

ABSTRACT
Enterococcus faecium is a difficult-to-treat pathogen with emerging resistance to most clinically available antibiotics. Daptomycin (DAP) is the standard of care, but even high DAP doses (12 mg/kg body weight/day) failed to eradicate some vancomycin-resistant strains. Combination DAP-ceftaroline (CPT) may increase β-lactam affinity for target penicillin binding proteins (PBP); however, in a simulated endocardial vegetation (SEV) pharmacokinetic/pharmacodynamic (PK/PD) model, DAP-CPT did not achieve therapeutic efficacy against a DAP-nonsusceptible (DNS) vancomycin-resistant E. faecium (VRE) isolate. Phage-antibiotic combinations (PAC) have been proposed for resistant high-inoculum infections. We aimed to identify PAC with maximum bactericidal activity and prevention/reversal of phage and antibiotic resistance in an SEV PK/PD model against DNS isolate R497. Phage-antibiotic synergy (PAS) was evaluated with modified checkerboard MIC and 24-h time-kill analyses (TKA). Human-simulated antibiotic doses of DAP and CPT with phages NV-497 and NV-503-01 were then evaluated in 96-h SEV PK/PD models against R497. Synergistic and bactericidal activity was identified with the PAC of DAP-CPT combined with phage cocktail NV-497–NV-503-01, demonstrating a significant reduction in viability down to 3-log10 CFU/g (–Δ, 5.77-log10 CFU/g; P < 0.001). This combination also demonstrated isolate resensitization to DAP. Evaluation of phage resistance post-SEV demonstrated prevention of phage resistance for PACs containing DAP-CPT. Our results provide novel data highlighting bactericidal and synergistic activity of PAC against a DNS E. faecium isolate in a high-inoculum ex vivo SEV PK/PD model with subsequent DAP resensitization and prevention of phage resistance.
IMPORTANCE Our study supports the additional benefit of standard-of-care antibiotics combined with a phage cocktail compared to antibiotic alone against a daptomycin-nonsusceptible (DNS) E. faecium isolate in a high-inoculum simulated endocardial vegetation ex vivo PK/PD model. E. faecium is a leading cause of hospital-acquired infections and is associated with significant morbidity and mortality. Daptomycin is considered the first-line therapy for vancomycin-resistant E. faecium (VRE), but the highest published doses have failed to eradicate some VRE isolates. The addition of a β-lactam to daptomycin may result in synergistic activity, but previous in vitro data demonstrate that daptomycin plus ceftaroline failed to eradicate a VRE isolate. Phage therapy as an adjunct to antibiotic therapy has been proposed as a salvage therapy for high-inoculum infections; however, pragmatic clinical comparison trials for endocarditis are lacking and difficult to design, reinforcing the timeliness of such analysis.
KEYWORDS: bacteriophage, Enterococcus, antibiotic resistance, infective endocarditis
INTRODUCTION
Vancomycin-resistant enterococci (VRE) are the leading cause of hospital-acquired infections in immunocompromised patients and those with high comorbidity burden (1–3). VRE bacteremia is associated with mortality rates as high as 20%, which is strongly influenced by vancomycin resistance, the presence of infective endocarditis (IE), and microbiological failure (3–6).
Moreover, the Centers for Disease Control and Prevention (CDC) and World Health Organization consider VRE to be a high-priority bacterium for which new therapies are urgently needed (7).
Daptomycin (DAP) is the agreed upon first-line therapy to treat resistant Enterococcus faecium infections, due to a lack of other clinically effective agents. However, shortly following FDA approval, treatment failures were reported for infections caused by daptomycin-nonsusceptible (DNS) enterococci, often accompanied by bacteremia (8, 9). Much of it is attributed to substitutions in the LiaFSR system involved in the stress response system, although a recent study suggests other stress response pathways may also play a role (10–12). Due to the increasing incidence of DNS clinical isolates, emergence of resistance in enterococci during DAP treatment, and DAP-related treatment failure of resistant E. faecium infections, the Clinical and Laboratory Standards Institute (CLSI) Subcommittee on Antimicrobial Susceptibility Testing updated breakpoints for DAP and enterococci (13–15). While in vitro data also suggest that exposures achieved with doses of 10 to 12 mg/kg/day are needed to prevent these types of resistance emergence, including in strains with LiaFSR substitutions but wild-type MICs, uncertainties remain regarding DAP use for the treatment of VRE blood infections and IE, particularly since even the highest published doses (12 mg/kg body weight/day) have demonstrated the inability to eradicate some wild-type isolates of E. faecium (10, 16). An E. faecium murine thigh infection model with human simulated exposures of DAP 10 mg/kg/day achieved a 1 log kill rate for E. faecium strains with MICs up to 8 mg/L (17). These data call into question the current DAP susceptibility breakpoint and whether it limits therapeutic optimization for enterococcal bacteremia and IE.
The addition of ceftaroline (CPT) to DAP to treat resistant enterococci infections may serve as a therapeutic alternative for VR-E. faecium secondary to the enhanced binding affinity of CPT to penicillin binding protein 5 (PBP5) that assists DAP in overcoming cell membrane repulsion, a common mechanism of resistance in DAP-nonsusceptible E. faecium strains (18–21). In previous 4-day in vitro model experiments, CPT was the most effective β-lactam in reducing the DAP MIC against E. faecium (18). Although this combination is promising, previous evaluations of DAP in combination with CPT using the simulated endocardial vegetation (SEV) ex vivo pharmacokinetic/pharmacodynamic (PK/PD) model, validated with a rat model of IE, demonstrated that the combination of DAP plus CPT did not achieve therapeutic efficacy and failed to prevent elevation of DAP and CPT MICs from baseline in DNS E. faecium clinical blood isolate R497 (22).
Phage therapy, in which bacterial viruses are used to treat bacterial infections, has been proposed as a salvage therapy, especially in the context of high inoculum-infections and those caused by multidrug-resistant pathogens (23). The available evidence suggests that phage therapy may be an effective complementary strategy to antibiotics for the treatment of VR-E. faecium infections, including IE, in phage-antibiotic combinations (PACs) (23–25). As comparative clinical trials are difficult to design in the field of IE, this study aimed to evaluate phage-antibiotic synergy (PAS) with PACs containing DAP and CPT in vitro and efficacy in 96-h ex vivo simulated endocardial vegetation (SEV) PK/PD models with an assessment of antibiotic and phage susceptibility compared to baseline.
RESULTS
Modified checkerboard MIC.
To evaluate PAS and identify the optimal phage multiplicity of infection (MOI) in PAC prior to 24-h time-kill analyses (TKA) and ex vivo SEV PK/PD models, bacterial growth was assessed against serial dilutions of DAP and/or CPT and phage titer using a modified checkerboard (CB) MIC microtiter plate assay system in which the optical density of each well was measured following 24 h of incubation (Fig. 1). Well absorbance was read using spectrophotometric analysis and converted to a heat map of percent growth compared to the growth control. Against DNS E. faecium R497 in a modified CB MIC assay, DAP-CPT combined with single-phage NV-497 and two-phage cocktail NV-497–NV-503-01 (each phage at an MOI of 0.1) demonstrated synergy with a fractional inhibitory concentration (FIC) index of 0.5 (Fig. 1C1 and C3, yellow outline), while the addition of single-phage NV-503-01 (MOI, 0.1) to DAP-CPT was additive with an FIC index of 1 (Fig. 1C2, orange outline).
FIG 1.

(A to C) Modified checkerboard MIC assay of (A) daptomycin, (B) ceftaroline, and (C) daptomycin plus ceftaroline combined with single phages NV-497 (A1-C1) and NV-503-01 (A2-C2) or two-phage cocktail NV-497 plus NV-503-01 (A3-C3) against daptomycin-nonsusceptible E. faecium. Phage was added to each non-GC and non-MC well in panels C1 to C3 at a phage MOI of 0.1. Squares within the yellow outline (C1 and C3) demonstrated synergy with a fractional inhibitory concentration (FIC) of 0.5. Squares within the orange outline demonstrated additivity with an FIC of 1. Percent growth was compared to the control via spectrophotometric analysis. GC, growth control; MC, medium control; MOI, multiplicity of infection; DAP, daptomycin; CPT, ceftaroline.
FIG 2.
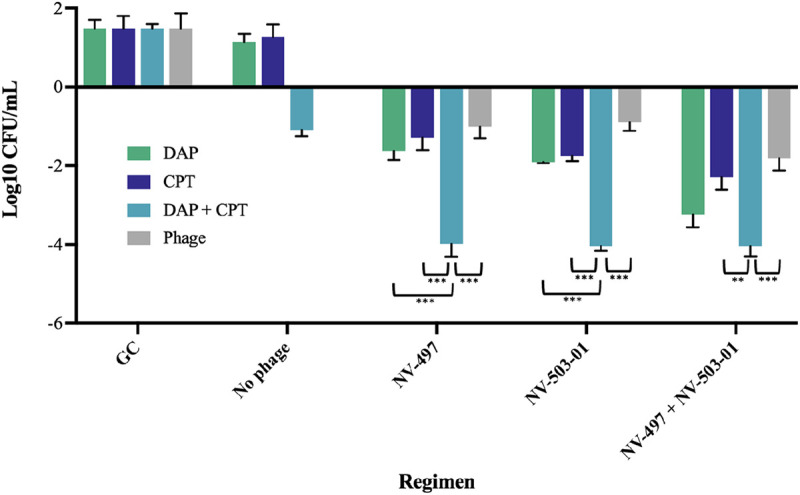
Bacterial quantification in 24-h time-kill experiments of daptomycin (0.5× MIC), ceftaroline (peak total concentration, 21.3), or daptomycin plus ceftaroline combined with phages NV-497 and NV-503-01 (each at a theoretical multiplicity of infection [MOI] of 0.1) against daptomycin nonsusceptible E. faecium R497. The error bars indicate standard deviation. P values were determined using a one-way ANOVA and Tukey’s post hoc test. **, P < 0.05; ***, P < 0.001. GC, growth control; DAP, daptomycin; CPT, ceftaroline.
Time-kill analyses.
DNS E. faecium R497 was then evaluated in a 24-h TKA against DAP (0.5× MIC), CPT (peak total concentration, 21.3 μg/mL), and phages NV-497 and NV-503-01 (each at an MOI of 0.1 based on modified CB MIC results). Combination DAP-CPT with single phages NV-497 and NV-503-01 or a cocktail of the two demonstrated bactericidal activity (–Δ, 4 log10 CFU/mL; P < 0.001) with killing to the detection limit of 2 log10 CFU/mL (Fig. 2). The combination of DAP-CPT with either the single phage NV-497 or NV-503-01 also demonstrated synergistic activity compared to the next best regimen of DAP plus either phage (–Δ, ≥2 log10 CFU/mL; P < 0.001). DAP with phage cocktail NV-497 plus NV-503-01 was also bactericidal with a 3.34 log10 CFU/mL reduction compared to the growth control.
SEV PK/PD ex vivo model.
In total, 12 separate 96-h ex vivo SEV PK/PD models were completed in duplicate with DNS E. faecium isolate R497 (Fig. 3). Humanized doses of DAP 10 mg/kg every 24 h and CPT 600 mg every 8 h were administered alone and in combination with single phages NV-497 and NV-503-01 or as a phage cocktail (each at an MOI of 1) (Table 1). Selection of the phage MOI for use in the high-inoculum SEV models was based on CB MIC results demonstrating enhanced killing for all tested PACs at a minimum MOI of 1 (Fig. 1). In the ex vivo SEV models against R497, the combination of DAP-CPT with the 2-phage cocktail NV-497–NV-503-01 demonstrated bactericidal and synergistic activity with a significant reduction in viability from the starting inoculum of 8.77 log10 CFU/g down to 3 log10 CFU/g (–Δ, 5.77 log10 CFU/g) compared to the next most effective regimen consisting of DAP-CPT-NV-503-01 (P < 0.001) (Fig. 3). Regimens with DAP-CPT and the single phage NV-497 or NV-503-01 demonstrated initial bactericidal activity with bacterial burdens decreasing to 4.22 and 4.50 log10 CFU/g (–Δ, 4.93 and 4.65 log10 CFU/g), respectively, at 32 h, followed by regrowth from 48 to 96 h to a final inoculum of 7.40 and 7 log10 CFU/g, respectively.
FIG 3.

Efficacy of bacteriophage-antibiotic combinations in a simulated endocardial vegetation (SEV) pharmacokinetic/pharmacodynamic (PK/PD) model (bacterial quantification) over a 96-h period. The error bars indicate standard deviation. GC, growth control; DAP, daptomycin; CPT, ceftaroline.
Table 1.
Pharmacokinetic parameters for antibiotics used in ex vivo SEV modelsa
| Antibiotic (dose) | Target |
Achieved |
|||
|---|---|---|---|---|---|
| Cmax (μg/mL) | t1/2 (h) | Cmax (μg/mL) | t1/2 (h) | AUC0–24 (μg*h/mL) | |
| DAP (10 mg/kg) | 141.1 | 8 | 142.1 ± 0.7 | 8.12 ± 0.08 | 1,764.1 ± 22.4 |
| CPT (600 mg) | 21.3 | 2.66 | 22.6 ± 0.2 | 2.7 ± 0.2 | 81.3 ± 1.9 |
SEV, simulated endocardial vegetation; DAP, daptomycin; CPT, ceftaroline; Cmax, maximum concentration; t1/2, half-life; AUC0–24, area under the concentration-time curve over the 24-h dosing interval.
Humanized DAP 10 mg/kg/day doses used in the ex vivo models were a total maximum drug concentration (Cmax) of 142.1 ± 0.7 μg/mL (target total Cmax, 141.1 μg/mL) and half-life (t1/2) of 8.12 ± 0.08 h (target t1/2, 8 h) and an area under the curve (AUC) from 0 to 24 h of 1,764.1 ± 22.4 μg · h/mL (Table 1). The PK parameters for CPT in the model were a total drug Cmax of 22.6 ± 0.2 μg/mL (target total Cmax, 21.3 μg/mL), a t1/2 of 2.7 ± 0.2 h (target t1/2, 2.66 h), and an AUC from 0 to 24 h (AUC0–24) of 81.3 ± 1.9 μg · h/mL.
Post-SEV model susceptibility and sensitivity tests.
(i) Antibiotic MIC. To further evaluate the findings of the SEV model and examine whether antibiotic resensitization was present, we conducted MIC testing for DNS E. faecium isolate R497 in each 96-h SEV sample for DAP, CPT, and other standard-of-care antibiotics in triplicate using broth microdilution following CLSI guidelines (26). If a 96-h sample demonstrated an MIC change of ≥2 dilutions from baseline, then samples were assessed for resistance in a backward stepwise manner from 72 h to earlier time points until a ≤1 dilution change in MIC was identified for the sample. At 96 h of the SEV model, the DAP MIC demonstrated resensitization for R497 (16 μg/mL to 4 μg/mL) for the PAC DAP-CPT combined with phage cocktail NV-497 and NV-503-01 (Table 2). Upon further analysis, DAP resensitization to an MIC of 4 μg/mL was identified in 72-h and 48-h samples; however, the DAP-resensitization was not stable upon MIC retesting 6 months postmodel; thus, additional analyses using whole-genome sequencing (WGS) were not feasible. All other 96-h SEV samples demonstrated stable antibiotic MICs relative to the premodel baseline MICs for DAP, CPT, and other antibiotics tested, including ampicillin, linezolid, minocycline, and vancomycin.
Table 2.
Antibiotic MIC for daptomycin-nonsusceptible E. faecium isolate R497 in 96-h SEV samplesa
| SEV model regimen | Antibiotic MIC (μg/mL)a |
|||||
|---|---|---|---|---|---|---|
| Daptomycin | Ceftaroline | Ampicillin | Linezolid | Minocycline | Vancomycin | |
| Baseline | 16 | 128 | 256 | 4 | 16 | 2 |
| Growth control | ||||||
| DAP | ||||||
| No phage | 16 | 128 | 256 | 4 | 16 | 2 |
| NV-497 | ||||||
| NV-503-01 | ||||||
| NV-497–NV-503-01 | ||||||
| CPT | ||||||
| No phage | 16 | 128 | 256 | 4 | 16 | 2 |
| NV-497 | ||||||
| NV-503-01 | ||||||
| NV-497–NV-503-01 | ||||||
| DAP-CPT | ||||||
| No phage | 16 | 128 | 256 | 4 | 16 | 2 |
| NV-497 | ||||||
| NV-503-01 | ||||||
| NV-497–NV-503-01 | 4 | |||||
SEV, simulated endocardial vegetation; DAP, daptomycin; CPT, ceftaroline.
(ii) Phage sensitivity. To evaluate whether there was a change in phage sensitivity in 96-h SEV samples compared to baseline, the frequency of apparent resistance (FoR) (Table 3), followed by confirmatory testing of single FoR colonies in triplicate (Fig. 4), was evaluated as previously described for phages used in the SEV model (e.g., NV-497 and NV-503-01) (27). Phage sensitivity in 96 h SEV samples was also evaluated for seven other E. faecium phages previously evaluated against E. faecium isolate R497 using spot testing, as previously described (Fig. 5) (28, 29). Upon evaluation of the two phages used in the SEV models (NV-497 and NV-503-01), phage resistance was identified in 96-h SEV model samples for both phages with the following regimens: (i) DAP or CPT monotherapy plus the phage cocktail NV-497–NV-503-01 and (ii) NV-497–NV-503-01 phage cocktail without antibiotic (Fig. 4). Neither phage NV-497 nor NV-503-01 demonstrated decreased sensitivity for R497 in 96-h SEV samples for regimens that included combination DAP-CPT. Spot test phage sensitivity results for other E. faecium phages tested did not demonstrate changes in phage sensitivity compared to baseline (Fig. 5).
Table 3.
Frequency of apparent resistance in E. faecium R497 sensitive to each phagea
| Treatment | Timepoint (h) | Colony count | Mean CFU/g in sample | Apparent FoRb | LoDc (1/CFU on plate) |
|---|---|---|---|---|---|
| No antibiotic | |||||
| NV-497–NV-503-01 | 0 | 0 | 3.35 × 108 | <LoD | 2.99 × 108 |
| 96 | Est. 1,000 | 4.78 × 108 | 2.09 × 10−6 | 2.09 × 108 | |
| DAP | |||||
| NV-497 | 0 | 0 | 5.15 × 108 | <LoD | 1.94 × 108 |
| 96 | 579 | 7.46 × 108 | 7.76 × 10−7 | 1.34 × 108 | |
| NV-503-01 | 0 | 0 | 3.77 × 108 | <LoD | 2.65 × 108 |
| 96 | 665 | 6.86 × 108 | 6.69 × 10−7 | 1.46 × 108 | |
| NV-497–NV-503-01 | 0 | 0 | 5.14 × 108 | <LoD | 1.95 × 108 |
| 96 | 236 | 4.48 × 108 | 5.27 × 10−7 | 2.23 × 108 | |
| CPT | |||||
| NV-497 | 0 | 0 | 2.65 × 108 | <LoD | 3.77 × 108 |
| 96 | 788 | 6.26 × 108 | 1.26 × 10−6 | 1.59 × 108 | |
| NV-503-01 | 0 | 0 | 2.13 × 108 | <LoD | 4.69 × 108 |
| 96 | Est. 1,000 | 5.07 × 108 | 1.97 × 10−6 | 1.97 × 108 | |
| NV-497–NV-503-01 | 0 | 0 | 6.25 × 108 | <LoD | 1.61 × 108 |
| 96 | Est. 1,000 | 2.89 × 108 | 3.46 × 10−6 | 3.46 × 108 | |
| DAP-CPT | |||||
| NV-497 | 0 | 0 | 7.33 × 108 | <LoD | 1.36 × 108 |
| 96 | 272 | 2.93 × 108 | 9.28 × 10−7 | 3.41 × 108 | |
| NV-503-01 | 0 | 0 | 3.27 × 108 | <LoD | 3.06 × 108 |
| 96 | 304 | 5.29 × 108 | 5.75 × 10−7 | 1.89 × 108 | |
| NV-497–NV-503-01 | 0 | 0 | 7.41 × 108 | <LoD | 1.34 × 108 |
| 96 | 39 | 5.07 × 108 | 7.69 × 10−8 | 1.97 × 108 |
Est., estimated counts; DAP, daptomycin; CPT, ceftaroline.
Frequency of resistance ([colonies that arise on the resistance plate]/[CFU per milliliter used to create the plate]).
LoD, limit of detection. If <LoD is indicated, then the apparent frequency of resistance equals 0.
FIG 4.

Final resistance in E. faecium R497 sensitive to each phage. Numbers are the PFU per milliliter for each combination of phage and bacterial isolate. Green, orange, and red cells indicate apparent, faint, and no bacterial lawn clearing, respectively. The lack of observed defined plaques is indicated by a titer of zero. S, sensitive; R, resistant; DAP, daptomycin; CPT, ceftaroline.
FIG 5.

Postmodel sensitivity of bacteriophage not used in the SEV ex vivo models for daptomycin-nonsusceptible E. faecium isolate R497. Phage sensitivity was classified as high, medium, or low based on PFU counts as follows: >107 PFU/mL was defined as high (green), ≥103 to ≤107 PFU/mL was defined as median (orange), and <103 was defined as low (yellow). Phage was classified as nonsusceptible (NS) if no PFU were identified (red). SEV, simulated endocardial vegetation.
DISCUSSION
To the authors’ knowledge, this is the first study to evaluate the efficacy of adding a phage cocktail to standard-of-care antibiotics in a high-inoculum PK/PD SEV ex vivo model against a DNS E. faecium isolate with the aim to assess fundamental phage-antibiotic synergy components, including bactericidal activity, antibiotic resensitization, and prevention of phage and/or antibiotic treatment-emergent resistance. Specifically, DAP and CPT were evaluated alone and in various combinations with two E. faecium phages, NV-497 and NV-503-01, against a DAP-nonsusceptible E. faecium clinical blood isolate, R497, harboring common LiaRS substitutions.
These data expand on the authors’ previous work evaluating the E. faecium isolate R497 against DAP and β-lactam combinations in both 24-h in vitro TKA and 14-day ex vivo SEV models (30–32). In the 14-day SEV models, DAP plus CPT showed no activity against R497 with selection of resistant isolates with corresponding elevated MICs as high as >64 μg/mL. However, in 24-h TKA in both planktonic and biofilm states, it was demonstrated that DAP and CPT in combination with a single phage demonstrated bactericidal activity and were synergistic compared to DAP plus CPT at 24 h.
The results of the current study provide novel data highlighting bactericidal and synergistic activity of DAP-CPT combined with a two-phage cocktail (NV-497–NV-503-01) against DNS E. faecium isolate R497 in a high-inoculum SEV PK/PD model with subsequent DAP resensitization and prevention of phage resistance. Phage-antibiotic synergy has been documented multiple times previously, including in the current study with addition of a two-phage cocktail to DAP and CPT; however, the mechanism behind it has not been fully elucidated. It has been hypothesized that phage-antibiotic synergy may be due, in part, to phage-induced selective pressure on bacterial strains, with a fitness cost of regaining antibiotic susceptibility (33). Regarding the resensitization of R497 to DAP following 48 to 96 h of isolate exposure to the phage-antibiotic combination DAP-CPT with phage cocktail NV-497–NV-503-01, as well as isolate reversion back to DAP nonsusceptibility, this phenomenon has not been previously reported to the authors’ knowledge. However, the reversion of a DAP-resistant S. aureus isolate to DAP susceptible has been reported to occur clinically and is hypothesized to be related to the downregulation of the two-compartment systems that control cell wall biosynthesis-related genes, including the downregulation of VraSR, yycG, and GraS (34). It is tempting to speculate that removal of this phage-induced selective pressure restored the fitness advantage of the strain and reconstituted the DAP nonsusceptibility (NS) after extended storage. Further studies are warranted to evaluate whether similar system downregulation occurs in E. faecium in the presence of phage-antibiotic combinations.
Limitations to this study include those reflective of its study design. Infective endocarditis is a disease state that is treated for an extended period of approximately 6 weeks. Thus, we are unable to exclude the possibility that a longer duration model may result in variability in regimen differences compared to that seen in the present study of 96-h ex vivo models. Additional limitations include the evaluation of a single E. faecium strain, lack of dose escalation/de-escalation experiments, and the lack of whole-genome sequencing analysis to identify genetic mechanisms of antibiotic and phage resistance.
Due to the increasing incidence of resistant enterococcal infections, including infective endocarditis, and the resultant suboptimal antibiotic target attainment and poor clinical outcomes, future studies are warranted to evaluate the effectiveness of DAP combined with other standard-of-care β-lactams and phage therapy. In summary, the addition of a two-phage cocktail to DAP and CPT was bactericidal and highly synergistic against DNS E. faecium, demonstrated DAP resensitization compared to baseline, and protected against treatment-emergent phage resistance.
MATERIALS AND METHODS
Bacterial strain.
E. faecium R497, a DAP-NS (DAP MIC, 16 mg/L) human blood isolate with LiaSR mutations, was evaluated in this study (35–38).
Phage isolation and sensitivity assessment.
Bacteriophages (phages) specific to E. faecium R497, NV-497, and NV-503-01, were isolated from wastewater in Maryland. Phage isolation, purification, concentration, and replication were carried out by standard methods as described elsewhere (39). Phage sensitivity was previously evaluated with the small-drop overlay or “spot testing” on brain heart infusion (BHI) plates with results reported previously (28, 40). Two phages, NV-497 and NV-503-01, were selected for use in the current study based on their demonstrated high phage susceptibility (>107 PFU) for DNS E. faecium clinical blood isolates.
Antimicrobial agents and media.
Daptomycin was obtained commercially from Xellia Pharmaceuticals (Buffalo Grove, IL, USA), and CPT analytical powder was obtained from AbbVie, Inc. (North Chicago, IL, USA). Brain heart infusion (BHI) broth (Difco, Detroit, MI, USA) supplemented with calcium and magnesium (50 and 12.5 μg/mL, respectively) was used for modified CB MIC analysis and 24 h TKA. Mueller-Hinton II broth (MHB) in ex vivo SEV models was supplemented to a final calcium concentration of 75 μg/mL to account for the dependency of DAP antimicrobial activity on calcium and calcium binding due to albumin (41). Colony counts were performed on BHI agar (Difco).
Antibiotic Susceptibility Testing.
MICs of study antimicrobials were determined in duplicate by manual broth microdilution (BMD) at approximately 106 CFU/mL according to the Clinical and Laboratory Standards Institute (CLSI) (26). Due to raised MIC values of CPT for this E. faecium strain, combination MIC values for DAP were determined by supplementing the broth with concentrations of CPT at its biological free peak concentration (fCmax); (21.3 μg/mL) to validate the ability of CPT to reduce the DAP MIC. The DAP MIC fold reduction from baseline was calculated by dividing DAP MICs by DAP MICs in the presence of CPT.
Modified checkerboard MIC assay.
Antibiotic and phage synergy against R497 was initially evaluated using modified CB MIC analyzing the activity of antibiotics and phage in combination compared to their individual activities, as previously described (28, 30). In brief, 100 μL of BHI was added to each well of a 96-well round-bottom microtiter plate. Next, 100 μL of DAP or CPT at 2-fold the MIC was added to column 1 and serially diluted to column 10, like MIC testing. For checkerboards analyzing synergy between two antibiotics, 100 μL of the other antibiotic was added to the first row in columns 1 to 10 and diluted 2-fold through row 7. For PAS screening, 10 μL of phage (MOI, 10) instead of antibiotic was added to the first row in columns 1 to 10 with phage dilutions performed 10-fold rather than 2-fold to achieve the desired range of MOI. Next, 100 μL of BHI inoculated with 5 × 105 starting inoculum of test organism was then added to each well of the 96-well plate except for the last column. The microtiter plate was then placed on the new plate and incubated at 37°C for 24 h and then read at an optical density of 570 nm (OD570). In triple-therapy checkerboards assessing the synergy of DAP plus CPT in the presence of constant phage, phage was added to each well, excluding controls, at a subinhibitory MOI determined in single phage-antibiotic CB. Synergy, additive activity, and antagonism were defined as a calculated fractional inhibitory concentration (FIC) index of ≤0.5, 1 to 4, and >4, respectively (42, 43).
Time-kill analyses in planktonic state.
Time-kill analyses of E. faecium R497 were performed in microwell plates against DAP, CPT, and/or phages NV-497 and NV-503-01 at a starting bacterial inoculum of 106 CFU/mL, as previously described (33). DAP was added at 0.5× the MIC value and CPT was added at the peak total concentration (CPT peak total concentration, 21.3 μg/mL). Phage at an MOI of 0.1 (phage:target organism) was based on modified CB results. In wells containing phage and antibiotic, antibiotic was added first, followed directly by phage. Synergy was defined as a ≥2-log10 CFU/mL kill rate compared to the most effective agent (or double-combination regimen), and bactericidal activity was defined as a ≥3-log10 CFU/mL reduction from baseline at 24 h.
Ex vivo PK/PD model.
Ex vivo SEV PK/PD models were conducted (44–46), and SEV clots were prepared as previously documented (27, 41). The models were incubated at 37°C for the duration of each 96-h model. MHB was cycled into and out of each model at a rate to simulate antibiotic t1/2, as appropriate, with humanized doses of DAP 10 mg/kg and CPT 600 mg injected into each model every 24 and 8 h, respectively. When included in the regimens, each phage was injected into the models every 24 h at an MOI of 1. Once SEV samples were removed in duplicate from each model at hour time points 0, 4, 8, 24, 32, 48, 72, and 96, each SEV clot was homogenized and then centrifuged twice, with supernatant replaced with normal saline following each centrifuge cycle to eliminate antibiotic and phage carryover.
Pharmacokinetic analysis.
Pharmacokinetic samples were obtained in duplicate through the injection port of each infection model at 0, 4, 8, 24, 32, 48, 72, and 96 h for verification of target antibiotic concentrations as previously described (46). All samples were stored at −80°C until ready for analysis. CPT concentrations were determined by bioassay using Bacillus subtilis ATCC 6633 (47). Blank 1/4-in. disks were spotted with 10 μL of standard concentrations or samples. Each standard was tested in duplicate by placing the disk on agar plates (antibiotic medium 11) inoculated with a 0.5 McFarland suspension of the test organism. This assay demonstrated an intraday coefficient of variance of less than 4.7% for high, medium, and low broth standards. The plates were incubated for 24 h at 37°C, at which time the zone sizes were measured using a ProtoCOL plate reader (Microbiology International, Frederick, MD, USA). DAP concentrations were determined using a validated high-performance liquid chromatography (HPLC) assay that conforms to the guidelines set forth by the College of American Pathologists and demonstrated an intraday coefficient of variance of less than 2% for high, medium, and low standards. The half-lives, peak concentrations, and area under the curve (AUC) (by trapezoidal method), or time spent above the MIC (T > MIC), were determined as appropriate for all antimicrobials utilizing PK Analyst software (version 1.10; MicroMath Scientific Software, Salt Lake City, UT, USA).
Post-SEV antibiotic susceptibility and phage sensitivity testing.
(i) Antibiotic susceptibility. Changes in antibiotic susceptibility compared to baseline were evaluated in each 96-h SEV sample using broth microdilution MICs following CLSI guidelines (26). If samples demonstrated MIC changes of ≥2 dilutions from baseline, then samples were assessed for resistance in a backward stepwise manner from 72 h to earlier time points until a ≤1 dilution change in MIC was identified for the sample. Antibiotics tested included those used in the SEV models (e.g., daptomycin and ceftaroline) and other antibiotics used clinically in the treatment of various infections caused by E. faecium, including ampicillin, linezolid, minocycline, and vancomycin.
(ii) Phage sensitivity. The development of phage resistance for phages used in the SEV models (e.g., NV-497 and NV-503-01) was tested in triplicate populations of the post-SEV R497 strains in a manner similar to that previously described (29, 48). Prior to SEV exposure, the frequency of spontaneous phage resistance in R497 populations was estimated to be approximately 10−8. First, surviving colonies were streak-purified on BHI plates followed by an evaluation of phage sensitivity using the double-drop method on BHI plates as previously described (29). The apparent frequency of resistance was calculated as the number of colonies on each test plate divided by the bacterial replicate number. The development of phage resistance for phages not used in the SEV models but previously tested with R497 (e.g., NV-503-02, NV-S447-01, NV-S447-02, and ATCC phages 113, 9181, 9183, and 9184) was also tested in triplicate populations of the post-SEV R497 strains using spot testing as previously described (28, 29).
Statistical analysis.
Statistical analyses were conducted with SPSS version 29 (IBM Corp., Armonk, NY, USA) software and Prism software version 9.2.0 (GraphPad, La Jolla, CA, USA). Bacterial burden reduction differences between phage and/or antibiotic regimens were compared by one-way analysis of variance (ANOVA) with Tukey’s post hoc test. A P value of ≤0.05 was considered significant. PK Analyst software version 1.10 (MicroMath, Salt Lake City, UT, USA) was used for the pharmacokinetic analysis of antibiotics used in the SEV models (e.g., daptomycin and ceftaroline) including t1/2, total Cmax concentrations, and AUC0–24.
ACKNOWLEDGMENTS
A.J.K.C., K.S., A.E.G., R.K., B.B., M.W., M.V.D., T.T.T., C.A.A., and M.J.R. were involved in the conceptualization, methodology, validation, formal analysis, investigation, writing–original draft preparation, and writing–review and editing of this manuscript. M.W. and M.V.D. contributed to isolation, amplification, and purification of the phages. All authors have read and agreed to the published version of the manuscript.
This research received no external funding. M.J.R. is supported by NIH grant R21 AI163726. C.A.A. is supported by NIH grants K24AI121296, R01AI134637, R01AI48342, and P01AI152999.
A.J.K.C., K.S., A.E.G., R.K., B.B., M.W., M.V.D., T.T.T., C.A.A. and M.J.R. have nothing to declare. M.J.R. has received grant support and has consulted or spoken on behalf of Allergan, Melinta, Merck, Paratek, Shionogi, Spero, and Tetraphase. C.A.A. has received grant support from Merck Pharmaceuticals, Entasis Pharmaceuticals and MeMed Diagnostics and is a cofounder and Entasis Therapeutics shareholder of Ancilia Biosciences.
The views expressed in this article reflect the results of research conducted by the authors and do not necessarily reflect the official policy or position of the Department of the Navy, Department of Defense, or the United States government.
This work was supported by work unit number A1417.
Contributor Information
Michael J. Rybak, Email: m.rybak@wayne.edu.
Brian Conlon, University of North Carolina at Chapel Hill.
REFERENCES
- 1.Weber S, Hogardt M, Reinheimer C, Wichelhaus TA, Kempf VAJ, Kessel J, Wolf S, Serve H, Steffen B, Scheich S. 2019. Bloodstream infections with vancomycin-resistant enterococci are associated with a decreased survival in patients with hematological diseases. Ann Hematol 98:763–773. doi: 10.1007/s00277-019-03607-z. [DOI] [PubMed] [Google Scholar]
- 2.Kramer TS, Remschmidt C, Werner S, Behnke M, Schwab F, Werner G, Gastmeier P, Leistner R. 2018. The importance of adjusting for enterococcus species when assessing the burden of vancomycin resistance: a cohort study including over 1000 cases of enterococcal bloodstream infections. Antimicrob Resist Infect Control 7:133. doi: 10.1186/s13756-018-0419-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Papanicolaou GA, Ustun C, Young J-AH, Chen M, Kim S, Woo Ahn K, Komanduri K, Lindemans C, Auletta JJ, Riches ML, CIBMTR Infection and Immune Reconstitution Working Committee . 2019. Bloodstream infection due to vancomycin-resistant Enterococcus is associated with increased mortality after hematopoietic cell transplantation for acute leukemia and myelodysplastic syndrome: a multicenter, retrospective cohort study. Clin Infect Dis 69:1771–1779. doi: 10.1093/cid/ciz031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Prematunge C, MacDougall C, Johnstone J, Adomako K, Lam F, Robertson J, Garber G. 2016. VRE and VSE bacteremia outcomes in the era of effective VRE therapy: a systematic review and meta-analysis. Infect Control Hosp Epidemiol 37:26–35. doi: 10.1017/ice.2015.228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Hefazi M, Damlaj M, Alkhateeb HB, Partain DK, Patel R, Razonable RR, Gastineau DA, Al-Kali A, Hashmi SK, Hogan WJ, Litzow MR, Patnaik MM. 2016. Vancomycin-resistant Enterococcus colonization and bloodstream infection: prevalence, risk factors, and the impact on early outcomes after allogeneic hematopoietic cell transplantation in patients with acute myeloid leukemia. Transpl Infect Dis 18:913–920. doi: 10.1111/tid.12612. [DOI] [PubMed] [Google Scholar]
- 6.Contreras GA, Munita JM, Simar S, Luterbach C, Dinh AQ, Rydell K, Sahasrabhojane PV, Rios R, Diaz L, Reyes K, Zervos M, Misikir HM, Sanchez-Petitto G, Liu C, Doi Y, Abbo LM, Shimose L, Seifert H, Gudiol C, Barberis F, Pedroza C, Aitken SL, Shelburne SA, van Duin D, Tran TT, Hanson BM, Arias CA. 2022. Contemporary clinical and molecular epidemiology of vancomycin-resistant enterococcal bacteremia: a prospective multicenter cohort study (VENOUS I). Open Forum Infect Dis 9:ofab616. doi: 10.1093/ofid/ofab616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Boucher HW, Talbot GH, Bradley JS, Edwards JE, Gilbert D, Rice LB, Scheld M, Spellberg B, Bartlett J. 2009. Bad bugs, no drugs: no ESKAPE! an update from the Infectious Diseases Society of America. Clin Infect Dis 48:1–12. doi: 10.1086/595011. [DOI] [PubMed] [Google Scholar]
- 8.Prater AG, Mehta HH, Beabout K, Supandy A, Miller WR, Tran TT, Arias CA, Shamoo Y. 2021. Daptomycin resistance in Enterococcus faecium can be delayed by disruption of the LiaFSR stress response pathway. Antimicrob Agents Chemother 65:e01317-20. doi: 10.1128/AAC.01317-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Humphries RM. 2019. The new, new daptomycin breakpoint for Enterococcus spp. J Clin Microbiol 65:e00600-19. doi: 10.1128/JCM.00600-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Turnidge J, Kahlmeter G, Cantón R, MacGowan A, Giske CG, European Committee on Antimicrobial Susceptibility Testing . 2020. Daptomycin in the treatment of enterococcal bloodstream infections and endocarditis: a EUCAST position paper. Clin Microbiol Infect 26:1039–1043. doi: 10.1016/j.cmi.2020.04.027. [DOI] [PubMed] [Google Scholar]
- 11.Munita JM, Arias CA. 2016. Mechanisms of antibiotic resistance. Microbiol Spectr 4:15. doi: 10.1128/microbiolspec.VMBF-0016-2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Miller WR, Tran TT, Diaz L, Rios R, Khan A, Reyes J, Prater AG, Panesso D, Shamoo Y, Arias CA. 2019. LiaR-independent pathways to daptomycin resistance in Enterococcus faecalis reveal a multilayer defense against cell envelope antibiotics. Mol Microbiol 111:811–824. doi: 10.1111/mmi.14193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Munita JM, Mishra NN, Alvarez D, Tran TT, Diaz L, Panesso D, Reyes J, Murray BE, Adachi JA, Bayer AS, Arias CA. 2019. Failure of high-dose daptomycin for bacteremia caused by daptomycin-susceptible Enterococcus faecium harboring LiaSR substitutions. Clin Infect Dis 59:1277–1280. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shukla BS, Shelburne S, Reyes K, Kamboj M, Lewis JD, Rincon SL, Reyes J, Carvajal LP, Panesso D, Sifri CD, Zervos MJ, Pamer EG, Tran TT, Adachi J, Munita JM, Hasbun R, Arias CA. 2016. Influence of minimum inhibitory concentration in clinical outcomes of Enterococcus faecium bacteremia treated with daptomycin: is it time to change the breakpoint? Clin Infect Dis 62:1514–1520. doi: 10.1093/cid/ciw173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Satlin MJ, Nicolau DP, Humphries RM, Kuti JL, Campeau SA, Lewis JS II, Weinstein MP, Jorgensen JH. 2020. Development of daptomycin susceptibility breakpoints for Enterococcus faecium and revision of the breakpoints for other enterococcal species by the Clinical and Laboratory Standards Institute. Clin Infect Dis 70:1240–1246. doi: 10.1093/cid/ciz845. [DOI] [PubMed] [Google Scholar]
- 16.Werth BJ, Steed ME, Ireland CE, Tran TT, Nonejuie P, Murray BE, Rose WE, Sakoulas G, Pogliano J, Arias CA, Rybak MJ. 2016. Defining daptomycin resistance prevention exposures in vancomycin-resistant Enterococcus faecium and E. faecalis. Antimicrob Agents Chemother 58:5253–5261. doi: 10.1128/AAC.00098-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Kidd JM, Abdelraouf K, Asempa TE, Humphries RM, Nicolau DP. 2018. Pharmacodynamics of daptomycin against Enterococcus faecium and Enterococcus faecalis in the murine thigh infection model. Antimicrob Agents Chemother 62:e00506-18. doi: 10.1128/AAC.00506-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Smith JR, Barber KE, Raut A, Rybak MJ. 2015. β-Lactams enhance daptomycin activity against vancomycin-resistant Enterococcus faecalis and Enterococcus faecium in in vitro pharmacokinetic/pharmacodynamic models. Antimicrob Agents Chemother 59:2842–2848. doi: 10.1128/AAC.00053-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Smith JR, Barber KE, Raut A, Aboutaleb M, Sakoulas G, Rybak MJ. 2015. β-Lactam combinations with daptomycin provide synergy against vancomycin-resistant Enterococcus faecalis and Enterococcus faecium. J Antimicrob Chemother 70:1738–1743. doi: 10.1093/jac/dkv007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Miller WR, Murray BE, Rice LB, Arias CA. 2020. Resistance in vancomycin-resistant enterococci. Infect Dis Clin North Am 34:751–771. doi: 10.1016/j.idc.2020.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Miller WR, Munita JM, Arias CA. 2014. Mechanisms of antibiotic resistance in enterococci. Expert Rev Anti Infect Ther 12:1221–1236. doi: 10.1586/14787210.2014.956092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Smith D, Singh K, Tran T, Arias C, Murray B. 2016. Daptomycin plus ceftaroline against daptomycin-resistant Enterococcus faecium in a rat model of experimental endocarditis. Open Forum Infect Dis 3:1980. doi: 10.1093/ofid/ofw172.1528. [DOI] [Google Scholar]
- 23.Suh GA, Lodise TP, Tamma PD, Knisely JM, Alexander J, Aslam S, Barton KD, Bizzell E, Totten KMC, Campbell JL, Chan BK, Cunningham SA, Goodman KE, Greenwood-Quaintance KE, Harris AD, Hesse S, Maresso A, Nussenblatt V, Pride D, Rybak MJ, Sund Z, van Duin D, Van Tyne D, Patel R, for the Antibacterial Resistance Leadership Group . 2022. Considerations for the use of phage therapy in clinical practice. Antimicrob Agents Chemother 66:e02071-21. doi: 10.1128/aac.02071-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rubalskii E, Ruemke S, Salmoukas C, Boyle EC, Warnecke G, Tudorache I, Shrestha M, Schmitto JD, Martens A, Rojas SV, Ziesing S, Bochkareva S, Kuehn C, Haverich A. 2020. Bacteriophage therapy for critical infections related to cardiothoracic surgery. Antibiotics 9:232. doi: 10.3390/antibiotics9050232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Onsea J, Soentjens P, Djebara S, Merabishvili M, Depypere M, Spriet I, De Munter P, Pirnay J-P, Lavigne R, Metsemakers W-J. 2019. Bacteriophage application for difficult-to-treat musculoskeletal infections: development of a standardized multidisciplinary treatment protocol. Viruses 11:891. doi: 10.3390/v11100891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Humphries RM, Ambler J, Mitchell SL, Castanheira M, Dingle T, Hindler JA, Koeth L, Sei K, CLSI Methods Development and Standardization Working Group of the Subcommittee on Antimicrobial Susceptibility Testing . 2018. CLSI Methods Development and Standardization Working Group best practices for evaluation of antimicrobial susceptibility tests. J Clin Microbiol 56:e01934-17. doi: 10.1128/JCM.01934-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.McGrath BJ, Kang SL, Kaatz GW, Rybak MJ. 1994. Bactericidal activities of teicoplanin, vancomycin, and gentamicin alone and in combination against Staphylococcus aureus in an in vitro pharmacodynamic model of endocarditis. Antimicrob Agents Chemother 38:2034–2040. doi: 10.1128/AAC.38.9.2034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Kunz Coyne A, Stamper K, Kebriaei R, Holger DJ, El Ghali A, Morrisette T, Biswas B, Wilson M, Deschenes MV, Canfield GS, Duerkop BA, Arias CA, Rybak MJ. 2022. Phage cocktails with daptomycin and ampicillin eradicates biofilm-embedded multidrug-resistant Enterococcus faecium with preserved phage susceptibility. Antibiotics 11:1175. doi: 10.3390/antibiotics11091175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lehman S, Mearns G, Rankin D, Cole R, Smrekar F, Branston S, Morales S. 2019. Design and preclinical development of a phage product for the treatment of antibiotic-resistant Staphylococcus aureus infections. Viruses 11:88. doi: 10.3390/v11010088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lev K, Kunz Coyne AJ, Kebriaei R, Morrisette T, Stamper K, Holger DJ, Canfield GS, Duerkop BA, Arias CA, Rybak MJ. 2022. Evaluation of bacteriophage-antibiotic combination therapy for biofilm-embedded MDR Enterococcus faecium. Antibiotics 11:392. doi: 10.3390/antibiotics11030392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Morrisette T, Lev KL, Kebriaei R, Abdul-Mutakabbir JC, Stamper KC, Morales S, Lehman SM, Canfield GS, Duerkop BA, Arias CA, Rybak MJ. 2020. Bacteriophage-antibiotic combinations for Enterococcus faecium with varying bacteriophage and daptomycin susceptibilities. Antimicrob Agents Chemother 64:e00993-20. doi: 10.1128/AAC.00993-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Morrisette T, Lev KL, Canfield GS, Duerkop BA, Kebriaei R, Stamper KC, Holger D, Lehman SM, Willcox S, Arias CA, Rybak MJ. 2022. Evaluation of bacteriophage cocktails alone and in combination with daptomycin against daptomycin-nonsusceptible Enterococcus faecium. Antimicrob Agents Chemother 66:e0162321. doi: 10.1128/AAC.01623-21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kebriaei R, Lev KL, Stamper KC, Lehman SM, Morales S, Rybak MJ. 2020. Bacteriophage AB-SA01 cocktail in combination with antibiotics against MRSA-VISA strain in an in vitro pharmacokinetic/pharmacodynamic model. Antimicrob Agents Chemother 65:e01863-20. doi: 10.1128/AAC.01863-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Iwata Y, Satou K, Tsuzuku H, Furuichi K, Senda Y, Sakai-Takemori Y, Wada T, Fujita S, Miyake T, Yasuda H, Sakai N, Kitajima S, Toyama T, Shinozaki Y, Sagara A, Miyagawa T, Hara A, Shimizu M, Kamikawa Y, Kaneko S, Wada T. 2017. Down-regulation of the two-component system and cell-wall biosynthesis-related genes was associated with the reversion to daptomycin susceptibility in daptomycin non-susceptible methicillin-resistant Staphylococcus aureus. Eur J Clin Microbiol Infect Dis 36:1839–1845. doi: 10.1007/s10096-017-2999-3. [DOI] [PubMed] [Google Scholar]
- 35.Reyes J, Panesso D, Tran TT, Mishra NN, Cruz MR, Munita JM, Singh KV, Yeaman MR, Murray BE, Shamoo Y, Garsin D, Bayer AS, Arias CA. 2015. A liaR deletion restores susceptibility to daptomycin and antimicrobial peptides in multidrug-resistant Enterococcus faecalis. J Infect Dis 211:1317–1325. doi: 10.1093/infdis/jiu602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Tran TT, Panesso D, Gao H, Roh JH, Munita JM, Reyes J, Diaz L, Lobos EA, Shamoo Y, Mishra NN, Bayer AS, Murray BE, Weinstock GM, Arias CA. 2013. Whole-genome analysis of a daptomycin-susceptible Enterococcus faecium strain and its daptomycin-resistant variant arising during therapy. Antimicrob Agents Chemother 57:261–268. doi: 10.1128/AAC.01454-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Munita JM, Tran TT, Diaz L, Panesso D, Reyes J, Murray BE, Arias CA, 2022. A liaF codon deletion abolishes daptomycin bactericidal activity against vancomycin-resistant Enterococcus faecalis. Antimicrob Agents Chemother 57:2831–2833. 10.1128/AAC.00021-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Diaz L, Tran TT, Munita JM, Miller WR, Rincon S, Carvajal LP, Wollam A, Reyes J, Panesso D, Rojas NL, Shamoo Y, Murray BE, Weinstock GM, Arias CA. 2014. Whole-genome analyses of Enterococcus faecium isolates with diverse daptomycin MICs. Antimicrob Agents Chemother 58:4527–4534. doi: 10.1128/AAC.02686-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Estrella LA, Quinones J, Henry M, Hannah RM, Pope RK, Hamilton T, Teneza-Mora N, Hall E, Biswajit B. 2016. Characterization of novel Staphylococcus aureus lytic phage and defining their combinatorial virulence using the OmniLog system. Bacteriophage 6:e1219440. doi: 10.1080/21597081.2016.1219440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bonilla N, Rojas MI, Netto Flores Cruz G, Hung SH, Rohwer F, Barr JJ. 2016. Phage on tap–a quick and efficient protocol for the preparation of bacteriophage laboratory stocks. PeerJ 4:e2261. doi: 10.7717/peerj.2261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kebriaei R, Rice SA, Singh KV, Stamper KC, Dinh AQ, Rios R, Diaz L, Murray BE, Munita JM, Tran TT, Arias CA, Rybak MJ. 2018. Influence of inoculum effect on the efficacy of daptomycin monotherapy and in combination with β-lactams against daptomycin-susceptible Enterococcus faecium harboring LiaSR substitutions. Antimicrob Agents Chemother 62:e00315-18. doi: 10.1128/AAC.00315-18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Rodriguez-Gonzalez RA, Leung CY, Chan BK, Turner PE, Weitz JS. 2020. Quantitative models of phage-antibiotic combination therapy. mSystems 5:e00756-19. doi: 10.1128/mSystems.00756-19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhvania P, Hoyle NS, Nadareishvili L, Nizharadze D, Kutateladze M. 2017. Phage therapy in a 16-year-old boy with netherton syndrome. Front Med (Lausanne) 4:94. doi: 10.3389/fmed.2017.00094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Kang SL, Rybak MJ. 2012. Pharmacodynamics of RP 59500 alone and in combination with vancomycin against Staphylococcus aureus in an in vitro-infected fibrin clot model. Antimicrob Agents Chemother 35:1505–1511. doi: 10.1128/AAC.39.7.1505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Hall AD, Steed ME, Arias CA, Murray BE, Rybak MJ. 2012. Evaluation of standard- and high-dose daptomycin versus linezolid against vancomycin-resistant Enterococcus isolates in an in vitro pharmacokinetic/pharmacodynamic model with simulated endocardial vegetations. Antimicrob Agents Chemother 56:3174–3180. doi: 10.1128/AAC.06439-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Hall Snyder A, Werth BJ, Barber KE, Sakoulas G, Rybak MJ. 2014. Evaluation of the novel combination of daptomycin plus ceftriaxone against vancomycin-resistant enterococci in an in vitro pharmacokinetic/pharmacodynamic simulated endocardial vegetation model. J Antimicrob Chemother 69:2148–2154. doi: 10.1093/jac/dku113. [DOI] [PubMed] [Google Scholar]
- 47.Werth BJ, Sakoulas G, Rose WE, Pogliano J, Tewhey R, Rybak MJ. 2013. Ceftaroline increases membrane binding and enhances the activity of daptomycin against daptomycin-nonsusceptible vancomycin-intermediate Staphylococcus aureus in a pharmacokinetic/pharmacodynamic model. Antimicrob Agents Chemother 57:66–73. doi: 10.1128/AAC.01586-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.O’Flynn G, Ross RP, Fitzgerald GF, Coffey A. 2004. Evaluation of a cocktail of three bacteriophages for biocontrol of Escherichia coli O157:H7. Appl Environ Microbiol 70:3417–3424. doi: 10.1128/AEM.70.6.3417-3424.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]