

Abstract
Multidrug antibiotic resistance is an urgent public health concern. Multiple strategies have been suggested to alleviate this problem, including the use of antibiotic combinations and cyclic therapies. We examine how adaptation to (1) combinations of drugs affects resistance to individual drugs, and to (2) individual drugs alters responses to drug combinations. To evaluate this, we evolved multiple strains of drug resistant Staphylococcus epidermidis in the lab. We show that evolving resistance to four highly synergistic combinations does not result in cross‐resistance to all of their components. Likewise, prior resistance to one antibiotic in a combination does not guarantee survival when exposed to the combination. We also identify four 3‐step and four 2‐step treatments that inhibit bacterial growth and confer collateral sensitivity with each step, impeding the development of multidrug resistance. This study highlights the importance of considering higher‐order drug combinations in sequential therapies and how antibiotic interactions can influence the evolutionary trajectory of bacterial populations.
Keywords: antibiotic combinations, antibiotic resistance, antimicrobial, bacterial evolution
1. INTRODUCTION
Antibiotic resistance is a problem facing humanity on a global scale (Ahmad & Khan, 2019; Coates et al., 2020; Hernando‐Amado et al., 2019; Maillard et al., 2020; Ventola, 2015). Not only is drug resistance to a single drug a threat to public health, but even more concerning, multidrug resistance in bacteria is reducing the number of viable treatment options. Some estimates show that by 2050, there will no longer be any effective antibiotic options unless new drugs are developed or discovered (Jin et al., 2020). Multiple strategies have been suggested to mitigate this ever‐growing problem, including cycling through antibiotic treatments (Nichol et al., 2015) and the use of combinational drug therapy (e.g., using multiple antibiotics simultaneously or combining antibiotics with alternative therapies, such as phage therapy) (Vivas et al., 2019). In designing new therapies, major considerations include whether or not to use two or more antibiotics at the same time, in succession, or both, as well as how much of each drug to use. We are especially interested in determining whether particular strategies (out of a world of possibilities) can reduce the emergence of antibiotic resistance. This requires investigating if and how adaptation to individual antibiotics differs from adaptation to drug combinations, and what patterns of resistance this confers on evolved populations.
Resistance of bacteria to single antibiotics has long been a topic of study. Typically, for a single antibiotic, resistance is quantified by the minimum inhibitory concentration (MIC). The MIC is when the concentration of an antibiotic that reduces growth to 0%–10% when compared to growing in an antibiotic free‐environment, depending on the specific method used when evaluating growth (Eagle & Musselman, 1948; Garrod, 1935; Haight & Finland, 1952; Thomson & Sanders, 1994). However, when multiple antibiotics with multiple mechanisms of action are used in combination, there are added complexities and nonlinearities (Beppler et al., 2016, 2017; Loewe, 1953; Tekin et al., 2016). Now, the cell is no longer facing one specific mechanism to evolve resistance to but rather multiple mechanisms, which may require multiple mutations that confer resistance.
Antibiotic resistance is acquired in two primary ways: through spontaneous mutation or through horizontal gene transfer (Blair et al., 2015). No matter how resistance evolves initially, the subsequent mutations can affect the outcomes of future antibiotic exposures (Barbosa et al., 2017; Gomez et al., 2017; Munck et al., 2014; Nichol et al., 2015, 2019; Santos‐Lopez et al., 2019). If specific mutations gained from adapting to one antibiotic confer a higher resistance to a different antibiotic, then a bacterial strain has gained cross‐resistance (Haight & Finland, 1952; Obolski et al., 2015; Sanders, 2001). If these mutations instead reduce a strain's resistance to a different antibiotic, then it is termed collateral sensitivity (Obolski et al., 2015; Pál et al., 2015). Exploiting these relationships to find combinations of antibiotics to cycle through different treatment regimen can be one potential strategy to address the antibiotic resistance problem (Brown & Nathwani, 2005; Kavanaugh et al., 2021; Kollef, 2001; Masterton, 2005; Nichol et al., 2015, 2019). This is because one goal of the cycle is always to have at least one viable antibiotic regimen available, thus locking the bacterial population in a constant state of susceptibility to current drug options.
Another proposed method for addressing multidrug antibiotic resistance is through the use of simultaneous antibiotic combinations (Vivas et al., 2019). Antibiotic combinations can be categorized by three main types of interactions resulting from their combined effect. (1) An additive combination's effects are directly predictable from combining the known, individual effects of the drugs employed. (2) A synergistic combination yields an effect that is stronger than the additive effect (Yeh et al., 2009). Synergistic combinations impose stronger selection, making it more likely that resistance mutations will sweep quickly through a population (Orr, 2000; Pepin & Wichman, 2008), and can cause faster adaptation rates (Hegreness et al., 2008). (3) Finally, an antagonistic combination yields an effect weaker than the additive effect. These antagonistic combinations require larger doses to have the same killing efficiency as synergistic combinations (Chait et al., 2007; Hegreness et al., 2008; Michel et al., 2008; Yeh et al., 2009).
The different ways in which antibiotics interact may have distinct implications for the evolution of resistance. Intriguingly, antagonistic combinations have been shown to have several advantages for slowing the evolution of resistant strains (Hegreness et al., 2008; Maillard et al., 2020; Michel et al., 2008; Ventola, 2015). Synergistic two‐drug combinations have been shown to increase the likelihood of drug resistance evolving via spontaneous mutations (Michel et al., 2008). Conversely, antagonistic combinations show a decrease in the likelihood of resistance evolving (Michel et al., 2008). Additionally, Hegreness et al. (2008) showed that antagonistic two‐drug combinations slowed the rate of resistance evolution when compared to the rate of resistance evolution to two‐drug synergistic combinations. These findings highlight how antibiotic interactions can be leveraged to slow the evolution of resistance to multidrug antibiotic combinations. Most studies have focused on two‐drug combinations, creating a knowledge gap around higher‐order combinations, that is, combinations with three or more drugs (Tekin et al., 2017; Tekin, Yeh, et al., 2018).
In multidrug combinations, many factors contribute to the overall fitness effect of the combination. The first factors to consider are the effects of each drug acting alone. Next, we must consider the effects of the additive interactions between the smaller sub‐sets of antibiotics and the other single antibiotics (or combination of antibiotics) in the mix. Finally, we need to examine the highest‐order emergent effect—that is, the effect of the interaction between all drugs present in the combination. All these factors influence how a combination will ultimately affect a bacterial population.
Numerous studies have examined the evolutionary effects of cross‐resistance and collateral sensitivity among single antibiotics in multiple species (Barbosa et al., 2017; Gomez et al., 2017; Munck et al., 2014; Nichol et al., 2015). The trajectory of resistance evolution can be influenced using knowledge of collateral effects to create a specific sequence or cycle of antibiotics to steer it away from resistance evolution (Nichol et al., 2015). In addition, some mutations, such as those affecting the ribosome, increase the evolution of multidrug resistance (Gomez et al., 2017). Yet other studies have questioned just how predictable are the collateral effects of resistance evolution. These studies have found high amounts of stochasticity and variability of these collateral effects in replicate populations (Barbosa et al., 2017; Nichol et al., 2019). Studies have also expanded into evaluating how pairwise combinations align with cross‐resistance and collateral sensitivity (Munck et al., 2014; Raymond, 2019). These studies found that bacterial populations exposed to collaterally sensitive antibiotic combinations are less likely to evolve resistance to the combination. Conversely, antibiotic pairs with similar or the same cellular/physiological targets that result in cross‐resistance tend to increase the likelihood of evolution of resistance to the combination (Munck et al., 2014).
When antibiotics are used in combinations, they are typically used at concentrations near or above the MIC (Kleine et al., 2017; Martin‐Loeches et al., 2010; Paul et al., 2014). It was previously thought that weakening the selection pressure using lower antibiotic concentrations might curb the rapid evolution of resistance. However, for some antibiotic combinations, the opposite is true. Short‐term higher doses are more effective at preventing resistance evolution compared to weaker doses (Bollenbach, 2015). When exposed to one antibiotic, at sub‐inhibitory concentrations, an SOS response system is induced in bacteria. This SOS response allows for damaged DNA to be bypassed by the cell which in turn allows for mutation rates to increase (Chow et al., 2021). Thus, the effect of combinations could also be dependent on the dosages.
Our study focuses on Staphylococcus epidermidis, a gram‐positive bacterium that colonizes skin and mucosa. S. epidermidis was previously considered an innocuous commensal microorganism on the human skin. However, it has recently become an opportunistic pathogen that results in nosocomial infections, particularly in indwelling medical devices such as catheters (Otto, 2009). We investigate the effects of resistance evolution to a single antibiotic and combinations of antibiotic used simultaneously. We examine four highly synergistic three‐drug combinations where all antibiotics are at sub‐inhibitory concentrations (<30% inhibition). These combinations are made up of piperacillin and tetracycline, with a third antibiotic of either: chloramphenicol, doxycycline, erythromycin, or neomycin. Specifically, we address the following questions: (1) How does evolved resistance to a three‐drug combination affect subsequent sensitivity to individual components of the combination? (2) Conversely, how does evolved resistance to individual components of a three‐drug combination affect sensitivity to the combination? (3) How can the interactions within a three‐drug combination change after bacteria evolve resistance to one part of the antibiotic combination?
2. MATERIALS AND METHODS
2.1. Creation and isolation of resistant mutants
Eight strains of resistant S. epidermidis (ATCC 14990) were independently evolved in a stepwise manner to each of six antibiotics (Table 1) and each of four focal three‐drug combinations known to be highly synergistic (Table 2). From here onward, individual antibiotics will be spelled out while combinations of antibiotics will be listed using their abbreviations (Table 1). For example, a combination consisting of piperacillin, tetracycline, and erythromycin will be listed as PIP + TET + ERY. We use the term drug regimen to be a single antibiotic or combination of antibiotics used simultaneously to inhibit bacterial growth and the term treatment to encompass a set of drug regimens (in sequential order or not) that can decrease bacterial growth.
TABLE 1.
Antibiotic list and properties on the ancestral strain of Staphylococcus epidermidis (ATCC 14990).
| Antibiotic (abbreviation) | Class | Mechanism of action | Concentration used (μM) | Relative fitness to No drug control | MIC (μM) |
|---|---|---|---|---|---|
| Chloramphenicol (CHL) | Chloramphenicol | Protein Synthesis, 50S | 90 | 0.989 | 1352.621 |
| Doxycycline (DOX) | Tetracycline | Protein Synthesis, 30S | 0.6 | 0.780 | 257.522 |
| Erythromycin (ERY) | Macrolide | Protein Synthesis, 50S | 0.05 | 1.000 | 0.511 |
| Neomycin (NEO) | Aminoglycoside | Protein Synthesis, 30S | 0.35 | 0.803 | 13.280 |
| Piperacillin (PIP) | β‐Lactam | β‐Lactam, Cell wall | 0.6 | 0.966 | 1.627 |
| Tetracycline (TET) | Tetracycline | Protein Synthesis, 30S | 20 | 0.713 | 205.137 |
TABLE 2.
Drug concentrations of synergistic three‐drug combinations with net interaction (DA) and emergent interaction (E 3) values are based on the ancestral strain of Staphylococcus epidermidis (ATCC 14990).
| Three‐drug combination | Drugs in combination (abbreviation) | Single drug concentration (μM) | Fitness effect of the combination | Net interaction (DA) | Emergent interaction (E 3) |
|---|---|---|---|---|---|
| 1 | (A) Piperacillin (PIP) | 0.6 | 0.004 | −0.97 | −0.09 |
| (B) Tetracycline (TET) | 20 | ||||
| (C) Chloramphenicol (CHL) | 90 | ||||
| 2 | (A) Piperacillin (PIP) | 0.6 | 0 | −1 | 1 |
| (B) Tetracycline (TET) | 20 | ||||
| (C) Doxycycline (DOX) | 0.6 | ||||
| 3 | (A) Piperacillin (PIP) | 0.6 | 0 | −1 | 0.98 |
| (B) Tetracycline (TET) | 20 | ||||
| (C) Erythromycin (ERY) | 0.05 | ||||
| 4 | (A) Piperacillin (PIP) | 0.6 | 0 | −1 | −0.08 |
| (B) Tetracycline (TET) | 20 | ||||
| (C) Neomycin (NEO) | 0.35 |
To start the initial populations, we prepared a highly dense cell culture of S. epidermidis by pinning the parental strain of S. epidermidis into 200 μL per well of a 96‐well plate of Lysogeny Broth (LB) media (10 g tryptone, 5 g yeast extract, and 10 g NaCl) and incubated the culture for ~16 h at 37°C shaking at 130 rpm. To evolve resistance to a single antibiotic (those listed in Table 1), the highly dense cell culture was then pinned over to a 96‐well plate containing 200 μL per well of LB and the antibiotic concentration for day 1, beginning with 50% of the parental minimum inhibitory concentration (MIC). The antibiotic concentration was continually doubled every 48 h over 10 days, which is roughly 100 generations, resulting in a final drug concentration of 800% of the parental MIC. To evolve combination ‐resistant strains, we also began with the parental strain of S. epidermidis. Similar to the creation of our single‐drug‐resistant mutants, we utilized a high‐density cell culture of the parental strain. These high‐density cell cultures were then pin‐transferred into wells of a new 96‐well plate filled with 200 μL of LB media with one of the four, three‐drug combinations, as listed in Table 2. These 96‐well plates were then incubated at 37°C for approximately 24 h. Pinning the cell cultures into fresh media and antibiotics occurred every 24 h and continued for 10 days or approximately 100 generations. Drug concentrations for all different drug combinations remained at constant levels for the entire time of the evolution experiment. This was done to keep the interactions within a combination as constant as possible allowing only for adaptations in the populations to influence how an interaction may change (Berenbaum et al., 1983).
Once the 10‐days experimental evolution component concluded, we isolated a single colony from each independently evolved population (each well). For the antibiotic chloramphenicol, resistant mutants were collected at 400% of the parental MIC, since growth would not occur at any higher concentrations. We isolated and confirmed resistance by streak purifying onto LB agar with 800% parental MIC of the respective antibiotic (400% parental MIC for chloramphenicol) or the respective antibiotic combination. We took the selected colonies from the streak purification, grew them in 2 mL Luria Broth and incubated the culture for ~16 h at 37°C shaking at 160 rpm. This cell culture was then re‐purified again on LB agar with the respective 800% (or 400% for CHL) parental MIC of the respective drug or the respective antibiotic combination. Master tubes with aliquots were made from a single colony selected from each of the second purifications that were cultured and stored at −80°C (in 25% glycerol).
2.2. Determination of the dose–response curve
The dose–response curves were determined using a 20‐step, two‐fold serial dilution of the antibiotics starting at 2000 μg/mL. The layout for determining the dose–response curves of all bacterial strains is seen below in Figure 1a. Each well had a total working volume of 200 μL, was inoculated with 50 μL 1 × 105 CFU/mL, and incubated at 37°C for 22 h (Reller et al., 2009). A total of four technical replicates for each dose–response curve determination were done. The data from all four replicates were pooled together into the “drc” package in R to model the dose–response curve (Ritz et al., 2015, 2016). This model was then used to estimate the MIC and the concentration of antibiotics needed to inhibit growth by 50% (IC50). We also included negative controls on each of the 96‐well plates to confirm that there was no contamination of our media.
FIGURE 1.
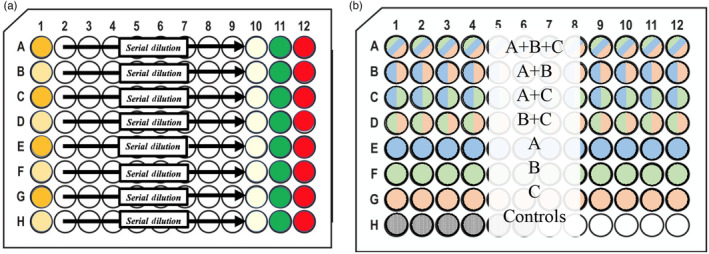
Plate Layouts for the MIC estimates and Deep Well Plates. (a) This figure shows each drug in concentration value in the 96‐well plates during the trials for evaluating the MIC of each bacterial strain. (b) The diagram above illustrates an example of the layout used—a single deep well plate—during the trials. This figure includes the locations for the high‐ and lower‐order drug combinations and their controls all of which are required to determine emergent interactions. These deep well plates were incubated for 22 h at 37°C and later transferred onto a flat bottom 96‐well plates for OD reading.
2.3. Combination interaction determination
The single‐drug resistant mutants showed a phenotype that aggregates at the bottom of the standard flat well plates. This is not uncommon when resistance evolves (Cushnie et al., 2007; Dastgheyb et al., 2015; Haaber et al., 2012; Lozano‐Huntelman et al., 2019; Ritz et al., 2015; Secor et al., 2018). This aggregation did not allow for an accurate optical density, OD, reading that correlates to the number of cells in the culture. To obtain accurate OD readings, we used deep 96‐well plates for the drug combinations so we could resuspend the cells in the media without creating bubbles. The plate set‐up includes all higher‐ and lower‐order combinations along with positive and negative controls as a means to compare the relative growth of bacteria (Figure 1b) which is required for the framework used to determine the interaction values (see next paragraph for more information on this framework). The total working volume of these deep well plates was 400 μL and each well was comprised of 100 μL of LB and 100 μL of the drug combination at their specified concentration, which is described in Table 2. Additionally, these deep‐well plates were inoculated with 200 μL 1 × 105 CFU/mL and incubated for 24 h at 37°C. After the 24‐h incubation period was complete, 200 μL of the culture was transferred to a flat bottom 96‐well plate to gather OD readings.
To calculate net and emergent interactions, we followed the Rescaled Bliss Independence (RBI) framework outlined in Beppler et al. (2016) and Tekin et al. (2016). Other approaches have also been used, such as Loewe Additivity (Loewe, 1953), which, unlike RBI, assumes that a drug cannot interact with itself. However, the RBI framework was chosen for its unique ability to determine the emergent E 3 value (described below) that quantifies how much of the interaction is emerging from having all three drugs in combination, rather than from a strong interaction from a two‐drug combination (Beppler et al., 2016; Tekin et al., 2016). Briefly, in the RBI framework the net deviation from additivity, DA, is determined by only removing the fitness effects contributed by each drug alone () from the overall fitness effect () assuming Bliss independence (Equation 1) (Bliss, 1939). Once the net DA is determined, the process can be done again, removing not only the additive contributions of each drug but also the effects of all lower‐order interactions, leaving only the emergent effect (Equation 2) (Beppler et al., 2016).
| (1) |
| (2) |
After the initial interaction value is determined, a rescaling process is used to better distinguish between interaction types (Tekin et al., 2016). The interaction values were rescaled following the same framework and methodology as used in Tekin, White, et al. (2018).
2.4. Whole genome sequencing and quality control
To ensure strain purity, each strain was grown overnight in the antibiotic regimens they evolved resistance to (either a single drug or in a corresponding combination). DNA extractions were performed using the “Quick‐DNA™ Fungal/Bacterial Miniprep Kit” from Zymo Research (Catalog number: D6005). We followed the manufacturer's protocols which included adding beta‐mercaptoethanol to the genomic lysis buffer. Once DNA was extracted, samples were sent to Novogene for paired‐end sequencing using Illumina's NovaSeq PE150 and paired‐end read quality was evaluated using FastQC (v0.11.9). Additionally, all paired‐end samples were subjected to metagenomic taxonomic classification using the Kraken2 algorithm (Wood et al., 2019). Samples with extensive (>5%) unclassified reads were removed from further analysis. Under this criterion, PIP1, DOX1, DOX6, and DOX7 samples were discarded from downstream analyses.
2.5. Variant analysis
Sample‐specific paired‐end read sets were aligned against the ancestral strain using the Burrows‐Wheeler Alignment BWA‐MEM algorithm and single or multiple nucleotide polymorphisms (SNP/MNP) mutations were detected across mutant genomes using the FreeBayes SNP caller v1.3.2‐1 (Garrison & Marth, 2012). To boost confidence in detected mutations, all gene variants with a Phred score beneath 20—corresponding to a 99% accuracy threshold—were removed from further analysis. Additionally, given the interest in identifying mutations that directly impact known protein function in evolved resistant strains, synonymous mutations and mutations residing in hypothetical gene products were excluded from downstream analysis.
2.6. Statistical analysis
We assessed how the evolution of resistance can change both combination susceptibility and individual antibiotic sensitivity. We measured relative growth (Equation 3) as the growth of a population exposed to an antibiotic(s) normalized to the growth of the same population grown in a drug‐free environment.
| (3) |
When exposing bacterial populations to the combinations, if relative growth was at or below 5%, we considered the strain to be susceptible to the combination. These definitions of relative growth and susceptibility to a combination are used consistently throughout the remainder of the paper.
To assess how the evolution of resistance to a combination (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO) can change individual antibiotic sensitivity, we evaluated the inhibitory concentration at 50% inhibition (IC50). We evaluated the fold change in IC50 (IC50 of resistant strain/IC50 of ansestrial strain) by performing a two‐tailed, one‐sample t‐test defining H0 as μ = 1. We then determined statistical significance through a Holm–Bonferroni correction. To assess how the evolution of resistance to a single drug can change the susceptibility to a combination (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO), we evaluated if resistance to a single drug resulted in more than 5% relative growth by performing a two‐tailed, one‐sample t‐tests to (using H0: μ = 0.05). We then adjusted for multiple comparisons via a Holm–Bonferroni correction.
We also examined how the interaction values may have changed when resistance to a single antibiotic evolved. We took the difference between the interaction value of an evolved strain and the interaction value of the ancestral strain, these values were then compared to zero via a two‐tailed, one‐sample t‐test (H0: μ = 0).
3. RESULTS
We cultivated eight independently evolved resistant strains of S. epidermidis for each individual antibiotic (tetracycline, piperacillin, chloramphenicol, doxycycline, erythromycin, or neomycin). We also created eight independently evolved resistant strains for each of the four 3‐drug combinations: PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, and PIP + TET + NEO. These combinations are all highly synergistic and show at least 95% inhibition of the ancestral strain (ATCC 14990).
3.1. Resistance to combinations leads to changes in single antibiotic susceptibility
We first examined the collateral effects of how the evolution of resistance to the combinations can influence the IC50 of all single antibiotics tested in this study. Over 80% of the observed changes in susceptibility (71 of 86) reflected increases in susceptibility rather than resistance (Figure 2). Further examination of strain‐specific fold changes also yielded more instances of increased sensitivity (Table 3).
FIGURE 2.
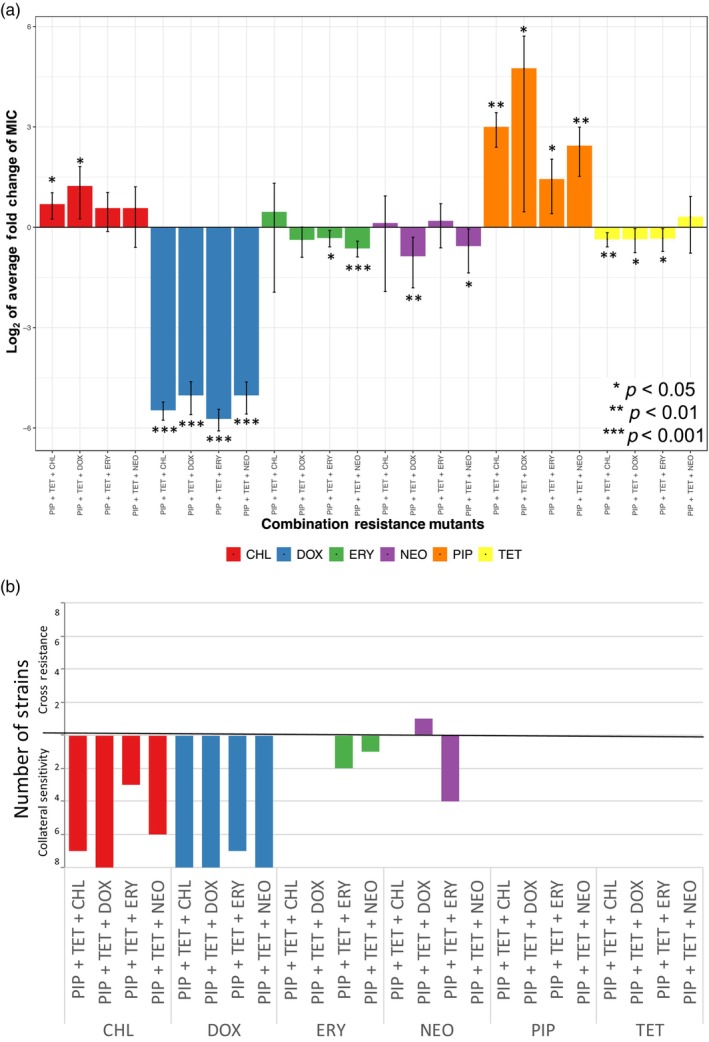
Resistance to synergistic drug combinations results in more incidences of collateral sensitivity than cross‐resistance. (a) The fold change in MIC values (MICresistant/MICancestral) when strains resistance to synergistic drug combinations are exposed to a single antibiotic. (b) The number of strains that had a change in susceptibility to a single antibiotic with a p‐value < 0.002. This indicates a significant change in the MIC after correcting for multiple comparisons. Strains that showed cross‐resistance are above the 0 line while collaterally sensitive strains are below the 0 line.
TABLE 3.
Strain‐specific relative fold change of concentration that inhibits 50% growth with the corresponding p‐value.
| Independently evolved strain | CHL | DOX | ERY | NEO | PIP | TET |
|---|---|---|---|---|---|---|
| PIP + TET + CHL | ||||||
| 1 | Fold change: 0.383 | Fold change: 0.087 | Fold change: 1.096 | Fold change: 1.746 | Fold change: 0.548 | Fold change: 3.34 |
| p = 0 | p = 0 | p = 0.565 | p = 0.074 | p = 0.821 | p = 0.024 | |
| 2 | Fold change: 0.251 | Fold change: 0.043 | Fold change: 1.360 | Fold change: 2.249 | Fold change: 0.581 | Fold change: 1.564 |
| p = 0 | p = 0 | p = 0.127 | p = 0.012 | p = 0.883 | p = 0.203 | |
| 3 | Fold change: 0.324 | Fold change: 0.080 | Fold change: 2.740 | Fold change: 2.144 | Fold change: 0.416 | Fold change: 1.604 |
| p = 0 | p = 0 | p = 0.009 | p = 0.016 | p = 0.698 | p = 0.186 | |
| 4 | Fold change: 0.280 | Fold change: 0.063 | Fold change: 1.720 | Fold change: 1.562 | Fold change: 0.289 | Fold change: 1.320 |
| p = 0 | p = 0 | p = 0.035 | p = 0.249 | p = 0.468 | p = 0.279 | |
| 5 | Fold change: 0.319 | Fold change: 0.052 | Fold change: 2.216 | Fold change: 1.728 | Fold change: 0.470 | Fold change: 2.235 |
| p = 0 | p = 0 | p = 0.027 | p = 0.03 | p = 0.732 | p = 0.055 | |
| 6 | Fold change: 0.548 | Fold change: 0.088 | Fold change: 0.995 | Fold change: 1.522 | Fold change: 0.492 | Fold change: 1.257 |
| p = 0 | p = 0 | p = 0.973 | p = 0.071 | p = 0.752 | p = 0.362 | |
| 7 | Fold change: 0.245 | Fold change: 0.108 | Fold change: 1.495 | Fold change: 1.615 | Fold change: 0.385 | Fold change: 1.425 |
| p = 0 | p = 0 | p = 0.029 | p = 0.102 | p = 0.567 | p = 0.207 | |
| 8 | Fold change: 0.279 | Fold change: 0.061 | Fold change: 0.977 | Fold change: 1.330 | Fold change: 0.218 | Fold change: 1.185 |
| p = NA | p = 0 | p = 0.885 | p = 0.391 | p = 0.011 | p = 0.486 | |
| PIP + TET + DOX | ||||||
| 1 | Fold change: 0.245 | Fold change: 0.124 | Fold change: 0.935 | Fold change: 5.379 | Fold change: 10.379 | Fold change: 2.759 |
| p = 0 | p = 0 | p = 0.646 | p = 0.001 | p = NA | p = 0.012 | |
| 2 | Fold change: 0.270 | Fold change: 0.102 | Fold change: 1.019 | Fold change: 1.712 | Fold change: 0.466 | Fold change: 1.292 |
| p = 0 | p = 0 | p = 0.905 | p = 0.101 | p = 0.856 | p = 0.333 | |
| 3 | Fold change: 0.268 | Fold change: 0.057 | Fold change: 1.126 | Fold change: 1.583 | Fold change 0.263 | Fold change: 1.178 |
| p = 0 | p = 0 | p = 0.565 | p = 0.046 | p = NA | p = 0.502 | |
| 4 | Fold change: 0.280 | Fold change: 0.210 | Fold change 0.894 | Fold change: 1.513 | Fold change: 0.226 | Fold change: 1.386 |
| p = 0 | p = 0 | p = 0.402 | p = 0.065 | p = 0.141 | p = 0.277 | |
| 5 | Fold change: 5.050 | Fold change: 0.084 | Fold change: 0.727 | Fold change: 2.140 | Fold change: 0.438 | Fold change: 1.100 |
| p = 0.377 | p = 0 | p = 0.008 | p = 0.058 | p = 0.748 | p = 0.97 | |
| 6 | Fold change: 0.269 | Fold change: 0.089 | Fold change: 0.847 | Fold change: 3.309 | Fold change: 0.326 | Fold change: 1.171 |
| p = 0 | p = 0 | p = 0.256 | p = 0.01 | p = 0.649 | p = 0.543 | |
| 7 | Fold change: 0.810 | Fold change: 0.087 | Fold change: 0.835 | Fold change: 1.373 | Fold change: 0.244 | Fold change: 2.065 |
| p = 0.002 | p = 0 | p = 0.176 | p = 0.104 | p = 0.065 | p = 0.072 | |
| 8 | Fold change: 0.290 | Fold change: 0.047 | Fold change: 0.755 | Fold change: 4.556 | Fold change: 0.314 | Fold change: 1.324 |
| p = 0 | p = 0 | p = 0.026 | p = 0.011 | p = 0.07 | p = 0.315 | |
| PIP + TET + ERY | ||||||
| 1 | Fold change: 0.134 | Fold change: 0.041 | Fold change: 0.667 | Fold change: 0.612 | Fold change: 0.166 | Fold change: 1.941 |
| p = NA | p = 0 | p = 0.001 | p = 0.002 | p = 0.012 | p = 0.048 | |
| 2 | Fold change: 0.659 | Fold change: 0.086 | Fold change: 1.035 | Fold change: 0.221 | Fold change: 0.073 | Fold change: 1.502 |
| p = NA | p = 0 | p = 0.831 | p = 0 | p = NA | p = 0.249 | |
| 3 | Fold change: 0.324 | Fold change: 0.057 | Fold change: 0.775 | Fold change: 0.532 | Fold change: 0.260 | Fold change: 1.619 |
| p = 0 | p = 0 | p = 0.052 | p = 0 | p = 0.097 | p = 0.618 | |
| 4 | Fold change: 0.601 | Fold change: 0.060 | Fold change: 0.737 | Fold change: 0.589 | Fold change: 0.559 | Fold change: 1.350 |
| p = NA | p = 0 | p = 0.024 | p = 0 | p = 0.698 | p = 0.293 | |
| 5 | Fold change: 0.325 | Fold change: 0.077 | Fold change: 0.622 | Fold change: 0.587 | Fold change: 1.408 | Fold change: 1.699 |
| p = NA | p = NA | p = 0 | p = 0.004 | p = 0.898 | p = 0.186 | |
| 6 | Fold change: 0.600 | Fold change: 0.081 | Fold change: 0.830 | Fold change: 0.616 | Fold change: 10.437 | Fold change: 1.119 |
| p = NA | p = 0 | p = 0.18 | p = 0.003 | p = 0.727 | p = 0.663 | |
| 7 | Fold change: 0.259 | Fold change: 0.058 | Fold change: 0.821 | Fold change: 1.010 | Fold change: 0.394 | Fold change: 1.180 |
| p = 0 | p = 0 | p = 0.114 | p = 0.963 | p = 0.438 | p = 0.516 | |
| 8 | Fold change: 0.271 | Fold change: 0.099 | Fold change: 1.660 | Fold change: 1.895 | Fold change: 0.547 | Fold change: 1.611 |
| p = 0 | p = 0 | p = 0.032 | p = 0.074 | p = 0.714 | p = 0.185 | |
| PIP + TET + NEO | ||||||
| 1 | Fold change: 0.664 | Fold change: 0.065 | Fold change: 0.920 | Fold change: 1.116 | Fold change: 0.248 | Fold change: 2.336 |
| p = 0 | p = 0 | p = 0.559 | p = 0.635 | p = 0.055 | p = 0.037 | |
| 2 | Fold change: 0.595 | Fold change: 0.053 | Fold change: 0.821 | Fold change: 1.323 | Fold change: 0.241 | Fold change: 1.130 |
| p = 0 | p = 0 | p = 0.129 | p = 0.345 | p = NA | p = 0.648 | |
| 3 | Fold change: 0.645 | Fold change: 0.080 | Fold change: 0.920 | Fold change: 1.078 | Fold change: 0.288 | Fold change: 1.106 |
| p = 0 | p = 0 | p = 0.575 | p = 0.724 | p = 0.399 | p = 0.739 | |
| 4 | Fold change: 0.213 | Fold change: 0.089 | Fold change: 0.904 | Fold change: 1.013 | Fold change: 0.273 | Fold change: 1.205 |
| p = 0 | p = 0 | p = 0.479 | p = 0.952 | p = 0.399 | p = 0.506 | |
| 5 | Fold change: 0.650 | Fold change: 0.080 | Fold change: 0.881 | Fold change: 1.635 | Fold change: 0.305 | Fold change: 1.055 |
| p = 0.0003 | p = 0 | p = 0.325 | p = 0.179 | p = 0.419 | p = NA | |
| 6 | Fold change: 0.688 | Fold change: 0.057 | Fold change: 0.576 | Fold change: 1.214 | Fold change: 15.883 | Fold change: 1.145 |
| p = NA | p = 0 | p = 0 | p = 0.437 | p = 0.564 | p = 0.652 | |
| 7 | Fold change: 0.626 | Fold change: 0.082 | Fold change: 0.781 | Fold change: 1.053 | Fold change: 0.150 | Fold change: 1.577 |
| p = NA | p = 0 | p = 0.050 | p = 0.797 | p = 0.011 | p = 0.335 | |
| 8 | Fold change: 0.196 | Fold change: 0.041 | Fold change: 0.886 | Fold change: 0.916 | Fold change: 0.405 | Fold change: 1.289 |
| p = 0 | p = 0 | p = 0.374 | p = 0.661 | p = 0.593 | p = 0.414 | |
We found that resistance to any of the combination regimens (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO) led to significant changes in the IC50 concentration of doxycycline or chloramphenicol. Nearly all strains resistant to any one of the combinations (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO) showed collateral sensitivity to doxycycline and chloramphenicol (Table 3, Figure 2). To explore potential mechanistic links between resistance to these combination regimens and the resulting observed collateral sensitivities, we used whole genome sequencing to identify highly conserved genetic variants that acquired nonsynonymous mutations independently in combination‐resistant, doxycycline‐resistant, and chloramphenicol‐resistant replicate sets (Figure S1A–F). Within both the combination and collaterally sensitive doxycycline and chloramphenicol strains, highly conserved missense mutations were found in genes that encode mobile element protein and members of the IS200 transposase family (Table 4, Figure 3a,b). Specifically, within a gene for a mobile element protein SE0090, missense 134T>C and 221A>G point mutations, corresponding to Leu45Ser and His74Arg amino acid substitutions were found among the combination and collaterally sensitive replicate sets, in addition to a dipeptide substitution at TyrGln66HisLys (Table 4). Within the SE1292 gene, which encodes a member of the IS200 transposase family, a conserved 181G>A point mutation—corresponding to an Asp61Asn amino acid substitution—was also detected in the combination and collaterally sensitive replicate sets (Table 4). Overlap in mutations between the combination and doxycycline‐resistant sets was also observed in the amino acid/proton symporter YbeC gene and SE0629, which encodes a member of the IS4 transposase family (Table 4, Figure 3a). Within the YbeC gene, a 603G>C mutation and resulting Met201Ile substitution was observed, whereas SE0629 experienced an 802G>A (Asp268Asn) substitution.
TABLE 4.
Highly conserved nonsynonymous mutations across combinations and individual doxycycline‐resistant and chloramphenicol‐resistant Staphylococcus epidermidis strains.
| Resistance | Associated gene/plasmid | Strains | Mutation type | Chromosomal position | Nucleotide substitution | Amino acid substitution |
|---|---|---|---|---|---|---|
| Piperacillin | SE1292 | PIP: 1, 3–8 | Missense | 429 | 181G>A | Asp61Asn |
| PIP: 1, 3–8 | Missense | 545 | 65T>C | Val22Ala | ||
| SE1038 | PIP: 3–5, 8 | Frameshift/Del | 272012 | 451delC | Leu151fs | |
| PIP: 6–7 | Missense | 272162 | 302G>A | Gly101Asp | ||
| Doxycycline | SE0623 | DOX: 2–5, 8 | Missense | 326421 | 104G>A | Gly35Asp |
| YbeC | DOX: 2–5, 8 | Missense | 107568 | 603G>C | Met201Ile | |
| SE0090 | DOX: 2–5, 8 | Missense | 40114 | 221A>G | Hi74Arg | |
| 40136 | 196TATC>CATA | TyrGln66HisLys | ||||
| 40201 | 134T>C | Leu45Ser | ||||
| SE0629 | DOX: 2–5, 8 | Missense | 754 | 802G>A | Asp268Asn | |
| SE1292 | DOX: 2–5, 8 | Missense | 429 | 181G>A | Asp61Asn | |
| DOX: 2, 4–5, 8 | Missense | 545 | 65T>C | Val22Ala | ||
| Chloramphenicol | SE0090 | CHL: 1–8 | Missense | 40114 | 221A>G | His74Arg |
| CHL: 1–5, 7–8 | Missense | 40136 | 196TATC>CATA | TyrGln66HisLys | ||
| Missense | 40201 | 134T>C | Leu45Ser | |||
| SE1292 | CHL: 1–8 | Missense | 208 | 402G>T | Glu134Asp | |
| CHL: 2–8 | Missense | 545 | 65T>C | Val22Ala | ||
| SE0161 | CHL: 1–8 | Missense | 273 | 264AG>GA | GluGly88GluSer | |
| PIP + TET + CHL | YbeC | PIP + TET + CHL: 1–8 | Missense | 107568 | 603G>C | Met201Ile |
| SE0691 | PIP + TET + CHL: 1, 3–5, 7–8 | Stop Gained | 116330 | 766G>T | Glu256* | |
| PIP + TET + CHL: 6 | Stop Gained | 116225 | 661G>T | Glu221* | ||
| PIP + TET + CHL: 1–8 | Missense | 115658 | 94C>T | Pro32Ser | ||
| SE0090 | PIP + TET + CHL: 1–8 | Missense | 40114 | 221A>G | His74Arg | |
| Missense | 40136 | 196TATC>CATA | TyrGln66HisLys | |||
| Missense | 40201 | 134T>C | Leu45Ser | |||
| SE0629 | PIP + TET + CHL: 1–8 | Missense | 754 | 802G>A | Asp268Asn | |
| SE1292 | PIP + TET + CHL: 1–2, 4–8 | Missense | 429 | 181G>A | Asp61Asn | |
| Missense | 545 | 65T>C | Val22Ala | |||
| PIP + TET + CHL: 6 | Missense | 208 | 402G>T | Glu134Asp | ||
| SE0243 | PIP + TET + CHL: 1, 3–5, 7–8 | Missense | 45631 | 363C>G | Asp121Glu | |
| PIP + TET + DOX | YbeC | PIP + TET + CHL: 1–8 | Missense | 107568 | 603G>C | Met201Ile |
| SE0691 | PIP + TET + CHL: 1–5, 6–8 | Stop Gained | 116330 | 766G>T | Glu256* | |
| PIP + TET + CHL: 5 | Missense | 115658 | 94C> | Pro32Ser | ||
| SE0090 | PIP + TET + CHL: 1–8 | Missense | 40114 | 221A>G | His74Arg | |
| Missense | 40136 | 196TATC>CATA | TyrGln66HisLys | |||
| Missense | 40201 | 134T>C | Leu45Ser | |||
| SE0629 | PIP + TET + CHL: 1–8 | Missense | 754 | 802G>A | Asp268Asn | |
| SE1292 | PIP + TET + CHL: 1–7 | Missense | 429 | 181G>A | Asp61Asn | |
| Missense | 545 | 65T>C | Val22Ala | |||
| PIP + TET + CHL: 8 | Missense | 754 | 802G>A | Asp268Asn | ||
| SE0243 | PIP + TET + CHL: 1–4, 6–8 | Missense | 45631 | 363C>G | Asp121Glu | |
| PIP + TET + ERY | YbeC | PIP + TET + ERY: 1–8 | Missense | 107568 | 603G>C | Met201Ile |
| SE0691 | PIP + TET + ERY: 1–6 | Stop Gained | 116330 | 766G>T | Glu256* | |
| PIP + TET + ERY: 7 | Missense | 115943 | 379GC>AA | Ala127Asn | ||
| PIP + TET + ERY: 7 | Missense | 115952 | 388G>T | Asp130Tyr | ||
| PIP + TET + ERY: 8 | Frameshift/Del | 116060 | 503delT | Phe168fs | ||
| SE0090 | PIP + TET + ERY: 1–8 | Missense | 40114 | 221A>G | His74Arg | |
| Missense | 40136 | 196TATC>CATA | TyrGln66HisLys | |||
| Missense | 40201 | 134T>C | Leu45Ser | |||
| SE0629 | PIP + TET + ERY: 1–8 | Missense | 754 | 802G>A | Asp268Asn | |
| SE1292 | PIP + TET + ERY: 1–8 | Missense | 429 | 181G>A | Asp61Asn | |
| Missense | 545 | 65T>C | Val22Ala | |||
| SE0243 | PIP + TET + ERY: 1–6 | Missense | 45631 | 363C>G | Asp121Glu | |
| SE0161 | PIP + TET + ERY: 1–2, 4–6, 8 | Missense | 273 | 264AG>GA | GluGly88GluSer | |
| PIP + TET + NEO | SE0090 | PIP + TET + NEO: 1–8 | Missense | 40114 | 221A>G | His74Arg |
| Missense | 40136 | 196TATC>CATA | TyrGln66HisLys | |||
| Missense | 40201 | 134T>C | Leu45Ser | |||
| SE0629 | PIP + TET + NEO: 1–8 | Missense | 754 | 802G>A | Asp268Asn | |
| SE1292 | PIP + TET + NEO: 1–8 | Missense | 429 | 181G>A | Asp61Asn | |
| Missense | 545 | 65T>C | Val22Ala | |||
| YbeC | PIP + TET + NEO: 1–4, 7–8 | Missense | 107568 | 603G>C | Met201Ile |
Note: All mutations listed were found in ≥75% of the evaluated replicate sets and most mutations conferring missense variants or early termination in translation were consistently found in combination and doxycycline‐resistant and chloramphenicol‐resistant strains.
FIGURE 3.
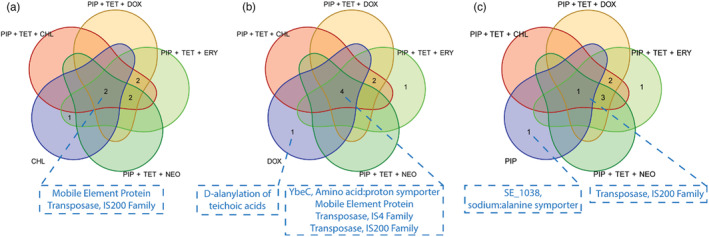
Overlap in highly conserved nonsynonymous genetic variant sets for synergistic drug combinations and collaterally sensitive (a) chloramphenicol‐resistant, (b) doxycycline‐resistant, and (c) piperacillin‐resistant strains. Missense mutations found in genes encoding mobile element protein and members of the IS200 transposase family were found in all drug combination‐resistant strains as well as the individual doxycycline‐resistant or chloramphenicol‐resistant strains. Furthermore, doxycycline resistance coincided with missense mutations in the amino acid‐proton symporter YbeC gene, and members of the IS4 transposase family, which were also found in all combination‐resistant strains. Overlap in nonsynonymous protein‐coding genetic variant sets for synergistic drug combinations and doxycycline resistance. Missense mutations found in genes encoding mobile element protein and the members of the IS4 and IS200 transposase families were conserved across all combination‐resistant and doxycycline‐resistant strains and coincide with the collateral sensitivity of Staphylococcus epidermidis to doxycycline. Mutations exclusive to the conserved doxycycline variant set include a 104G>A nucleotide point mutation, corresponding to a Gly35Asp substitution in the resulting D‐alanine‐poly (phosphoribitol) ligase, subunit one protein product.
Piperacillin and tetracycline are both in all the combinations we studied (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, and PIP + TET + NEO) yet susceptibility to piperacillin and tetracycline did not significantly change in strains that evolved resistance to these combinations (Figure 2, Table 3).
Collateral sensitivity to erythromycin was observed in three of the 32 combination‐resistant strains. One of the PIP + TET + DOX resistance strains showed cross‐resistance to neomycin and four strains of PIP + TET + ERY became collaterally sensitive to neomycin (Table 3, Figure 2, Table S1).
3.2. Resistance to single drugs leads to changes in susceptibility to combinations
Evolving resistance to any single antibiotic typically resulted in an increase in resistance to the combinations to some degree (Table 5, Figure 4). Piperacillin‐resistant strains uniquely remained susceptible (i.e., not significantly greater than 5% relative growth) to all combinations—having the lowest estimated growth means compared to all other single‐drug resistant strains (Figure 4). We performed WGS for these strains to identify highly conserved nonsynonymous variant sets (Figure 3c, Figure S1G). Here, we detected conserved frameshift deletion and missense mutations in the SE1038 gene, encoding the putative amino acid transporter (Table 4). Similar Asp61Asn and Val22Ala missense mutations from the IS200 transposase family member encoding SE1292 gene were also detected here.
TABLE 5.
Comparison of relative growth between ancestral strain and strains with evolved resistance to one of the single drug components.
| Strain | Exposure to combination | Estimate | 95% CI | t‐statistic | p‐value | n | |
|---|---|---|---|---|---|---|---|
| 2.50% | 97.50% | ||||||
| CHL | PIP + TET + CHL | 0.219 | −0.246 | 0.685 | 0.860 | 0.418 | 7 |
| DOX | PIP + TET + CHL | 0.533 | 0.109 | 0.957 | 2.694 | 0.031 | 7 |
| ERY | PIP + TET + CHL | 1.295 | 0.877 | 1.713 | 7.044 | 0.000 | 7 |
| NEO | PIP + TET + CHL | 1.157 | 0.686 | 1.627 | 5.565 | 0.001 | 7 |
| PIP | PIP + TET + CHL | 0.045 | −0.018 | 0.107 | −0.195 | 0.851 | 7 |
| TET | PIP + TET + CHL | 0.433 | −0.021 | 0.886 | 1.996 | 0.086 | 7 |
| CHL | PIP + TET + DOX | 0.048 | 0.006 | 0.090 | −0.108 | 0.917 | 7 |
| DOX | PIP + TET + DOX | 0.459 | 0.104 | 0.814 | 2.726 | 0.030 | 7 |
| ERY | PIP + TET + DOX | 1.226 | 1.002 | 1.451 | 12.844 | 0.000 | 6 |
| NEO | PIP + TET + DOX | 1.054 | 0.594 | 1.515 | 5.161 | 0.001 | 7 |
| PIP | PIP + TET + DOX | 0.035 | −0.007 | 0.078 | −0.826 | 0.436 | 7 |
| TET | PIP + TET + DOX | 0.340 | −0.027 | 0.708 | 1.931 | 0.102 | 6 |
| CHL | PIP + TET + ERY | 0.225 | 0.028 | 0.421 | 2.104 | 0.073 | 7 |
| DOX | PIP + TET + ERY | 0.458 | 0.201 | 0.715 | 3.759 | 0.007 | 7 |
| ERY | PIP + TET + ERY | 1.054 | 0.781 | 1.328 | 8.694 | 0.000 | 7 |
| NEO | PIP + TET + ERY | 0.717 | 0.456 | 0.978 | 6.043 | 0.001 | 7 |
| PIP | PIP + TET + ERY | 0.024 | −0.009 | 0.057 | −1.876 | 0.103 | 7 |
| TET | PIP + TET + ERY | 0.434 | 0.115 | 0.754 | 2.844 | 0.025 | 7 |
| CHL | PIP + TET + NEO | 0.152 | 0.003 | 0.301 | 1.623 | 0.149 | 7 |
| DOX | PIP + TET + NEO | 0.533 | 0.236 | 0.831 | 3.838 | 0.006 | 7 |
| ERY | PIP + TET + NEO | 1.125 | 0.750 | 1.500 | 7.011 | 0.000 | 6 |
| NEO | PIP + TET + NEO | 0.607 | 0.246 | 0.968 | 3.773 | 0.009 | 6 |
| PIP | PIP + TET + NEO | 0.036 | −0.005 | 0.076 | −0.824 | 0.437 | 7 |
| TET | PIP + TET + NEO | 0.351 | 0.130 | 0.573 | 3.213 | 0.015 | 7 |
Note: Significant differences after correcting for multiple tests (via Holm–Bonferroni, α = 0.0016) are shown in bold.
FIGURE 4.
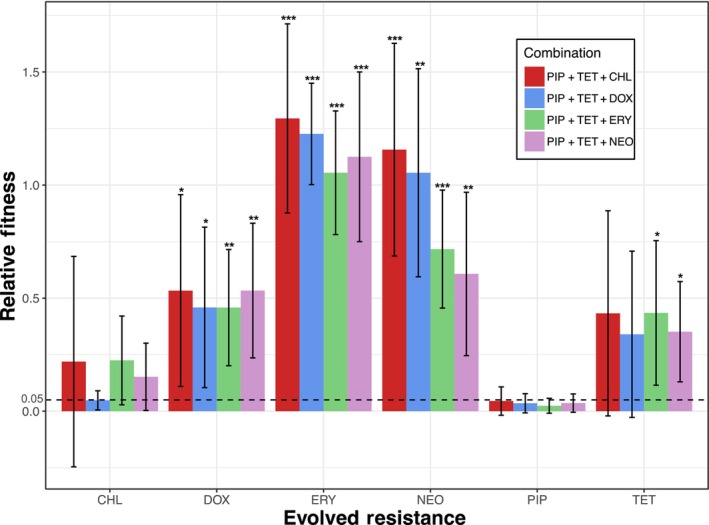
The evolution of resistance to some individual drug components results in a loss in susceptibility to the originally highly synergistic combination. The dashed line indicates a relative growth of 5%. Growth significantly higher than 5% gained some level of resistance to the combination. Otherwise, there was no significant impact on the strain's susceptibility to the highly synergistic combination. Error bars show 95% confidence intervals of the mean (n = 7). *p < 0.05, **p < 0.01, ***p < 0.001.
The chloramphenicol‐resistant strains were still susceptible to PIP + TET + DOX and were highly variable to PIP + TET + CHL, PIP + TET + ERY, and PIP + TET + NEO regimens. Tetracycline‐resistant strains showed mixed susceptibility and high variability when exposed to each combination. They remained susceptible to PIP + TET + CHL and PIP + TET + DOX but also showed decreased susceptibility to PIP + TET + ERY and PIP + TET + NEO. Doxycycline‐resistant strains showed less variation in growth than tetracycline‐resistant strains, yet relative growth did not change when exposed to any of the drug combinations (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO).
All other single antibiotic‐resistant strains (erythromycin‐resistant and neomycin‐resistant strains) led to the three‐drug combinations (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, and PIP + TET + NEO) no longer being effective at suppressing growth. The erythromycin‐resistant and the neomycin‐resistant strains showed the overall strongest resistance to these combinations with some individual strains growing better in the presence of drug combinations than when antibiotics were absent. The strains resistant to a tetracycline‐class antibiotic—the tetracycline‐resistant strains and doxycycline‐resistant strains—showed high variability in growth when exposed to each combination. Although the mean values for the tetracycline‐resistant strains showed an increase in growth (over 5% relative growth), there was a large variation among each strain (Table 5, Figure 4).
3.3. Patterns of cross resistance and collateral sensitivity
We examined patterns of the cross‐resistant and collateral sensitivity networks for all the evolved strains (Figure 5a). The cross‐resistant and collateral sensitivity networks can change among specific replicate populations and even show contrasting outcomes when evolving resistance to the same antibiotic or combination. Thus, we focus our attention on the trends we see among most or all of our independently evolved biological replicates (Figure 5b). Strain‐by‐strain fold changes in the MIC can be found in Figure 2a. For instance, despite the synergistic interaction between the antibiotic combinations, the collateral effects varied depending on the specific antibiotic. The relationships between piperacillin, doxycycline, chloramphenicol, and all the combinations tested in this study, are mostly consistent across a large majority of the independently evolved biological replicates (Figure 5b).
FIGURE 5.
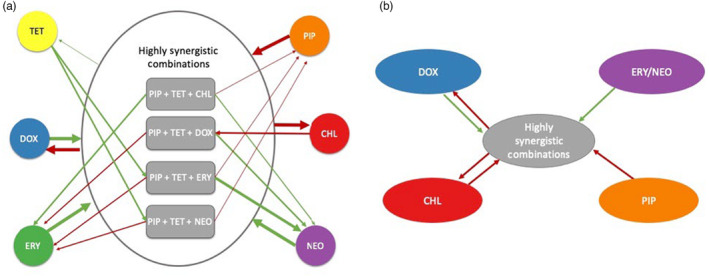
Patterns in the relationships between the four highly synergistic combinations and their individual components. Green arrows show a positive relationship for the bacteria: resistance to the combination/individual drug showed cross‐resistance or loss of sensitivity to the individual drug/combination. Red arrows show a negative relationship for the bacteria: resistance to the combination/individual drug showed collateral sensitivity or remained completely susceptible to the individual drug/combination. (a) Arrow weight shows how consistent the relationship is; the heavier the weight the more likely it is to observe the relationship. (b) Possible viable antibiotic sequence, resulting in less resistance evolving.
We also calculated the MICs of the single‐drug resistant strains to exposure to all other single drugs to consider the patterns of cross‐resistance and collateral sensitivities of single drug resistance among the single drugs (Figure S3). MIC values can be found in Figure S2. Resistance to any of the single drugs usually led to cross‐resistance to erythromycin. In contrast, resistance to any of the single drugs did not result in cross‐resistance—there was either no significant change in MIC or the strain became collaterally sensitive—to tetracycline. Collateral effects to piperacillin were mixed depending on the resistant strain (Figure S3).
3.4. Change in net and emergent interactions values
Drug interactions were measured using both the ancestral strain and the evolved resistant strains to examine whether resistance to a single drug or resistance to the three‐drug combination changes the nature and effects of drug interactions. The interaction values of the combinations for the ancestral strain are listed in Table 2 and the degree of the change in the interaction values is shown in Figure 6 (interaction values for each strain for all two‐drug and three‐drug combinations can be found in Table S1).
FIGURE 6.
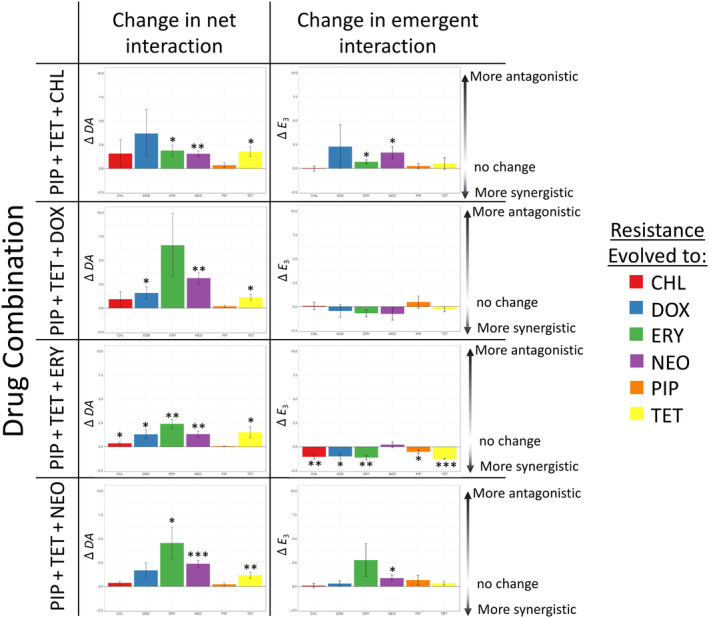
Net interactions are more likely than emergent interactions to be affected by the evolution of antibiotic resistance to a single antibiotic within the combination. Positive values indicate that the interaction is now more antagonistic with the evolved resistance than the ancestral strain. Negative values indicate that the interaction is now more synergistic with the evolved resistance than the ancestral strain (two‐tailed, one‐sample t‐test, μ = 0). Of the combinations tested, PIP + TET + ERY typically shows the most change in interaction values. *p < 0.05, **p < 0.01, ***p < 0.001.
We will first examine the net effects (DA), which are the overall effects that are due to all possible interactions within a combination. The net effects moved away from synergy and toward antagonism (the values became more positive) in cases that resulted in the three‐drug combinations no longer being effective at largely inhibiting growth. Figure 6 shows just how frequently the net effects significantly changed. Piperacillin‐resistant mutants were the only mutants to consistently have unchanged net effects (DA values). This means that the drug combinations remained highly synergistic (p < 0.05, two‐tailed, one‐sample t‐test, H0: μ = 0).
In contrast to the ever‐changing net effects (DA), the emergent effects (E 3) were much more robust. The emergent effects are the effects of the interaction that is solely due to all three antibiotics being present in the combination. The emergent effects have fewer significant changes to the emergent interaction value (eight changes in the emergent effect vs. 14 changes in net effect out of a total possible 24 opportunities to change) (Figure 6). The changes in the emergent effect varied between becoming more antagonistic (positive) or synergistic (negative) with no clear trend. The combination of PIP + TET + ERY was the most susceptible to changes in interaction values, both net and emergent, effects when antibiotic resistance evolves.
When examining the strains resistant to the combinations themselves we saw the same trends as we saw with the single‐drug resistant strains. There was a shift toward antagonist values (more positive values). There were also more significant changes in the net effect than in the emergent effects (Figure S4, Table S1).
4. DISCUSSION
We show that evolving resistance to a combination of antibiotics results in varying types of collateral effects to the individual antibiotics within the combination (Figure 2). This suggests resistance to a combination is not only the result of a stepwise accumulation of mutations offering enhanced resistance to constituent antibiotics. Initially, predictive models for the evolution of resistance to a multidrug combination assumed that the mechanisms of resistance are independent of each other (Johnston et al., 2007; Komarova, 2006; Saputra et al., 2018). That is, a fully sensitive cell cannot become resistant to all drugs in the combination without undergoing multiple mutation events (Komarova, 2006). However, empirically it has been shown that these assumptions were not always supported in two‐drug combinations (Gifford et al., 2019). Work has been done to try to incorporate these findings into predictive evolutionary models (Berríos‐Caro et al., 2021). Furthermore, other factors such as tolerance—the ability for bacterial cells to survive but not actively grow in the presence of antibiotics—have also been shown to influence the predictions of these models (Liu et al., 2020). Even with the progress made and knowledge gained for two‐drug combinations, higher‐order combinations (3+ antibiotics) are not as well studied which is why they are the focus of our study. This study provides further empirical evidence to support the idea that the evolution of resistance to a combination, including higher‐order combinations, is not exclusively the result of a stepwise accumulation of resistance mutations to each antibiotic component within the combination.
Our results show that the evolution of resistance to a single component of a three‐drug combination does not always make the combination ineffective (Figure 4). These findings suggest that it could be crucial to consider combinations of drugs in addition to individual drugs when searching for a more viable antibiotic treatments. Others have also examined how the collateral effects of sequential treatment and the interaction between antibiotics relate for two‐drug combinations. Combinations that are made up of pairs that have the potential for collateral sensitivities tend to slow the rate of antibiotic resistance adaptations (Barbosa et al., 2018). Antagonistic combinations have been shown to slow rates of adaptation and result in resistance profiles that are different from those only resistant to one component of the combination (Dean et al., 2020). While synergistic combinations can have higher efficacy, they can also increase the selective advantage of resistance mutations over wild‐type strains (Torella et al., 2010).
In this study, we incorporated all components of the four 3‐drug combinations (piperacillin, tetracycline, chloramphenicol, doxycycline, erythromycin, neomycin) and the combinations themselves (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, PIP + TET + NEO) to create a collateral effects network (Figure 4a). Using collateral effect networks—and even cellular hysteresis which has a much more rapid timescale (Roemhild et al., 2018)—to develop sequential/cyclical antibiotic treatments has been suggested as one approach to mitigate the problem of antibiotic resistance. In chronic infections such as cystic fibrosis, it may be effective to use phenotypes that, after initial treatment and the following evolved resistance, are still susceptible to other antibiotic options (Imamovic et al., 2018).
However, the success of using a collateral effect network to treat infections is often determined by the pair of antibiotics involved. Sequential use of antibiotics that make up a synergistic pair has been correlated with increased cross‐resistance between the two drugs (Fuentes‐Hernandez et al., 2015; Rodriguez de Evgrafov et al., 2015). However, using collateral sensitivities in principal may be leveraged in a clinical environment (Imamovic & Sommer, 2013). For example, sequential treatment of Escherichia coli with collaterally sensitive drug pairs can have higher efficacy at lower doses than using both antibiotics simultaneously (Fuentes‐Hernandez et al., 2015). In sequential treatments, collateral sensitives have even been shown to regenerate sensitivities to drug regimens that populations were previously resistant to (Barbosa et al., 2019; Dhawan et al., 2017). Antagonistic combinations have been shown to select against resistance adaptations when the bacteria has already adapted to one of the antibiotics within the combinations (Chait et al., 2007). Our findings suggest four potential three‐step treatments and four potential two‐step treatments where each step independently can result in collateral sensitivities.
In our examination of four possible three‐step treatments, we observed that if a bacterial population evolved resistance to piperacillin, it likely became susceptible to any one of the highly synergistic combinations, PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO. The evolution of resistance to a combination leads to collateral sensitivity to doxycycline. The sequence of these treatments began with piperacillin, then any one of the combinations (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO), and ends with doxycycline. This order can promote collateral sensitivity as a population evolved resistance to each step of the sequence. But reversing the order, by evolving resistance to doxycycline first, resulted in the population becoming cross‐resistant to any of the combinations (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO).
Additionally, we investigated four treatments involving only two‐drug regimens: any one of the four 3‐drug combinations (PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, or PIP + TET + NEO) and chloramphenicol. Evolving resistance to any of these three‐drug combinations typically resulted in collateral sensitivities to chloramphenicol, and evolving resistance to chloramphenicol did not significantly influence susceptibility to the combinations. Thus, these treatments may continually select for collateral sensitivity to the other treatment in the pair.
We evaluated the collateral effect networks (Figure 5) assuming that: (1) there is little effect from epistatic interactions among the accumulated adaptive mutations as a population progresses through a sequence of antibiotics and (2) resistance adaptations give similar collateral effects. Epistasis has been identified as a mechanism enabling compensatory mutations to reduce the fitness cost of multidrug resistance (Das et al., 2020; Moura de Sousa et al., 2017) and resistance mutations for the same antibiotic can lead to varying collateral effects (Ardell & Kryazhimskiy, 2021; Maltas & Wood, 2019).
This study has possibly identified four two‐step treatments that are susceptible in both directions. That is, resistance to any one of four drug combinations tested in this study leads to higher susceptibility to chloramphenicol, and resistance to chloramphenicol still results in susceptibility to any of the synergistic drug combinations. Chloramphenicol resistance is typically mediated by monoacetylation and diacetylation by the chloramphenicol acetyltransferase enzyme. In staphylococci, two genes have been associated with this resistance—cfr and fexA (Kehrenberg & Schwarz, 2006). The gene cfr can mediate resistance to multiple antibiotics, such as clindamycin and florfenicol in addition to chloramphenicol (Kehrenberg et al., 2005). In contrast, the fexA gene represents a novel efflux protein that only accepts florfenicol and chloramphenicol (Kehrenberg & Schwarz, 2005).
All of the combinations examined in this study use tetracycline and piperacillin and a third antibiotic allowing for a characterization of the interaction between single‐drug resistance evolution and resistance evolution to these four synergistic combinations. Depending on (1) the type of drug regimen (a combination or an individual antibiotic) first driving populations to evolve resistance and (2) the order of subsequent exposures, the collateral effects show opposing trends where one direction promotes collateral sensitivity and the other promotes cross‐resistance (Figure 5b). For example, if a bacterial population evolves resistance to piperacillin, the collateral effect would not change the susceptibility to any one of the combinations—that is, there would be no collateral effect. The collateral effect of the evolution of resistance to a combination would be collateral sensitivity, in this case to doxycycline. This sequential order could promote continual sensitivity as a population evolves resistance to each step of the sequence. However, if the order is reversed—that is, evolving resistance to doxycycline first, then any one of the highly synergistic combinations, and ending with evolving resistance to piperacillin—the collateral effects of the sequence may lead to cross resistance across each step.
Let us first examine the mechanism of resistance evolution to doxycycline and piperacillin individually. Doxycycline is part of the tetracycline class, a family of antibiotics that inhibit protein synthesis. This is done by preventing the attachment of aminoacyl‐tRNA to the ribosomal acceptor (A) site (Chopra & Roberts, 2001). Evolving resistance to a tetracycline is a textbook example of the removal of the antibiotic through efflux pumps to keep intracellular concentrations low, rendering the antibiotic ineffective (Speer et al., 1992). This strategy has been associated with multidrug resistance across multiple classes due to the non‐specific nature of the efflux pump (Blanco et al., 2016; Webber & Piddock, 2003). However, the doxycycline‐resistant strains cultivated in this study have a mutation in one gene that is conserved that is unique to doxycycline resistance, SE0623. This gene codes for a component that is involved in the D‐alanylation of teichoic acids. The D‐alanylation of teichoic acids has been shown to provide some levels of antimicrobial peptide resistance in streptococcus (Kristian et al., 2005) and is a possible target pathway to inactivate in order to overcome multidrug resistance in MRSA (methicillin‐resistant Staphylococcus aureus) (Coupri et al., 2021). It is possible that evolving more general resistance mechanisms such as the D‐alanylation of teichoic acids may be enough to gain cross resistance to the highly synergistic combinations of this study: PIP + TET + CHL, PIP + TET + DOX, PIP + TET + ERY, PIP + TET + NEO (Figure 4).
Piperacillin is included in all combinations and is a part of the penicillin class, and is a β‐lactam. The main mechanism of resistance for this class is through the increased production of penicillin‐binding proteins (PBPs), proteins that are specific to β‐lactam resistance (Dever & Dermody, 1991). Piperacillin specifically evokes a paradox where it primarily selects for one variant of PBPs (PBP2b) even though its most reactive target is a different PBP (PBP2x). Both of these PBPs are essential and involved in peptidoglycan assembly resulting in resistance and leading to a change in morphology (Philippe et al., 2015). This change in morphology may account for some of the variability observed in the combination resistant strains, which are suspected of evolving piperacillin resistance (Figure 4), due to noise in optical density readings (Stevenson et al., 2016). Our results show that strains with evolved resistance to piperacillin both remained susceptible to the drug combinations and possessed a conserved mutation in genes SE1292 and SE1038. This would suggest that the evolution of a more drug‐specific resistance adaptation, such as the PBPs selected for piperacillin resistance, is not enough to gain cross‐resistance to the synergistic combinations (Figure 5).
We show here that resistance to any of the antibiotic combinations in this study consistently results in collateral sensitivity to doxycycline and chloramphenicol. Whole genome sequencing revealed sets of highly conserved genetic variants that were found in combination‐resistant strains, and strains resistant to doxycycline and chloramphenicol, thus providing insight into potential evolutionary routes that S. epidermidis takes to acquire resistance. Here, highly conserved point mutations in genes encoding mobile genetic elements and transposases were found in combination‐resistant, doxycycline‐resistant, and chloramphenicol‐resistant strains (Table 4, Figure 3a,b), suggesting potential gene transfer mechanisms that are altered to overcome antibiotic stresses. Furthermore, within the doxycycline‐resistant and the combination‐resistant strains, which displayed the most consistent collateral sensitivity to doxycycline, conserved mutations in YbeC implicate a path toward resistance to the combinations involving serine/threonine metabolism. Importantly, these observations are made with the assumption that the mechanistic links between multidrug resistance and collateral sensitivity toward doxycycline and chloramphenicol exposure exist in protein‐coding regions of the S. epidermidis genome. Further investigation into non‐coding regions of the genome must be explicitly taken to rule out their roles in multidrug resistance.
Historically, net interactions (DA) have been the focus of evolutionary biologists (Chen et al., 2008; Cokol et al., 2017; Katzir et al., 2019; Zimmer et al., 2016) and ecologists (McCoy et al., 2012; Sitvarin & Rypstra, 2014; Sokol‐Hessner & Schmitz, 2002) because of the difficulties in determining and describing higher‐order emergent interactions accurately. Within the past 15 years, there has been substantial progress in determining and analyzing higher‐order interactions (Beppler et al., 2016, 2017; Cokol et al., 2017; Lozano‐Huntelman et al., 2021; Palmer et al., 2015; Tekin et al., 2016; Tekin, White, et al., 2018; Yeh et al., 2009; Zimmer et al., 2016). In both net and emergent interactions, the effects of an interaction are not primarily due to the chemical interactions between the compounds but rather due to how each individual component affects the physiology of the cell and how those effects interact (Bollenbach, 2015; Bollenbach et al., 2009; Mason et al., 2017). To add to the potential complexities of antibiotic interactions, interaction types may change when the concentration of the antibiotic changes, even if they are kept in the same ratios (Berenbaum et al., 1983), and a shift in specific dose combination can lead to different evolutionary outcomes (Gjini & Wood, 2021). Once the physiology of the cell is altered through evolved resistance adaptations, these interactions have the potential to change.
This study is the first to examine how both net and emergent interactions of drug combinations change in response to antibiotic resistance evolution. By testing synergistic combinations in both the ancestral susceptible strain and the single drug‐resistant strains, our results show that net interactions are more prone to change when a population evolves (i.e., adapted antibiotic resistance) (Figure 6). This makes the net interactions a more dynamic factor in response to the accumulation, loss, or change of physiological functions throughout the evolutionary history of a population. In contrast, emergent interactions appear to be more robust to these physiological changes and adaptations. Understanding how these emergent interactions can affect the evolutionary trajectory of populations will be key to creating long‐term plans to assess antibiotic resistance in natural and clinical populations. This is because although populations will continue to evolve and adapt to new and changing environments (Fitzgerald, 2019; Santos‐Lopez et al., 2019) the effects of emergent interactions may be more likely to persist due to their higher robustness, as shown in Figure 6. The ability to fine‐tune selection pressures to control evolutionary trajectories has been a goal for evolutionary biologists (Fischer et al., 2015; Iram et al., 2021) and we suggest that emergent interactions may be something to consider as a contributing factor.
In conclusion, we show that: (1) Evolving resistance to a combination of antibiotics being used simultaneously does not often lead to cross‐resistance to all of the components of that combination. (2) The evolution of resistance to one component of a combination does not always lead to cross‐resistance to the combination. (3) The evolution of resistance to a single antibiotic affects the net interaction more often than the emergent interaction of a combination. Our findings suggest that it is important to consider antibiotic combinations in addition to individual antibiotics when measuring and examining cross‐resistance and collateral sensitivity networks. Using those methods, we identified four possible two‐step treatments that confer susceptibility and collateral sensitivity to each other and four possible three‐step treatments where the sequence of the drug regimens can promote both types of collateral effects. This framework—examining both collateral sensitivity and cross‐resistance across whole networks of interactions at both the additive and emergent interaction levels—could allow researchers to uncover more viable sequential/cyclical treatment options that would extend the useful life span of antibiotics currently available. We encourage future studies to examine not just these possible treatments but to dive further into the genomic analysis to better identify possible antibiotic resistance genes that may be the target for these antibiotic combinations aside from those associated with each individual drug.
CONFLICT OF INTEREST STATEMENT
The authors declare no conflicts of interest.
Supporting information
Data S1
ACKNOWLEDGMENTS
We thank Iris Yang, Tony Li, Cynthia White, and Tina Manzhu Kang for their aid in experiments identifying the antibiotic combinations examined here. We also thank Portia Mira and Nicholas Lozano for helpful comments that improved the manuscript. We are grateful for funding from the Hellman Foundation (P.Y.) and a KL2 Fellowship (P.Y.) through the NIH/National Center for Advancing Translational Sciences (NCATS) UCLA CTSI Grant Number UL1TR001881.
Lozano‐Huntelman, N. A. , Bullivant, A. , Chacon‐Barahona, J. , Valencia, A. , Ida, N. , Zhou, A. , Kalhori, P. , Bello, G. , Xue, C. , Boyd, S. , Kremer, C. , & Yeh, P. J. (2023). The evolution of resistance to synergistic multi‐drug combinations is more complex than evolving resistance to each individual drug component. Evolutionary Applications, 16, 1901–1920. 10.1111/eva.13608
DATA AVAILABILITY STATEMENT
The raw data will be available through Mendeley Data at https://data.mendeley.com/datasets/cfdk3y9skf/1. Sequencing data is available via the National Center for Biotechnology Information under BioProject ID PRJNA1033048.
REFERENCES
- Ahmad, M. , & Khan, A. U. (2019). Global economic impact of antibiotic resistance: A review. Journal of Global Antimicrobial Resistance, 19, 313–316. [DOI] [PubMed] [Google Scholar]
- Ardell, S. M. , & Kryazhimskiy, S. (2021). The population genetics of collateral resistance and sensitivity. eLife, 10, e73250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbosa, C. , Beardmore, R. , Schulenburg, H. , & Jansen, G. (2018). Antibiotic combination efficacy (ACE) networks for a Pseudomonas aeruginosa model. PLoS Biology, 16(4), e2004356. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbosa, C. , Römhild, R. , Rosenstiel, P. , & Schulenburg, H. (2019). Evolutionary stability of collateral sensitivity to antibiotics in the model pathogen Pseudomonas aeruginosa . eLife, 8, e51481. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbosa, C. , Trebosc, V. , Kemmer, C. , Rosenstiel, P. , Beardmore, R. , Schulenburg, H. , & Jansen, G. (2017). Alternative evolutionary paths to bacterial antibiotic resistance cause distinct collateral effects. Molecular Biology and Evolution, 34(9), 2229–2244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beppler, C. , Tekin, E. , Mao, Z. , White, C. , McDiarmid, C. , Vargas, E. , Miller, J. H. , Savage, V. M. , & Yeh, P. J. (2016). Uncovering emergent interactions in three‐way combinations of stressors. Journal of the Royal Society Interface, 13(125), 20160800. 10.1098/rsif.2016.0800 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beppler, C. , Tekin, E. , White, C. , Mao, Z. , Miller, J. H. , Damoiseaux, R. , Savage, V. M. , & Yeh, P. J. (2017). When more is less: Emergent suppressive interactions in three‐drug combinations. BMC Microbiology, 17(1), 107. 10.1186/s12866-017-1017-3 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berenbaum, M. C. , Yu, V. L. , & Felegie, T. P. (1983). Synergy with double and triple antibiotic combinations compared. Journal of Antimicrobial Chemotherapy, 12(6), 555–563. [DOI] [PubMed] [Google Scholar]
- Berríos‐Caro, E. , Gifford, D. R. , & Galla, T. (2021). Competition delays multi‐drug resistance evolution during combination therapy. Journal of Theoretical Biology, 509, 110524. [DOI] [PubMed] [Google Scholar]
- Blair, J. M. , Webber, M. A. , Baylay, A. J. , Ogbolu, D. O. , & Piddock, L. J. (2015). Molecular mechanisms of antibiotic resistance. Nature Reviews Microbiology, 13(1), 42–51. 10.1038/nrmicro3380 [DOI] [PubMed] [Google Scholar]
- Blanco, P. , Hernando‐Amado, S. , Reales‐Calderon, J. A. , Corona, F. , Lira, F. , Alcalde‐Rico, M. , Bernardini, A. , Sanchez, M. B. , & Martinez, J. L. (2016). Bacterial multidrug efflux pumps: Much more than antibiotic resistance determinants. Microorganisms, 4(1), 14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bliss, C. (1939). The toxicity of poisons applied jointly. Annals of Applied Biology, 26(3), 585–615. [Google Scholar]
- Bollenbach, T. (2015). Antimicrobial interactions: Mechanisms and implications for drug discovery and resistance evolution. Current Opinion in Microbiology, 27, 1–9. [DOI] [PubMed] [Google Scholar]
- Bollenbach, T. , Quan, S. , Chait, R. , & Kishony, R. (2009). Nonoptimal microbial response to antibiotics underlies suppressive drug interactions. Cell, 139(4), 707–718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, E. M. , & Nathwani, D. (2005). Antibiotic cycling or rotation: A systematic review of the evidence of efficacy. Journal of Antimicrobial Chemotherapy, 55(1), 6–9. [DOI] [PubMed] [Google Scholar]
- Chait, R. , Craney, A. , & Kishony, R. (2007). Antibiotic interactions that select against resistance. Nature, 446(7136), 668–671. [DOI] [PubMed] [Google Scholar]
- Chen, C. Y. , Hathaway, K. M. , Thompson, D. G. , & Folt, C. L. (2008). Multiple stressor effects of herbicide, pH, and food on wetland zooplankton and a larval amphibian. Ecotoxicology and Environmental Safety, 71(1), 209–218. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chopra, I. , & Roberts, M. (2001). Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiology and Molecular Biology Reviews, 65(2), 232–260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chow, L. K. , Ghaly, T. M. , & Gillings, M. R. (2021). A survey of sub‐inhibitory concentrations of antibiotics in the environment. Journal of Environmental Sciences, 99, 21–27. [DOI] [PubMed] [Google Scholar]
- Coates, A. R. , Hu, Y. , Holt, J. , & Yeh, P. (2020). Antibiotic combination therapy against resistant bacterial infections: Synergy, rejuvenation and resistance reduction. Expert Review of Anti‐Infective Therapy, 18(1), 5–15. [DOI] [PubMed] [Google Scholar]
- Cokol, M. , Kuru, N. , Bicak, E. , Larkins‐Ford, J. , & Aldridge, B. B. (2017). Efficient measurement and factorization of high‐order drug interactions in mycobacterium tuberculosis. Science Advances, 3(10), e1701881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coupri, D. , Verneuil, N. , Hartke, A. , Liebaut, A. , Lequeux, T. , Pfund, E. , & Budin‐Verneuil, A. (2021). Inhibition of d‐alanylation of teichoic acids overcomes resistance of methicillin‐resistant Staphylococcus aureus . Journal of Antimicrobial Chemotherapy, 76(11), 2778–2786. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cushnie, T. , Hamilton, V. , Chapman, D. , Taylor, P. , & Lamb, A. (2007). Aggregation of Staphylococcus aureus following treatment with the antibacterial flavonol galangin. Journal of Applied Microbiology, 103(5), 1562–1567. [DOI] [PubMed] [Google Scholar]
- Das, S. G. , Direito, S. O. , Waclaw, B. , Allen, R. J. , & Krug, J. (2020). Predictable properties of fitness landscapes induced by adaptational tradeoffs. eLife, 9, e55155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dastgheyb, S. , Parvizi, J. , Shapiro, I. M. , Hickok, N. J. , & Otto, M. (2015). Effect of biofilms on recalcitrance of staphylococcal joint infection to antibiotic treatment. The Journal of Infectious Diseases, 211(4), 641–650. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dean, Z. , Maltas, J. , & Wood, K. B. (2020). Antibiotic interactions shape short‐term evolution of resistance in E. faecalis . PLoS Pathogens, 16(3), e1008278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dever, L. A. , & Dermody, T. S. (1991). Mechanisms of bacterial resistance to antibiotics. Archives of Internal Medicine, 151(5), 886–895. [PubMed] [Google Scholar]
- Dhawan, A. , Nichol, D. , Kinose, F. , Abazeed, M. E. , Marusyk, A. , Haura, E. B. , & Scott, J. G. (2017). Collateral sensitivity networks reveal evolutionary instability and novel treatment strategies in ALK mutated non‐small cell lung cancer. Scientific Reports, 7(1), 1–9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eagle, H. , & Musselman, A. (1948). The rate of bactericidal action of penicillin in vitro as a function of its concentration, and its paradoxically reduced activity at high concentrations against certain organisms. The Journal of Experimental Medicine, 88(1), 99–131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer, A. , Vázquez‐García, I. , & Mustonen, V. (2015). The value of monitoring to control evolving populations. Proceedings of the National Academy of Sciences of the United States of America, 112(4), 1007–1012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fitzgerald, D. M. (2019). Bacterial evolution: The road to resistance. eLife, 8, e52092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fuentes‐Hernandez, A. , Plucain, J. , Gori, F. , Pena‐Miller, R. , Reding, C. , Jansen, G. , Schulenburg, H. , Gudelj, I. , & Beardmore, R. (2015). Using a sequential regimen to eliminate bacteria at sublethal antibiotic dosages. PLoS Biology, 13(4), e1002104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garrison, E. , & Marth, G. (2012). Haplotype‐based variant detection from short‐read sequencing. arXiv. 10.48550/arXiv.1207.3907 [DOI]
- Garrod, L. P. (1935). The effect of bacterial numbers on minimum bacteriostatic concentrations. The Journal of Infectious Diseases, 57, 247–251. [Google Scholar]
- Gifford, D. R. , Berríos‐Caro, E. , Joerres, C. , Galla, T. , & Knight, C. G. (2019). Mutators drive evolution of multi‐resistance to antibiotics. bioRxiv. 10.1101/643585 [DOI] [PMC free article] [PubMed]
- Gjini, E. , & Wood, K. B. (2021). Price equation captures the role of drug interactions and collateral effects in the evolution of multidrug resistance. eLife, 10, e64851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomez, J. E. , Kaufmann‐Malaga, B. B. , Wivagg, C. N. , Kim, P. B. , Silvis, M. R. , Renedo, N. , Ioerger, T. R. , Ahmad, R. , Livny, J. , Fishbein, S. , Sacchettini, J. C. , Carr, S. A. , & Hung, D. T. (2017). Ribosomal mutations promote the evolution of antibiotic resistance in a multidrug environment. eLife, 6, e20420. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haaber, J. , Cohn, M. T. , Frees, D. , Andersen, T. J. , & Ingmer, H. (2012). Planktonic aggregates of Staphylococcus aureus protect against common antibiotics. PLoS One, 7(7), e41075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haight, T. H. , & Finland, M. (1952). Resistance of bacteria to erythromycin. Proceedings of the Society for Experimental Biology and Medicine, 81(1), 183–188. [DOI] [PubMed] [Google Scholar]
- Hegreness, M. , Shoresh, N. , Damian, D. , Hartl, D. , & Kishony, R. (2008). Accelerated evolution of resistance in multidrug environments. Proceedings of the National Academy of Sciences of the United States of America, 105(37), 13977–13981. 10.1073/pnas.0805965105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hernando‐Amado, S. , Coque, T. M. , Baquero, F. , & Martínez, J. L. (2019). Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nature Microbiology, 4(9), 1432–1442. [DOI] [PubMed] [Google Scholar]
- Imamovic, L. , Ellabaan, M. M. H. , Dantas Machado, A. M. , Citterio, L. , Wulff, T. , Molin, S. , Krogh Johansen, H. , & Sommer, M. O. A. (2018). Drug‐driven phenotypic convergence supports rational treatment strategies of chronic infections. Cell, 172(1–2), 121–134.e14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imamovic, L. , & Sommer, M. O. (2013). Use of collateral sensitivity networks to design drug cycling protocols that avoid resistance development. Science Translational Medicine, 5(204), 204ra132. [DOI] [PubMed] [Google Scholar]
- Iram, S. , Dolson, E. , Chiel, J. , Pelesko, J. , Krishnan, N. , Güngör, Ö. , Kuznets‐Speck, B. , Deffner, S. , Ilker, E. , Scott, J. G. , & Hinczewski, M. (2021). Controlling the speed and trajectory of evolution with counterdiabatic driving. Nature Physics, 17(1), 135–142. [Google Scholar]
- Jin, M. , Liu, L. , Wang, D.‐N. , Yang, D. , Liu, W.‐L. , Yin, J. , Yang, Z. W. , Wang, H. R. , Qiu, Z. G. , Shen, Z. Q. , Shi, D. Y. , Li, H. B. , Guo, J. H. , & Li, J. W. (2020). Chlorine disinfection promotes the exchange of antibiotic resistance genes across bacterial genera by natural transformation. The ISME Journal, 14(7), 1847–1856. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Johnston, M. D. , Edwards, C. M. , Bodmer, W. F. , Maini, P. K. , & Chapman, S. J. (2007). Mathematical modeling of cell population dynamics in the colonic crypt and in colorectal cancer. Proceedings of the National Academy of Sciences of the United States of America, 104(10), 4008–4013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Katzir, I. , Cokol, M. , Aldridge, B. B. , & Alon, U. (2019). Prediction of ultra‐high‐order antibiotic combinations based on pairwise interactions. PLoS Computational Biology, 15(1), e1006774. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kavanaugh, L. , Harrison, S. , Flanagan, J. , & Steck, T. (2021). Antibiotic cycling reverts extensive drug resistant Burkholderia multivorans . Antimicrobial Agents and Chemotherapy, 65, e0061121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kehrenberg, C. , & Schwarz, S. (2005). Florfenicol‐chloramphenicol exporter gene fexA is part of the novel transposon Tn 558. Antimicrobial Agents and Chemotherapy, 49(2), 813–815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kehrenberg, C. , & Schwarz, S. (2006). Distribution of florfenicol resistance genes fexA and cfr among chloramphenicol‐resistant Staphylococcus isolates. Antimicrobial Agents and Chemotherapy, 50(4), 1156–1163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kehrenberg, C. , Schwarz, S. , Jacobsen, L. , Hansen, L. H. , & Vester, B. (2005). A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: Methylation of 23S ribosomal RNA at A2503. Molecular Microbiology, 57(4), 1064–1073. [DOI] [PubMed] [Google Scholar]
- Kleine, C.‐E. , Schlabe, S. , Hischebeth, G. T. , Molitor, E. , Pfeifer, Y. , Wasmuth, J.‐C. , & Spengler, U. (2017). Successful therapy of a multidrug‐resistant extended‐spectrum β‐lactamase–producing and fluoroquinolone‐resistant Salmonella enterica subspecies enterica serovar Typhi infection using combination therapy of meropenem and fosfomycin. Clinical Infectious Diseases, 65(10), 1754–1756. [DOI] [PubMed] [Google Scholar]
- Kollef, M. H. (2001). Is there a role for antibiotic cycling in the intensive care unit? Critical Care Medicine, 29(4), N135–N142. [DOI] [PubMed] [Google Scholar]
- Komarova, N. (2006). Stochastic modeling of drug resistance in cancer. Journal of Theoretical Biology, 239(3), 351–366. [DOI] [PubMed] [Google Scholar]
- Kristian, S. A. , Datta, V. , Weidenmaier, C. , Kansal, R. , Fedtke, I. , Peschel, A. , Gallo, R. L. , & Nizet, V. (2005). D‐alanylation of teichoic acids promotes group a streptococcus antimicrobial peptide resistance, neutrophil survival, and epithelial cell invasion. Journal of Bacteriology, 187(19), 6719–6725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, J. , Gefen, O. , Ronin, I. , Bar‐Meir, M. , & Balaban, N. Q. (2020). Effect of tolerance on the evolution of antibiotic resistance under drug combinations. Science, 367(6474), 200–204. [DOI] [PubMed] [Google Scholar]
- Loewe, S. (1953). The problem of synergism and antagonism of combined drugs. Arzneimittel‐Forschung, 3, 285–290. [PubMed] [Google Scholar]
- Lozano‐Huntelman, N. A. , Singh, N. , Valencia, A. , Mira, P. , Sakayan, M. , Boucher, I. , Tang, S. , Brennan, K. , Gianvecchio, C. , Fitz‐Gibbon, S. , & Yeh, P. J. (2019). Evolution of antibiotic cross‐resistance and collateral sensitivity in Staphylococcus epidermidis using the mutant prevention concentration and the mutant selection window. Evolutionary Applications, 13, 808–823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lozano‐Huntelman, N. A. , Zhou, A. , Tekin, E. , Cruz‐Loya, M. , Østman, B. , Boyd, S. , Savage, V. M. , & Yeh, P. (2021). Hidden suppressive interactions are common in higher‐order drug combinations. iScience, 24, 102355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maillard, J.‐Y. , Bloomfield, S. F. , Courvalin, P. , Essack, S. Y. , Gandra, S. , Gerba, C. P. , Rubino, J. R. , & Scott, E. A. (2020). Reducing antibiotic prescribing and addressing the global problem of antibiotic resistance by targeted hygiene in the home and everyday life settings: A position paper. American Journal of Infection Control, 48, 1090–1099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maltas, J. , & Wood, K. B. (2019). Pervasive and diverse collateral sensitivity profiles inform optimal strategies to limit antibiotic resistance. PLoS Biology, 17(10), e3000515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martin‐Loeches, I. , Lisboa, T. , Rodriguez, A. , Putensen, C. , Annane, D. , Garnacho‐Montero, J. , Restrepo, M. I. , & Rello, J. (2010). Combination antibiotic therapy with macrolides improves survival in intubated patients with community‐acquired pneumonia. Intensive Care Medicine, 36(4), 612–620. [DOI] [PubMed] [Google Scholar]
- Mason, D. J. , Stott, I. , Ashenden, S. , Weinstein, Z. B. , Karakoc, I. , Meral, S. , Kuru, N. , Bender, A. , & Cokol, M. (2017). Prediction of antibiotic interactions using descriptors derived from molecular structure. Journal of Medicinal Chemistry, 60(9), 3902–3912. [DOI] [PubMed] [Google Scholar]
- Masterton, R. G. (2005). Antibiotic cycling: More than it might seem? Journal of Antimicrobial Chemotherapy, 55(1), 1–5. [DOI] [PubMed] [Google Scholar]
- McCoy, M. W. , Stier, A. C. , & Osenberg, C. W. (2012). Emergent effects of multiple predators on prey survival: The importance of depletion and the functional response. Ecology Letters, 15(12), 1449–1456. [DOI] [PubMed] [Google Scholar]
- Michel, J.‐B. , Yeh, P. J. , Chait, R. , Moellering, R. C. , & Kishony, R. (2008). Drug interactions modulate the potential for evolution of resistance. Proceedings of the National Academy of Sciences of the United States of America, 105(39), 14918–14923. 10.1073/pnas.0800944105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moura de Sousa, J. , Balbontín, R. , Durão, P. , & Gordo, I. (2017). Multidrug‐resistant bacteria compensate for the epistasis between resistances. PLoS Biology, 15(4), e2001741. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Munck, C. , Gumpert, H. K. , Wallin, A. I. N. , Wang, H. H. , & Sommer, M. O. (2014). Prediction of resistance development against drug combinations by collateral responses to component drugs. Science Translational Medicine, 6(262), 262ra156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichol, D. , Jeavons, P. , Fletcher, A. G. , Bonomo, R. A. , Maini, P. K. , Paul, J. L. , Gatenby, R. A. , Anderson, A. R. , & Scott, J. G. (2015). Steering evolution with sequential therapy to prevent the emergence of bacterial antibiotic resistance. PLoS Computational Biology, 11(9), e1004493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nichol, D. , Rutter, J. , Bryant, C. , Hujer, A. M. , Lek, S. , Adams, M. D. , Jeavons, P. , Anderson, A. R. A. , Bonomo, R. A. , & Scott, J. G. (2019). Antibiotic collateral sensitivity is contingent on the repeatability of evolution. Nature Communications, 10(1), 1–10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Obolski, U. , Stein, G. Y. , & Hadany, L. (2015). Antibiotic restriction might facilitate the emergence of multi‐drug resistance. PLoS Computational Biology, 11(6), e1004340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orr, H. A. (2000). The rate of adaptation in asexuals. Genetics, 155(2), 961–968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otto, M. (2009). Staphylococcus epidermidis—The ‘accidental’ pathogen. Nature Reviews Microbiology, 7(8), 555–567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pál, C. , Papp, B. , & Lázár, V. (2015). Collateral sensitivity of antibiotic‐resistant microbes. Trends in Microbiology, 23(7), 401–407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmer, A. C. , Toprak, E. , Baym, M. , Kim, S. , Veres, A. , Bershtein, S. , & Kishony, R. (2015). Delayed commitment to evolutionary fate in antibiotic resistance fitness landscapes. Nature Communications, 6(1), 1–8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paul, M. , Lador, A. , Grozinsky‐Glasberg, S. , & Leibovici, L. (2014). Beta lactam antibiotic monotherapy versus beta lactam‐aminoglycoside antibiotic combination therapy for sepsis. Cochrane Database of Systematic Reviews, 2014(1), CD003344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pepin, K. M. , & Wichman, H. A. (2008). Experimental evolution and genome sequencing reveal variation in levels of clonal interference in large populations of bacteriophage φX174. BMC Evolutionary Biology, 8(1), 85. 10.1186/1471-2148-8-85 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Philippe, J. , Gallet, B. , Morlot, C. , Denapaite, D. , Hakenbeck, R. , Chen, Y. , Vernet, T. , & Zapun, A. (2015). Mechanism of β‐lactam action in Streptococcus pneumoniae: The piperacillin paradox. Antimicrobial Agents and Chemotherapy, 59(1), 609–621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raymond, B. (2019). Five rules for resistance management in the antibiotic apocalypse, a road map for integrated microbial management. Evolutionary Applications, 12(6), 1079–1091. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reller, L. B. , Weinstein, M. , Jorgensen, J. H. , & Ferraro, M. J. (2009). Antimicrobial susceptibility testing: A review of general principles and contemporary practices. Clinical Infectious Diseases, 49(11), 1749–1755. [DOI] [PubMed] [Google Scholar]
- Ritz, C. , Baty, F. , Streibig, J. C. , & Gerhard, D. (2015). Dose‐response analysis using R. PLoS One, 10(12), e0146021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ritz, C. , Strebig, J. C. , & Ritz, M. C. (2016). Package ‘drc’ . Creative Commons.
- Rodriguez de Evgrafov, M. , Gumpert, H. , Munck, C. , Thomsen, T. T. , & Sommer, M. O. (2015). Collateral resistance and sensitivity modulate evolution of high‐level resistance to drug combination treatment in Staphylococcus aureus . Molecular Biology and Evolution, 32(5), 1175–1185. [DOI] [PubMed] [Google Scholar]
- Roemhild, R. , Gokhale, C. S. , Dirksen, P. , Blake, C. , Rosenstiel, P. , Traulsen, A. , Andersson, D. I. , & Schulenburg, H. (2018). Cellular hysteresis as a principle to maximize the efficacy of antibiotic therapy. Proceedings of the National Academy of Sciences of the United States of America, 115(39), 9767–9772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders, C. C. (2001). Mechanisms responsible for cross‐resistance and dichotomous resistance among the quinolones. Clinical Infectious Diseases, 32(Supplement_1), S1–S8. [DOI] [PubMed] [Google Scholar]
- Santos‐Lopez, A. , Marshall, C. W. , Scribner, M. R. , Snyder, D. J. , & Cooper, V. S. (2019). Evolutionary pathways to antibiotic resistance are dependent upon environmental structure and bacterial lifestyle. eLife, 8, e47612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saputra, E. C. , Huang, L. , Chen, Y. , & Tucker‐Kellogg, L. (2018). Combination therapy and the evolution of resistance: The theoretical merits of synergism and antagonism in cancer. Cancer Research, 78(9), 2419–2431. [DOI] [PubMed] [Google Scholar]
- Secor, P. R. , Michaels, L. A. , Ratjen, A. , Jennings, L. K. , & Singh, P. K. (2018). Entropically driven aggregation of bacteria by host polymers promotes antibiotic tolerance in Pseudomonas aeruginosa . Proceedings of the National Academy of Sciences of the United States of America, 115(42), 10780–10785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sitvarin, M. I. , & Rypstra, A. L. (2014). The importance of intraguild predation in predicting emergent multiple predator effects. Ecology, 95(10), 2936–2945. [Google Scholar]
- Sokol‐Hessner, L. , & Schmitz, O. J. (2002). Aggregate effects of multiple predator species on a shared prey. Ecology, 83(9), 2367–2372. [Google Scholar]
- Speer, B. S. , Shoemaker, N. B. , & Salyers, A. A. (1992). Bacterial resistance to tetracycline: Mechanisms, transfer, and clinical significance. Clinical Microbiology Reviews, 5(4), 387–399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevenson, K. , McVey, A. F. , Clark, I. B. , Swain, P. S. , & Pilizota, T. (2016). General calibration of microbial growth in microplate readers. Scientific Reports, 6(1), 1–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tekin, E. , Beppler, C. , White, C. , Mao, Z. , Savage, V. M. , & Yeh, P. J. (2016). Enhanced identification of synergistic and antagonistic emergent interactions among three or more drugs. Journal of the Royal Society Interface, 13(119), 20160332. 10.1098/rsif.2016.0332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tekin, E. , Savage, V. M. , & Yeh, P. J. (2017). Measuring higher‐order drug interactions: A review of recent approaches. Current Opinion in Systems Biology, 4, 16–23. [Google Scholar]
- Tekin, E. , White, C. , Kang, T. M. , Singh, N. , Cruz‐Loya, M. , Damoiseaux, R. , Savage, V. M. , & Yeh, P. J. (2018). Prevalence and patterns of higher‐order drug interactions in Escherichia coli . NPJ Systems Biology and Applications, 4(1), 31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tekin, E. , Yeh, P. J. , & Savage, V. M. (2018). General form for interaction measures and framework for deriving higher‐order emergent effects. Frontiers in Ecology and Evolution, 6, 166. [Google Scholar]
- Thomson, K. S. , & Sanders, C. C. (1994). Dissociated resistance among fluoroquinolones. Antimicrobial Agents and Chemotherapy, 38(9), 2095–2100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torella, J. P. , Chait, R. , & Kishony, R. (2010). Optimal drug synergy in antimicrobial treatments. PLoS Computational Biology, 6(6), e1000796. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ventola, C. L. (2015). The antibiotic resistance crisis: Part 1: Causes and threats. Pharmacy and Therapeutics, 40(4), 277. [PMC free article] [PubMed] [Google Scholar]
- Vivas, R. , Barbosa, A. A. T. , Dolabela, S. S. , & Jain, S. (2019). Multidrug‐resistant bacteria and alternative methods to control them: An overview. Microbial Drug Resistance, 25(6), 890–908. [DOI] [PubMed] [Google Scholar]
- Webber, M. , & Piddock, L. (2003). The importance of efflux pumps in bacterial antibiotic resistance. Journal of Antimicrobial Chemotherapy, 51(1), 9–11. [DOI] [PubMed] [Google Scholar]
- Wood, D. E. , Lu, J. , & Langmead, B. (2019). Improved metagenomic analysis with Kraken 2. Genome Biology, 20(1), 1–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeh, P. , Hegreness, M. , Aiden, A. P. , & Kishony, R. (2009). Drug interactions and the evolution of antibiotic resistance. Nature Reviews Microbiology, 7, 460–466. 10.1038/nrmicro2133 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zimmer, A. , Katzir, I. , Dekel, E. , Mayo, A. E. , & Alon, U. (2016). Prediction of multidimensional drug dose responses based on measurements of drug pairs. Proceedings of the National Academy of Sciences of the United States of America, 113(37), 10442–10447. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data S1
Data Availability Statement
The raw data will be available through Mendeley Data at https://data.mendeley.com/datasets/cfdk3y9skf/1. Sequencing data is available via the National Center for Biotechnology Information under BioProject ID PRJNA1033048.