

Abstract
Pulsatile gonadotropin-releasing hormone (GnRH) release is critical for reproduction. Disruptions to GnRH secretion patterns may contribute to polycystic ovary syndrome (PCOS). Prenatally androgenized (PNA) female mice recapitulate many neuroendocrine abnormalities observed in PCOS patients. PNA and development induce changes in spontaneous GnRH neuron firing rate, response to synaptic input, and the afterhyperpolarization potential of the action potential. We hypothesized potassium currents are altered by PNA treatment and/or development. Whole-cell patch-clamp recordings were made of transient and residual potassium currents of GnRH neurons in brain slices from 3-week-old and adult control and PNA females. At 3-weeks of age, PNA treatment increased transient current density versus controls. Development and PNA altered voltage-dependent activation and inactivation of the transient current. In controls, transient current activation and inactivation were depolarized at 3-weeks of age versus in adulthood. In GnRH neurons from 3-week-old mice, transient current activation and inactivation were more depolarized in control than PNA mice. Development and PNA treatment interacted to shift the time-dependence of inactivation and recovery from inactivation. Notably, in cells from adult PNA females, recovery was prolonged compared to all other groups. Activation of the residual current occurred at more depolarized membrane potentials in 3-week-old than adult controls. PNA depolarized activation of the residual current in adults. These findings demonstrate the properties of GnRH neuron potassium currents change during typical development, potentially contributing to puberty, and further suggest PNA treatment may both alter some typical developmental changes and induce additional modifications, which together may underlie aspects of the PNA phenotype.
Introduction
Gonadotropin-releasing hormone (GnRH) neurons produce the final central output for the control of reproduction. GnRH release occurs in an episodic pattern, termed pulses1,2. GnRH acts at the anterior pituitary where low-frequency GnRH pulses favor the secretion of follicle-stimulating hormone (FSH), and high-frequency pulses favor the secretion of luteinizing hormone (LH)3,4. Disruptions to the secretion patterns of GnRH, LH and/or FSH result in altered reproductive function5–8. Polycystic ovary syndrome (PCOS) is the leading cause of infertility in females9. Patients with hyperandrogenemic PCOS have persistent high-frequency LH pulse secretion and relative FSH deficiency9–12, suggesting GnRH pulses are also persistently high frequency. Interestingly, PCOS-like symptoms have been observed in peripubertal girls, suggesting the neuroendocrine system is altered even before puberty is complete13,14.
The etiology of PCOS remains largely unknown. In addition to possible genetic causes15,16, it has been hypothesized that in utero exposure to high circulating levels of androgens from the mother might reprogram the reproductive neuroendocrine system. This postulate has been supported by animal models including non-human primates, rats, sheep, and mice17–22. In these models, treatment of pregnant dams with androgens produces offspring that develop neuroendocrine phenotypes similar to those observed in females with hyperandrogenemic PCOS. Adult female prenatally androgenized (PNA) mice exhibit increased circulating testosterone levels, increased LH pulse frequency, and disrupted estrous cycles20,22–24. PNA causes early vaginal opening, an external index of pubertal onset25, suggesting PNA alters reproductive development. Use of mouse models makes it possible to measure neurophysiologic parameters. Spontaneous GnRH neuron firing rate changes with development in controls and is altered by PNA treatment in both prepubertal and adult mice. Specifically, in PNA females, spontaneous GnRH neuron firing rate is low at 3-weeks of age compared to controls but is increased in adulthood20,23. The developmental and PNA-induced changes in spontaneous GnRH neuron activity could be the result of altered neurotransmission to these cells and/or changes in their intrinsic properties. PNA treatment increases the frequency of GABAergic neurotransmission to GnRH neurons in 3-week-old and adult females17,24. Unlike most neurons, GnRH neurons maintain higher intracellular chloride concentration and activation of GABAA receptors can thus excite these neurons26 27. Despite receiving more frequent GABAergic transmission, however, GnRH neuron activity in 3-week-old PNA females is lower than controls23, suggesting that GnRH neurons from PNA females at this age respond differently to GABAergic input. Supporting this postulate, local GABA application increased firing in only about 30% of GnRH neurons from 3-week-old PNA females compared to 100% of neurons from 3-week-old control females and control and PNA adults24. The reduced response to GABA in 3-week-old PNA females is not attributable to differences from 3-week-old control in the baseline membrane potential or intracellular chloride concentration24. Together these findings suggest changes intrinsic to GnRH neurons might regulate how they respond to synaptic input and thus contribute to the altered firing rate.
In this regard, GnRH neurons from control and PNA adults fire more action potentials than control and PNA females at 3-weeks of age in response to the same amount of current injection28. Further, in adulthood the afterhyperpolarization potential of the action potential is larger and delayed relative to prepubertal mice28. The ionic changes underlying these differences are unknown. Here, we focus on voltage-gated potassium currents, which play a major role in determining the response of neurons to the inputs they receive, in shaping firing rate and in the repolarization characteristics of neurons29–32. GnRH neurons display two primary voltage-gated potassium currents, a fast transient current and a residual, delayed-rectifier-like current33. We tested the hypotheses that their properties change with age and with PNA treatment.
Materials and Methods
All chemicals were acquired from Sigma-Aldrich (St. Louis, MO, USA) unless noted otherwise.
Animals.
GnRH-GFP (Tg(Gnrh1-EGFP)51 Sumo MGI:6158457) mice were bred in our colony34. All mice were provided with ad libitum water and Harlan 2916 (non-breeders) or 2919 (breeders) chow. Mice were held on a 14:10 light/dark cycle with lights on at 3AM Eastern Standard Time. To generate female PNA mice, female GnRH-GFP transgenic mice on a C57Bl/6J background and a CD1 female were bred with a C57Bl/6J male. The CD1 dam assists in providing maternal care and nutrition. Females were monitored daily for a copulatory plug (day 1 of pregnancy). On days 16–18 of pregnancy, GnRH-GFP dams were injected subcutaneously with 225μg/day of dihydrotestosterone (DHT) in 50μl sesame oil for PNA treatment or sesame oil for controls. Litter sizes were adjusted to 4–15 pups by culling the CD1 pups, identifiable by size and coat color, to standardize nutrition. Adult female mice were studied on the morning of diestrus, determined by vaginal cytology and confirmed by uterine mass. All procedures were approved by the Institutional Animal Care and Use Committee of the University of Michigan.
Verification of PNA phenotype.
Electrophysiological studies were done on 3-week-old (18–21 d, before vaginal opening, VO) and adult (84–130 d) diestrous females. PNA-induced differences were confirmed for the prepubertal mice in littermates raised to adulthood, as the PNA phenotype is consistent among littermates. PNA status was measured by monitoring age at which VO occurred, estrous cyclicity via vaginal lavage for 14 consecutive days and measuring anogenital distance (AGD) between 70–73d of age (average of three successive daily measures).
Brain slice preparation.
All solutions were bubbled with 95% O2/5% CO2 for at least 15 min before use and throughout experimental recordings. Brain slices were prepared as described35. Brains were removed and placed in ice-cold sucrose saline containing the following (in mM): 250 sucrose, 3.5 KCl, 26 NaHCO3, 10 D-glucose, 1.25 Na2HPO4, 1.2 MgSO4, and 3.8 MgCl2 (350mOsm). Coronal slices (300μm) through the preoptic and anterior hypothalamic region were cut with a Leica VT1200S Microtome (Leica Biosystems, Buffalo Grove, IL). Slices were incubated for 30 min at room temperature (~21–23°C) in 50% sucrose saline and 50% artificial cerebrospinal fluid (ACSF, containing (in mM): 135 NaCl, 3.5 KCl, 26 NaHCO3, 10 D-glucose, 1.25 Na2HPO4, 1.2 MgSO4, 2.5 CaCl2, 315 mOsm, pH 7.4). Slices were then transferred to 100% ACSF at room temperature for 0.5–5 h before recording. No changes in properties were associated with time from slice preparation to recording. A minimum of five mice from at least four litters were studied per group; up to three recordings were used per mouse.
Recording solutions and data acquisition.
Brain slices were transferred to a recording chamber continuously perfused with carboxygenated ACSF (3mL/min) maintained at 30–31°C with an inline-heating unit (Warner Instruments Model SH-27B). Potassium currents were pharmacologically isolated by blocking fast sodium (2μM tetrodotoxin; Tocris Bioscience or Abcam) and calcium (300μM NiCl2) channels as well as ionotropic receptors for GABA (100μM picrotoxin) and glutamate (20μM D-APV and 10μM CNQX; Tocris Bioscience). The pipet solution contained (in mM): 120 K gluconate, 20 KCl, 10 HEPES, 5 EGTA, 0.1 CaCl2, 4 MgATP, and 0.4 NaGTP (305 mOsm, 7.2 pH with NaOH). This solution was based on the native intracellular chloride concentrations in GnRH neurons that was determined using gramicidin perforated-patch recordings26. A 14.5mV liquid junction potential was negated online before each recording. GFP-positive GnRH neurons were visualized with infrared differential interference contrast and fluorescence microscopy on an Olympus BX51WI microscope. Recordings were made using an EPC-10 patch-clamp amplifier and a computer running PatchMaster software (HEKA Elektronik). All current measurements were acquired at 20kHz and filtered at 10kHz. Recording quality and passive properties were monitored throughout experiments from the averaged membrane current response to 16 hyperpolarizing voltage steps from −70mV (5mV, 20ms). Data were analyzed using WaveMetrics IgorPro (Sutter Instruments). Only recordings with an input resistance of >500MΩ, stable compensated series resistance of <15MΩ and a stable capacitance (8pF-20pF) were used for analysis.
Activation/inactivation of voltage-gated potassium currents.
GnRH neurons display two major voltage-gated potassium currents, a fast transient (IA-like) current and a slow residual (IK-like) current. The distinct voltage dependence of inactivation of the transient and residual currents was used to isolate them. GnRH neurons were given a prepulse step to −100mV or −40mV for 500ms. The −40mV or −100mV prepulse was followed by a family of voltage steps from −100mV to +50mV (500ms) in 10mV increments, and then a final step to −10mV for 50ms. Pilot studies showed that a 500ms prepulse at −40mV results in full inactivation of the transient component, leaving the residual current mostly unchanged, and the prepulse at −100mV for 500ms completely removes IA inactivation, yielding total current. The transient current was mathematically isolated by subtracting the current with the −40mV prepulse from that obtained with the −100mV prepulse at each voltage step. Activation of the transient current was estimated using the peak current during the final test pulse to −10mV that followed the family of voltage steps. Leak current was calculated from an average of 4 sweeps at 1/8 amplitude (modified P/−8) from a baseline potential of −70mV36 and subtracted online.
Time dependence of inactivation and recovery following inactivation for transient current.
To measure the time dependence of inactivation, a 500ms prepulse at −100mV was used to remove inactivation. Then, membrane potential was stepped to −40mV for 1, 2, 4, 8, 16, 32, 64, 128, 256, 512, and 1024ms, followed by a test pulse at −10mV to measure the peak current. The non-inactivating component at −10mV after the 1024ms inactivation step was subtracted from all of the other traces to isolate the transient current. To measure the time dependence of recovery from inactivation of the transient current, membrane potential was held at −40mV (500ms) to fully inactivate the fast transient component. Then, membrane potential was stepped to −100mV for 1, 2, 4, 8, 16, 32, 64, 129, 512, and 1024ms, followed by a test pulse at −10mV to measure the peak current. The non-inactivating component at 1ms recovery was subtracted from each of the other traces to isolate the transient component.
Data analysis.
Current density was calculated by dividing the maximum current by the capacitance for each recording. The driving force was calculated using the Goldman-Hodgkin-Katz (GHK) equation37,38:
where V is the step potential and EK was calculated to be −92.6mV after activity corrections39. Potassium conductance was estimated by dividing peak current by driving force. Conductance values were used to plot the voltage-dependent activation of transient and sustained potassium current components.
These data were plotted as a function of voltage and fit with the Boltzmann equation:
where V is the command membrane potential of the step, V0.5 is the potential at which half the current was activated or inactivated, and k is the “slope factor”, F is the Faraday constant, R is the gas constant, and T is the absolute temperature. For inactivation, the driving force is the same for all cells because it is measured at the same membrane potential (test pulse 0mV); because of this, there is no need to convert to conductance for comparison. The Boltzmann equation was thus fit to the normalized peak current as a function of the inactivating prepulse (−100mV to −30mV). The Boltzmann fit was used to estimate the V0.5 activation, V0.5 inactivation and the slope factors.
To characterize the time dependence of inactivation and recovery, the relevant current was normalized to the maximum peak current for each cell and plotted as a function of duration of the prepulse. This was fit to a double exponential equation to account for a fast and slow components:
In this equation, Ystart is the Y value when X (time; ms) is zero; this value is held constant either at 1 for time-dependence of inactivation or 0 for the time-dependence of recovery from inactivation. Yend is the y-value when X is 1024ms; this is held constant at either 0 for the time-dependence of inactivation or 1 for the time-dependence of recovery from inactivation. Ffast, the percent of the fast component, was held constant at 90% for both time-dependence fits. Kfast and Kslow are the rate constants, expressed in the reciprocal of the X-axis time units. Values for the fast (τfast) and slow (τslow) time constants are reported.
Statistics.
Data were tested for normal distribution with the Shapiro-Wilk test. Data are reported as the mean ± SEM or median ± IQR, as appropriate, with the individual values shown when practical. Statistical tests were chosen based on experimental design and data distribution and are specified in the results. Statistical comparisons and all graphs were made in Prism 10.1.1 (GraphPad Software), except for the statistical analysis of the proportion of time spent in each estrous cycle phase, for which RStudio was used40–42 as Prism is unable to run the appropriate chi-squared tests43,44. Analysis in R (R version 4.3.1 “Beagle Scouts”) using RStudio (RStudio Team, 2023) and a combination of open-sourced packages including, rstatix43 and flextable44, for which the “chisq_test”, “chisq_descriptives”, “row_wise_fishers_test”, and the “p.adjust.method” function were used to run the chi-squared tests. Significance was set at p<0.05.
Results
Verification of prenatal androgenization phenotype.
PNA-induced differences were confirmed in this study. Consistent with previous observations17,20,23,28,45 vaginal opening (VO) occurred at a younger age (Mann Whitney U-test) and a lower body mass (Mann Whitney U-test) in PNA (n=24) than control (n=28) females (Figure 1A, B; Table 1). Anogenital distance (AGD) was increased in adult PNA females (two-tailed, unpaired Student’s t-test, Figure 1C; Table 1) and estrous cycles were disrupted. Specifically, PNA females spent more time in diestrus and less time in proestrus and estrus than controls (χ2, Figures 1D, E; Table 1).
Figure 1.

Confirmation of PNA phenotype. A-C. Individual values and median + the interquartile range (IQR) for age at vaginal opening (VO; A) and body mass at VO (B). Individual values and mean ± SEM for anogenital distance in adulthood (AGD, C). D, Representative estrous cycles over 14d. P, proestrus; D, diestrus; E, estrus (top, vehicle; bottom, PNA). E, Individual values and mean ± SEM for days in each cycle over 14d. Statistical parameters are in Table 1.
Table 1.
Statistical parameters characterizing the PNA phenotype.
| Property | VEH median and interquartile range (IQR) | PNA median and interquartile range (IQR) | Mann-Whitney U test | Two-tailed p-value |
|---|---|---|---|---|
| age at vaginal opening (d) | 31.0 (IQR 29.25–32.5) | 24.0 (IQR 21.25–27.00) | U=43 | p<0.0001 |
| body mass (g) at vaginal opening | 14.7 (IQR 14.13–15.4) | 10.15 (IQR 9.38–13.08) | U=71.50 | p<0.0001 |
| Property | Mean ± SEM for AGD | ||
|---|---|---|---|
| AGD (mm) | VEH | PNA | |
| 4.996 ± 0.07 | 6.195 ± 0.08 | ||
| Unpaired, two-tailed Student’s t test | mean difference (VEH-PNA) | Effect size r 2 | |
| t(10.98), df=48; p<0.0001 | Diff[Cl,0.979,1.418] 1.198 ± 0.11 | r2=0.72 | |
| Mean ± SEM for days spent across estrous cycle state | ||
|---|---|---|
| Cycle Stage | VEH | PNA |
| estrus | 6.00 ± 0.381 | 4.583 ± 0.645 |
| diestrus | 5.833 ± 0.333 | 9.292 ± 0.636 |
| proestrus | 2.125 ± 0.202 | 0.125 ± 0.069 |
| Property | χ2 test | ||
|---|---|---|---|
| Estrous cycle stage distribution | χ2= 66.194, n=671, df=2, p<0.0001 | ||
| Estrus | Diestrus | Proestrus | |
| Std. residual | 2.736304 | −6.388093 | 6.823246 |
| Fisher’s exact test; Bonferroni adjusted | p=0.02 | p<0.0001 | p<0.0001 |
Bold indicates p<0.05.
Recording quality parameters and passive properties of GnRH neurons.
Passive properties and series resistance were used to assess the quality of electrophysiological recordings (Figure 2). There were no differences among these parameters among any of the recording protocols, and all recordings were thus combined for quality assessment (two-way ANOVA; Table 2). There were no differences in passive properties or series resistance among groups.
Figure 2.

Recording quality and passive property parameters. A-D, Individual values and mean ± SEM for input resistance (A), compensated series resistance (B), capacitance (C), and holding current (D). Numbers and statistical parameters are in Table 2.
Table 2.
Descriptive statistics and statistical parameters from two-way ANOVA for recording quality parameters and passive properties.
| Protocol | VEH 3-weeks | PNA 3-weeks | VEH adults | PNA adults |
|---|---|---|---|---|
| Voltage-dependence of activation & inactivation (transient & residual currents) | n=14 cells from 7 mice from 5 litters | n=16 cells from 10 mice from 7 litters | n=14 cells from 7 mice from 6 litters | n=14 cells from 10 mice from 10 litters |
| time dependence of inactivation (transient current) | n=13 from 8 mice from 4 litters | n=13 cells from 8 mice from 7 litters | n=13 cells from 7 mice from 6 litters | n=15 cells from 10 mice from 10 litters |
| Time dependence of recovery following inactivation (transient current) | n=13 cells from 8 mice from 4 litters | n=13 cells from 8 mice from 5 litters | n=13 cells from 8 mice from 7 litters | n=14 cells from 9 mice from 9 litters |
| Descriptive Statistics (mean ± SEM) | ||||
|---|---|---|---|---|
| Property | VEH 3-weeks | PNA 3-weeks | VEH adults | PNA adults |
| series resistance (MΩ) | 10.343 ± 0.316 | 9.449 ± 0.319 | 10.376 ± 0.278 | 10.141 ± 0.420 |
| input resistance (MΩ) | 877.413 ± 89.167 | 830.047 ± 62.029 | 841.480 ± 89.866 | 821.913 ± 67.695 |
| capacitance (pF) | 13.529 ± 0.443 | 12.717 ± 0.701 | 13.813 ± 0.740 | 14.691 ± 0.591 |
| holding current (pA) | −43.060 ± 4.929 | −43.564 ± 5.130 | −32.774 ± 5.355 | −45.236 ± 5.546 |
| Two-way ANOVA | |||
|---|---|---|---|
| Property | Age | Treatment | Interaction |
| series resistance (MΩ) | Diff, −0.3643 [Cl, −1.541, 0.2098] F(1,62)=1.176, p=0.2824 |
Diff, 0.5641 [Cl, −0.1040, 1.232] F(1,62)=2.848; p=0.0965 |
Diff, 0.6590 [Cl, −0.6774, 1.995] F(1,62)=0.9716, p=0.3281 |
| input resistance (MΩ) | Diff, 22.03 [Cl, −136.2, 180.3] F(1,62)=0.08615; p=0.7749 |
Diff, 34.47 [Cl, −124.8, 191.7] F(1,62)=0.1788; p=0.6739 |
Diff, 27.80 [Cl, −288.7, 344.3] F(1,62)=0.03083; p=0.8612 |
| capacitance (pF) | Diff, −1.129 [Cl, −2.380, 0.1214] F(1,62)=3.258; p=0.0759 |
Diff, −0.03315 [Cl, −1.284, 1.217] F(1,62)=0.002808; p= 0.9579 |
Diff, 1.690 [Cl, −0.8115, 4.191] F(1,62)=1.824; p=0.1818 |
| holding current (pA) | Diff, −4.307 [Cl, −14.78, 6.163] F(1,62)=0.6762; p=0.4141 |
Diff, 6.483 [Cl, −3.986, 16.95] F(1,62)=1.532; p=0.2204 |
Diff, −11.96 [Cl, −32.90, 8.890] F(1,62)=1.303; p=0.2580 |
Voltage-dependent properties of the transient potassium current change with development and PNA treatment.
First, we characterized the amplitude, density, and voltage-dependences of activation and inactivation of the transient current (Figure 3, Table 3). Figure 3A shows representative traces. Neither development nor PNA treatment alter the maximum peak current measured in GnRH neurons at −10 mV (Figure 3B; two-way ANOVA/Fishers least significant difference (LSD). There was, however, an interaction between development and PNA treatment on transient current density. This difference appears to be driven by a PNA-induced increase in this parameter at 3 weeks of age (Figure 3C two-way ANOVA/Fisher’s LSD). Figure 3D shows the normalized current inactivation (left) and normalized conductance activation (right) as a function of voltage for the transient current. The V0.5 of activation and V0.5 of inactivation were more depolarized in cells from 3-week-old than adult control mice (Figures 3E, 3F; two-way ANOVA/Fisher’s LSD). PNA treatment hyperpolarized the V0.5 of inactivation at 3 weeks of age. Neither development nor PNA treatment altered the slope factor of activation or inactivation of the transient current (Figures 3G, 3H; two-way ANOVA/Fisher’s LSD).
Figure 3.
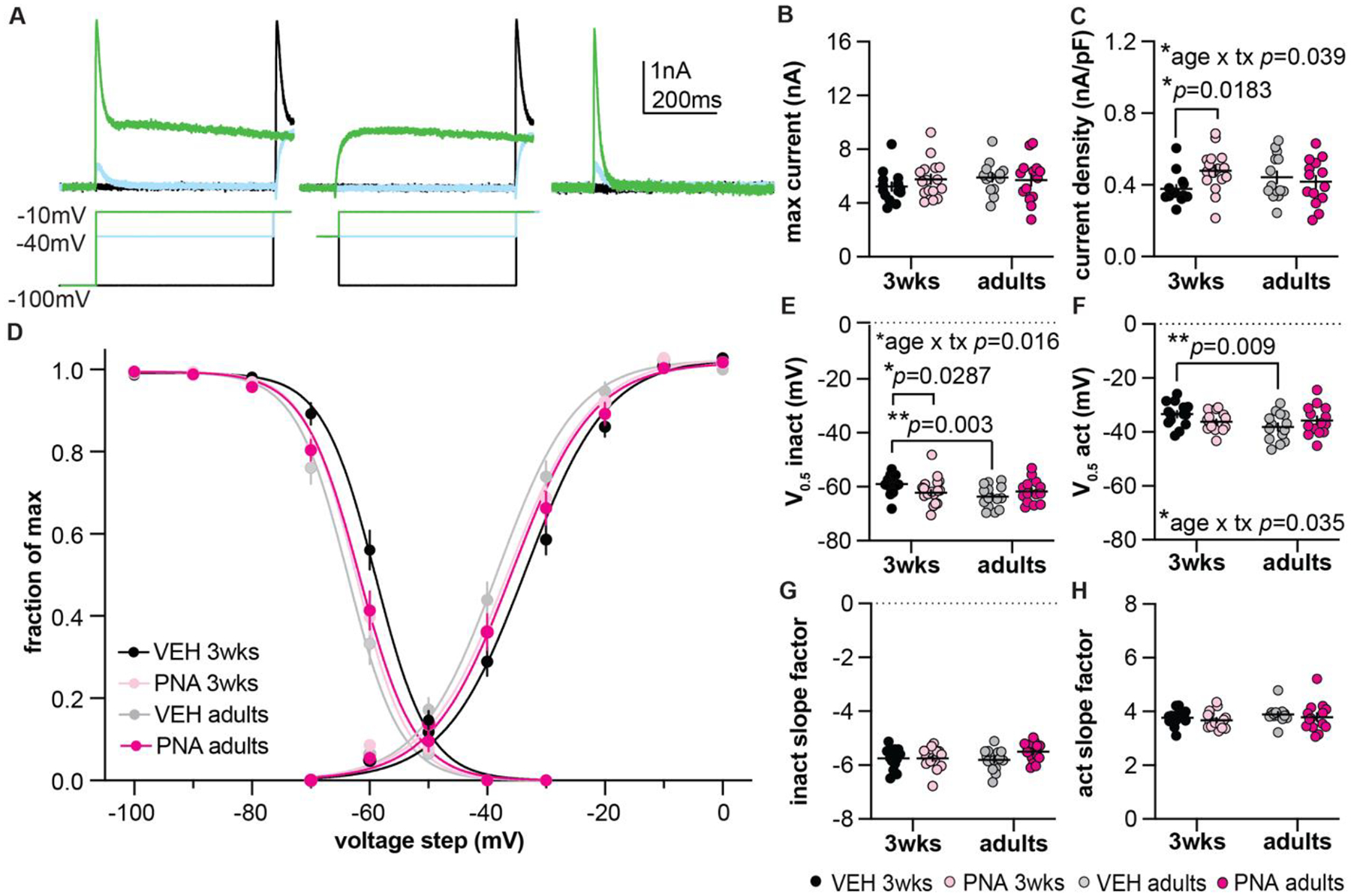
Characterization of the transient potassium current. A, Representative traces illustrating mathematical isolation of the transient current (right) by subtracting the −40mV prepulse traces in the middle panel from the −100mV prepulse traces in the left panel. Only three voltage-steps are shown for clarity. B, C, Individual values and mean±SEM for maximum current (B) and current density (C). D, Voltage-dependence of inactivation (left) and activation (right). Fraction of maximum refers to normalized current for inactivation and normalized conductance for activation. Boltzmann fits of the mean data are shown for each group. E-H, Individual values and mean±SEM V0.5 activation (V0.5 act, E), V0.5 inactivation (V0.5 inact, F), inactivation slope factor (G), and activation slope factor (H). For B, C and E-H, two-way ANOVA/Fisher’s LSD; statistical parameters are in Table 3; tx = treatment. Error bars are often within the size of the symbol.
Table 3.
Descriptive statistics and statistical parameters from two-way ANOVA/Fisher’s LSD for the voltage-dependent properties of the transient current.
| Descriptive Statistics (mean ± SEM) | ||||
|---|---|---|---|---|
| Property | VEH 3-weeks | PNA 3-weeks | VEH adults | PNA adults |
| V0.5 activation (mV) | −33.427 ± 1.181 | −36.269 ± 0.841 | −38.167 ± 1.367 | −35.744 ± 1.464 |
| Activation slope factor | 3.765 ± 0.295 | 3.674 ± 0.320 | 3.888 ± 0.321 | 3.789 ± 0.543 |
| V0.5 inactivation (mV) | −58.976 ± 0.005 | −62.269 ± 0.942 | −63.671 ± 1.119 | −61.736 ± 1.158 |
| Inactivation slope factor | −5.750 ± 0.103 | −5.750 ± 0.095 | −5.804 ± 0.106 | −5.504 ± 0.095 |
| Maximum current (pA) | 5.222 ± 0.314 | 5.771 ± 0.346 | 5.901 ± 0.310 | 5.710 ± 0.432 |
| Maximum current density (pA/pF) | 0.377 ± 0.022 | 0.479 ± 0.029 | 0.442 ± 0.033 | 0.417 ± 0.034 |
| Two-way ANOVA | |||||
|---|---|---|---|---|---|
| Property | Age | Treatment | Interaction | ||
| V0.5 activation (mV) | Diff, 2.108 [Cl, −0.3297, 4.564] F(1,54) =3.006 ; p=0.0887 |
Diff, 0.2094 [Cl, −2.228, 2.647] F(1,54) = 0.029965; p=0.8638 |
Diff, 5.265 [Cl, 0.3891, 10.14] F(1,54) = 4.687; p=0.0348 |
||
| Fisher’s LSD | 3-week VEH versus 3-week PNA | 3-week VEH versus adult VEH | 3-week PNA versus adult PNA | adult VEH versus adult PNA | |
| p=0.0988 | p=0.0089 | p=0.7578 | p=0.1712 | ||
| Activation slope factor | Diff, −0.1189 [Cl, −0.3199, 0.08204] F(1,54) = 1.408; p=0.2406 |
Diff, 0.09526 [Cl, −0.1057, 0.2962] F(1,54) = 0.9030, p=0.3462 |
Diff, −0.007502 [Cl, −0.3944, 0.4094] F(1,54) = 0.0014, p=0.9703 |
||
| V0.5 inactivation (mV) | Diff, 2.081 [Cl, −0.0300, 4.192] F(1,54) = 3.906; p=0.0532 |
Diff, 0.6790 [Cl, −1432, 2.790] F(1,54) = 0.4158; p=0.5218 |
Diff, 5.228 [Cl, 1.005, 9.450] F(1,43) = 6.162; p=0.0162 |
||
| Fisher’s LSD | 3-week VEH versus 3-week PNA | 3-week VEH versus adult VEH | 3-week PNA versus adult PNA | adult VEH versus adult PNA | |
| p=0.0287 | p=0.0030 | p=0.7175 | p=0.2064 | ||
| Inactivation slope factor | Diff, −0.09627 [Cl, −0.2965, 0.1040] F(1,54) = 0.9292, p=0.3394 |
Diff, −0.1496 [Cl, −0.3498, 0.05064] F(1,54) = 2.243, p=0.1400 |
Diff, 0.2993 [Cl, −0.1011, 0.6998] F(1,54) = 2.246, p=0.1398 |
||
| Maximum current (pA) | Diff, −0.3096 [Cl, −01.020, 0.4009] F(1,54) = 0.7634, p=0.3861 |
Diff, −0.1793 [Cl, −0.8898, 0.5312] F(1,53) = 0.2559, p=0.6150 |
Diff, −0.7400 [Cl, −2.161, 0.6810] F(1,54) = 1.0909, p=0.3011 |
||
| Maximum current density (pA/pF) | Diff, 0–0.0018 [Cl, −0.06185, 0.05832] F(1,54) = 0.0035, p=0.9532 |
Diff, −0.03836 [Cl, −0.09844, 0.02172] F(1,54) = 1.6838, p=0.2060 |
Diff, −0.1283 [Cl, −0.2464, −0.0061] F(1,54) = 4.439, p=0.0398 |
||
| Fisher’s LSD | 3-week VEH versus 3-week PNA | 3-week VEH versus adult VEH | 3-week PNA versus adult PNA | adult VEH versus adult PNA | |
| p=0.0183 | p=0.1376 | p=0.1468 | p=0.5674 | ||
Bold indicates p<0.05.
Time-dependence of inactivation and recovery from inactivation of the transient potassium current change with development and PNA treatment.
Next, we characterized the time-dependence of inactivation (Figure 4A–D) and recovery from inactivation (Figure 4E–H) of the transient current (Table 4). Representative traces are in Figures 4A and E, and the time-dependence of inactivation and removal of inactivation in Figures 4B and F. Neither age nor PNA treatment altered the fast time constant of inactivation (Figures 4C, D). In contrast, there was an increase in the fast time constant for recovery in PNA adults compared to both 3-week-old PNA mice and adult controls (Figures 4G, H; two-way ANOVA/Fisher’s LSD). Neither the slow time constant for inactivation nor recovery were affected by any treatment.
Figure 4.

Characterization of the time dependence of inactivation and recovery from inactivation of the transient current in GnRH neurons. A, E, Representative traces illustrating the time dependence of transient current inactivation (A) and recovery from inactivation (E). Only three voltage-steps are shown for clarity (bottom). B, F, Current was normalized to the maximum and plotted as a function of time (B, inactivation; F, recovery). The solid lines represent the double exponential equation evaluated using the mean fast and slow time constants for each group. C, D, G, H, Individual values and mean±SEM for the fast time constant of inactivation (inact; C), the slow time constant of inactivation (inact; D), the fast time constant of recovery (recov; G), and the slow time constant of recovery (recov; H). C, D, G, H, two-way ANOVA/Fisher’s LSD; statistical parameters are in Table 4.
Table 4.
Descriptive statistics and statistical parameters from two-way ANOVA/Fisher’s LSD for the time-dependence of inactivation and recovery from inactivation for the transient current.
| Descriptive Statistics (mean ± SEM) | ||||
|---|---|---|---|---|
| Property | VEH 3-weeks | PNA 3-weeks | VEH adults | PNA adults |
| fast inactivation time constant (ms) | 61.065 ± 4.456 | 54.449 ± 5.563 | 48.896 ± 4.670 | 56.888 ± 5.254 |
| slow inactivation time constant (ms) | 439.708 ± 29.126 | 325.946 ± 61.406 | 329.105 ± 64.934 | 301.855 ± 46.740 |
| fast recovery time constant (ms) | 14.802 ± 0.682 | 16.295 ± 1.039 | 16.088 ± 0.978 | 20.023 ± 1.389 |
| slow recovery time constant (ms) | 61.596 ± 13.174 | 73.841 ± 26.838 | 62.848 ± 11.798 | 92.188 ± 19.666 |
| Two-way ANOVA | ||||||
|---|---|---|---|---|---|---|
| Property | Age | Treatment | Interaction | |||
| fast inactivation time constant (ms) | Diff, 2.185 [Cl, −7.626, 12.00] F(1,45)=0.2102; p=0.6559 |
Diff, 1.992 [Cl, −7.819, 11.80] F(1,45)=0.1672; p=0.6845 |
Diff, 19.97 [Cl, 0.3468, 39.59] F(1,45)=4.201; p=0.0462 |
|||
| Fisher’s LSD | 3-week VEH versus 3-week PNA | 3-week VEH versus VEH adult | 3-week PNA versus PNA adult | VEH adult versus PNA adult | ||
| p=0.0854 | p=0.0876 | p=0.2581 | p=0.2574 | |||
| slow inactivation time constant (ms) | Diff, 67.35 [Cl, −35.48,173.5] F(1,45)=1.734; p=0.1945 |
Diff, 70.51 [Cl, −32.49, 173.5] F(1,45)=1.901; p=0.1748 |
Diff, 86.51 [Cl, −119.5,292.5] F(1,45)=0.7155; p=0.4021 |
|||
| fast recovery time constant (ms) | Diff, −2.507 [Cl, −4.602,−0.4113] F(1,45)=5.805; p=0.0201 |
Diff, −2.714 [Cl,−4.810,−0.6188] F(1,45)=6.806; p=0.0123 |
Diff, 2.443 [Cl, −1.748, 6.634] F(1,45)=1.379; p=0.2465 |
|||
| Fisher’s LSD | 3-week VEH versus 3-week PNA | 3-week VEH versus VEH adult | 3-week PNA versus PNA adult | VEH adult versus PNA adult | ||
| p=0.3110 | p=0.3824 | p=0.0158 | p=0.0111 | |||
| slow recovery time constant (ms) | Diff, −9.799 [Cl, −47.37,27.77] F(1,45)=0.2760; p=0.6019 |
Diff, −20.79 [Cl, −58.36,16.78] F(1,45)=1.242; p=0.2709 |
Diff, 17.10 [Cl, −58,04, 92.24] F(1,45)=0.2100; p=0.6490 |
|||
Bold indicates p<0.05.
Development and PNA treatment interact to alter the voltage-dependence of activation of the residual current.
We next tested if development and/or PNA treatment alter the residual current (Figure 5A). Neither maximum peak current nor current density were altered by development or PNA treatment alone (Figures 5B, C; two-way ANOVA, Table 5). Figure 5D shows the normalized conductance as a function of voltage estimated from the residual current. There was an age by treatment interaction on the V0.5 of activation. This was attributable to a development-related hyperpolarization of V0.5 activation in control mice and a PNA-induced depolarization of V0.5 activation in adult mice (Figure 5E). Neither development nor PNA treatment altered the slope factor of activation of the residual current (Figure 5F; two-way ANOVA).
Figure 5.

Characterization of the residual potassium current in GnRH neurons. A, Representative traces illustrating the activation of the residual current (top). Only three voltage-steps are shown for clarity (bottom). Individual values and mean±SEM for maximum current at −10 mV. (B), current density (C). D, Voltage-dependence of activation plotted as fraction of maximum conductance; lines are Boltzmann fits of the mean data for each group. Individual values and mean±SEM for V0.5 activation (V0.5 act, E), and activation (act) slope factor (F). B, C, E, F two-way ANOVA/Fisher’s LSD. Statistical parameters are in Table 5.
Table 5.
Descriptive statistics and statistical parameters from two-way ANOVA/Fisher’s LSD for the voltage-gated properties of the residual current.
| Descriptive Statistics (mean ± SEM) | ||||
|---|---|---|---|---|
| Property | VEH 3-weeks | PNA 3-weeks | VEH adults | PNA adults |
| V0.5 activation (mV) | −8.352 ± 0.905 | −10.842 ± 0.937 | −11.838 ± 1.423 | −8.193 ± 1.172 |
| Activation slope factor | 3.724 ± 0.061 | 3.850 ± 0.077 | 3.632 ± 0.097 | 3.640 ± 0.137 |
| Maximum current (pA) | 4.337 ± 0.248 | 4.492 ± 0.224 | 4.731 ± 0.318 | 4.769 ± 0.236 |
| Maximum. current density (pA/pF) | 0.313 ± 0.017 | 0.376 ± 0.024 | 0.351 ± 0.027 | 0.329 ± 0.023 |
| Two-way ANOVA | |||||
|---|---|---|---|---|---|
| Property | Age | Treatment | Interaction | ||
| V0.5 activation (mV) | Diff, 0.4182 [Cl, −1.826, 2.663] F(1,54) = 1396, p=0.702 |
Diff, −0.5773 [Cl, −2.882, 1.667] F(1,54) = 0.2660, p=0.6082 |
Diff, 6.135 [Cl, 1.646, 10.62] F(1,54) = 7.508, p=0.0083 |
||
| Fisher’s LSD | 3-week VEH versus 3-week PNA | 3-week VEH versus adult VEH | 3-week PNA versus adult PNA | adult VEH versus adult PNA | |
| p=0.1157 | p=0.0347 | p=0.0947 | p=0.0275 | ||
| Activation slope factor | Diff, 0.1510 [Cl, −0.04228, 0.3444] F(1,54) = 2.454, p=0.1231 |
Diff, −0.06698 [Cl, −0.2603, 0.1264] F(1,54) = 0.4824, p=0.4903 |
Diff, −0.1181 [Cl, −0.5047, 0.2686] F(1,54) = 0.3749, p=0.5429 |
||
| Maximum current (pA) | Diff, −0.3357 [Cl, −0.8520, 0.1806] F(1,54) = 01.700,p=0.1979 |
Diff, −0.09675 [Cl, −0.6131, 0.4195] F(1,54) = 0.1412, p=0.7086 |
Diff, −0.1177 [Cl, −1.150, 0.9149] F(1,54) = 0.05218, p=0.8202 |
||
| Maximum. current density (pA/pF) | Diff, 0.004584 [Cl, −0.04177, 0.05094] F(1,54) = 0.0393, p=0.8436 |
Diff, −0.02063 [Cl, −0.06698, 0.02573] F(1,54) = 7961, p=0.3762 |
Diff, −0.08477 [Cl, −0.1775, 0.0079939] F(1,54) = 3.361, p=0.0723 |
||
Bold indicates p<0.05.
Discussion
The pattern of GnRH/LH release can change in ways that promote reproduction, such as through the pubertal transition, and in ways that impair reproduction, such as persistent high-frequency release in women with PCOS. Changes in GnRH neuron firing rate are likely to underlie changes in GnRH release46, and potassium currents play a major role in sculpting neuronal firing activity29–32. Here we demonstrate that changes in voltage-dependent potassium currents of GnRH neurons occur during pubertal development and that PNA treatment further alter some characteristics of these currents.
GnRH neurons express a fast transient potassium current33,35 that responds to subthreshold changes in voltage and plays a role in regulating neuronal excitability and firing properties47. Transient current density was increased in GnRH neurons from PNA mice at 3-weeks of age compared to 3-week-old controls. This finding extends prior work in this PNA model that showed that GnRH neurons from PNA females have a lower spontaneous firing rate than controls at 3-weeks of age by providing a possible mechanistic explanation; greater transient potassium current density typically resists depolarization48, which may lead to cells initiating firing bouts less frequently in the absence of other changes. Further, the increased transient current density in 3-week-old PNA females could contribute to the blunted firing response to exogenous GABA exposure observed in GnRH neurons from PNA mice compared to controls at 3-weeks of age. Of interest, the increased transient current density in 3-week-old PNA females may be a compensatory mechanism initiated in GnRH neurons by the increased excitatory GABAergic neurotransmission at this age as a way of providing neuroprotection in PNA mice49–52.
In addition to current density, the voltage and time dependence of transient potassium currents help sculpt firing in excitable cells. Regarding the former, a clear development-associated hyperpolarization of both the inactivation and activation of the transient current occur in control mice. Hyperpolarization of inactivation in adults would tend to make less current available in the subthreshold range, perhaps contributing to increased excitability of adult GnRH neurons compared to those from prepubertal mice28. The hyperpolarization of activation in adult controls may sculpt aspects of the action potential spike, including the increased afterhyperpolarization potential28. In PNA females, there were no changes in either voltage-dependence of inactivation or activation with development. In contrast to the effects of development on voltage dependence of inactivation, PNA treatment had a more marked effect on its time dependence. Most notably, recovery from inactivation took longer in PNA adults in comparison to other groups. This may be a contributing factor to the increased spontaneous activity observed in GnRH neurons from PNA mice in adulthood23. Together these data indicate changes in transient current properties occur concomitant with pubertal development, and that prenatal androgen exposure both disrupts typical developmental changes in this current and induces an independent shift in properties.
The speculative attribution of the effects of specific properties of the transient current to prior observations in GnRH neuron physiology in these same animal models is done with caution for several reasons. First, different properties of the same macroscopic current can have different predicted effects on neuronal output in the same animal model. For example, changes in voltage-dependence of inactivation of the transient current from 3-weeks of age to adults in control mice might increase excitability, whereas the lack of difference in transient current density between these same groups might predict no change. Second, the direction of change induced by a particular property in isolation is a postulate. For example, increased transient current can be associated with reduced excitability35 but can also induce membrane potential changes that enhance recovery from inactivation of the fast sodium current and thus increase firing rate53. Third, the effects of the transient potassium current on firing and excitability do not occur in isolation but are integrated with changes in other GnRH neuron intrinsic properties that may occur as the animal goes through puberty and/or in response to PNA treatment. In this regard, we observed age-dependent changes in the activation of the residual potassium current in controls but PNA treatment-dependent shifts in this property in adults. Activation of this current typically occurs at the depolarized membrane potentials achieved during an action potential. The residual current is important in repolarizing the membrane, sculpting action potential shape54,55 and determining the generation of subsequent action potentials in a burst54. In adult control females, hyperpolarized residual current activation might contribute to reduced spontaneous firing activity of GnRH neurons in the group compared to 3-week-old controls and to adult PNA females. Fourth, there are documented changes in GABAergic fast synaptic input with age and PNA treatment that will also contribute to determining the overall firing rate of these cells17,24. Finally, other factors, such as neuromodulator input, may also differ among groups. Of interest in this regard, no difference was observed in the spontaneous firing rate of arcuate kisspeptin neurons among these same four groups56, but interactions with other neuromodulatory neurons and with non-neuronal cells could alter GnRH neuron output in a PNA-treatment and/or age-dependent manner. Computational modeling would be one way to address these interactions, but there are no data on the other intrinsic properties of GnRH neurons in these animal models, limiting the utility of this approach at present.
The observed changes in the voltage- and time-dependent properties of potassium currents in this study extend work in other brain regions indicating these currents change across development. For example, in acutely-dissociated large aspiny neurons, transient current inactivation is depolarized and occurs faster with increasing age; this may contribute to reduced spike latency in mature versus young neurons57. Developmental changes in voltage-dependent potassium currents are associated with establishing reliable spiking in the calyx of Held58. Potassium current alterations can also underlie neuropathological conditions, including pain59, neurodegeneration60, and epilepsy61. Within the reproductive neuroendocrine system, changes in potassium currents, in particular the transient current, are associated with changes in GnRH and arcuate kisspeptin neuron physiology. In a model for estradiol positive feedback in which GnRH neurons are more active and more excitable62,63, transient current density is suppressed and other properties shifted in comparison to a gonadectomized open feedback loop condition35. In contrast, no differences were found between the open loop and a model of negative feedback; there were also no differences in excitability between these groups33. Estradiol also reduces transient current amplitude in arcuate kisspeptin neurons; in these cells, reducing this current enhances synaptic response and is associated with altered firing activity64. Of note, most prior studies on GnRH and kisspeptin neuron currents were done in adults, thus no developmental data are available. The steroid regulation of these currents is nonetheless applicable to the present work. The increased production of sex steroids that occurs with puberty might be the catalyst behind the developmental change observed in potassium currents observed in cells from control mice, as well as excitability and the GnRH neuron transcriptome28,45. Steroidogenesis is also altered in adult PNA mice, which have increased testosterone levels17,21,65. While many studies have examined estrogen effects on potassium currents in neuronal populations33,35,55,64,66, we found no studies of androgen effects on central neurons. Work in the cardiovascular system suggests androgen-associated changes in potassium currents. Cardiovascular disease, including arrhythmias, are more common in men67. Electrocardiogram QTc intervals are similar among prepubertal males and female and adult females, whereas adult males have shorter QTc intervals68–70. In rat cardiomyocytes, dihydrotestosterone (DHT), a non-aromatizable androgen, upregulates the delayed-rectified potassium channel and shortens the electrocardiogram QT interval, which correspond to the action potential duration in ventricular cardiomyocytes, compared to castrated males without DHT treatment71. In immortalized pituitary GH3 cells, androgens rapidly and reversibly block a sustained current72, and in cancer cells, voltage-gated potassium currents are associated with cell proliferation73. These studies support the postulate that age and PNA-induced changes in sex steroid milieu in the present studies could be the catalysts underlying changes observed in voltage-gated potassium currents.
The changes in voltage-dependent potassium currents in GnRH neurons described in this study likely play a role in shaping the activity of GnRH neurons through typical pubertal development and the changes induced by prenatal androgen exposure. Untangling these underlying mechanisms has implications for increased GnRH/LH release frequency and altered pubertal progression in hyperandrogenemic adolescents and individuals with PCOS.
Acknowledgements:
We thank Laura Burger, J. Rudolph Starrett and Elizabeth Wagenmaker for expert technical assistance, and Laura Burger, Xi Chen, Amanda Gibson, J. Rudolph Starrett and Elizabeth Wagenmaker, for editorial comments.
Grant Support:
National Institute of Health (NIH)/Eunice Kennedy Shriver National Institute of Child Health and Human Development Grant R01HD104345 (to S.M.M.). Support for J.J. was provided by NIH T32HD079342 and F31HD110279.
Abbreviations:
- GnRH
Gonadotropin-releasing hormone
- PNA
prenatal androgenization
- LH
luteinizing hormone
- FSH
follicle stimulating hormone
- PCOS
polycystic ovary syndrome
- DHT
dihydrotestosterone
- GABA
gamma aminobutyric acid
- GFP
green-fluorescent protein
- VO
vaginal opening
- AGD
anogenital distance
- LSD
least significant difference
- tx
treatment
- act
activation
- inact
inactivation
- recov
recovery
Footnotes
Conflict of interest: The authors declare no competing financial interests.
Literature cited
- 1.Clarke IJ & Cummins JT The temporal relationship between gonadotropin releasing hormone (GnRH) and luteinizing hormone (LH) secretion in ovariectomized ewes. Endocrinology 111, 1737–1739 (1982). https://doi.org: 10.1210/endo-111-5-1737 [DOI] [PubMed] [Google Scholar]
- 2.Moenter SM, Brand RM, Midgley AR & Karsch FJ Dynamics of gonadotropin-releasing hormone release during a pulse. Endocrinology 130, 503–510 (1992). https://doi.org: 10.1210/endo.130.1.1727719 [DOI] [PubMed] [Google Scholar]
- 3.Wildt L et al. Frequency and amplitude of gonadotropin-releasing hormone stimulation and gonadotropin secretion in the rhesus monkey. Endocrinology 109, 376–385 (1981). https://doi.org: 10.1210/endo-109-2-376 [DOI] [PubMed] [Google Scholar]
- 4.Haisenleder DJ, Dalkin AC, Ortolano GA, Marshall JC & Shupnik MA A pulsatile gonadotropin-releasing hormone stimulus is required to increase transcription of the gonadotropin subunit genes: evidence for differential regulation of transcription by pulse frequency in vivo. Endocrinology 128, 509–517 (1991). https://doi.org: 10.1210/endo-128-1-509 [DOI] [PubMed] [Google Scholar]
- 5.Legro RS Diagnostic criteria in polycystic ovary syndrome. Semin Reprod Med 21, 267–275 (2003). https://doi.org: 10.1055/s-2003-43304 [DOI] [PubMed] [Google Scholar]
- 6.Tsutsumi R & Webster NJ GnRH pulsatility, the pituitary response and reproductive dysfunction. Endocr J 56, 729–737 (2009). https://doi.org: 10.1507/endocrj.k09e-185 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sidhoum VF et al. Reversal and relapse of hypogonadotropic hypogonadism: resilience and fragility of the reproductive neuroendocrine system. J Clin Endocrinol Metab 99, 861–870 (2014). https://doi.org: 10.1210/jc.2013-2809 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Burt Solorzano CM et al. Neuroendocrine dysfunction in polycystic ovary syndrome. Steroids 77, 332–337 (2012). https://doi.org: 10.1016/j.steroids.2011.12.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.McCartney CR, Campbell RE, Marshall JC & Moenter SM The role of gonadotropin-releasing hormone neurons in polycystic ovary syndrome. J Neuroendocrinol 34, e13093 (2022). https://doi.org: 10.1111/jne.13093 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Yen SS, Vela P & Rankin J Inappropriate secretion of follicle-stimulating hormone and luteinizing hormone in polycystic ovarian disease. J Clin Endocrinol Metab 30, 435–442 (1970). https://doi.org: 10.1210/jcem-30-4-435 [DOI] [PubMed] [Google Scholar]
- 11.Kazer RR, Kessel B & Yen SS Circulating luteinizing hormone pulse frequency in women with polycystic ovary syndrome. J Clin Endocrinol Metab 65, 233–236 (1987). https://doi.org: 10.1210/jcem-65-2-233 [DOI] [PubMed] [Google Scholar]
- 12.Broekmans FJ & Fauser BC Diagnostic criteria for polycystic ovarian syndrome. Endocrine 30, 3–11 (2006). https://doi.org: 10.1385/ENDO:30:1:3 [DOI] [PubMed] [Google Scholar]
- 13.Apter D, Bützow T, Laughlin GA & Yen SS Accelerated 24-hour luteinizing hormone pulsatile activity in adolescent girls with ovarian hyperandrogenism: relevance to the developmental phase of polycystic ovarian syndrome. J Clin Endocrinol Metab 79, 119–125 (1994). https://doi.org: 10.1210/jcem.79.1.8027216 [DOI] [PubMed] [Google Scholar]
- 14.Yoo RY et al. Increased luteinizing hormone pulse frequency in obese oligomenorrheic girls with no evidence of hyperandrogenism. Fertil Steril 85, 1049–1056 (2006). https://doi.org: 10.1016/j.fertnstert.2005.09.037 [DOI] [PubMed] [Google Scholar]
- 15.Barber TM & Franks S Genetics of polycystic ovary syndrome. Front Horm Res 40, 28–39 (2013). https://doi.org: 10.1159/000341682 [DOI] [PubMed] [Google Scholar]
- 16.Dapas M et al. Distinct subtypes of polycystic ovary syndrome with novel genetic associations: An unsupervised, phenotypic clustering analysis. PLoS Med 17, e1003132 (2020). https://doi.org: 10.1371/journal.pmed.1003132 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Sullivan SD & Moenter SM Prenatal androgens alter GABAergic drive to gonadotropin-releasing hormone neurons: implications for a common fertility disorder. Proc Natl Acad Sci U S A 101, 7129–7134 (2004). https://doi.org: 10.1073/pnas.0308058101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Foecking EM, Szabo M, Schwartz NB & Levine JE Neuroendocrine consequences of prenatal androgen exposure in the female rat: absence of luteinizing hormone surges, suppression of progesterone receptor gene expression, and acceleration of the gonadotropin-releasing hormone pulse generator. Biol Reprod 72, 1475–1483 (2005). https://doi.org: 10.1095/biolreprod.105.039800 [DOI] [PubMed] [Google Scholar]
- 19.Mahoney MM & Padmanabhan V Developmental programming: impact of fetal exposure to endocrine-disrupting chemicals on gonadotropin-releasing hormone and estrogen receptor mRNA in sheep hypothalamus. Toxicol Appl Pharmacol 247, 98–104 (2010). https://doi.org: 10.1016/j.taap.2010.05.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Roland AV & Moenter SM Prenatal androgenization of female mice programs an increase in firing activity of gonadotropin-releasing hormone (GnRH) neurons that is reversed by metformin treatment in adulthood. Endocrinology 152, 618–628 (2011). https://doi.org: 10.1210/en.2010-0823 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Witham EA, Meadows JD, Shojaei S, Kauffman AS & Mellon PL Prenatal exposure to low levels of androgen accelerates female puberty onset and reproductive senescence in mice. Endocrinology 153, 4522–4532 (2012). https://doi.org: 10.1210/en.2012-1283 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Moore AM, Prescott M, Marshall CJ, Yip SH & Campbell RE Enhancement of a robust arcuate GABAergic input to gonadotropin-releasing hormone neurons in a model of polycystic ovarian syndrome. Proc Natl Acad Sci U S A 112, 596–601 (2015). https://doi.org: 10.1073/pnas.1415038112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Dulka EA & Moenter SM Prepubertal Development of Gonadotropin-Releasing Hormone Neuron Activity Is Altered by Sex, Age, and Prenatal Androgen Exposure. Endocrinology 158, 3943–3953 (2017). https://doi.org: 10.1210/en.2017-00768 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Berg T, Silveira MA & Moenter SM Prepubertal Development of GABAergic Transmission to Gonadotropin-Releasing Hormone (GnRH) Neurons and Postsynaptic Response Are Altered by Prenatal Androgenization. J Neurosci 38, 2283–2293 (2018). https://doi.org: 10.1523/JNEUROSCI.2304-17.2018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Caligioni CS Assessing reproductive status/stages in mice. Curr Protoc Neurosci Appendix 4, Appendix 4I (2009). https://doi.org: 10.1002/0471142301.nsa04is48 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.DeFazio RA, Heger S, Ojeda SR & Moenter SM Activation of A-type gamma-aminobutyric acid receptors excites gonadotropin-releasing hormone neurons. Mol Endocrinol 16, 2872–2891 (2002). https://doi.org: 10.1210/me.2002-0163 [DOI] [PubMed] [Google Scholar]
- 27.Herbison AE & Moenter SM Depolarising and hyperpolarising actions of GABA(A) receptor activation on gonadotrophin-releasing hormone neurones: towards an emerging consensus. J Neuroendocrinol 23, 557–569 (2011). https://doi.org: 10.1111/j.1365-2826.2011.02145.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jaime J & Moenter SM GnRH neuron excitability and action potential properties change with development but are not affected by prenatal androgen exposure. eNeuro 9 (2022). https://doi.org: 10.1523/ENEURO.0362-22.2022 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Feria Pliego JA & Pedroarena CM Kv1 potassium channels control action potential firing of putative GABAergic deep cerebellar nuclear neurons. Sci Rep 10, 6954 (2020). https://doi.org: 10.1038/s41598-020-63583-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Johnston J, Forsythe ID & Kopp-Scheinpflug C Going native: voltage-gated potassium channels controlling neuronal excitability. J Physiol 588, 3187–3200 (2010). https://doi.org: 10.1113/jphysiol.2010.191973 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Mendonça PRF, Kyle V, Yeo SH, Colledge WH & Robinson HPC Kv4.2 channel activity controls intrinsic firing dynamics of arcuate kisspeptin neurons. J Physiol 596, 885–899 (2018). https://doi.org: 10.1113/JP274474 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Storm JF Action potential repolarization and a fast after-hyperpolarization in rat hippocampal pyramidal cells. J Physiol 385, 733–759 (1987). https://doi.org: 10.1113/jphysiol.1987.sp016517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.DeFazio RA & Moenter SM Gonadotropin-Releasing Hormone (GnRH) Neuron Potassium Currents and Excitability in Both Sexes Exhibit Minimal Changes upon Removal of Negative Feedback. eNeuro 8 (2021). https://doi.org: 10.1523/ENEURO.0126-21.2021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Suter KJ et al. Genetic targeting of green fluorescent protein to gonadotropin-releasing hormone neurons: characterization of whole-cell electrophysiological properties and morphology. Endocrinology 141, 412–419 (2000). [DOI] [PubMed] [Google Scholar]
- 35.DeFazio RA & Moenter SM Estradiol feedback alters potassium currents and firing properties of gonadotropin-releasing hormone neurons. Mol Endocrinol 16, 2255–2265 (2002). https://doi.org: 10.1210/me.2002-0155 [DOI] [PubMed] [Google Scholar]
- 36.Bezanilla F & Armstrong CM Inactivation of the sodium channel. I. Sodium current experiments. J Gen Physiol 70, 549–566 (1977). https://doi.org: 10.1085/jgp.70.5.549 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Clay JR Determining K+ channel activation curves from K+ channel currents. Eur Biophys J 29, 555–557 (2000). https://doi.org: 10.1007/s002490000091 [DOI] [PubMed] [Google Scholar]
- 38.Clay JR Determining k channel activation curves from k channel currents often requires the goldman-hodgkin-katz equation. Front Cell Neurosci 3, 20 (2009). https://doi.org: 10.3389/neuro.03.020.2009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Kielland J Individual Activity Coefficients of Ions in Aqueous Solutions. Journal of the American Chemical Society 59, 1675–1678 (1937). [Google Scholar]
- 40.Verzani J, Loukides MK & Romano R Getting started with RStudio. First edition. edn, (O’Reilly, 2012). [Google Scholar]
- 41.Cohen Y & Cohen JY 1 online resource (619 p.) (Wiley, Chichester, U.K., 2008). [Google Scholar]
- 42.Pace L 1 online resource (321 p.) (Apress : Imprint: Apress, Berkeley, CA, 2012). [Google Scholar]
- 43.Kassambara A rstatix: Pipe-Friendly Framework for Basic Statistical Tests. R package version 0.7.2, <https://rpkgs.datanovia.com/rstatix/> (2023).
- 44.Gohel D flextable: functions for tabular reporting (R package version 0.5.11), <https://cran.r-project.org/package=flextable.> (2020).
- 45.Burger LL, Wagenmaker ER, Phumsatitpong C, Olson DP & Moenter SM Prenatal Androgenization Alters the Development of GnRH Neuron and Preoptic Area RNA Transcripts in Female Mice. Endocrinology 161 (2020). https://doi.org: 10.1210/endocr/bqaa166 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chen X & Moenter SM Gonadal Feedback Alters the Relationship between Action Potentials and Hormone Release in Gonadotropin-Releasing Hormone Neurons in Male Mice. J Neurosci 43, 6717–6730 (2023). https://doi.org: 10.1523/JNEUROSCI.2355-22.2023 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Noh W, Pak S, Choi G & Yang S Transient Potassium Channels: Therapeutic Targets for Brain Disorders. Front Cell Neurosci 13, 265 (2019). https://doi.org: 10.3389/fncel.2019.00265 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kita T, Kita H & Kitai ST Effects of 4-aminopyridine (4-AP) on rat neostriatal neurons in an in vitro slice preparation. Brain Res 361, 10–18 (1985). https://doi.org: 10.1016/0006-8993(85)91269-7 [DOI] [PubMed] [Google Scholar]
- 49.Misonou H Homeostatic regulation of neuronal excitability by K(+) channels in normal and diseased brains. Neuroscientist 16, 51–64 (2010). https://doi.org: 10.1177/1073858409341085 [DOI] [PubMed] [Google Scholar]
- 50.Marder E & Prinz AA Current compensation in neuronal homeostasis. Neuron 37, 2–4 (2003). https://doi.org: 10.1016/s0896-6273(02)01173-x [DOI] [PubMed] [Google Scholar]
- 51.Marder E & Goaillard JM Variability, compensation and homeostasis in neuron and network function. Nat Rev Neurosci 7, 563–574 (2006). https://doi.org: 10.1038/nrn1949 [DOI] [PubMed] [Google Scholar]
- 52.Shruti S, Clem RL & Barth AL A seizure-induced gain-of-function in BK channels is associated with elevated firing activity in neocortical pyramidal neurons. Neurobiol Dis 30, 323–330 (2008). https://doi.org: 10.1016/j.nbd.2008.02.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Klyachko VA, Ahern GP & Jackson MB cGMP-mediated facilitation in nerve terminals by enhancement of the spike afterhyperpolarization. Neuron 31, 1015–1025 (2001). https://doi.org: 10.1016/s0896-6273(01)00449-4 [DOI] [PubMed] [Google Scholar]
- 54.Manis PB in Encyclopedia of Computational Neuroscience (eds Jaeger Dieter & Jung Ranu) 1–16 (Springer; New York, 2013). [Google Scholar]
- 55.Du J et al. Effects of estradiol on voltage-gated potassium channels in mouse dorsal root ganglion neurons. J Membr Biol 247, 541–548 (2014). https://doi.org: 10.1007/s00232-014-9670-z [DOI] [PubMed] [Google Scholar]
- 56.Gibson AG, Jaime J, Burger LL & Moenter SM Prenatal androgen treatment does not alter the firing activity of hypothalamic arcuate kisspeptin neurons in female mice. eNeuro (2021). https://doi.org: 10.1523/ENEURO.0306-21.2021 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Deng P, Pang Z, Zhang Y & Xu ZC Developmental changes of transient potassium currents in large aspiny neurons in the neostriatum. Brain Res Dev Brain Res 153, 97–107 (2004). https://doi.org: 10.1016/j.devbrainres.2004.08.001 [DOI] [PubMed] [Google Scholar]
- 58.Nakamura Y & Takahashi T Developmental changes in potassium currents at the rat calyx of Held presynaptic terminal. J Physiol 581, 1101–1112 (2007). https://doi.org: 10.1113/jphysiol.2007.128702 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Wang W, Gu J, Li YQ & Tao YX Are voltage-gated sodium channels on the dorsal root ganglion involved in the development of neuropathic pain? Mol Pain 7, 16 (2011). https://doi.org: 10.1186/1744-8069-7-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Shah NH & Aizenman E Voltage-gated potassium channels at the crossroads of neuronal function, ischemic tolerance, and neurodegeneration. Transl Stroke Res 5, 38–58 (2014). https://doi.org: 10.1007/s12975-013-0297-7 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Köhling R & Wolfart J Potassium Channels in Epilepsy. Cold Spring Harb Perspect Med 6 (2016). https://doi.org: 10.1101/cshperspect.a022871 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Christian CA, Mobley JL & Moenter SM Diurnal and estradiol-dependent changes in gonadotropin-releasing hormone neuron firing activity. Proc Natl Acad Sci U S A 102, 15682–15687 (2005). https://doi.org: 10.1073/pnas.0504270102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Adams C, Stroberg W, DeFazio RA, Schnell S & Moenter SM Gonadotropin-Releasing Hormone (GnRH) Neuron Excitability Is Regulated by Estradiol Feedback and Kisspeptin. J Neurosci 38, 1249–1263 (2018). https://doi.org: 10.1523/JNEUROSCI.2988-17.2017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.DeFazio RA, Navarro MA, Adams CE, Milescu LS & Moenter SM Estradiol Enhances the Depolarizing Response to GABA and AMPA Synaptic Conductances in Arcuate Kisspeptin Neurons by Diminishing Voltage-Gated Potassium Currents. J Neurosci 39, 9532–9545 (2019). https://doi.org: 10.1523/JNEUROSCI.0378-19.2019 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Silva MSB et al. Activation of arcuate nucleus GABA neurons promotes luteinizing hormone secretion and reproductive dysfunction: Implications for polycystic ovary syndrome. EBioMedicine 44, 582–596 (2019). https://doi.org: 10.1016/j.ebiom.2019.05.065 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Power EM, Ganeshan D & Iremonger KJ Estradiol regulates voltage-gated potassium currents in corticotropin-releasing hormone neurons. J Exp Biol 226 (2023). https://doi.org: 10.1242/jeb.245222 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Makkar RR, Fromm BS, Steinman RT, Meissner MD & Lehmann MH Female gender as a risk factor for torsades de pointes associated with cardiovascular drugs. JAMA 270, 2590–2597 (1993). https://doi.org: 10.1001/jama.270.21.2590 [DOI] [PubMed] [Google Scholar]
- 68.Lehmann MH et al. Age-gender influence on the rate-corrected QT interval and the QT-heart rate relation in families with genotypically characterized long QT syndrome. J Am Coll Cardiol 29, 93–99 (1997). https://doi.org: 10.1016/s0735-1097(96)00454-8 [DOI] [PubMed] [Google Scholar]
- 69.Argenziano M et al. Arrhythmogenic effect of androgens on the rat heart. J Physiol Sci 67, 217–225 (2017). https://doi.org: 10.1007/s12576-016-0459-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Rautaharju PM et al. Sex differences in the evolution of the electrocardiographic QT interval with age. Can J Cardiol 8, 690–695 (1992). [PubMed] [Google Scholar]
- 71.Masuda K et al. Testosterone-mediated upregulation of delayed rectifier potassium channel in cardiomyocytes causes abbreviation of QT intervals in rats. J Physiol Sci 68, 759–767 (2018). https://doi.org: 10.1007/s12576-017-0590-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Suárez L, Bilal U, Bordallo J, Cantabrana B & Sánchez M Androgens block outward potassium currents and decrease spontaneous action potentials in GH3 cells. Naunyn Schmiedebergs Arch Pharmacol 388, 67–78 (2015). https://doi.org: 10.1007/s00210-014-1057-2 [DOI] [PubMed] [Google Scholar]
- 73.Rao VR, Perez-Neut M, Kaja S & Gentile S Voltage-gated ion channels in cancer cell proliferation. Cancers (Basel) 7, 849–875 (2015). https://doi.org: 10.3390/cancers7020813 [DOI] [PMC free article] [PubMed] [Google Scholar]