

ABSTRACT
The gut microbiota exerts a mutualistic interaction with the host in a fragile ecosystem and the host intestinal, neural, and immune cells. Perturbations of the gastrointestinal track composition after stress have profound consequences on the central nervous system and the immune system. Reciprocally, brain signals after stress affect the gut microbiota highlighting the bidirectional communication between the brain and the gut. Here, we focus on the potential role of inflammation in mediating stress-induced gut-brain changes and discuss the impact of several immune cells and inflammatory molecules of the gut-brain dialogue after stress. Understanding the impact of microbial changes on the immune system after stress might provide new avenues for therapy.
KEYWORDS: Stress, Th17 cells, microbiota, behaviors, immune cells, cytokines, quorum sensing molecules
Stress
Stress is a part of modern life in developed societies. Although the definition of stress varies from individual to individual, it is considered to involve external encounters including mental or mechanical challenges, which can be actual or perceived. It is accepted that a low level of stress often is beneficial for the individual boosting the immune system, whereas continuous exposure to high level of stress can have detrimental consequences for the organism, leading to mental illness or other serious medical conditions.1,2 The consequences of stress often differ from individual to individual. And it has been proposed that the timing, magnitude and duration of the encounter stress might be critical to prime individual to the potential consequences of a subsequent stress exposure.3 Thus, on one hand, childhood trauma is strongly associated with later in life development of depression, whereas on the other hand, repeated exposure to stress can be associated with resilience.
Information about the stressor is often perceived by sensory systems in periphery that relay the information to the brain. Many mediators of stress have been uncovered including the canonical hypothalamic pituitary adrenal axis (HPA)4 or the adrenaline/noradrenaline fight or flight response.5 These pathways aim at preparing the body to respond to the insult. In this review, we will focus on the effects of stress on the gut microbiome and reciprocally the effects of the gut microbiome on the stress response in the context of inflammation.
Stress affects the physiology of the entire body and significantly impacts the gut in an unpleasant way, often leading to dysbiosis, and leaky gut.
The gut microbiome is a fragile ecosystem composed of bacteria, protozoans, virus, fungus, archaea. Most research has concentrated on the bacteria, but new knowledge of virus and fungi in the context of the microbiome and health has also emerged. The definition of a healthy microbiome remains to be determined. Overall, the gut mycobiome is less taxonomically diverse and heterogeneous than the gut bacteriome and the virome between individuals. The geography accounts for 30.8%, the ethnicity for 15.6%, the diet for 9.8% and host factors and environmental factors account for the remaining variations of the microbiome variations across healthy individuals.6,7
Bacteria
The human intestine harbors ~ 100 trillion bacteria that are crucial for health,8 they provide nutrients, process complex polysaccharides from the diet, and are required for a healthy immune system and neurodevelopment.9,10 Bacteria in the gut form their own ecosystem but are also directly influenced by human physiological signals and vice versa. The composition of the gut microbiota varies among individuals but is estimated of ~ 1,000 species and > 7,000 strains. Bacteroidetes and Firmicutes are predominant, while Proteobacteria, Actinobacteria, Fusobacteria, Verrucomicrobiota and others are also present. Microbiota changes have been reported in depression (see section C).
Fungi
Fungi are an integral part of the human gut and represent a small (0.1%) but crucial component of the microbiome and although their function in the gut is not well-established, they regulate host homeostasis and pathogenesis.11,12 Fungi are early colonizers of the gut and are influenced by diet intake and environment. The mycobiome is highly variable between individuals and over time, compared to the gut microbiota.11,13 Some fungi are beneficial, such as Saccharomyces boulardii, which is used as probiotics, while others are associated with diseases such as autoimmune14–16 or neurological disorders.17,18 In diseases, it appears that the increased abundance of Candida albicans is associated with inflammatory bowel disease,14,16,19 asthma,20 liver disease,21 schizophrenia22,23 and COVID-19.24 Candida albicans is an interesting example of an interaction between a fungus and the host. Candida albicans drives anti-fungal T helper (Th) 17 cell-mediated immune responses. Mannans present in the wall of fungi are sufficient to increase Th17 differentiation and IL-23 responses, as well as toll-like receptor-, C-type lectin receptor-, and Nucleotide-binding oligomerization domain-containing protein (NOD)-like receptor-mediated anti-fungal actions.25–27 Recent findings show that the mycobiome is altered in children and adolescents with depression.28 Overall, it remains to be determined how fungi affect the host and whether their interactions with bacteria are critical for their role in diseases (e.g., fungi colonization in germ-free mice did not affect the host physiology,29 and even less is known about the role of fungi in the stress response.
Viruses
Healthy human guts are also colonized by a great variety of viruses30 including bacteriophages that infect bacteria, viruses that infect archaea, viruses that infect human cells or viruses present in the food. It seems that viruses resident of the gut do not have an envelope.
Like the bacteria and fungi, there is an inter-individual variation in the composition of the virome. Depressed patients have increased levels of Siphoviridae and Microviridae.31 Shifts in Caudovirales bacteriophage are also present in MDD,32 suggesting that stress, which is a major contributor to depression, might also affect phage biology. Although the relationship between viruses and stress is less studied than the relationship between bacteria and stress, it is accepted that stress by suppressing the immune system allows the resurging of latent viruses. Thus, for example, virus-specific antibodies and T cell responses to hepatitis B are suppressed by acute stress.33 This might underline the known positive association between psychological stress and increased risk of viral infections.34,35 But it is also possible that phages by modifying bacteria, shape bacterial communities which influence host physiology. Cocktails of phages have been used to treat bacterial infections,36 and prophages seem responsible for the effects of food (e.g., sweeteners) in inhibiting bacterial growth for example.37
Because stress is dependent on various factors and individual physiology is different, it is difficult to draw the line between beneficial and detrimental effects of stress for each individual, which might explain the complexity of the effects of stress on the gut-brain axis.
Stress and dysbiosis
Dysbiosis is defined as changes of the bacterial composition, bacterial metabolism, or bacterial distribution. This can translate into a loss of beneficial bacteria, an outgrowth of pathogenic bacteria, an overall reduction of bacterial diversity or any combination of those,38 and a loss of bacterial diversity is often considered harmful to the host.
After stress, the blood including the immune system, and the autonomic systems brings stress signals to the gut that favor the bloom of pathogenic bacteria that promote dysbiosis and gut barrier permeability.39 Thus, stressed people have unpredictable dysbiosis,40 less health beneficial gut bacteria as exemplified in university students after chronic stress,41 and increased gut permeability after the speech stressor test.42 Furthermore, couples with hostile relationships have greater gut leakiness than couples with less hostile relationships,43 suggesting that chronic stress can induce detrimental effects on the gut and the microbiome. In addition, stress affects multiple regions and habitats of the intestine within the lumen and the mucosal lining of the gut, showing a wide effect of stress in the gut.
The causes of dysbiosis are multiple and involve host factors such as genetics, health issues (e.g., inflammation or infections), lifestyle and environmental factors (e.g., diet, medications, hygiene). Diet is one of the major factor contributing to dysbiosis, and is affected by stress, offering a way whereby stress promotes dysbiosis. As an example, diet enriched in simple sugars is sufficient to disrupt gut permeability, induce inflammation and negatively change the host metabolism, and these effects are dependent on the microbiota as microbial depletion abolishes these effects.44 It is in the common belief that people eat sugary treats when feeling stressed. Negative emotion is associated with increased taste-based eating45 and unhealthy eating is promoted by mild stressors.46 For example, cities whose NFL football team lost on Sunday saw an increased intake of saturated fat on Monday compared to a decrease in cities whose team won, and no change in cities whose NFL team did not play, or without a NFL team.46 It has been proposed that the executive function in response to food cues is deactivated after stress favoring comfort foods.47 Reciprocally, high consumption of sugars has been associated with cognitive impairments, amplifying the unhealthy eating induced by stress. In addition to the diet, stress also affects the host metabolism, which also impacts the gut microbiome.48
The change of gut permeability after stress is amplified by inflammation, since the loss of gut permeability after stress allows the entry of commensal and pathogenic bacteria and food antigens to the lamina propria inducing local inflammation which often propagates to the blood circulation, inducing systemic inflammation.49 However, the mechanisms, whereby stress promotes dysbiosis remain mainly unknown.
Stress hormones and the gut
Stress hormones (i.e., corticosterone metabolites, catecholamines) have been found in the stools after various stressors.50 Stressed germ-free mice have increased level of corticosterone even though the basal level of corticosterone is unchanged compared to specific pathogen free mice in unstressed mice.51–54 Stress hormones are released into the gut from the blood, but some are also produced in the gut by the enteric neurons, the gut epithelial cells or the enterochromaffin cells.55 Stress hormones directly affect bacteria level. For example, the serum level of cortisol is a negative predictor of the level of Rumunococcus spp. in pigs.56 Similarly, saliva level of cortisol is negatively correlated with the alpha and beta diversity of the gut microbiota in humans after stress57. Catecholamines increase the bacterial growth of certain gram-negative strains55 to the point that certain bacteria levels are increased by a 10,000 fold and infectiousness increases within 14 h in vitro.57 Various studies have shown the effects of stress hormones on the gut microbes.55,56,58,59 HPA activation such as maternal separation in rhesus monkeys or early-life stress in rats results in a reduction of the Lactobacilli Gram-negative bacteria,60,61 and a change of the composition of the gut microbiota,62,63 respectively.
Although glucocorticoid receptors have not been identified on bacteria, catecholamines receptors (QseBC) are present on certain bacteria, such as E. coli,64 suggesting that the effects of stress hormones can directly influence bacterial metabolism (Figure 3). Indeed, in response to catecholamines, QseBC induces the transcription of flagellar regulon in E. coli.65 Cortisol also impacts the community-wide transcriptome of the oral microbiome.66 Other receptors such as BasRS and CpxAR67,68 or KdpD and VicK (WalK)69,70 have also been described as putative adrenergic receptors in bacteria. Catecholamines have antimicrobial activity on selective bacteria in vitro71 and in vivo72 reducing the growth of Staphylococcus aureus or Candida albicans, for example.71 However, growth of other bacteria is increased by catecholamines in minimum medium- or low-iron medium,73,74 suggesting a role of catecholamines on iron utilization and amino acid biosynthesis. Catecholamines also affect bacterial chemotaxis, or their ability to migrate where conditions are beneficial for their growth.75–82 The effect on motility seems dose dependent as low dose of noradrenaline decreases motility genes of P. aeruginosa PA14 whereas a high concentration of noradrenaline increases them,78 showing the importance of the concentration of stress hormones for downstream effects on microbes. Catecholamines also influence the formation of bacterial biofilm, which is required for the bacteria to colonize the host and induce pathogenicity,83–85 promoting the biofilm’s thickness of E. coli for example.86 Other properties of bacteria are affected by catecholamines, such as sensitivity to antibiotics,87 or production of metabolites.88–90 In sum, the effects of stress hormones on bacteria are large, offering a possible explanation of the inter-individual variability of stress effects on microbiome.
Figure 3.
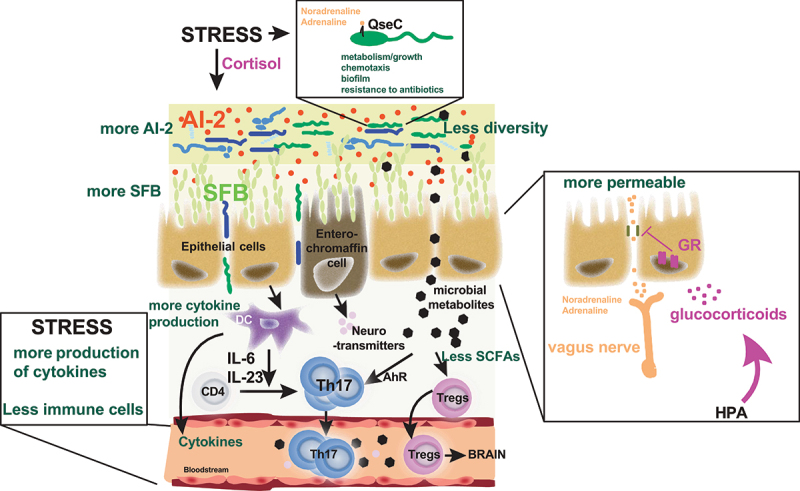
Effects of stress on the microbiota and the immune response.
Stress hormones such as norepinephrine, epinephrine, and cortisol, impact the microbiota diversity and metabolites. Associated with stress, there is an increase of cytokine production but a decrease in various populations of immune cells. The increase of proinflammatory cytokines leads to increased differentiation of Th17 cells.
In addition to modulate the composition and the properties of the gut microbiota, stress hormones also affect the host gastrointestinal properties. Thus, noradrenaline has been shown to affect colonic contractions.91 Increased gut permeability is only present in those with elevated levels of cortisol and besides cortisol, mast cells are also necessary to weaken the gut barriers to promote bacterial leakage. In addition, the adherence of bacteria to the gut mucosal lining and the uptake of bacteria into the Peyer’s patches are increased by adrenocorticotropic hormone (ACTH), cortisol and catecholamines.92–95 Taken together, the stress hormones have a profound impact on the gut microbiome.
Microbiome and stress-related behaviors
Several reports point out to the role of the microbiota in stress-related behaviors such as anxiety and depression. For example, germ-free mice52,96,97 or mice treated with antibiotics98 exhibit reduced anxiety-like behaviors, whereas germ-free mice receiving stress-prone microbiota exhibit depressive-like behaviors.99 Importantly, it has been demonstrated that stress exposure early in life or during adulthood affects the microbiota composition, suggesting that microbial populations shape the stress responsiveness of an organism.62,100–104 Although there is not a consensus about which bacteria are affected by stress, exposure to certain bacteria changes behaviors. Thus, exposure to Citrobacter rodentium, Trichuris muris, and Campylobacter jejuni increases anxious behavior in rodents.105,106 Segmented filamentous bacterium promotes depressive-like behaviors,107 whereas exposure to Lactobacillus spp. and Bifidobacterium spp. reduces anxiety-like and depression-like behaviors.108–111 Furthermore, diets that modify the microbiota such as prebiotics and probiotics have been shown to reduce stress-related behaviors.112–117
Consistent with, changes of the microbiome composition have been associated with psychiatric diseases, in particular depression and anxiety106,118,119 and administration of the probiotic Lactobacillus farciminis prevents the gut leakage and the activation of the HPA axis in depressed patients,111,120 pointing to a role of the microbiota in depression. Furthermore, there is an increased risk of severe mental illnesses after antibiotic use, which reduces the gut microbiota diversity,121–123 including a direct association between antibiotics use and subsequent development of depression.124 Similarly, exposure to environmental factors or unhealthy diet known to impact the microbiome increases the incidence of depression.125,126 Although modulation of the microbiota affects both cognition and mood,106,116,127–132 the mechanisms whereby the gut microbiota promotes depression remains to be fully understood. The findings about the changes of gut microbiota composition in depressed patients are inconsistent. Overall, depressed patients seem to have a shift toward pro-inflammatory bacteria,133,134 with an enhanced Bacteroidetes/Firmicutes ratio.135,136 One of the most consistent findings in depressed patients is an enrichment of the genus Bacteroides and Eggerthella and a depletion of the genera Blautia, Faecalibacterium, Coprococcus, and Sutterella.32,133,134,137,138 Transfer of human fecal microbiota from depressed patients to germ-free mice confers depressive-like behaviors to the recipient mice compared to mice receiving the microbiota from healthy patients, showing that the gut microbiome is sufficient for inducing depressive-like behaviors in mice.136,139
Gut metabolites are also released by gut bacteria or by the host in response to microbial changes.140 These include host molecules modified by bacteria such as bile acids or products of bacteria.140 Thus, because the composition of the microbiome is changed in depressed patients, the microbial metabolome is also affected. For example, the level of the bile acids modulated by the microbiota is inversely correlated with the severity of depression symptoms,141 suggesting that bile acids could provide anti-depressant actions. Furthermore, in depressed patients, microbial metabolites such as short chain fatty acids (SCFA) including acetate, propionate, and butyrate, are reduced.136,142 Butyrate, for example, has been shown to exert antidepressant actions when administered to rodents, reducing gut permeability and stress responsiveness.143,144 Other microbial metabolites, such as increases of trimethylamine-N-oxide from choline, lipopolysaccharide (LPS), lactate and B vitamins have been associated with depression.145–148 Neurotransmitters [e.g., serotonin or γ-aminobutyric acid (GABA)] are also produced by microbes and can modulate behaviors.144 This suggests that in addition to the microbiome composition, the metabolism and the metabolites of the microbiota are critical to modulate the stress response. Although some fungi metabolites can be detrimental for health such as mycotoxins,149 others have beneficial effects on health such as antibiotics,150 immunomodulatory metabolites such as cyclosporin A151 or mood modulator such as psilocybin.152
Quorum sensing molecules and stress
Chemical signaling molecules called quorum sensing molecules (QSMs) are used by bacteria to communicate, sense, and respond to environmental changes dependent on bacterial cell density.153 They regulate synchronicity of bacterial group behaviors, known as quorum sensing.154 In the gut, they regulate many of the interactions of the bacteria with the host environment to promote bacterial survival, including facilitation of bacterial attachment, nutritional supply, competition, and motility.155 Quorum sensing signaling molecules (autoinducers) are differently produced between Gram-negative and Gram-positive bacteria. N-acyl homoserine lactones (AHL) are mainly produced by Gram-negative bacteria (Figure 1), while autoinducer peptides (AIP) are mainly used by Gram-positive bacteria.156 AHL have been found in the upper intestine of both human and mice.157 Autoinducer (AI)-2 is considered a universal QSM produced by both Gram-negative and Gram-positive bacteria and is produced by > 50% of sequenced bacterial species. AI-2 mediates interspecies communication throughout the bacterial kingdom,158–160 is synthetized by LuxS161 (Figure 1) and engineered increase of AI-2 level in the gut favors the expansion of the Firmicutes phylum.155 AI-2 regulates niche-specific behaviors (e.g., cell division, virulence, motility, and biofilm formation) of both commensal and pathogenic bacteria.162,163 In the gut, Firmicutes use quorum sensing mediated by peptides [e.g., peptides produced as secondary signaling molecules after degradation of lipoproteins into the extracellular environment.164 To survive in the gut, bacteria adapt their metabolism through metabolite-sensing mechanisms in a mutual relationship with the mammalian host.165 Consistent with this, QSMs have been implicated in human diseases such as cancer and autism107,166–170 and more recently in the stress response.107 Thus, AI-2, but not AHL is sufficient to promote susceptibility to stress-related behaviors in mice by acting on Segmented filamentous bacterium.107 It is important to note that some bacteria can either not produce the autoinducer but can detect it, or can produce it but not detect it, the latter being especially true for AI-2. Altogether, stress has a profound impact on the gut microbiome, and reciprocally the gut microbiome is sensitive to stress signals.
Figure 1.
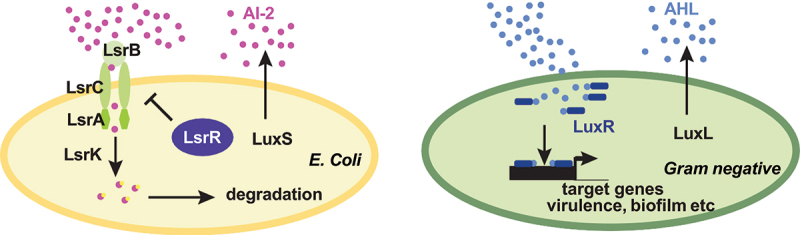
AI-2 system in E. coli and AHL system in gram negative bacteria.
In E. coli, AI-2 is produced by LuxS, while AHL in Gram negative bacteria is produced by LuxL. In E coli, AI-2 is sensed by LsrB, which captures AI-2 into the bacteria. AI-2 is then phosphorylated by LsrK. Phosphorylation triggers AI-2 degradation. AHLs, in contrast, bind to LuxR to modulate the expression of genes required for virulence, biofilm formation, etc.
Gut-brain axis, immunity, and stress
Early colonization of the gut is pivotal for the maturation of the immune system.171 The gut is exposed to commensal bacteria and their by-products, food and environmental antigens and exhibits a 3-layers barrier to prevent these to reach the lamina propria or the circulation: 1) a thick layer of mucus composed of hyperglycosylated mucin-2, produced by goblet cells, which provide protection by forming a shielding barrier, 2) a monolayer of enterocytes linked with tight junctions to restrict trans-epithelial passage of microbes and metabolites and 3) a gut vascular layer of endothelial cells, pericytes and glial cells.172 Associated with those layers, the gut hosts more than 70% of the total immune cells of the body including innate and adaptive immune cells173 (Figure 2). Dendritic cells in particular help maintaining the compartmentalization of the enteric microbiota.174
Figure 2.
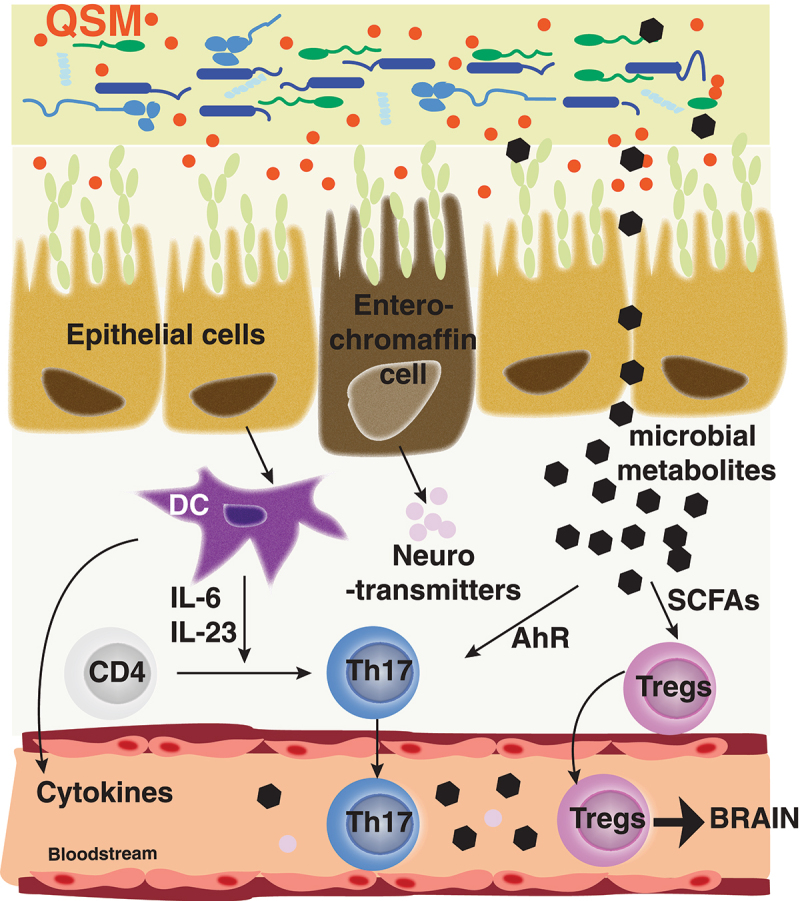
Effects of the microbiota on intestinal immune response.
Signals (QSM, bacteria, microbial metabolites, neurotransmitters, neuropeptides) originating from the microbiota affect the intestinal epithelial cells, enterochromaffin cells, or diffuse through the intestinal barrier, to modulate immune cells (T cells, dendritic cells, etc.) present in the lamina propria. Microbial signals and immune cells often reach the bloodstream and migrate to the brain. Here are presented cells relevant for the stress behavioral effects.
The importance of the microbiota for maintaining immune responses in the gut is shown using microbiota-depleted mice or germ-free mice. Germ-free mice have immunological defects such as abnormal germinal center, smaller Peyer’s patches and lymphoid follicles175 and reduced protective IgA.176 These defects are corrected when a specific-pathogen-free single mouse is placed in the cage of germ-free mice. Because the gut immune cells monitor potential infections and are in contact with microbial antigens, gut bacteria promote the development of T helper (Th) 17 cells,177 regulatory T cells (Tregs)178 and memory T cells.179–181 Thus, Segmented filamentous bacterium promotes Th17 cell differentiation178 while Clostridium spp. induce Tregs.182,183 In addition, commensal bacterial antigens shape immunoglobulin repertoires in the gut,184 interacting for example, with the carbohydrate antigens of the ABO blood group to stimulate IgM antibody response185 and protect from induction of mucosal IgE, which is associated with susceptibility to allergies.186 Toll-like receptor (TLR)5 recognizes flagellin of bacteria and is responsible for containing colonization by flagellated bacteria during the neonatal period.187 It is also thought that these interactions occur early in life to shape the immune response in adult.
However, the long-term impact of subtler dysbiosis early in life remains to be determined.
Neurons and astrocytes also participate in intestinal immunity. Thus, for example, astrocytes promotes group 3 innate lymphoid cell (ILC3) production of interleukin-22 and reciprocally immune cells confer survival signals to neurons to prevent inflammation-dependent bowel dysmotility.188
There is a bidirectional communication between the brain and the gut during homeostasis, which involves multiple pathways including endocrine (HPA axis), neural (vagus nerve) and immune (mucosal, peripheral and central) pathways to detect noxious stimuli which trigger local and systemic inflammation.189,190 The orchestration of the bidirectional communication in the stress response remains to be fully elucidated, including the descending pathways after stress affecting the gut microbial composition (Figure 3). Several ascending pathways controlling the communication from the gut to the brain have been identified. Thus, for example, microbiota changes often trigger local inflammatory responses, ranging from production of cytokines to differentiation of immune cells and production of antibodies or anti-microbial peptides (Figure 2). The immune cells are programmed to recognize microbes, and any inflammatory response will be induced when the immune system is in contact with microbes even commensal bacteria. One example is the lipopolysaccharide (LPS), present on Gram-negative bacteria, also known as endotoxin. Usually contained within the gut by the intestinal barriers,191 leakage of LPS into the lamina propria usually activates macrophages through TLR4,192 and TLR4 also mediates stress-related behaviors.107,193 TLR4 might be one of the pathways whereby stress by modifying the composition of the gut microbiota, increases pro-inflammatory cytokines (e.g., IL-6, IL-1, TNF and IFN) and chemokines,194 often leading to the disruption of the gut barrier which increases gut permeability and the release of LPS into the circulation.195,196 This is reversed by probiotic agents.61,112,196 This finding is particularly relevant for the stress response, as circulating inflammatory signals (e.g., cytokines and LPS) are sufficient to promote stress-related behaviors in mice193 and promote depressive symptoms in human.197
Other receptors such as NOD-like receptors shape the microbiota. Thus, NOD1 assists adaptive lymphoid cells to maintain intestinal homeostasis.198 NOD2 is a bacterial sensor of intestinal epithelial cells and immune cells recognizing peptidoglycan-conserved motifs and induces anti-microbial and anti-inflammatory responses.199 Once activated by commensal bacteria, NOD2 triggers survival and regeneration of the intestinal epithelium,200 controls the activation of antigen specific CD4 T cells including Th17 cells201 and suppresses the expansion of proinflammatory bacteria.199 NOD1 and NOD2 synergize with TLR4 to exacerbate sickness behaviors,202 whereas NOD1 depletion promotes susceptibility to stress-related behaviors.203
The Aryl hydrocarbon receptor (AhR) is expressed on intestinal epithelial cells but also immune cells. AhR is a ligand-dependent transcription factor, essential to detect bacterial metabolites. It is important to note that AhR binds various ligands besides microbial metabolites, such as phytochemicals, AhR-active tryptophan metabolites, dietary or pharmaceutical components.204–207 AhR has been recently shown to sense stress and to induce anxiety-like behaviors,208 suggesting that the change of microbiome after stress might be detected by AhR. Some AhR modulators act as antidepressant in female mice,209 although the mechanisms whereby they induce antidepressant actions remain to be determined.
Metabolite sensing G protein coupled receptors (GPCRs) are expressed on intestinal immune and nonimmune cells. GPCRs recognize microbiota-derived metabolites such as SCFA, amino acids, bile acids, lactate and promote the integrity of the intestinal barrier, inducing anti-inflammatory response and reducing the recruitment of pro-inflammatory immune cells by enhancing Tregs promotion. Overall, both the intestine and the immune system have a variety of receptors to sense the microbiome.
Consistent with this, transplantation of the microbiota of mice exposed to a stressor into germ-free mice induced exacerbated inflammatory responses to Citrobacter rodentium infection.210 Some have proposed a role for regulatory T cells and the secretion of IL-10 in mediating the probiotic effects of microorganisms,211–213 while other have focused on proinflammatory T cells to promote stress-related behaviors. Thus, stress-induced dysbiosis triggers the release of the proinflammatory cytokine, IL-17A, by Th17 cells to amplify the stress response.214 The commensal bacteria, Segmented filamentous bacterium, for example, is sufficient to promote Th17 cell- induced depressive-like behaviors in mice and Segmented filamentous bacterium-specific Th17 cells have been shown to migrate to the hippocampus after stress.107 Recently, Lactobacillus- dependent colonic IL-17A-producing T cells were shown to promote chronic social defeat215 and IL-17A-producing T cells also promote anxiety-like behaviors,216 pointing to the importance of IL-17A in promoting microbial-dependent stress-related behaviors. The importance of Th17 cells in mediating the effect of the microbiota after stress was further demonstrated using fecal transfer. Thus, fecal transfer of stools of depressed patients into germ-free-like mice induces depressive-like behaviors in the recipient mice,217 whereas this is abolished in germ-free-like mice deficient of Th17 cells,31 confirming the importance of Th17 cells in mediating microbiota-dependent depressive-like behaviors. In addition to T cell change, stress also affects the number of myeloid cells in the blood218 and this is dependent on the microbiota,219 since the microbiota promotes the production of monocytes220–223 and the trafficking of monocytes.143
Phages from pathogenic bacteria also interact with the immune system. They are engulfed by dendritic cells, monocytes, and B cells, to induce type I interferon responses through TLR3 signaling.224 E. coli phages isolated from human feces induce Th1 and Tc1 responses in germ-free mice at mucosal sites while phages from other bacteria stimulate cytokine production via TLR9,225 suggesting that the changes of bacteriophages observed in depressed patients might also impact the immune responses of these patients.
Bacterial metabolism furthermore impacts the immune response and ultimately the brain. SCFAs effects on inflammation differ depending on the type and concentration of SCFAs. Butyrate, for example, has anti-inflammatory properties.226,227 SCFAs levels strongly correlated with better cognitive test scores after stress in adolescents.228 SCFAs ameliorate depressive-like behaviors induced by chronic stress143,229 or reward seeking behavior after psychosocial stress.143 Thus for example, microbes through the activation of the SCFA-GPCR pathway230,231 promote the production of glial-cell derived neurotrophic factor (GDNF),232 which enhances the production of ILC3 and IL-22 to provide protection and restoration of the intestinal barriers.233 Bacterial metabolites induce the release by intestinal cells of local peptide neurotransmitters, including peptide tyrosine-tyrosine (PYY) and serotonin,234,235 in addition to bacteria producing nitric oxide,236 acetylcholine,237 noradrenaline,238 GABA239,240 and dopamine. Enteric neurons express specific receptors for some of these neurotransmitters,234,235 while other neurotransmitters have been shown to affect directly the brain.239,240 Several neurotransmitter released by gut microbes induce TLR signaling on epithelial, immune, and neuronal cells,241,242 suggesting that once activated by microbes the immune system could influence neurotransmitter levels that affect the stress response. In addition, microbial metabolites alter the vagus nerve signaling to the brain, representing another route whereby bacteria by modulating cytokines and neurotransmitters send signals to the brain. Taken together, the microbiota-inflammation axis is critical to promote stress-related behaviors impacting well-being, and possibly influencing other health outcomes.
Impact of inflammation and the microbiota on the brain physiology
In addition to understand the impact of the microbiota on the immune system, germ-free mice have also been instrumental in understanding the impact of the microbiota on brain physiology ranging from early brain development to behavioral tasks.243–245 Thus, structurally, the brains of microbiota-depleted mice exhibit hypermyelination of the prefrontal cortex,246,247 increased hippocampal concentration of serotonin,52 altered expression of brain-derived neurotrophic factor (BDNF) in the hippocampus, cortex and amygdala,51,96 increased neurogenesis in the hippocampus,10 reduced hippocampal dendritic spine density but increased basolateral amygdala dendritic spine density.248
Inflammation and changes of the microbiota have been shown to modulate the permeability of the blood-brain barrier. The depletion of the gut microbiota in germ-free mice is associated with decreased expression of tight junction proteins leading to structural alterations of the blood-brain barrier (BBB) and leaky BBB.249 Of note, colonizing germ-free mice with stool of specific-pathogen-free mice, decreases the permeability of the BBB,249 showing the importance of the gut microbes for the BBB integrity. Depletion of the gut microbiota with antibiotics in mice results in novel object recognition impairment250 and a decrease of hippocampal neurogenesis and memory retention.223 These impairments are reversed by administration of probiotic bacteria. Moreover, colonization with the butyrate producer Clostridium tyrobutyricum or treatment with sodium butyrate of germ-free mice increase tight junction protein expression improving BBB integrity.249 Sodium butyrate also prevents BBB breakdown after traumatic brain injury and promotes neurogenesis.251–253
The neurotransmitters produced by the gut microbiota could also affect brain function. Thus, Lactobacillus and Bifidobacterium spp. metabolize the most abundant amino acid: glutamate, which is an excitatory neurotransmitter in the brain, to produce GABA, an inhibitory neurotransmitter.240 The conversion of tryptophan present in various food254 to the neurotransmitter tryptamine is achieved by decarboxylases secreted by Clostridium sporogenes. Yet, tryptamine is necessary for the release of serotonin by intestinal cells.255,256 Consequently, germ-free mice have higher plasma tryptophan levels than specific-pathogen-free mice. The level of tryptophan in germ-free mice are normalized after colonization with the microbiota of a specific-pathogen-free mouse.52 Similarly, colonization with Bifidobacterium infantis increases plasma levels of tryptophan.257 The consequences of the conversion of tryptophan to indoles or other metabolites258 (e.g., kynurenic acid) by bacteria are anti-inflammatory259 and neuroprotective.260 For example, indoles are ligands of AhR that attenuates disease severity in the experimental autoimmune encephalomyelitis model.261 In contrast, the production of quinolinic acid has neurotoxic effects disrupting the BBB and has been implicated in the etiology of psychiatric disorders and neurodegenerative diseases.262,263 Therefore, understanding the role of tryptophan and glutamate-metabolizing bacteria may open new therapeutic avenues to 1) control BBB permeability, 2) enhance anti-inflammatory effects and 3) induce brain protection.
Microbiota is essential for the microglia development and function.264,265 Microglia respond to factor produced by the microbiota as shown by the difference in structure and function of microglia of germ-free and specific-pathogen-free mice.9,266,267 Microglia of germ-free mice are more ramified indicative of a resting state. Microglial morphology and density are also affected in germ-free mice and there is an attenuated inflammatory response,268 with male offspring microglia in early development being more affected than female offspring microglia.266 Yet, stress also affects microglia promoting their activation (e.g., increased branching of the microglia, production of proinflammatory cytokines) and activated microglia is associated with depressive-like behaviors.269 Whether microglia mediate microbiota-induced depressive-like behaviors remains to be determined.
Although the mechanisms whereby the microbiota affects the brain remain to be fully elucidated, there is convergent evidence for a role of the microbiota like the role of the immune system in shaping the brain function throughout the lifespan. Whether these changes by the microbiota require immune modulation remains to be determined.
Conclusions
Our understanding of the role of the microbiome in health and disease has advanced tremendously. Yet the characteristics, function and the composition of a healthy microbiome remains to be fully understood. Although the impacts of the microbiota in the stress response are starting to be uncovered, the molecular pathways responsible for these effects have not been yet fully understood. It is now clear that modulations of the microbiome have profound consequences on the stress response. Yet the bacteria responsible for stress effects are started to be identified, whereas other microbiome components such as phages, virus or fungus remain understudied. Many pathways converge on the gut microbiome, and it is unlikely that supplementation with only a cocktail of probiotics will be sufficient to provide relief from stress-related disorders. Diet appears as one of the modifier factor that might be able to control some of the negative effects of the stress response. Yet, with the climate change, it is projected that warmer climate will modify the soil-born fungi and viruses, which will likely impact the gut microbiome, and the stress response. The host response to microbial change involves a variety of metabolic and signaling pathways that could open new avenue of research to develop a new generation of microbiome-targeting drugs for both disease and prevention.
Acknowledgments
Work in Dr. Beurel’s lab was supported by the NIH (MH104656, MH110415). The author has nothing to disclose.
Funding Statement
This work was supported by the National institue of Health [MH104656, MH110415].
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- 1.Koolhaas JM, Bartolomucci A, Buwalda B, de Boer SF, Flügge G, Korte SM, Meerlo P, Murison R, Olivier B, Palanza P. et al. Stress revisited: a critical evaluation of the stress concept. Neurosci Biobehav Rev. 2011;35(5):1291–23. doi: 10.1016/j.neubiorev.2011.02.003. [DOI] [PubMed] [Google Scholar]
- 2.Salleh MR. Life event, stress and illness. Malays J Med Sci. 2008;15:9–18. [PMC free article] [PubMed] [Google Scholar]
- 3.McEwen BS. Protective and damaging effects of stress mediators: central role of the brain. Dialogues Clin Neurosci. 2006;8(4):367–381. doi: 10.31887/DCNS.2006.8.4/bmcewen. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stephens MA, Wand G. Stress and the HPA axis: role of glucocorticoids in alcohol dependence. Alcohol Res. 2012;34:468–483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Goldstein DS. Adrenal responses to stress. Cell Mol Neurobiol. 2010;30(8):1433–1440. doi: 10.1007/s10571-010-9606-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Sun Y, Zuo T, Cheung CP, Gu W, Wan Y, Zhang F, Chen N, Zhan H, Yeoh YK, Niu J. et al. Population-level configurations of gut mycobiome across 6 ethnicities in urban and rural China. Gastroenterology. 2021;160(1):272–286.e11. doi: 10.1053/j.gastro.2020.09.014. [DOI] [PubMed] [Google Scholar]
- 7.Zuo T, Sun Y, Wan Y, Yeoh YK, Zhang F, Cheung CP, Chen N, Luo J, Wang W, Sung JJY. et al. Human-gut-DNA virome variations across geography, ethnicity, and urbanization. Cell Host Microbe. 2020;28(5):741–751.e4. doi: 10.1016/j.chom.2020.08.005. [DOI] [PubMed] [Google Scholar]
- 8.Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA. Diversity of the human intestinal microbial flora. Science. 2005;308(5728):1635–1638. doi: 10.1126/science.1110591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Erny D, Hrabě de Angelis AL, Jaitin D, Wieghofer P, Staszewski O, David E, Keren-Shaul H, Mahlakoiv T, Jakobshagen K, Buch T. et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat Neurosci. 2015;18(7):965–977. doi: 10.1038/nn.4030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ogbonnaya ES, Clarke G, Shanahan F, Dinan TG, Cryan JF, O’Leary OF. Adult hippocampal neurogenesis is regulated by the microbiome. Biol Psychiatry. 2015;78(4):e7–9. doi: 10.1016/j.biopsych.2014.12.023. [DOI] [PubMed] [Google Scholar]
- 11.Nash AK, Auchtung TA, Wong MC, Smith DP, Gesell JR, Ross MC, Stewart CJ, Metcalf GA, Muzny DM, Gibbs RA. et al. The gut mycobiome of the human microbiome project healthy cohort. Microbiome. 2017;5(1):153. doi: 10.1186/s40168-017-0373-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sender R, Fuchs S, Milo R. Revised estimates for the Number of Human and Bacteria Cells in the body. PLoS Biol. 2016;14(8):e1002533. doi: 10.1371/journal.pbio.1002533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, Wolfe BE, Ling AV, Devlin AS, Varma Y, Fischbach MA. et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature. 2014;505(7484):559–563. doi: 10.1038/nature12820. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Liguori G, Lamas B, Richard ML, Brandi G, da Costa G, Hoffmann TW, Di Simone MP, Calabrese C, Poggioli G, Langella P. et al. Fungal dysbiosis in mucosa-associated microbiota of Crohn’s disease patients. J Crohns Colitis. 2016;10(3):296–305. doi: 10.1093/ecco-jcc/jjv209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Nelson A, Stewart CJ, Kennedy NA, Lodge JK, Tremelling M, Probert CS, Parkes M, Mansfield JC, Smith DL, Hold GL. et al. The impact of NOD2 genetic variants on the gut mycobiota in Crohn’s disease patients in remission and in individuals without gastrointestinal inflammation. J Crohns Colitis. 2021;15(5):800–812. doi: 10.1093/ecco-jcc/jjaa220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Sokol H, Leducq V, Aschard H, Pham H-P, Jegou S, Landman C, Cohen D, Liguori G, Bourrier A, Nion-Larmurier I. et al. Fungal microbiota dysbiosis in IBD. Gut. 2017;66(6):1039–1048. doi: 10.1136/gutjnl-2015-310746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Strati F, Cavalieri D, Albanese D, De Felice C, Donati C, Hayek J, Jousson O, Leoncini S, Renzi D, Calabrò A. et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome. 2017;5(1):24. doi: 10.1186/s40168-017-0242-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Zou R, Wang Y, Duan M, Guo M, Zhang Q, Zheng H. Dysbiosis of gut fungal microbiota in children with autism spectrum disorders. J Autism Dev Disord. 2021;51(1):267–275. doi: 10.1007/s10803-020-04543-y. [DOI] [PubMed] [Google Scholar]
- 19.Hoarau G, Mukherjee PK, Gower-Rousseau C, Hager C, Chandra J, Retuerto MA, Neut C, Vermeire S, Clemente J, Colombel JF. et al. Bacteriome and mycobiome interactions underscore microbial dysbiosis in familial Crohn’s disease. mBio. 2016;7(5):10–128. doi: 10.1128/mBio.01250-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Van Dyken SJ, Garcia D, Porter P, Huang X, Quinlan PJ, Blanc PD, Corry DB, Locksley RM. Fungal chitin from asthma-associated home environments induces eosinophilic lung infiltration. J Immunol. 2011;187(5):2261–2267. doi: 10.4049/jimmunol.1100972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Chu H, Duan Y, Lang S, Jiang L, Wang Y, Llorente C, Liu J, Mogavero S, Bosques-Padilla F, Abraldes JG. et al. The Candida albicans exotoxin candidalysin promotes alcohol-associated liver disease. J Hepatol. 2020;72(3):391–400. doi: 10.1016/j.jhep.2019.09.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Severance EG, Alaedini A, Yang S, Halling M, Gressitt KL, Stallings CR, Origoni AE, Vaughan C, Khushalani S, Leweke FM. et al. Gastrointestinal inflammation and associated immune activation in schizophrenia. Schizophr Res. 2012;138(1):48–53. doi: 10.1016/j.schres.2012.02.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Severance EG, Gressitt KL, Stallings CR, Katsafanas E, Schweinfurth LA, Savage CL, Adamos MB, Sweeney KM, Origoni AE, Khushalani S. et al. Candida albicans exposures, sex specificity and cognitive deficits in schizophrenia and bipolar disorder. NPJ Schizophr. 2016;2(1):16018. doi: 10.1038/npjschz.2016.18. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zuo T, Zhan H, Zhang F, Liu Q, Tso EYK, Lui GCY, Chen N, Li A, Lu W, Chan FKL. et al. Alterations in fecal fungal microbiome of patients with COVID-19 during time of hospitalization until discharge. Gastroenterology. 2020;159(4):1302–1310.e5. doi: 10.1053/j.gastro.2020.06.048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Richard ML, Sokol H. The gut mycobiota: insights into analysis, environmental interactions and role in gastrointestinal diseases. Nat Rev Gastroenterol Hepatol. 2019;16:331–345. doi: 10.1038/s41575-019-0121-2. [DOI] [PubMed] [Google Scholar]
- 26.Kasper L, König A, Koenig P-A, Gresnigt MS, Westman J, Drummond RA, Lionakis MS, Groß O, Ruland J, Naglik JR. et al. The fungal peptide toxin Candidalysin activates the NLRP3 inflammasome and causes cytolysis in mononuclear phagocytes. Nat Commun. 2018;9(1):4260. doi: 10.1038/s41467-018-06607-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gil ML, Gozalbo D. Role of Toll-like receptors in systemic Candida albicans infections. Front Biosci. 2009;14:570–582. doi: 10.2741/3263. [DOI] [PubMed] [Google Scholar]
- 28.Hao SR, Zhang Z, Zhou Y-Y, Zhang X, Sun W-J, Yang Z, Zhao J-H, Jiang H-Y. Altered gut bacterial–fungal interkingdom networks in children and adolescents with depression. J Affect Disord. 2023;332:64–71. doi: 10.1016/j.jad.2023.03.086. [DOI] [PubMed] [Google Scholar]
- 29.van Tilburg Bernardes E, Pettersen VK, Gutierrez MW, Laforest-Lapointe I, Jendzjowsky NG, Cavin J-B, Vicentini FA, Keenan CM, Ramay HR, Samara J. et al. Intestinal fungi are causally implicated in microbiome assembly and immune development in mice. Nat Commun. 2020;11(1):2577. doi: 10.1038/s41467-020-16431-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Liang G, Bushman FD. The human virome: assembly, composition and host interactions. Nat Rev Microbiol. 2021;19(8):514–527. doi: 10.1038/s41579-021-00536-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Medina-Rodriguez EM, Watson J, Reyes J, Trivedi M, Beurel E. Th17 cells sense microbiome to promote depressive-like behaviors. Microbiome. 2023;11(1):92. doi: 10.1186/s40168-022-01428-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yang J, Zheng P, Li Y, Wu J, Tan X, Zhou J, Sun Z, Chen X, Zhang G, Zhang H. et al. Landscapes of bacterial and metabolic signatures and their interaction in major depressive disorders. Sci Adv. 2020;6(49):eaba8555. doi: 10.1126/sciadv.aba8555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Glaser R, Kiecolt-Glaser JK, Malarkey WB, Sheridan JF. The influence of psychological stress on the Immune Response to vaccines a. Ann NY Acad Sci. 1998;840(1):649–655. doi: 10.1111/j.1749-6632.1998.tb09603.x. [DOI] [PubMed] [Google Scholar]
- 34.Cohen S, Frank E, Doyle WJ, Skoner DP, Rabin BS, Gwaltney JM. Types of stressors that increase susceptibility to the common cold in healthy adults. Health Psychol. 1998;17(3):214–223. doi: 10.1037/0278-6133.17.3.214. [DOI] [PubMed] [Google Scholar]
- 35.Cohen S, Tyrrell DA, Smith AP. Psychological stress and susceptibility to the common cold. N Engl J Med. 1991;325(9):606–612. doi: 10.1056/NEJM199108293250903. [DOI] [PubMed] [Google Scholar]
- 36.Kortright KE, Chan BK, Koff JL, Turner PE. Phage therapy: a renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe. 2019;25(2):219–232. doi: 10.1016/j.chom.2019.01.014. [DOI] [PubMed] [Google Scholar]
- 37.Boling L, Cuevas DA, Grasis JA, Kang HS, Knowles B, Levi K, Maughan H, McNair K, Rojas MI, Sanchez SE. et al. Dietary prophage inducers and antimicrobials: toward landscaping the human gut microbiome. Gut Microbes. 2020;11(4):721–734. doi: 10.1080/19490976.2019.1701353. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bien J, Palagani V, Bozko P. The intestinal microbiota dysbiosis and clostridium difficile infection: is there a relationship with inflammatory bowel disease? Therap Adv Gastroenterol. 2013;6(1):53–68. doi: 10.1177/1756283X12454590. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Zeng MY, Inohara N, Nunez G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017;10(1):18–26. doi: 10.1038/mi.2016.75. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zaneveld JR, McMinds R, Vega Thurber R. Stress and stability: applying the Anna Karenina principle to animal microbiomes. Nat Microbiol. 2017;2(9):17121. doi: 10.1038/nmicrobiol.2017.121. [DOI] [PubMed] [Google Scholar]
- 41.Knowles SR, Nelson EA, Palombo EA. Investigating the role of perceived stress on bacterial flora activity and salivary cortisol secretion: a possible mechanism underlying susceptibility to illness. Biol Psychol. 2008;77(2):132–137. doi: 10.1016/j.biopsycho.2007.09.010. [DOI] [PubMed] [Google Scholar]
- 42.Vanuytsel T, van Wanrooy S, Vanheel H, Vanormelingen C, Verschueren S, Houben E, Salim Rasoel S, Tόth J, Holvoet L, Farré R. et al. Psychological stress and corticotropin-releasing hormone increase intestinal permeability in humans by a mast cell-dependent mechanism. Gut. 2014;63(8):1293–1299. doi: 10.1136/gutjnl-2013-305690. [DOI] [PubMed] [Google Scholar]
- 43.Kiecolt-Glaser JK, Wilson SJ, Bailey ML, Andridge R, Peng J, Jaremka LM, Fagundes CP, Malarkey WB, Laskowski B, Belury MA. et al. Marital distress, depression, and a leaky gut: translocation of bacterial endotoxin as a pathway to inflammation. Psychoneuroendocrinology. 2018;98:52–60. doi: 10.1016/j.psyneuen.2018.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Schulz MD, Atay Ç, Heringer J, Romrig FK, Schwitalla S, Aydin B, Ziegler PK, Varga J, Reindl W, Pommerenke C. et al. High-fat-diet-mediated dysbiosis promotes intestinal carcinogenesis independently of obesity. Nature. 2014;514(7523):508–512. doi: 10.1038/nature13398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Reichenberger J, Kuppens P, Liedlgruber M, Wilhelm FH, Tiefengrabner M, Ginzinger S, Blechert J. No haste, more taste: an EMA study of the effects of stress, negative and positive emotions on eating behavior. Biol Psychol. 2018;131:54–62. doi: 10.1016/j.biopsycho.2016.09.002. [DOI] [PubMed] [Google Scholar]
- 46.Cornil Y, Chandon P. From fan to fat? Vicarious losing increases unhealthy eating, but self-affirmation is an effective remedy. Psychol Sci. 2013;24(10):1936–1946. doi: 10.1177/0956797613481232. [DOI] [PubMed] [Google Scholar]
- 47.Tryon MS, Carter CS, Decant R, Laugero KD. Chronic stress exposure may affect the brain’s response to high calorie food cues and predispose to obesogenic eating habits. Physiol Behav. 2013;120:233–242. doi: 10.1016/j.physbeh.2013.08.010. [DOI] [PubMed] [Google Scholar]
- 48.Boulange CL, Neves AL, Chilloux J, Nicholson JK, Dumas ME. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016;8(1):42. doi: 10.1186/s13073-016-0303-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Vanuytsel T, Tack J, Farre R. The role of intestinal permeability in gastrointestinal disorders and Current methods of evaluation. Front Nutr. 2021;8:717925. doi: 10.3389/fnut.2021.717925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Cinque C, Zinni M, Zuena AR, Giuli C, Alemà SG, Catalani A, Casolini P, Cozzolino R. Faecal corticosterone metabolite assessment in socially housed male and female wistar rats. Endocr Connect. 2018;7(2):250–257. doi: 10.1530/EC-17-0338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Sudo N, Chida Y, Aiba Y, Sonoda J, Oyama N, Yu X-N, Kubo C, Koga Y. Postnatal microbial colonization programs the hypothalamic–pituitary–adrenal system for stress response in mice. J Physiol. 2004;558(1):263–275. doi: 10.1113/jphysiol.2004.063388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Clarke G, Grenham S, Scully P, Fitzgerald P, Moloney RD, Shanahan F, Dinan TG, Cryan JF. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol Psychiatry. 2013;18(6):666–673. doi: 10.1038/mp.2012.77. [DOI] [PubMed] [Google Scholar]
- 53.Crumeyrolle-Arias M, Jaglin M, Bruneau A, Vancassel S, Cardona A, Daugé V, Naudon L, Rabot S. Absence of the gut microbiota enhances anxiety-like behavior and neuroendocrine response to acute stress in rats. Psychoneuroendocrinology. 2014;42:207–217. doi: 10.1016/j.psyneuen.2014.01.014. [DOI] [PubMed] [Google Scholar]
- 54.Luo Y, Zeng B, Zeng L, Du X, Li B, Huo R, Liu L, Wang H, Dong M, Pan J. et al. Gut microbiota regulates mouse behaviors through glucocorticoid receptor pathway genes in the hippocampus. Transl Psychiatry. 2018;8(1):187. doi: 10.1038/s41398-018-0240-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lyte M, Ernst S. Catecholamine induced growth of gram negative bacteria. Life Sci. 1992;50(3):203–212. doi: 10.1016/0024-3205(92)90273-R. [DOI] [PubMed] [Google Scholar]
- 56.Mudd AT, Berding K, Wang M, Donovan SM, Dilger RN. Serum cortisol mediates the relationship between fecal ruminococcus and brain N-acetylaspartate in the young pig. Gut Microbes. 2017;8(6):589–600. doi: 10.1080/19490976.2017.1353849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Freestone PP, Williams PH, Haigh RD, Maggs AF, Neal CP, Lyte M. Growth stimulation of intestinal commensal Escherichia coli by catecholamines: a possible contributory factor in trauma-induced sepsis. Shock. 2002;18(5):465–470. doi: 10.1097/00024382-200211000-00014. [DOI] [PubMed] [Google Scholar]
- 58.Keskitalo A, Aatsinki A-K, Kortesluoma S, Pelto J, Korhonen L, Lahti L, Lukkarinen M, Munukka E, Karlsson H, Karlsson L. et al. Gut microbiota diversity but not composition is related to saliva cortisol stress response at the age of 2.5 months. Stress. 2021;24(5):551–560. doi: 10.1080/10253890.2021.1895110. [DOI] [PubMed] [Google Scholar]
- 59.Lyte M, Vulchanova L, Brown DR. Stress at the intestinal surface: catecholamines and mucosa–bacteria interactions. Cell Tissue Res. 2011;343(1):23–32. doi: 10.1007/s00441-010-1050-0. [DOI] [PubMed] [Google Scholar]
- 60.Desbonnet L, Garrett L, Clarke G, Kiely B, Cryan JF, Dinan TG. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience. 2010;170(4):1179–1188. doi: 10.1016/j.neuroscience.2010.08.005. [DOI] [PubMed] [Google Scholar]
- 61.Gareau MG, Jury J, MacQueen G, Sherman PM, Perdue MH. Probiotic treatment of rat pups normalises corticosterone release and ameliorates colonic dysfunction induced by maternal separation. Gut. 2007;56(11):1522–1528. doi: 10.1136/gut.2006.117176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.O’Mahony SM, Marchesi JR, Scully P, Codling C, Ceolho A-M, Quigley EMM, Cryan JF, Dinan TG. Early life stress alters behavior, immunity, and microbiota in rats: implications for irritable bowel syndrome and psychiatric illnesses. Biol Psychiatry. 2009;65(3):263–267. doi: 10.1016/j.biopsych.2008.06.026. [DOI] [PubMed] [Google Scholar]
- 63.Barouei J, Moussavi M, Hodgson DM, Heimesaat MM. Effect of maternal probiotic intervention on HPA axis, immunity and gut microbiota in a rat model of irritable bowel syndrome. PloS One. 2012;7(10):e46051. doi: 10.1371/journal.pone.0046051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Weigel WA, Demuth DR. QseBC, a two-component bacterial adrenergic receptor and global regulator of virulence in Enterobacteriaceae and Pasteurellaceae. Mol Oral Microbiol. 2016;31(5):379–397. doi: 10.1111/omi.12138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Clarke MB, Hughes DT, Zhu C, Boedeker EC, Sperandio V. The QseC sensor kinase: a bacterial adrenergic receptor. Proc Natl Acad Sci USA. 2006;103(27):10420–10425. doi: 10.1073/pnas.0604343103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Duran-Pinedo AE, Solbiati J, Frias-Lopez J. The effect of the stress hormone cortisol on the metatranscriptome of the oral microbiome. Npj Biofilms Microbiomes. 2018;4(1):25. doi: 10.1038/s41522-018-0068-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Karavolos MH, Bulmer DM, Spencer H, Rampioni G, Schmalen I, Baker S, Pickard D, Gray J, Fookes M, Winzer K. et al. Salmonella typhi sense host neuroendocrine stress hormones and release the toxin haemolysin E. EMBO Rep. 2011;12(3):252–258. doi: 10.1038/embor.2011.4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Karavolos MH, Spencer H, Bulmer DM, Thompson A, Winzer K, Williams P, Hinton J, Khan CA. Adrenaline modulates the global transcriptional profile of Salmonella revealing a role in the antimicrobial peptide and oxidative stress resistance responses. BMC Genomics. 2008;9(1):458. doi: 10.1186/1471-2164-9-458. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Borrel V, Thomas P, Catovic C, Racine P-J, Konto-Ghiorghi Y, Lefeuvre L, Duclairoir-Poc C, Zouboulis CC, Feuilloley MGJ. Acne and stress: impact of catecholamines on Cutibacterium acnes. Front Med. 2019;6:155. doi: 10.3389/fmed.2019.00155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Cambronel M, Nilly F, Mesguida O, Boukerb AM, Racine P-J, Baccouri O, Borrel V, Martel J, Fécamp F, Knowlton R. et al. Influence of catecholamines (Epinephrine/Norepinephrine) on biofilm formation and adhesion in pathogenic and probiotic strains of Enterococcus faecalis. Front Microbiol. 2020;11:1501. doi: 10.3389/fmicb.2020.01501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Cuvas Apan O, Apan TZ, Apan A. In vitro antimicrobial activity of commonly used vasoactive drugs. J Clin Anesth. 2016;34:407–411. doi: 10.1016/j.jclinane.2016.05.008. [DOI] [PubMed] [Google Scholar]
- 72.Kesici S, Demirci M, Kesici U. Antibacterial effects of lidocaine and adrenaline. Int Wound J. 2019;16(5):1190–1194. doi: 10.1111/iwj.13182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Freestone PP, Haigh RD, Williams PH, Lyte M. Stimulation of bacterial growth by heat-stable, norepinephrine-induced autoinducers. FEMS Microbiol Lett. 1999;172(1):53–60. doi: 10.1111/j.1574-6968.1999.tb13449.x. [DOI] [PubMed] [Google Scholar]
- 74.Williams PH, Rabsch W, Methner U, Voigt W, Tschäpe H, Reissbrodt R. Catecholate receptor proteins in Salmonella enterica: role in virulence and implications for vaccine development. Vaccine. 2006;24(18):3840–3844. doi: 10.1016/j.vaccine.2005.07.020. [DOI] [PubMed] [Google Scholar]
- 75.Chet I, Henis Y, Mitchell R. Effect of biogenic amines and cannabinoids on bacterial chemotaxis. J Bacteriol. 1973;115(3):1215–1218. doi: 10.1128/jb.115.3.1215-1218.1973. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Biaggini K, Barbey C, Borrel V, Feuilloley M, Déchelotte P, Connil N. The pathogenic potential of Pseudomonas fluorescens MFN1032 on enterocytes can be modulated by serotonin, substance P and epinephrine. Arch Microbiol. 2015;197(8):983–990. doi: 10.1007/s00203-015-1135-y. [DOI] [PubMed] [Google Scholar]
- 77.Cambronel M, Tortuel D, Biaggini K, Maillot O, Taupin L, Réhel K, Rincé I, Muller C, Hardouin J, Feuilloley M. et al. Epinephrine affects motility, and increases adhesion, biofilm and virulence of Pseudomonas aeruginosa H103. Sci Rep. 2019;9(1):20203. doi: 10.1038/s41598-019-56666-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hegde M, Wood TK, Jayaraman A. The neuroendocrine hormone norepinephrine increases Pseudomonas aeruginosa PA14 virulence through the las quorum-sensing pathway. Appl Microbiol Biotechnol. 2009;84(4):763–776. doi: 10.1007/s00253-009-2045-1. [DOI] [PubMed] [Google Scholar]
- 79.Bearson BL, Bearson SM. The role of the QseC quorum-sensing sensor kinase in colonization and norepinephrine-enhanced motility of Salmonella enterica serovar typhimurium. Microb Pathog. 2008;44(4):271–278. doi: 10.1016/j.micpath.2007.10.001. [DOI] [PubMed] [Google Scholar]
- 80.Bansal T, Englert D, Lee J, Hegde M, Wood TK, Jayaraman A. Differential effects of epinephrine, norepinephrine, and indole on Escherichia coli O157: H7 chemotaxis, colonization, and gene expression. Infect Immun. 2007;75(9):4597–4607. doi: 10.1128/IAI.00630-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Cogan TA, Thomas AO, Rees LEN, Taylor AH, Jepson MA, Williams PH, Ketley J, Humphrey TJ. Norepinephrine increases the pathogenic potential of Campylobacter jejuni. Gut. 2007;56(8):1060–1065. doi: 10.1136/gut.2006.114926. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Yang Q, Zou P, Cao Z, Wang Q, Fu S, Xie G, Huang J. QseC inhibition as a novel antivirulence strategy for the prevention of acute hepatopancreatic necrosis disease (AHPND)-causing Vibrio parahaemolyticus. Front Cell Infect Microbiol. 2020;10:594652. doi: 10.3389/fcimb.2020.594652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Costerton JW, Stewart PS, Greenberg EP. Bacterial biofilms: a common cause of persistent infections. Science. 1999;284(5418):1318–1322. doi: 10.1126/science.284.5418.1318. [DOI] [PubMed] [Google Scholar]
- 84.Donlan RM, Costerton JW. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev. 2002;15(2):167–193. doi: 10.1128/CMR.15.2.167-193.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Parsek MR, Singh PK. Bacterial biofilms: an emerging link to disease pathogenesis. Annu Rev Microbiol. 2003;57(1):677–701. doi: 10.1146/annurev.micro.57.030502.090720. [DOI] [PubMed] [Google Scholar]
- 86.Yang K, Meng J, Huang Y-C, Ye L-H, Li G-J, Huang J, Chen H-M. The role of the QseC quorum-sensing sensor kinase in epinephrine- enhanced motility and biofilm formation by Escherichia coli. Cell Biochem Biophys. 2014;70(1):391–398. doi: 10.1007/s12013-014-9924-5. [DOI] [PubMed] [Google Scholar]
- 87.Freestone PP, Hirst RA, Sandrini SM, Sharaff F, Fry H, Hyman S, O’Callaghan C. Pseudomonas aeruginosa-catecholamine inotrope interactions: a contributory factor in the development of ventilator-associated pneumonia? Chest. 2012;142(5):1200–1210. doi: 10.1378/chest.11-2614. [DOI] [PubMed] [Google Scholar]
- 88.Roberts A, Matthews JB, Socransky SS, Freestone PPE, Williams PH, Chapple ILC. Stress and the periodontal diseases: effects of catecholamines on the growth of periodontal bacteria in vitro. Oral Microbiol Immunol. 2002;17(5):296–303. doi: 10.1034/j.1399-302X.2002.170506.x. [DOI] [PubMed] [Google Scholar]
- 89.Calil CM, Oliveira GM, Cogo K, Pereira AC, Marcondes FK, Groppo FC. Effects of stress hormones on the production of volatile sulfur compounds by periodontopathogenic bacteria. Braz Oral Res. 2014;28(1):1–8. doi: 10.1590/1807-3107BOR-2014.vol28.0008. [DOI] [PubMed] [Google Scholar]
- 90.de Lima PO, Nani BD, Almeida B, Marcondes FK, Groppo FC, de Moraes ABA, Franz‐Montan M, Cogo‐Müller K. Stress-related salivary proteins affect the production of volatile sulfur compounds by oral bacteria. Oral Dis. 2018;24(7):1358–1366. doi: 10.1111/odi.12890. [DOI] [PubMed] [Google Scholar]
- 91.Kurahashi M, Kito Y, Hara M, Takeyama H, Sanders KM, Hashitani H. Norepinephrine has dual effects on human colonic contractions through distinct subtypes of alpha 1 adrenoceptors. Cell Mol Gastroenterol Hepatol. 2020;10(3):658–671.e1. doi: 10.1016/j.jcmgh.2020.04.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Chen C, Brown DR, Xie Y, Green BT, Lyte M. Catecholamines modulate Escherichia coli O157:H7 adherence to murine cecal mucosa. Shock. 2003;20(2):183–188. doi: 10.1097/01.shk.0000073867.66587.e0. [DOI] [PubMed] [Google Scholar]
- 93.Green BT, Lyte M, Chen C, Xie Y, Casey MA, Kulkarni-Narla A, Vulchanova L, Brown DR. Adrenergic modulation of Escherichia coli O157: H7 adherence to the colonic mucosa. Am J Physiol Gastrointest Liver Physiol. 2004;287(6):G1238–G1246. doi: 10.1152/ajpgi.00471.2003. [DOI] [PubMed] [Google Scholar]
- 94.Schreiber KL, Brown DR. Adrenocorticotrophic hormone modulates Escherichia coli O157:H7 adherence to porcine colonic mucosa. Stress. 2005;8(3):185–190. doi: 10.1080/10253890500188732. [DOI] [PubMed] [Google Scholar]
- 95.Unsal H, Balkaya M, Ünsal C, Bıyık H, Başbülbül G, Poyrazoğlu E. The short-term effects of different doses of dexamethasone on the numbers of some bacteria in the ileum. Dig Dis Sci. 2008;53(7):1842–1845. doi: 10.1007/s10620-007-0089-6. [DOI] [PubMed] [Google Scholar]
- 96.Diaz Heijtz R, Wang S, Anuar F, Qian Y, Björkholm B, Samuelsson A, Hibberd ML, Forssberg H, Pettersson S. Normal gut microbiota modulates brain development and behavior. Proc Natl Acad Sci U S A. 2011;108(7):3047–3052. doi: 10.1073/pnas.1010529108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Neufeld KA, Kang N, Bienenstock J, Foster JA. Effects of intestinal microbiota on anxiety-like behavior. Commun Integr Biol. 2011;4(4):492–494. doi: 10.4161/cib.15702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Desbonnet L, Clarke G, Traplin A, O’Sullivan O, Crispie F, Moloney RD, Cotter PD, Dinan TG, Cryan JF. Gut microbiota depletion from early adolescence in mice: implications for brain and behaviour. Brain Behav Immun. 2015;48:165–173. doi: 10.1016/j.bbi.2015.04.004. [DOI] [PubMed] [Google Scholar]
- 99.Bercik P, Denou E, Collins J, Jackson W, Lu J, Jury J, Deng Y, Blennerhassett P, Macri J, McCoy KD. et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology. 2011;141(2):599–609.e3. doi: 10.1053/j.gastro.2011.04.052. [DOI] [PubMed] [Google Scholar]
- 100.Bailey MT, Dowd SE, Galley JD, Hufnagle AR, Allen RG, Lyte M. Exposure to a social stressor alters the structure of the intestinal microbiota: implications for stressor-induced immunomodulation. Brain Behav Immun. 2011;25(3):397–407. doi: 10.1016/j.bbi.2010.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Bharwani A, Mian, MF, Foster, JA, Surette, MG, Bienenstock J, Forsythe P. Structural & functional consequences of chronic psychosocial stress on the microbiome & host. Psychoneuroendocrinology. 2016;63:217–227. doi: 10.1016/j.psyneuen.2015.10.001. [DOI] [PubMed] [Google Scholar]
- 102.De Palma G, Blennerhassett P, Lu J, Deng Y, Park AJ, Green W, Denou E, Silva MA, Santacruz A, Sanz Y. et al. Microbiota and host determinants of behavioural phenotype in maternally separated mice. Nat Commun. 2015;6(1):7735. doi: 10.1038/ncomms8735. [DOI] [PubMed] [Google Scholar]
- 103.Golubeva AV, Crampton S, Desbonnet L, Edge D, O’Sullivan O, Lomasney KW, Zhdanov AV, Crispie F, Moloney RD, Borre YE. et al. Prenatal stress-induced alterations in major physiological systems correlate with gut microbiota composition in adulthood. Psychoneuroendocrinology. 2015;60:58–74. doi: 10.1016/j.psyneuen.2015.06.002. [DOI] [PubMed] [Google Scholar]
- 104.Jasarevic E, Howerton CL, Howard CD, Bale TL. Alterations in the vaginal microbiome by maternal stress are associated with metabolic reprogramming of the offspring gut and brain. Endocrinology. 2015;156(9):3265–3276. doi: 10.1210/en.2015-1177. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Lyte M, Varcoe JJ, Bailey MT. Anxiogenic effect of subclinical bacterial infection in mice in the absence of overt immune activation. Physiol Behav. 1998;65(1):63–68. doi: 10.1016/S0031-9384(98)00145-0. [DOI] [PubMed] [Google Scholar]
- 106.Stilling RM, Dinan TG, Cryan JF. Microbial genes, brain & behaviour – epigenetic regulation of the gut–brain axis. Genes Brain Behav. 2014;13(1):69–86. doi: 10.1111/gbb.12109. [DOI] [PubMed] [Google Scholar]
- 107.Medina-Rodriguez EM, Madorma D, O’Connor G, Mason BL, Han D, Deo SK, Oppenheimer M, Nemeroff CB, Trivedi MH, Daunert S. et al. Identification of a signaling mechanism by which the microbiome regulates Th17 Cell-Mediated depressive-like behaviors in mice. Am J Psychiatry. 2020;177(10):974–990. doi: 10.1176/appi.ajp.2020.19090960. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Arseneault-Breard J, Rondeau I, Gilbert K, Girard S-A, Tompkins TA, Godbout R, Rousseau G. Combination of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 reduces post-myocardial infarction depression symptoms and restores intestinal permeability in a rat model. Br J Nutr. 2012;107(12):1793–1799. doi: 10.1017/S0007114511005137. [DOI] [PubMed] [Google Scholar]
- 109.Bercik P, Park AJ, Sinclair D, Khoshdel A, Lu J, Huang X, Deng Y, Blennerhassett PA, Fahnestock M, Moine D. et al. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol Motil. 2011;23(12):1132–1139. doi: 10.1111/j.1365-2982.2011.01796.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Bravo JA, Dinan TG, Cryan JF. Alterations in the central CRF system of two different rat models of comorbid depression and functional gastrointestinal disorders. Int J Neuropsychopharmacol. 2011;14(5):666–683. doi: 10.1017/S1461145710000994. [DOI] [PubMed] [Google Scholar]
- 111.Messaoudi M, Lalonde R, Violle N, Javelot H, Desor D, Nejdi A, Bisson J-F, Rougeot C, Pichelin M, Cazaubiel M. et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br J Nutr. 2011;105(5):755–764. doi: 10.1017/S0007114510004319. [DOI] [PubMed] [Google Scholar]
- 112.Ait-Belgnaoui A, Durand H, Cartier C, Chaumaz G, Eutamene H, Ferrier L, Houdeau E, Fioramonti J, Bueno L, Theodorou V. et al. Prevention of gut leakiness by a probiotic treatment leads to attenuated HPA response to an acute psychological stress in rats. Psychoneuroendocrinology. 2012;37(11):1885–1895. doi: 10.1016/j.psyneuen.2012.03.024. [DOI] [PubMed] [Google Scholar]
- 113.Davis DJ, Doerr HM, Grzelak AK, Busi SB, Jasarevic E, Ericsson AC, Bryda EC. Lactobacillus plantarum attenuates anxiety-related behavior and protects against stress-induced dysbiosis in adult zebrafish. Sci Rep. 2016;6(1):33726. doi: 10.1038/srep33726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Liang S, Wang T, Hu X, Luo J, Li W, Wu X, Duan Y, Jin F. Administration of Lactobacillus helveticus NS8 improves behavioral, cognitive, and biochemical aberrations caused by chronic restraint stress. Neuroscience. 2015;310:561–577. doi: 10.1016/j.neuroscience.2015.09.033. [DOI] [PubMed] [Google Scholar]
- 115.Pusceddu MM, El Aidy S, Crispie F, O’Sullivan O, Cotter P, Stanton C, Kelly P, Cryan JF, Dinan TG. N-3 polyunsaturated fatty acids (PUFAs) reverse the impact of early-life stress on the gut microbiota. PLoS One. 2015;10(10):e0139721. doi: 10.1371/journal.pone.0139721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Schmidt C. Mental health: thinking from the gut. Nature. 2015;518(7540):S12–15. doi: 10.1038/518S13a. [DOI] [PubMed] [Google Scholar]
- 117.Tarr AJ, Galley JD, Fisher S, Chichlowski M, Berg BM, Bailey MT. The prebiotics 3′Sialyllactose and 6′Sialyllactose diminish stressor-induced anxiety-like behavior and colonic microbiota alterations: evidence for effects on the gut–brain axis. Brain Behav Immun. 2015;50:166–177. doi: 10.1016/j.bbi.2015.06.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Medina-Rodriguez EM, Cruz AA, De Abreu JC, Beurel E. Stress, inflammation, microbiome and depression. Pharmacol Biochem Behav. 2023;227-228:173561. doi: 10.1016/j.pbb.2023.173561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Liu L, Wang H, Chen X, Zhang Y, Zhang H, Xie P. Gut microbiota and its metabolites in depression: from pathogenesis to treatment. EBioMedicine. 2023;90:104527. doi: 10.1016/j.ebiom.2023.104527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Messaoudi M, Violle N, Bisson J-F, Desor D, Javelot H, Rougeot C. Beneficial psychological effects of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in healthy human volunteers. Gut Microbes. 2011;2(4):256–261. doi: 10.4161/gmic.2.4.16108. [DOI] [PubMed] [Google Scholar]
- 121.Kohler O, Petersen L, Mors O, Mortensen PB, Yolken RH, Gasse C, Benros ME. Infections and exposure to anti-infective agents and the risk of severe mental disorders: a nationwide study. Acta Psychiatr Scand. 2017;135(2):97–105. doi: 10.1111/acps.12671. [DOI] [PubMed] [Google Scholar]
- 122.Lavebratt C, Yang LL, Giacobini M, Forsell Y, Schalling M, Partonen T, Gissler M. Early exposure to antibiotic drugs and risk for psychiatric disorders: a population-based study. Transl Psychiatry. 2019;9(1):317. doi: 10.1038/s41398-019-0653-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Lurie I, Yang YX, Haynes K, Mamtani R, Boursi B. Antibiotic exposure and the risk for depression, anxiety, or psychosis: a nested case-control study. J Clin Psychiatry. 2015;76(11):1522–1528. doi: 10.4088/JCP.15m09961. [DOI] [PubMed] [Google Scholar]
- 124.Pouranayatihosseinabad M, Bezabih Y, Hawrelak J, Peterson GM, Veal F, Mirkazemi C. Antibiotic use and the development of depression: a systematic review. J Psychosom Res. 2023;164:111113. doi: 10.1016/j.jpsychores.2022.111113. [DOI] [PubMed] [Google Scholar]
- 125.Marx W, Lane MM, Hockey M, Aslam H, Walder K, Borsini A, Firth J, Pariante CM, Berding K, Cryan JF. et al. Diet and depression: future needs to unlock the potential. Mol Psychiatry. 2022;27(2):778–780. doi: 10.1038/s41380-021-01360-2. [DOI] [PubMed] [Google Scholar]
- 126.Herselman MF, Bailey S, Bobrovskaya L. The effects of stress and diet on the “Brain–gut” and “gut–Brain” pathways in animal models of stress and depression. Int J Mol Sci. 2022;23(4):2013. doi: 10.3390/ijms23042013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Collins SM, Bercik P. The relationship between intestinal microbiota and the central nervous system in normal gastrointestinal function and disease. Gastroenterology. 2009;136(6):2003–2014. doi: 10.1053/j.gastro.2009.01.075. [DOI] [PubMed] [Google Scholar]
- 128.Cryan JF, Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. 2012;13(10):701–712. doi: 10.1038/nrn3346. [DOI] [PubMed] [Google Scholar]
- 129.Foster JA, McVey Neufeld KA. Gut–brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. 2013;36(5):305–312. doi: 10.1016/j.tins.2013.01.005. [DOI] [PubMed] [Google Scholar]
- 130.Kaplan BJ, Rucklidge JJ, Romijn AR, Dolph M. A randomised trial of nutrient supplements to minimise psychological stress after a natural disaster. Psychiatry Res. 2015;228(3):373–379. doi: 10.1016/j.psychres.2015.05.080. [DOI] [PubMed] [Google Scholar]
- 131.Mayer EA, Knight R, Mazmanian SK, Cryan JF, Tillisch K. Gut microbes and the brain: paradigm shift in neuroscience. J Neurosci. 2014;34(46):15490–15496. doi: 10.1523/JNEUROSCI.3299-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Rhee SH, Pothoulakis C, Mayer EA. Principles and clinical implications of the brain–gut–enteric microbiota axis. Nat Rev Gastroenterol Hepatol. 2009;6(5):306–314. doi: 10.1038/nrgastro.2009.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Simpson CA, Diaz-Arteche C, Eliby D, Schwartz OS, Simmons JG, Cowan CSM. The gut microbiota in anxiety and depression – a systematic review. Clin Psychol Rev. 2021;83:101943. doi: 10.1016/j.cpr.2020.101943. [DOI] [PubMed] [Google Scholar]
- 134.Liu L, Wang H, Zhang H, Chen X, Zhang Y, Wu J, Zhao L, Wang D, Pu J, Ji P. et al. Toward a deeper understanding of gut microbiome in depression: the promise of clinical applicability. Adv Sci (Weinh). 2022;9(35):e2203707. doi: 10.1002/advs.202203707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Zheng P, Yang J, Li Y, Wu J, Liang W, Yin B, Tan X, Huang Y, Chai T, Zhang H. et al. Gut Microbial Signatures Can Discriminate Unipolar from Bipolar Depression. Adv Sci (Weinh). 2020;7(7):1902862. doi: 10.1002/advs.201902862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 136.Zheng P, Zeng B, Zhou C, Liu M, Fang Z, Xu X, Zeng L, Chen J, Fan S, Du X. et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol Psychiatry. 2016;21(6):786–796. doi: 10.1038/mp.2016.44. [DOI] [PubMed] [Google Scholar]
- 137.Nikolova VL, Smith MRB, Hall LJ, Cleare AJ, Stone JM, Young AH. Perturbations in gut microbiota composition in psychiatric disorders: a review and meta-analysis. JAMA Psychiarty. 2021;78(12):1343–1354. doi: 10.1001/jamapsychiatry.2021.2573. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 138.McGuinness AJ, Davis JA, Dawson SL, Loughman A, Collier F, O’Hely M, Simpson CA, Green J, Marx W, Hair C. et al. A systematic review of gut microbiota composition in observational studies of major depressive disorder, bipolar disorder and schizophrenia. Mol Psychiatry. 2022;27(4):1920–1935. doi: 10.1038/s41380-022-01456-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Medina-Rodriguez EM, Watson J, Reyes J, Trivedi M, Beurel E. Th17 cells sense microbiome to promote depressive-like behaviors. Microbiome. 2023;11(1):1–4. in press. doi: 10.1186/s40168-022-01428-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 140.Nicolas GR, Chang PV. Deciphering the chemical lexicon of Host–gut microbiota interactions. Trends Pharmacol Sci. 2019;40(6):430–445. doi: 10.1016/j.tips.2019.04.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 141.Sun N, Zhang J, Wang J, Liu Z, Wang X, Kang P, Yang C, Liu P, Zhang K. Abnormal gut microbiota and bile acids in patients with first-episode major depressive disorder and correlation analysis. Psychiatry Clin Neurosci. 2022;76(7):321–328. doi: 10.1111/pcn.13368. [DOI] [PubMed] [Google Scholar]
- 142.Skonieczna-Zydecka K, Grochans E, Maciejewska D, Szkup M, Schneider-Matyka D, Jurczak A, Łoniewski I, Kaczmarczyk M, Marlicz W, Czerwińska-Rogowska M. et al. Faecal short chain fatty acids profile is changed in Polish Depressive Women. Nutrients. 2018;10(12):1939. doi: 10.3390/nu10121939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.van de Wouw M, Boehme M, Lyte JM, Wiley N, Strain C, O’Sullivan O, Clarke G, Stanton C, Dinan TG, Cryan JF. et al. Short-chain fatty acids: microbial metabolites that alleviate stress-induced brain–gut axis alterations. J Physiol. 2018;596(20):4923–4944. doi: 10.1113/JP276431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Caspani G, Kennedy S, Foster JA, Swann J. Gut microbial metabolites in depression: understanding the biochemical mechanisms. Microb Cell. 2019;6(10):454–481. doi: 10.15698/mic2019.10.693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Meinitzer S, Baranyi A, Holasek S, Schnedl WJ, Zelzer S, Mangge H, Herrmann M, Meinitzer A, Enko D. Sex-Specific Associations of Trimethylamine-N-Oxide and Zonulin with signs of depression in carbohydrate malabsorbers and nonmalabsorbers. Dis Markers. 2020;2020:1–6. doi: 10.1155/2020/7897240. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Li T, Zheng LN, Han XH. Fenretinide attenuates lipopolysaccharide (LPS)-induced blood-brain barrier (BBB) and depressive-like behavior in mice by targeting nrf-2 signaling. Biomed Pharmacother. 2020;125:109680. doi: 10.1016/j.biopha.2019.109680. [DOI] [PubMed] [Google Scholar]
- 147.Rudzki L, Stone TW, Maes M, Misiak B, Samochowiec J, Szulc A. Gut microbiota-derived vitamins – underrated powers of a multipotent ally in psychiatric health and disease. Prog Neuropsychopharmacol Biol Psychiatry. 2021;107:110240. doi: 10.1016/j.pnpbp.2020.110240. [DOI] [PubMed] [Google Scholar]
- 148.Ortega MA, Alvarez-Mon MA, García-Montero C, Fraile-Martinez O, Guijarro LG, Lahera G, Monserrat J, Valls P, Mora F, Rodríguez-Jiménez R. et al. Gut microbiota metabolites in Major depressive disorder—deep insights into their pathophysiological role and potential translational applications. Metabolites. 2022;12(1):50. doi: 10.3390/metabo12010050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Magnussen A, Parsi MA. Aflatoxins, hepatocellular carcinoma and public health. World J Gastroenterol. 2013;19(10):1508–1512. doi: 10.3748/wjg.v19.i10.1508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Bills GF, Gloer JB, Heitman J, Howlett BJ, Stukenbrock EH. Biologically active secondary metabolites from the fungi. Microbiol Spectr. 2016;4(6):4–6. doi: 10.1128/microbiolspec.FUNK-0009-2016. [DOI] [PubMed] [Google Scholar]
- 151.von Wartburg A, Traber R. Cyclosporins, fungal metabolites with immunosuppressive activities. Prog Med Chem. 1988;25:1–33. [DOI] [PubMed] [Google Scholar]
- 152.Lowe H, Toyang N, Steele B, Valentine H, Grant J, Ali A, Ngwa W, Gordon L. The therapeutic potential of psilocybin. Molecules. 2021;26(10):2948. doi: 10.3390/molecules26102948. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Mukherjee S, Bassler BL. Bacterial quorum sensing in complex and dynamically changing environments. Nat Rev Microbiol. 2019;17(6):371–382. doi: 10.1038/s41579-019-0186-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Rutherford ST, Bassler BL. Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb Perspect Med. 2012;2(11):a012427–a012427. doi: 10.1101/cshperspect.a012427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Thompson JA, Oliveira RA, Djukovic A, Ubeda C, Xavier KB. Manipulation of the quorum sensing signal AI-2 affects the antibiotic-treated gut microbiota. Cell Rep. 2015;10(11):1861–1871. doi: 10.1016/j.celrep.2015.02.049. [DOI] [PubMed] [Google Scholar]
- 156.Ng WL, Bassler BL. Bacterial quorum-sensing network architectures. Annu Rev Genet. 2009;43(1):197–222. doi: 10.1146/annurev-genet-102108-134304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Valdes AM, Walter J, Segal E, Spector TD. Role of the gut microbiota in nutrition and health. BMJ. 2018;361:k2179. doi: 10.1136/bmj.k2179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Chen X, Schauder S, Potier N, Van Dorsselaer A, Pelczer I, Bassler BL, Hughson FM. Structural identification of a bacterial quorum-sensing signal containing boron. Nature. 2002;415(6871):545–549. doi: 10.1038/415545a. [DOI] [PubMed] [Google Scholar]
- 159.Miller ST, Xavier KB, Campagna SR, Taga ME, Semmelhack MF, Bassler BL, Hughson FM. Salmonella typhimurium recognizes a chemically distinct form of the bacterial quorum-sensing signal AI-2. Mol Cell. 2004;15(5):677–687. doi: 10.1016/j.molcel.2004.07.020. [DOI] [PubMed] [Google Scholar]
- 160.Pereira CS, Thompson JA, Xavier KB. AI-2-mediated signalling in bacteria. FEMS Microbiol Rev. 2013;37(2):156–181. doi: 10.1111/j.1574-6976.2012.00345.x. [DOI] [PubMed] [Google Scholar]
- 161.Surette MG, Miller MB, Bassler BL. Quorum sensing in Escherichia coli, salmonella typhimurium, and Vibrio harveyi: a new family of genes responsible for autoinducer production. Proc Natl Acad Sci USA. 1999;96(4):1639–1644. doi: 10.1073/pnas.96.4.1639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Federle MJ. Autoinducer-2-based chemical communication in bacteria: complexities of interspecies signaling. Contrib Microbiol. 2009;16:18–32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Hammer BK, Bassler BL. Quorum sensing controls biofilm formation in Vibrio cholerae. Mol Microbiol. 2003;50(1):101–104. doi: 10.1046/j.1365-2958.2003.03688.x. [DOI] [PubMed] [Google Scholar]
- 164.Perez-Pascual D, Monnet V, Gardan R. Bacterial Cell-Cell Communication in the Host via RRNPP Peptide-Binding Regulators. Front Microbiol. 2016;7:706. doi: 10.3389/fmicb.2016.00706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Dominguez-Bello MG, Godoy-Vitorino F, Knight R, Blaser MJ. Role of the microbiome in human development. Gut. 2019;68(6):1108–1114. doi: 10.1136/gutjnl-2018-317503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Lustri BC, Sperandio V, Moreira CG, Andrews-Polymenis HL. Bacterial chat: intestinal metabolites and signals in host-microbiota-pathogen interactions. Infect Immun. 2017;85(12):10–128. doi: 10.1128/IAI.00476-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Wynendaele E, Verbeke F, D’Hondt M, Hendrix A, Van De Wiele C, Burvenich C, Peremans K, De Wever O, Bracke M, De Spiegeleer B. et al. Crosstalk between the microbiome and cancer cells by quorum sensing peptides. Peptides. 2015;64:40–48. doi: 10.1016/j.peptides.2014.12.009. [DOI] [PubMed] [Google Scholar]
- 168.Durre P, Hollergschwandner C. Initiation of endospore formation in Clostridium acetobutylicum. Anaerobe. 2004;10(2):69–74. doi: 10.1016/j.anaerobe.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 169.Gaida MM, Mayer C, Dapunt U, Stegmaier S, Schirmacher P, Wabnitz GH, Hänsch GM. Expression of the bitter receptor T2R38 in pancreatic cancer: localization in lipid droplets and activation by a bacteria-derived quorum-sensing molecule. Oncotarget. 2016;7(11):12623–12632. doi: 10.18632/oncotarget.7206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 170.De Spiegeleer B, Verbeke F, D’Hondt M, Hendrix A, Van De Wiele C, Burvenich C, Peremans K, De Wever O, Bracke M, Wynendaele E. et al. The quorum sensing peptides PhrG, CSP and EDF promote angiogenesis and invasion of breast cancer cells in vitro. PloS One. 2015;10(3):e0119471. doi: 10.1371/journal.pone.0119471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Gensollen T, Iyer SS, Kasper DL, Blumberg RS. How colonization by microbiota in early life shapes the immune system. Science. 2016;352(6285):539–544. doi: 10.1126/science.aad9378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Spadoni I, Zagato E, Bertocchi A, Paolinelli R, Hot E, Di Sabatino A, Caprioli F, Bottiglieri L, Oldani A, Viale G. et al. A gut-vascular barrier controls the systemic dissemination of bacteria. Science. 2015;350(6262):830–834. doi: 10.1126/science.aad0135. [DOI] [PubMed] [Google Scholar]
- 173.Vighi G, Marcucci F, Sensi L, Di Cara G, Frati F. Allergy and the gastrointestinal system. Clin Exp Immunol. 2008;Suppl 153(Supplement_1):3–6. doi: 10.1111/j.1365-2249.2008.03713.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 174.Macpherson AJ, Uhr T. Induction of protective IgA by intestinal dendritic cells carrying commensal bacteria. Science. 2004;303(5664):1662–1665. doi: 10.1126/science.1091334. [DOI] [PubMed] [Google Scholar]
- 175.Macpherson AJ, Harris NL. Interactions between commensal intestinal bacteria and the immune system. Nat Rev Immunol. 2004;4(6):478–485. doi: 10.1038/nri1373. [DOI] [PubMed] [Google Scholar]
- 176.Hapfelmeier S, Lawson MAE, Slack E, Kirundi JK, Stoel M, Heikenwalder M, Cahenzli J, Velykoredko Y, Balmer ML, Endt K. et al. Reversible microbial colonization of germ-free mice reveals the dynamics of IgA immune responses. Science. 2010;328(5986):1705–1709. doi: 10.1126/science.1188454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 177.Yang Y, Torchinsky MB, Gobert M, Xiong H, Xu M, Linehan JL, Alonzo F, Ng C, Chen A, Lin X. et al. Focused specificity of intestinal TH17 cells towards commensal bacterial antigens. Nature. 2014;510(7503):152–156. doi: 10.1038/nature13279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Ivanov II, Atarashi K, Manel N, Brodie EL, Shima T, Karaoz U, Wei D, Goldfarb KC, Santee CA, Lynch SV. et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell. 2009;139(3):485–498. doi: 10.1016/j.cell.2009.09.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Campion SL, Brodie TM, Fischer W, Korber BT, Rossetti A, Goonetilleke N, McMichael AJ, Sallusto F. Proteome-wide analysis of HIV-specific naive and memory CD4(+) T cells in unexposed blood donors. J Exp Med. 2014;211(7):1273–1280. doi: 10.1084/jem.20130555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 180.Round JL, O’Connell RM, Mazmanian SK. Coordination of tolerogenic immune responses by the commensal microbiota. J Autoimmun. 2010;34(3):J220–J225. doi: 10.1016/j.jaut.2009.11.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Su LF, Kidd BA, Han A, Kotzin JJ, Davis MM. Virus-specific CD4(+) memory- phenotype T cells are abundant in unexposed adults. Immunity. 2013;38(2):373–383. doi: 10.1016/j.immuni.2012.10.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y, Cheng G, Yamasaki S, Saito T, Ohba Y. et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. 2011;331(6015):337–341. doi: 10.1126/science.1198469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 183.Nagano Y, Itoh K, Honda K. The induction of Treg cells by gut-indigenous clostridium. Curr Opin Immunol. 2012;24(4):392–397. doi: 10.1016/j.coi.2012.05.007. [DOI] [PubMed] [Google Scholar]
- 184.Wesemann DR, Portuguese AJ, Meyers RM, Gallagher MP, Cluff-Jones K, Magee JM, Panchakshari RA, Rodig SJ, Kepler TB, Alt FW. et al. Microbial colonization influences early B-lineage development in the gut lamina propria. Nature. 2013;501(7465):112–115. doi: 10.1038/nature12496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 185.Kappler K, Hennet T. Emergence and significance of carbohydrate-specific antibodies. Genes Immun. 2020;21(4):224–239. doi: 10.1038/s41435-020-0105-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 186.Cahenzli J, Koller Y, Wyss M, Geuking MB, McCoy KD. Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host Microbe. 2013;14(5):559–570. doi: 10.1016/j.chom.2013.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 187.Fulde M, Sommer F, Chassaing B, van Vorst K, Dupont A, Hensel M, Basic M, Klopfleisch R, Rosenstiel P, Bleich A. et al. Neonatal selection by Toll-like receptor 5 influences long-term gut microbiota composition. Nature. 2018;560(7719):489–493. doi: 10.1038/s41586-018-0395-5. [DOI] [PubMed] [Google Scholar]
- 188.Huh JR, Veiga-Fernandes H. Neuroimmune circuits in inter-organ communication. Nat Rev Immunol. 2020;20(4):217–228. doi: 10.1038/s41577-019-0247-z. [DOI] [PubMed] [Google Scholar]
- 189.Mowat AM. Historical perspective: Metchnikoff and the intestinal microbiome. J Leukoc Biol. 2021;109(3):513–517. doi: 10.1002/JLB.4RI0920-599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 190.Jacobson A, Yang D, Vella M, Chiu IM. The intestinal neuro-immune axis: crosstalk between neurons, immune cells, and microbes. Mucosal Immunol. 2021;14(3):555–565. doi: 10.1038/s41385-020-00368-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Ghosh SS, Wang J, Yannie PJ, Ghosh S. Intestinal barrier dysfunction, LPS translocation, and disease development. J Endocr Soc. 2020;4(2):bvz039. doi: 10.1210/jendso/bvz039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Janssen AW, Kersten S. Potential mediators linking gut bacteria to metabolic health: a critical view. J Physiol. 2017;595(2):477–487. doi: 10.1113/JP272476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Dantzer R, O’Connor JC, Freund GG, Johnson RW, Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci. 2008;9(1):46–56. doi: 10.1038/nrn2297. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Maslanik T, Tannura K, Mahaffey L, Loughridge AB, Benninson L, Ursell L, Greenwood BN, Knight R, Fleshner M. Commensal bacteria and MAMPs are necessary for stress-induced increases in IL-1β and IL-18 but not IL-6, IL-10 or MCP-1. PloS One. 2012;7(12):e50636. doi: 10.1371/journal.pone.0050636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 195.Garcia-Rodenas CL, Bergonzelli GE, Nutten S, Schumann A, Cherbut C, Turini M, Ornstein K, Rochat F, Corthésy‐Theulaz I. Nutritional approach to restore impaired intestinal barrier function and growth after neonatal stress in rats. J Pediatr Gastroenterol Nutr. 2006;43(1):16–24. doi: 10.1097/01.mpg.0000226376.95623.9f. [DOI] [PubMed] [Google Scholar]
- 196.Teitelbaum AA, Gareau MG, Jury J, Yang PC, Perdue MH. Chronic peripheral administration of corticotropin-releasing factor causes colonic barrier dysfunction similar to psychological stress. Am J Physiol Gastrointest Liver Physiol. 2008;295(3):G452–G459. doi: 10.1152/ajpgi.90210.2008. [DOI] [PubMed] [Google Scholar]
- 197.Reichenberg A, Yirmiya R, Schuld A, Kraus T, Haack M, Morag A, Pollmächer T. Cytokine-associated emotional and cognitive disturbances in humans. Arch Gen Psychiatry. 2001;58(5):445–452. doi: 10.1001/archpsyc.58.5.445. [DOI] [PubMed] [Google Scholar]
- 198.Bouskra D, Brézillon C, Bérard M, Werts C, Varona R, Boneca IG, Eberl G. Lymphoid tissue genesis induced by commensals through NOD1 regulates intestinal homeostasis. Nature. 2008;456(7221):507–510. doi: 10.1038/nature07450. [DOI] [PubMed] [Google Scholar]
- 199.Ramanan D, Tang MS, Bowcutt R, Loke P, Cadwell K. Bacterial sensor Nod2 prevents inflammation of the small intestine by restricting the expansion of the commensal bacteroides vulgatus. Immunity. 2014;41(2):311–324. doi: 10.1016/j.immuni.2014.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Nigro G, Rossi R, Commere PH, Jay P, Sansonetti PJ. The cytosolic bacterial peptidoglycan sensor Nod2 affords stem cell protection and links microbes to gut epithelial regeneration. Cell Host Microbe. 2014;15(6):792–798. doi: 10.1016/j.chom.2014.05.003. [DOI] [PubMed] [Google Scholar]
- 201.Napier RJ, Lee EJ, Davey MP, Vance EE, Furtado JM, Snow PE, Samson KA, Lashley SJ, Brown BR, Horai R. et al. T cell-intrinsic role for Nod2 in protection against Th17-mediated uveitis. Nat Commun. 2020;11(1):5406. doi: 10.1038/s41467-020-18961-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Farzi A, Reichmann F, Meinitzer A, Mayerhofer R, Jain P, Hassan AM, Fröhlich EE, Wagner K, Painsipp E, Rinner B. et al. Synergistic effects of NOD1 or NOD2 and TLR4 activation on mouse sickness behavior in relation to immune and brain activity markers. Brain Behav Immun. 2015;44:106–120. doi: 10.1016/j.bbi.2014.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 203.Pusceddu MM, Barboza M, Keogh CE, Schneider M, Stokes P, Sladek JA, Kim HJD, Torres‐Fuentes C, Goldfild LR, Gillis SE. et al. Nod-like receptors are critical for gut–brain axis signalling in mice. J Physiol. 2019;597(24):5777–5797. doi: 10.1113/JP278640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 204.Stockinger B, Shah K, Wincent E. AHR in the intestinal microenvironment: safeguarding barrier function. Nat Rev Gastroenterol Hepatol. 2021;18(8):559–570. doi: 10.1038/s41575-021-00430-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 205.Safe S, Jin UH, Park H, Chapkin RS, Jayaraman A. Aryl Hydrocarbon Receptor (AHR) Ligands as Selective AHR Modulators (SAhRMs). Int J Mol Sci. 2020;21(18):6654. doi: 10.3390/ijms21186654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Safe S, Han H, Goldsby J, Mohankumar K, Chapkin RS. Aryl Hydrocarbon Receptor (AhR) Ligands as Selective AhR Modulators: Genomic Studies. Curr Opin Toxicol. 2018;11-12:10–20. doi: 10.1016/j.cotox.2018.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 207.Grifka-Walk HM, Jenkins BR, Kominsky DJ. Amino acid trp: the far out impacts of host and commensal tryptophan metabolism. Front Immunol. 2021;12:653208. doi: 10.3389/fimmu.2021.653208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Madison CA, Hillbrick L, Kuempel J, Albrecht GL, Landrock KK, Safe S, Chapkin RS, Eitan S. Intestinal epithelium aryl hydrocarbon receptor is involved in stress sensitivity and maintaining depressive symptoms. Behav Brain Res. 2023;440:114256. doi: 10.1016/j.bbr.2022.114256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 209.Madison CA, Kuempel J, Albrecht GL, Hillbrick L, Jayaraman A, Safe S, Chapkin RS, Eitan S. 3,3′-diindolylmethane and 1,4-dihydroxy-2-naphthoic acid prevent chronic mild stress induced depressive-like behaviors in female mice. J Affect Disord. 2022;309:201–210. doi: 10.1016/j.jad.2022.04.106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 210.Willing BP, Vacharaksa A, Croxen M, Thanachayanont T, Finlay BB, Bereswill S. Altering host resistance to infections through microbial transplantation. PloS One. 2011;6(10):e26988. doi: 10.1371/journal.pone.0026988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Dinan TG, Stanton C, Cryan JF. Psychobiotics: a novel class of psychotropic. Biol Psychiatry. 2013;74(10):720–726. doi: 10.1016/j.biopsych.2013.05.001. [DOI] [PubMed] [Google Scholar]
- 212.Bilbo SD, Schwarz JM. The immune system and developmental programming of brain and behavior. Front Neuroendocrinol. 2012;33(3):267–286. doi: 10.1016/j.yfrne.2012.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Macpherson AJ, Uhr T. Gut flora--mechanisms of regulation. Eur J Surg Suppl. 2002;1(587):53–57. [PubMed] [Google Scholar]
- 214.Xu C, Lee SK, Zhang D, Frenette PS. The gut microbiome regulates psychological-stress-induced inflammation. Immunity. 2020;53(2):417–428.e4. doi: 10.1016/j.immuni.2020.06.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Zhu X, Sakamoto S, Ishii C, Smith MD, Ito K, Obayashi M, Unger L, Hasegawa Y, Kurokawa S, Kishimoto T. et al. Dectin-1 signaling on colonic γδ T cells promotes psychosocial stress responses. Nat Immunol. 2023;24(4):625–636. doi: 10.1038/s41590-023-01447-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 216.Alves de Lima K, Rustenhoven J, Da Mesquita S, Wall M, Salvador AF, Smirnov I, Martelossi Cebinelli G, Mamuladze T, Baker W, Papadopoulos Z. et al. Meningeal γδ T cells regulate anxiety-like behavior via IL-17a signaling in neurons. Nat Immunol. 2020;21(11):1421–1429. doi: 10.1038/s41590-020-0776-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 217.Kelly JR, Clarke G, Cryan JF, Dinan TG. Brain-gut-microbiota axis: challenges for translation in psychiatry. Ann Epidemiol. 2016;26(5):366–372. doi: 10.1016/j.annepidem.2016.02.008. [DOI] [PubMed] [Google Scholar]
- 218.Dhabhar FS, Malarkey WB, Neri E, McEwen BS. Stress-induced redistribution of immune cells—from barracks to boulevards to battlefields: a tale of three hormones – Curt Richter award winner. Psychoneuroendocrinology. 2012;37(9):1345–1368. doi: 10.1016/j.psyneuen.2012.05.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.van de Wouw M, Lyte JM, Boehme M, Sichetti M, Moloney G, Goodson MS, Kelley-Loughnane N, Dinan TG, Clarke G, Cryan JF. et al. The role of the microbiota in acute stress-induced myeloid immune cell trafficking. Brain Behav Immun. 2020;84:209–217. doi: 10.1016/j.bbi.2019.12.003. [DOI] [PubMed] [Google Scholar]
- 220.Balmer ML, Schürch CM, Saito Y, Geuking MB, Li H, Cuenca M, Kovtonyuk LV, McCoy KD, Hapfelmeier S, Ochsenbein AF. et al. Microbiota-derived compounds drive steady-state granulopoiesis via MyD88/TICAM signaling. J Immunol. 2014;193(10):5273–5283. doi: 10.4049/jimmunol.1400762. [DOI] [PubMed] [Google Scholar]
- 221.Khosravi A, Yáñez A, Price J, Chow A, Merad M, Goodridge H, Mazmanian S. Gut microbiota promote hematopoiesis to control bacterial infection. Cell Host Microbe. 2014;15(3):374–381. doi: 10.1016/j.chom.2014.02.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Karbach SH, Schönfelder T, Brandão I, Wilms E, Hörmann N, Jäckel S, Schüler R, Finger S, Knorr M, Lagrange J. et al. Gut Microbiota Promote Angiotensin II–Induced Arterial Hypertension and Vascular Dysfunction. J Am Heart Assoc. 2016;5(9):e003698. doi: 10.1161/JAHA.116.003698. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 223.Mohle L, Mattei D, Heimesaat M, Bereswill S, Fischer A, Alutis M, French T, Hambardzumyan D, Matzinger P, Dunay I. et al. Ly6C(hi) monocytes provide a link between Antibiotic-Induced Changes in gut microbiota and adult hippocampal neurogenesis. Cell Rep. 2016;15(9):1945–1956. doi: 10.1016/j.celrep.2016.04.074. [DOI] [PubMed] [Google Scholar]
- 224.Sweere JM, Van Belleghem JD, Ishak H, Bach MS, Popescu M, Sunkari V, Kaber G, Manasherob R, Suh GA, Cao X. et al. Bacteriophage trigger antiviral immunity and prevent clearance of bacterial infection. Science. 2019;363(6434):eaat9691. doi: 10.1126/science.aat9691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 225.Gogokhia L, Buhrke K, Bell R, Hoffman B, Brown DG, Hanke-Gogokhia C, Ajami NJ, Wong MC, Ghazaryan A, Valentine JF. et al. Expansion of bacteriophages is linked to aggravated intestinal inflammation and colitis. Cell Host Microbe. 2019;25(2):285–299 e288. doi: 10.1016/j.chom.2019.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 226.Segain JP, Kornprobst M, Siavoshian S. Butyrate inhibits inflammatory responses through NFkappaB inhibition: implications for Crohn’s disease. Gut. 2000;47(3):397–403. doi: 10.1136/gut.47.3.397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 227.Luhrs H, Gerke T, Schauber J, Dusel G, Melcher R, Scheppach W, Menzel T. Cytokine-activated degradation of inhibitory κB protein α is inhibited by the short-chain fatty acid butyrate. Int J Colorectal Dis. 2001;16(4):195–201. doi: 10.1007/s003840100295. [DOI] [PubMed] [Google Scholar]
- 228.Provensi G, Schmidt SD, Boehme M, Bastiaanssen TFS, Rani B, Costa A, Busca K, Fouhy F, Strain C, Stanton C. et al. Preventing adolescent stress-induced cognitive and microbiome changes by diet. Proc Natl Acad Sci USA. 2019;116(19):9644–9651. doi: 10.1073/pnas.1820832116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 229.Tang CF, Wang C-Y, Wang J-H, Wang Q-N, Li S-J, Wang H-O, Zhou F, Li J-M. Short-chain fatty acids ameliorate depressive-like behaviors of high fructose-fed mice by rescuing hippocampal neurogenesis decline and blood–brain barrier damage. Nutrients. 2022;14(9):1882. doi: 10.3390/nu14091882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 230.Alkahtani SH. The steroidal Na+/K+ ATPase inhibitor 3-[(R)-3-pyrrolidinyl]oxime derivative (3-R-POD) induces potent pro-apoptotic responses in colonic tumor cells. Anticancer Res. 2014;34:2967–2971. [PubMed] [Google Scholar]
- 231.Kabouridis PS, Lasrado R, McCallum S, Chng SH, Snippert HJ, Clevers H, Pettersson S, Pachnis V. The gut microbiota keeps enteric glial cells on the move; prospective roles of the gut epithelium and immune system. Gut Microbes. 2015;6(6):398–403. doi: 10.1080/19490976.2015.1109767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 232.Rosenbaum C, Schick MA, Wollborn J, Heider A, Scholz C-J, Cecil A, Niesler B, Hirrlinger J, Walles H, Metzger M. et al. Activation of Myenteric Glia during acute inflammation in vitro and in vivo. PLoS One. 2016;11(3):e0151335. doi: 10.1371/journal.pone.0151335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 233.Ibiza S, García-Cassani B, Ribeiro H, Carvalho T, Almeida L, Marques R, Misic AM, Bartow-McKenney C, Larson DM, Pavan WJ. et al. Glial-cell-derived neuroregulators control type 3 innate lymphoid cells and gut defence. Nature. 2016;535(7612):440–443. doi: 10.1038/nature18644. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 234.Jackerott M, Larsson LI. Immunocytochemical localization of the NPY/PYY Y1 receptor in enteric neurons, endothelial cells, and endocrine-like cells of the rat intestinal tract. J Histochem Cytochem. 1997;45(12):1643–1650. doi: 10.1177/002215549704501207. [DOI] [PubMed] [Google Scholar]
- 235.Terry N, Margolis KG. Serotonergic mechanisms regulating the GI Tract: experimental evidence and therapeutic relevance. Handb Exp Pharmacol. 2017;239:319–342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 236.Ji XB, Hollocher TC. Reduction of nitrite to nitric oxide by enteric bacteria. Biochem Biophys Res Commun. 1988;157(1):106–108. doi: 10.1016/S0006-291X(88)80018-4. [DOI] [PubMed] [Google Scholar]
- 237.Stanaszek PM, Snell JF, O’Neill JJ. Isolation, extraction, and measurement of acetylcholine from Lactobacillus plantarum. Appl Environ Microbiol. 1977;34(2):237–239. doi: 10.1128/aem.34.2.237-239.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 238.Tsavkelova EA, Botvinko IV, Kudrin VS, Oleskin AV. Detection of neurotransmitter amines in microorganisms with the use of high-performance liquid chromatography. Dokl Biochem. 2000;372:115–117. [PubMed] [Google Scholar]
- 239.Spielman LJ, Gibson DL, Klegeris A. Unhealthy gut, unhealthy brain: the role of the intestinal microbiota in neurodegenerative diseases. Neurochem Int. 2018;120:149–163. doi: 10.1016/j.neuint.2018.08.005. [DOI] [PubMed] [Google Scholar]
- 240.Barrett E, Ross RP, O’Toole PW, Fitzgerald GF, Stanton C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J Appl Microbiol. 2012;113(2):411–417. doi: 10.1111/j.1365-2672.2012.05344.x. [DOI] [PubMed] [Google Scholar]
- 241.Barajon I, Serrao G, Arnaboldi F, Opizzi E, Ripamonti G, Balsari A, Rumio C. Toll-like receptors 3, 4, and 7 are expressed in the enteric nervous system and dorsal root ganglia. J Histochem Cytochem. 2009;57(11):1013–1023. doi: 10.1369/jhc.2009.953539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 242.Brun P, Giron MC, Qesari M, Porzionato A, Caputi V, Zoppellaro C, Banzato S, Grillo AR, Spagnol L, De Caro R. et al. Toll-like receptor 2 regulates intestinal inflammation by controlling integrity of the enteric nervous system. Gastroenterology. 2013;145(6):1323–1333. doi: 10.1053/j.gastro.2013.08.047. [DOI] [PubMed] [Google Scholar]
- 243.Luczynski P, McVey Neufeld K-A, Oriach CS, Clarke G, Dinan TG, Cryan JF. Growing up in a bubble: using germ-free animals to assess the influence of the gut microbiota on brain and behavior. Int J Neuropsychopharmacol. 2016;19(8):pyw020. doi: 10.1093/ijnp/pyw020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 244.Gareau MG. Microbiota-gut-brain axis and cognitive function. Adv Exp Med Biol. 2014;817:357–371. [DOI] [PubMed] [Google Scholar]
- 245.Sampson TR, Mazmanian SK. Control of brain development, function, and behavior by the microbiome. Cell Host Microbe. 2015;17(5):565–576. doi: 10.1016/j.chom.2015.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 246.Hoban AE, Stilling RM, Ryan FJ, Shanahan F, Dinan TG, Claesson MJ, Clarke G, Cryan JF. Regulation of prefrontal cortex myelination by the microbiota. Transl Psychiatry. 2016;6(4):e774. doi: 10.1038/tp.2016.42. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 247.Gacias M, Gaspari S, Santos PMG, Tamburini S, Andrade M, Zhang F, Shen N, Tolstikov V, Kiebish MA, Dupree JL. et al. Microbiota-driven transcriptional changes in prefrontal cortex override genetic differences in social behavior. Elife. 2016;5. doi: 10.7554/eLife.13442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 248.Luczynski P, Whelan SO, O’Sullivan C, Clarke G, Shanahan F, Dinan TG, Cryan JF. Adult microbiota-deficient mice have distinct dendritic morphological changes: differential effects in the amygdala and hippocampus. Eur J Neurosci. 2016;44(9):2654–2666. doi: 10.1111/ejn.13291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 249.Braniste V, Al-Asmakh M, Kowal C, Anuar F, Abbaspour A, Tóth M, Korecka A, Bakocevic N, Ng LG, Kundu P. et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci Transl Med. 2014;6(263):263ra158. doi: 10.1126/scitranslmed.3009759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 250.Frohlich EE, Farzi A, Mayerhofer R, Reichmann F, Jačan A, Wagner B, Zinser E, Bordag N, Magnes C, Fröhlich E. et al. Cognitive impairment by antibiotic-induced gut dysbiosis: analysis of gut microbiota-brain communication. Brain Behav Immun. 2016;56:140–155. doi: 10.1016/j.bbi.2016.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 251.Li H, Sun J, Du J, Wang F, Fang R, Yu C, Xiong J, Chen W, Lu Z, Liu J. et al. Clostridium butyricum exerts a neuroprotective effect in a mouse model of traumatic brain injury via the gut-brain axis. Neurogastroenterol Motil. 2018;30(5):e13260. doi: 10.1111/nmo.13260. [DOI] [PubMed] [Google Scholar]
- 252.Li H, Sun J, Wang F, Ding G, Chen W, Fang R, Yao Y, Pang M, Lu Z-Q, Liu J. et al. Sodium butyrate exerts neuroprotective effects by restoring the blood-brain barrier in traumatic brain injury mice. Brain Res. 2016;1642:70–78. doi: 10.1016/j.brainres.2016.03.031. [DOI] [PubMed] [Google Scholar]
- 253.Kim HJ, Rowe M, Ren M, Hong J-S, Chen P-S, Chuang D-M. Histone deacetylase inhibitors exhibit anti-inflammatory and neuroprotective effects in a rat permanent ischemic model of stroke: multiple mechanisms of action. J Pharmacol Exp Ther. 2007;321(3):892–901. doi: 10.1124/jpet.107.120188. [DOI] [PubMed] [Google Scholar]
- 254.Agus A, Planchais J, Sokol H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe. 2018;23(6):716–724. doi: 10.1016/j.chom.2018.05.003. [DOI] [PubMed] [Google Scholar]
- 255.Williams BB, Van Benschoten A, Cimermancic P, Donia M, Zimmermann M, Taketani M, Ishihara A, Kashyap P, Fraser J, Fischbach M. et al. Discovery and characterization of gut microbiota decarboxylases that can produce the neurotransmitter tryptamine. Cell Host Microbe. 2014;16(4):495–503. doi: 10.1016/j.chom.2014.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 256.Yano JM, Yu K, Donaldson G, Shastri G, Ann P, Ma L, Nagler C, Ismagilov R, Mazmanian S, Hsiao E. et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. 2015;161(2):264–276. doi: 10.1016/j.cell.2015.02.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 257.Desbonnet L, Garrett L, Clarke G, Bienenstock J, Dinan TG. The probiotic bifidobacteria infantis: an assessment of potential antidepressant properties in the rat. J Psychiatr Res. 2008;43(2):164–174. doi: 10.1016/j.jpsychires.2008.03.009. [DOI] [PubMed] [Google Scholar]
- 258.Alkhalaf LM, Ryan KS. Biosynthetic manipulation of tryptophan in bacteria: pathways and mechanisms. Chem Biol. 2015;22(3):317–328. doi: 10.1016/j.chembiol.2015.02.005. [DOI] [PubMed] [Google Scholar]
- 259.Kaszaki J, Érces D, Varga G, Szabó A, Vécsei L, Boros M. Kynurenines and intestinal neurotransmission: the role of N-methyl-D- aspartate receptors. J Neural Transm (Vienna). 2012;119(2):211–223. doi: 10.1007/s00702-011-0658-x. [DOI] [PubMed] [Google Scholar]
- 260.Stone TW, Darlington LG. The kynurenine pathway as a therapeutic target in cognitive and neurodegenerative disorders. Br J Pharmacol. 2013;169(6):1211–1227. doi: 10.1111/bph.12230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 261.Rothhammer V, Mascanfroni ID, Bunse L, Takenaka MC, Kenison JE, Mayo L, Chao C-C, Patel B, Yan R, Blain M. et al. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat Med. 2016;22(6):586–597. doi: 10.1038/nm.4106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 262.Saito K, Markey SP, Heyes MP. Effects of immune activation on quinolinic acid and neuroactive kynurenines in the mouse. Neuroscience. 1992;51(1):25–39. doi: 10.1016/0306-4522(92)90467-G. [DOI] [PubMed] [Google Scholar]
- 263.Fujigaki H, Yamamoto Y, Saito K. L-Tryptophan-kynurenine pathway enzymes are therapeutic target for neuropsychiatric diseases: focus on cell type differences. Neuropharmacology. 2017;112:264–274. doi: 10.1016/j.neuropharm.2016.01.011. [DOI] [PubMed] [Google Scholar]
- 264.Mayer EA, Tillisch K, Gupta A. Gut/brain axis and the microbiota. J Clin Invest. 2015;125(3):926–938. doi: 10.1172/JCI76304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 265.Fung TC, Olson CA, Hsiao EY. Interactions between the microbiota, immune and nervous systems in health and disease. Nat Neurosci. 2017;20(2):145–155. doi: 10.1038/nn.4476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 266.Thion MS, Low D, Silvin A, Chen J, Grisel P, Schulte-Schrepping J, Blecher R, Ulas T, Squarzoni P, Hoeffel G. et al. Microbiome Influences Prenatal and Adult Microglia in a Sex-Specific Manner. Cell. 2018;172(3):500–516.e16. doi: 10.1016/j.cell.2017.11.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 267.Matcovitch-Natan O, Winter DR, Giladi A, Vargas Aguilar S, Spinrad A, Sarrazin S, Ben-Yehuda H, David E, Zelada González F, Perrin P. et al. Microglia development follows a stepwise program to regulate brain homeostasis. Science. 2016;353(6301):aad8670. doi: 10.1126/science.aad8670. [DOI] [PubMed] [Google Scholar]
- 268.Castillo-Ruiz A, Mosley M, George AJ, Mussaji LF, Fullerton EF, Ruszkowski EM, Jacobs AJ, Gewirtz AT, Chassaing B, Forger NG. et al. The microbiota influences cell death and microglial colonization in the perinatal mouse brain. Brain Behav Immun. 2018;67:218–229. doi: 10.1016/j.bbi.2017.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 269.Li B, Yang W, Ge T, Wang Y, Cui R. Stress induced microglial activation contributes to depression. Pharmacol Res. 2022;179:106145. doi: 10.1016/j.phrs.2022.106145. [DOI] [PubMed] [Google Scholar]