

Abstract
The central circadian clock in the brain controls the time-of-the-day variations in acute meal responses, with a low glycemic response but a high satiety/thermogenic response to meals consumed at waking compared to other time points. Consistently, studies show that consuming a significant proportion of calories, particularly carbohydrates, in breakfast is beneficial for the chronic management of obesity and its associated metabolic syndrome, compared to consuming identical meals at dinner. Conversely, breakfast skipping or/and late dinner can have unfavorable metabolic outcomes. It remains controversial how meal frequency affects metabolic health. In contrast, irregular meals, especially irregular breakfasts, show consistent adverse metabolic consequences. Time-restricted feeding (TRF), with all calories consumed within less than 12-h per day, can improve metabolism and extend lifespan. A major component of TRF in humans is caloric restriction, which contributes significantly to the beneficial effects of TRF in humans. By comparison, TRF effects in rodents can be independent of caloric restriction and show day/night phase specificity. TRF could alleviate metabolic abnormalities due to circadian disruption, but its effects appear independent of the circadian clock in rodents. Understanding neuroendocrine mechanisms underlying clock-mediated metabolic regulation will shed light on the metabolic effects of temporal meal patterns.
Keywords: cardiometabolic, circadian rhythm, daily variation, diabetes, meal response, suprachiasmatic nuclei
Introduction
We are what we eat. In addition to “what we eat”, “when we eat” is increasingly recognized as another important factor that determines metabolic health. Since eating is generally a repetitive behavior on a daily basis in humans, it is intrinsically connected to the daily 24-h rhythm of the neuroendocrine system. We will focus on the temporal aspects of meals in this review and will not discuss macronutrient compositions or caloric restriction. Given the vast literature on circadian disruptions and sleep disorders due to changes in light schedules, we will focus on studies on the normal light/dark schedule. For the same reason, we will not discuss genetic mouse models of the circadian clock unless it provides direct insights into the 24-h physiological rhythms under normal light/dark or feeding conditions. Lastly, we will focus on metabolic outcomes related to obesity, diabetes, fatty liver, and other cardiometabolic risk factors.
Circadian clock
The molecular circadian clock generates cue-independent 24-h rhythms in behavior and metabolism physiology. The molecular circadian clock operates in mammalian cells and consists of several interlocked transcriptional-translational feedback loops [1, 2]. Heterodimers of the transcription factors Brain and Muscle ARNT-Like 1 (BMAL1) and CLOCK bind to the E-box elements in the promoter/enhancer regions of the target genes and activate their transcription. Among BMAL1/CLOCK target genes are core clock genes Period (PER) and Cryptochrome (CRY) (Figure 1). PER and CRY proteins form heterodimers that interact with BMAL1/CLOCK and counteract BMAL1/CLOCK-mediated transcription activation. As a result, the levels of PER and CRY transcripts and proteins would decay until the brake on BMAL1/CLOCK is released, followed by another rise of the PER and CRY expression with a 24-h period [1, 2]. The casein kinase controls the rate at which the PER/CRY complex is degraded or enters the nucleus. A loss-of-function mutation in a casein kinase shortens the intrinsic period of the clock in mice and gives rise to sleep phase disorders in humans [3]. Pharmacological modulation of the casein kinases can alter the period by modulating PER localization and stability [4].
Figure 1:
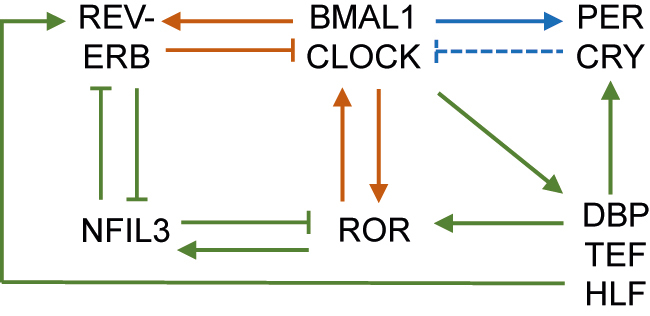
Diagram of the molecular circadian clock machinery.
The blue, red, and green depicts the first, second, and third feedback loops, respectively. The solid lines indicate transcriptional regulation, while the dotted line indicates posttranslational regulation. BMAL1, brain and muscle ARNT-like 1; PER, period; CRY, cryptochrome; ROR, retinoic-acid-receptor-related orphan receptors; NFIL3, nuclear factor interleukin 3 regulated; DBP, albumin D-box binding protein; TEF, thyrotroph embryonic factor; HLF, human hepatic leukemia factor.
While the PER and CRY constitute the primary negative feedback loop, REV-ERB and retinoic-acid-receptor-related orphan receptors (ROR) constitute a second negative loop within the molecular clock machinery. BMAL1/CLOCK stimulates the transcription of nuclear receptors REV-ERB and ROR (Figure 1). The REV-ERB/ROR proteins compete for the ROR elements in the promoter/enhancer regions of the target genes, such as BMAL1, where REV-ERB proteins inhibit BMAL1 transcription and ROR proteins activate it [1, 2]. The REV-ERB/ROR loop also drives the transcription of nuclear factor, interleukin 3 regulated (NFIL3). NFIL3 binds to D-box elements in the promoter/enhancer regions of REV-ERB and ROR to repress their transcription. BMAL1/CLOCK drives other D-box binding proteins, including albumin D-box binding protein (DBP), thyrotroph embryonic factor (TEF), and human hepatic leukemia factor (HLF), creating another transcription loop [1, 2]. These interlocked transcriptional feedback loops of the clock machinery cooperatively regulate the rhythmic expression of clock-controlled genes that serve as the output pathways of the clock to confer temporal cues to various physiological processes.
The molecular clock machinery is expressed in most tissues and organs in mammals. The central clock refers to the clock in the hypothalamic suprachiasmatic nuclei (SCN), while peripheral clocks refer to those expressed in peripheral tissues and other parts of the central nervous system [5]. The SCN clock is kept in alignment with the external photic cues through the retinohypothalamic tract (RHT) that connects the light-sensing melanopsin-expressing intrinsically photoreceptive retinal ganglion cells (ipRGCs) to the SCN [6]. The RHT releases excitatory glutamate as its primary neurotransmitter to depolarize neurons within the SCN [7]. At the molecular level, glutamate receptor activation leads to a post-synaptic increase of intracellular calcium levels, which activates cAMP-responsive element-binding protein (CREB) and then transactivates clock genes, such as PERs. These mechanisms allow the SCN central clock to be aligned with the external photic cues [8], [9], [10].
Output of the SCN clock
The SCN central clock relays temporal signals to other regions of the brain, which coordinates the diurnal rhythm of circulating hormone levels or the autonomic nervous system (ANS) [11, 12]. The SCN is primarily composed of neurons using gamma-aminobutyric acid (GABA) as the neurotransmitter [13] and projects to several hypothalamic nuclei that regulate metabolism, including the subparaventricular zone (SPZ), paraventricular nucleus (PVN), arcuate nucleus (ARC), dorsomedial hypothalamic nucleus (DMH), and medial preoptic area (MPOA) [14, 15].
The SPZ is the main efferent target of neural projections from the SCN and an essential relay for the circadian timing system. Early studies have found that electrolytic lesion of the SPZ abolishes the circadian rhythmicity of the eating and drinking behaviors in rats and reduces the amplitude of curling-up behaviors during sleep [16]. Restricted lesion of subregions of SPZ reveals that separate neuronal populations in the SPZ play distinct roles in circadian rhythms of sleep and body temperature. Ventral SPZ lesion causes substantial loss of circadian rhythms of sleep and locomotor activity but has less effect on body temperature. On the contrary, dorsal SPZ lesion reduces the amplitude of the body temperature rhythm but not that of the locomotor activity or sleep [17]. Thus, ventral SPZ regulates the rhythm of the sleep/wake cycle, whereas dorsal SPZ controls body temperature oscillation. The SCN output to the SPZ extends dorsocaudally to the PVN, a pivotal feeding center. The neurocircuit from the SCN to PVN contributes to the light-mediated suppression of feeding behavior [18]. PVN relays the SCN signal to the pineal gland to control the nocturnal secretion of melatonin [19]. PVN also regulates the daily oscillation of adrenocorticotropic hormone (ACTH), glucocorticoid, and oxytocin [11]. The SCN-to-PVN circuit regulates the ANS and hepatic glucose production [20]. Activation of neuronal activity in the PVN induces time-dependent hyperglycemia only during the light cycle, an effect that is abolished in SCN-ablated animals lacking GABAergic input from the SCN to the PVN [21]. Apart from its connection with the SCN, the PVN also has its own circadian oscillator, as specific deletion of BMAL1 in PVN neurons in mice reduces diurnal rhythmicity in oxygen consumption, feeding, and locomotor activities and promotes diet-induced obesity [22].
The ARC is a hypothalamic nucleus that senses nutrient availability and regulates hunger or satiety [23]. The ARC is located close to the median eminence (ME) that is free of the blood–brain barrier. Therefore, the ARC has access to metabolites and hormones in the circulation [24]. The SCN sends direct projections to ARC neurons expressing α melanocyte-stimulating hormone (αMSH), which influences the ARC neural activity [25]. The bilateral SCN lesion can ablate the nocturnal peak of the ARC neural activity at zeitgeber time ZT22 (ZT0 is when light is on, and ZT12 is when light is off) [25]. Disruption of the SCN-ARC connection in rats results in ARC desynchronization and a loss of rhythmicity in locomotor activity, corticosterone levels, and body temperature without affecting the SCN clock gene expression rhythmicity, suggesting that the neural projection between the SCN and ARC is essential for multiple physiological rhythms [26]. A recent study revealed that SCN input drives the daily regulation of the permeability of the ME-ARC barrier. The SCN enhances the penetrability of glucose from cerebrospinal fluid (CSF) into the ARC by inducing glucose transporter 1 (GLUT1) expression in tanycytes at ZT22 before sleep in rats, which promotes glucose counter-regulation [27].
The SCN projection to the DMH influences various behavioral circadian rhythms [28]. The DMH lesion in rats causes a marked reduction in the circadian rhythms of wakefulness, locomotor activity, feeding, and blood corticosteroid levels [28]. The DMH relays the signals from the SCN to other sleep-regulating regions. For example, the DMH sends GABAergic projections to the ventrolateral preoptic nucleus (VLPO) to inhibit sleep and sends glutamate/thyrotropin-releasing projections to the lateral hypothalamus to promote wakefulness [28]. MPOA also receives projections from the SCN and contributes to the indirect neuronal pathway in circadian control of the sleep/wake cycle [29]. In addition to receiving projections from the SCN, the DMH also sends inhibitory projections to the SCN, which plays a role in food anticipatory activity [30]. The DMH neuronal activity increases while the SCN neuronal activity decreases when animals anticipate food [30].
Daily variations in acute responses to meals
The SCN clock not only controls food eating or food-anticipating behaviors but also regulates responses to meals. The acute glycemic responses to a meal, or glucose tolerance, show a robust diurnal rhythm. In healthy individuals, the same meal at breakfast induces a lower glucose excursion than at dinner in healthy human subjects [31], [32], [33], [34], [35], [36] (Table 1). Similarly, mice or rats have the best glucose tolerance at waking or the early active phase compared to the other times of the day, associated with the best systemic insulin sensitivity [37], [38], [39], [40]. Insulin is secreted in response to dietary carbohydrates to stimulate glucose uptake from the blood and suppress endogenous glucose production from the liver [41]. Hyperinsulinemic euglycemic clamp analysis demonstrates that the diurnal rhythm of insulin sensitivity is attributable to the heightened sensitivity to insulin-mediated suppression of hepatic glucose production at waking [39, 40]. The SCN lesion abolishes the diurnal rhythm of glucose tolerance, suggesting that the SCN central clock is responsible for these time-of-the-day variations in glycemic responses to meals [42]. Knockout of BMAL1 in mice abolishes the diurnal rhythm in systemic insulin sensitivity, suggesting that the circadian clock is responsible for this temporal pattern [38]. Depletion of REV-ERBα/β in the GABAergic neuron in mice (REV-GABA-KO) abolishes the diurnal neural firing pattern of the SCNGABA neurons and disrupts the diurnal rhythm of glycemic responses to meals [40]. Chemogenetic manipulation of SCNGABA neurons can mimic or abolish the waking-specific glucose intolerance in REV-GABA-KO mice, demonstrating that the SCN clock controls the diurnal rhythm of insulin-mediated suppression of hepatic glucose production [40]. Inducible phase-specific re-expression of REV-ERBα in the SCNGABA neurons in REV-GABA-KO mice can rescue the ZT-dependent glucose intolerance [40]. The SCN might relay the temporal information to the liver through neuronal or endocrine mediators without affecting the overall behavioral rhythm under the normal light-dark cycles [43]. These results suggest that the SCN clock regulates the diurnal rhythm of acute glycemic responses to meals and results in better glycemic control at breakfast in healthy subjects. Thus, it is likely better to consume meals with high glycemic indices at breakfast as compared to consuming the same meal at other times of the day.
Table 1:
The effects of temporal meal patterns on metabolism.
| Test subjects | Groups | Duration | Outcomes | Reference, PMID |
|---|---|---|---|---|
| Intrinsic acute responses to meals | ||||
| Humans: healthy men | Identical meal at breakfast vs. noon (n = 6) | Acute | Less postprandial glucose and insulin levels, higher postprandial TG levels | Burdge 2003 [31] 14647218 |
| Human: healthy adults | Isoenergetic big meals at breakfast vs. dinner (n = 6) | Acute | Lower postprandial glucose and better insulin sensitivity indices | Morgan 2012 [32] 22176632 |
| Human: healthy adults | Identical meals at 8am vs. 8pm (n = 20) | Acute | Lower postprandial glucose, insulin, and FFAs levels; higher postprandial metabolic rate | Bo 2015 [33] 26219416 |
| Human: healthy adults | Breakfast vs. dinner (n = 14) | Acute | Lower postprandial glucose levels | Morris 2015 [34] 25870289 |
| Human: healthy men | Meal at morning (9am) vs. evening 5pm) (n = 19) | Acute | Lower postprandial glucose and GIP levels, higher AUC of 5 glycolysis, TCA, and nucleotide-related metabolites, and 18 amino acid-related metabolites | Takahashi 2018 [35] 30441841 |
| Humans: meta-analysis | Identical meal during the day (7am-4pm) vs. night (8pm-4am) | Acute | Lower postprandial glucose levels after identical meals preceded by at least 3 h fast | Leung 2020 [36] 31782659 |
| Rat | 6 time-points (ZT2, 8, 11, 14, 18, 22) | Acute | Lowest meal-induced glucose and insulin increase at ZT14 | Fleur 2001 [37] 11375322 |
| Mouse | 4 time-points (ZT1, 7, 13, 19) | Acute | Highest insulin sensitivity at waking (ZT13) by insulin clamp | Shi 2013 [38] 23434278 |
| Mouse | Active phase (ZT18) vs. sleep (ZT6) | Acute | Higher insulin sensitivity by insulin clamp | Coomans 2013 [39] 23303208 |
| Mouse | Waking (ZT12) vs. sleep (ZT6) or before sleep (ZT0) | Acute | Higher insulin sensitivity by insulin clamp | Ding 2021 [40] 33762728 |
| Humans: healthy adults | Identical meal at noon vs. midnight (n = 25) | Acute | Lower postprandial blood TG levels in men, no difference in women | Sopowski 2001 [44] 11407787 |
| Rat | Oral fat challenge at the beginning of the active phase vs. rest phase | Acute | Lower plasma TG; higher TG uptake in muscle and brown adipose; no differences in the rate of intestinal TG secretion, abolished by SCN lesion | Moran-Ramos 2017 [45] 29113012 |
| Mouse: effect of fish oil on top of a high-sucrose diet | Fish oil at around the activity onset (breakfast) vs. offset (dinner) | Acute | Lower plasma and liver TG and total cholesterol; higher plasma PUFAs | Oishi 2018 [46] 29149647 |
| Human: healthy adults | Identical meals at 8am vs. 8pm (n = 20) | Acute | Higher postprandial epinephrine/norepinephrine and lower acylated ghrelin | Bo 2017 [47] 29255175 |
| Human: healthy adults | Breakfast vs. dinner (n = 13) | Acute | Higher diet-induced thermogenesis | Morris 2015 [48] 26414564 |
| Human: overweight adults | Post-breakfast vs. post-lunch or post-dinner (n = 14) | Acute | Higher thermogenic effects of the meal | Ruddick-Collins 2022 [49] 34473293 |
|
| ||||
| Big breakfast | ||||
| Human: adults | High-carb intake in the morning vs. control (n = 1488) | Observational correlation | Lower incidence of metabolic syndrome using multivariate nutrient density logistic models | Almoosawi 2013 [50] 22777542 |
| Human: adolescents | Morning snacking (n = 180) vs. evening snacking (n = 28) | Cross-sectional observation | Smaller chance of overweight | Bo 2014 [51] 24897170 |
| Human: non-obese adults | Early high-carbohydrate (<1:30 pm), late high-fat (4:30–10:00 pm) vs. inverse order (n = 29) | 4 weeks | Lower whole-day glucose levels in subjects with impaired glucose tolerance | Kessler 2017 [52] 28272464 |
| Human: overweight or obese women | Big breakfast vs. big dinner (n = 36–38 in each group) | 12 weeks of weight loss program | Greater reduction in weight, waist circumference, blood glucose, insulin, TG levels, HOMA-IR, and glucose tolerance; equal calorie intake | Jakubowicz 2013 [53] 23512957 |
| Human: women with PCOS | Big breakfast vs. big dinner (n = about 30 each group) | 12 weeks of weight loss program | Greater reduction in blood glucose, testosterone, and HOMA-IR; higher ovulation rate, despite equal total calorie intake | Jakubowicz 2013 [54] 23688334 |
| Human: adults with type 2 diabetes | Big breakfast vs. big dinner (n = about 20–30 each group) | 12 weeks of weight loss program | Greater reductions in HbA1c, glucose levels, blood pressure, and hunger scores; equal body weight and calorie intake | Rabinovitz 2014 [55] 24311451 |
| Human: obese adults | Big vs. normal breakfast on top of the Mediterranean diet (n = 18) | 12 weeks | Lower body weight, glucose, and lipid levels despite equal total calorie | Lombardo 2014 [56] 24809437 |
| Human: adults with type 2 diabetes | Metformin with big breakfast vs. metformin with big dinner (n = 22) | 7 days | Greater improvement in glucose tolerance, greater increase in insulin secretion, equal total calorie intake | Jakubowicz 2015 [57] 25724569 |
| Human: adults with type 2 diabetes | Low-carb high-fat breakfast vs. normal breakfast (n = 23) | 24 h | Lower glucose levels and glucose variability as measured by CGM | Chang 2019 [58] 30968140 |
|
| ||||
| Breakfast skipping and late dinner | ||||
| Human: men | Breakfast skipping vs. normal 3 meals (n = 29,206) | Observational correlation | Higher risk of type 2 diabetes | Mekary 2012 [60] 22456660 |
| Human: non-shift-working type 2 diabetes patients | Breakfast skipping vs. normal (n = 194) | Observational correlation | Higher HbA1c levels even after adjusting multiple factors | Reutrakul 2014 [61] 24094031 |
| Human: adults with type 2 diabetes | Skip breakfast vs. non-skipping (n = 22) | Acute | Higher postprandial glucose levels after lunch/dinner, lower insulin secretion | Jakubowicz 2015 [62] 26220945 |
| Human: healthy adults | Skip breakfast vs. skip dinner (n = 17) | 24-h | Higher insulin levels and fat oxidation, equal total calorie intake | Nas 2017 [63] 28490511 |
| Human: healthy men | Skip breakfast vs. normal 3 meals (n = 10) | 6 days | Higher mean glucose levels and glycemic variability, no change in energy expenditure | Ogata 2019 [64] 31095288 |
| Human: healthy adults | Skip breakfast vs. normal eating | 6 weeks | No difference in body weight or indexes of cardiovascular health, less glycemic variability | Betts 2014 [65] 24898233 |
| Human: healthy adults | Skip breakfast vs. normal breakfast (n = about 15) | 6 weeks | Same response to food in appetite, glucose, insulin, and thermogenesis | Chowdhury 2018 [66] 29378040 |
| Human: healthy adults | Late dinner vs. early dinner vs. (n = 6) | 3 days | higher RER after breakfast, higher glucose variability | Nakamura 2021 [67] 34371933 |
| Human: overweight/obese women | Late dinner (11pm) vs. early dinner (8 pm) (n = 40) | acute | Impaired glucose tolerance in MTNR1B risk carriers and not in the non-risk carriers. | Lopez-Minguez 2018 [68] 28455106 |
| Human: overweight or obese women | Late eater (lunch after 3 pm) vs. early eater (lunch before 3 pm) (n = about 200) | 20 weeks of weight loss program | Lose less weight; no change in energy intake, diet composition, energy expenditure, appetite hormones, or sleep duration | Garaulet 2013 [69] 23357955 |
| Human: healthy men | Late meals vs. early meals (n = 10) | 13 days | Lower average glucose levels; no change in subjective hunger, sleepiness, or clock markers | Wehrens 2017 [70] 28578930 |
| Human: type 2 diabetes patients | Early dinner before 6pm or skipping dinner vs. habitual eating (n = 20) | 3 days | More nocturnal hypoglycemia incidences | King 2015 [71] 25370327 |
| Human | Self-reported meal timing (n = 133) | Correlation | No correlation with BMI | Marinac 2019 [72] 30798690 |
| Rat | Skip breakfast or dinner vs. no skip | 5 weeks | Lower blood TG, cholesterol, and glucose in the active phase | Wu 2011 [73] 22080734 |
| Human: adults with type 2 diabetes | Night eating vs. non-night eating (n = 194) | Correlation | Poorer glycemic control and depressive symptoms | Hood 2014 [75] 24751916 |
| Human: Pregnant women | Night-eating vs. non-night eating (n = 148) | Correlation | Higher indices of insulin resistance and HbA1c level | Deniz 2019 [76] 31360584 |
| Human: overweight or obese adults | Night-eating vs. non-night eating (n = 615) | Correlation | Higher BMI; higher blood pressure in women; higher waist circumference and blood TG in men | Gallant 2014 [77] 24854802 |
| Human: overweight or obese adults | Night-eating vs. non-night eating (n = 388 total) | Correlation follow-up | Bodyweight gain in non-obese subjects; reduced bodyweight in severely obese subjects | Gallant 2015 [78] 25797608 |
| Human: adults with type 2 diabetes. | Night eaters vs. non-night eater (n = 85) | Correlation | Lower heart rate variability in the low and very-low-frequency bands, and poorer sleep quality | Bermúdez-Millán 2022 [79] 35534102 |
| Human: adults | Dinner immediately before bed or/and snacks after dinner vs. control (n = 8,153) | Correlation | Higher chances of obesity in both sexes, higher chances of metabolic syndrome in women | Yoshida 2018 [80] 30537972 |
| Human: healthy adult participants | Night eating vs. non-night eating (n = 7,771 total) | Correlation | Positive correlation with arterial stiffness in women, but not in men | Zhang 2020 [81] 32954888 |
| Human: adults with type 1 diabetes | Late-night snacking vs. control (n = 148 total) | Correlation | No change in glycemic control in subjects with continuous subcutaneous insulin infusion | Matejko 2015 [83] 26221525 |
|
| ||||
| Meal frequency | ||||
| Human: healthy adults | Meal frequency correlation with BMI (n > 10,000) | Correlation | Eating less frequently and big breakfasts are correlated with lower long-term weight gain | Kahleova 2017 [84] 28701389 |
| Human: type 2 diabetes adult patients | 2 meals (breakfast and lunch) vs. 6 smaller meals (n = 27 each) | 12 weeks on top of oral hypoglycemic agents | Greater reduction in body weight, hepatic lipid content, fasting glucose, C-peptide, glucagon, and insulin resistance; equal caloric intake | Kahleova 2014 [85] 24838678 |
| Human: type 2 diabetes adult patients | 2 meals (breakfast and lunch) vs. 6 smaller meals (n = 27 each) | 12 weeks on top of oral hypoglycemic agents | Higher fasting ghrelin levels despite similar postprandial responses of leptin, GIP, and other appetite hormones and equal total caloric intake | Belinova 2017 [86] 28369078 |
| Human: adults with type 2 diabetes | 3 meals per day vs. 6 meals per day (n = about 14) | 12 weeks | Greater reduction in body weight, HbA1c, appetite, glucose, and insulin; equal total calorie intake | Jakubowicz 2019 [87] 31548244 |
| Human: obese adults | 3 vs. 6 meals per day (n = 13) | 3 days | No change in glucose total AUC; increased postprandial insulin responses | Kanaley 2014 [88] 25231499 |
| Human: healthy adults | 1 vs. 3 meals per day (n = 15) | 8 weeks | Higher fasting glucose levels, impaired morning glucose tolerance, delayed insulin response | Carlson 2007 [89] 17998028 |
| Human: adults with impaired glucose tolerance | 3 vs. 6 eucaloric meals per day (n = 47) | 12 weeks | Less glucose tolerance, same total calorie intake | Papakonstantinou 2018 [90] 29680359 |
| Human: women with PCOS | 3 vs. 6 isocaloric meals per day (n = 24) | 24 weeks | Less post-OGTT insulin sensitivity (Matsuda index), same total calorie intake | Papakonstantinou 2016 [91] 26862008 |
|
| ||||
| Meal regularity | ||||
| Human: overweight or obese adults | High vs. low daily variability in breakfast timing (n = 73 total) | Observational correlation |
High body weight and HbA1c levels | Zhao 2021 [94] 34348822 |
| Human: school children | Irregular vs. regular meal pattern (n = 604) | Observational correlation |
Higher BMI, although breakfast was the only single meal associated with BMI. | Lehto 2011 [95] 21129237 |
| Human: adults | Irregular vs. regular meal pattern (n = about 1400) | Observational, 10 and 17 year follow-up, correlation | Higher BMI, TG, blood pressure, especially with irregular breakfast or lunch | Pot 2016 [96] 26548599 |
| Human: adults | Irregular vs. regular meal pattern (n = 1768) | Observational correlation |
Higher BMI and metabolic syndrome risk with irregular breakfast | Pot 2014 [97] 24675713 |
| Human: adolescent | Irregular vs. regular meal pattern (n = 889) | 27 years follow-up | Higher prevalence of BMI and metabolic syndrome in later life, only with irregular breakfast | Wennberg 2015 [98] 25936413 |
| Human: healthy adult women | Variable meal frequency (3–9 meals/day) vs. 6 regular meals per day (n = 11 total) | 2 weeks | Higher postprandial insulin and fasting total/LDL cholesterol; lower postprandial energy expenditure and thermogenesis; no change in total calorie intake | Farshchi 2004–2005 [99], [100], [101] 15220950 15085170 15640455 |
| Human: normal-weight adult women | Variable meal frequency (3–9 meals/day) vs. 6 regular meals (n = 11) | 2 weeks | Less thermic effect of food, higher glucose AUC, no difference in gut hormones | Alhussain 2016 [102] 27305952 |
|
| ||||
| Time-restricted feeding (TRF) | ||||
| Human: healthy young men | TRF (11 h) vs. baseline (n = 27) | 2 weeks | Lower body weight | LeCheminant 2013 [106] 23702187 |
| Human: overweight or obese adults | TRF (10 h) vs. baseline (before TRF) (n = 8) | 16 weeks | Lower calorie intake, lower body weight, improved sleep | Gill 2015 [107] 26411343 |
| Human: Strength-trained men | TRF (8 h) vs. control (n = 17 per group) | 8 weeks | Lower fat mass, glucose, insulin, TG, and leptin levels | Moro 2016 [108] 27737674 |
| Human: men with prediabetes | Early TRF (6 h, dinner before 3 pm) vs. habitual (n = 8) | 5 weeks | Better insulin sensitivity, lower blood pressure, less oxidative stress, no weight loss | Sutton 2018 [109] 29754952 |
| Human: healthy adults (mostly females) | TRF (delay breakfast and advance dinner by 1.5 h each) vs. habitual (n = 6–7) | 10 weeks | Reduced total calorie intake, reduced adiposity and fasting glucose levels | Antoni 2018 [110] doi.org/10.1017/jns.2018.13 |
| Human: obese adults | TRF (8 h) vs. baseline (before TRF) (n = 23) | 12 weeks | Reduced blood pressure; no change in body weight, glucose, insulin, or lipid levels | Gabel K 2018 [111] 29951594 |
| Human: obese adults | TRF (8 h) vs. baseline (before TRF) (n = 40) | 12 weeks | Lower body weight and HbA1c | Kesztyus 2019 [112] 31766465 |
| Human: overweight older subjects | TRF (8 h) vs. baseline (before TRF) | 4 weeks | Reduced body weight | Anton 2019 [113] 31262054 |
| Human: overweight adults | TRF (8 h) vs. control habitual eating (n = about 10) | 12 weeks | Lower body weight and fat mass, no change in physical activity, glucose, or lipid levels | Chow 2020 [114] 32270927 |
| Human: adults with metabolic disorder | TRF (10 h) vs. before TRF (n = 19) | 12 weeks | Lower body weight, total cholesterol, and blood pressure | Wilkinson 2020 [115] 31813824 |
| Human: healthy young adults | TRF (8 h) vs. baseline (before TRF) (n = 33) | 4 weeks | Lower body weight, insulin levels, no difference in sleep | Park 2021 [116] 34202475 |
| Human: healthy runners | TRF (8 h) vs. habitual eating (n = 7–10) | 8 weeks | Lower body mass and energy intake, no change in endurance performance or metabolism | Brady 2021 [117] 32796255 |
| Human: obese adults | TRF (4 h or 6 h) vs. habitual (n = 20/group) | 8 weeks | Lower body weight, reduced energy intake | Cienfuegos 2020 [118] 32673591 |
| Human: healthy, physically active men | TRF (8 h) vs. isocaloric control (n = 22) | 4 weeks | Lower body weight, fat mass, and blood pressure; no change in glucose, insulin, or lipids | McAllister 2020 [119] 31955013 |
| Human: obese adults | TRF (10 h) vs. TRF (12 h) (n = 39) | 8 weeks | Lower body weight and glucose levels | Peeke PM 2021 [120] 33446635 |
| Human: healthy adults | early TRF (early part of the day) vs. mid-day TRF (n = 82) | 5 weeks | More effective at improving insulin sensitivity; lower glucose, adiposity, and inflammation | Xie 2022 [121] 35194047 |
| Human: overweight adults | early TRF (8 h) vs. habitual eating (n = 11) | 4 days | Lower mean 24 h glucose levels | Jameshed 2019 [122] 31151228 |
| Human: overweight adults | early TRF (8am – 2pm) vs. control (8am – 8pm) (n = about 10) | 4 days | increase protein oxidation, decreased indices of appetite, equal energy expenditure | Ravussin E. 2019 [123] 31339000 |
| Human: obese men | early TRF (8am–5pm) or late TRF (12–9pm) vs. baseline (n = 15) | 7 days | Lower body weight and mean glucose levels in both early TRF and delayed TRF | Hutchison 2019 [124] 31002478 |
| Human: overweight/obese adults | TRF (8 h) vs. habitual eating (n = 11) | 5 days | Lower nocturnal mean glucose levels | Parr 2020 [125] 32079327 |
| Jamshed 2022 [126] 35939311 | ||||
| Human: Strength-trained men | TRF vs. habitual (n = 9 per group) | 8 weeks | No change in lean mass retention or muscle function; reduced energy intake | Tinsley 2017 [127] 27550719 |
| Human: healthy men | TRF vs. habitual (n = 10–20 per group) | 12 weeks | No change in body composition or muscle function, decreased hematocrit, white blood cells, lymphocytes, and neutrophils | Gasmi 2018 [128] 29571007 |
| Human: resistance-trained women | TRF (12–8pm) vs. habitual (n = 17) | 8 weeks | No effects on training-induced muscle hypertrophy or muscular performance improvements | Tinsley 2019 [129] 31268131 |
| Human: healthy adults | TRF (8 h) vs. habitual (n = 22) | 6 weeks | No change in body weight, cardiovascular functions, glucose | Martens 2020 [130] 31975053 |
| Human: type 2 diabetes | TRF (9 h) vs. habitual (n = 19) | 4 weeks | No change in HbA1c or body weight; reduced calorie intake | Parr 2020 [131] 33105701 |
| Human: adults with metabolic disorder | TRF (12 h) vs. standard dietary advice (n = 54) | 6 months | Equal reduction in body weight | Phillips 2021 [132] 33807102 |
| Human: obese adults | TRF 8 h (8am-4pm) vs. equal CR (25%) without TRF (n = 70) | 1 year | Equally effective in reducing body weight, body fat, blood pressure, glucose, dyslipidemia | Liu 2022 [133] 35443107 |
| Human: overweight or adult adults | TRF (2–8pm) vs. 3 structured meals (n = about 60) | 12 weeks | No effects on body weight, fat mass, insulin, glucose, HbA1c levels, or energy expenditure | Lowe 2020 [134] 32986097 |
| Human: obese men without diabetes | TRF (10 h) vs. CR without TRF (n = 15) | 8 weeks | No change in glycemic responses to meals | Zhao 2022 [135] 35150947 |
| Human: at risk of type 2 diabetes | TRF (10 h) vs. habitual eating | 12 weeks | Ongoing | Quist 2020 [136] 32847912 |
| Human: firefighters on a diet program | TRF (10 h) vs. habitual (n = about 75) | 1 year | Ongoing | Manoogian 2021 [137] 34135038 |
| Human: Metabolic syndrome patients | TRF (10 h) vs. habitual | 12 weeks | Ongoing | Świątkiewicz 2021 [138] 33498955 |
| Mouse: high-fat diet | TRF (8 h of the active cycle) vs. ad libitum | 20 weeks | Reduced obesity, insulin, hepatic steatosis and inflammation; improved motor coordination; equal total calorie intake |
Hatori 2012 [139] 22608008 |
| Mouse: high-fat diet | TRF (4 h in the sleep cycle) vs. ad libitum | 18 weeks | Lower body weight, cholesterol, and TNFα levels; improved insulin sensitivity; lower total calorie intake | Sherman 2012 [140] 22593546 |
| Mouse: various diets | TRF (8–9 h) vs. ad libitum | 24 weeks | Attenuated or reverse metabolic diseases on different obesogenic diets, even after temporarily interruptions | Chaix 2014 [141] 25470547 |
| Mouse: high-fat diet | TRF (8–12 h) vs. ad libitum | 9 weeks | Reduce obesity and inflammatory cytokines | Sundaram 2016 [142] 27188906 |
| Mice: high-fat diet | TRF (10 h of the active cycle) vs. ad libitum | 8 weeks | Lower body weight, lower pro-inflammatory genes expression in white adipose tissue | Lee 2021 [143] 34836036 |
| Mice: high-fat diet | TRF (9 h) vs. ad libitum | 12 weeks | Reduce fatty liver and glucose intolerance in both sexes; lower body weight only in males | Chaix 2021 [144] 34407415 |
| Mouse: high-fat diet | Feed in the active vs. sleep cycle | 6 weeks | Less body weight gain | Arble 2009 [145] 19730426 |
| Mouse: high-fat diet | Feed at the early vs. late phase of the active cycle | 12 weeks | Less weight gain, glucose intolerance, insulin, TG, and leptin levels; equal total calories | Bray 2010 [146] 20351731 |
| Mouse: high-fat high-sucrose diet | TRF in the active vs. sleep cycles | 1 week | Lower blood insulin and leptin levels, lower liver lipid contents, but higher glucose levels | Yasumoto 2016 [147] 27085778 |
| Rat | TRF (12 h in the active cycle) vs. TRF (12 h during sleep) | 8 weeks | Smaller adipose tissue, more total food intake, altered gene expression in multiple tissues | Opperhuizen 2016 [148] 27562056 |
| Rat | TRF (10 h in the active cycle) vs. TRF (10 h during sleep) | 4 weeks | Improved glucose tolerance, equal body weight and total calorie intake | De Goede 2019 [149] 31496992 |
| Mouse | TRF (12 h in the active cycle) vs. TRF (12 h during sleep) | >3 years | Better longevity-promoting effects | Acosta-Rodriguez 2022 [150] 35511946 |
| Rat: shiftwork model (sleep deprivation for 8 h in the sleep cycle) | TRF (12 h in the active cycle) vs. TRF (12 h in sleep) or ad libitum | 4 weeks | Less obesity, better metabolic profiles, equal total calorie intake | Salgado-Delgado 2010 [153] 20080873 |
| Mouse: shift work (6 h advance twice weekly) | TRF (fixed 12 h period every 24 h) vs. ad libitum. | 14 weeks | Lower body weight gain, improved glucose tolerance, equal total calorie intake | Oike 2015 [154] 26297949 |
| Mouse: Lacking the circadian clock | TRF (10 h) vs. ad libitum | 12 weeks | Prevents obesity and metabolic disorders in mice with whole-body Cry1/2 KO or liver-specific Bmal1 or Rev-erbα/β KO | Chaix 2019 [155] 30174302 |
| Mouse: SCN clock deficient mice | TRF (12 h) vs. ad libitum on SCN-BMAL1 KO mouse | 10 weeks in constant darkness | Normalize glucose intolerance and weight gain in SCN-Bmal1 KO mice | Kolbe 2019 [156] 31767165 |
TG, triglycerides; FFAs, free fatty acids; GIP, gastric inhibitory peptide; AUC, area under the curve; TCA, tricarboxylic acid cycle; ZT, Zeitgeber time; PUFA, polyunsaturated fatty acids; OGTT, oral glucose tolerance test; HOMA-IR, homeostatic model assessment for insulin resistance; HbA1c, hemoglobin A1C; CGM, continuous glucose monitoring; RER, respiratory exchange ratio; BMI, body mass index; CR, caloric restriction; KO, knockout.
In addition to glycemic responses to meals, triglycerides (TG) levels also respond differently to identical meals at different times of the day, although the difference is less well characterized compared to glycemic responses (Table 1). In healthy men, identical meals at noon result in higher postprandial blood TG levels than breakfast [31] or dinner [44]. The difference between noon and dinner was not observed in women [44]. However, the oral fat challenge in rats at the beginning of the active cycle (equivalent to breakfast) results in lower blood TG levels compared to the early sleep phase [45]. Consistent with the rat study, mice showed lower blood and lipid TG levels after the fish oil challenge at the activity onset (breakfast) compared to activity offset (dinner) [46]. These results suggest different phases between humans and rodents in TG responses to meals.
In addition to TG responses to meals, the satiety and thermogenic effects of the meals also show diurnal rhythms. Identical meals consumed at breakfast produce more postprandial epinephrine and norepinephrine, less acylated ghrelin, and higher diet-induced thermogenesis than the meal consumed at dinner time in healthy human subjects [47], [48], [49] (Table 1). These results suggest that consuming a large proportion of calories at breakfast could be better than at dinner if one aims to control body weight by reducing calorie intake or increasing energy expenditure. In the following sections, we will review human and animal studies that examine the chronic effects of different temporal meal patterns on metabolic health.
Big breakfast
A higher proportion of carbohydrate intake in the morning is associated with a reduced incidence of metabolic syndrome in adults [50, 51] (Table 1). Consistently, morning snacking is associated with smaller chances of being overweight in school adolescents compared to evening snacking [51]. Particularly, the high-carbohydrate breakfast, high-fat dinner meal pattern has a better overall glucose profile than the high-fat breakfast, high-carbohydrate dinner meal pattern in non-obese subjects with impaired glucose tolerance [52]. In patients with metabolic disorders such as obesity, diabetes, or polycystic ovary syndrome (PCOS), dietary intervention with high caloric breakfast and low caloric dinner promoted more robust weight loss compared to low caloric breakfast and high caloric dinner, with a more significant reduction in blood glucose, HbA1c, insulin levels, and insulin resistance indices, without changing the total daily calorie intake [53], [54], [55], [56] (Table 1). Big breakfasts also resulted in greater improvement in glucose tolerance than big dinners in combination with metformin therapy, despite equal total caloric intake [57]. Another study suggests that low-carbohydrate, high-fat breakfast lowered glucose levels and glucose variability in patients with type 2 diabetes [58]. In summary, the current evidence suggests that consuming a significant proportion of calories, particularly carbohydrates, in breakfast is beneficial for the chronic management of obesity and its associated metabolic syndrome compared to consuming the same amount of calories or carbohydrates at dinner.
Breakfast skipping and late dinner
Compared to altering calorie or macronutrient distributions among different meals, skipping some meals altogether is a more drastic change in temporal meal pattern [59]. Breakfast skipping is associated with a higher risk of type 2 diabetes in the general population [60] and with higher HbA1C levels among type 2 diabetes patients [61]. Breakfast skipping elevates postprandial or mean glucose levels, increases glycemic variability, and lowers insulin secretion in humans in some studies [62], [63], [64] (Table 1). Other studies did not observe changes in glucose/insulin levels or even observed less glycemic variability after breakfast skipping [65, 66].
Breakfast skipping is usually associated with a later chronotype, which contributes to late dinner. Late dinner leads to higher glycemic variability [67] and impaired glucose tolerance in subjects that are genetically predisposed to diabetes [68]. In a 20-week weight-loss trial, overweight women grouped in the late lunch eaters lost less weight and had a slower weight loss rate than early eaters despite similar energy intake, dietary composition, energy expenditure, appetite hormones, and sleep duration [69]. However, healthy men who eat identical meals 5 h later have decreased average glucose than those of an earlier schedule in another study [70]. Early dinner or dinner skipping was found to increase nocturnal hypoglycemia incidences [71] (Table 1). No correlation was found between the early or late meal timing with BMI [72]. Rats that skip the first or the last meal in the active cycle show similar levels of reduction in blood levels of glucose, TG, and cholesterol [73]. Meal skipping on top of the 3-meal/day pattern in small rodents could involve energy imbalance due to their high baseline metabolic rate. Therefore, rodent models are not particularly helpful in addressing the controversy. The results overall suggest that breakfast skipping or/and late dinner can have unfavorable metabolic outcomes.
An extreme case of delayed eating behaviors is night eating syndrome (NES) [74]. The NES is characterized by consuming > 25% of the total daily calorie after the last meal and/or at least two episodes of nocturnal awakening per week with food ingestion, morning anorexia, a strong urge to eat after supper and before initiation of the sleep or upon awakening from the sleep, insomnia, depression [74]. Patients with NES showed phase delay or amplitude reduction of oscillatory hormones involved in appetite regulation and nutrient metabolism, including insulin, ghrelin, leptin, melatonin, and glucocorticoid. Positive correlations were found between NES and poor glycemic control, higher HbA1c, and higher indices of insulin resistance [75, 76] (Table 1). NES was also shown to be correlated with poor cardiometabolic parameters, including high BMI, higher blood pressure, higher blood TG levels, and arterial stiffness [77], [78], [79], [80], [81]. The correlations appear more robust in women than men [80, 81]. However, the relationship between NES and BMI is mixed [82], and some studies did not find a correlation between late-night snacking and glycemic control [83].
Meal frequency and regularity
In addition to the phase of the meals (early vs. late), another aspect of the temporal pattern is the frequency. Shall we eat multiple smaller meals or fewer bigger meals? The frequency and phase are two independent questions theoretically but are intrinsically intertwined in reality because less frequent meals need to be usually eaten at a specific phase, either early or late. In a population-wide observational correlation study, eating less frequently with big breakfasts is associated with lower long-term body weight [84]. Interventional studies on human subjects with type 2 diabetes show that 2–3 meals per day for 12 weeks can cause a greater reduction in body weight, blood glucose and HbA1c levels, and insulin resistance compared to 6 meals per day without affecting total caloric intake [85], [86], [87] (Table 1). However, several studies found no change or even higher glucose levels and less glucose tolerance after reducing meal frequency as compared to 6 meals per day, with equal total caloric intake [88], [89], [90], [91] (Table 1). A meta-analysis found that higher meal frequency may lower total cholesterol and LDL cholesterol levels without affecting glycemic control [92]. The controversy about the effects of altering meal frequency may be attributable to the fact that ‘less meal frequency’ is a mixed entity and involves many variables that need to be carefully controlled during studies.
Compared to meal frequency, more consistent results were reported about meal regularity. Consuming meals irregularly is correlated with higher cardiometabolic risks [93]. Higher daily meal variability is associated with higher BMI, HbA1c, TG, cholesterol, glucose, blood pressure, and thermogenic effects in multiple observational studies in both adults and adolescents [94], [95], [96], [97], [98] (Table 1). Interventional studies also showed that variable meal frequency with 3–9 meals per day for 2 weeks increases cholesterol, glucose, insulin, and lower thermic effects of food as compared to regular 6 meals per day in healthy adults [99], [100], [101], [102] (Table 1). In many studies, the effects of meal irregularity are the most robust or only observed for irregular breakfasts [95], [96], [97], [98] (Table 1). Therefore, irregular meals, especially irregular breakfasts, have adverse metabolic outcomes.
Time-restricted feeding
Time-restricted feeding (TRF) is a dietary intervention limiting daily food intake to a finite time window of 12-h or less every day [103, 104]. By comparison, intermittent fasting (IF) refers to fasting for a more extended period, usually 16–48 h, with intervening periods of normal food intake, on a recurring basis. Periodic fasting (PF) refers to an even longer time scale with fasting or a ‘fasting-mimicking diet’ lasting more than 2 days. IF and PF involves calorie restriction (CR) while TRF may not because more than 8 h of access to food every day is enough for mice to eat the same calorie as ad libitum feeding [105], although TRF in humans tends to lead to reduced caloric intake. As a study tool in animals, TRF can be used to dissect the role of meal timing or misalignment between the central clock and peripheral clocks without the confounding effects of calorie restriction in rodent models. As a therapeutic intervention in humans, TRF also has better patient compliance than IF or PF.
Daily TRF within 6–11 h for 2–12 weeks can reduce caloric intake and lower body weight, glucose levels, and blood lipid levels compared to habitual eating patterns without TRF in human subjects [106], [107], [108], [109], [110], [111], [112], [113], [114], [115], [116], [117], [118], [119], [120], [121] (Table 1). The reduction in body fat, glucose, and lipids can be independent of the total caloric intake [119]. TRF within 10 h was shown to have a more robust reduction in body weight and glucose levels compared to TRF of 12-h [120]. Even 4–7 days of TRF, especially early TRF that restricts eating in the morning and early afternoon, can lower body weight, glucose levels, blood pressure and HbA1c levels in obese human subjects [121], [122], [123], [124], [125], [126] (Table 1). Several studies did not observe TRF-mediated changes in body composition, body weight, or blood metabolites compared to the habitual eating pattern in healthy subjects [127], [128], [129], [130], [131] (Table 1). TRF also did not show superior benefit to caloric restriction or standard dietary guidelines in patients with diabetes or weight or obesity [132], [133], [134], [135], but did show more robust effects in lowering body weight in a recent study [126] (Table 1). Several ongoing studies aim to provide more conclusive evidence about the health outcome of long-term TRF [136], [137], [138] (Table 1). In summary, the current data suggest that a major component of TRF in humans is caloric restriction, which contributes significantly to the beneficial effects of TRF in human subjects.
In contrast to humans, TRF in rodent models can occur without altering the total caloric intake. TRF on obese mice reduces obesity, insulin resistance, glucose levels, liver lipid content, and inflammatory markers compared to ad libitum without altering the total caloric intake [139], [140], [141], [142], [143], [144] (Table 1). When compared to TRF in the sleep cycle, TRF in the active cycle showed better metabolic outcomes, including lower weight gain, fat mass, glucose, and TG levels in mice or rats [145], [146], [147], [148], [149], [150] (Table 1), demonstrating that the phase of TRF matters. In particular, TRF with 35% caloric restriction within the active cycle prolonged lifespan more profoundly than the same degree of caloric restriction spread out in the day or within the sleep cycle in mice, which was associated with the restoration of the liver gene expression rhythm that was dampened by aging. These results demonstrate the phase-specific TRF effect in rodent animals [150]. In support of this notion, 30% calorie restriction by dietary dilution, in which mice ate all day to compensate for the low energy density of the diet, had no beneficial effects on lifespan [151] even though dietary dilution was shown to be slightly more effective in reducing body fat than calorie restriction [152] (Table 1). In addition to combating overnutrition, TRF was also efficacious in correcting the metabolic disruptions due to circadian disruption. TRF in the active cycle or in the fixed time in mice or rats on top of the shiftwork models can reduce obesity and improve glucose tolerance [153, 154]. TRF in genetic mouse models lacking core clock genes can also reduce body weight gain or improve glucose tolerance [155, 156]. These results suggest that TRF could be a dietary intervention for circadian disruption. However, the results also demonstrate that the effects of TRF in rodent species were independent of the presence of the intact circadian clock. The available animal studies provide limited information regarding whether or which hypothalamic nuclei are involved in the metabolic regulatory effects of TRF.
Conclusions
The central clock relays the temporal cues through neural or endocrine systems to metabolic organs throughout the body. The central clock controls the time-of-the-day variations in meal-induced responses, with a low glycemic response but a high satiety response to meals consumed at waking compared to that before sleep. These physiologic rhythms of acute meal responses suggest that the best time to consume meals, especially those high in calories or carbohydrates, is at the early phase of the wake cycle, i.e., breakfast. Consistently, the current studies on humans support that consuming a significant proportion of calories, particularly carbohydrates, in breakfast is beneficial for the chronic management of obesity and its associated metabolic syndrome compared to consuming the same amount of calories or carbohydrates at dinner. Conversely, breakfast skipping or/and late dinner can have unfavorable metabolic outcomes. Night eating syndrome, a more severe form of late eating, is found by some studies to be correlated with adiposity, especially in women. It remains controversial how altering meal frequency affects metabolic health, probably because ‘less meal frequency’ is a mixed entity and involves many variables that need to be carefully controlled during studies. In contrast, irregular meals, especially irregular breakfasts, show consistent adverse metabolic outcomes. TRF can improve systemic metabolism and extend lifespan. The current data suggest that a major component of TRF in humans is caloric restriction, which contributes significantly to the beneficial effects of TRF in human subjects. In contrast, TRF effects in rodent models can be separated from caloric restriction and show day/night phase specificity. TRF could be a dietary intervention for circadian disruption. However, the results also demonstrate that the effects of TRF in rodent species were not necessarily dependent on the presence of the intact circadian clock. Future investigation of neuroendocrine mechanisms underlying central clock connection with systemic metabolism will help understand the etiology of time-related metabolic conditions and provide insights into therapeutic dietary interventions that manipulate temporal meal patterns.
Footnotes
Research funding: The investigators are supported by NIH (DK111436, HL153320, AG069966, and ES027544), the John S. Dunn Foundation, the Mrs. Clifford Elder White Graham Endowed Research Fund, the Dan L Duncan Comprehensive Cancer Center (P30CA125123), the Texas Medical Center Digestive Diseases Center (P30DK056338), the SPORE program in lymphoma at Baylor College of Medicine (P50 CA126752), and the Gulf Coast Center for Precision Environmental Health (P30ES030285).
Author contributions: All authors have accepted responsibility for the entire content of this manuscript and approved its submission.
Competing interests: Authors state no conflict of interest.
Informed consent: Informed consent was obtained from all individuals included in this study.
Ethical approval: The local Institutional Review Board deemed the study exempt from review.
References
- 1.Patke A, Young MW, Axelrod S. Molecular mechanisms and physiological importance of circadian rhythms. Nat Rev Mol Cell Biol. 2020;21:67–84. doi: 10.1038/s41580-019-0179-2. [DOI] [PubMed] [Google Scholar]
- 2.Partch CL, Green CB, Takahashi JS. Molecular architecture of the mammalian circadian clock. Trends Cell Biol. 2014;24:90–9. doi: 10.1016/j.tcb.2013.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Xu Y, Padiath QS, Shapiro RE, Jones CR, Wu SC, Saigoh N, et al. Functional consequences of a CKIdelta mutation causing familial advanced sleep phase syndrome. Nature. 2005;434:640–4. doi: 10.1038/nature03453. [DOI] [PubMed] [Google Scholar]
- 4.Narasimamurthy R, Virshup DM. The phosphorylation switch that regulates ticking of the circadian clock. Mol Cell. 2021;81:1133–46. doi: 10.1016/j.molcel.2021.01.006. [DOI] [PubMed] [Google Scholar]
- 5.Mohawk JA, Green CB, Takahashi JS. Central and peripheral circadian clocks in mammals. Annu Rev Neurosci. 2012;35:445–62. doi: 10.1146/annurev-neuro-060909-153128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.LeGates TA, Fernandez DC, Hattar S. Light as a central modulator of circadian rhythms, sleep and affect. Nat Rev Neurosci. 2014;15:443–54. doi: 10.1038/nrn3743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hastings MH, Maywood ES, Brancaccio M. Generation of circadian rhythms in the suprachiasmatic nucleus. Nat Rev Neurosci. 2018;19:453–69. doi: 10.1038/s41583-018-0026-z. [DOI] [PubMed] [Google Scholar]
- 8.Colwell CS. Linking neural activity and molecular oscillations in the SCN. Nat Rev Neurosci. 2011;12:553–69. doi: 10.1038/nrn3086. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sakamoto K, Norona FE, Alzate-Correa D, Scarberry D, Hoyt KR, Obrietan K. Clock and light regulation of the CREB coactivator CRTC1 in the suprachiasmatic circadian clock. J Neurosci. 2013;33:9021–7. doi: 10.1523/jneurosci.4202-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Hastings MH, Brancaccio M, Maywood ES. Circadian pacemaking in cells and circuits of the suprachiasmatic nucleus. J Neuroendocrinol. 2014;26:2–10. doi: 10.1111/jne.12125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kalsbeek A, Palm IF, La Fleur SE, Scheer Fa. JL, Perreau-Lenz S, Ruiter M, et al. SCN outputs and the hypothalamic balance of life. J Biol Rhythm. 2006;21:458–69. doi: 10.1177/0748730406293854. [DOI] [PubMed] [Google Scholar]
- 12.Kalsbeek A, Bruinstroop E, Yi CX, Klieverik LP, La Fleur SE, Fliers E. Hypothalamic control of energy metabolism via the autonomic nervous system. Ann N Y Acad Sci. 2010;1212:114–29. doi: 10.1111/j.1749-6632.2010.05800.x. [DOI] [PubMed] [Google Scholar]
- 13.Ono D, Honma K, Yanagawa Y, Yamanaka A, Honma S. GABA in the suprachiasmatic nucleus refines circadian output rhythms in mice. Commun Biol. 2019;2:1–12. doi: 10.1038/s42003-019-0483-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Leak RK, Moore RY. Topographic organization of suprachiasmatic nucleus projection neurons. J Comp Neurol. 2001;433:312–34. doi: 10.1002/cne.1142. [DOI] [PubMed] [Google Scholar]
- 15.Morin LP. Neuroanatomy of the extended circadian rhythm system. Exp Neurol. 2013;243:4–20. doi: 10.1016/j.expneurol.2012.06.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Cipolla-Neto J, Afeche SC, Menna-Barreto L, Marques N, Benedito-Silva AA, Fortunato G, et al. Lack of similarity between the effect of lesions of the suprachiasmatic nucleus and subparaventricular hypothalamic zone on behavioral circadian rhythms. Braz J Med Biol Res. 1988;21:653–4. [PubMed] [Google Scholar]
- 17.Lu J, Zhang Y-H, Chou TC, Gaus SE, Elmquist JK, Shiromani P, et al. Contrasting effects of ibotenate lesions of the paraventricular nucleus and subparaventricular zone on sleep–wake cycle and temperature regulation. J Neurosci. 2001;21:4864–74. doi: 10.1523/jneurosci.21-13-04864.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Santoso P, Nakata M, Ueta Y, Yada T. Suprachiasmatic vasopressin to paraventricular oxytocin neurocircuit in the hypothalamus relays light reception to inhibit feeding behavior. Am J Physiol Endocrinol Metab. 2018;315:E478–88. doi: 10.1152/ajpendo.00338.2016. [DOI] [PubMed] [Google Scholar]
- 19.Moore RY, Danchenko RL. Paraventricular–subparaventricular hypothalamic lesions selectively affect circadian function. Chronobiol Int. 2002;19:345–60. doi: 10.1081/cbi-120002876. [DOI] [PubMed] [Google Scholar]
- 20.Kalsbeek A. Suprachiasmatic GABAergic inputs to the paraventricular nucleus control plasma glucose concentrations in the rat via sympathetic innervation of the liver. J Neurosci. 2004;24:7604–13. doi: 10.1523/jneurosci.5328-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kalsbeek A, Foppen E, Schalij I, Heijningen CV, van der Vliet J, Fliers E, et al. Circadian control of the daily plasma glucose rhythm: an interplay of GABA and glutamate. PLoS One. 2008;3:e3194. doi: 10.1371/journal.pone.0003194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kim ER, Xu Y, Cassidy RM, Lu Y, Yang Y, Tian J, et al. Paraventricular hypothalamus mediates diurnal rhythm of metabolism. Nat Commun. 2020;11:3794. doi: 10.1038/s41467-020-17578-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Joly-Amado A, Cansell C, Denis RGP, Delbes A-S, Castel J, Martinez S, et al. The hypothalamic arcuate nucleus and the control of peripheral substrates. Best Pract Res Clin Endocrinol Metab. 2014;28:725–37. doi: 10.1016/j.beem.2014.03.003. [DOI] [PubMed] [Google Scholar]
- 24.Rodríguez EM, Blázquez JL, Guerra M. The design of barriers in the hypothalamus allows the median eminence and the arcuate nucleus to enjoy private milieus: the former opens to the portal blood and the latter to the cerebrospinal fluid. Peptides. 2010;31:757–76. doi: 10.1016/j.peptides.2010.01.003. [DOI] [PubMed] [Google Scholar]
- 25.Guzmán-Ruiz M, Saderi N, Cazarez-Márquez F, Guerrero-Vargas NN, Basualdo MC, Acosta-Galván G, et al. The suprachiasmatic nucleus changes the daily activity of the arcuate nucleus α-MSH neurons in male rats. Endocrinology. 2014;155:525–35. doi: 10.1210/en.2013-1604. [DOI] [PubMed] [Google Scholar]
- 26.Buijs FN, Guzmán-Ruiz M, León-Mercado L, Basualdo MC, Escobar C, Kalsbeek A, et al. Suprachiasmatic nucleus interaction with the arcuate nucleus; essential for organizing physiological rhythms. eNeuro. 2017;24:ENEURO.0028–17. doi: 10.1523/ENEURO.0028-17.2017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rodríguez-Cortés B, Hurtado-Alvarado G, Martínez-Gómez R, León-Mercado LA, Prager-Khoutorsky M, Buijs RM. Suprachiasmatic nucleus-mediated glucose entry into the arcuate nucleus determines the daily rhythm in blood glycemia. Curr Biol. 2022;32:796–805.e4. doi: 10.1016/j.cub.2021.12.039. [DOI] [PubMed] [Google Scholar]
- 28.Chou TC, Scammell TE, Gooley JJ, Gaus SE, Saper CB, Lu J. Critical role of dorsomedial hypothalamic nucleus in a wide range of behavioral circadian rhythms. J Neurosci. 2003;23:10691–702. doi: 10.1523/jneurosci.23-33-10691.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Deurveilher S, Burns J, Semba K. Indirect projections from the suprachiasmatic nucleus to the ventrolateral preoptic nucleus: a dual tract-tracing study in rat. Eur J Neurosci. 2002;16:1195–213. doi: 10.1046/j.1460-9568.2002.02196.x. [DOI] [PubMed] [Google Scholar]
- 30.Acosta-Galvan G, Yi C-X, van der Vliet J, Jhamandas JH, Panula P, Angeles-Castellanos M, et al. Interaction between hypothalamic dorsomedial nucleus and the suprachiasmatic nucleus determines intensity of food anticipatory behavior. Proc Natl Acad Sci U S A. 2011;108:5813–8. doi: 10.1073/pnas.1015551108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Burdge GC, Jones AE, Frye SM, Goodson L, Wootton SA. Effect of meal sequence on postprandial lipid, glucose and insulin responses in young men. Eur J Clin Nutr. 2003;57:1536–44. doi: 10.1038/sj.ejcn.1601722. [DOI] [PubMed] [Google Scholar]
- 32.Morgan LM, Shi JW, Hampton SM, Frost G. Effect of meal timing and glycaemic index on glucose control and insulin secretion in healthy volunteers. Br J Nutr. 2012;108:1286–91. doi: 10.1017/s0007114511006507. [DOI] [PubMed] [Google Scholar]
- 33.Bo S, Fadda M, Castiglione A, Ciccone G, De Francesco A, Fedele D, et al. Is the timing of caloric intake associated with variation in diet-induced thermogenesis and in the metabolic pattern? A randomized cross-over study. Int J Obes. 2015;39:1689–95. doi: 10.1038/ijo.2015.138. [DOI] [PubMed] [Google Scholar]
- 34.Morris CJ, Yang JN, Garcia JI, Myers S, Bozzi I, Wang W, et al. Endogenous circadian system and circadian misalignment impact glucose tolerance via separate mechanisms in humans. Proc Natl Acad Sci U S A. 2015;112:E2225–34. doi: 10.1073/pnas.1418955112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Takahashi M, Ozaki M, Kang M-I, Sasaki H, Fukazawa M, Iwakami T, et al. Effects of meal timing on postprandial glucose metabolism and blood metabolites in healthy adults. Nutrients. 2018;10:E1763. doi: 10.3390/nu10111763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Leung GKW, Huggins CE, Ware RS, Bonham MP. Time of day difference in postprandial glucose and insulin responses: systematic review and meta-analysis of acute postprandial studies. Chronobiol Int. 2020;37:311–26. doi: 10.1080/07420528.2019.1683856. [DOI] [PubMed] [Google Scholar]
- 37.la Fleur SE, Kalsbeek A, Wortel J, Fekkes ML, Buijs RM. A daily rhythm in glucose tolerance: a role for the suprachiasmatic nucleus. Diabetes. 2001;50:1237–43. doi: 10.2337/diabetes.50.6.1237. [DOI] [PubMed] [Google Scholar]
- 38.Shi S, Ansari TS, McGuinness OP, Wasserman DH, Johnson CH. Circadian disruption leads to insulin resistance and obesity. Curr Biol. 2013;23:372–81. doi: 10.1016/j.cub.2013.01.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Coomans CP, van den Berg SAA, Houben T, van Klinken JB, van den Berg R, Pronk ACM, et al. Detrimental effects of constant light exposure and high-fat diet on circadian energy metabolism and insulin sensitivity. FASEB J. 2013;27:1721–32. doi: 10.1096/fj.12-210898. [DOI] [PubMed] [Google Scholar]
- 40.Ding G, Li X, Hou X, Zhou W, Gong Y, Liu F, et al. REV-ERB in GABAergic neurons controls diurnal hepatic insulin sensitivity. Nature. 2021;592:763–7. doi: 10.1038/s41586-021-03358-w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Saltiel AR, Kahn CR. Insulin signalling and the regulation of glucose and lipid metabolism. Nature. 2001;414:799–806. doi: 10.1038/414799a. [DOI] [PubMed] [Google Scholar]
- 42.Fleur SEla, Kalsbeek A, Wortel J, Fekkes ML, Buijs RM. A daily rhythm in glucose tolerance: a role for the suprachiasmatic nucleus. Diabetes. 2001;50:1237–43. doi: 10.2337/diabetes.50.6.1237. [DOI] [PubMed] [Google Scholar]
- 43.Peng F, Li X, Xiao F, Zhao R, Sun Z. Circadian clock, diurnal glucose metabolic rhythm, and dawn phenomenon. Trends Neurosci. 2022;45:471–82. doi: 10.1016/j.tins.2022.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Sopowski MJ, Hampton SM, Ribeiro DC, Morgan L, Arendt J. Postprandial triacylglycerol responses in simulated night and day shift: gender differences. J Biol Rhythm. 2001;16:272–6. doi: 10.1177/074873001129001881. [DOI] [PubMed] [Google Scholar]
- 45.Moran-Ramos S, Guerrero-Vargas NN, Mendez-Hernandez R, Basualdo MDC, Escobar C, Buijs RM. The suprachiasmatic nucleus drives day-night variations in postprandial triglyceride uptake into skeletal muscle and brown adipose tissue. Exp Physiol. 2017;102:1584–95. doi: 10.1113/ep086026. [DOI] [PubMed] [Google Scholar]
- 46.Oishi K, Konishi T, Hashimoto C, Yamamoto S, Takahashi Y, Shiina Y. Dietary fish oil differentially ameliorates high-fructose diet-induced hepatic steatosis and hyperlipidemia in mice depending on time of feeding. J Nutr Biochem. 2018;52:45–53. doi: 10.1016/j.jnutbio.2017.09.024. [DOI] [PubMed] [Google Scholar]
- 47.Bo S, Broglio F, Settanni F, Parasiliti Caprino M, Ianniello A, Mengozzi G, et al. Effects of meal timing on changes in circulating epinephrine, norepinephrine, and acylated ghrelin concentrations: a pilot study. Nutr Diabetes. 2017;7:303. doi: 10.1038/s41387-017-0010-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Morris CJ, Garcia JI, Myers S, Yang JN, Trienekens N, Scheer FAJL. The human circadian system has a dominating role in causing the morning/evening difference in diet-induced thermogenesis. Obesity. 2015;23:2053–8. doi: 10.1002/oby.21189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Ruddick-Collins LC, Flanagan A, Johnston JD, Morgan PJ, Johnstone AM. Circadian rhythms in resting metabolic rate account for apparent daily rhythms in the thermic effect of food. J Clin Endocrinol Metab. 2022;107:e708–15. doi: 10.1210/clinem/dgab654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Almoosawi S, Prynne CJ, Hardy R, Stephen AM. Time-of-day and nutrient composition of eating occasions: prospective association with the metabolic syndrome in the 1946 British birth cohort. Int J Obes. 2013;37:725–31. doi: 10.1038/ijo.2012.103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bo S, De Carli L, Venco E, Fanzola I, Maiandi M, De Michieli F, et al. Impact of snacking pattern on overweight and obesity risk in a cohort of 11- to 13-year-old adolescents. J Pediatr Gastroenterol Nutr. 2014;59:465–71. doi: 10.1097/mpg.0000000000000453. [DOI] [PubMed] [Google Scholar]
- 52.Kessler K, Hornemann S, Petzke KJ, Kemper M, Kramer A, Pfeiffer AFH, et al. The effect of diurnal distribution of carbohydrates and fat on glycaemic control in humans: a randomized controlled trial. Sci Rep. 2017;7:44170. doi: 10.1038/srep44170. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Jakubowicz D, Barnea M, Wainstein J, Froy O. High caloric intake at breakfast vs. dinner differentially influences weight loss of overweight and obese women. Obesity. 2013;21:2504–12. doi: 10.1002/oby.20460. [DOI] [PubMed] [Google Scholar]
- 54.Jakubowicz D, Barnea M, Wainstein J, Froy O. Effects of caloric intake timing on insulin resistance and hyperandrogenism in lean women with polycystic ovary syndrome. Clin Sci (Lond) 2013;125:423–32. doi: 10.1042/cs20130071. [DOI] [PubMed] [Google Scholar]
- 55.Rabinovitz HR, Boaz M, Ganz T, Jakubowicz D, Matas Z, Madar Z, et al. Big breakfast rich in protein and fat improves glycemic control in type 2 diabetics. Obesity. 2014;22:E46–54. doi: 10.1002/oby.20654. [DOI] [PubMed] [Google Scholar]
- 56.Lombardo M, Bellia A, Padua E, Annino G, Guglielmi V, D’Adamo M, et al. Morning meal more efficient for fat loss in a 3-month lifestyle intervention. J Am Coll Nutr. 2014;33:198–205. doi: 10.1080/07315724.2013.863169. [DOI] [PubMed] [Google Scholar]
- 57.Jakubowicz D, Wainstein J, Ahrén B, Bar-Dayan Y, Landau Z, Rabinovitz HR, et al. High-energy breakfast with low-energy dinner decreases overall daily hyperglycaemia in type 2 diabetic patients: a randomised clinical trial. Diabetologia. 2015;58:912–9. doi: 10.1007/s00125-015-3524-9. [DOI] [PubMed] [Google Scholar]
- 58.Chang CR, Francois ME, Little JP. Restricting carbohydrates at breakfast is sufficient to reduce 24-hour exposure to postprandial hyperglycemia and improve glycemic variability. Am J Clin Nutr. 2019;109:1302–9. doi: 10.1093/ajcn/nqy261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Lopez-Minguez J, Gómez-Abellán P, Garaulet M. Timing of breakfast, lunch, and dinner. effects on obesity and metabolic risk. Nutrients. 2019;11:E2624. doi: 10.3390/nu11112624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Mekary RA, Giovannucci E, Willett WC, van Dam RM, Hu FB. Eating patterns and type 2 diabetes risk in men: breakfast omission, eating frequency, and snacking. Am J Clin Nutr. 2012;95:1182–9. doi: 10.3945/ajcn.111.028209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Reutrakul S, Hood MM, Crowley SJ, Morgan MK, Teodori M, Knutson KL. The relationship between breakfast skipping, chronotype, and glycemic control in type 2 diabetes. Chronobiol Int. 2014;31:64–71. doi: 10.3109/07420528.2013.821614. [DOI] [PubMed] [Google Scholar]
- 62.Jakubowicz D, Wainstein J, Ahren B, Landau Z, Bar-Dayan Y, Froy O. Fasting until noon triggers increased postprandial hyperglycemia and impaired insulin response after lunch and dinner in individuals with type 2 diabetes: a randomized clinical trial. Diabetes Care. 2015;38:1820–6. doi: 10.2337/dc15-0761. [DOI] [PubMed] [Google Scholar]
- 63.Nas A, Mirza N, Hägele F, Kahlhöfer J, Keller J, Rising R, et al. Impact of breakfast skipping compared with dinner skipping on regulation of energy balance and metabolic risk. Am J Clin Nutr. 2017;105:1351–61. doi: 10.3945/ajcn.116.151332. [DOI] [PubMed] [Google Scholar]
- 64.Ogata H, Kayaba M, Tanaka Y, Yajima K, Iwayama K, Ando A, et al. Effect of skipping breakfast for 6 days on energy metabolism and diurnal rhythm of blood glucose in young healthy Japanese males. Am J Clin Nutr. 2019;110:41–52. doi: 10.1093/ajcn/nqy346. [DOI] [PubMed] [Google Scholar]
- 65.Betts JA, Richardson JD, Chowdhury EA, Holman GD, Tsintzas K, Thompson D. The causal role of breakfast in energy balance and health: a randomized controlled trial in lean adults. Am J Clin Nutr. 2014;100:539–47. doi: 10.3945/ajcn.114.083402. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Chowdhury EA, Richardson JD, Tsintzas K, Thompson D, Betts JA. Postprandial metabolism and appetite do not differ between lean adults that eat breakfast or morning fast for 6 weeks. J Nutr. 2018;148:13–21. doi: 10.1093/jn/nxx004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Nakamura K, Tajiri E, Hatamoto Y, Ando T, Shimoda S, Yoshimura E. Eating dinner early improves 24-h blood glucose levels and boosts lipid metabolism after breakfast the next day: a randomized cross-over trial. Nutrients. 2021;13:2424. doi: 10.3390/nu13072424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Lopez-Minguez J, Saxena R, Bandín C, Scheer FA, Garaulet M. Late dinner impairs glucose tolerance in MTNR1B risk allele carriers: a randomized, cross-over study. Clin Nutr. 2018;37:1133–40. doi: 10.1016/j.clnu.2017.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Garaulet M, Gómez-Abellán P, Alburquerque-Béjar JJ, Lee Y-C, Ordovás JM, Scheer Fa. JL. Timing of food intake predicts weight loss effectiveness. Int J Obes. 2013;37:604–11. doi: 10.1038/ijo.2012.229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Wehrens SMT, Christou S, Isherwood C, Middleton B, Gibbs MA, Archer SN, et al. Meal timing regulates the human circadian system. Curr Biol. 2017;27:1768–75.e3. doi: 10.1016/j.cub.2017.04.059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.King AB, Clark D. Omitting late-night eating may cause hypoglycemia in ‘well controlled’ basal insulin-treated type 2 diabetes. Endocr Pract. 2015;21:280–5. doi: 10.4158/ep14334.or. [DOI] [PubMed] [Google Scholar]
- 72.Marinac CR, Quante M, Mariani S, Weng J, Redline S, Cespedes Feliciano EM, et al. Associations between timing of meals, physical activity, light exposure, and sleep with body mass index in free-living adults. J Phys Act Health. 2019;16:214–21. doi: 10.1123/jpah.2017-0389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Wu T, Sun L, ZhuGe F, Guo X, Zhao Z, Tang R, et al. Differential roles of breakfast and supper in rats of a daily three-meal schedule upon circadian regulation and physiology. Chronobiol Int. 2011;28:890–903. doi: 10.3109/07420528.2011.622599. [DOI] [PubMed] [Google Scholar]
- 74.Shoar S, Naderan M, Mahmoodzadeh H, Shoar N, Lotfi D. Night eating syndrome: a psychiatric disease, a sleep disorder, a delayed circadian eating rhythm, and/or a metabolic condition? Expert Rev Endocrinol Metab. 2019;14:351–8. doi: 10.1080/17446651.2019.1657006. [DOI] [PubMed] [Google Scholar]
- 75.Hood MM, Reutrakul S, Crowley SJ. Night eating in patients with type 2 diabetes. Associations with glycemic control, eating patterns, sleep, and mood. Appetite. 2014;79:91–6. doi: 10.1016/j.appet.2014.04.009. [DOI] [PubMed] [Google Scholar]
- 76.Deniz ÇD, Özler S, Sayın FK, Eryılmaz MA. Associations between night eating syndrome and metabolic parameters in pregnant women. Turk J Obstet Gynecol. 2019;16:107–11. doi: 10.4274/tjod.galenos.2019.77864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Gallant A, Drapeau V, Allison KC, Tremblay A, Lambert M, O’Loughlin J, et al. Night eating behavior and metabolic heath in mothers and fathers enrolled in the QUALITY cohort study. Eat Behav. 2014;15:186–91. doi: 10.1016/j.eatbeh.2014.01.002. [DOI] [PubMed] [Google Scholar]
- 78.Gallant A, Lundgren J, O’Loughlin J, Allison K, Tremblay A, Henderson M, et al. Night-eating symptoms and 2-year weight change in parents enrolled in the QUALITY cohort. Int J Obes. 2015;39:1161–5. doi: 10.1038/ijo.2015.36. [DOI] [PubMed] [Google Scholar]
- 79.Bermúdez-Millán A, Pérez-Escamilla R, Lampert R, Feinn R, Damio G, Segura-Pérez S, et al. Night eating among latinos with diabetes: exploring associations with heart rate variability, eating patterns, and sleep. J Nutr Educ Behav. 2022;54:449–54. doi: 10.1016/j.jneb.2022.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Yoshida J, Eguchi E, Nagaoka K, Ito T, Ogino K. Association of night eating habits with metabolic syndrome and its components: a longitudinal study. BMC Publ Health. 2018;18:1366. doi: 10.1186/s12889-018-6262-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Zhang X, Wu Y, Na M, Lichtenstein AH, Xing A, Chen S, et al. Habitual night eating was positively associated with progress of arterial stiffness in Chinese adults. J Am Heart Assoc. 2020;9:e016455. doi: 10.1161/JAHA.120.016455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Bruzas MB, Allison KC. A review of the relationship between night eating syndrome and body mass index. Curr Obes Rep. 2019;8:145–55. doi: 10.1007/s13679-019-00331-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Matejko B, Kiec-Wilk B, Szopa M, Trznadel Morawska I, Malecki MT, Klupa T. Are late-night eating habits and sleep duration associated with glycemic control in adult type 1 diabetes patients treated with insulin pumps? J Diabetes Investig. 2015;6:460–4. doi: 10.1111/jdi.12320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Kahleova H, Lloren JI, Mashchak A, Hill M, Fraser GE. Meal frequency and timing are associated with changes in body mass index in adventist health study 2. J Nutr. 2017;147:1722–8. doi: 10.3945/jn.116.244749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Kahleova H, Belinova L, Malinska H, Oliyarnyk O, Trnovska J, Skop V, et al. Eating two larger meals a day (breakfast and lunch) is more effective than six smaller meals in a reduced-energy regimen for patients with type 2 diabetes: a randomised crossover study. Diabetologia. 2014;57:1552–60. doi: 10.1007/s00125-014-3253-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Belinova L, Kahleova H, Malinska H, Topolcan O, Windrichova J, Oliyarnyk O, et al. The effect of meal frequency in a reduced-energy regimen on the gastrointestinal and appetite hormones in patients with type 2 diabetes: a randomised crossover study. PLoS One. 2017;12:e0174820. doi: 10.1371/journal.pone.0174820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Jakubowicz D, Landau Z, Tsameret S, Wainstein J, Raz I, Ahren B, et al. Reduction in glycated hemoglobin and daily insulin dose alongside circadian clock upregulation in patients with type 2 diabetes consuming a three-meal diet: a randomized clinical trial. Diabetes Care. 2019;42:2171–80. doi: 10.2337/dc19-1142. [DOI] [PubMed] [Google Scholar]
- 88.Kanaley JA, Heden TD, Liu Y, Fairchild TJ. Alteration of postprandial glucose and insulin concentrations with meal frequency and composition. Br J Nutr. 2014;112:1484–93. doi: 10.1017/s0007114514002128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Carlson O, Martin B, Stote KS, Golden E, Maudsley S, Najjar SS, et al. Impact of reduced meal frequency without caloric restriction on glucose regulation in healthy, normal-weight middle-aged men and women. Metabolism. 2007;56:1729–34. doi: 10.1016/j.metabol.2007.07.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Papakonstantinou E, Kontogianni MD, Mitrou P, Magriplis E, Vassiliadi D, Nomikos T, et al. Effects of 6 vs 3 eucaloric meal patterns on glycaemic control and satiety in people with impaired glucose tolerance or overt type 2 diabetes: a randomized trial. Diabetes Metab. 2018;44:226–34. doi: 10.1016/j.diabet.2018.03.008. [DOI] [PubMed] [Google Scholar]
- 91.Papakonstantinou E, Kechribari I, Mitrou P, Trakakis E, Vassiliadi D, Georgousopoulou E, et al. Effect of meal frequency on glucose and insulin levels in women with polycystic ovary syndrome: a randomised trial. Eur J Clin Nutr. 2016;70:588–94. doi: 10.1038/ejcn.2015.225. [DOI] [PubMed] [Google Scholar]
- 92.Abdollahi S, Kazemi A, de Souza RJ, Clark CCT, Soltani S. The effect of meal frequency on biochemical cardiometabolic factors: a systematic review and meta-analysis of randomized controlled trials. Clin Nutr. 2021;40:3170–81. doi: 10.1016/j.clnu.2020.12.038. [DOI] [PubMed] [Google Scholar]
- 93.Pot GK, Almoosawi S, Stephen AM. Meal irregularity and cardiometabolic consequences: results from observational and intervention studies. Proc Nutr Soc. 2016;75:475–86. doi: 10.1017/s0029665116000239. [DOI] [PubMed] [Google Scholar]
- 94.Zhao L, Teong XT, Liu K, Liu B, Melaku YA, Vincent A, et al. Eating architecture in adults at increased risk of type 2 diabetes: associations with body fat and glycaemic control. Br J Nutr. 2021:1–10. doi: 10.1017/S0007114521002944. [DOI] [PubMed] [Google Scholar]
- 95.Lehto R, Ray C, Lahti-Koski M, Roos E. Meal pattern and BMI in 9-11-year-old children in Finland. Public Health Nutr. 2011;14:1245–50. doi: 10.1017/s1368980010003034. [DOI] [PubMed] [Google Scholar]
- 96.Pot GK, Hardy R, Stephen AM. Irregularity of energy intake at meals: prospective associations with the metabolic syndrome in adults of the 1946 British birth cohort. Br J Nutr. 2016;115:315–23. doi: 10.1017/s0007114515004407. [DOI] [PubMed] [Google Scholar]
- 97.Pot GK, Hardy R, Stephen AM. Irregular consumption of energy intake in meals is associated with a higher cardiometabolic risk in adults of a British birth cohort. Int J Obes. 2014;38:1518–24. doi: 10.1038/ijo.2014.51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Wennberg M, Gustafsson PE, Wennberg P, Hammarström A. Irregular eating of meals in adolescence and the metabolic syndrome in adulthood: results from a 27-year prospective cohort. Public Health Nutr. 2016;19:667–73. doi: 10.1017/s1368980015001445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Farshchi HR, Taylor MA, Macdonald IA. Regular meal frequency creates more appropriate insulin sensitivity and lipid profiles compared with irregular meal frequency in healthy lean women. Eur J Clin Nutr. 2004;58:1071–7. doi: 10.1038/sj.ejcn.1601935. [DOI] [PubMed] [Google Scholar]
- 100.Farshchi HR, Taylor MA, Macdonald IA. Decreased thermic effect of food after an irregular compared with a regular meal pattern in healthy lean women. Int J Obes Relat Metab Disord. 2004;28:653–60. doi: 10.1038/sj.ijo.0802616. [DOI] [PubMed] [Google Scholar]
- 101.Farshchi HR, Taylor MA, Macdonald IA. Beneficial metabolic effects of regular meal frequency on dietary thermogenesis, insulin sensitivity, and fasting lipid profiles in healthy obese women. Am J Clin Nutr. 2005;81:16–24. doi: 10.1093/ajcn/81.1.16. [DOI] [PubMed] [Google Scholar]
- 102.Alhussain MH, Macdonald IA, Taylor MA. Irregular meal-pattern effects on energy expenditure, metabolism, and appetite regulation: a randomized controlled trial in healthy normal-weight women. Am J Clin Nutr. 2016;104:21–32. doi: 10.3945/ajcn.115.125401. [DOI] [PubMed] [Google Scholar]
- 103.Manoogian ENC, Chow LS, Taub PR, Laferrère B, Panda S. Time-restricted eating for the prevention and management of metabolic diseases. Endocr Rev. 2022;43:405–36. doi: 10.1210/endrev/bnab027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Świątkiewicz I, Woźniak A, Taub PR. Time-restricted eating and metabolic syndrome: current status and future perspectives. Nutrients. 2021;13:221. doi: 10.3390/nu13010221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Mattson MP, Longo VD, Harvie M. Impact of intermittent fasting on health and disease processes. Ageing Res Rev. 2017;39:46–58. doi: 10.1016/j.arr.2016.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.LeCheminant JD, Christenson E, Bailey BW, Tucker LA. Restricting night-time eating reduces daily energy intake in healthy young men: a short-term cross-over study. Br J Nutr. 2013;110:2108–13. doi: 10.1017/s0007114513001359. [DOI] [PubMed] [Google Scholar]
- 107.Gill S, Panda S. A smartphone app reveals erratic diurnal eating patterns in humans that can be modulated for health benefits. Cell Metab. 2015;22:789–98. doi: 10.1016/j.cmet.2015.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Moro T, Tinsley G, Bianco A, Marcolin G, Pacelli QF, Battaglia G, et al. Effects of eight weeks of time-restricted feeding (16/8) on basal metabolism, maximal strength, body composition, inflammation, and cardiovascular risk factors in resistance-trained males. J Transl Med. 2016;14:290. doi: 10.1186/s12967-016-1044-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Sutton EF, Beyl R, Early KS, Cefalu WT, Ravussin E, Peterson CM. Early time-restricted feeding improves insulin sensitivity, blood pressure, and oxidative stress even without weight loss in men with prediabetes. Cell Metab. 2018;27:1212–21.e3. doi: 10.1016/j.cmet.2018.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Antoni R, Robertson TM, Robertson MD, Johnston JD. A pilot feasibility study exploring the effects of a moderate time-restricted feeding intervention on energy intake, adiposity and metabolic physiology in free-living human subjects. J Food Sci. 2018;7:E22. [Google Scholar]
- 111.Gabel K, Hoddy KK, Haggerty N, Song J, Kroeger CM, Trepanowski JF, et al. Effects of 8-hour time restricted feeding on body weight and metabolic disease risk factors in obese adults: a pilot study. Nutr Healthy Aging. 2018;4:345–53. doi: 10.3233/nha-170036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Kesztyüs D, Cermak P, Gulich M, Kesztyüs T. Adherence to time-restricted feeding and impact on abdominal obesity in primary care patients: results of a pilot study in a pre-post design. Nutrients. 2019;11:E2854. doi: 10.3390/nu11122854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Anton SD, Lee SA, Donahoo WT, McLaren C, Manini T, Leeuwenburgh C, et al. The effects of time restricted feeding on overweight, older adults: a pilot study. Nutrients. 2019;11:E1500. doi: 10.3390/nu11071500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Chow LS, Manoogian ENC, Alvear A, Fleischer JG, Thor H, Dietsche K, et al. Time-restricted eating effects on body composition and metabolic measures in humans who are overweight: a feasibility study. Obesity. 2020;28:860–9. doi: 10.1002/oby.22756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Wilkinson MJ, Manoogian ENC, Zadourian A, Lo H, Fakhouri S, Shoghi A, et al. Ten-hour time-restricted eating reduces weight, blood pressure, and atherogenic lipids in patients with metabolic syndrome. Cell Metab. 2020;31:92–104.e5. doi: 10.1016/j.cmet.2019.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Park S-J, Yang J-W, Song Y-J. The effect of four weeks dietary intervention with 8-hour time-restricted eating on body composition and cardiometabolic risk factors in young adults. Nutrients. 2021;13:2164. doi: 10.3390/nu13072164. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Brady AJ, Langton HM, Mulligan M, Egan B. Effects of 8 wk of 16:8 time-restricted eating in male middle- and long-distance runners. Med Sci Sports Exerc. 2021;53:633–42. doi: 10.1249/mss.0000000000002488. [DOI] [PubMed] [Google Scholar]
- 118.Cienfuegos S, Gabel K, Kalam F, Ezpeleta M, Wiseman E, Pavlou V, et al. Effects of 4- and 6-h time-restricted feeding on weight and cardiometabolic health: a randomized controlled trial in adults with obesity. Cell Metab. 2020;32:366–78.e3. doi: 10.1016/j.cmet.2020.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.McAllister MJ, Pigg BL, Renteria LI, Waldman HS. Time-restricted feeding improves markers of cardiometabolic health in physically active college-age men: a 4-week randomized pre-post pilot study. Nutr Res. 2020;75:32–43. doi: 10.1016/j.nutres.2019.12.001. [DOI] [PubMed] [Google Scholar]
- 120.Peeke PM, Greenway FL, Billes SK, Zhang D, Fujioka K. Effect of time restricted eating on body weight and fasting glucose in participants with obesity: results of a randomized, controlled, virtual clinical trial. Nutr Diabetes. 2021;11:6. doi: 10.1038/s41387-021-00149-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Xie Z, Sun Y, Ye Y, Hu D, Zhang H, He Z, et al. Randomized controlled trial for time-restricted eating in healthy volunteers without obesity. Nat Commun. 2022;13:1003. doi: 10.1038/s41467-022-28662-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Jamshed H, Beyl RA, Della Manna DL, Yang ES, Ravussin E, Peterson CM. Early time-restricted feeding improves 24-hour glucose levels and affects markers of the circadian clock, aging, and autophagy in humans. Nutrients. 2019;11:1234. doi: 10.3390/nu11061234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Ravussin E, Beyl RA, Poggiogalle E, Hsia DS, Peterson CM. Early time-restricted feeding reduces appetite and increases fat oxidation but does not affect energy expenditure in humans. Obesity. 2019;27:1244–54. doi: 10.1002/oby.22518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Hutchison AT, Regmi P, Manoogian ENC, Fleischer JG, Wittert GA, Panda S, et al. Time-restricted feeding improves glucose tolerance in men at risk for type 2 diabetes: a randomized crossover trial. Obesity. 2019;27:724–32. doi: 10.1002/oby.22449. [DOI] [PubMed] [Google Scholar]
- 125.Parr EB, Devlin BL, Radford BE, Hawley JA. A delayed morning and earlier evening time-restricted feeding protocol for improving glycemic control and dietary adherence in men with overweight/obesity: a randomized controlled trial. Nutrients. 2020;12:E505. doi: 10.3390/nu12020505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Jamshed H, Steger FL, Bryan DR, Richman JS, Warriner AH, Hanick CJ, et al. Effectiveness of early time-restricted eating for weight loss, fat loss, and cardiometabolic health in adults with obesity: a randomized clinical trial. JAMA Intern Med. 2022;e223050 doi: 10.1001/jamainternmed.2022.3050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Tinsley GM, Forsse JS, Butler NK, Paoli A, Bane AA, La Bounty PM, et al. Time-restricted feeding in young men performing resistance training: a randomized controlled trial. Eur J Sport Sci. 2017;17:200–7. doi: 10.1080/17461391.2016.1223173. [DOI] [PubMed] [Google Scholar]
- 128.Gasmi M, Sellami M, Denham J, Padulo J, Kuvacic G, Selmi W, et al. Time-restricted feeding influences immune responses without compromising muscle performance in older men. Nutrition. 2018;51–2:29–37. doi: 10.1016/j.nut.2017.12.014. [DOI] [PubMed] [Google Scholar]
- 129.Tinsley GM, Moore ML, Graybeal AJ, Paoli A, Kim Y, Gonzales JU, et al. Time-restricted feeding plus resistance training in active females: a randomized trial. Am J Clin Nutr. 2019;110:628–40. doi: 10.1093/ajcn/nqz126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Martens CR, Rossman MJ, Mazzo MR, Jankowski LR, Nagy EE, Denman BA, et al. Short-term time-restricted feeding is safe and feasible in non-obese healthy midlife and older adults. Geroscience. 2020;42:667–86. doi: 10.1007/s11357-020-00156-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Parr EB, Devlin BL, Lim KHC, Moresi LNZ, Geils C, Brennan L, et al. Time-restricted eating as a nutrition strategy for individuals with type 2 diabetes: a feasibility study. Nutrients. 2020;12:E3228. doi: 10.3390/nu12113228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Phillips NE, Mareschal J, Schwab N, Manoogian ENC, Borloz S, Ostinelli G, et al. The effects of time-restricted eating versus standard dietary advice on weight, metabolic health and the consumption of processed food: a pragmatic randomised controlled trial in community-based adults. Nutrients. 2021;13:1042. doi: 10.3390/nu13031042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Liu D, Huang Y, Huang C, Yang S, Wei X, Zhang P, et al. Calorie restriction with or without time-restricted eating in weight loss. N Engl J Med. 2022;386:1495–504. doi: 10.1056/nejmoa2114833. [DOI] [PubMed] [Google Scholar]
- 134.Lowe DA, Wu N, Rohdin-Bibby L, Moore AH, Kelly N, Liu YE, et al. Effects of time-restricted eating on weight loss and other metabolic parameters in women and men with overweight and obesity: the TREAT randomized clinical trial. JAMA Intern Med. 2020;180:1491–9. doi: 10.1001/jamainternmed.2020.4153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Zhao L, Hutchison AT, Liu B, Yates CL, Teong XT, Wittert GA, et al. Time-restricted eating improves glycemic control and dampens energy-consuming pathways in human adipose tissue. Nutrition. 2022;96:111583. doi: 10.1016/j.nut.2021.111583. [DOI] [PubMed] [Google Scholar]
- 136.Quist JS, Jensen MM, Clemmensen KKB, Pedersen H, Bjerre N, Størling J, et al. Protocol for a single-centre, parallel-group, randomised, controlled, superiority trial on the effects of time-restricted eating on body weight, behaviour and metabolism in individuals at high risk of type 2 diabetes: the REStricted Eating Time (RESET) study. BMJ Open. 2020;10:e037166. doi: 10.1136/bmjopen-2020-037166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Manoogian ENC, Zadourian A, Lo HC, Gutierrez NR, Shoghi A, Rosander A, et al. Protocol for a randomised controlled trial on the feasibility and effects of 10-hour time-restricted eating on cardiometabolic disease risk among career firefighters doing 24-hour shift work: the Healthy Heroes Study. BMJ Open. 2021;11:e045537. doi: 10.1136/bmjopen-2020-045537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.Świątkiewicz I, Mila-Kierzenkowska C, Woźniak A, Szewczyk-Golec K, Nuszkiewicz J, Wróblewska J, et al. Pilot clinical trial of time-restricted eating in patients with metabolic syndrome. Nutrients. 2021;13:346. doi: 10.3390/nu13020346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Hatori M, Vollmers C, Zarrinpar A, DiTacchio L, Bushong EA, Gill S, et al. Time-restricted feeding without reducing caloric intake prevents metabolic diseases in mice fed a high-fat diet. Cell Metab. 2012;15:848–60. doi: 10.1016/j.cmet.2012.04.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Sherman H, Genzer Y, Cohen R, Chapnik N, Madar Z, Froy O. Timed high-fat diet resets circadian metabolism and prevents obesity. FASEB J. 2012;26:3493–502. doi: 10.1096/fj.12-208868. [DOI] [PubMed] [Google Scholar]
- 141.Chaix A, Zarrinpar A, Miu P, Panda S. Time-restricted feeding is a preventative and therapeutic intervention against diverse nutritional challenges. Cell Metab. 2014;20:991–1005. doi: 10.1016/j.cmet.2014.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Sundaram S, Yan L. Time-restricted feeding reduces adiposity in mice fed a high-fat diet. Nutr Res. 2016;36:603–11. doi: 10.1016/j.nutres.2016.02.005. [DOI] [PubMed] [Google Scholar]
- 143.Lee Y, Kim Y, Lee M, Wu D, Pae M. Time-restricted feeding restores obesity-induced alteration in adipose tissue immune cell phenotype. Nutrients. 2021;13:3780. doi: 10.3390/nu13113780. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Chaix A, Deota S, Bhardwaj R, Lin T, Panda S. Sex- and age-dependent outcomes of 9-hour time-restricted feeding of a Western high-fat high-sucrose diet in C57BL/6J mice. Cell Rep. 2021;36:109543. doi: 10.1016/j.celrep.2021.109543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Arble DM, Bass J, Laposky AD, Vitaterna MH, Turek FW. Circadian timing of food intake contributes to weight gain. Obesity. 2009;17:2100–2. doi: 10.1038/oby.2009.264. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Bray MS, Tsai J-Y, Villegas-Montoya C, Boland BB, Blasier Z, Egbejimi O, et al. Time-of-day-dependent dietary fat consumption influences multiple cardiometabolic syndrome parameters in mice. Int J Obes. 2010;34:1589–98. doi: 10.1038/ijo.2010.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 147.Yasumoto Y, Hashimoto C, Nakao R, Yamazaki H, Hiroyama H, Nemoto T, et al. Short-term feeding at the wrong time is sufficient to desynchronize peripheral clocks and induce obesity with hyperphagia, physical inactivity and metabolic disorders in mice. Metab Clin Exp. 2016;65:714–27. doi: 10.1016/j.metabol.2016.02.003. [DOI] [PubMed] [Google Scholar]
- 148.Opperhuizen A-L, Wang D, Foppen E, Jansen R, Boudzovitch-Surovtseva O, de Vries J, et al. Feeding during the resting phase causes profound changes in physiology and desynchronization between liver and muscle rhythms of rats. Eur J Neurosci. 2016;44:2795–806. doi: 10.1111/ejn.13377. [DOI] [PubMed] [Google Scholar]
- 149.de Goede P, Foppen E, Ritsema WIGR, Korpel NL, Yi C-X, Kalsbeek A. Time-restricted feeding improves glucose tolerance in rats, but only when in line with the circadian timing system. Front Endocrinol. 2019;10:554. doi: 10.3389/fendo.2019.00554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Acosta-Rodríguez V, Rijo-Ferreira F, Izumo M, Xu P, Wight-Carter M, Green CB, et al. Circadian alignment of early onset caloric restriction promotes longevity in male C57BL/6J mice. Science. 2022;376:1192–202. doi: 10.1126/science.abk0297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Solon-Biet SM, McMahon AC, Ballard JWO, Ruohonen K, Wu LE, Cogger VC, et al. The ratio of macronutrients, not caloric intake, dictates cardiometabolic health, aging, and longevity in ad libitum-fed mice. Cell Metab. 2014;19:418–30. doi: 10.1016/j.cmet.2014.02.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Liu X, Jin Z, Summers S, Derous D, Li M, Li B, et al. Calorie restriction and calorie dilution have different impacts on body fat, metabolism, behavior, and hypothalamic gene expression. Cell Rep. 2022;39:110835. doi: 10.1016/j.celrep.2022.110835. [DOI] [PubMed] [Google Scholar]
- 153.Salgado-Delgado R, Angeles-Castellanos M, Saderi N, Buijs RM, Escobar C. Food intake during the normal activity phase prevents obesity and circadian desynchrony in a rat model of night work. Endocrinology. 2010;151:1019–29. doi: 10.1210/en.2009-0864. [DOI] [PubMed] [Google Scholar]
- 154.Oike H, Sakurai M, Ippoushi K, Kobori M. Time-fixed feeding prevents obesity induced by chronic advances of light/dark cycles in mouse models of jet-lag/shift work. Biochem Biophys Res Commun. 2015;465:556–61. doi: 10.1016/j.bbrc.2015.08.059. [DOI] [PubMed] [Google Scholar]
- 155.Chaix A, Lin T, Le HD, Chang MW, Panda S. Time-restricted feeding prevents obesity and metabolic syndrome in mice lacking a circadian clock. Cell Metab. 2019;29:303–19.e4. doi: 10.1016/j.cmet.2018.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Kolbe I, Leinweber B, Brandenburger M, Oster H. Circadian clock network desynchrony promotes weight gain and alters glucose homeostasis in mice. Mol Metab. 2019;30:140–51. doi: 10.1016/j.molmet.2019.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]