mRNA‐loaded DCs vaccine is used to treat glioblastoma, and it can increase long‐term progression‐free survival and prolong overall survival. 138 All these studies show that mRNA‐based vaccines may be safe and effective alternatives to cancer immunotherapy. In addition, many mRNA‐based cancer vaccines have been evaluated for their effectiveness.
2.3. Genome editing by mRNA
In recent years, genome editing has become the most potent strategy in the field of gene therapy. Zinc finger nuclease (ZFN), transcriptional activator‐like effect nuclease (TALEN), and clustered regularly interspaced short palindromic repeat sequence (CRISPR)‐related protein 9 (Cas9) systems have been developed as powerful tools for locus‐specific modification of genomes, providing a promising strategy for diseases caused by gene defects. 139 , 140 However, these approaches have disadvantages of the risk of off‐target mutagenesis. 141 Consequently, mRNA‐based encoding ZFNs, TALENs, and CRISPR/Cas9 has been successfully applied for genome editing in vivo and in vitro, providing an attractive alternative due to their transient expression. 141
mRNA‐based genome editing open the opportunity to be used in various therapeutic field as a novel drug candidate. For example, ZFNs‐encoding mRNA is used to target the human C‐C chemokine receptor 5 (CCR5) for the treatment of HIV. 49 , 142 Moreover, TALEN‐encoding mRNA treats pyruvate kinase deficiency (PKD) by correcting the pyruvate kinase L/R gene (PKLR) in hematopoietic stem cells. 143 In addition, nanoparticle‐mediated CRISPR/Cas9‐encoding mRNA delivery is the most frequently used gene‐editing technology (Figure 3). Several studies have shown that in non‐human primate models, CRISPR/Cas9‐mediated CCR5 disruption is used to give resistance to HIV/SIV infection. 144 Therefore, mRNA‐based genome editing will be a promising modality for gene therapy, and several of them are undergoing clinical studies.
FIGURE 3.
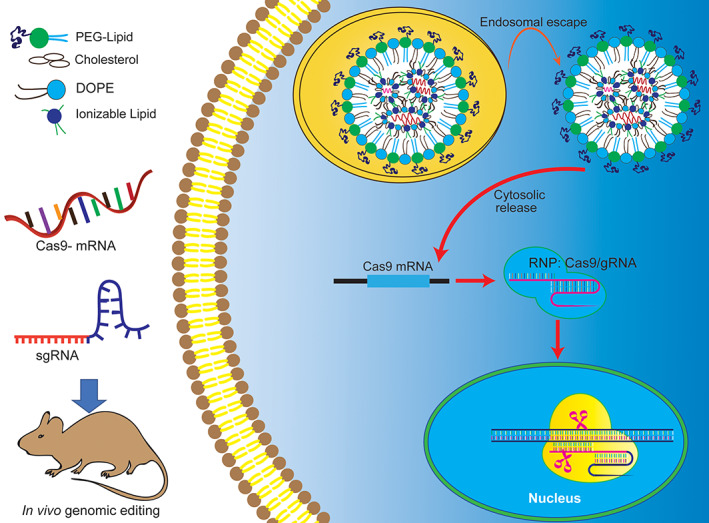
Delivery of mRNA for genomic editing by nanoparticles. The CRISPR/Cas9 components (sgRNA, Cas9 mRNA) are encapsulated in nanocarriers, which need to escape from endocytosis/lysosomes after cellular uptake. Then, it will be translated directly into proteins in the cytoplasm and transported to the nucleus so that the CRISPR machinery can act on the genomic DNA of the cell. mRNA delivery and translation are transient and thus avoid the concern of permanent integration of CRISPR genes into the host genome.
3. NANOPARTICLES FOR MRNA DELIVERY
mRNA must to enter the host cytoplasm and express specific antigens to remain functional. However, it is difficult for negatively charged mRNA macromolecules to penetrate the anionic lipid bilayer on the cell membrane. Moreover, the single‐chain mRNA is fragile and easily degraded by extracellular ribonucleases in skin and blood. 9 Therefore, an ideal mRNA delivery system not only enhances efficient cellular uptake by host cells but also protects them from degradation. In the following sections, we describe various mRNA delivery systems developed and applied.
3.1. Lipid and LNPs for mRNA delivery
The lipid can be cationic, neutral, and zwitterionic, which is an important class of vector material for mRNA delivery. 145 However, cationic lipids represent the widely applied and studied materials for mRNA delivery. 145 For example, N‐[1‐(2,3‐dioleyloxy) propyl]‐N,N,N‐trimethy‐lammonium chloride (DOTMA), as the first synthetic cationic lipids, is used to delivery mRNA to several cell lines. 146 Unfortunately, toxicity and immunogenicity may be the main obstacles to its clinical application. 147 The cationic lipids, combined with other components forming nanoparticles, are named LNPs. 148 LNPs are the most extensively investigated and clinically advanced delivery system for mRNA‐based therapy. 13 , 149 LNPs are nanocarriers with a monolayer structure of phospholipids, which can protect the mRNA in the lipid core from degradation. Usually, LNPs are synthesized by cholesterol, polyethylene glycol‐modified (PEGylated) lipids, helper phospholipids, and ionizable lipids. 13 Figure 4 represents the structural classes of lipids components of LNPs tested for mRNA‐based vaccines and treatments. In addition, each component of LNPs exerts a different function in mRNA delivery. Cholesterol can enhance the stability of lipid nanoparticles, assist membrane fusion, and facilitate the entry of mRNA into cytoplasm. 150 The helper phospholipids are generally saturated phospholipids, which can improve the overall phase transition temperature and stability of nanoparticles. 151 PEGylated lipids can potentially improve the manufacturability and stability of LNPs and prevent macrophage‐mediated degradation. 152 Ionizable lipids are the most critical excipient and decisive factor in releasing mRNAs in the cytoplasm through ionization. 153 , 154 Therefore, LNPs have been widely utilized as the vehicle for mRNA drugs. For instance, LNPs‐mediated delivery of IL‐10 mRNA can promote the expression of IL‐10 in inflammatory leukocytes and attenuate the dextran sodium sulfate (DSS)‐induced colitis in experimental mouse. 155 Moreover, LNPs mediated delivery of mRNA vaccines against COVID‐19 has confirmed encouraging results and shown a perfect protective efficacy across different population. 60 , 127 In addition, LNPs mediated delivery of Cas9 mRNA is to exert cancer immunotherapy or genome editing. 156 , 157 , 158
FIGURE 4.
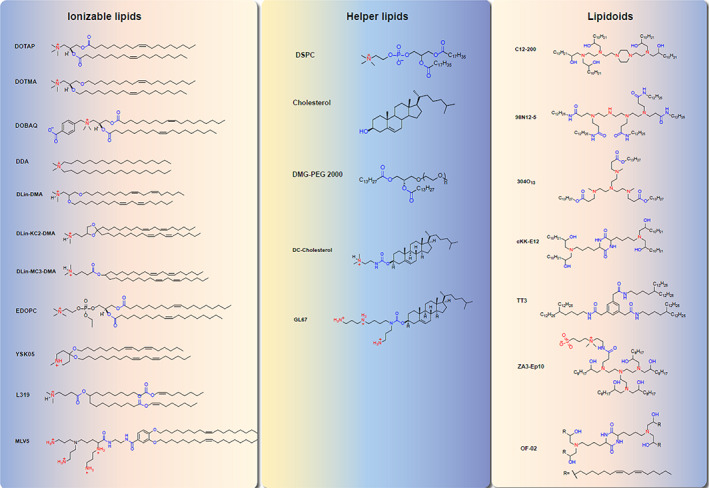
The chemical structural formula of major lipids utilized by lipid nanoparticles for mRNA‐Based therapy.
Despite their unprecedented potential, several studies have demonstrated that LNPs still have potential limitations that hinder them practical application. One limitation is the safety of LNPs due to their potential toxicological effects and the tissue distribution of their payloads. 159 , 160 Another limitation is immunogenicity of LNPs because LNPs constituents can induce immune response, promote neutrophil infiltration, and boost the secretion of proinflammatory factor and reactive oxygen species (ROS). 161 Moreover, LNPs induced side effects, such as pain, redness, and fever, have been reported. 162 , 163 Therefore, further studies on the cytotoxicity and immunogenicity of LNPs delivery system are urgently needed.
3.2. Lipopolyplex for mRNA delivery
The lipopolyplex (LPP) nano‐delivery platform is a bilayer structure with a polymer‐encapsulated mRNA molecule as the core, which is wrapped in a phospholipid‐coated bilayer shell to protect the mRNA molecule in the polyplex core structure from RNase degradation. As a nonviral gene delivery vector, LPP combines the advantages of polymers and liposomes, exhibiting superior stability, reduced cytotoxicity, high gene transfection efficiency, and the ability to release mRNA molecules progressively with polymer degradation. For example, Persano et al. have developed a lipopolysaccharide (LPS) mRNA vaccine consisting of a poly(β‐amino acid) polymeric mRNA core wrapped inside a lipid shell. Intraperitoneal injection of LPP carrying ovalbumin antigen and tyrosinase‐related protein 2 (TRP2) mRNA can target DCs and activated T‐cell immune responses and melanoma lung metastases in a mouse model of the tumor, with over 90% reduction in tumor nodules nodule size were observed. 164 In contrast to liposome delivery of mRNA, LNP delivery of N1 methyl pseudouridine‐modified mRNA cannot induce type I IFN for effective T‐cell immunity. This mode of action can reduce the inflammatory response without impeding T‐cell immunity. This approach provides a new way to explore effective and low inflammatory mRNA lipid polymorphs. 165
3.3. RNA delivery with virus‐like nanoparticle
Virus‐like nanoparticle (VLP) delivery system is an intermediate between viral and nonviral vectors. These particles contain most components from viral vector, such as envelope and capsid but no viral genome. 166 Specific recognition between mRNA stem‐loop structure and viral capsid structure capsid protein can provide VLP‐mRNA formation by virus engineering technology. In particularly, VLP‐mRNA can efficiently infect cells with the help of viral capsid and transient expression of mRNA itself. It has been shown that Cas9 mRNA delivered by VLP‐mRNA can be present for only 72 h compared to viral systems expressing Cas9 for a long time. Therefore, the off‐target effects can be significantly reduced. VLP delivery of cas9 mRNA has successfully developed Vegfa and reduced neovascularization area by 63% as a significant therapeutic approach in a mouse model of age‐related macular degeneration. Sequence results show that VLP‐mRNA does not induce off‐target effects. 167 These experimental results strongly support the potential of VLP for clinical application in delivering CRISPR gene therapy. 168 Yadav et al. have fused an aptamer‐binding protein (ABP) to the N‐terminal of the HIV Gag protein. The interaction between RNA aptamers and ABPs can recruit Cas9 mRNA or RNP for enhanced vector production and gene expression. 169 Segel et al. have found that one of the retroviral‐like proteins (PEG10) directly binds to an extracellular viral‐like capsid and secretes its mRNA into the vesicle. The mRNA cargo using PEG10 can be engineered to package, secrete, and deliver specific RNAs by linking the target mRNA to the UTR of PEG10, and PEG10‐based VLP have low immunogenicity compared to currently available viral vectors, dramatically expanding the range of applications for nucleic acid therapy. 170 As a naturally occurring biodegradable nanomaterial, VLP avoids disrupting the genome integrity raised by retroviruses. With the further development of gene technology, VLPs are being developed as safe and effective vaccines and gene editing delivery tools.
3.4. Membrane‐coated nanoparticles/biomimetic nanoparticles
Cell membrane‐coated vehicles can mimic the properties of cell membranes, which combine the properties of natural cell membranes with those of nanomaterials, thus significantly improving biocompatibility while enabling long circulation and targeted delivery in vivo. A series of nano‐drug carriers have been developed using immune cell membranes, such as leukocytes, macrophages, neutrophils, and other “camouflage.” In addition, tumor cells and bacteria can be used to prepare cell membrane‐camouflaged nanocarriers. Given the specific proteins on the surface of tumor cells and bacteria that activate the immune system and improve adhesion, these new drug carriers are more diverse in function delivery of mRNA drug than traditional nanocarriers. Moreover, the utilization of specific recognition proteins on cell membranes can achieve targeted drug delivery to lesion sites, offering a novel strategy for significantly improving the therapeutic effect. Despite the apparent advantages of membrane‐coated nanoparticles (CM‐NPs), technical preparation for such CM‐NPs remains challenging. 171 Therefore, much work remains to be explored before their clinical implementation.
Meanwhile, virus‐mimicking cell CM‐NPs promise to improve cellular delivery efficiency. Previous studies have shown that the hemagglutinin (HA) protein or HA2 peptide derived from the surface of the influenza virus mediates membrane fusion in the endosomal acidic pH environment. 172 Therefore, researchers genetically modified cells to express the HA protein or insert HA2 peptide on the cell membrane to create such nanoparticles. They have then isolated cell membranes coated mRNA/miRNA nanoparticles inside. 173 This virus‐mimicking cell MC‐NP can trigger the endosomal escape and release its mRNA/miRNA load to reach intracellular localization for protein regulation. 174
3.5. Polymer‐based delivery system
The polymer‐based delivery system consists of another prominent family of mRNA delivery carrier and has excellent potential for mRNA delivery. 175 Moreover, a polymer‐based delivery system is comprised three kinds of polymer, including cationic polymer, dendrimer, and polysaccharide polymer. 13 Cationic polymers have the advantages of easy synthesis and modification. In addition, cationic polymer can be complex with mRNA by electrostatic attraction and hydrophobic interaction, contributing to a more stable polyplex. 176 Polyethyleneimine (PEI) is one of the most widely used network polymers and has high efficiency of mRNA delivery. 177 However, PEI has the disadvantages of poor biodegradability and high toxicity, limiting its broad clinical application. 147 Fortunately, with the development of chemical modification, the toxicity of PEI is reduced while maintaining its protonatable properties and high delivery efficiency. 178 For example, Li et al. have found that linking cyclodextrin to PEI not only reduces the cytotoxicity of cationic PEI polymer but also maintains a large number of protonatable groups. Moreover, they have further discovered that modified PEI‐based mRNA encoding HIV‐1 gp120 can be dramatically up‐taken by lymphoid tissue, resulting in anti‐HIV immune response in experimental mice. 178
Dendrimer polymer is a vital delivery vehicle with high tolerability, which contains a core and multiple regularly hyperbranched monomers. 179 , 180 Results from Xiong et al. show that the delivery system based on dendrimer polymer can successfully transport many kinds of mRNAs to tumor tissue, which provides convenient for tumor radiography and treatment. 181 Moreover, the etiology of hepatorenal tyrosinemia type I (HT‐1) is involved in the mutation of fumarylacetoacetate hydrolase (FAH), which prevents catalysis of the terminal step in the tyrosine catabolic pathway, resulting in an aggregation of toxic metabolites and the disfunction of liver and kidney. 182 Interestingly, the delivery of therapeutic FAH mRNA based on dendritic molecules can restore liver function and prolong the survival time of HT‐1 mice. 183 Therefore, these results provide a dendrimer polymer delivery system with great potential to improve mRNA delivery and treat genetic diseases.
Polysaccharides, as relatively common natural biomaterials, can be easily chemically modified for efficient delivery with high biocompatibility and little toxicity and are mainly composed of chitosan, alginate, dextran, and hyaluronic acid. 184 Chitosan is one of the most common polysaccharides to deliver mRNA. It has been reported that chitosan‐based deliver therapeutic surfactant protein B (SP‐B) mRNA can replace SP‐B deficiency and experimental asthma in mouse models and prolonged life in treated mice. 185 Cystic fibrosis (Cf) is a genetic disorder that originates in an alteration in the cystic fibrosis transmembrane conduction regulator (CFTR) gene. 186 , 187 Fortunately, chitosan‐based delivery of CFTR mRNA can restore CFTR function and provide great potential for treating CF. 188 Alginate, as an important polysaccharide, is derived from brown algae and bacteria and consists of a‐l‐guluronic acid and b‐d‐mannuronic acid building blocks linearly linked by 1,4‐glycosidic linkages. 189 Moreover, alginate has biodegradable, nontoxic, and abundant features. 190 It has been reported that delivery of alginate‐based neurotrophin‐3 (NT‐3) mRNA can enhance nerve regeneration. 191 Dextran, as a water‐soluble, naturally degradable polysaccharide, originates from bacterial metabolites and consists of (1,6)‐a‐d‐glucose with various ratios of linkages and branches. 190 It has been demonstrated that dextran‐based mRNA delivery can induce high gene expression and low cytotoxicity and provide a promising strategy to treat breast cancer. 192 Hyaluronic acid, as a nonsulfated glycosaminoglycan, is composed of glucuronic acid and N‐acetyl‐d‐glucosamine bound linked beta‐linkages with good bio‐compatibility and biodegradability. 193 Yan et al. have found that hyaluronic acid‐based Wingless and the name Int‐1 (WNT) 16 mRNA can be delivered to human cartilage explants and antagonize canonical β‐catenin/WNT3a signaling, contributing to increased lubricin production and decreased chondrocyte apoptosis in osteoarthritis (OA). 194 Therefore, polysaccharides polymer may be a promising substitute to deliver mRNA in treating human diseases.
3.6. Inorganic‐based nanomaterial for mRNA delivery
Inorganic nanomaterials as delivery vehicles have unique physicochemical properties, such as excellent storage stability, good biocompatibility, and easy preparation, making them ideal platforms for mRNA delivery. Currently, inorganic nanostructures, including quantum dots, silica nanoparticles (SNPs), gold nanoparticles (GNPs), and carbon‐based and magnetic iron oxide‐based nanostructures, are the most popular types in the field of nanomedicine. AuNPs exhibit unique properties for delivering nucleic acid drugs. Through covalent or noncovalent conjugation, functional moieties, such as nucleic acid drugs and targeting ligands, can covalently attached to the gold nanoparticle core through a thiol moiety. AuNPs are expected to be attractive scaffolds for mRNA delivery due to their tunable size, simple functionalization, nontoxicity and immunologically inert properties. Yeom et al. have observed that mRNA encoding the pro‐apoptotic factor Bcl‐2‐associated X (BAX) protein is encapsulated by AuNPs, showing highly efficient mRNA delivery and inhibiting xenograft tumors. 195 The ligand length, density, particle size, hydrophobicity, and affinity of AuNPs need to be considered and optimized for efficient, targeted delivery of mRNA nanoparticles. Iron oxide nanoparticles are nanoscale magnetic materials mainly composed of iron‐based oxides, which have been widely used in biomedical research due to their superparamagnetic properties at specific sizes, biocompatibility, and easy‐to‐modify surfaces for biologics. For example, iron oxide nanoparticles coated with nucleic acid‐loaded lipidoids exhibit efficient RNA and DNA delivery in vitro. Most recently, Erasmus JH et al. developed a lipid in organic nanoparticle (LION) consisting of inorganic superparamagnetic iron oxide (SPIO) nanoparticles for delivery saRNA, which is undergoing clinical trials for the protection against SARS‐CoV‐2 infection. 196 This formulation is highly stable for at least 3 months when stored between 4 and 25°C. The showcases the role of SPIO nanoparticles in enhancing the stability, delivery, and immunogenicity of mRNA vaccines. However, the long‐term, nonspecific accumulation of these inorganic nanomaterials in organisms may cause toxicity, which hinders their large‐scale clinical applications. Detailed toxicological assessments (genotoxicity and oxidative stress) are therefore needed. It is crucial to develop biodegradable and scavengable inorganic nanomaterials to minimize the short‐ and long‐term cytotoxicity of inorganic nanoparticles. Besides, these inorganic nanomaterial carriers must be targeted to specific tissues or organs to minimize side effects.
3.7. Exosome‐based nanoparticles for mRNA delivery
Exosomes are engendered by the inward budding of the endosomal membrane forming Intraluminal vesicles (ILVs) within multivesicular bodies (MVBs). When MVBs fuse with the plasma membrane, the ILVs are released into the periplasmic space referred to as exosomes (Figure 5). As a group of small vesicles (50–150 nm), exosomes contain large amounts of specific proteins, lipids, DNA, mRNA, noncoding microRNAs (microRNAs and miRNAs), and enzymes, which are a new mode of intercellular communication. Therefore, exosomes can function as an mRNA cargo delivery system. 197 Table 2 summarizes the recent progress of exosome‐based mRNA delivery systems for treating various diseases. In contrast to synthesized nanoparticles, native exosomes possess outstanding properties, such as excellent biocompatibility and low immunogenicity. In addition, due to the small size of exosomes, it can inhibit the clearance by mononuclear phagocytes and has high permeability and retention effects at solid tumor sites, which can achieve drug accumulation at target sites. Exosomes themselves can hijack the blood–brain barrier (BBB) to deliver drugs into the brain. Especially, intranasal administration can promote the drug to reach the brain lesions quickly, providing the possibility for noninvasive treatment of brain diseases. Using the advantages of exosomes across the BBB, Kojima et al. have encapsulated catalase mRNA in HEK293T‐derived exosome to effectively attenuate neuroinflammation in the model of Parkinson's disease or LPS‐induced rats. 203 Yang et al. have employed exosomes to encapsulate phosphatase and tensin homolog (PTEN) as tumor suppressor for brain cancer treatment. 202 In this way, gliomas‐targeting peptides are anchored to the N‐terminus of exosomal membrane protein CD47, which remarkably enhances the cellular uptake of exosomes by glioma cells. These modified glioma‐targeted exosomes exhibit a significant inhibitory effect on tumor cell dissemination without any side effects on other normal tissues.
FIGURE 5.
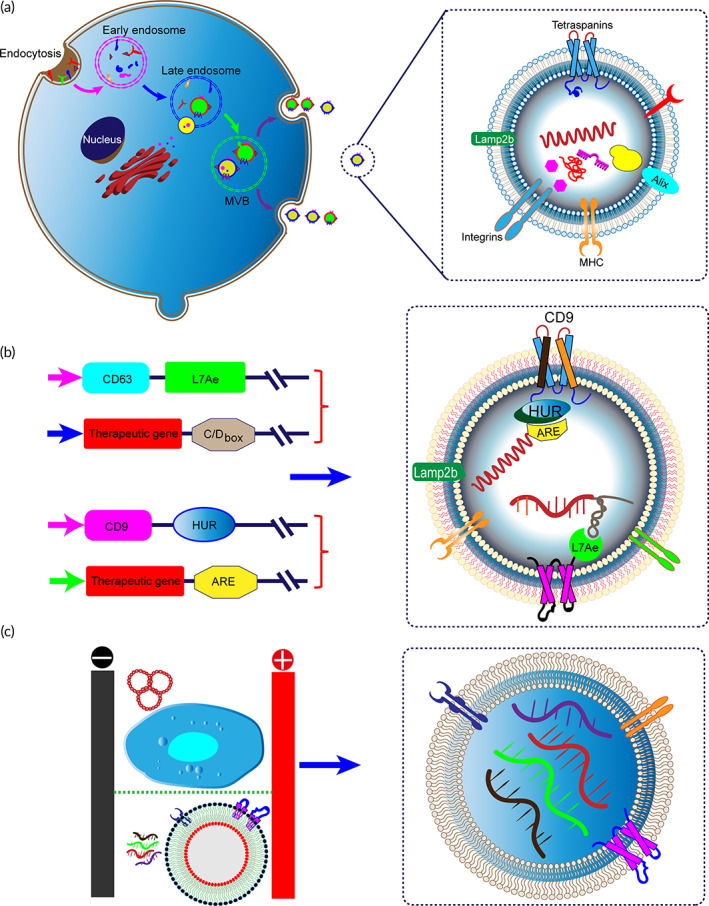
Exosome‐mediated mRNA delivery for gene therapy. (a) Exosomes are biogenesis from the invagination of the cell membrane and mature into late endosomes/MVBs. Then, MVBs fuse with the cellular membrane and are released into the extracellular space. (b) Packaging and delivery of RNAs via binding with RNA‐binding protein (HUR or L7Ae) that recognizes specific sequences on RNA. (c) Cellular‐nanoporation technology for large‐scale generation of exosomes containing therapeutic mRNAs
TABLE 2.
Recent advances in exosome‐mediated mRNA therapy and genomic edit
| Donor cells | Cargo | Loading method | Function | References |
|---|---|---|---|---|
| HEK293T RAW264.7 | Interleukin‐10 mRNA | Cells were transfected with IRES‐IL‐10 expressing vector plasmids | Reduces atherosclerotic lesion formation | 198 |
| AML12 cells | Ldlr mRNA | Cells were transfected with Ldlr plasmids | Ameliorate the phenotype of atherosclerosis | 199 |
| Bone marrow‐derived MSCs | LDLR mRNA | Ldlr‐expressing virus vector | Homozygous familial hypercholesterolemia patients | Clinical trials: NCT05043181 |
| HEK293 | Nerve growth factor (NGF) mRNA | Cells were transfected with NGF plasmid | Reduce inflammation and rescue ischemic injury | 200 |
| HEK‐293 T | UPRT mRNA | Transfected cell with pCD‐UPRT‐EGFP plasmid | Activate pro‐drug and inhibit schwannoma tumor growth | 201 |
| MEFs, MSCs, DCs or HEK‐293 T cells | PTEN mRNA | Cellular nanoporation | Restore tumor‐suppressor function, produce larger amount of PTEN mRNAs into exosome | 202 |
| HEK‐293 T (EXOtic devices) | Catalase mRNA | Cell were transfected plasmids encoding catalase mRNA (PhCMV‐Catalase‐C/Dbox‐pA) | Protection against neurotoxicity and neuroinflammation | 203 |
| 293FT | HCHrR6 mRNA | Transfection of cells with the XPort/HChrR6 encoding plasmid | Targeted delivery and inhibit tumor growth in vivo | 204 |
| 293F | SARS‐CoV‐2 spike and nucleocapsid proteins | Incubation with lipid‐coated mRNAs | Induce long‐lasting cellular and humoral responses | 205 |
| Red blood cell‐derived EVs (RBCEVs) | Cas9 mRNA | Electroporation | Improve genome editing | 206 |
| SK‐LU‐1 Lung adenocarcinoma cells | p53 mRNA | Transfected with wt‐p53 expressing plasmid DNA | Support antitumor environment leading to decreased oncogenesis | 207 |
| HEK293T cells | CRISPR/dCas9 mRNA | CD9‐HuR and AU rich elements | Effectively deliver CRISPR/Cas9 system and reduced C/ebpα expression in the liver | 208 |
| HEK293T cells |
GFP mRNA p53 mRNA |
Co‐transfected with ARRDC1‐TAT and TAR‐GFP/TAR‐p53 | Efficiently package and deliver mRNA | 209 |
| HEK 293 T cells human lung spheroid cells |
Spike‐mRNA GFP‐mRNA RFP‐mRNA |
Electroporation | Dry powder available drug‐delivery vehicles inhalation of mRNA | 210 |
Mizrak et al. have exploited the genetically engineered microvesicles (MVs) derived from parent cell overexpressing suicidal CD‐UPRT mRNA. They have found that MVs deliver CD‐UPRT mRNA into schwannomas via direct intratumoral administration, which can achieve high‐level expression of functional proteins and promote tumor cell regression following systemic treatment with the prodrug 5‐FC. 203 Exosomes can package synthetic mRNA from overexpression of RNA in parental cells by DNA transfection or lentiviral transduction. Therefore, mRNA can be passively loaded into exosomes by transient transfection of IL‐10 in parent cell. Exosome‐based engineered IL‐10 mRNA offers an efficient drug delivery vector target to macrophages in plaques of experimental atherosclerosis and alleviates atherosclerosis in ApoE−/− mice. Therefore, it provides a safe and efficient approach to treat atherosclerosis. 198 According to a recent study by Wang et al., exosomes were used as carriers to improve the delivery of mRNA encoding HChrR6 to target cells (HER2+ cells). 204 Li et al. have demonstrated that exosomes from Ldlr overexpressing liver cells can protect against atherosclerosis. For the applicable clinical trial, they have used GMP‐compliant normal donor bone marrow‐derived MSCs to yield Ldlr mRNA‐enriched exosomes. They plan to inject exosomes via ultrasound‐guided paracentesis as a first‐in‐human study to evaluate the safety of this exosome product for gene therapy. This study is expected to provide a new therapeutic strategy for patients with homozygous familial hypercholesterolemia (Clinical Trials: NCT05043181).
Exosomes have received increasing attention as promising vehicles for the delivery of mRNA vaccines, especially those against tumors and Covid‐19, as DC‐derived extracellular vesicles containing MHCs on their surface can enhance T‐cell immune responses in patient. 211 In the further development of exosome as vaccines, the delivery of mRNA of pathogenic proteins into DC‐derived may provide a rich source of antigens. 211 In this approach, total tumor RNA is loaded onto DC‐derived extracellular vesicles. Exosome‐mediated mRNA delivery is safe and has high clinical potential for inducing SARS‐CoV‐2 immune function. Compared with LNPs that cause significant cytotoxicity, exosomes have no side effects in vitro or in vivo at any dose tested. Furthermore, exosomes medicate functional mRNA to human cells showing better performance than mRNA‐loaded LNPs. Exosomes incorporated with mRNA encoding spike protein of SARS‐CoV‐2 can significantly induce long‐term immune responses. 205 Collectively, these results reveal that exosomes can be utilized as deliver carrier for mRNA against COVID‐19.
Furthermore, traditional transfection methods can directly load the purified exosomes with synthetic mRNA molecules. After the electroporation, small micropores appear on the surface of exosomes, and the mRNA can enter the vesicle along the transient small hole. Electroporation has been widely used as a technology for loading exogenous mRNA. For example, extracellular vesicles from human erythrocytes have also been electroporated with mRNA‐encoded Cas9 and gRNA targeting MiR‐125b‐2 locus, showing genome editing in leukemia cells 206 (Figure 5a). However, the mRNA of the gene‐editing protein Cas9, with a size of nearly 5000 nt, makes it challenging to encapsulate CRISPR/Cas9 into exosomes by electroporation or other strategies. For specific enrichment of CRISPR/Cas9 mRNAs into extracellular vesicles, the exosomal transmembrane protein CD9 is fused with HuR protein, which can facilitate the sorting of RNAs with AU‐rich elements into exosomes. Li et al. have shown that Cas9 mRNA engineered with AREs (AU‐rich elements) can be efficiently loaded into exosomes and significantly reduce the expression of C/ebpα in the liver. 208 , 212 , 213 Previous evidence has suggested that the ribosomal protein L7Ae forms a classic kink‐turn structure with the C/D box RNA structure, 214 and the resulting protein–RNA complex can serve as a unique way to load mRNA (Figure 5b). Kojima et al. have assembled L7Ae before the N‐terminus of CD63 and fuse the C/D box at the end of the 3′‐UTR of the target gene. Interactions between RNA‐binding protein and the box C/D motif can specifically recruit RNAs into exosome. 203 Another approach to precisely manipulate exosomes utilizes a specific subset of MVs termed “arrestin domain‐containing protein 1 (ARRDC1)‐mediated MVs (ARMM).” In this method, the C‐terminus of ARRDC1 is fused to a TAT peptide that mainly reacts with TAR and a stem‐loop‐containing transactivation response (TAR) element. RNA cargo molecule can be efficiently loaded into ARMM. GFP and p53 mRNAs are successfully delivered into exosomes by this strategy and expressed in recipient cells. 209 To improve the mRNA‐loading efficiency of exosomes, James Lee's group have reported a cellular nanoporation method to produce large amounts of therapeutic mRNA‐containing exosomes 202 (Figure 5c). The production of RNA‐loaded exosomes can be further promoted by mechanically extruding the vesicles through a micrometer pore filter using cells expressing of the desired mRNA. 215
In addition to exosomes as emerging carriers for mRNA drug delivery, surface functionalization of exosomes can achieve target‐specific delivery. 216 For example, we used targeted peptide expressed with Lamp2b on the external surface of exosomes to target chondrocytes or mesenchymal stem cells. 217 , 218 , 219 Thereby, modified exosome can precisely delivery mRNA molecules to target cells or organs. Overall, exosomes are highly biocompatible and have substantial clinical application potential, opening up a new avenue for mRNA drug delivery.
4. CONCLUSION AND FUTURE PERSPECTIVES
The rapid development of mRNA therapy has brought unprecedented prospects and opportunities in the biomedical field. We comprehensively reviewed the latest progress in mRNA delivery, which gives essential value for designing mRNA delivery systems. Addressing the poor stability of mRNA for in vitro and in vivo target delivery is a crucial aspect of its clinical application. Meanwhile, mRNA has a large molecular weight and is negatively charged, making it challenging for exogenous mRNA to enter the cells. However, clinical trials have shown that physical methods are often harmful to cells and are unsuitable for in vivo application. Although viral vector‐based delivery of mRNA is highly efficient and usually shows better transfection efficiency, the undesirable properties of viral vectors, including potential carcinogenicity and high immunogenicity, hinder its broad clinical application. Until the outbreak of the COVID‐19 pandemic in 2020, mRNA‐based therapy was used for the first time in clinical practice. All approved mRNA vaccines use the lipid‐based nanoparticle delivery system. Several preclinical and clinical trial studies of mRNA vaccines for infectious disease and cancer therapy have also confirmed the reliability of nano‐delivered mRNA‐based therapeutic platforms. For mRNA delivery purposes, nanoparticle should possess several features: effectively encapsulate and protect mRNA from degradation before reaching the target site; and carriers effectively help mRNA delivery into target cells and release mRNA to the lysosome promptly. To this purpose, the commonly used carriers for mRNA delivery, including lipid nanoparticles, LNP, polymer nanoparticles, exosomes, and MVs, are evolved as a promising class of nanomaterials for RNA delivery. We summarized the strengths and weaknesses of various nanomaterials in mRNA delivery (Table 3).
TABLE 3.
Several advantages and challenges for mRNA delivery
| Delivery method | Advantages | Limitations | References |
|---|---|---|---|
| Viral Vectors | Escape the endosome, high transduction efficiency | Large‐scale production, high production costs, potential to induce undesired insertional mutagenesis and immunogenicity | 117, 220 |
| Physical methods (microinjection, electroporation, acoustic, laser and magnetic forces) | Low cost, high delivery efficiency | Potential tissue and cellular damage limitations for in vivo use | 220, 221 |
| Liposomes | High cargo capacity, escaping phagocytosis | Require complex production, difficult scalability, lack of selectivity for target tissues | 48, 222 |
| Lipid nanoparticles | Generally biocompatible, fast and scalability preparation, high reproducibility, high encapsulate efficiency, efficient condensation of mRNA, better kinetic stability | Potential cytotoxicity, relatively short circulation, rapid renal clearance, shorter half‐life blood circulation time, trigger immunogenic response | 48 |
| Polymer‐based nanoparticles | Good biocompatibility, easy for fabricate and functionalization, effectively encapsulate and release mRNA, protects RNA from degradation, biodegradable, industrial scale‐up | Safety risks, inherent toxicity of the components, insufficient targeting capabilities, limited efficiency of drug load, possible drug leakage | 223 |
| Inorganic nanoparticles | High encapsulation ability, simulate imaging, surface‐engineered, high delivery efficiency, highly stable, improved circulation time | Poor degradability, liver bioaccumulation; low‐tolerability, induce potentially immune responses | 224 |
| Lipopolyplex (LPP) | Stability in blood, protectant mRNA from degradation, highly encapsulation efficiency, dendritic cell targeting, endosomal escape | Design and formulation of lipopolyplexes are complex | 225, 226 |
| Virus‐like particle (VLP) | Modified for targeted delivery, cost‐effective for large‐scale production, avoid off‐target effects | Unknown immunogenicity, poor stability, extravasate from bloodstream, off‐organ accumulation | 166, 169 |
| Virus‐mimicking membrane‐coated nanoparticles | Biosafety, slow elimination, promote the target efficiency | Immunogenicity, unknown escape mechanisms, membrane destabilization | 173, 174 |
| Membrane‐coated vehicles | Biocompatible, improve the circulation time, enhance cell encapsulation efficiency | Perturbations in membrane integrity, limit size of the cargo, technologies problem (cell membrane coating, fresh isolation of native cell membranes, co‐extrusion with drug carriers | 227 |
| Extracellular vesicle (EVs) | Low‐toxicity, less immunogenicity, biocompatibility, high penetration and retention effect, conveniently cross physiological barriers, intrinsic homing properties, suitable for targeting modification | Heterogeneity of EVs, RNA‐loading capacity is low, low yields, characteristic complexity, lack of access to large‐scale production | 216, 228, 229 |
mRNA must cross several tissue barrier, extracellular barrier, intracellular endosomal escape, and intracellular immunity before it arrives at the intracellular target site. Therefore, it remains challenging to deliver mRNA precisely into the target cells. Most of the existing LNPs are accumulated in the liver after being injected into the body. Therefore, how to precisely delivery sufficient mRNA to target organs when using intramuscular LNP‐RNA is of great importance. This problem can be overcome by selective organ targeting (SORT) strategies. The key to organ‐specific delivery is manipulating the internal or external charges of LNPs. For example, altering the disulfide bonds between the long lipid chains leads to selective accumulation of mRNA formulations in the liver. 230 While increasing the percentage of permanently positively charged DDAB or EPC can shift tissue tropism from the liver to the lung and introducing the negatively charged SORT molecule 1,2‐dioleoyl‐sn‐glycero‐3‐phosphate (18PA) into LNP can lead to spleen‐specific delivery. 231 Alternatively, mimetic lipids can be used to synthesize LNP for organ‐targeted delivery. For example, certain neurotransmitters can cross the BBB, and they encapsulate its derived lipidoids to gain access throughout the brain. 232 Both the modulation of lipid chemistry and nanoparticle fractions allow for organ selectivity, offering the potential for the development of next‐generation mRNA‐targeted delivery vectors.
However, the safety of repeated administration of LNPs still remains a challenge for their clinical translation. Exosomes possess great potential as a new type of carrier for mRNA therapy. In particular, exosomes are naturally occurring nanovesicles, making them a safe vehicle for targeted drug delivery. Since exosomes express the surface proteins and receptors of the parent cell, they are advantageous in targeting recipient cells in the same tissue, such as the use of lung spheroid cell‐derived exosome for targeted delivery of vaccines to the lung tissue, as we mentioned in Figure 6. In addition, exosomes can be genetically engineered, chemically surface‐modified, or enzymatically engineered to introduce peptides or proteins to enhance its targeting. 216 , 228 Due to their high biocompatibility, bioavailability, and ability to cross biological barriers, there is increasing interest in exosomes as promising carriers for mRNA drug delivery. Exosomes can either encapsulate the transcripted mRNA from parent cells or be directly loaded with synthetic mRNA by conventional transfection methods. In addition, exosomes exhibit great stability in blood, allowing them to transport over long distances in vivo under both physiological and pathological conditions. More importantly, exosomes can be lyophilized as dry powder for inhalation. Accumulating evidence has demonstrated the efficacy and stability of lyophilized lung‐exos at room temperature for up to 28 days. 210 Current mRNA vaccine products require storage temperatures as low as −20°C or −70°C to ensure stability. Therefore, the use of exosomes as lyophilizable vaccines can achieve a longer shelf life, reduce transport costs, and facilitate vaccine distribution.
FIGURE 6.
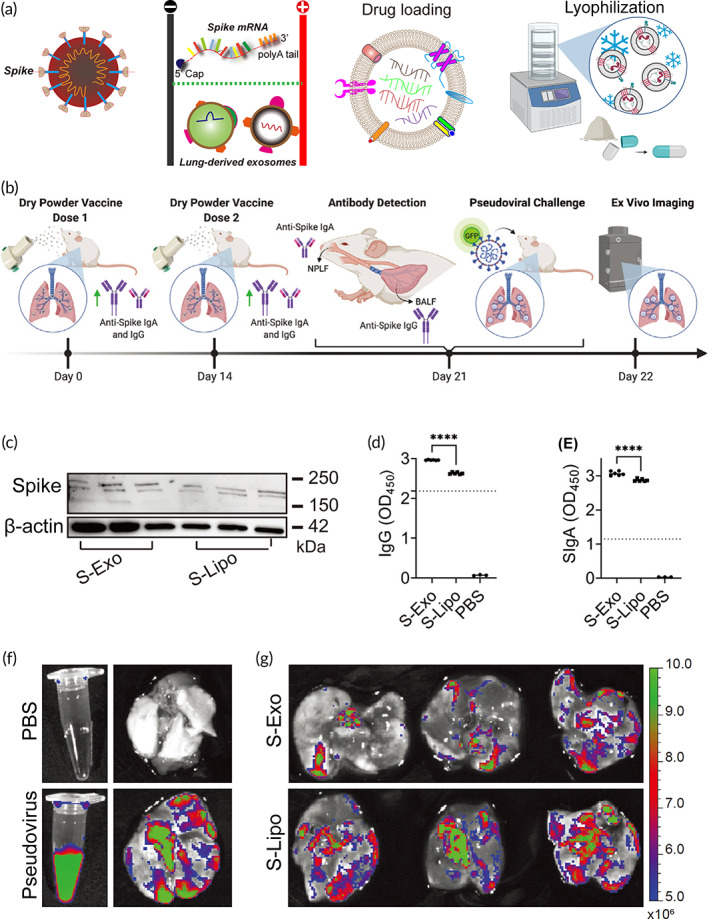
Inhalation of the exosome‐mediated mRNA vaccine elicits immune responses. (a) Pulmonary cell‐derived exosomes are loaded with SARS‐CoV‐2S‐protein mRNA by electro‐transferred and lyophilized to obtain a dry powder formulation. (b) Schematic representation of inhalation process of exosome mRNA, antibody production against S‐protein, and pseudoviral challenge. (c) Identification of spike protein expression in the upper and lower respiratory tracts by western blotting analysis. (d, e) ELISA shows significantly elevated IgG and IgM in both S‐Exos and S‐Lipos. (f) Images of lung tissues from PBS or pseudovirus infection. (g) Biodistribution of the clearance of pseudoviral infection after inhalation of S‐Exo or S‐Lipo vaccine. Source: Reproduced with permission. 210 Copyright 2022, COVID‐19 resource center, Elsevier.
One of the leading ongoing challenges for exosomes is the commercially available methods. However, their active substances in exosomes are relatively complex and face a certain degree of inherent biological variation, which may lead to product heterogeneity between different batches. Cell culture is currently one of the main sources of exosomes, while its production and scale are still challenging. In terms of scale‐up production, several strategies have been developed, including the use of bioreactors, 233 while the cost/yield is still economically unsuitable for clinical application. Alternative sources, including blood, milk, and plant, to obtain exosomes or exosome‐like vesicles may allow for lower production costs and shorter production times. 234 , 235 In the future, exosomes are expected to bring breakthroughs in the field of mRNA drugs when technical difficulties are solved.
For the clinical application of nano‐mediated mRNA delivery, it is also necessary to optimize the production and purification methods to obtain a population of suitable size and uniform distribution. In addition, the dose of mRNA delivered by nano‐mediated approaches should also be optimized based on the material. The size, quantity, and loading efficiency should be quantified. At present, most of the research on nanoparticles‐based mRNA therapy remains in the lab bench, further research is needed at the patient's bedside.
AUTHOR CONTRIBUTIONS
De‐feng Li: Conceptualization (lead); writing – original draft (lead); writing – review and editing (lead). Qi‐song Liu: Investigation (equal); methodology (equal). Mei‐feng Yang: Investigation (equal); methodology (equal). Hao‐ming Xu: Data curation (equal); investigation (equal); resources (equal). Min‐zheng Zhu: Investigation (equal); methodology (equal). Yuan Zhang: Resources (equal); validation (equal). Jing Xu: Project administration (equal); supervision (equal); visualization (equal). Cheng‐mei Tian: Data curation (equal); formal analysis (equal). Jun Yao: Supervision (equal); validation (equal); writing – review and editing (equal). Li‐sheng Wang: Project administration (equal); supervision (equal); writing – review and editing (equal). Yu‐jie Liang: Conceptualization (equal); funding acquisition (equal); supervision (equal); writing – review and editing (equal).
FUNDING INFORMATION
This work was funded by the Science and Technology Innovation Committee of Shenzhen (No. JCYJ20200109150700942, No. CXFZ20211020164543006), Shenzhen Clinical Research Center for Mental Disorders (20210617155253001), the Guangdong Basic and Applied Basic Research Foundation (2020A1515011581), the Key‐Area Research and Development Program of Guangdong Province (2019B030335001), the Shenzhen Fund for Guangdong Provincial High Level Clinical Key Specialties (No. SZGSP013), the Shenzhen Key Medical Discipline Construction Fund (No. SZXK042), and Shenzhen Science and Technology Projects (No. KCXFZ20211020164543006, No. JCYJ20150403101028164, No. JCYC20170307100911479, No. JCYJ20190807145617113 and JCYJ20210324113802006).
CONFLICT OF INTEREST
The authors declare no competing interests.
PEER REVIEW
The peer review history for this article is available at https://publons.com/publon/10.1002/btm2.10492.
Li D, Liu Q, Yang M, et al. Nanomaterials for mRNA‐based therapeutics: Challenges and opportunities. Bioeng Transl Med. 2023;8(3):e10492. doi: 10.1002/btm2.10492
De‐feng Li, Qi‐song Liu, and Mei‐feng Yang contributed equally to this study.
Contributor Information
Jun Yao, Email: yao.jun@szhospital.com.
Li‐sheng Wang, Email: wanglsszrmyy@163.com.
Yu‐jie Liang, Email: liangyjie@126.com.
DATA AVAILABILITY STATEMENT
There are no supplementary data available for this article.
REFERENCES
- 1. Iavarone C, O'Hagan DT, Yu D, et al. Mechanism of action of mRNA‐based vaccines. Expert Rev Vaccines. 2017;16:871‐881. [DOI] [PubMed] [Google Scholar]
- 2. Prieve MG, Harvie P, Monahan SD, et al. Targeted mRNA therapy for ornithine transcarbamylase deficiency. Mol Ther. 2018;26:801‐813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Tsakiri M, Zivko C, Demetzos C, Mahairaki V. Lipid‐based nanoparticles and RNA as innovative neuro‐therapeutics. Front Pharmacol. 2022;13:900610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Magadum A, Kaur K, Zangi L. mRNA‐based protein replacement therapy for the heart. Mol Ther. 2019;27:785‐793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Wolff JA, Malone RW, Williams P, et al. Direct gene transfer into mouse muscle in vivo. Science. 1990;247:1465‐1468. [DOI] [PubMed] [Google Scholar]
- 6. Kowalski PS, Rudra A, Miao L, Anderson DG. Delivering the messenger: advances in technologies for rherapeutic mRNA delivery. Mol Ther. 2019;27:710‐728. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Sullenger BA, Nair S. From the RNA world to the clinic. Science. 2016;352:1417‐1420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Mukai H, Ogawa K, Kato N, Kawakami S. Recent advances in lipid nanoparticles for delivery of nucleic acid, mRNA, and gene editing‐based therapeutics. Drug Metab Pharmacokinet. 2022;44:100450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Li M, Li Y, Li S, et al. The nano delivery systems and applications of mRNA. Eur J Med Chem. 2022;227:113910. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Yan Y, Liu XY, Lu A, Wang XY, Jiang LX, Wang JC. Non‐viral vectors for RNA delivery. J Control Release. 2022;342:241‐279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Byun MJ, Lim J, Kim SN, et al. Advances in nanoparticles for effective delivery of RNA therapeutics. Biochip J. 2022;16:128‐145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Li B, Zhang X, Dong Y. Nanoscale platforms for messenger RNA delivery. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2019;11:e1530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Wang Y, Zhang R, Tang L, Yang L. Nonviral delivery systems of mRNA vaccines for cancer gene therapy. Pharmaceutics. 2022;14:512. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Ibba ML, Ciccone G, Esposito CL, Catuogno S, Giangrande PH. Advances in mRNA non‐viral delivery approaches. Adv Drug Deliv Rev. 2021;177:113930. [DOI] [PubMed] [Google Scholar]
- 15. Steinle H, Weber J, Stoppelkamp S, et al. Delivery of synthetic mRNAs for tissue regeneration. Adv Drug Deliv Rev. 2021;179:114007. [DOI] [PubMed] [Google Scholar]
- 16. Xu S, Yang K, Li R, Zhang L. mRNA vaccine era‐mechanisms, drug platform and clinical prospection. Int J Mol Sci. 2020;21:6582. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Halloy F, Biscans A, Bujold KE, et al. Innovative developments and emerging technologies in RNA therapeutics. RNA Biol. 2022;19:313‐332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Ouranidis A, Vavilis T, Mandala E, et al. mRNA therapeutic modalities design, formulation and manufacturing under pharma 4.0 principles. Biomedicine. 2021;10:50. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Beck JD, Reidenbach D, Salomon N, et al. mRNA therapeutics in cancer immunotherapy. Mol Cancer. 2021;20:69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Yin H, Kanasty RL, Eltoukhy AA, Vegas AJ, Dorkin JR, Anderson DG. Non‐viral vectors for gene‐based therapy. Nat Rev Genet. 2014;15:541‐555. [DOI] [PubMed] [Google Scholar]
- 21. Lundstrom K. Viral vectors in gene therapy. Diseases. 2018;6:42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22. Paul A, Collins MG, Lee HY. Gene therapy: the next‐generation therapeutics and their delivery approaches for neurological disorders. Front Genome Ed. 2022;4:899209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23. Finer M, Glorioso J. A brief account of viral vectors and their promise for gene therapy. Gene Ther. 2017;24:1‐2. [DOI] [PubMed] [Google Scholar]
- 24. Prasad V. Immunotherapy: Tisagenlecleucel ‐ the first approved CAR‐T‐cell therapy: implications for payers and policy makers. Nat Rev Clin Oncol. 2018;15:11‐12. [DOI] [PubMed] [Google Scholar]
- 25. Waehler R, Russell SJ, Curiel DT. Engineering targeted viral vectors for gene therapy. Nat Rev Genet. 2007;8:573‐587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Thomas CE, Ehrhardt A, Kay MA. Progress and problems with the use of viral vectors for gene therapy. Nat Rev Genet. 2003;4:346‐358. [DOI] [PubMed] [Google Scholar]
- 27. Billingsley MM, Singh N, Ravikumar P, Zhang R, June CH, Mitchell MJ. Ionizable lipid nanoparticle‐mediated mRNA delivery for human CAR T cell engineering. Nano Lett. 2020;20:1578‐1589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28. Islam MA, Reesor EK, Xu Y, et al. Biomaterials for mRNA delivery. Biomater Sci. 2015;3:1519‐1533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Pack DW, Hoffman AS, Pun S, Stayton PS. Design and development of polymers for gene delivery. Nat Rev Drug Discov. 2005;4:581‐593. [DOI] [PubMed] [Google Scholar]
- 30. Sahin U, Oehm P, Derhovanessian E, et al. An RNA vaccine drives immunity in checkpoint‐inhibitor‐treated melanoma. Nature. 2020;585:107‐112. [DOI] [PubMed] [Google Scholar]
- 31. Anselmo AC, Mitragotri S. Nanoparticles in the clinic: an update. Bioeng Transl Med. 2019;4:e10143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Cafri G, Gartner JJ, Zaks T, et al. mRNA vaccine‐induced neoantigen‐specific T cell immunity in patients with gastrointestinal cancer. J Clin Invest. 2020;130:5976‐5988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Bozec A, Culié D, Poissonnet G, Demard F, Dassonville O. Current therapeutic strategies in paients with oropharyngeal squamous cell carcinoma: impact of the tumor HPV status. Cancers (Basel). 2021;13:5456. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. III HAB, Patel MR, Cho DC, et al. A phase 1, open‐label, multicenter study to assess the safety, tolerability, and immunogenicity of mRNA‐4157 alone in subjects with resected solid tumors and in combination with pembrolizumab in subjects with unresectable solid tumors (Keynote‐603). J Glob Oncol. 2019;5:93. [Google Scholar]
- 35. Kirchhammer N, Trefny MP, Auf der Maur P, Läubli H, Zippelius A. Combination cancer immunotherapies: emerging treatment strategies adapted to the tumor microenvironment. Sci Transl Med. 2022;14:eabo3605. [DOI] [PubMed] [Google Scholar]
- 36. Aldrich C, Leroux‐Roels I, Huang KB, et al. Proof‐of‐concept of a low‐dose unmodified mRNA‐based rabies vaccine formulated with lipid nanoparticles in human volunteers: a phase 1 trial. Vaccine. 2021;39:1310‐1318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Fotin‐Mleczek M, Zanzinger K, Heidenreich R, et al. Highly potent mRNA based cancer vaccines represent an attractive platform for combination therapies supporting an improved therapeutic effect. J Gene Med. 2012;14:428‐439. [DOI] [PubMed] [Google Scholar]
- 38. Schneider K, Grønhøj C, Hahn CH, von Buchwald C. Therapeutic human papillomavirus vaccines in head and neck cancer: a systematic review of current clinical trials. Vaccine. 2018;36:6594‐6605. [DOI] [PubMed] [Google Scholar]
- 39. John S, Yuzhakov O, Woods A, et al. Multi‐antigenic human cytomegalovirus mRNA vaccines that elicit potent humoral and cell‐mediated immunity. Vaccine. 2018;36:1689‐1699. [DOI] [PubMed] [Google Scholar]
- 40. Feldman RA, Fuhr R, Smolenov I, et al. mRNA vaccines against H10N8 and H7N9 influenza viruses of pandemic potential are immunogenic and well tolerated in healthy adults in phase 1 randomized clinical trials. Vaccine. 2019;37:3326‐3334. [DOI] [PubMed] [Google Scholar]
- 41. Bahl K, Senn JJ, Yuzhakov O, et al. Preclinical and clinical demonstration of immunogenicity by mRNA vaccines against H10N8 and H7N9 influenza viruses. Mol Ther. 2017;25:1316‐1327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. de Alwis R, Gan ES, Chen S, et al. A single dose of self‐transcribing and replicating RNA‐based SARS‐CoV‐2 vaccine produces protective adaptive immunity in mice. Mol Ther. 2021;29:1970‐1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43. Rauch S, Roth N, Schwendt K, Fotin‐Mleczek M, Mueller SO, Petsch B. mRNA‐based SARS‐CoV‐2 vaccine candidate CVnCoV induces high levels of virus‐neutralising antibodies and mediates protection in rodents. NPJ Vaccines. 2021;6:57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44. Vogel AB, Kanevsky I, Che Y, et al. BNT162b vaccines protect rhesus macaques from SARS‐CoV‐2. Nature. 2021;592:283‐289. [DOI] [PubMed] [Google Scholar]
- 45. Zhang NN, Li XF, Deng YQ, et al. A thermostable mRNA vaccine against COVID‐19. Cell. 2020;182:1271‐1283.e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Baden LR, El Sahly HM, Essink B, et al. Efficacy and safety of the mRNA‐1273 SARS‐CoV‐2 vaccine. N Engl J Med. 2021;384:403‐416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Corbett KS, Edwards DK, Leist SR, et al. SARS‐CoV‐2 mRNA vaccine design enabled by prototype pathogen preparedness. Nature. 2020;586:567‐571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48. Hou X, Zaks T, Langer R, Dong Y. Lipid nanoparticles for mRNA delivery. Nat Rev Mater. 2021;6:1078‐1094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49. Gomez‐Aguado I, Rodriguez‐Castejon J, Vicente‐Pascual M, et al. Nanomedicines to deliver mRNA: state of the art and future perspectives. Nanomaterials (Basel). 2020;10:364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50. Cullis PR, Hope MJ. Lipid nanoparticle systems for enabling gene therapies. Mol Ther. 2017;25:1467‐1475. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Jackson LA, Anderson EJ, Rouphael NG, et al. An mRNA vaccine against SARS‐CoV‐2 ‐ preliminary report. N Engl J Med. 2020;383:1920‐1931. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Kaczmarek JC, Kowalski PS, Anderson DG. Advances in the delivery of RNA therapeutics: from concept to clinical reality. Genome Med. 2017;9:60. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Gupta A, Kafetzis KN, Tagalakis AD, Yu‐Wai‐Man C. RNA therapeutics in ophthalmology ‐ translation to clinical trials. Exp Eye Res. 2021;205:108482. [DOI] [PubMed] [Google Scholar]
- 54. Mollocana‐Lara EC, Ni M, Agathos SN, Gonzales‐Zubiate FA. The infinite possibilities of RNA therapeutics. J Ind Microbiol Biotechnol. 2021;48:9‐10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines ‐ a new era in vaccinology. Nat Rev Drug Discov. 2018;17:261‐279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Sahu I, Haque A, Weidensee B, et al. Recent developments in mRNA‐based protein supplementation therapy to target lung diseases. Mol Ther. 2019;27:803‐823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Qin S, Tang X, Chen Y, et al. mRNA‐based therapeutics: powerful and versatile tools to combat diseases. Signal Transduct Target Ther. 2022;7:166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Brenner S, Jacob F, Meselson M. An unstable intermediate carrying information from genes to ribosomes for protein synthesis. Nature. 1961;190:576‐581. [DOI] [PubMed] [Google Scholar]
- 59. Heiser A, Coleman D, Dannull J, et al. Autologous dendritic cells transfected with prostate‐specific antigen RNA stimulate CTL responses against metastatic prostate tumors. J Clin Invest. 2002;109:409‐417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Patel R, Kaki M, Potluri VS, Kahar P, Khanna D. A comprehensive review of SARS‐CoV‐2 vaccines: Pfizer Moderna & Johnson & Johnson. Hum Vaccin Immunother. 2022;18:2002083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Strenkowska M, Grzela R, Majewski M, et al. Cap analogs modified with 1,2‐dithiodiphosphate moiety protect mRNA from decapping and enhance its translational potential. Nucleic Acids Res. 2016;44:9578‐9590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Eckmann CR, Rammelt C, Wahle E. Control of poly(A) tail length. Wiley Interdiscip Rev RNA. 2011;2:348‐361. [DOI] [PubMed] [Google Scholar]
- 63. Zinckgraf JW, Silbart LK. Modulating gene expression using DNA vaccines with different 3'‐UTRs influences antibody titer, seroconversion and cytokine profiles. Vaccine. 2003;21:1640‐1649. [DOI] [PubMed] [Google Scholar]
- 64. Hinnebusch AG, Ivanov IP, Sonenberg N. Translational control by 5′‐untranslated regions of eukaryotic mRNAs. Science. 2016;352:1413‐1416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65. Adibzadeh S, Fardaei M, Takhshid MA, et al. Enhancing stability of destabilized green fluorescent protein using chimeric mRNA containing human Beta‐globin 5′ and 3' untranslated regions. Avicenna J Med Biotechnol. 2019;11:112‐117. [PMC free article] [PubMed] [Google Scholar]
- 66. Sonenberg N, Hinnebusch AG. Regulation of translation initiation in eukaryotes: mechanisms and biological targets. Cell. 2009;136:731‐745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Beckert B, Masquida B. Synthesis of RNA by in vitro transcription. Methods Mol Biol. 2011;703:29‐41. [DOI] [PubMed] [Google Scholar]
- 68. Beissert T, Perkovic M, Vogel A, et al. A trans‐amplifying RNA vaccine strategy for induction of potent protective immunity. Mol Ther. 2020;28:119‐128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Trepotec Z, Lichtenegger E, Plank C, Aneja MK, Rudolph C. Delivery of mRNA therapeutics for the treatment of hepatic diseases. Mol Ther. 2019;27:794‐802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Tang X, Zhang S, Fu R, et al. Therapeutic prospects of mRNA‐based gene therapy for glioblastoma. Front Oncol. 2019;9:1208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Zhang C, Maruggi G, Shan H, Li J. Advances in mRNA vaccines for infectious diseases. Front Immunol. 2019;10:594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Martini PGV, Guey LT. A new era for rare genetic diseases: messenger RNA therapy. Hum Gene Ther. 2019;30:1180‐1189. [DOI] [PubMed] [Google Scholar]
- 73. Berraondo P, Martini PGV, Avila MA, Fontanellas A. Messenger RNA therapy for rare genetic metabolic diseases. Gut. 2019;68:1323‐1330. [DOI] [PubMed] [Google Scholar]
- 74. Granot‐Matok Y, Kon E, Dammes N, Mechtinger G, Peer D. Therapeutic mRNA delivery to leukocytes. J Control Release. 2019;305:165‐175. [DOI] [PubMed] [Google Scholar]
- 75. Pastor F, Berraondo P, Etxeberria I, et al. An RNA toolbox for cancer immunotherapy. Nat Rev Drug Discov. 2018;17:751‐767. [DOI] [PubMed] [Google Scholar]
- 76. Scheel B, Teufel R, Probst J, et al. Toll‐like receptor‐dependent activation of several human blood cell types by protamine‐condensed mRNA. Eur J Immunol. 2005;35:1557‐1566. [DOI] [PubMed] [Google Scholar]
- 77. Thran M, Mukherjee J, Ponisch M, et al. mRNA mediates passive vaccination against infectious agents, toxins, and tumors. EMBO Mol Med. 2017;9:1434‐1447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78. Rybakova Y, Kowalski PS, Huang Y, et al. mRNA delivery for therapeutic anti‐HER2 antibody expression In vivo. Mol Ther. 2019;27:1415‐1423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79. Roth GA, Mensah GA, Johnson CO, et al. Global burden of cardiovascular diseases and risk factors, 1990‐2019: update from the GBD 2019 study. J Am Coll Cardiol. 2020;76:2982‐3021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80. GBD 2019 Diseases and Injuries Collaborators . Global burden of 369 diseases and injuries in 204 countries and territories, 1990‐2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet. 2020;396:1204‐1222. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81. Kaur K, Zangi L. Modified mRNA as a therapeutic tool for the heart. Cardiovasc Drugs Ther. 2020;34:871‐880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82. Kaur K, Sultana N, Hadas Y, et al. Delivery of modified mRNA in a myocardial infarction mouse model. J Vis Exp. 2020;160:e60832. [DOI] [PubMed] [Google Scholar]
- 83. Lockney NA, Zhang M, Lu Y, et al. Pyruvate kinase muscle Isoenzyme 2 (PKM2) expression is associated with overall survival in pancreatic ductal adenocarcinoma. J Gastrointest Cancer. 2015;46:390‐398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84. Magadum A, Singh N, Kurian AA, et al. Pkm2 regulates cardiomyocyte cell cycle and promotes cardiac regeneration. Circulation. 2020;141:1249‐1265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85. Huang CL, Leblond AL, Turner EC, et al. Synthetic chemically modified mrna‐based delivery of cytoprotective factor promotes early cardiomyocyte survival post‐acute myocardial infarction. Mol Pharm. 2015;12:991‐996. [DOI] [PubMed] [Google Scholar]
- 86. Zangi L, Oliveira MS, Ye LY, et al. Insulin‐like growth factor 1 receptor‐dependent pathway drives epicardial adipose tissue formation after myocardial injury. Circulation. 2017;135:59‐72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87. Zangi L, Lui KO, von Gise A, et al. Modified mRNA directs the fate of heart progenitor cells and induces vascular regeneration after myocardial infarction. Nat Biotechnol. 2013;31:898‐907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88. Strauss KA, Ahlfors CE, Soltys K, et al. Crigler‐Najjar syndrome type 1: pathophysiology, natural history, and therapeutic frontier. Hepatology. 2020;71:1923‐1939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89. Ebrahimi A, Rahim F. Crigler‐Najjar syndrome: current perspectives and the application of clinical genetics. Endocr Metab Immune Disord Drug Targets. 2018;18:201‐211. [DOI] [PubMed] [Google Scholar]
- 90. Apgar JF, Tang JP, Singh P, et al. Quantitative systems pharmacology model of hUGT1A1‐modRNA encoding for the UGT1A1 enzyme to treat Crigler‐Najjar syndrome type 1. CPT Pharmacometrics Syst Pharmacol. 2018;7:404‐412. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91. Lu Y, Wang LR, Lee J, et al. The unfolded protein response to PI*Z alpha‐1 antitrypsin in human hepatocellular and murine models. Hepatol Commun. 2022;6:2354‐2367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92. Huntington JA, Read RJ, Carrell RW. Structure of a serpin‐protease complex shows inhibition by deformation. Nature. 2000;407:923‐926. [DOI] [PubMed] [Google Scholar]
- 93. Afsharinasab M, Akbari AH, Mirzaei V, et al. The investigation of the frequency of the alpha‐1‐antitrypsin phenotype in patients with liver cirrhosis. Horm Mol Biol Clin Investig. 2022;43:397‐403. [DOI] [PubMed] [Google Scholar]
- 94. Patel D, Teckman JH. Alpha‐1‐antitrypsin deficiency liver disease. Clin Liver Dis. 2018;22:643‐655. [DOI] [PubMed] [Google Scholar]
- 95. Connolly B, Isaacs C, Cheng L, Asrani KH, Subramanian RR. SERPINA1 mRNA as a treatment for Alpha‐1 antitrypsin deficiency. J Nucleic Acids. 2018;2018:8247935‐8247937. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96. Liu‐Chen S, Connolly B, Cheng L, Subramanian RR, Han Z. mRNA treatment produces sustained expression of enzymatically active human ADAMTS13 in mice. Sci Rep. 2018;8:7859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97. Roseman DS, Khan T, Rajas F, et al. G6PC mRNA therapy positively regulates fasting blood glucose and decreases liver abnormalities in a mouse model of glycogen storage disease 1a. Mol Ther. 2018;26:814‐821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98. Jiang L, Berraondo P, Jerico D, et al. Systemic messenger RNA as an etiological treatment for acute intermittent porphyria. Nat Med. 2018;24:1899‐1909. [DOI] [PubMed] [Google Scholar]
- 99. DeRosa F, Guild B, Karve S, et al. Therapeutic efficacy in a hemophilia B model using a biosynthetic mRNA liver depot system. Gene Ther. 2016;23:699‐707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100. Mays LE, Ammon‐Treiber S, Mothes B, et al. Modified Foxp3 mRNA protects against asthma through an IL‐10‐dependent mechanism. J Clin Invest. 2013;123:1216‐1228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101. Zhu X, Yin L, Theisen M, et al. Systemic mRNA therapy for the treatment of Fabry disease: preclinical studies in wild‐type mice, Fabry mouse model, and wild‐type non‐human primates. Am J Hum Genet. 2019;104:625‐637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102. Golombek S, Pilz M, Steinle H, et al. Intradermal delivery of synthetic mRNA using hollow microneedles for efficient and rapid production of exogenous proteins in skin. Mol Ther Nucleic Acids. 2018;11:382‐392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103. Phua KK, Leong KW, Nair SK. Transfection efficiency and transgene expression kinetics of mRNA delivered in naked and nanoparticle format. J Control Release. 2013;166:227‐233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104. Elangovan S, Khorsand B, Do AV, et al. Chemically modified RNA activated matrices enhance bone regeneration. J Control Release. 2015;218:22‐28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105. Balmayor ER, Geiger JP, Aneja MK, et al. Chemically modified RNA induces osteogenesis of stem cells and human tissue explants as well as accelerates bone healing in rats. Biomaterials. 2016;87:131‐146. [DOI] [PubMed] [Google Scholar]
- 106. Andre FE, Booy R, Bock HL, et al. Vaccination greatly reduces disease, disability, death and inequity worldwide. Bull World Health Organ. 2008;86:140‐146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107. Minor PD. Live attenuated vaccines: historical successes and current challenges. Virology. 2015;479–480:379‐392. [DOI] [PubMed] [Google Scholar]
- 108. Minor PD. The polio‐eradication programme and issues of the end game. J Gen Virol. 2012;93:457‐474. [DOI] [PubMed] [Google Scholar]
- 109. Rubin LG, Levin MJ, Ljungman P, et al. 2013 IDSA clinical practice guideline for vaccination of the immunocompromised host. Clin Infect Dis. 2014;58:e44‐e100. [DOI] [PubMed] [Google Scholar]
- 110. Li C, Lee A, Grigoryan L, et al. Mechanisms of innate and adaptive immunity to the Pfizer‐BioNTech BNT162b2 vaccine. Nat Immunol. 2022;23:543‐555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111. Awasthi S, Friedman HM. An mRNA vaccine to prevent genital herpes. Transl Res. 2022;242:56‐65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112. Jawalagatti V, Kirthika P, Lee JH. Oral mRNA vaccines against infectious diseases—a bacterial perspective [invited]. Front Immunol. 2022;13:884862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113. Duan LJ, Wang Q, Zhang C, Yang DX, Zhang XY. Potentialities and challenges of mRNA vaccine in cancer immunotherapy. Front Immunol. 2022;13:923647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114. Rosa SS, Prazeres DMF, Azevedo AM, Marques MPC. mRNA vaccines manufacturing: challenges and bottlenecks. Vaccine. 2021;39:2190‐2200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115. He Q, Gao H, Tan D, Zhang H, Wang JZ. mRNA cancer vaccines: advances, trends and challenges. Acta Pharm Sin B. 2022;12:2969‐2989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116. Wang Y, Zhang Z, Luo J, Han X, Wei Y, Wei X. mRNA vaccine: a potential therapeutic strategy. Mol Cancer. 2021;20:33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117. Wadhwa A, Aljabbari A, Lokras A, Foged C, Thakur A. Opportunities and challenges in the delivery of mRNA‐based vaccines. Pharmaceutics. 2020;12:102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118. Nitika WJ, Hui AM. The development of mRNA vaccines for infectious diseases: recent updates. Infect Drug Resist. 2021;14:5271‐5285. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119. Huff AL, Jaffee EM, Zaidi N. Messenger RNA vaccines for cancer immunotherapy: progress promotes promise. J Clin Invest. 2022;132:e156211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120. Wesselhoeft RA, Kowalski PS, Anderson DG. Engineering circular RNA for potent and stable translation in eukaryotic cells. Nat Commun. 2018;9:2629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121. Qu L, Yi Z, Shen Y, et al. Circular RNA vaccines against SARS‐CoV‐2 and emerging variants. Cell. 2022;185:1728‐1744.e16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122. Liu X, Zhang Y, Zhou S, Dain L, Mei L, Zhu G. Circular RNA: an emerging frontier in RNA therapeutic targets, RNA therapeutics, and mRNA vaccines. J Control Release. 2022;348:84‐94. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123. Abdolmaleki G, Taheri MA, Paridehpour S, et al. A comparison between SARS‐CoV‐1 and SARS‐CoV2: an update on current COVID‐19 vaccines. Daru. 2022;30:379‐406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124. Talukder B, van Loon GW, Hipel KW. Planetary health & COVID‐19: a multi‐perspective investigation. One Health. 2022;15:100416. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125. Wiersinga WJ, Rhodes A, Cheng AC, Peacock SJ, Prescott HC. Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID‐19): a review. JAMA. 2020;324:782‐793. [DOI] [PubMed] [Google Scholar]
- 126. Corlett RT, Primack RB, Devictor V, et al. Impacts of the coronavirus pandemic on biodiversity conservation. Biol Conserv. 2020;246:108571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127. Farnudian‐Habibi A, Mirjani M, Montazer V, et al. Review on approved and inprogress COVID‐19 vaccines. Iran J Pharm Res. 2022;21:e124228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128. Bruxvoort KJ, Sy LS, Qian L, et al. Real‐world effectiveness of the mRNA‐1273 vaccine against COVID‐19: interim results from a prospective observational cohort study. Lancet Reg Health Am. 2022;6:100134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129. Chodick G, Tene L, Rotem RS, et al. The effectiveness of the two‐dose BNT162b2 vaccine: analysis of real‐world data. Clin Infect Dis. 2022;74:472‐478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130. Lauring AS, Tenforde MW, Chappell JD, et al. Clinical severity of, and effectiveness of mRNA vaccines against, covid‐19 from omicron, delta, and alpha SARS‐CoV‐2 variants in the United States: prospective observational study. BMJ. 2022;376:e069761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131. Chen J, Chen J, Xu Q. Current developments and challenges of mRNA vaccines. Annu Rev Biomed Eng. 2022;24:85‐109. [DOI] [PubMed] [Google Scholar]
- 132. Eralp Y. Application of mRNA technology in cancer therapeutics. Vaccines (Basel). 2022;10:1262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133. Sobhani N, Scaggiante B, Morris R, et al. Therapeutic cancer vaccines: from biological mechanisms and engineering to ongoing clinical trials. Cancer Treat Rev. 2022;109:102429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134. Deng Z, Tian Y, Song J, An G, Yang P. mRNA vaccines: the Dawn of a new era of cancer immunotherapy. Front Immunol. 2022;13:887125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135. Papachristofilou A, Hipp MM, Klinkhardt U, et al. Phase Ib evaluation of a self‐adjuvanted protamine formulated mRNA‐based active cancer immunotherapy, BI1361849 (CV9202), combined with local radiation treatment in patients with stage IV non‐small cell lung cancer. J Immunother Cancer. 2019;7:38. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136. Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392:245‐252. [DOI] [PubMed] [Google Scholar]
- 137. Palucka K, Banchereau J. Dendritic‐cell‐based therapeutic cancer vaccines. Immunity. 2013;39:38‐48. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138. Batich KA, Reap EA, Archer GE, et al. Long‐term survival in glioblastoma with cytomegalovirus pp65‐targeted vaccination. Clin Cancer Res. 2017;23:1898‐1909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139. Hockemeyer D, Wang H, Kiani S, et al. Genetic engineering of human pluripotent cells using TALE nucleases. Nat Biotechnol. 2011;29:731‐734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140. Mali P, Yang L, Esvelt KM, et al. RNA‐guided human genome engineering via Cas9. Science. 2013;339:823‐826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141. Weng Y, Li C, Yang T, et al. The challenge and prospect of mRNA therapeutics landscape. Biotechnol Adv. 2020;40:107534. [DOI] [PubMed] [Google Scholar]
- 142. Badia R, Pauls E, Riveira‐Munoz E, Clotet B, Esté JA, Ballana E. Zinc finger endonuclease targeting PSIP1 inhibits HIV‐1 integration. Antimicrob Agents Chemother. 2014;58:4318‐4327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143. Quintana‐Bustamante O, Fananas‐Baquero S, Orman I, et al. Gene editing of PKLR gene in human hematopoietic progenitors through 5′ and 3' UTR modified TALEN mRNA. PLoS One. 2019;14:e0223775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144. Schmidt JK, Reynolds MR, Golos TG, Slukvin II. CRISPR/Cas9 genome editing to create nonhuman primate models for studying stem cell therapies for HIV infection. Retrovirology. 2022;19:17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145. Wahane A, Waghmode A, Kapphahn A, Dhuri K, Gupta A, Bahal R. Role of lipid‐based and polymer‐based non‐viral vectors in nucleic acid delivery for next‐generation gene therapy. Molecules. 2020;25:2866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146. Malone RW, Felgner PL, Verma IM. Cationic liposome‐mediated RNA transfection. Proc Natl Acad Sci U S A. 1989;86:6077‐6081. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147. Lv H, Zhang S, Wang B, Cui S, Yan J. Toxicity of cationic lipids and cationic polymers in gene delivery. J Control Release. 2006;114:100‐109. [DOI] [PubMed] [Google Scholar]
- 148. Das R, Kanjilal P, Medeiros J, et al. What's next after lipid nanoparticles? A perspective on enablers of nucleic acid therapeutics. Bioconjug Chem. 2022;33:1996‐2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149. Tenforde MW, Patel MM, Ginde AA, et al. Effectiveness of SARS‐CoV‐2 mRNA vaccines for preventing Covid‐19 hospitalizations in the United States. medRxiv. 2021;07:21259776. [Google Scholar]
- 150. Patel S, Ashwanikumar N, Robinson E, et al. Naturally‐occurring cholesterol analogues in lipid nanoparticles induce polymorphic shape and enhance intracellular delivery of mRNA. Nat Commun. 2020;11:983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151. Cheng X, Lee RJ. The role of helper lipids in lipid nanoparticles (LNPs) designed for oligonucleotide delivery. Adv Drug Deliv Rev. 2016;99:129‐137. [DOI] [PubMed] [Google Scholar]
- 152. Kalyanram P, Puri A, Gupta A. Thermotropic effects of PEGylated lipids on the stability of HPPH‐encapsulated lipid nanoparticles (LNP). J Therm Anal Calorim. 2022;147:6337‐6348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153. Liu S, Cheng Q, Wei T, et al. Membrane‐destabilizing ionizable phospholipids for organ‐selective mRNA delivery and CRISPR‐Cas gene editing. Nat Mater. 2021;20:701‐710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154. Meyer RA, Hussmann GP, Peterson NC, Santos JL, Tuesca AD. A scalable and robust cationic lipid/polymer hybrid nanoparticle platform for mRNA delivery. Int J Pharm. 2022;611:121314. [DOI] [PubMed] [Google Scholar]
- 155. Veiga N, Goldsmith M, Granot Y, et al. Cell specific delivery of modified mRNA expressing therapeutic proteins to leukocytes. Nat Commun. 2018;9:4493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156. Fenton OS, Kauffman KJ, Kaczmarek JC, et al. Synthesis and biological evaluation of Ionizable lipid materials for the In vivo delivery of messenger RNA to B lymphocytes. Adv Mater. 2017;29:1606944. [DOI] [PubMed] [Google Scholar]
- 157. Fenton OS, Kauffman KJ, McClellan RL, et al. Bioinspired alkenyl amino alcohol Ionizable lipid materials for highly potent In vivo mRNA delivery. Adv Mater. 2016;28:2939‐2943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158. Rosenblum D, Gutkin A, Kedmi R, et al. CRISPR‐Cas9 genome editing using targeted lipid nanoparticles for cancer therapy. Sci Adv. 2020;6:eabc9450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159. Kozma GT, Shimizu T, Ishida T, Szebeni J. Anti‐PEG antibodies: properties, formation, testing and role in adverse immune reactions to PEGylated nano‐biopharmaceuticals. Adv Drug Deliv Rev. 2020;154‐155:163‐175. [DOI] [PubMed] [Google Scholar]
- 160. McSweeney MD, Mohan M, Commins SP, et al. Anaphylaxis to Pfizer/BioNTech mRNA COVID‐19 vaccine in a patient with clinically confirmed PEG allergy. Front Allergy. 2021;2:715844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161. Kedmi R, Ben‐Arie N, Peer D. The systemic toxicity of positively charged lipid nanoparticles and the role of toll‐like receptor 4 in immune activation. Biomaterials. 2010;31:6867‐6875. [DOI] [PubMed] [Google Scholar]
- 162. Wadman M. Public needs to prep for vaccine side effects. Science. 2020;370:1022. [DOI] [PubMed] [Google Scholar]
- 163. Ndeupen S, Qin Z, Jacobsen S, et al. The mRNA‐LNP platform's lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. Science. 2021;24:103479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164. Persano S, Guevara ML, Li Z, et al. Lipopolyplex potentiates anti‐tumor immunity of mRNA‐based vaccination. Biomaterials. 2017;125:81‐89. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165. Van der Jeught K, De Koker S, Bialkowski L, et al. Dendritic cell targeting mRNA Lipopolyplexes combine strong antitumor T‐cell immunity with improved inflammatory safety. ACS Nano. 2018;12:9815‐9829. [DOI] [PubMed] [Google Scholar]
- 166. Rohovie MJ, Nagasawa M, Swartz JR. Virus‐like particles: next‐generation nanoparticles for targeted therapeutic delivery. Bioeng Transl Med. 2017;2:43‐57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167. Yin D, Ling S, Wang D, et al. Targeting herpes simplex virus with CRISPR‐Cas9 cures herpetic stromal keratitis in mice. Nat Biotechnol. 2021;39:567‐577. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168. Yin J, Spillman E, Cheng ES, et al. Brain‐specific lipoprotein receptors interact with astrocyte derived apolipoprotein and mediate neuron‐glia lipid shuttling. Nat Commun. 2021;12:2408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169. Yadav M, Atala A, Lu B. Developing all‐in‐one virus‐like particles for Cas9 mRNA/single guide RNA co‐delivery and aptamer‐containing lentiviral vectors for improved gene expression. Int J Biol Macromol. 2022;209:1260‐1270. [DOI] [PubMed] [Google Scholar]
- 170. Segel M, Lash B, Song J, et al. Mammalian retrovirus‐like protein PEG10 packages its own mRNA and can be pseudotyped for mRNA delivery. Science. 2021;373:882‐889. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171. Liu Y, Luo J, Chen X, Liu W, Chen T. Cell membrane coating technology: a promising strategy for biomedical applications. Nano‐Micro Lett. 2019;11:100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172. Zhou Z, Liu Y, Wu L, Li L, Huang Y. Enhanced nuclear delivery of anti‐cancer drugs using micelles containing releasable membrane fusion peptide and nuclear‐targeting retinoic acid. J Mater Chem B. 2017;5:7175‐7185. [DOI] [PubMed] [Google Scholar]
- 173. Park JH, Mohapatra A, Zhou J, et al. Virus‐mimicking cell membrane‐coated nanoparticles for cytosolic delivery of mRNA. 2022;61:e202113671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174. Park JH, Mohapatra A, Zhou J, et al. Virus‐mimicking cell membrane‐coated nanoparticles for cytosolic delivery of mRNA. Angew Chem Int Ed Engl. 2022;61:e202113671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 175. Kaczmarek JC, Kauffman KJ, Fenton OS, et al. Optimization of a degradable polymer‐lipid nanoparticle for potent systemic delivery of mRNA to the lung endothelium and immune cells. Nano Lett. 2018;18:6449‐6454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 176. Shuai Q, Zhu F, Zhao M, Yan Y. mRNA delivery via non‐viral carriers for biomedical applications. Int J Pharm. 2021;607:121020. [DOI] [PubMed] [Google Scholar]
- 177. Rejman J, Tavernier G, Bavarsad N, Demeester J, de Smedt SC. mRNA transfection of cervical carcinoma and mesenchymal stem cells mediated by cationic carriers. J Control Release. 2010;147:385‐391. [DOI] [PubMed] [Google Scholar]
- 178. Li M, Zhao M, Fu Y, et al. Enhanced intranasal delivery of mRNA vaccine by overcoming the nasal epithelial barrier via intra‐ and paracellular pathways. J Control Release. 2016;228:9‐19. [DOI] [PubMed] [Google Scholar]
- 179. Knauer N, Arkhipova V, Li G, et al. In vitro validation of the therapeutic potential of dendrimer‐based nanoformulations against tumor stem cells. Int J Mol Sci. 2022;23:5691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180. Chen J, Zhu D, Liu X, Peng L. Amphiphilic dendrimer vectors for RNA delivery: state‐of‐the‐art and future perspective. Acc Mater Res. 2022;3:484‐497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181. Xiong H, Liu S, Wei T, Cheng Q, Siegwart DJ. Theranostic dendrimer‐based lipid nanoparticles containing PEGylated BODIPY dyes for tumor imaging and systemic mRNA delivery in vivo. J Control Release. 2020;325:198‐205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182. Chinsky JM, Singh R, Ficicioglu C, et al. Diagnosis and treatment of tyrosinemia type I: a US and Canadian consensus group review and recommendations. Genet Med. 2017;19:1380‐1395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183. Cheng Q, Wei T, Jia Y, et al. Dendrimer‐based lipid nanoparticles deliver therapeutic FAH mRNA to normalize liver function and extend survival in a mouse model of hepatorenal tyrosinemia type I. Adv Mater. 2018;30:e1805308. [DOI] [PubMed] [Google Scholar]
- 184. Yang X, Du H, Liu J, et al. Advanced nanocarriers based on heparin and its derivatives for cancer management. Biomacromolecules. 2015;16:423‐436. [DOI] [PubMed] [Google Scholar]
- 185. Mahiny AJ, Dewerth A, Mays LE, et al. In vivo genome editing using nuclease‐encoding mRNA corrects SP‐B deficiency. Nat Biotechnol. 2015;33:584‐586. [DOI] [PubMed] [Google Scholar]
- 186. Gelzo M, Sica C, Elce A, et al. Reduced absorption and enhanced synthesis of cholesterol in patients with cystic fibrosis: a preliminary study of plasma sterols. Clin Chem Lab Med. 2016;54:1461‐1466. [DOI] [PubMed] [Google Scholar]
- 187. Singh VK, Schwarzenberg SJ. Pancreatic insufficiency in cystic fibrosis. J Cyst Fibros. 2017;16:S70‐S78. [DOI] [PubMed] [Google Scholar]
- 188. Kolonko AK, Efing J, Gonzalez‐Espinosa Y, et al. Capsaicin‐loaded chitosan nanocapsules for wtCFTR‐mRNA delivery to a cystic fibrosis cell line. Biomedicine. 2020;8:364. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189. Paques JP, van der Linden E, van Rijn CJ, et al. Preparation methods of alginate nanoparticles. Adv Colloid Interface Sci. 2014;209:163‐171. [DOI] [PubMed] [Google Scholar]
- 190. Plucinski A, Lyu Z, Schmidt B. Polysaccharide nanoparticles: from fabrication to applications. J Mater Chem B. 2021;9:7030‐7062. [DOI] [PubMed] [Google Scholar]
- 191. Yang Z, Yang Y, Xu Y, et al. Biomimetic nerve guidance conduit containing engineered exosomes of adipose‐derived stem cells promotes peripheral nerve regeneration. Stem Cell Res Ther. 2021;12:442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192. Tang Q, Cao B, Lei X, Sun B, Zhang Y, Cheng G. Dextran‐peptide hybrid for efficient gene delivery. Langmuir. 2014;30:5202‐5208. [DOI] [PubMed] [Google Scholar]
- 193. Machado V, Morais M, Medeiros R. Hyaluronic acid‐based nanomaterials applied to cancer: where are we now? Pharmaceutics. 2022;14:2092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194. Yan H, Hu Y, Akk A, et al. Induction of WNT16 via peptide‐mRNA nanoparticle‐based delivery maintains cartilage homeostasis. Pharmaceutics. 2020;12:73. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195. Yeom JH, Ryou SM, Won M, Park M, Bae J, Lee K. Inhibition of xenograft tumor growth by gold nanoparticle‐DNA oligonucleotide conjugates‐assisted delivery of BAX mRNA. PLoS One. 2013;8:e75369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196. Erasmus JH, Khandhar AP, O'Connor MA, et al. An alphavirus‐derived replicon RNA vaccine induces SARS‐CoV‐2 neutralizing antibody and T cell responses in mice and nonhuman primates. Sci Transl Med. 2020;12:eabc9396. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197. Duan L, Xu L, Xu X, et al. Exosome‐mediated delivery of gene vectors for gene therapy. Nanoscale. 2021;13:1387‐1397. [DOI] [PubMed] [Google Scholar]
- 198. Bu T, Li Z, Hou Y, et al. Exosome‐mediated delivery of inflammation‐responsive Il‐10 mRNA for controlled atherosclerosis treatment. Theranostics. 2021;11:9988‐10000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 199. Li Z, Zhao P, Zhang Y, et al. Exosome‐based Ldlr gene therapy for familial hypercholesterolemia in a mouse model. Theranostics. 2021;11:2953‐2965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200. Yang J, Wu S, Hou L, et al. Therapeutic effects of simultaneous delivery of nerve growth factor mRNA and protein via exosomes on cerebral ischemia. Mol Ther Nucleic Acids. 2020;21:512‐522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 201. Mizrak A, Bolukbasi MF, Ozdener GB, et al. Genetically engineered microvesicles carrying suicide mRNA/protein inhibit schwannoma tumor growth. Mol Ther. 2013;21:101‐108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202. Yang Z, Shi J, Xie J, et al. Large‐scale generation of functional mRNA‐encapsulating exosomes via cellular nanoporation. Nat Biomed Eng. 2020;4:69‐83. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203. Kojima R, Bojar D, Rizzi G, et al. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson's disease treatment. Nat Commun. 2018;9:1305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204. Wang JH, Forterre AV, Zhao J, et al. Anti‐HER2 scFv‐directed extracellular vesicle‐mediated mRNA‐based gene delivery inhibits growth of HER2‐positive human breast tumor xenografts by prodrug activation. Mol Cancer Ther. 2018;17:1133‐1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205. Tsai SJ, Atai NA, Cacciottolo M, et al. Exosome‐mediated mRNA delivery in vivo is safe and can be used to induce SARS‐CoV‐2 immunity. J Biol Chem. 2021;297:101266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206. Usman WM, Pham TC, Kwok YY, et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat Commun. 2018;9:2359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207. Trivedi M, Talekar M, Shah P, Ouyang Q, Amiji M. Modification of tumor cell exosome content by transfection with wt‐p53 and microRNA‐125b expressing plasmid DNA and its effect on macrophage polarization. Oncogenesis. 2016;5:e250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208. Li Z, Zhou X, Wei M, et al. In vitro and in vivo RNA inhibition by CD9‐HuR functionalized exosomes encapsulated with miRNA or CRISPR/dCas9. Nano Lett. 2019;19:19‐28. [DOI] [PubMed] [Google Scholar]
- 209. Li J, Huang Y, Song J, et al. Cartilage regeneration using arthroscopic flushing fluid‐derived mesenchymal stem cells encapsulated in a one‐step rapid cross‐linked hydrogel. Acta Biomater. 2018;79:202‐215. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210. Popowski KD, Moatti A, Scull G, et al. Inhalable dry powder mRNA vaccines based on extracellular vesicles. Matter. 2022;5:2960‐2974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211. Escudier B, Dorval T, Chaput N, et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived‐exosomes: results of thefirst phase I clinical trial. J Transl Med. 2005;3:10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 212. Duan L, Ouyang K, Wang J, et al. Exosomes as targeted delivery platform of CRISPR/Cas9 for therapeutic genome editing. Chembiochem. 2021;22:3360‐3368. [DOI] [PubMed] [Google Scholar]
- 213. Liang Y, Iqbal Z, Wang J, et al. Cell‐derived extracellular vesicles for CRISPR/Cas9 delivery: engineering strategies for cargo packaging and loading. Biomater Sci. 2022;10:4095‐4106. [DOI] [PubMed] [Google Scholar]
- 214. Moore T, Zhang Y, Fenley MO, Li H. Molecular basis of box C/D RNA‐protein interactions; cocrystal structure of archaeal L7Ae and a box C/D RNA. Structure. 2004;12:807‐818. [DOI] [PubMed] [Google Scholar]
- 215. Jang HL, Jin K, Lee J, et al. Revisiting whitlockite, the second most abundant biomineral in bone: nanocrystal synthesis in physiologically relevant conditions and biocompatibility evaluation. ACS Nano. 2014;8:634‐641. [DOI] [PubMed] [Google Scholar]
- 216. Liang Y, Duan L, Lu J, Xia J. Engineering exosomes for targeted drug delivery. Theranostics. 2021;11:3183‐3195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217. Liang Y, Xu X, Li X, et al. Chondrocyte‐targeted microRNA delivery by engineered exosomes toward a cell‐free osteoarthritis therapy. ACS Appl Mater Interfaces. 2020;12:36938‐36947. [DOI] [PubMed] [Google Scholar]
- 218. Liang Y, Xu X, Xu L, et al. Chondrocyte‐specific genomic editing enabled by hybrid exosomes for osteoarthritis treatment. Theranostics. 2022;12:4866‐4878. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219. Xu X, Liang Y, Li X, et al. Exosome‐mediated delivery of kartogenin for chondrogenesis of synovial fluid‐derived mesenchymal stem cells and cartilage regeneration. Biomaterials. 2021;269:120539. [DOI] [PubMed] [Google Scholar]
- 220. Fajrial AK, He QQ, Wirusanti NI, Slansky JE, Ding X. A review of emerging physical transfection methods for CRISPR/Cas9‐mediated gene editing. Theranostics. 2020;10:5532‐5549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 221. Kim HJ, Seo SK, Park HY. Physical and chemical advances of synthetic delivery vehicles to enhance mRNA vaccine efficacy. J Control Release. 2022;345:405‐416. [DOI] [PubMed] [Google Scholar]
- 222. Guan S, Rosenecker J. Nanotechnologies in delivery of mRNA therapeutics using nonviral vector‐based delivery systems. Gene Ther. 2017;24:133‐143. [DOI] [PubMed] [Google Scholar]
- 223. Li DF, Yang MF, Xu HM, et al. Nanoparticles for oral delivery: targeted therapy for inflammatory bowel disease. J Mater Chem B. 2022;10:5853‐5872. [DOI] [PubMed] [Google Scholar]
- 224. Duan L, Ouyang K, Xu X, et al. Nanoparticle delivery of CRISPR/Cas9 for genome editing. Front Genet. 2021;12:673286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225. Perche F, Clemençon R, Schulze K, Ebensen T, Guzmán CA, Pichon C. Neutral lipopolyplexes for in vivo delivery of conventional and replicative RNA vaccine. Mol Ther Nucleic Acids. 2019;17:767‐775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226. Rezaee M, Oskuee RK, Nassirli H, Malaekeh‐Nikouei B. Progress in the development of lipopolyplexes as efficient non‐viral gene delivery systems. J Control Release. 2016;236:1‐14. [DOI] [PubMed] [Google Scholar]
- 227. Luk BT, Zhang L. Cell membrane‐camouflaged nanoparticles for drug delivery. J Control Release. 2015;220:600‐607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 228. Liang Y, Iqbal Z, Lu J, et al. Cell‐derived nanovesicle‐mediated drug delivery to the brain: principles and strategies for vesicle engineering. Mol Ther. 2022;31:00620‐7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229. O'Brien K, Breyne K, Ughetto S, et al. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat Rev Mol Cell Biol. 2020;21:585‐606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230. Qiu M, Glass Z, Chen J, et al. Lipid nanoparticle‐mediated codelivery of Cas9 mRNA and single‐guide RNA achieves liver‐specific in vivo genome editing of Angptl3. Proc Natl Acad Sci U S A. 2021;118:e2020401118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 231. Cheng Q, Wei T, Farbiak L, Johnson LT, Dilliard SA, Siegwart DJ. Selective organ targeting (SORT) nanoparticles for tissue‐specific mRNA delivery and CRISPR‐Cas gene editing. Nat Nanotechnol. 2020;15:313‐320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232. Ma F, Yang L, Sun Z, et al. Neurotransmitter‐derived lipidoids (NT‐lipidoids) for enhanced brain delivery through intravenous injection. Sci Adv. 2020;6:eabb4429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233. Duan L, Li X, Xu X, et al. Large‐scale preparation of synovial fluid mesenchymal stem cell‐derived exosomes by 3D bioreactor culture. J Vis Exp. 2022;185:e62221. [DOI] [PubMed] [Google Scholar]
- 234. Shinge SAU, Xiao Y, Xia J, Liang Y, Duan L. New insights of engineering plant exosome‐like nanovesicles as a nanoplatform for therapeutics and drug delivery. EVCNA. 2022;3:150‐162. [Google Scholar]
- 235. Xu L, Liang Y, Xu X, et al. Blood cell‐derived extracellular vesicles: diagnostic biomarkers and smart delivery systems. Bioengineered. 2021;12:7929‐7940. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
There are no supplementary data available for this article.