

Abstract
Cyclic adenosine monophosphate (cAMP) and Ca2+ levels may oscillate in harmony within excitable cells; a mathematical oscillation loop model, the Cooper model, of these oscillations was developed two decades ago. However, in that model all adenylyl cyclase (AC) isoforms were assumed to be inhibited by Ca2+, and it is now known that the heart expresses multiple AC isoforms, among which the type 5/6 isoforms are Ca2+-inhibitable whereas the other five (AC2, 3, 4, 7, and 9) are not. We used a computational systems biology approach with CellDesigner simulation software to develop a comprehensive graphical map and oscillation loop model for cAMP and Ca2+. This model indicated that Ca2+-mediated inhibition of AC is essential to create oscillations of Ca2+ and cAMP, and the oscillations were not altered by incorporation of phosphodiesterase-mediated cAMP hydrolysis or PKA-mediated inhibition of AC into the model. More importantly, they were created but faded out immediately in the co-presence of Ca2+-noninhibitable AC isoforms. Because the subcellular locations of AC isoforms are different, spontaneous cAMP and Ca2+ oscillations may occur within microdomains containing only Ca2+-inhibitable isoforms in cardiac myocytes, which might be necessary for fine tuning of excitation–contraction coupling.
Electronic supplementary material
The online version of this article (doi:10.1007/s12576-014-0354-3) contains supplementary material, which is available to authorized users.
Keywords: Adenlyl cyclase, Subtype, Computational analysis, Oscillation, Cyclic AMP, Calcium
Introduction
β-Adrenergic receptor (β-AR) signaling is of crucial importance in regulating normal cardiac function, and abnormality of β-AR signaling contributes to the development of heart failure via altered cyclic AMP (cAMP) and calcium (Ca2+) signaling [1–3]. Cardiac excitation–contraction coupling (E–C coupling) is the process that links electrical excitation of cardiac myocytes to contraction of heart muscle. Ca2+ is essential for cardiac electrical activity and is a direct activator of myofilaments, causing both contraction and relaxation [4–6]. Dysregulation of cAMP and the subsequent Ca2+ oscillation are fundamental causes of both contractile and diastolic dysfunction, and arrhythmia among heart failure patients [1, 3, 5, 7–10].
Adenylyl cyclase (AC) is a membrane-bound enzyme that catalyzes the conversion of ATP to cAMP [1, 11]. cAMP, an intracellular second messenger, activates protein kinase A (PKA), leading to phosphorylation of multiple molecules involved in cardiac contraction, including the L-type Ca2+-channel [1]. Phosphorylation of the L-type Ca2+-channel is known to increase the influx of Ca2+, resulting in increased intracellular levels of Ca2+. In the 1990s, AC isoforms directly inhibited by Ca2+ were identified, i.e., types 5 and 6 AC isoforms (AC5 and AC6) [11–14]. Because these isoforms are dominantly expressed in the heart, it was proposed that cAMP levels may oscillate in harmony with Ca2+ levels; an increase in cAMP, as generated by AC5/6, phosphorylates L-type Ca2+ channels, and induces influx of Ca2+ into the cytosol [11]. An increase of cytosolic Ca2+ inhibits AC5/6 and reduces phosphorylation of the L-type Ca2+ channel. Thus, the activity of AC5/6 and the L-type Ca2+ channel may work synergistically to generate an oscillation loop of cAMP and Ca2+ in cardiac myocytes [11].
It is now well known that the heart expresses not only AC5 and AC6, but also many other AC isoforms (AC2, 3, 4, 5, 6, 7, and 9) [2]. Although these isoforms are all expressed in the heart, recent studies have indicated they may have different subcellular locations [15, 16]. AC5 is a major cardiac isoform in adults, and AC6 is a fetal or neonatal cardiac AC isoform [1, 17–19]. AC5 and AC6 share most, if not all, of their biochemical properties, and are inhibited not only by Ca2+, but also by Gi and PKA [2, 11, 13, 20–22]. In contrast, the other AC isoforms (AC2, 3, 4, 7, and 9), which are ubiquitously expressed throughout the body, are not inhibited by Ca2+, Gi, or PKA [13], and their involvement in cAMP and Ca2+ oscillations is poorly understood.
We therefore examined the involvement of AC isoforms in cAMP and Ca2+ oscillations [1, 12]. Because it is difficult to examine these issues by means of traditional in vitro or in vivo biochemical approaches, we used a computational systems biology approach with CellDesigner software, a recently developed, structure diagram editor for drawing gene-regulatory and biochemical networks [23–25].
Materials and methods
CellDesigner version 4.2 (http://www.celldesigner.org/) enables users to describe molecular interactions by using well-defined and consistent graphical notions and to create a comprehensive model incorporating positive feedforward or negative feedback loops among AC, cAMP, Ca2+, phosphodiesterase (PDE), and PKA within the β-AR signaling pathway [23–25].
The CellDesigner notation used in this paper is briefly illustrated for a simple reaction scheme in Supplemental Fig. 1. Protein A is transformed to protein B and protein C promotes this transition (Supplemental Fig. 1a). Supplemental Fig. 1b shows the notation for degradation (upper) or production (lower) of protein A; their balance determines the concentration of protein A in cells under physiological conditions.
In this study, most of the variables were the same as in Cooper’s original model [12]; in future work, it would be desirable to optimize the variables used in the oscillation loop model to match physiological conditions.
The formulas and values used to generate the oscillation models shown in the figures can be found in the online Supplemental methods and Supplemental Tables 1–3, available on http://www.link.springer.com/journal/12576.
Results
Cooper’s model mimicked by CellDesigner
We first mimicked Cooper’s original model by using CellDesigner to create a graphical comprehensive map (Fig. 1a) and oscillation loop model of cAMP and Ca2+ (Fig. 1b). We also incorporated the activity of PKA and AC. For AC, we used AC5/6 because they are the major cardiac isoforms and are directly inhibited by Ca2+ [2, 13]. We obtained stable and spontaneous oscillation curves for cAMP and Ca2+, as demonstrated in the original model [12]. The activity of AC and PKA also oscillated (Fig. 1b).
Fig. 1.
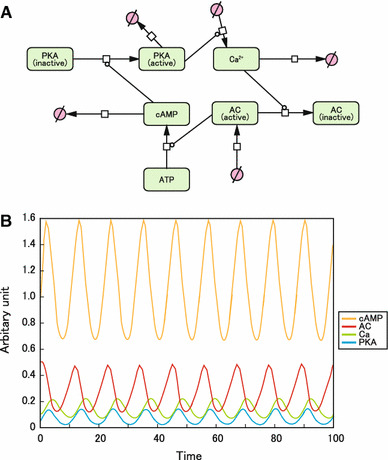
Cooper’s model mimicked by CellDesigner. a Graphical notations used in CellDesigner to depict Cooper’s model. b Computational oscillation loop of β-AR signaling molecules. cAMP, Ca2+, AC, and PKA formed a stable and persistent negative feedback loop. The intracellular concentration of cAMP at baseline (0 min) was taken as 1 (arbitrary units)
The formulas and values used to generate this oscillation model are shown in online Supplemental methods and Supplemental Table 1.
Incorporation of the PDE-mediated cAMP hydrolysis
Intracellular cAMP concentration is determined by the balance between its production via AC and its hydrolysis via PDE in the heart under physiological and pathological conditions [26]. Because PDE is activated by cAMP, we incorporated its activity into the model (Supplemental Fig. 2a). We found that oscillations of the molecules involved in the β-AR signaling pathway were maintained, even though their amplitudes were increased by approximately 1.6-fold, compared with those of Cooper’s original model (Supplemental Fig. 2b). Thus, PDE-mediated cAMP hydrolysis did not appear to change the behavior of the oscillation, but exaggerated its amplitude.
The formulas and values used to generate this oscillation model are shown in online Supplemental methods and Supplemental Table 2.
Effect of Ca2+-mediated inhibition of AC
The heart expresses multiple AC isoforms (AC2, 3, 4, 5, 6, 7, and 9) [2], of which AC5 and AC6 are directly inhibited by submicromolar Ca2+ [2, 13]. Thus, to examine the effect of Ca2+-mediated inhibition of AC, we modeled the situation in which all AC isoforms in the heart are not Ca2+-inhibitable in the heart (Fig. 2a). As shown in Fig. 2b, we found that no oscillation appeared. This result indicates that the presence of Ca2+-inhibitable AC isoforms is essential for stable and spontaneous cAMP and Ca2+ oscillations to occur.
Fig. 2.
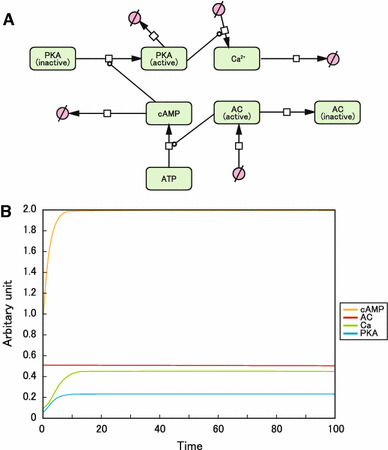
Effects of Ca2+-mediated inhibition of AC. a Graphical notation used in CellDesigner to depict β-AR signaling molecules AC, cAMP, Ca2+, and PKA. Ca2+-mediated inhibition of AC was deleted from Cooper’s original model. b Computational oscillation loop of β-AR signaling molecules. cAMP, Ca2+, AC, and PKA did not form a negative feedback loop. The intracellular concentration of cAMP at baseline (0 min) was taken as 1 (arbitrary units)
The formulas and values used to generate this oscillation model are shown in online supplemental methods and supplemental Table 1.
Incorporation of PKA-mediated inhibition of cardiac AC isoforms into the model
Recent studies have indicated that AC5 and AC6 are inhibited not only by Ca2+, but also by PKA [13, 20–22]. Therefore, we next incorporated PKA-mediated inhibition into Cooper’s model (Fig. 3a). Oscillations of cAMP and Ca2+, as well as AC and PKA, were observed, but the amplitudes were reduced by approximately 13 %, compared with those in Cooper’s original model (Fig. 3b). Thus, PKA-mediated inhibition of cardiac AC isoforms did not seem to change the behavior of the oscillation, but reduced its amplitude.
Fig. 3.
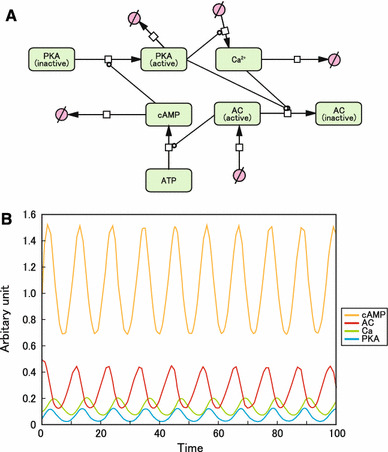
Effects of PKA-mediated inhibition of AC. a Graphical notation used in CellDesigner to depict β-AR signaling molecules AC, cAMP, Ca2+, and PKA. PKA-mediated inhibition of AC was incorporated into Cooper’s original model. b Computational oscillation loop of β-AR signaling molecules. cAMP, Ca2+, AC, and PKA formed a stable and spontaneous negative feedback loop. The intracellular concentration of cAMP at baseline (0 min) was taken as 1 (arbitrary units)
The formulas and values used to generate this oscillation model are shown in online Supplemental methods and Supplemental Table 1.
Incorporation of Ca2+-mediated inhibition of type 5/6 AC isoforms into the model
We then examined the model incorporating both Ca2+-inhibitable (AC5/6) and non-inhibitable AC isoforms (AC2, 3, 4, 7, and 9) (Fig. 4a). Studies with transgenic mouse models in vivo have demonstrated that AC2, 3, 4, 7, and 9 contribute significantly to the total AC activity in the heart [2, 27], but, unlike AC5/6, are not subjected to PKA-mediated inhibition [13]. Oscillations of cAMP, Ca2+, PKA, PDE, and AC5/6 were observed, and then faded. However, essentially, no oscillation of AC4/7 occurred (Fig. 4b). Thus, when both Ca2+-inhibitable and Ca2+-non-inhibitable AC isoforms coexist, continuous oscillation is not usually observed for cellular cAMP and Ca2+.
Fig. 4.
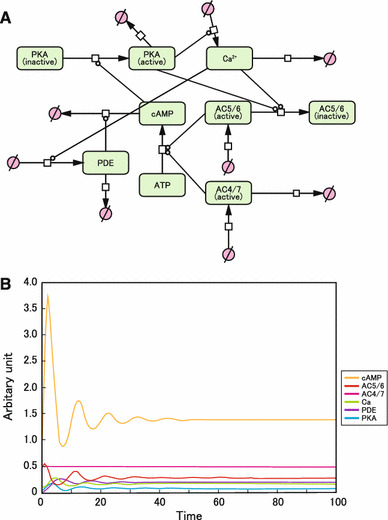
Incorporation of Ca2+-mediated inhibition of type 5/6 AC into the model. a Graphical notation used by CellDesigner to depict β-AR signaling molecules: cardiac AC subtypes (AC5/6), non-cardiac AC subtypes (AC4/7), cAMP, Ca2+, and PKA. A negative feedback loop was not observed. b Computational oscillation loop of β-AR signaling molecules: cAMP, Ca2+, AC5/6, PDE, and PKA did not form a continuous negative feedback loop, and then faded. However, essentially no oscillation occurred. The intracellular concentration of cAMP at baseline (0 min) was taken as 1 (arbitrary units)
The formulas and values used to generate this oscillation model are shown in online Supplemental methods and Supplemental Table 3.
Discussion
Control systems in vivo are dynamic and complex, and it is very difficult to predict systems behavior on the basis of biochemical studies of individual molecules. However, use of systems biology tools, for example CellDesigner, makes it feasible to simulate complex biochemical networks flexibly [28–30]. In this study, this software enabled us to study Ca2+ and cAMP oscillations under different conditions, e.g., in the presence or absence of Ca2+-inhibitable and Ca2+-non-inhibitable AC isoforms and other regulatory molecules, in silico, without the need for experimental assays [25].
First, we confirmed that the original oscillation model of cAMP and Ca2+ developed by Cooper in 1995 [12] can be effectively simulated by use of CellDesigner [23–25]. When the signaling pathway contained only Ca2+-inhibitable AC isoforms (AC5/6), we found that stable and spontaneous oscillations occurred.
Inclusion of PDE-mediated cAMP hydrolysis or PKA-mediated inhibition of AC5/6 into Cooper’s model did not seem to change the oscillation behavior, but altered the amplitude to a greater or lesser extent [13, 20–22, 26].
Inclusion of PKA-mediated inhibition of AC5/6 induced a decrease of the amplitude by approximately 13 % (Fig. 3), whereas inclusion of PDE-mediated cAMP hydrolysis increased the amplitude by approximately 1.6-fold (Supplemental Fig. 2), compared with those of Cooper’s original model (Fig. 1).
Phosphorylation of the L-type Ca2+-channel increases the Ca2+ concentration, and might form an ascending loop. In turn, cardiac AC isoforms (AC5/6) are inhibited by Ca2+ and this might form a descending loop. Importantly, AC5/6 are inhibited by PKA, which might reduce the amplitude of the oscillation loop [13, 22]. Conversely, PDE, in association with Ca2+-mediated inhibition, forms the descending phase of cAMP and Ca2+ oscillation in cardiac myocytes [31, 32]. These data, together with our current findings, indicate that PKA may have both positive and negative regulatory effects on the amplitude of the cAMP and Ca2+ oscillation loop, whereas PDE may have a positive regulatory effect on the amplitude [31, 32].
In contrast, when AC isoforms were not Ca2+-inhibitable, no oscillation occurred. Interestingly, when both Ca2+-non-inhibitable and Ca2+-inhibitable AC isoforms were included in the model, oscillation occurred, but decayed very rapidly. Inclusion of PDE-mediated cAMP hydrolysis did not change this behavior. Therefore, our simulations indicate that for stable and spontaneous oscillation, the presence of Ca2+-inhibitable AC isoforms and the absence of Ca2+-non-inhibitable AC isoforms are both required.
Further studies will be required to incorporate the effects of newly identified AC5/6-associated proteins, including Snapin, a SNAP25-binding protein, and PAM, a protein associated with Myc, on the cAMP and Ca2+ oscillations, because the findings of this study show that Ca2+-inhibitable AC isoforms (AC5/6) are essential for the oscillations of cAMP and Ca2+ [2, 13, 15].
Because the heart expresses seven AC isoforms [1, 7], including both Ca2+-inhibitable and Ca2+-non-inhibitable isoforms, continuous cAMP and Ca2+ oscillation in cardiac myocytes may not always occur, on the basis of the above findings (Fig. 4). Studies using AC5-deficient mice from our laboratory have shown that nearly half of the AC activity within the heart may be because of Ca2+-non-inhibitable AC isoforms [1, 2, 7, 9, 33, 34]. However, microenvironments in which only Ca2+-inhibitable AC5/6 are accumulated, such as lipid rafts or caveolae, are believed to exist [15, 16]. Indeed, it has been reported that Ca2+-sensitive AC isoforms (AC1, 5, 6, and 8) and their associated proteins, such as PKA, A-kinase anchoring proteins (AKAPs), anchored PDEs, non-anchored PDE, and transient receptor potential (TRP) 1/3, are present in lipid rafts in many cell types, including cardiac myocytes, whereas the Ca2+-insensitive AC 2, 4, and 7 are excluded from the rafts [15]. Because intracellular cAMP and Ca2+ mediate a diverse array of cellular functions, oscillation of cAMP and Ca2+ concentration might be involved in receptor-mediated signal transduction, not only in excitable cells, for example cardiac myocytes, but also in non-excitable cells [35]. Further, the occurrence or disturbance of cAMP and Ca2+ oscillations might contribute to the development of cardiac dysfunction or arrhythmia.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgments
This study was supported in part by grants from the Ministry of Health, Labor, and Welfare (Dr Ishikawa), a Grant-in-Aid for Scientific Research on Innovative Areas (22136009), and Grants from the Japanese Ministry of Education, Culture, Sports, Science, and Technology (Drs Ishikawa, Okumura, Fujita, Sato, Yokoyama, Mototani), the National Cerebral and Cardiovascular Center (Dr Ishikawa), the Takeda Science Foundation (Dr Okumura), the Yokohama Foundation for Advancement of Medical Science (Dr Okumura), the Yokohama Academic Foundation (Dr Ohnuki), the Research Foundation for Community Medicine (Dr Okumura), and the Suzuken Memorial Foundation (Dr Okumura).
Conflict of interest
The authors declare no potential conflicts of interest.
Contributor Information
Satoshi Okumura, Phone: 81-45-580-8476, Email: okumura-s@tsuruni-u.ac.jp.
Yoshihiro Ishikawa, Phone: 81-45-787-2575, Email: yishikaw@yokohama-cu.ac.jp.
References
- 1.Okumura S, Suzuki S, Ishikawa Y. New aspects for the treatment of cardiac diseases based on the diversity of functional controls on cardiac muscles: effects of targeted disruption of the type 5 adenylyl cyclase gene. J Pharmacol Sci. 2009;109:354–359. doi: 10.1254/jphs.08R26FM. [DOI] [PubMed] [Google Scholar]
- 2.Okumura S, Kawabe J, Yatani A, Takagi G, Lee MC, Hong C, Liu J, Takagi I, Sadoshima J, Vatner DE, Vatner SF, Ishikawa Y. Type 5 adenylyl cyclase disruption alters not only sympathetic but also parasympathetic and calcium-mediated cardiac regulation. Circ Res. 2003;93:364–371. doi: 10.1161/01.RES.0000086986.35568.63. [DOI] [PubMed] [Google Scholar]
- 3.Okumura S, Fujita T, Cai W, Jin M, Namekata I, Mototani Y, Jin H, Ohnuki Y, Tsuneoka Y, Bai Y, Suzuki S, Hidaka Y, Umemura M, Ichikawa Y, Yokoyama U, Sato M, Ishikawa F, Izumi-Nakaseko H, Adachi-Akahane S, Tanaka H, Ishikawa Y. EPAC1-dependent phospholamban phosphorylation mediates the cardiac response to stresses. J Clin Invest. 2014;124:2785–2801. doi: 10.1172/JCI64784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ohnuki Y, Nishimura S, Sugiura S, Saeki Y. Phosphorylation status of regulatory proteins and functional characteristics in myocardium of dilated cardiomyopathy of Syrian hamsters. J Physiol Sci. 2008;58:15–20. doi: 10.2170/physiolsci.RP012807. [DOI] [PubMed] [Google Scholar]
- 5.Bers DM. Calcium and cardiac rhythms: physiological and pathophysiological. Circ Res. 2002;90:14–17. [PubMed] [Google Scholar]
- 6.Ohnuki Y, Yamada T, Mototani Y, Umeki D, Shiozawa K, Fujita T, Saeki Y, Okumura S. Effects of protein kinase a on the phosphorylation status and transverse stiffness of cardiac myofibrils. J Pharmacol Sci. 2013;123:279–283. doi: 10.1254/jphs.13110SC. [DOI] [PubMed] [Google Scholar]
- 7.Okumura S, Takagi G, Kawabe J, Yang G, Lee MC, Hong C, Liu J, Vatner DE, Sadoshima J, Vatner SF, Ishikawa Y. Disruption of type 5 adenylyl cyclase gene preserves cardiac function against pressure overload. Proc Natl Acad Sci USA. 2003;100:9986–9990. doi: 10.1073/pnas.1733772100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Marks AR. Calcium cycling proteins and heart failure: mechanisms and therapeutics. J Clin Invest. 2013;123:46–52. doi: 10.1172/JCI62834. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Okumura S, Vatner DE, Kurotani R, Bai Y, Gao S, Yuan Z, Iwatsubo K, Ulucan C, Kawabe J, Gjosh K, Vatner SF, Ishikawa Y. Disruption of type 5 adenylyl cyclase enhances desensitization of cyclic adenosine monophosphate signal and increases Akt signal with chronic catecholamine stress. Circulation. 2007;116:1776–1783. doi: 10.1161/CIRCULATIONAHA.107.698662. [DOI] [PubMed] [Google Scholar]
- 10.Ohnuki Y, Umeki D, Mototani Y, Jin H, Cai W, Shiozawa K, Suita K, Fujita T, Ishikawa Y, Okumura S. Role of cyclic AMP sensor epac1 in masseter muscle hypertrophy and myosin heavy chain transition induced by β2-adrenoceptor stimulation. J Physiol. 2014;592:5461–5475. doi: 10.1113/jphysiol.2014.282996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ishikawa Y, Homcy CJ. The adenylyl cyclases as integrators of transmembrane signal transduction. Circ Res. 1997;80:297–304. doi: 10.1161/01.RES.80.3.297. [DOI] [PubMed] [Google Scholar]
- 12.Cooper DM, Mons N, Karpen JW. Adenylyl cyclases and the interaction between calcium and cAMP signalling. Nature. 1995;374:421–424. doi: 10.1038/374421a0. [DOI] [PubMed] [Google Scholar]
- 13.Hanoune J, Defer N. Regulation and role of adenylyl cyclase isoforms. Annu Rev Pharmacol Toxicol. 2001;41:145–174. doi: 10.1146/annurev.pharmtox.41.1.145. [DOI] [PubMed] [Google Scholar]
- 14.Katsushika S, Chen L, Kawabe J, Nilakantan R, Halnon NJ, Homcy C, Ishikawa Y. Cloning and characterization of a sixth adenylyl cyclase isoform: types V and VI constitute a subgroup within the mammalian adenylyl cyclase family. Proc Natl Acad Sci USA. 1992;89:8774–8778. doi: 10.1073/pnas.89.18.8774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Willoughby D, Cooper DM. Organization and Ca2+ regulation of adenylyl cyclases in cAMP microdomains. Physiol Rev. 2007;87:965–1010. doi: 10.1152/physrev.00049.2006. [DOI] [PubMed] [Google Scholar]
- 16.Timofeyev V, Myers RE, Kim HJ, Woltz RL, Sirish P, Heiserman JP, Li N, Singapuri A, Tang T, Yarov-Yarovoy V, Yamoah EN, Hammond HK, Chiamvimonvat N. Adenylyl cyclase subtype-specific compartmentalization: differential regulation of L-type Ca2+ current in ventricular myocytes. Circ Res. 2013;112:1567–1576. doi: 10.1161/CIRCRESAHA.112.300370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ishikawa Y, Katsushika S, Chen L, Halnon NJ, Kawabe J, Homcy CJ. Isolation and characterization of a novel cardiac adenylylcyclase cDNA. J Biol Chem. 1992;267:13553–13557. [PubMed] [Google Scholar]
- 18.Espinasse I, Iourgenko V, Defer N, Samson F, Hanoune J, Mercadier JJ. Type V, but not type VI, adenylyl cyclase mRNA accumulates in the rat heart during ontogenic development. Correlation with increased global adenylyl cyclase activity. J Mol Cell Cardiol. 1995;27:1789–1795. doi: 10.1016/0022-2828(95)90002-0. [DOI] [PubMed] [Google Scholar]
- 19.Tobise K, Ishikawa Y, Holmer SR, Im MJ, Newell JB, Yoshie H, Fujita M, Susannie EE, Homcy CJ. Changes in type VI adenylyl cyclase isoform expression correlate with a decreased capacity for cAMP generation in the aging ventricle. Circ Res. 1994;74:596–603. doi: 10.1161/01.RES.74.4.596. [DOI] [PubMed] [Google Scholar]
- 20.Premont RT, Jacobowitz O, Iyengar R. Lowered responsiveness of the catalyst of adenylyl cyclase to stimulation by GS in heterologous desensitization: a role for adenosine 3′,5′-monophosphate-dependent phosphorylation. Endocrinology. 1992;131:2774–2784. doi: 10.1210/endo.131.6.1332848. [DOI] [PubMed] [Google Scholar]
- 21.Chen Y, Harry A, Li J, Smit MJ, Bai X, Magnusson R, Pieroni JP, Weng G, Iyengar R. Adenylyl cyclase 6 is selectively regulated by protein kinase A phosphorylation in a region involved in Galphas stimulation. Proc Natl Acad Sci USA. 1997;94:14100–14104. doi: 10.1073/pnas.94.25.14100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Iwami G, Kawabe J, Ebina T, Cannon PJ, Homcy CJ. Regulation of adenylyl cyclase by protein kinase A. J Biol Chem. 1995;270:12481–12484. doi: 10.1074/jbc.270.21.12481. [DOI] [PubMed] [Google Scholar]
- 23.Kitano H, Funahashi A, Matsuoka Y, Oda K. Using process diagrams for the graphical representation of biological networks. Nat Biotechnol. 2005;23:961–966. doi: 10.1038/nbt1111. [DOI] [PubMed] [Google Scholar]
- 24.Funahashi A, Jouraku A, Matsuoka Y, Kitano H. Integration of Cell Designer and SABIO-RK. In Silico Biol. 2007;7:S81–S90. [PubMed] [Google Scholar]
- 25.Matsuoka Y, Funahashi A, Ghosh S, Kitano H. Modeling and simulation using Cell Designer. Methods Mol Biol. 2014;1164:121–145. doi: 10.1007/978-1-4939-0805-9_11. [DOI] [PubMed] [Google Scholar]
- 26.Sato N, Asai K, Okumura S, Takagi G, Shannon RP, Fujita-Yamaguchi Y, Ishikawa Y, Vatner SF, Vatner DE, et al. Mechanisms of desensitization to a PDE inhibitor (milrinone) in conscious dogs with heart failure. Am J Physiol . 1999;276:H1699–H1705. doi: 10.1152/ajpheart.1999.276.5.H1699. [DOI] [PubMed] [Google Scholar]
- 27.Tang T, Gao MH, Lai NC, Firth AL, Takahashi T, et al. Adenylyl cyclase type 6 deletion decreases left ventricular function via impaired calcium handling. Circulation. 2008;117:61–69. doi: 10.1161/CIRCULATIONAHA.107.730069. [DOI] [PubMed] [Google Scholar]
- 28.Autiero I, Costantini S, Colonna G. Modeling of the bacterial mechanism of methicillin-resistance by a systems biology approach. PLoS One. 2009;4:e6226. doi: 10.1371/journal.pone.0006226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Grieco L, Calzone L, Bernard-Pierrot I, Radvanyi F, Kahn-Perles B, Thieffy D. Integrative modelling of the influence of MAPK network on cancer cell fate decision. PLoS Comput Biol. 2013;9:e1003286. doi: 10.1371/journal.pcbi.1003286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wu G, Zhu L, Dent JE, Nardini C. A comprehensive molecular interaction map for rheumatoid arthritis. PLoS One. 2010;5:e10137. doi: 10.1371/journal.pone.0010137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mika D, Richter W, Westenbroek RE, Catterall WA, Conti M. PDE4B mediates local feedback regulation of β1-adrenergic cAMP signaling in a sarcolemmal compartment of cardiac myocytes. J Cell Sci. 2014;127:1033–1042. doi: 10.1242/jcs.140251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Masaki N, Fujimoto K, Honda-Kitahara M, Hada E, Sawai S. Robustness of self-organizing chemoattractant field arising from precise pulse induction of its breakdown enzyme: a single-cell level analysis of PDE expression in Dictyostelium. Biophys J. 2013;104:1191–1202. doi: 10.1016/j.bpj.2013.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Okumura S, Tsunematsu T, Bai Y, Jiao Q, Ono S, Suzuki S, Kurotani R, Sato M, Minamisawa S, Umemura S, Ishikawa Y. Type 5 adenylyl cyclase plays a major role in stabilizing heart rate in response to microgravity induced by parabolic flight. J Appl Physiol 1985. 2008;105:173–179. doi: 10.1152/japplphysiol.01166.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bai Y, Tsunematsu T, Jiao Q, Ohnuki Y, Mototani Y, Shiozawa K, Jin M, Cai W, Jin HL, Fujita T, Ichikawa Y, Suita K, Kurotani R, Yokoyama U, Sato M, Iwatsubo K, Isikawa Y. Pharmacological stimulation of type 5 adenylyl cyclase stabilizes heart rate under both microgravity and hypergravity induced by parabolic flight. J Pharmacol Sci. 2012;119:381–389. doi: 10.1254/jphs.12102FP. [DOI] [PubMed] [Google Scholar]
- 35.Dolmetsch RE, Xu K, Lewis RS. Calcium oscillations increase the efficiency and specificity of gene expression. Nature. 1998;392:933–936. doi: 10.1038/31960. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.