

Abstract
Acyl carrier proteins (ACPs) are universally conserved proteins amongst different species and are involved in fatty acid synthesis. Bacteria utilize ACPs as acyl carriers and donors for the synthesis of products such as endotoxins or acyl homoserine lactones (AHLs), which are used in quorum sensing mechanisms. In this study, wehave expressed isotopically labeled holo-ACP from Burkholderia mallei in Escherichia coli to assign 100% of non-proline backbone amide (HN) resonances, 95.5% of aliphatic carbon resonances and 98.6% of aliphatic hydrogen sidechain resonances.
Keywords: Acyl carrier proteins, Quorum sensing, NMR assignment
Biological context
Quorum sensing in bacteria is defined as the ability to adapt gene expression as a function of cell-population density. Gram-negative bacteria count N-acyl-l-homoserine lactone (AHL) molecules to estimate population density. As cell density increases, chemical signal molecules produced by bacteria increase in concentration until they reach a threshold after which they alter gene expression. This process can be used to regulate physiological activities such as biofilm formation, sporulation, virulence, production of antibiotics, motility and many more.(Duerkop et al. 2009; Hammer et al., 2003; Smith et al., 2003) To establish efficiency in the counting process, AHL signals are made specifically by evolved AHL synthases to avoid recognition of non-native signals and ensure native substrate recognition for that specific bacterium. In this context, ACPs serve as intermediate cargo carriers which present acyl chains to AHLs and provide specificity through individual protein interfaces. (Agarwal et al. 2012; Lu et al. 2006; Mindrebo et al. 2020)
Bacterial ACPs are small, acidic four-helix bundle proteins. Helix II harbors most of the conserved residues, including a highly conserved ‘DSL’ motif (Zhang et al. 2003). The functional holo-ACP is formed when a 4′-phosphopantetheine moiety from coenzyme A (CoA) is attached to this serine (Chan et al., 2010). For production of AHL molecules during quorum sensing, acyl chains are attached to the free thiol of the phosphopantetheine moiety to be recognized by AHL-synthases (Montebello et al. 2014; Raychaudhuri et al. 2005).
The pathogen Burkholderia mallei, which has been shown to be involved in the development of Glanders disease utilizes the two AHL synthases BmaI1 and BmaI3 to recognize octanoyl-ACP (C8-ACP) and 3-hydroxyoctanoyl-ACP (3-hydroxy-C8-ACP), respectively (Duerkop et al. 2007, 2008; Mayerczyk et al., 2013; Withlock et al., 2007). In the case of BmaI1, for instance, the AHL synthesis is facilitated by the transfer of the acyl chain from octanoyl-ACP to the primary amine in S-adenosyl-methionine (SAM). Intramolecular lactonization releases the AHL molecule octanoyl homoserine lactone (C8-HSL) and 5’-deoxy-5’-(methylthio)adenosine (MTA). To achieve substrate specificity, BmaI1 recognizes a large interface on octanoyl-ACP that extends over helix II, loops 2 and 3 and helices III and IV. AHL synthases from other bacteria can achieve substrate specificity through other mechanisms, such as specific hydrogen bonding between functionalized acyl chains and AHL synthase in the case of EsaI from Pantoea stewartii (Fischer et al. 2023). These examples demonstrate that, despite the multitude of available ACP structures, the dynamics, and interactions of acylated ACPs still remain a largely unexplored field. In this work we report the assignments of holo-ACP and C8i-ACP from Burkholderia mallei. C8i-modification of ACP is realized by the attachment of a C8-carbon chain to the phosphopantetheine moiety via thioether linkage, to circumvent enzymatic reactions with AHL synthases in future experiments. As NMR represents an ideal tool to study these small, well-behaved proteins (i.e., well-expressing and stable over extensive periods of time) our report on the production, purification, modification, and assignment of ACPs will greatly aid towards future work in deciphering the mechanisms of bacterial quorum sensing signal biosynthesis.
Extent of assignments and data deposition
Figure 1A shows a superposition of the 1H-15N-HSQC spectra of holo-ACPbm and C8i-ACPbm. Both proteins are well structured, as the resonances are dispersed in the proton dimension. In the case of holo-ACPbm, following standard triple resonance experiments, 100% of hydrogen, 100% of nitrogen and 100% of carbon (C’, Cα) backbone resonances were assigned. Furthermore, 95.5% of sidechain carbons and 98.6% of sidechain hydrogens were assigned. Regarding C8i-ACPbm, 98.7% of backbone amide resonances were assigned. This was achieved by comparison with assigned spectra of holo-ACPbm and E.coli C8i-ACP (Fischer et al. 2023), using Cα backbone resonance in case of ambiguity. No sidechain assignment was carried out for this protein. Chemical shift perturbation (CSP) values were calculated between the HSQC peaks in the spectra of holo-ACPbm and C8i-ACPbm (Fig. 1B). CSP values greater than the mean value plus one standard deviation were classified as significant and are highlighted in two different hues of red. In Fig. 1C, the location of these changes is mapped onto a crystal structure of E. coli C7-ACP bound (PDB: 2FAD). Notably, for the residues within helix I, most CSPs remain below average, indicating a magnetic environment that is hardly affected by the binding of the C8i cargo. Similar observations are made for the outermost C-terminus. Indeed, helix I and the C-terminal portion of helix IV are not directly involved in the formation of the binding cleft. On the other hand, significant perturbations occur in widespread locations throughout the rest of the sequence. The hotspots lie within helices II-IV which form the cargo binding cleft and the adjacent loop I, connecting helices I and II. Surprisingly, the CSPs do not point out one distinct region being in close contact with the C8i cargo that is absent from the holo-ACP. These data suggest rather widespread structural alterations of the binding site upon cargo loading that remain to be deciphered in future studies. In conclusion, these assignments will provide a starting point for further NMR studies utilizing different isoforms of ACPs with and without cargo.
Fig. 1.
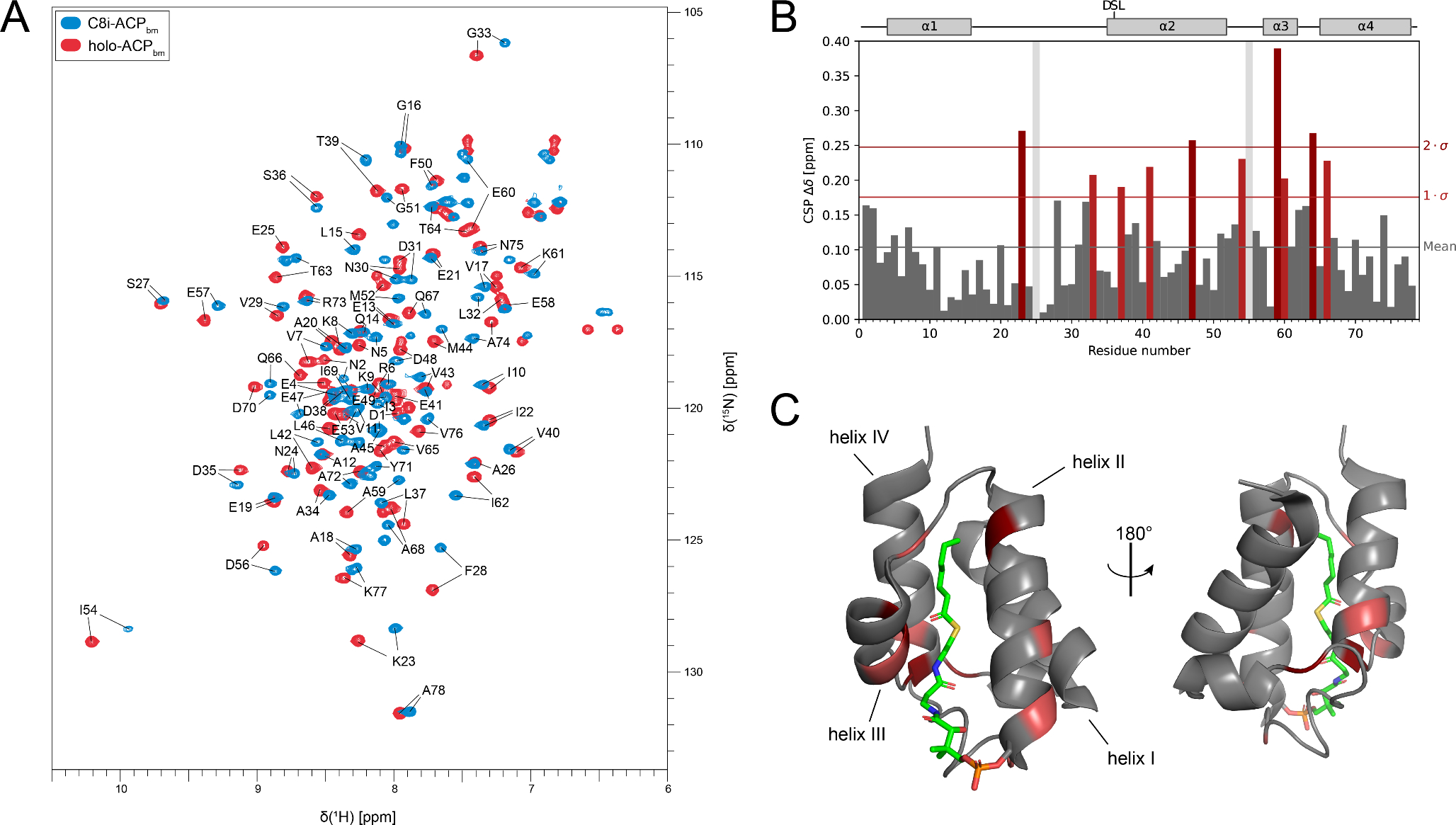
A: 15 N-HSQC spectra of C8i-ACPbm and holo-ACPbm including assignments recorded at pH = 6.8, 298 K, respectively. B: Chemical shift perturbation (CSP) bar plot for the two spectra in A. The mean CSP value is marked using a grey line, deviations by one and two standard deviations (σ) are marked using red and dark red lines. CSP greater than (mean + σ) are colored in red. C: Crystal structure of E. coli C7-ACP (PDB: 2FAD) colored by CSP values using the scheme from B
Secondary structure prediction
Using DANGLE (Cheung et al. 2010) a chemical shift and sequence query of holo-ACPbm was matched to a structural database of known PHI/PSI angles (Fig. 2). This predicted secondary structure data is consistent with the reported four helix bundle structures of ACPs.
Fig. 2.
Secondary structure prediction based on chemical shifts (DANGLE) of holo-ACP from Burkholderia mallei compared to deposited structures of acyl carrier proteins from other bacteria
Methods and experiments
Materials
Isotopes for protein labeling were purchased from Cambridge Isotope Laboratories, Inc. (Boston, MA).
Unless otherwise noted, all other reagents were purchased from Fisher Scientific (Hampton, NH).
Protein expression and purification
[U- 15 N, 13 C] ACPbm
The Burkholderia mallei Acyl-carrier protein (ACPbm) was expressed recombinantly in a pET vector containing an N-terminal HRV 3 C protease-cleavable His6 tag. An overnight culture of freshly transformed E. coli BL21 (DE3) was grown in 10 mL LB/Kan+ medium at 37 °C and subsequently used to inoculate 1 L M9 minimal medium containing 6 g/L Na2HPO4, 3 g/L KH2PO4, 0.5 g/L NaCl, 2 mM MgSO4, and 0.1 mM CaCl2 in H2O supplemented with vitamin mix, trace elements, 2 g/L 13 C-glucose, and 1 g/L 15NH4Cl. Cells were grown at 37 °C to an OD600 of 0.7. Next, protein expression was induced by adding 0.5 mM isopropyl β-D-1-thiogalactopyranoside (IPTG) and carried out at 20 °C for 18 h. The culture was harvested through centrifugation at 6,000 × g for 30 min and subsequently resuspended in 40 mL of lysis buffer (20 mM Tris-HCl pH = 8.0, 150 mM NaCl, 10 mM imidazole) supplemented with 2.5 mM β-mercaptoethanol, lysozyme, benzonase and protease inhibitor (cOmplete EDTA-free). Cells were disrupted by sonication (power level 6, 5 min, 1s on, 2 s off) and centrifuged at 37,000 × g for 45 min to remove cell debris. The supernatant was collected and used for nickel affinity chromatography. To this end, it was incubated with 5 mL (10 mL of a 50% slurry) of pre-equilibrated Ni-NTA resin for 1.5 h. The resin was washed with 50 mL of lysis buffer and subsequently eluted in an identical buffer containing 350 mM imidazole. The eluate was dialyzed over night against cleavage buffer (20 mM Tris-HCl pH = 8.0, 150 mM NaCl, 1 mM EDTA, 2 mM DTT). HRV 3 C protease was added to the eluate to cleave off the His6-tag. The dialysate was buffer exchanged and purified further using size exclusion chromatography on a HiLoad pg Superdex 75 16/600 column against NMR buffer (50 mM MES pH 6.8, 10 mM MgCl2, 1 mM TCEP). ACPbm-containing fractions were identified using SDS-PAGE and pooled. Eventually, the protein was concentrated to 1 mM for NMR spectroscopy. Mass spectroscopy and NMR analysis were used to identify the purified protein as holo-ACP.
[U- 15 N, 13 C] C8i-ACPbm
Octyl-ACP (C8i-ACPbm) was produced by first converting holo-ACPbm to apo-ACPbm. This was achieved by incubation of a solution of holo-ACPbm (~ 200 μM) with 5 μM ACPH in 50 mM Tris-HCl buffer (pH 8) in 15 mM MgCl2, 1 mM MnCl2, and 100 mM NaCl overnight (Fischer et al. 2023). Following conversion into apo-ACPbm, the protein was concentrated to ~ 500 μM in ACP NMR buffer (50 mM MES, pH = 6.8, 10 mM MgCl2, 1 mM TCEP-HCl) and combined with 1 μM purified Sfp and 1.25 equivalents (with respect to apo-ACP) of synthesized C8iCoA (C8-CoA in which the C8-alkyl chain is connected to phosphopantetheine via thioether linkage rather than thioester). The reaction mixture was allowed to incubate at 25 °C for 1 h and reaction completion was confirmed by 15 N-1 H-HSQC NMR spectra.
Intact Mass Spectrometry Analysis
Protein (5 μg) was injected onto a self-packed column (5 cm POROS 50R2, packed into 500 μm internal diameter fused silica), desalted for 4 min with solvent A (0.1 M acetic acid in water) and then eluted with an HPLC gradient (0–100% B in 1 min, B = 0.1 M acetic acid in acetonitrile) into an LTQ ion trap mass spectrometer (ThermoFisher Scientific, San Jose, CA). The mass spectrometer collected ESI (spray voltage = 5 kV) mass spectra from m/z 300–2000 in profile mode. Mass spectra were deconvoluted using MagTran version 1.03b2 (Zhang et al., 1998). Average 13 C, 15 N labeled protein mass was calculated using mzStudio (Ficarro et al. 2017). The calculated mass for holo-ACPbm, assuming 100% 13 C, 15 N incorporation, is 9689. The observed mass was 9685.2, confirming presence of holo-ACPbm.
NMR spectroscopy
Samples for NMR spectroscopy were either 1 mM holo-ACPbm or 0.5 mM C8-ACPbm uniformly 15 N and/or C labeled in NMR buffer (50 mM MES, pH = 6.8, 10 mM MgCl2, 1 mM TCEP-HCl), supplemented with 5% D2O for lock purposes. All spectra were acquired at 298 K on a Bruker AVANCE III NMR spectrometer equipped with a cryogenic TXO style probe, operating at 800 MHz proton frequency. Sequential assignments of backbone NH, Cα, Cβ and carbonyl resonances were carried out using standard set of HNCA, HNCO, HN(CA)CO, HN(CO)CA experiments from the Bruker pulse program library and a modified HNCACB experiment (N-C transfer time set to 6.9 ms to evolve Cβ only). Sidechain hydrogen and carbon assignments were carried out using H(CCO)NH and CC(CO)NH experiments from the Bruker pulse sequence library. In the H(CCO)NH experiment modifications were made to change the frequency to the aliphatic region during the indirect 1 H evolution period and jump back to the frequency of water (4.7 ppm), thus optimizing the spectral width. All 3D spectra were acquired using non-uniform sampling (NUS) with sampling 10% of the Nyquist grid using Poisson Gap Sampling (Hyberts et al. 2010). The NUS spectra were reconstructed and processed with hmsIST (Hyberts et al. 2012). NMR data was processed using NMRPipe (Delaglio et al. 1995) and analyzed using the CcpNmr Analysis software (version 2.4.1) (Vranken et al. 2005).
Funding
Financial support for this project came from NSF CHE #1905311 (RN), NIH R01GM136859 (HA), NIH INBRE grants P20 RR016454 and P20 GM103408 (NIGMS).
Footnotes
Competing interests The authors declare no competing interests or other interests that might be perceived to influence the results and/or discussion reported in this paper.
Ethics approval and consent to participate N/A.
Consent for publication Yes.
Data availability
The complete chemical shift assignments of Burkholderia mallei holo-ACP and backbone amide assignments of Burkholderia mallei C8i-ACP have been deposited at the Biological Magnetic Resonance Data Bank under the accession code 51,800 and 51,813, respectively.
References
- Agarwal V et al. (2012) Structure of the enzyme-acyl carrier protein (ACP) substrate gatekeeper complex required for biotin synthesis. Proc Natl Acad Sci U S A 109(43):17406–17411 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan DI, Vogel HJ (2010) Current understanding of fatty acid biosynthesis and the acyl carrier protein. Biochem J 430(1):1–19 [DOI] [PubMed] [Google Scholar]
- Cheung MS et al. (2010) DANGLE: a bayesian inferential method for predicting protein backbone dihedral angles and secondary structure. J Magn Reson 202(2):223–233 [DOI] [PubMed] [Google Scholar]
- Delaglio F et al. (1995) NMRPipe: a multidimensional spectral processing system based on UNIX pipes. J Biomol NMR 6(3):277–293 [DOI] [PubMed] [Google Scholar]
- Duerkop BA, Ulrich RL, Greenberg EP (2007) Octanoyl-homoserine lactone is the cognate signal for Burkholderia mallei BmaR1-BmaI1 quorum sensing. J Bacteriol 189(14):5034–5040 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duerkop BA et al. (2008) The Burkholderia mallei BmaR3-BmaI3 quorum-sensing system produces and responds to N-3-hydroxyoctanoyl homoserine lactone. J Bacteriol 190(14):5137–5141 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duerkop BA et al. (2009) Quorum-sensing control of antibiotic synthesis in Burkholderia thailandensis. J Bacteriol 191(12):3909–3918 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ficarro SB, Alexander WM, Marto JA (2017) mzStudio: a Dynamic Digital Canvas for user-driven interrogation of Mass Spectrometry Data.Proteomes, 5(3) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer PD et al. (2023) Carrier protein mediated cargo sensing in quorum signal synthases. Chem Commun (Camb) 59(8):1014–1017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hammer BK, Bassler BL (2003) Quorum sensing controls biofilm formation in Vibrio cholerae. Mol Microbiol 50(1):101–104 [DOI] [PubMed] [Google Scholar]
- Hyberts SG, Takeuchi K, Wagner G (2010) Poisson-gap sampling and forward maximum entropy reconstruction for enhancing the resolution and sensitivity of protein NMR data. J Am Chem Soc 132(7):2145–2147 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyberts SG et al. (2012) Application of iterative soft thresholding for fast reconstruction of NMR data non-uniformly sampled with multidimensional Poisson gap scheduling. J Biomol NMR 52(4):315–327 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lu YJ et al. (2006) Acyl-phosphates initiate membrane phospholipid synthesis in Gram-positive pathogens. Mol Cell 23(5):765–772 [DOI] [PubMed] [Google Scholar]
- Majerczyk C et al. (2013) Virulence of Burkholderia mallei quorum-sensing mutants. Infect Immun 81(5):1471–1478 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mindrebo J et al. (2020) 1.04-Structural basis of acyl-carrier protein interactions in fatty acid and Polyketide Biosynthesis. Comprehensive Natural Products III, p 61 [Google Scholar]
- Montebello AN et al. (2014) Acyl-ACP substrate recognition in Burkholderia mallei BmaI1 acyl-homoserine lactone synthase. Biochemistry 53(39):6231–6242 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raychaudhuri A, Jerga A, Tipton PA (2005) Chemical mechanism and substrate specificity of RhlI, an acylhomoserine lactone synthase from Pseudomonas aeruginosa. Biochemistry 44(8):2974–2981 [DOI] [PubMed] [Google Scholar]
- Smith RS, Iglewski BH (2003) P. aeruginosa quorum-sensing systems and virulence. Curr Opin Microbiol 6:56–60 [DOI] [PubMed] [Google Scholar]
- Vranken WF et al. (2005) The CCPN data model for NMR spectros-copy: development of a software pipeline. Proteins 59(4):687–696 [DOI] [PubMed] [Google Scholar]
- Whitlock GC, Estes DM, Torres AG (2007) Glanders: off to the races with Burkholderia mallei. FEMS Microbiol Lett 277(2):115–122 [DOI] [PubMed] [Google Scholar]
- Zhang Z, Marshall AG (1998) A universal algorithm for fast and automated charge state deconvolution of electrospray mass-to-charge ratio spectra. J Am Soc Mass Spectrom 9(3):225–233 [DOI] [PubMed] [Google Scholar]
- Zhang YM et al. (2003) The application of computational methods to explore the diversity and structure of bacterial fatty acid synthase. J Lipid Res 44(1):1–10 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The complete chemical shift assignments of Burkholderia mallei holo-ACP and backbone amide assignments of Burkholderia mallei C8i-ACP have been deposited at the Biological Magnetic Resonance Data Bank under the accession code 51,800 and 51,813, respectively.