

Abstract
Proteins are highly labile molecules, thus requiring the presence of appropriate solvents and excipients in their liquid milieu to keep their stability and biological activity. In this field, ionic liquids (ILs) have gained momentum in the past years, with a relevant number of works reporting their successful use to dissolve, stabilize, extract, and purify proteins. Different approaches in protein-IL systems have been reported, namely, proteins dissolved in (i) neat ILs, (ii) ILs as co-solvents, (iii) ILs as adjuvants, (iv) ILs as surfactants, (v) ILs as phase-forming components of aqueous biphasic systems, and (vi) IL-polymer-protein/peptide conjugates. Herein, we critically analyze the works published to date and provide a comprehensive understanding of the IL-protein interactions affecting the stability, conformational alteration, unfolding, misfolding, and refolding of proteins while providing directions for future studies in view of imminent applications. Overall, it has been found that the stability or purification of proteins by ILs is bispecific and depends on the structure of both the IL and the protein. The most promising IL-protein systems are identified, which is valuable when foreseeing market applications of ILs, e.g., in “protein packaging” and “detergent applications”. Future directions and other possibilities of IL-protein systems in light-harvesting and biotechnology/biomedical applications are discussed.
1. Introduction
Proteins are among the most incredible creations of nature, performing activities from generation to operation, from the beginning to the end of life.1 Their in vivo operation has inspired their ex vivo applications, such as in healthcare, food,2,3 paper pulp bleaching,4 leather processing,5 biocatalysis,6 detergency,7 and materials.8 However, these applications require proteins in a bioactive form, particularly if storage is envisaged for long periods. Several strategies for the in vitro packaging of proteins have been developed,9−11 such as protein lyophilization with disaccharides; maintenance of their conformational stability in water using salts, osmolytes, or buffers; ligand binding; and interfacial stabilization promoted by surfactants, colloidal stabilization, aerogels, protein cohabitation, pharmacological chaperones, chemical modification, site-directed mutagenesis, protein staple, and use of ionic liquids (ILs).9−11 Among the several strategies investigated, in the past two decades ILs have been a hot topic of research in the field of protein stabilization, dissolution, extraction, and purification.
ILs are salts with an organic cation and organic/inorganic anions, thus having a low lattice energy and low melting points when compared to inorganic salts. Due to their organic nature, ILs are able to establish a wider range of interactions when compared to Coulombic-dominated salts,12 resulting in a series of unique properties. Aprotic ILs, if properly designed, have a negligible vapor pressure at ambient conditions and a high thermal and chemical stability. However, one of their most relevant properties is their designer solvent ability, i.e., solvation customization by choosing suitable cation–anion combinations,13 which has contributed to the observed continuous growth in IL applications.14 These include protein dissolution, stabilization, extraction, and purification in native ILs and their mixtures with molecular solvents.15−17
The ability of ILs and their water mixtures to solvate and to keep the protein’s integrity depends on their physicochemical properties, such as viscosity, polarity, hydrogen-bond donor or acceptor ability, ionicity or ion charge density, and IL concentration, among other factors.18−27 Neat ILs can exist as ion-pairs, directional hydrogen bond networks, ion-clusters, and/or self-assembled nanostructures,27 and all possibilities have a relevant impact on protein solvation and stabilization. In addition to the IL characteristics, the proteins structure, molecular weight, isoelectric point, and conformation are key determinants for their solubility and stability in given ILs. If we apply the “like dissolves like” concept, most proteins are expected to display similar solubility in neat ILs since both exhibit polar/nonpolar domains; yet, this is only true for a few proteins (Lipase, Cytochrome c, Cellulase, Zein, Keratin) and in scarce ILs.28−34 Therefore, both the IL and protein contributions should be taken into account when dealing with neat ILs as promising solvents for the molecular packaging of proteins.
Beyond molecular packaging, protein dispersions in neat ILs have been applied to carry out high-temperature biocatalysis.35,36 Most reports to date on neat IL-protein systems are focused on enzyme biocatalysis, a topic that has been comprehensively reviewed from 2002 to 2021.15,37−52 Because this is beyond the scope of this review, it is here only briefly discussed. These reviews summarized developments in enzyme biocatalysis, achieved mainly with imidazolium-based ILs, highlighting key impediments for practical applications, like high-end cost, the toxicity of the ILs employed, and poor fundamental understanding of IL-enzyme interactions to increase stability and activity of the enzyme. Commonly, less viscous, more hydrophobic, and surface-active ILs, or ILs composed of “kosmotropic” anions and “chaotropic” cations (in accordance with the Hofmeister series53), enhance the activity and stability of enzymes.49 Overall, the thermal stability of ILs is the key property being exploited in biocatalysis, allowing the increase of the reaction rate by causing more collisions between the enzyme and the substrate at high temperatures. In biocatalysis, enzymes need not be dissolved in ILs and can function in their dispersed form if the active sites are accessible.
As stated before, neat ILs exist in hydrogen-bonded supramolecular structures consisting of polar/nonpolar domains.24,54 Upon small dilution with water, marked changes in the physicochemical properties of ILs occur,55,56 which then affect their ability to solubilize and stabilize proteins.57−75 The main advantage of aqueous concentrated-IL solutions (A-ILCSs) for protein packaging is based on the inherent natural environment, in the form of the surrounding water, in addition to the thermal stability afforded by the presence of IL clusters.27,54,76 Nevertheless, opposite findings have been reported, namely on the deleterious effects of ILs on proteins when moving from neat ILs (NILs) to A-ILCS.57,77 Furthermore, in highly diluted solutions, IL ions are expected to behave similarly to conventional inorganic electrolytes; however, inconsistent perceptions have been reported,78−83 in which the associative or dissociative nature of ILs in dilute solutions seems to depend on their hydrophobicity.79 For instance, by molecular dynamics studies, water molecules were reported to be confined at the boundary of the polar and nonpolar of ([C8mim][NO3])/water mixtures, wherein the IL retained its nanodomain structure.81 In contrast, in a later report,83 carried out with 1H NMR studies with quaternary ammonium- and imidazolium-based ILs, water molecules have been shown to be in close proximity and to be confined inside the IL. These inconsistencies further have implications on how ILs and their solutions with other solvents behave and interact with proteins.
When considering the dilute solution of ILs, attempts have been carried out to better understand the effects of ILs on proteins’ stability based on the Hofmeister series and related concepts.84−86 According to the Hofmeister concept, more hydrating (“kosmotrope”) anions, like SO42– and CO32–, and less hydrating (“chaotrope”) cations, like NH4+ and (CH3)4N+, stabilize proteins in aqueous solution, whereas the opposite effect is observed for “kosmotropic” cations (Li+ and Mg2+) and “chaotropic” anions (SCN– and ClO4–).87−102 It should be noted that caution should be taken with the “kosmotropic”/“chaotropic” concepts, which are based on the stabilization or destabilization of the water structure.103 In this field, recent spectroscopic and computer simulation studies have refuted these concepts and tried to justify protein stabilization or destabilization based on the ion-specific interactions of salts with the protein surface rather than by ion–water interactions.97−102 In-depth discussion on the Hofmeister series and its validity based on the “kosmotropic”/“chaotropic” concept is done later in this review. Most of the reported cations of ILs are “chaotropes”, an inherent result of their large organic cation structure. Accordingly, their Hofmeister-based effect on proteins has been scrutinized by considering ion pairs of “chaotropic” cations and “chaotropic” anions or “chaotropic” cations and “kosmotropic” anions. Interestingly, in addition to the traditional specific ion–water interaction phenomenon, the Hofmeister effect of ILs on proteins has been interpreted in the purview of modifying the protein–water interactions, IL-protein interactions, and the effect of IL cations and anions on protein stability and activity.15,16,86 However, contradictions persist to universalize the Hofmeister ranking of IL ions, which is mainly due to the contrasting results obtained with different proteins, thus reinforcing the need for further and systematic revision in the field.
When considering the concept of protein stability, the common perception is that the native conformation of a given protein is thermodynamically the most stable and consequently the most biologically active one.104 However, whether the thermodynamically most stable form is the catalytically most active one is still a disputed point.105 This is because it is determined by the totality of interatomic interactions and hence by the amino acid sequences.106−109 Regarding the thermodynamic vs kinetic control of protein stability in ILs, when proteins are packaged for a long time, thermodynamic control plays a key role since kinetic control depends solely on initial conditions. When short periods are considered instead, for example in biocatalysis, the protein stability is under kinetic control. This is supported by the fact that certain ILs lead to an increase in the activity of enzymes, though their structures are different from their native conformation. ILs provide enough kinetic energy in the system to cross the energy barrier to achieve the most active conformation, which is not at the global energy minima.110
When considering ILs composed of hydrophobic/fluorinated anions, such as [TfO]−, [Tf2N]−, and [PF6]−, it has been shown that local hydrophobic interactions play a major role in protein stability.16 On the other hand, when increasing the hydrophobicity of ILs by increasing the alkyl side chain length of either the IL cation or anion, different behaviors upon the IL ion’s interaction with proteins have been identified.111−140 ILs with an alkyl side chain length with more than 8 carbon atoms (n) have been classified as surface-active ILs (SAILs).141 Some SAILs exhibit superior properties compared to their nonsurface-active counterparts in terms of adsorption behavior, emulsifying tendency, and low aggregation concentration.141,142 Since surfactants exhibit wide applications with proteins,143,144 specifically in detergent and pharma industries, SAIL-protein colloidal formulations were also studied in light of their possible applications in these directions, and they are discussed in a separate section in this review.111−140,145−149 In addition to these systems, investigations on proteins fibrillation, PEGylation, and the formation of protein/peptide–polymer conjugates as ways of improving the solubility and stability of proteins in NILs, either for nonaqueous biocatalysis or kinetic storage, have also been discussed.150−153
ILs have been used as well to form liquid two-phase systems with water or organic solvents, while envisioning the development of separation processes for proteins/enzymes.154,155 Among these, IL-based aqueous biphasic systems (IL-ABS, ternary systems typically composed of water, ILs, and salts) hold advantages over other systems based on more hydrophobic ILs due to their large water content, a beneficial aspect when dealing with most proteins. Therefore, progress made with IL-ABS in pursuit of protein extraction processes156−158 is also reviewed in this work.
Before the inception of the IL era, there were already strategies available for the stabilization and packaging of proteins. Therefore, the first section of this review is focused on providing a rational understanding behind the protein’s stability afforded by more conventional strategies. The stronger and weaker links of these strategies are revealed, and limitations that can be overcome by ILs are discussed. The following section covers IL-protein systems in view of molecular-level mechanisms and interactions established between ILs and proteins, in parallel with those occurring in well-known protein stabilizers to come up with a rational perspective of the ILs’ promise. The last section of the current review provides the main conclusions on the suitability of ILs for different protein-based applications, with special emphasis on protein packaging and purification. Future prospects of ILs are finally provided, including the packaging of proteins, as well as in light-harvesting systems, protein/antibody purification, in vitro biomimicking of biological fluids (molecular crowding), and 3-D cell culture, among others.
2. Rational Understanding behind Protein Stability
In biological fluids, the native conformation of proteins is stabilized by minerals, sugars, lipids, amino acids, and other biomolecules.159 Consequently, some of these molecules have been used to stabilize proteins in vitro, employing various strategies and making use of several solvents. Below, we briefly review these strategies and solvents used, aiming to better understand and expose the molecular-level mechanisms responsible for protein stability.
2.1. Proteins in Pure Water
The role of water to drive the structure, dynamics, and biological functions of proteins is well-known.160−172 Proteins have a high affinity for water, where “about 0.3–0.7 g of bound water remains associated per gram of a dry protein in an aqueous solution.”160−162 Upon solubilization in water, proteins adopt a compact structure, driven by H-bonding with the protein-charged sites and hydrophobic solvation of the nonpolar core, where 83% of the protein’s nonpolar side chains and 82% of the peptide groups are buried.165 Compared to bulk water, the hydration shell of a protein has been evidenced to be 10–25% denser due to the clustering of water molecules, as determined by translational diffusion coefficients.160,164 The hydration process or the cover of the polar charged groups at the protein surface by water assists proteins to attain a high biological activity, sometimes described as a “lubricating effect” on proteins.167 The hydration layer of proteins, also called “biological water”,165 is composed of bound and free water, which remain in dynamic equilibrium. Free water molecules diffuse into the hydration layer from the bulk, and this represents a “feedback” mechanism of the hydration layer, wherein the bound and free water undergo reversible transitions..165 A scheme showing the interactions between water and proteins and the “feedback” mechanism of hydration layer is depicted in Figure 1.
Figure 1.

Water molecules in the hydration layer of proteins, showing the “feedback mechanism” of dynamical exchange between free and bound water along with continuous diffusion from the bulk water. Adapted with permission from ref (165). Copyright 2004 American Chemical Society.
Excellent reviews since the 1950s have highlighted the developments in the understanding of the solvation dynamics of water, in both the hydration shell and bulk, along with the water role in protein stability and activity and protein–protein interactions.160,163,165,167−170 Water–protein interactions have been discussed taking into account five main issues: (i) bound water molecules inside a protein, which have shown to provide flexibility to proteins to interact with different strands during folding or collapse;171 (ii) presence of water molecules in protein reactions, such as proton transfer in Bacteriorhodopsin (BR) membrane proteins to convert light into chemical energy; (iii) water at the protein surface (biological water), which assists in the stabilization of proteins and acts as an antifreeze agent to sustain life at low temperatures; (iv) solvent role in the protein glass transition, disclosing water–protein interactions and the dynamic transition of free and bulk water; and (v) motions in proteins induced by the translational diffusion of water.172
The long-term aqueous stability of proteins at room temperature against aggregation-induced denaturation is still a tricky issue, considering the fact that the native state of proteins is just 2–10 kcal·mol–1 more stable than the denatured form.168 The major force responsible for protein destabilization is conformational entropy.173 Being colloidal solutes, proteins undergo Brownian motion, thus allowing the occurrence of protein–protein interactions that may lead to aggregation and induced denaturation via unfolding pathways. The protein’s stability in water is controlled by five major forces: covalent peptide bonds, covalent disulfide bonds, electrostatic forces, dispersive-type interactions, and H-bonding. However, the total energy contributions of these forces to protein stability are easily overcome by denaturing processes in water, such as deamination, cysteine decomposition, and microbial growth causing denaturation by proteolysis, followed by aggregation. Still, the role of surrounding water in protein denaturation via misfolding and dimerization has also been reported.174 These findings reveal the dual nature of biological water in protein functioning and protein misfolding. Although water is the largest fraction of the protein’s biological environment, some proteins, and especially enzymes, have shown higher activity in organic solvents with low water content, with additional advantages, such as higher thermostability and avoidance of denaturation via microbial growth.174 Additionally, the stabilization of proteins in water can be further enhanced by the addition of additives (organic solvents, salts, osmolytes, and surfactants) or by their conjugation with polymers, as discussed below.
2.2. Proteins in Organic Solvents
Solubility limitations of some important fine chemicals and polymers, the need to reduce unwanted side reactions, and difficulties in product recovery are some of the major reasons behind the need for using organic solvents to solubilize proteins/enzymes.175 Organic solvents may additionally offer significant advantages over water, like pH control, protein imprinting, control of substrate specificity, regiospecificity, and enantioselectivity.175,176 Upon dispersion in organic solvents, proteins attain conformational rigidity, thus restricting their free mobility. But this does not mean that proteins do not require any water to display enhanced functions in organic solvents. As discussed in section 2.1, proteins attain full biological activity upon the formation of the first hydration layer. Accordingly, an additional question arises on how many water molecules are required. The biological activity of lysozyme is detectable after its hydration with 174 water molecules,177 corresponding to the water content required for the protein to acquire mobility, whereas chymotrypsin and subtilisin are active in organic solvents after hydration by 40–60 water molecules,178 which corresponds to the hydration of only the protein’s surface polar groups. Also, when lyophilized with specific substrates in water, proteins develop a special affinity for these molecules, even when later dissolved in organic solvents, called “molecular imprinting”.179,180 From these results, it is clear that in the absence of water, the protein’s charged groups get locked up with each other leading to an inactive conformation and that water is still required in the presence of organic solvents to maintain the protein stability and improved activity.
Serdakowski et al.181 reviewed the published works on enzyme activation in organic solvents in the purview of excipient (sugar and salts) addition during lyophilization. These excipients help in holding the water required to retain enzyme activity after lyophilization processes. They concluded that among sugars, disaccharides, like trehalose and sorbitol, are the best lyoprotectants. However, salts outperformed sugars. For instance, a 3,700-fold increase in the transesterification activity of subtilisin was reported in hexane when the enzyme lyophilization was carried out in the presence of KCl.182 Even more remarkably, the penicillin lyophilization in a 1:1 mixture of CsCl and KAc (or CH3COOK) leads to a 35,000-fold increase in the enzyme activity in hexane.183 The reasons given for such rises in activity were based on the formation of CsAc in solution, comprising a “kosmotropic” anion and a “chaotropic” cation.
Doukyu et al.184 reviewed the organic solvent tolerant enzymes and concluded that most of these enzymes are lipases, esterases, and proteases and that their solvent stability depends on the presence of disulfide bonds at their surface and secondary structure. Recently, Stepankova et al.185 reviewed the various strategies to stabilize enzymes in organic solvents, which are based on three main methods: (1) isolation of novel enzymes able to function under extreme conditions; (ii) modification of enzyme structures to increase their resistance toward nonconventional solvent media; and (iii) modification of the solvent environment to decrease its denaturing effect on enzymes. Though proteins/enzyme’s stability and activity in organic solvents exhibit certain advantages, this type of application is still limited to a few enzymes, and improvements are still required, either via protein engineering or by the finding of more adequate solvents.
2.3. Proteins in Aqueous Solution of Salts
It is imperative to understand the behavior of proteins in aqueous solutions of salts owing to their phylogenesis of modulating protein–protein interactions in vivo for solubility, aggregation, and function.186 However, due to the sheer complexity of salt ion-protein interactions, the related mechanism is still poorly understood.186−188 This is partly due to the labyrinthine nano anisotropic colloidal structure of proteins with an inhomogeneous surface charge density and differential polarity.186,189 Furthermore, protein–protein interactions involve electrostatic, hydrophobic, ion-dipole, and H-bonding.190 Salt ions can modulate these interactions (specifically the electrostatic interactions) subject to their concentrations, hydration, charge, and polarizability.191,192 The first important contribution to understanding protein behavior in an aqueous solution of salts was made by Franz Hofmeister.87 In 1887, while investigating regularities in the protein precipitating effects of salts and the relation of these effects with the physiological behavior of salts, Hofmeister found that the minimum normal concentration of salts required to precipitate egg globulin from an aqueous solution of egg white proteome follows an ion-specific order, known as the Hofmeister series (HS).87 The original HS, published in his 1887 seminal paper, is shown in Figure 2.
Figure 2.

Depiction of the original Hofmeister series published in 1887 by Franz Hofmeister.
In his next paper,88 Hofmeister explained this effect based on the water-absorbing ability of salts and concluded that more water-absorbing ions, like SO42– and Li+, induced fast precipitation compared to less water-absorbing ions, like NH4+ and ClO3–. This explanation was later framed into the 1930–1950s theory of water structure making (kosmotropes) and breaking (chaotropes) ions.89,90 The theory states that strongly hydrating kosmotropes can induce long-range water ordering beyond their first solvation shell, whereas weakly hydrating chaotropes lack this ability.89−91 Consequently, kosmotropes can easily withdraw water from the hydration shell of proteins to induce salting-out by promoting van der Waal’s and hydrophobic association of proteins at low concentrations, whereas chaotropes do not. Over the years, further experiments with a number of proteins and salts rendered improvements in the HS and extended the Hofmeister effect to protein stabilization and denaturing based on the same explanation of the kosmotropic or chaotropic nature of ions.91−95 Consequently, with few exceptions like lysozyme92 and γD-crystallins193 below their isoelectric point wherein a reversal of the order of ions was observed, the present-day Hofmeister series for protein stabilization and denaturation is summarized in Figure 3.91−95
Figure 3.

Modern Hofmeister series of ions for protein stability/denaturation.
Despite the veracity in the ion trend of the Hofmeister series, it should be remarked that recent spectroscopic and molecular dynamics simulation observations have demonstrated specific ion effects on the protein hydration shell, change in dielectric constant at the protein–water interface for adsorption of polarizable ions at the hydrophobic sites, ion–ion interactions at the protein surface, etc.96−101,194,195 The validity of these concepts is not in question in the current work, and as such, “kosmotropic” ions will be defined as those of high charge density and with strong salting-out ability, while “chaotropic” ions are those of low charge density displaying a salting-in nature.102
Due to their nanometer size and scattering nature, proteins in an aqueous solution are considered colloidal particles.196 Hence, apart from the Hofmeister effect, other theories (Poisson–Boltzmann;197 Debye–Huckel;198,199 and Derjaguin and Landau, Verwey and Overbeek (DLVO)200,201) have been used to explain the stability of proteins as colloidal particles in salt solutions. According to the Poisson–Boltzmann and Debye–Hückel theories, the colloidal stability of proteins is maintained by screening their surface charge due to the formation of electrical double layers of salt ions in the salt solutions causing electrostatic repulsion.197 Hence, electrostatic interactions dominate the van der Waals interactions. The DLVO theory, on the other hand, considers both electrostatic and van der Waals interactions between the charged protein particles for their colloidal stability in salt solutions.200,201
Currently, it is highly difficult to explain the colloidal stability of a given protein based on a simple hard sphere model since specific counterion-protein, and co-ion-protein interactions at the protein surface dominate their colloidal stability. Several small-angle X-ray (SAXS) and neutron scattering (SANS) studies with proteins (ovalbumin, α-crystallins, bovine serum albumin, and Apoferrtin) above their isoelectric point have shown that the effectiveness of counterions in screening electrostatic repulsive interactions between proteins depends on the co-ion and molecular weight of the protein when NaCl, YCl3, and CH3COONa are used as salts.202−204 Similarly, contrasting interactions of salts with proteins are reported at much below or slightly below their isoelectric points.205,206 A decrease in solubility due to enhanced protein–protein interactions of lysozyme was reported between pH 3.0 and 9.0, whereas an increase in solubility was reported at pH > 9.0 up to 1.2 M NaCl.205 Furthermore, at pH 9.4, the lysozyme showed inverse Hofmeister effect for the anions in liquid–liquid phase transition at low salt concentration, whereas the opposite effect was observed at high salt concentrations.84 Another important aspect to be considered here is the colloidal stability of proteins in buffer solution since they have been extensively used to stabilize proteins at a specific pH.207 Although not much is investigated about the mechanism of protein stabilization in buffers, the most accepted mechanism is the binding of buffer ions as ligands at the oppositely charged amino acids on the protein surface.207,208 In the case when the salt is added to the buffer solution of proteins, the salt ions compete with the buffer ions to adsorb at the specific sites.208 Accordingly, the mechanism of salt ion-protein interactions is very specific and strictly depends on a variety of physicochemical conditions, like solution pH, temperature, properties of ions, salt concentrations, protein structure, molecular weight, and surface charge density.
2.4. Proteins in Osmolytes Aqueous Solutions
Osmolytes are mainly organic solutes generally considered to protect the native conformation of proteins from external stress stimuli, such as variations in temperature, salinity, and pH.209−211 However, they can induce both stabilizing and denaturing effects, depending on their preferential interactions with proteins. Among the major osmolytes present in cellular fluids of eukaryotes, polyols, and disaccharides (glycerol and sucrose), some free amino acids and their derivatives (taurine and P-alanine, octopine), methylamines (trimethylamine-N-oxide (TMAO), betaine), and sarcosine, have been classified as protein stabilizers. On the other hand, urea, arginine, and guanidinium have been classified as protein destabilizers.209−211
In 1982, Yancey et al.209 investigated the evolution of osmolytes in various organisms (from prokaryotes to eukaryotes) against water stress and their compatibility with proteins. One of the author’s major conclusions relays on the “genetic simplicity in proteins”. The genetic simplicity and presence of compatible solutes allow proteins to function in the presence of various solutes and at their high/low concentrations, without any modification in the structure/confirmation of a number of proteins and assisting in the proper functioning of cells.209 The mechanisms of interaction of osmolytes with proteins have been explained in line with the Hofmeister effects, even with counteracting systems, e.g., urea:methylamine in a 2:1 molar ratio, wherein methylamine overwhelms the destabilizing action of urea.210 A summary of osmolyte behavior toward proteins is provided in Figure 4.
Figure 4.

Molecular structures of well-known osmolytes based on their stabilizing and destabilizing effect toward proteins.
Various other interaction mechanisms between osmolytes and proteins have been proposed over the years. However, if we consider it from a solution thermodynamic perspective, stabilizing osmolytes decreases the free energy of the protein’s unfolded state, favoring the folded population, whereas denaturing osmolytes lowers the free energy of the unfolded state, favoring the unfolded population of proteins. Takano et al.212 performed a molecular dynamics (MD) simulation on the effect of sarcosine on native RNase Sa, denatured RNase Sa, and four related proteins. The results gathered are in agreement with the osmophobic theory originally proposed by Bolen et al.,213 where “the osmolyte effect on protein stability is due to a solvophobic thermodynamic force which arises due to an unfavorable interaction of osmolytes mainly with the protein backbone.” These interactions can be determined from the transfer Gibbs energies of residue-specific contributions of proteins at the amino-acid level, from water to an osmolyte, based on solubility measurements.212,213 The stabilizing osmolytes get excluded from the vicinity of proteins in solution on account of unfavorable interactions, thus stabilizing it due to a “preferential hydration or preferential exclusion of solute” concept. This effect can be appraised by the preferential interaction parameter from equilibrium dialysis, which is positive for destabilizing osmolytes and negative for stabilizing osmolytes.214 This preferential exclusion concept is however supported by solvophobicity/solvophilicity, excluded volume, and surface tension (γ) effects. Solutes that increase the surface tension of the solution stabilize proteins, and vice versa, due to the preferential exclusion of solutes from the protein’s surface. Nevertheless, an opposing effect is observed with urea and TMAO, which respectively increase and decrease the surface tension of the solution and denature and stabilize proteins.210 Again, proteins are extremely complex molecules, and still there are many challenges to overcome aiming at finding a general theory capable of describing the reasons behind the protein’s stability/nonstability in solvents.
Street et al.215 stated that the thermodynamics concept is only descriptive and that there is no universal theory to explain the mechanisms by which osmolytes interact with proteins and accordingly affect their stability. The authors showed that there is no significant difference in the binding constant of stabilizing and denaturing osmolytes, thus attenuating the concept of preferential hydration or exclusion given above. A new model of osmolyte–protein–water interactions was proposed based on the transfer free energies of the protein’s backbone and osmolyte solution. This model was proposed based on the determination of transfer free energies (Δgtr) of backbone models from water to 1 M osmolyte solutions. Although side chains do play a role, it is primarily the backbone Δgtr that determines the extent to which osmolytes either stabilize (i.e., Δgtr > 0) or destabilize (i.e., Δgtr < 0) the protein. The validity of this concept was proved by experimentally determined and calculated Δgtr values.215 This model was further validated by Auton et al.,216 who proposed the additive contribution of both backbone and side chains to calculate Δgtr for 46 proteins and 9 different osmolytes, stating that the contribution of the side chain always opposes the backbone contribution.
In the latest development using Kirkwood–Buff integrals for protein–water interactions (G12) and protein-osmolyte interactions (G23), using sucrose, trehalose, sorbitol, and poly(ethylene glycol) as osmolytes and the antibody antistreptavidin immunoglobulin gamma-1 (AS-IgG1) as the protein, Barnett et al.217 concluded that the protective or denaturing action of osmolytes is concentration-dependent.217 It was also shown that the stabilization or destabilization tendency of osmolytes depends on the type of protein.217 Additionally, osmolytes such as betaine, citrulline, and proline have been reported to inhibit insulin fibrillation, operating via preferential exclusion of the osmolyte and polar interactions with the protein surface.218 The thermal stabilization of lysozyme by Glycine (GLY), N-methylglycine (NMG), N,N-dimethylglycine (DMG), N,N,N-trimethylglycine (TMG), and trimethyl-N-oxide (TMAO) has also been reported. Stabilization effects were explained due to favorable interactions between the amino protons of osmolytes with water, acting as proton donors in protein–water interactions.219 The stabilization propensity decreased with an increase in the alkyl chains in the amino groups,219 which attenuates the concept of the TMAO stabilization by preferential exclusion by steric reasons. In this regard, recent studies217,219 have presented reservations against the existing concepts of “preferential hydration or exclusion”, thus seeking new explanations based on novel mechanisms. Even though the molecular-level mechanisms behind the protein’s improved stability are not completely understood, the use of ILs as osmolytes, mainly due to their tunable features, may represent a promising option to impart better thermal and long-term stabilization of proteins, as will be discussed later.
2.5. Protein Formulations with Surfactants
Surfactants are a class of organic amphiphilic molecules that reduce the surface tension of molecular solvents or ILs upon adsorption at the air–liquid interface.26,220,221 Due to their amphiphilic nature, they can self-assemble into well-organized structures in the nanometer or micrometer scales, such as micelles, vesicles, and lamellar phases in solution.222 Depending upon having charge or not, and their position, surfactants can be classified into ionic, nonionic, zwitterionic, and gemini. Their relevance in protein formulation can be appraised from their applications in pharmaceuticals, laundry, cosmetics, paints, coatings, and biochemical reactions.143,144,223−229 If we go back into history, the sodium dodecyl sulfate (SDS)-induced denaturation of methemoglobin, evidenced by the protein color change, was the first report on surfactant-protein systems.230 It was followed by a series of studies on protein–surfactant chemistry, comprising features such as solubilization, dissociation, denaturing effects, and interactions occurring between proteins and surfactants, as summarized by Putnam231 and Steinhardt et al.232 Otzen224 reviewed the developments in surfactant-protein chemistry, delineating the existing mechanisms of interactions inducing the protein unfolding/refolding/misfolding.
From the protein stability perspective, ionic surfactants (cationic, anionic, and zwitterionic) have been reported to be 1,000-fold stronger denaturants, when compared to common denaturants like urea and guanidinium chloride. Due to their differences in chemical structures, surfactants interact with proteins via different mechanisms. Given that surfactants in solution may exist as monomers, aggregates, shared aggregates, and vesicles/micelles, their interaction mechanisms with proteins follow multiple equilibrium steps, which have been classified into four regimes: monomeric (0 → C1), aggregation (C1 → C2), shared aggregation (C2 → C3), and post aggregation (>C3).224,225 C1 is the critical aggregation concentration (CAC), which is a signature of the completion of the monomers (surfactants as individual molecules/ion) binding on proteins by electrostatic or hydrophobic interactions. It should be highlighted that CAC in the absence of protein, or any other polyelectrolyte, is equivalent to the CMC or CVC. Between C1 → C2, clusters of surfactants begin to form on proteins via cooperative binding due to hydrophobic interactions, thus causing the protein to unfold. These complexes are generally stabilized by interactions between different protein molecules forming shared micellar complexes between C2 → C3. The rate constant of unfolding in this regime has a linear dependence on the surfactant concentration. However, just below C3, proteins form individual complexes with surfactants, which can be quantified as the number of surfactants per protein. After CMC or CVC, surfactant aggregates individually form micelles/vesicles in solution, which is also composed of denatured proteins.233−236 Otzen224 reviewed the technical difficulties in characterizing surfactant-protein complexes and highlighted the problems above CMC/CVC, which could possibly be overcome using small-angle neutron scattering (SANS).237,238
Case studies with bovine serum albumin (BSA) and human serum albumin (HSA) allowed establishing the order of induced denaturation by ionic surfactants at low concentration, according to the rank gemini > cationic > anionic > zwitterionic.231,239,240 On the other hand, nonionic surfactants, with few exceptions, have been reported as stabilizers of proteins in all concentrations.231 Although these are overall trends, there are exceptions depending on the protein and surfactant, as observed before with organic solvents and osmolytes. For instance, the anionic surfactant SDS, the cationic surfactant tetradecyltrimethylammonium bromide (TTAB), and the uncharged surfactants octyl maltoside and octyl glucoside have been reported to activate the enzymes Thermomyces lanuginosus and cutinase, particularly at low concentrations and below their CMC.241,242 Still, it should be kept in mind that it is not feasible to compare denaturant/renaturant effects of surfactants over enzymes and nonenzymatic globular proteins, mainly because the structural alterations occurring in proteins are different from functional alterations.224 Moreover, the observed opposite effects of SDS and cetyltrimethylammonium chloride (CTAC) on the stability of BSA and HSA further vindicate the specific nature of surfactant-protein interactions.240
Apart from the surfactant charge, surfactant-protein interactions also depend on the solution pH, temperature, and stereochemistry.243,244 The stereochemical dependence of interactions opens the door to the design of surfactants for the selective extraction and stabilization of proteins. From the pharmaceutical point of view, many protein-based drugs need to be stored and shipped as solution formulations. The transportation causes severe agitation of the protein solutions, resulting in their interaction with container surfaces which can damage proteins due to the interfacial stress. Surfactants have been found to be the most suitable additives to avoid interfacial instability since they compete with proteins for the interfaces, upon migrating to the interfaces and protecting proteins during shaking or stirring.245 However, not all surfactants can stabilize proteins against interfacial damage since factors such as surfactant charge can act as a denaturant via ionic interactions with the protein surface. The generalized action of various types of surfactants toward proteins is shown in Figure 5.
Figure 5.
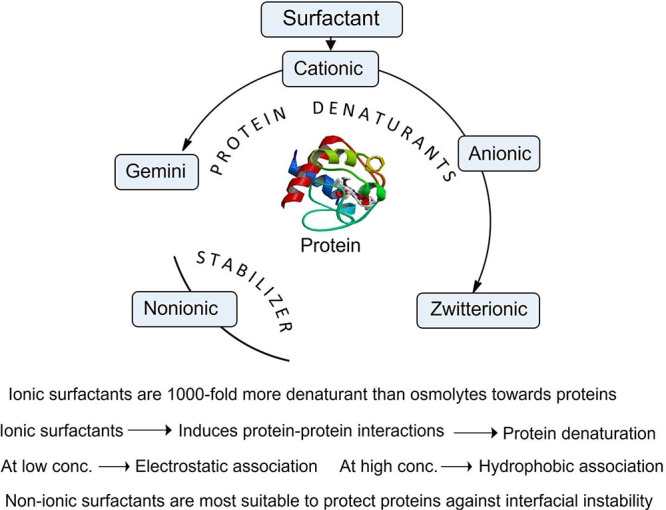
Schematic overview of the denaturation and stabilizing action of various types of surfactants toward proteins.
Irreversible aggregation of proteins is caused by disulfide shuffling or stable hydrophobic association. In the case of therapeutic proteins (TPs), this effect has a direct impact on drug efficacy and immunogenicity.246 The most common TPs stabilized by surfactants are monoclonal antibodies, Interleukin 2, cytokines, Human chorionic gonadotropin HGH, and fusion proteins.247−253 Most of the surfactants used to stabilize TPs are nonionic, while possessing long alkyl side chains, such as polysorbate 80 (PS 80), PS 60, PS 20, and Poloxamer 188, in the concentration range between 0.005 and 0.16%.247−253 In the bulk phase, surfactant-protein complexes prevent protein–protein interactions, which is one the major causes of their denaturation.
In 2011, Lee et al.254 reviewed the molecular origin of the surfactant-based protein stabilization potential and concluded that surfactants stabilize proteins by their preferential location at the interface and/or their association with proteins in solution. On the other hand, Khan et al.255 reviewed the key interactions occurring in formulations of surfactants and therapeutic proteins, raising reservations against the proposed mechanisms and warranting further work to develop a clearer picture of the phenomenon.255 More recently, ionic liquid surfactants (SAILs) with higher surface activity compared to their conventional counterparts have been proposed,111−140 which are discussed below as an independent section.
2.6. Protein-Polymer Conjugates
Frequently administered TPs are afforded in high concentrations due to their longer half-lives in the bloodstream. Therefore, they are required to be stable for long periods at high concentrations, up to 10 mg/mL or 10,000 ppm, against aggregation-induced denaturation.256 One of the most prevalent strategies to curb this challenge is their conjugation with polymers, such as by the PEGylation approach.256,257 The first report in this direction was published by Abuchowski et al.,258 who PEGylated the protein BSA, leading to lower immunogenic response and higher circulation time in blood.259 This pioneering work led to the rise in frequency of works in protein-PEGylation, with some strategies already approved by the FDA.260 Currently, there are 23 PEGylated protein therapeutics clinically used to treat a wide range of diseases.261 The high promise of this strategy is due to the special characteristics of PEG, such as its nonfouling nature and resistance to opsonin binding, which is the major protein initiating the phagocytosis of foreign molecules in the bloodstream. Since opsonin exhibits a higher affinity for hydrophobic and charged species, being a hydrophilic and neutral polymer, PEG overturns the opsonin’s phagocytosis effect and thus increases the biocirculation time of conjugated proteins. Biocirculation time is also increased due to an increase in the molecular weight of PEG attached to the TPs, therefore reducing kidney clearance.262−264 In 2015, Pelegri-O’Day et al.265 reviewed the developments in TP-polymer conjugates in the purview of PEGylation and beyond. The authors stated that besides the described specific advantages, the strategy also has shortcomings, such as hypersensitivity, nonbiodegradability causing accumulation in tissues, accelerated blood clearance by anti-PEG antibodies, and recognition of PEG by the immune system and immunological responses.266−269 Alternative polymers evolved to overturn these limitations, namely N-(2-hydroxypropyl)methacrylamide (HPMA), poly(2-ethyl-2-oxazoline), poly(glutamic acid), poly(ethylene glycol) methyl ether methacrylate (pPEGMA), poly(carboxybetaine), polyglycerols, hydroxyethyl starch, polysialic acid, poly(N-hydroxypropyl)methacrylamide (pHPMA), polyglycans, and glycopolymers with pendant trehalose.265 Zhao et al.270 reviewed the TP-polymer conjugates in terms of methods of their synthesis. However, further improvements to increase the activity of the proteins by specific orthogonal biological function are needed to design polymers with precise sequence control and monodispersity.265,270 The recent rise of polymeric ILs as a conjugated polymer can further improve the protein/TP solubility/stability,153,271 which is described later in this review. A pictorial summary of different designs, syntheses, and architectures of protein–polymer conjugates proposed for bioapplications is shown in Figure 6.
Figure 6.
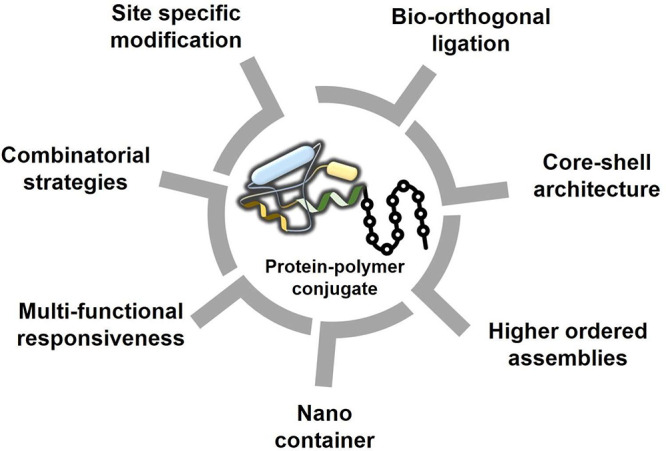
Overview of the design, synthesis, bioarchitectures, and bioactivities of protein–polymer conjugates for biobased applications.
3. Ionic Liquid-Protein Systems
The structural diversity of ILs, responsible for their tunable nature and properties, endowed them with the “luxury” to interact with proteins by different mechanisms. Some ILs have shown great promise as protein stabilizers and thus as potential protein packaging materials/solvents, which is a relevant application in the field of therapeutic proteins. Therefore, this section will cover the works published to date, divided according to the IL use, i.e., as native solvents, co-solvents, adjuvants, surfactants, IL-based aqueous biphasic systems (IL-ABS), and Poly(IL)-protein conjugates. The detailed mechanisms of interaction between ILs and proteins are discussed in parallel with those occurring in well-known protein stabilizers to come up with a rational perspective of the IL’s potential. Apart from protein packaging and relevance with TPs, other applications of IL-protein systems are addressed here.
3.1. Ionic Liquids as Native Solvents
Due to their ionic and hygroscopic nature, it is impossible to dry all ILs up to 100% as a ppm level of water always remains with ILs. Therefore, most of the marketed ILs are always tagged with a given percentage of water. However, despite the presence of water at ppm levels, generally considered enough to form hydration layers around proteins, only a scarce number of proteins are soluble in dry ILs or NILs,28−34 contrary to the larger number of organic solvents that act as good protein solvents. As discussed earlier, ILs comprise nanoscale heterogeneous domains, i.e., polar/nonpolar nanodomains,19−27 and as such must solvate proteins operating via different mechanisms. A summary of the proteins that have been solubilized in native ILs, along with their secondary structure and employed ILs, is shown in Table 1. The molecular structure of the most promising NILs for the stable solubilization of specific proteins is shown in Figure 7.
Table 1. Summary of Proteins Dissolved in Native ILs along with Their Secondary Structure.
| Protein | Secondary Structure | Ionic Liquid |
|---|---|---|
| CAL B28 | α/β | [C2mim][C2OSO3], [C4mim][Lac], [C2NH3][NO3], [C4mim][NO3], [(C2)3C1N][C1OSO3] |
| CAL B272 | α/β | [(C2)3C1N][C1OSO3] and [C4mim][C2N3] |
| CAL B274 | α/β | [HOPmim][NO3] |
| CAL B29 | α/β | [Me(OEt)2-Et-Im][OAc], [Me(OEt)3-Et-Im][OAc], [Me(Opr)3-Et-Im][OAc], [Me(Oet)3-Et3N][OAc], [C4mim][dca], [C2mim][OAc] [C8mim[OAc], [C4mim]HCOO], [Me(Oet)3-Et-Im][C2N3], [Me(Oet)3-Et3N][HCOO], [Me(Oet)2-Et3N][Oac], [Me(Oet)3-Bu-Im][Oac], [C2mim][C2OSO3], [(C4)4N][HCOO], [Amm110]Cl, [Me(Oet)7-Et-Im][OAc], [Me(Oet)3-MeOEtOMe-Im][OAc], [Me(Oet)3-Me-Et-Im][OAc] |
| CAL B275 | α/β | [C2OHmim][HOC1SO3], [C2OHmim][HOC2SO3], [C3OHTEA][HOC2SO3] |
| Cellulase30 | α/β barrel | [C1mim][Cl],[(OHC2H5)3C1N][C1OSO3] |
| Cytochrome c276 | All-α | [C2OHmim][Tf2N], [C3OHmim][Tf2N], [C2OC1mim][Tf2N], [C6OHmim][Tf2N], [C8OHmim][Tf2N] |
| Cytochrome c31 | All-α | [C2mim][C2OSO3] |
| Cytochrome c32 | All-α | [C4mim]Cl, [Amim]Cl |
| Silk Fibroin277 | Cross-β | [C4mim]Cl, [C4mim]Br, [C4mim]I, [C4(C1)2im]Cl, [C2mim]Cl |
| Keratin33 | α+β | [DMEA][HCOO] |
| Keratin278 | α+β | [C4mim][OAc], [Cho][TGA], [Cho][Pn], [C4mim]Cl, [TMG][Pn] |
| Zein279 | All-α | [C4mim] [C2N3], [C4mim]Cl |
| Zein34 | All-α | [C4mim][OAc], [C2mim][OAc], [C2mim][dca], [C4mim]Cl, [C1mim][OAc], [C1mim][HSO4], [C1mim][HCOO] |
| HSA280 | All-α | [C4mim][OAc], [C4mim][SCN] |
| Ovalbumin280 | α+β | [C6,6,6,14P]Cl [C2,4,4,4P][(C2)2OPO3], [C4mim][OAc], [C4mim][SCN], [C2mim][C2OSO3] |
| Myoglobin280 | All-α | [C2,4,4,4P][(C2)2OPO3], [C4mim][OAc], [C4mim][SCN] |
| α-Chymotrypsin280 | All-β | [C2,4,4,4P][(C2)2OPO3], [C4mim][OAc], [C4mim][SCN], [C2mim][C2OSO3] |
| Lysozyme280 | α+β | [C1,4Pyr][C2N3], [C4mim][OAc], [C4mim][SCN], [C2mim][C2OSO3] |
| Lactoferrin280 | α+β | [C4mim][OAc], [C4mim][SCN], [C2mim][C2OSO3] |
| Gelatin281 | Random coil | [CnNH3][NO3] (n = 2, 3, 4), [CnNH3][NO3] + [C4mim]Cl |
| Insulin282 | α+β | [Cho][gerenate] |
Figure 7.

Molecular structures of the most promising neat ionic liquids for both solubilization and stabilization of specific proteins.29,31,32,275,282
Different studies on NIL-protein systems have emphasized the role of covalent and noncovalent interactions both in protein solubility and stabilization. Lau et al.,28 in 2004, were the first to show the dissolution of the enzyme Candida antarctica lipase B in NILs, viz. [C2mim][C2OSO3], [C4mim][Lac], [C2NH3][NO3], [C4mim][NO3], and [(C2)3C1N][C1OSO3] at 40 °C. However, only [(C2)3C1N][C1OSO3] stabilized the enzyme, indicated by the retention of the secondary structure and trans-esterification activity, which is due to the H-bonding between the IL ions and the protein surface. The authors concluded the need for a balance between steric factors and hydrogen bond accepting/donating properties of ILs for protein solubilization and stabilization.28 In 2006, Sheldon and co-workers272 studied the comparative stability and activity of Candida antarctica lipase B and its cross-linked enzyme aggregate (CLEA) in [(C2)3C1N][C1OSO3] and [C4mim][C2N3].272 They found high stability of the enzyme in [(C2)3C1N][C1OSO3] compared to that in [C4mim][C2N3] due to the strong hydrogen bonding nature of the first IL. However, higher activity of CLEA was observed in [C4mim][C2N3], implying the role of protein engineering to increase enzyme activity in ILs.272
The relevance of H-bonding for protein stability in NILs was also demonstrated from simulation studies on the solvation of small cyclic hexapeptide in ILs, namely, [C4mim]Cl and [C4mim][PF6].273 The H-bonding between the hydroxyl groups of the peptide and the IL anion was shown to stabilize the peptide in [C4mim]Cl compared to that in [C4mim][PF6].273 Bermejo et al.274 also reported higher solubility of Candida antarctica lipase (12%) in [HOPmim][NO3] containing a cation with an H-bonding site. Zhao et al.29 investigated the dissolution and stabilization of Candida antarctica lipase B in 18, nonfunctionalized and ether-functionalized ammonium and imidazolium cation containing ILs, with [OAc], [HCOO], [dca], and [C2OSO3] anions. The order of solubility found was as follows: [Me(OEt)2-Et-Im][OAc] ≈ [Me(OEt)3-Et-Im][OAc] ≈ [Me(OPr)3-Et-Im][OAc] ≈ [Me(OEt)3-Et3N][OAc] > [C4mim][C2N3] ≈ [C2mim][OAc] ≈ [C8mim[OAc] ≈ [C4mim]HCOO] ≈ [Me(OEt)3-Et-Im][dca] > [Me(OEt)3-Et3N][HCOO] ≈ [Me(OEt)2-Et3N][OAc] ≈ [Me(OEt)3-Bu-Im][OAc] > [C2mim][C2OSO3] ≈ [(C4)4N][HCOO] > [Amm110]Cl ≈ [Me(OEt)7-Et-Im][OAc] ≈[Me(OEt)3-MeOEtOMe-Im][OAc] ≈ [Me(OEt)3-Me-Et-Im][OAc]. The ether-functionalized ILs, [Me(OEt)3-Et3N][OAc] and [Me(OEt)3-Et-Im][OAc], were found to be the most effective in preserving the secondary structure and activity of the enzyme.29 Ou et al.275 showed that the ionizing-dissociating abilities of ILs having hydroxyl functionalities paralleled the catalytic activity trend of lipases dissolved in these ILs. The studied ILs—[C2OHmim][HOC1SO3], [C2OHmim][HOC2SO3] and [C3OHTEA][HOC2SO3]—provide a nondenaturing and noninhibitory environment to the enzyme due to their ionizing-dissociating abilities. This evidence is critical for the development of ILs for the stabilization of not only TPs but as well to other relevant proteins with a moderate hydrophobic nature.
Besides Lipases, few other enzymes, like Cellulase and Cytochrome c, have been investigated in NILs. Bose et al.30 investigated the reactivity and stability of commercial Cellulases from Trichoderma reesei in eight ILs, reporting their structural and functional stability in [C1mim]Cl and [(OHC2H5)3C1N][C1OSO3]. However, HEMA imparted much higher thermal stability to Cellulase, demonstrated by its high melting temperature (Tm = 115–125 °C) compared to the Tm = 70–94 °C in [C1mim]Cl. Using an indirect strategy by complexation with dicyclohexano-18-crown-6, Shimojo et al.276 solubilized Cytochrome c in hydroxyl and oxy functional imidazolium-based ILs, paired with the bistriflamide anion. The enzyme solubility according to the IL cation follows the order [C2OHmim] > [C3OHmim] > [C2OC1mim] > [C6OHmim] > [C8OHmim]. The dissolution step, however, changed Cytochrome c from an electron transfer to a peroxidase enzyme due to the replacement of axial Met80 from the sixth coordination position by amino acid from the peptide chain.276
Bihari et al.31 reported the dissolution of Cytochrome c in [C2mim][C2OSO3], achieved by the complexation of the free heme coordination site imidazolium cation. The [C2mim][C2OSO3] preserved the secondary structure of the enzyme with an enhancement in its peroxidase activity.31 Tamura et al.32 explained the solubility of Cytochrome c in [C4mim]Cl and [Amim]Cl at 80 °C based on the Kamlet–Taft parameters (β and π*) of ILs. They stated that ILs with β and π* higher than 0.70 and 1.17, respectively, can dissolve Cytochrome c. Indeed, if we look back into the earlier report of Cytochrome c solubility in [C2mim][C2OSO3],31 we find that [C2mim][C2OSO3] has a β value of 0.71, thus supporting the Tamura et al.32 views. Hence, the role of H-bonding in solubilization and stability of enzymes is one common denominator for all the ILs discussed.
Other than enzymes, many structural and functional proteins have been dissolved in native ILs via different dissolution mechanisms. Phillips et al.277 reported the dissolution of various percentages of protein silk fibroin in [C4mim]Cl (13.2%), [C4mim]Br (0.7%), [C4mim]I (0.2%), [C4(C1)2im]Cl (8.3%), and [C2mim]Cl (23.3%), in which the dissolution is achieved by the disruption of hydrogen bonds in the crystalline domains of the cross-β structure.277 Chen et al.278 reported the dissolution of natural protein fibers such as wool, human hair, and silk in ILs, viz. [C4mim][OAc], [Choline][TGA], [Choline][Pn], [C4mim]Cl, and [TMG][Pn], occurring due to cuticle swelling or surface interactions.278 Idris et al.33 showed the dissolution of keratin in [DMEA][HCOO], yet without reporting any dissolution mechanism.
Biswas et al.279 reported the dissolution of zein in [C4mim][C2N3] and [C4mim]Cl and used the solution for successful acylation of the native protein. Tomlinson et al.34 further explored the solubility of zein in imidazolium-based ILs comprising the anions [OAc], Cl, [dca], [HCOO], and [HSO4]. They found the highest solubility in [C1mim][OAc] and [C2mim][C2N3] and explained the improved solubility based on the molar volume of ILs.
Strassburg et al.280 looked beyond the largely studied imidazolium-based ILs and reported the solubilization of BSA, HSA, Ovalbumin, Myoglobin, α-Chymotrypsin, Lysozyme, Cytochrome c, and Lactoferrin in quaternary phosphonium-, quaternary ammonium-, and pyrrolidinium-based ILs, alongside imidazolium-based counterparts for comparison purposes. Among these, [C6,6,6,14P]Cl dissolved ovalbumin; [C2,4,4,4P][(C2)2OPO3] dissolved BSA, Ovalbumin, Myoglobin, α-Chymotrypsin, and Cytochrome c; [C1,8,8,8N]Cl dissolved BSA; and [C1,4Pyr][dca] dissolved Lysozyme. However, imidazolium-based ILs still proved to be the first-choice cation for proteins, as [C4mim][OAc] and [C4mim][SCN] dissolved all the studied proteins. Nevertheless, further temperature-dependent stability analysis using DLS and FTIR revealed only small alterations in lysozyme structure upon heating at 80 °C in [C2,4,4,4P][C2OPO3] compared to that in [C2mim][C2OSO3].280
Mehta et al.281 reported high solubility (58 to 87%) of protein gelatin in neat alkyl ammonium nitrates ([CnNH3][NO3], n = 2, 3, 4) and their binary mixtures with [C4mim]Cl. The H-bonding of ILs with gelatin was suggested as the major driving force for protein solubility, as appraised by solubility experiments with an increasing [C4mim]Cl concentration and decreasing gelatin solubility in the [CnNH3][NO3] + [C4mim]Cl mixture.281
In a recent significant development, Banerjee et al.282 (Figure 8) reported the stable dissolution of Insulin (therapeutic protein) in choline geranate, [Cho][gerenate], as an oral insulin formulation. The 10 U/kg insulin-[Cho][gerenate] was orally delivered in enterically coated capsules using an oral gavage, resulting in a sustained decrease in blood glucose of up to 45%, thus establishing the relevant role of neat cytocompatible IL-protein formulations for pharmaceutical applications.282
Figure 8.

Illustration of the [Cho][gerenate]-insulin oral formulation.
3.1.1. Solubilization Mechanisms of Proteins in NILs
The above-discussed reports demonstrate that a relevant number of proteins, namely, CAL B, CRL, Cytochrome c, BSA, HSA, α-chymotrypsin, ovalbumin, myoglobin, lysozyme, lactoferrin, silk fibroin, Zein, keratin, and gelatin, can be solubilized in NILs. However, and with the exception of a few cases, solubilization in NILs resulted in alterations of the protein’s secondary and tertiary structure.28 Although all are connected, three different rationales have been used to describe the dominant molecular-level mechanisms responsible for the protein’s solubilization/stabilization: (1) H-bonding formation between the proteins and the ILs; (2) Kamlet–Taft parameters (α, β, and π*), which in turn show the hydrogen bond donor capacity (HBD) and hydrogen bond acceptor capacity (HBA) and dipolarity/polarizability of the IL;283 and (3) hydrogen bond disruption in proteins. Overall, from the works addressed, it can be seen that hydrogen-bonding of the IL ions with the protein surface has been the most accounted phenomenon observed with proteins bearing α/β or α+β secondary structural conformation. For proteins with an all-α conformation (Cytochrome c, BSA, HSA), higher β and π values of ILs have been accounted for as the reason for improved solubility, implying strong H-bonding (β value >0.7) between the IL ions and the protein. In the case of proteins with higher content of β-sheet structure, such as keratin and silk fibroin, disruption of intramolecular bonds of proteins by IL anions has been identified as the main reason for enhanced solubility. These views, proposed as ruling the solubilization of proteins in native ILs, are schematically presented in Figure 9. However, it is noted that H-bonding is dominant but not the sole force for the solubilization of proteins in NILs. Considering the broad polar heterogeneity of both proteins and ILs, protein dissolution/stabilization in native ILs involves secondary forces, such as Coulombic interactions, hydrophobic interactions of nonpolar groups, and van der Waal’s interactions. Experimental challenges for understanding of protein-neat ILs interaction arise due to the inapplicability of common techniques like in solution NMR and circular dichroism in near-pure ILs. Molecular dynamics (MD) simulations offer an alternative to better understand the interactions taking place.284 Shim et al.273 carried out MD studies on the solvation of cyclic hexa-peptides in the ILs [C4mim]Cl and [C4mim][PF6], revealing peptide structure distortion by electrostatic interactions occurring between the peptides and both ILs. The distortion was lower in the [C4mim]Cl due to the stabilization of the peptide by intermolecular H-bonding.273 Burney et al.285 simulated Candida rugosa lipase (CRL) in [C4mim][PF6] and [C4mim][NO3], concluding that ILs dampen protein dynamics, trapping the system near its initial structure due to electrostatic interactions.
Figure 9.

Dominant dissolution mechanism of proteins in a nano heterogeneous structure of native ILs. (a) Hydrogen bonding of protein with the anion of ILs, mainly observed for all-α proteins. (b) Hydrogen bonding of protein with both cation and anion of ILs, observed mainly in α/β or α+β proteins. (c) Disruption of the internal hydrogen bonds of proteins by the IL anion and steric interactions with the cation, mainly observed in all-β proteins. Besides the dominant H-bonding interactions, other interactions, like electrostatic and hydrophobic interactions, play a secondary role in improving the dissolution of proteins in ILs.
Klähn et al.286 studied the solvation and stability of CAL-B in various imidazolium and guanidinium cations using MD simulations. The authors concluded that the interaction of CAL-B with the IL anion is dominated by Coulombic interactions, whereas that with the cation is by van der Waal’s interactions. The authors also showed that smaller ions led to stronger electrostatic screening with the solvent and hence stronger interaction with the enzyme. On the other hand, ions with large size and more dispersed surface charge increase enzyme-IL interactions, leading to the destabilization of the enzyme with decreased solvation. Overall, MD simulations provide molecular-scale insights into IL-protein interactions, overcoming experimental limitations in studying protein stabilization in native ILs.
As stated in the beginning of this section, it must be noted that even in a dry state few water molecules coexist with ILs, which have been shown at the desired location around proteins. This phenomenon ultimately leads to the reference of H-bonding acting in a different direction. ILs exhibiting hydroxyl or oxy functionalities in both ions29,275,276 are improved solubilizers and stabilizers of proteins. Therefore, these ILs could represent the most efficient ones for the solubilization of proteins and TPs in NILs, while keeping their stability.
However, a special behavior has been observed with the ILs [C2mim][C2OSO3] and [C4mim][OAc], which have shown the ability to solubilize proteins of all kinds of secondary structures, namely, all-α (Cytochrome c, BSA, HSA, myoglobin), all-β (α-chymotrypsin, keratin), α/β (CAL B), and α+β (Ovalbumin, Lysozyme, and Lactoferrin). Because of their special behavior, we went into detail regarding their structures to uncover why these ILs are more suitable for protein solubilization, as discussed below. Comparing the β value (reflecting the hydrogen-bond basicity of the IL, and mainly dictated by the IL anion) they are on a suitable scale (>0.65)32,148 for dissolving biopolymers. Furthermore, [C4mim][OAc] has a higher hydrogen-bond basicity (β = 1.18)287 than [C2mim][C2OSO3] (β = 0.71),288 being thus a better solvent for proteins (Figure 10a).
Figure 10.

(a) Molecular structures of [C2mim][C2OSO3] and [C4mim][OAc], which solubilize all kinds of proteins, and [OHCn=2–8mim][NTf2] which dissolve Cytochrome c despite having β < 0.4. (b) depiction of the molecular mechanism of dual hydrogen bonding (DHB) by [C2mim][C2OSO3] and [C4mim][OAc] with folded proteins.
In the same line, imidazolium-based ILs having β > 0.65, such as those containing Cl and [C2N3] anions, are good candidates to dissolve proteins as well, as experimentally demonstrated.29,30,32,34,277,280 In contrast, few ILs such as [OHCn=2–8mim][NTf2] with β < 0.4 can also solubilize proteins.276 Therefore, β may not be the sole criterion for the special dissolving behavior observed by [C2mim][C2OSO3] and [C4mim][OAc], with electrostatic, hydrophobic, and van der Waal’s interactions also playing a role.
When addressing the electronic structure of the [C2mim][C2OSO3] ion pairs as a dry solvent, it has been reported to exist in three conformations, varying based on the H-bonding ratio between the ethylsulfate anion and hydrogens of the imidazolium cation.289 The lowest-energy conformer is involved in 6 hydrogen bonds between the cation and the anion, wherein the C2–H9, C5–H11, and C4–H10 bonds show bifurcated interactions. However, two other conformations have free acidic hydrogens at the C5 position. This hydrogen can particularly participate in H-bonding with carboxylate groups at the protein surface along with H-bonding between the anion and the amino groups, thus providing dual H-bonding sites like observed in [C2OHmim][HOC1SO3] stabilizing BSA.275 As far as the case of [C4mim][OAc] is concerned, a similar phenomenon could be depicted. Even though we could not find its electronic structure, [C2mim][OAc] also contains free acidic hydrogens at the C5 position289 and therefore could be contributing toward dual H-bonding and improved protein solubilization of different conformations. Figure 10b shows a schematic overview of the dual hydrogen-bonding phenomenon at the molecular level.
Overall, hydrogen bonding occurring between ILs and proteins providing a water-like environment at the surface of the protein, ideally in the form of dual hydrogen bonding as discussed above, seems to be the major governing force of protein solubility and stability in NILs. Further augmentation in this force could be sought by analyzing the effects of adding small amounts of water into NILs, in which ILs could act as co-solvents, as discussed in the following section.
3.2. Ionic Liquids as Co-solvents
Following the proteins that showed robustness when solubilized in NILs, a significant amount of work has followed with the addition of 2–50% water, resulting in A-ILCS. These IL-water mixtures, in which the IL could act as co-solvent, lead to given advantages, including (1) providing sufficient water to hydrate the protein and (2) imparting thermal stability to the proteins by IL ions present in the form of clusters in the hydration layer.24,27,54 The chemical structures and composition of the most promising water-IL systems for protein packaging and protection are provided in Figure 11.
Figure 11.

Most promising water-IL systems for protein packaging and protection.59,296,299,67,312,291,301
The mode of interaction, solvation, and stabilization of proteins by A-ILCS depends on the structure and polarity of the IL cation/anion and water content. For example, the earliest work in this direction was published by Baker et al.59 in 2004, who showed the stabilization of the protein monellin in A-ILCS with a hydrophobic IL, [C4mpy][Tf2N]:H2O (98:2 v/v) mixture, up to 105 °C compared to 40 °C in pure water. Using Trp fluorescence as an internal spectroscopic handle, the authors observed a blue shift in Trp fluorescence above 105 °C, which accounted for stripping off the water from the protein hydration layer by [C4mpy][Tf2N]. Moreover, calculated entropies of unfolding in [C4mpy][Tf2N]:water (136 J·K–1·mol–1) compared to water (250 J·K–1·mol–1) indicated more rigid solvation of the protein in the IL:water mixture.59 Diego et al.60 studied the structural stabilization of α-chymotrypsin in A-ILCS comprising [(C2)2mim][Tf2N]:1-propanol:H2O (85.5:12.5:2, v/v) using DSC, CD, and fluorescence techniques. From a thermodynamic point of view, the melting temperature of the protein increased by 10.4 °C, with an increase in the enthalpy of denaturation (dHcal) by 3-fold compared to water.
From a structural point of view, the [(C2)2mim][Tf2N] mixture stabilized the protein via the formation of a flexible and more compact 3D structure, while preserving the essential water shell. The all-β conformation of protein was preserved, with the rise in β-sheet from 33.4% to 47%, whereas the tertiary structural stabilization was reflected by a rise in Trp fluorescence indicating compaction of protein.60 Lozano et al.62 reported the structural and functional stability of enzymes, i.e., α-chymotrypsin in the [C2mim][NTf2]:1-propanol:H2O (85.5:12.5:2 (v/v/v) mixture. The mechanism of stabilization was explained based on the Dupont model of wet ILs.24,54 Enzymes reside in the hydrophilic gaps of the network, where the observed stabilization of enzymes could be attributed to the maintenance of this strong network around the protein. The extremely ordered supramolecular structure of ILs in the liquid phase was proposed to act as a “mold” in maintaining an active three-dimensional structure of enzymes in aqueous nanoenvironments, while avoiding the classical thermal unfolding.62 Therefore, A-ILCS of hydrophobic ILs form an IL solvent cage around proteins, allowing the stabilization of it against thermal denaturation (Figure 12).
Figure 12.

Illustration of the mechanism of protein stabilization by A-ILCS composed of hydrophobic ILs.
This structure stabilization model however changes significantly with the introduction of ILs with more interactive sites, polarity, or structure slightly similar to that of water. For instance, Falcioni et al.290 reported the stability of the conformation and activity of enzyme Subtilisin in A-ILCS of [(OHC2)2NH2]Cl:H2O (97.8:2.2, v/v mixtures), which was due to the presence of two hydroxyl groups in the cation that coordinate with the denaturing Cl anion, hence overturning its deleterious effect.290
Venkatesu and co-workers67,291−295 investigated the co-solvent effect of quaternary ammonium-based ILs on various proteins, with a special focus on α-chymotrypsin. In most of their reports, they showed the stabilization of proteins by the quaternary ammonium family of ILs, which was explained based on the mechanism of preferential exclusion of ILs ions from the protein surface. This is a phenomenon popular with the stability of proteins by osmolytes according to the osmophobic theory of Bolen213 and transfer Gibbs free energy concept (Figure 13).215,216
Figure 13.

Illustration of the mechanism of protein stabilization by the quaternary ammonium cation family of A-ILCS according to preferential exclusion from protein surface concept.
In their first work in the field, Attri et al.67 reported higher activity and stability of α-chymotrypsin in ILs: H2O (50:50, v/v) A-ILCS of [(C2H5)3NH][OAc] and [(C2H5)3NH][PO4] compared to [Bzmim]Cl, [Bzmim][BF4], and [(C4H9)4P][Br]. The phenomenon was explained based on unfavorable interactions of quaternary ammonium-based IL ions with the disulfide bonds and backbone of α-chymotrypsin.67 The [(C2H5)3NH][OAc]-based A-ILCS was also shown to refold α-chymotrypsin from the quenched thermally unfolded state. Further, Attri et al.291 reported that 3 molar of [(C2H5)3NH][OAc] in water attenuates the deleterious effect of urea on the denaturation of α-chymotrypsin due to the H-bond acceptor ability of acetate, strengthening the water-water and water-urea interactions and limiting the urea-α-chymotrypsin H-bonding interactions.291 To get deeper insights into the molecular mechanism of IL-protein interactions toward protein stability, the authors investigated the effect of A-ILCS of [Et2NH][OAc], [Et3NH][OAc], [Et2NH][dhp], [Et3NH][dhp], [Et2NH][HSO4], and [Et3NH][HSO4] on the stability of cyclic dipeptides, namely, cyclo(Gly-Gly), cyclo(Ala-Gly), cyclo(Ala-Ala), cyclo(Leu-Ala), and cyclo(Val-Val)292 based on the transfer free energy (Δgtr) concept.215,216 The positive values obtained revealed unfavorable interactions between ILs and cyclic dipeptides, leading to the stabilization of the native structure of cyclic dipeptides. The authors concluded that peptide bonds, the peptide backbone unit, the alanyl residue, and the valyl residue (containing amide) play a more relevant role in protein folding/unfolding compared to side chains of proteins.292 A similar mechanism was proposed for the stabilization of zwitterionic glycine peptides, namely, glycine (Gly), diglycine (Gly2), triglycine (Gly3), tetraglycine (Gly4), and cyclic glycylglycine (c(GG)), with a decreasing order of the m value, [(C2)2NH2][HSO4] > [(C2)3NH][OAc] > [(C2)3NH][HSO4] > [(C2)2NH2][OAc] > [(C1)3NH][OAc]>[(C1)3NH][dhp].293 Attri et al.294 also reported stability of succinylated ConA in A-ILCS of [(C2)2NH2][dhp], [(C2)3NH][dhp], [(C2)2NH2][HSO4], [(C1)3NH][dhp], and [(C2)3NH][HSO4] with 50% sodium acetate buffer (v/v).294 The studied IL cations failed the Hofmeister series to explain the stability of succinylated Con A, despite being “chaotropic”.294 Contrary to this 50% A-ILCS, [(CH3)3NH]+ with a “kosmotropic” anion perfectly follows the Hofmeister series (SO42– > HPO42– > CH3COO–) in regard to the succinylated Con A stability.295 Therefore, when explaining protein stability in A-ILCS based on the Hofmeister series, it is actually the cation series or new organic anions which should follow the order to be considered as part of the new development. Additionally, quaternary ammonium-based ILs cannot be universalized for protein stability and the effect can reverse either by changing the anion or protein structure. For example, quaternary ammonium-based A-ILCS with the OH– anion, viz ([(CH3)4N][OH], [(C2H5)4N][OH], [(C3H7)4N][OH], [(C4H9)4N][OH]): 50% sodium phosphate buffer, destabilized the structure of myoglobin and hemoglobin,68 which was due to the direct interaction of OH– with the protein surface, unlike the exclusion effect reported earlier.291,292 In contrast, the presence of the hydroxyl functionality was later cited as the reason for the stabilization of Cytochrome c in A-ILCS.61,63,66 Therefore, the protein-IL interactions are highly specific and vary from NIL to A-ILCS, with different proteins, different ILs, and particular specific IL-protein combinations.
Fujita et al.61 introduced the biocompatibility term to ILs and studied the temperature-dependent secondary structure stabilization of Cytochrome c in 80% A-ILCS of [Cho][dhp] and [P1,4][dhp] in 20% water. Aqueous mixtures of [P1,4][dhp] stabilized Cytochrome c up to 130 °C, compared to 100 °C in mixtures with [Cho][dhp]. The [dhp] anion bears an H-bond donor and an acceptor site, like that of neutral water, being responsible for the high protein stabilization effect. Interestingly, when the water content in A-ILCS was increased up to 80%, denaturation of protein was observed at much lower temperatures (77 and 62 °C for [Cho][dhp] and [P1,4][dhp]). Therefore, as a solute at low concentration, the [dhp] anion acted like a buffer solution in the thermal destabilization of the protein. This work demonstrated the opposite role exerted by IL ions at low and high concentrations of water, in which an optimized amount of water is beneficial to keep the stability of Cytochrome c.61 Working on the time-dependent stability of Cytochrome c, Fujita et al.63 reported excellent stability of the protein in 80% A-ILCSs (IL:H2O, 80:20, v/v) of cholinium-based ILs possessing different anions. The structural and functional activity of the enzyme was explained based on the kosmotropic order of the IL anions: [dhp] > [(C4H9)2 PO4] > [OAc] > [Lac] > [CH3SO3]. Still, the activity observed was highest for [Cho][dhp], which is the most suitable combination of a “chaotropic” cation with a “kosmotropic” anion. Remarkably, the long-term stabilization of Cytochrome c for up to 6–18 months was observed in A-ILCS with 80% [Cho][dhp] in water.296,63 This report showed the relevance of A-ILCS for a stable in vitro kinetic (long-term) packaging of proteins and TPs. Mazid et al.,297 from the same research group (MacFarlane and co-workers), investigated the biological structure and chemical stability of epidermal growth factor receptor monoclonal antibody (EGFR mAb) in cholinium-based buffered IL solutions, namely, [Cho][dhp]:H2O (20:80 and 50:50 v/v) stored at room temperature from 7 h to 7 days in the presence of proteinases. The EGFR mAb retained its α-helical structure and activity, as evidenced by successful binding to its cell receptor, while indicating the potential of this mixture for the packing of antibodies. Interestingly, a higher stability was observed at 50% [Cho][dhp] compared to 20% [Cho][dhp], which is in line with Fujita et al.61,63 However, both structural stability and functional activity showed a decrease with time. In 2010, Fujita et al.66 again showed the relevance of [Cho][dhp]:H2O (70:30, v/v) A-ILCS, but now on the enzymatic activity and thermal stability of several metalloproteins in addition to Cytochrome c, namely peroxidase, ascorbate oxidase, azurin, pseudoazurin, and fructose. Overall, this hydrated IL, i.e., [Cho][dhp], has indeed been one of the most promising ILs identified to keep the integrity of protein’s structure.298 The stability effect of [Cho][dhp] on different proteins in A-ILCS is illustrated in Figure 14.
Figure 14.

Stability effect of A-ILCS of [Cho][dhp] on different proteins.61,63,66,296,298,299
The relevance of [Cho][dhp] for protein stabilization proven by Fujita et al.61,63,66,296 was further exploited by Weaver et al.,299 who investigated in detail the effect of [Cho][dhp] on the thermodynamics, structure, and stability of lysozyme and Interleukin-2 as a TP. Both proteins were found to be soluble in [Cho][dhp] at all tested concentrations, but solubility was limited at 80% [Cho][dhp] in water. Increasing the amount of [Cho][dhp] present in the aqueous protein solution resulted in an increased thermal transition temperature (Tm), without significant changes in the protein’s tertiary structure for both proteins. Binding analysis with the lysozyme using isothermal titration calorimetry (ITC) showed that stability is not induced by the IL binding to the lysozyme. Thermal stabilization at low IL concentrations is due to shielding effects, where the net charge on the surface of the protein plays a key role; at high IL concentrations, stability is more dependent on solution properties. This work is especially promising since it addresses a TP, interleukin-2, paving the way for the use of IL-water mixtures to stabilize other TPs and other biopharmaceuticals. Using calorimetric and spectroscopic analysis Dhiman et al.300 reported enhanced thermal transition temperature (Tm) and 2 weeks of room-temperature structural stability of TP, immunoglobulin G (IgG) in A-ILCS of [Cho][OAc] and [Cho]Cl (up to 2.5 mM of ILs in water).300 Kumar et al.301 also showed the thermal stability of TPs, namely of insulin, in A-ILCS of protic quaternary ammonium ILs (from 20% to 80%), namely, [(C1)3NH][HSO4], [(C2)3NH][HSO4], [(C1)3NH][dhp], [(C2)3NH][dhp], and [(C2)3NH][OAc]. The authors showed that A-ILCS stabilizes insulin in its active monomeric form, revealing the role of compatible quaternary ammonium-based ILs in the packaging of TPs.
Other works regarding A-ILCS of quaternary ammonium-based ILs have shown the stability and enhanced activity of Cytochrome c.302−306 For instance, Papadopoulou et al.302 reported the stability and a 20-fold increase in the activity of Cytochrome c in [OHC2NH3][HCO2] (60% in water), resulting from the high “chaotropicity” of the cation. Bhakuni et al.303 showed the thermal stability and packaging of Cytochrome c for a month, with a 1.5-fold rise in activity in an IL-based medium comprising [Cho][dhp] (50%) and synthetic crowding agents, like Fycol + ethylene glycol + sucrose (5%). Matias et al.304 reported the retention of intrinsic redox properties of Cytochrome c in an aqueous solution of [Cho][dhp] and [Cho][MES]. Still, with cholinium-based ILs, Bisht et al.305 demonstrated the thermal and thermodynamic stability of Cytochrome in A-ILCS of [Cho][Glu]:water (50:50, w/w) under multiple stresses, such as temperature and H2O2. The authors reported a 50-fold increase in Cytochrome c activity, thermal stability up to 120 °C, and thermodynamic stability up to 5 months under ambient conditions, thus demonstrating the good packaging ability of the studied A-ILCS toward Cytochrome c. With a different perspective, Takekiyo et al.306 studied the cryopreservation effect of aqueous IL solutions of [C4mim][SCN] and [C2NH3][NO3] on Cytochrome c, concluding that although these ILs denature the Cytochrome c before and after cooling to −196 °C, >90% of the enzyme activity following cryopreservation was recovered. Overall, it is clear that it is important to analyze the structure of proteins in water after treatment or storage in A-ILCS to know their structural changes.
The described studies show that aqueous solutions of quaternary ammonium-based A-ILCS in combination with suitable anions allow the efficient packaging of Cytochrome c. However, this notion can change for other proteins with slightly different IL cation–anion combinations. For instance, A-ILCS of [Cho][Gly] led to poor stability of Stem bromelain.307 While the authors presumed H-bonding of the glycinate anion with the protein peptide backbone as the main reason for the observed effect, this is in stark contrast with the long-term stability of Cytochrome c in A-ILCS of the same IL, wherein the glutamate anion has more H-bonding sites. We do believe that the observed effect could be due to the specificity of IL-protein systems (Figure 15). Further evidence of specificity is found in the destabilization of α-chymotrypsin by A-ILCS of [Cho][OH] and [Cho][Cit]308 and hemoglobin and myoglobin by A-ILCS of [(CH3)4N][OH], [(C2H5)4N][OH], [(C3H7)4N][OH], and [(C4H9)4N][OH].68
Figure 15.

Specificity of quaternary ammonium-based A-ILCS-protein systems.301−307,68
Beyond packaging, several other applications of A-ILCS in protein unfolding-refolding, fibrillation, and crystallization were studied. For example, Fujita et al.309 reported that [Cho][dhp] with 4 water molecules per ion pair showed the best refolding tendency toward the aggregated recombinant cellulase obtained from Escherichia coli (CcCel6A). To understand the mechanism of refolding, the authors studied a series of A-ILCS with phosphonium and quaternary ammonium cations combined with bromide, chloride, and dihydrogen phosphate (dhp) anion. These mixtures were restricted to a limited number of water molecules per ion pair (3 to 15) on the solubilization and refolding of aggregated Concanavalin A (ConA).310 The authors concluded that the solubilization of aggregated ConA in the studied A-ILCS decreases with an increase in the water content, wherein [P44412][dhp] and [N8888][dhp] were found to be the most effective refolders due to the stronger H-bonding established between the anion and the protein.310 Constantinescu et al.311 investigated the patterns and equilibrium of unfolding-refolding and aggregation of RNase A in A-ILCS (0.5 to 4 M IL in water) employing the Lumry-Eyring scheme. The A-ILCS of 0.5 M [Chol][dhp] increased the thermal stability of RNase A, wherein the unfolding transition is about 66% completed at 70 °C compared to complete unfolding in an IL-free sample. Moreover, A-ILCS of [Cho][dhp] refolded 5 to 90% of the structure of heat-denatured RNase A, being the system found as the most efficient in suppressing the RNase A oligomerization.311 Byrne et al.312 improved lysozyme stabilization by mixing two concepts, namely, conventional sugar protection and proton activity of ILs, and reported aggregation protection against reversible folding-unfolding and multiyear (3 years) stabilization of lysozyme in A-ILCS of [CH3CH2NH3][NO3]. The solution developed is composed of sucrose (27 wt %), [CH3CH2NH3][NO3] (31 wt %), water (20 wt %), and lysozyme (22 wt %). However, a small decrease in the denaturation enthalpy (Tm = 74 °C), from 20.70 J·g–1 to 19.72 J·g–1, was observed up to the third cycle. When subjected to DSC measurements, no changes in the unfolding enthalpy (dHU) were observed for cold-stored lysozyme, whereas a 1/3 decrease in dHU was observed, indicating a 2/3 loss of protein to aggregation or hydrolytic decomposition. This work represents a significant development because protein samples usually stored in a refrigerator begin to aggregate after 7 days of storage. Therefore, the retention of 25% of the protein structural integrity after 3 years of incubation at room temperature is one outstanding achievement that should be further investigated, and the respective underlying molecular-level mechanism should be depicted.
Further understanding of the unfolding-refolding mechanism was sought by investigating the effect of proton activity (PA) of ILs on the denaturation temperature of lysozyme and RNase A, as a solution property parallel to that of pH in aqueous solution in the form of a two-state cooperative model (Native ↔ Unfolded).313 The authors showed that the unfolding temperature is altered by changing the PA, like what happens when altering the pH. An increase in the thermal stability of both the proteins was observed with an increase in the δ (N–H) shift. Finally, the authors developed a refolding index (RFI) metric to assess the stability of folded biomolecules in different solvent media and demarcated high RFI zones in hydrated PIL media using RNase A and lysozyme as model proteins.314 Wijaya et al.315 also used pH as a parameter to understand the effect of A-ILCS of [C2NH3][NO3] and [OHC2NH3][NO3] in 4–17% on the structure and activity of lysozyme. They found that the enzyme retains its α-helical structure, except in the acidic pH, and showed functional activity in the entire pH range (0–11) in the [C2NH3][NO3]-water mixture and from pH 4–11 in the [OHC2NH3][NO3]-water mixture. This is an important work considering that proteins are susceptible to pH-induced conformational changes. Mann et al.316 reported refolding of heat-denatured lysozyme in A-ILCS of [C2NH3][HCOO] and [C1OC2NH3][HCOO] at 25 wt %, where the A-ILCS of [OHC2NH3][HCOO] has shown to stabilize lysozyme against unfolding at high temperature due to strong H-bonding.316
Many proteins are prone to aggregation in an aqueous solution since the fibrillar state of a protein is the most thermodynamically stable, corresponding to its global energy minima. The best biological examples in this field are β-Amyloid, tau, and α-synuclein, responsible for neurodegenerative diseases, like Alzheimer’s and Parkinson’s diseases. Accordingly, A-ILCSs have been used as well to suppress protein fibrillation. Takekiyo et al.317 reported the suppression effect of insulin amyloid formation using aqueous solutions of the following ILs (from 0 to 30 mol %): [C4mim][SCN], [C2NH3][NO3], and [C3NH3][NO3]. Notably, at 30 mol % of IL, [C3NH3][NO3] showed a protective effect on the monomeric form of the protein. Byrne et al.152 also reported the fibrillation of lysozyme in PIL aqueous solutions and subsequent dissolution with the restoration of activity. The authors fibrillated the lysozyme with 80% NH4HSO4·20H2O and then redissolved the fibrils by the addition of [C2H5NH3][NO3], [(C2H5)3NH][CH3SO3], and [(C2H5)3NH][TfO]. Overall, [OHC2H5NH3][NO3] proved to be the most efficient IL in solubilizing the fibrils as it restored 72% of the activity, thus reinforcing the potential of this IL in biomedical applications targeting neurological diseases. However, besides the negative effects of protein fibrils, they can be beneficial when employed as biomaterials for applications in biotechnology, for example as a 3-D matrix for cell-grown and nanomedicines delivery. However, preparing fibrils makes use of pure and expensive proteins, further having long synthesis kinetics. Bharmoria et al.318 overcame this drawback by investigating the anion-specific interactions of ILs with the protein, allowing the instant fibrillation of the aqueous egg white proteome using an adequate IL, [Cho][Tos] (0.1 to 1 M in water). This IL contains interaction sites for ionic, hydrogen bonding, and π–π stacking interaction. Using several experimental techniques and computational studies, they found that H-bonding and hydrophobic interactions occurring between the IL and proteins are the driving forces to induce fibrillation, with additional beneficial effects afforded by ILs with aromatic anions by allowing the establishment of π···π interactions. The prepared fibrils were investigated for cytocompatibility by exposing the mouse fibroblast L929 cells to fibrils using a live–dead assay. The cells perfectly retained their membrane integrity and morphology, thus indicating cytocompatibility and possible application as a matrix for cell growth in biotechnology. The contrasting role of A-ILCS as fibril dissolving agents and fibril promoters, which is achievable due to the ILs’ designer solvents nature, is presented in Figure 16.
Figure 16.

Contrasting role of quaternary ammonium-based A-ILCS on protein refolding and fibril dissolution/promotion.
Beyond the quaternary ammonium family of ILs, several hydrophilic imidazolium-based A-ILCS were investigated to improve protein stability, denaturing/renaturing, and crystallization. In this field, Takekiyo et al.319 studied the effects of the nano heterogeneous structure of [C4mim][NO3] on protein conformation in water. Unfolded state of lysozyme was observed at 6 M [C4mim][NO3] due to specific interactions of the anion with lysozyme, and a partially globular state was observed at 10 M [C4mim][NO3] due to the reduced hydration of unfolded lysozyme.319 Yoshimura et al.320 reported the cryoprotection effect of A-ILCS of [C4mim]Cl and [C2NH3][NO3] from 0 to 30 mol % on lysozyme. A small loss in structure and activity of lysozyme occurred upon cooling to −196 °C, which was reversed upon returning to ambient temperature. Tavares et al.321 reported the maintenance of activity of laccase in A-ILCS of [C2mim][MDEGSO4], [C2mim][C2OSO3], and [C2mim][C1SO3] (10 to 75% in water) at pH 9.0, in addition to just a 10% loss in activity for 1 week of incubation in A-ILCS of [C2mim][MDEGSO4].321 Ferdjani et al.322 reported the activity and stability of a β-glycosidase (Thermus thermophilus) and two α-galactosidases (Thermotoga maritima and Bacillus stearothermophilus) in A-ILCS of [C1mim][C1OSO3] and [(C1)2mim][C1OSO3] (30–80%). Kim et al.323 reported a higher activity of Candida antarctica lipase B in A-ILCS of [C4mim][TfO] due to the stabilization of enzyme active sites. This concept was further vindicated by Nordwald et al.,324 who engineered the active site of lipase A (lipA) from Bacillus subtilis to increase resistance against 50% [C4mim]Cl in water.324 Using the SANS technique, Heller et al.71 reported dimeric to monomeric conformational transition of the green fluorescent protein (GFP), with a decrease in thermal denaturation in 25 and 50 vol % of [C4mim]Cl. Further analysis of IL-GFP interactions using a multitechnique approach in 1 M [C4mim]Cl, [C4mim][OAc], [C4C1py]Cl, [C4C1py][TfO], and [C4C1py][OAc] revealed direct interactions of IL ions with the protein surface, dominated by the type of anion to induce secondary/tertiary structural changes.325 In this direction, Lou et al.326 investigated the role of imidazolium IL paired anions ([BF4], [HSO4], Cl, [NO3], and [OAc]) on the structure and function of papain in A-ILCS (15% ILs in water). The authors concluded that the anion has a high impact on the structure, activity, and enantioselectivity of papain, since it was more stable in [Cnmim][BF4] (n = 2–6) containing systems compared to [C4mim][HSO4], [C4mim]Cl, [C4mim][NO3], or [C4mim][OAc]. The strong nucleophilicity of lower stabilizing anions caused higher interactions with positively charged sites on the enzyme and breaks internal H-bonding or forms new hydrogen bonds that perturb the enzyme structure. Additionally, due to the strong acidity, [C4mim][HSO4] broke the disulfide bond between Cys-56 and Cys-95 located in the helical domain of the papain molecule, leading to the denaturation of the protein.326 Jha et al.327 investigated the effect of the alkyl side chain length of imidazolium-based ILs; [Cnmim]Cl (n = 2, 4, 6, 10) from 0.01 to 1.5 M was applied to address the structural stability of Stem bromelain. It was concluded that their destabilizing effect increases with the cation alkyl chain length and consequent increase in hydrophobic interactions with the protein backbone.327 Figueiredo et al.328 reported that the hydrating ability of the IL anion and hydrophobicity of the cation dominates the stability of a small α-helical protein, when studied in 240 mM to 1000 mM IL ([C4mim]Cl, [C4mim][dca], [C2mim], and [C2mim][dca]) solution in water. Weakly hydrating anions, like [dca], dehydrate the positively charged residues on the protein backbone, whereas slightly more hydrophobic cations like [C4mim]+ cause higher denaturation. A summary of the main conclusions obtained from imidazolium-based A-ILCS on protein denaturation and unfolding is presented in Figure 17.
Figure 17.

Illustration of the main conclusions obtained from imidazolium-based A-ILCS on protein denaturation and unfolding.
Many reports used the internal fluorescence of tryptophan as a marker to investigate protein unfolding or denaturation. However, it is not a confirmatory technique since an increase or decrease in tryptophan fluorescence can happen due to multiple factors, like the inner filter effect and change in solvent polarity, other than quenching due to IL binding.69 Therefore, it is important to use advanced characterization tools such as SAXS, SANS, and fluorescence correlation spectroscopy to obtain correct information on IL-protein interactions. For example, from SANS and SAXS analysis, Baker et al.70 reported the denaturation of Cytochrome c and HSA to random coil conformation in 50% [C4mim]Cl in water, behaving like classical denaturants such as Guanidinium-hydrochloride and urea. In a series of works, Bhattacharya and co-workers112,113,329 exploited the fluorescence correlation spectroscopy technique to understand the protein dynamics in A-ILCS. Sasmal et al.329 reported the opposing effect of [C5mim]Br on conformational dynamics (unfolding and refolding) of HSA when in native or denatured form by fluorescence correlation spectroscopy (FCS). FCS is also a useful tool to measure hydrodynamic radii (Rh) of colored protein solutions whose absorption/emission interfere with the excitation laser wavelength of the dynamic light scattering (DLS) instrument. For example, A-ILCS of [C5mim]Br showed contrasting interaction behavior with human serum albumin (HSA), wherein it unfolded the native conformation and refolded the denatured conformation of HSA.329 During unfolding-refolding, the dynamics of the protein side-chain changed faster at τR= 3–40 μs, whereas interchain interactions occurred at a slow time scale of τR = 100–300 μs. The authors further investigated the change in the microenvironment of HSA in the presence of 1.5 M [C5mim]Br and 6 M GdnHCl from femtosecond up-conversion using 7-diethylamino-3-(4-maleimidophenyl)-4-methylcoumarin (CPM) as a covalently attached probe to cys-34. They found a decrease in solvation time (τs) of HSA from 650 to 260 ps (∼2.5 times) and 60 ps (∼11 times) in the presence of 1.5 M IL and GdnHCl, respectively, due to protein unfolding. Upon refolding analysis, a ∼2-fold faster solvent relaxation (τs ≈ 30 ps) compared to GdnHCl denatured HSA was found.112 Hence, different conformations of proteins can be easily identified by getting information on their solvation using FCS, which is an advance relative to the conventionally known Rh analysis-based DLS technique.112,113,329 A similar behavior was observed during FCS analysis of Cytochrome c in A-ILCS of [C5mim]Br using Alexa Fluor 488 as probe.113 The different relaxation of solvent around different conformations of Cytochromes could help to identify native, molten globule (MG-I and MG-II), and refolded conformation of Cytochrome c.114 These studies also indicate that IL-based refolded proteins never attain the native conformation since they show different solvation dynamics and may operate differently when used in different applications.
While A-ILCSs are important for protein packaging, another application of high interest is to promote protein crystallization by tuning the properties of water. Protein crystals hold high relevance when elucidating the correct structure of the protein. If IL ions are trapped during crystallization, it would give correct information about their binding sites. The most suitable water-IL systems reported are depicted in Figure 18.
Figure 18.

Most suitable water-IL systems for protein crystallization.330,333,334
Pusey et al.330 studied the crystallization tendency of A-ILCS comprising 0.4 M [C4mim]Cl, [C4mim][MDEGSO4], and [P1,4][dhp] on the crystallization of canavalin, β-lactoglobulin B, xylanase, and glucose isomerase. All protein-IL combinations produced at least one crystal, in which the ILs were proposed to act as a primary precipitant agent.330 Hekmat et al.331 reported enhanced lysozyme crystallization kinetics with reduced polymorphism in A-ILCSs (2.5 to 10% of ILs in water) of [OHC2NH3][HCO2], [C2NH3][NO3], [(CH3OC2H5)2NH2][OAc], [OHC2H5(CH3)2 NH] [Gly], and [Cho][dhp]. Soft anions such as formate or glycolate are superior to ILs with hard anions, like nitrate, in promoting crystallization. Wang et al.332 reported enhanced thermal resistance of lysozyme crystals prepared in 1% [C4mim]Cl, which is due to the change in lysozyme-lysozyme molecular interactions. This IL promoted the nucleation of crystals with small size, lower surface free energy, and improved morphology. Chen et al.333 reported that 3.5% [C4mim]Cl in water lowered supersaturation conditions to promote nucleation of lysozyme at ambient conditions. The X-ray diffraction pattern of the crystals showed the incorporation of the imidazolium cation into the crystal framework of the protein, where it is bound to the N and C atoms of Trp62, the C atoms of Trp63, and the O atom of Asp101.333 This is solid evidence of specific molecular interaction between IL ions and amino acid residues of proteins.
In depth mechanisms of the IL on protein’s crystallization was further investigated by Kowacz et al.,334 who studied the effect of the IL type and its concentration (0.0625 to 1 M) on the crystallization of lysozyme and RNase A. The ILs used were [C1mim][C1OPO3], [C2mim]Cl, [C2mim]Br, [C2mim][SCN], [C2mim][OAc], [C2mim][C2SO3], [C2mim][C4SO3], [C4mim][Cl, [C4mim][OAc], [Cho]Cl, [Ch][OAc], [Ch][Lac], and [Ch][C1SO3]. The authors explained that IL-protein molecular interactions drive protein crystallization, based on a combined effect of the intrinsic properties of ILs and their ionic strength. The relative salting-in/-out ability of ILs to assist crystallization changes upon increasing or decreasing their ionic strength. At low ionic strength, when the electrostatic interactions predominate, the ions screen the charge on the protein surface by preferential binding to avoid repulsive interactions between the similarly charged protein molecules, thus facilitating protein nucleation to promote crystallization. At high ionic strength, the IL ions influence the crystallization by altering the protein–water interfacial tension therein. ILs stabilize the protein in solution by lowering the protein–water interfacial tension via hydrophobic adsorption on the protein surface. As a result, anions that exert the highest salting-out effect at low ionic strength become the most prominent salting-in agents when hydration forces start to dominate at high ionic strength, thus inhibiting the seeding of protein crystallization.334 The molecular mechanism exerted by ILs on protein crystallization at low and high ionic strength is depicted in Figure 19.
Figure 19.

Schematic representation of interactions of a protein containing hydrophobic and cationic surface groups with its surroundings (IL ions, water, and inorganic ions). At low ionic strength, preferential binding of weakly hydrated anions to the cationic residues causes charge screen, resulting in the salting-out of the protein. The water-mediated interactions: electrostatic stabilization of hydration (salting in) and exchange of water molecules hydrating the protein with positively (salting-out) and negatively (salting-in) hydrated ions. At high ionic strength, nonspecific adsorption of weakly hydrated ions to the hydrophobic surface reduces the protein-water interfacial tension resulting in the salting-in of the protein. Adapted with permission from ref (334). 2012 The Royal Society of Chemistry.
Further evidence of the molecular mechanism of ion-protein interactions was obtained from simulation studies of A-ILCS-protein systems. From MD studies, Jaganathan et al.335 confirmed stable compaction of Cytochrome c, solvated by [C2NH3][NO3] (0.1 to 1 mM in the water), due to H-bonding between [C2NH3][NO3] and water, and electrostatic interactions between [C2NH3][NO3] and charged residues on the protein surface. This rationale supports the experimental results of A-ILCS-protein interactions for the quaternary ammonium family of ILs. Burney et al.336 reported stability of lysine to glutamate surface engineered Bos taurus α-chymotrypsin and Candida rugosa in A-ILCSs of 20% [C4mim]Cl and [C2mim][C2OSO3] in water, which was due to the clustering of cation and decreased anion interaction with the protein surface. The role of anion-specific interactions was further studied by Latif et al.337 on the dynamic properties of Candida antarctica lipase B and Candida rugosa lipase solvated by [C4mim[PF6], [C4mim[BF4], [C4mim]Cl, [C4mim[TfO], and [C4mim[Tf2N] (10% to 50% ILs in water). The ILs destabilized the enzymes by stripping off water from the hydration layer, in the order [Tf2N] < [PF6] < [TfO] < [BF4] < Cl. Li et al.338 reported competitive binding of [C4mim]+ at the active site of cellulase cellobiohydrolase I (CBH I) at high [C4mim]Cl concentration, inducing the inactivation of the enzyme. Manna et al.339 also reported a loss in activity of endoglucanase Cel12A from Rhodothermus marinus (RmCel12A) in 20%, 40%, and 60% aqueous solution of [C2mim][OAc] due to the intrusion of [C2mim] cation into the active site of the enzyme, thus affecting the dynamic motions around the active site.337 The cation-specific IL-protein interactions were further observed with A-ILCS of [C4mim]Cl on three α-helix bundles (the domain of protein A from Staphylococcus aureus (BdpA)). The authors cited the preferential accumulation of the [C4mim] cation at the protein surface as the reason for protein stabilization in 25%, 45%, and 65% of [C4mim]Cl in water. The accumulated [C4mim] cation reduces the H-bonding between water and the protein by displacing the water molecules from the protein surface, hence stabilizing the backbone hydrogen bonds.339 Using enhanced molecular dynamics simulations, Jaeger et al.340 reported closeness of HSA solvated in concentrated A-ILCS of [C4mim][BF4] and [Cho][dhp] with its crystal structure studied on time scales of hundreds of nanoseconds. Moreover, HSA solvated in 20% [Chol][dhp] in water was found to have similar structural stability to that found in water. The MD simulation studies thus support the specificity of IL-protein interactions for both stability and destabilization in A-ILCS.
3.2.1. Mechanisms of Interaction
From the discussed works involving A-ILCS and proteins, three major mechanisms of interaction have been found to rule the protein’s stability: (1) hydrophobic solvation; (2) dual-hydrogen-bonding; and (3) preferential exclusion of ions from the protein surface.
3.2.1.1. Hydrophobic Solvation
When considering hydrophobic ILs like [C4mpy][Tf2N], [(C2)2mim][Tf2N], [C2mim][Tf2N], [C4(C3)3N][Tf2N], [C4(C1)3N][[Tf2N], and [C2(C1)2C3N][[Tf2N], the stabilization of proteins is a result of their rigid solvation in their compact form via hydrophobic interactions and molding into the polar domains of A-ILCS (Figure 12).59,60,62 Solvation is driven by hydrophobic interactions with the hydrophobic patches on the protein surface. The water molecules dissolved in [Tf2N]-containing ILs behave as free water, interacting via H-bonds with the anions of the ILs, thus providing the desired aqueous-like microenvironment for protein solvation. Going by the secondary structure, three of these proteins (Lysozyme, CAL B, α-Chymotrypsin) possess α+β, and each one possesses α/β and all-β secondary structural conformation. Therefore, these proteins already contain a significant amount of β structure with lesser structural flexibility, which might be assisting in their stable compaction in hydrophobic ILs.
3.2.1.2. Preferential Exclusion
Preferential exclusion has been particularly observed in the quaternary ammonium family of ILs.67,291−295,301 The stabilization is caused by unfavorable interactions of the IL ions with the protein surface, due to which ions get excluded, thus causing stabilization by a phenomenon similar to the osmophobic theory of Bolen213 and the transfer Gibbs free energy concept (Figure 13).215,216 In such cases, IL ions behave similarly to normal osmolytes in preserving the protein structure.
3.2.1.3. Hydrogen Bonding
Hydrogen bonding between IL ions and the protein surface has accounted for both protein stability and destabilization. For example, the most promising IL identified in the set of works considered is [Cho][dhp], which stabilized, among other proteins, the structurally flexible all-α protein Cytochrome c for 18 months.61,63,66,296,298 The [dhp] anion was shown to interact with proteins both by donating and accepting protons through hydrogen bonding.61,63 Therefore, the water-like dual H-bonding character of [dhp] anion provides a beneficial environment to proteins, whereas the rigid matrix of the IL prevents aggregation-induced denaturation by restricting proteins at specific locations in the ionic matrix (Figure 20).
Figure 20.

Mechanisms of dual hydrogen bonding, such as with the [dhp] anion leading to higher stabilization of the protein structure.
These restrictions in the ionic matrix limit the protein’s diffusion and protein–protein interactions, which are the major cause of their aggregation. The stabilization effect of [Cho][dhp] is not restricted to Cytochrome c only and has also been applied to stabilize lysozyme299 and refolding of RNase A311 and cellulase,309 thus showing its broad-range application for proteins of different secondary structures. In contrast, anions other than [dhp], but still carrying hydrogen bond sites, such as [OAc], [CH3SO3], Cl, and [OH],68−70,305,319 have been reported to destabilize the protein structure. This is because these anions do not contain dual H-bonding donor sites like [dhp], and they mostly contain H-bond acceptor sites. In these ions, the ionic interaction with the cationic sites on proteins results in the disruption of the existing hydrogen bonding network of proteins to cause unfolding. Like [dhp], the [HSO4] and [Lac] anions contain dual H-bonding sites, but each one for H-bond donation and H-bond acceptation, which could be the possible reason for their denaturation effect on proteins.326 Interestingly, the high acidity of [HSO4] anions was also cited as the reason for breaking the disulfide bonds of the protein papain, inducing denaturation at high concentration (15% in buffer solution);326 however, it should be noted that [dhp] is also very acidic and stabilizes proteins.61,63 These results show that it is not just cation-protein or anion-protein interactions, but also the cation-anion interactions on the protein surface, that govern the overall effect on the stability of proteins in A-ILCSs.
3.3. Ionic Liquids as Adjuvants
Going further into the dilution of ILs in water, in which their structure and solution properties vary significantly, leads to the ILs here defined as adjuvants (concentrations lower than 2%). It should however be kept in mind that even in dilute solutions, ILs do not exist in the form of isolated ions as conventional inorganic electrolytes, but rather in associative forms, such as triple ions, contact ion pairs, solvent shared ion pairs, and loose ion pairs.24,27 These forms are reflected in a different interaction behavior with proteins than that observed with typical high-charge density salts. In the field of using ILs as adjuvants (A-ILAS), the systems have been studied in pursuit of understanding the mechanisms of interaction of ILs with proteins, based on Hofmeister effects, and other noncovalent interactions for protein stability, activity, fibrillation, etc.
Constantinescu et al.341 investigated the thermal denaturation tendency of ILs on RNase A and formulated a Hofmeister series in comparison to inorganic salts at pH 7.0. The cation series of Hofmeister toward RNase A denaturation follows the order K+ > Na+ > [C1,1,1,1N]+ > Li+ > [C2,2,2,2N]+ > [C2mim]+ > [C4mpyrr]+ > [C4mim]+ > [C3,3,3,3N]+ > [C6mim]+ > [C4,4,4,4N]+, whereas the order of anions is [SO4]2– > [HPO4]2– > Cl– > [C2OSO3]− > [BF4]− ≈ Br– > [C1OSO3]− > [TfO]− > [SCN] > [N(CN)2]− > [Tf2N]−. In comparison to inorganic cations and with the exception of [C1,1,1,1N]+, all organic cations destabilize RNase A despite their chaotropic nature, which is against the notion of the modern Hofmeister series wherein chaotropic cations are considered as protein stabilizers (Figure 3).91−95 Accordingly, there are forces beyond hydration effects which dictate IL-protein interactions in diluted aqueous solution. Weibels et al.342 investigated these forces by the ion specific activity analysis of yeast alcohol dehydrogenase in dilute aqueous solutions of ILs. The authors found hydrophobic interactions as a key factor governing the decreased activity of the enzyme, with the following Hofmeister series of cations and anions: Cl– ∥ Br > [C2OSO3] > [TfO] > [BF4] > [dca] > [SCN] and Na+ > [Me4N]+ > [Chol]+ > [C2mim]+ > [Et4N]+ > [Bu4N]+ > [Gdm]+ > [C4mim]+. Kumar et al.343 performed comparative analysis of the Hofmeister series of anions with sodium and the [C4mim]+ cation on α-chymotrypsin. The ILs studied were [C4mim][HSO4], [C4mim][SCN], [C4mim]I, [C4mim]Br, and [C4mim][OAc]. The comparative Hofmeister order of sodium- and [C4mim]-paired anions based on the stability against thermal denaturation follows the order SO42– > Br– > I– > SCN– > [OAc]− > Cl–, and [OAc]− > Br– > Cl– > HSO4– > SCN– > I–. The authors concluded that the Hofmeister anions combined with the sodium cation were a complete denaturant for the CT structure. On the other hand, a combination of the same anions with [C4mim]+ presented the reverse effect on the CT native structure. Therefore, the stabilization/destabilization effects may vary for cation/anion combination, protein, and pH.
Zhao et al.18 have reviewed the ion-specific effect of ILs toward protein stabilization/enzyme activity and concluded that in dilute aqueous-IL solutions, with several exceptions, the ion specificity of many enzymatic systems is in line with the traditional Hofmeister series/kosmotropicity; however, the specificity in concentrated or neat ILs is determined by β, the nucleophilicity of anions, hydrophobicity of IL, and other factors.18 Kumar et al.344 also reviewed the Hofmeister effect of ILs on proteins and concluded that the interactions are dominated by ion specificity and solvent environment related to H-bond disruption, nonpolar interactions, and electrostatic effect to stabilize or destabilize the protein. The bottom line is that the Hofmeister-type effect of ILs is inconsistent in the literature and cannot be generalized. This trend may be because ILs simply do not follow the concept on the grounds of hydration effects alone. Beyond the Hofmeister effect, the preferential electrostatic binding of [C4mim]+ on α-chymotrypsin and Candida rugosa lipase,345 and [Amim]+ on Hemoglobin346 surface, with exclusion of the counterion Cl into the bulk solution, was reported as the reason for protein conformation stability. Similarly, double layer clustering of [C6mim]Cl and [C8mim]Cl at the protein–water interface was reported as the reason for stable dispersion of proteins, as verified with BSA, HSA, IgG, β-Lg, and Gel-B around their isoelectric point, pH 5.0, in an electrolytic solution of IL.347 The surface selective binding of IL molecules to proteins screens their surface charge, leading to pH-independent dispersion stability by arresting their protonation and deprotonation in aqueous solution. This notion is very similar to the Poisson–Boltzmann, Debye–Hückel, and DLVO theories of colloidal stability of proteins in inorganic salt solutions.197−201 Therefore, these theories do find validation with organic salt solutions in the form of ILs as well.
Information availed from colloidal stability of proteins is limited to aggregation-induced destabilization effects, whereas molecular level information on IL-protein interactions could be obtained from spectroscopic assays or docking studies. For example, using small-angle neutron scattering, Heller et al.348 reported that the thermally driven unfolding and aggregation of HSA by GdnHCl share a common pathway with [C4mim]Cl and [C4mim][OAc], wherein [C4mim]+ and Gdn+ cations associate with the surface of the protein to induce more attractive protein–protein interactions and conformational changes. In these, the anion controls the temperature of unfolding. By combining fluorescence and docking studies, Shu et al.349 showed that cationic imidazolium moieties of ILs, viz. [C4mim][NO3], [C4mim]Cl, and [(C4)2im]Cl, enter the subdomains of BSA and interact with the hydrophobic residues of domain III to induce Trp quenching. Huang et al.350 reported static quenching of BSA fluorescence by ILs according to the following order: [C8mim]Cl > [C6mim]Cl > [C4mim]Cl. ILs interacting with tryptophan (Trp) and tyrosine (Tyr) residues lead to changes in the structure and internal hydrophobic conformation of BSA. An illustration of the comparative effects of ILs versus inorganic salts as electrolytes on proteins is shown in Figure 21.
Figure 21.

Comparative effects of ILs versus inorganic salts as electrolytes on proteins.
In a series of studies, Fan’s research group351−356 exploited the internal fluorescence of tryptophan as a tool to investigate the molecular-level interaction of ILs and the consequent effect on proteins’ secondary/tertiary structure and biological activity. In their first report, showing the [C8mim]Cl interaction with L-tryptophan in water,351 the authors demonstrated that [C8mim]Cl quenches Trp fluorescence by a static mechanism, wherein the IL is weakly associated with Trp via van der Waals interactions and hydrogen bonding. This report had implications on the understanding of the molecular-level interactions of the IL-Trp system, which is generally used as an informative tool to monitor protein unfolding upon interaction with ILs. Following this, the authors reported static quenching of papain’s fluorescence with the decrease in activity by [C8mim]Cl and [C4mim]Cl due to H-bonding and van der Waals interactions.352,353 In a subsequent work,354 they reported quenching of the Trp fluorescence of papain or pepsin by [NH2C2C4im]Br based on static or dynamic mechanisms, either by H-bonding and van der Waals or hydrophobic interactions with the IL, respectively. They also showed that [NH2C2C4im]Br had no obvious effect on the secondary structure of both enzymes, leading to a slight increase in their activities due to the chaotropicity and hydrogen bond donating ability of [NH2C2C4im]+.354 Later, the authors reported the ILs induced activity decrease of lipase due to hydrophobic interactions of the cation and H-bonding interactions of the IL anions and the active site of the enzyme.355 They classified the cations effect in three groups based on decreasing activity, as follows: (I) [C4mim]Cl < [Bzmim]Cl < [C7mim]Cl < [C8mim]Cl; (II) [C4mim]Br < [Bzmim]Br < [C7mim]Br < [C8mim]Br; (III) [C4Py]Br < [C8Py]Br. The order of anions was represented as [C4mim]CF3SO3] > [C4mim]N(CN)2 ≈ [C4mim][ClO4] ≈ [C4mim]Br > [C4mim]Cl > [C4mim][BF4] and [C4mim][TfO] > [C4mim][N(CN)2] ≈ [C4mim][ClO4] ≈ [C4mim]Br.355 To generalize this notion the authors also investigated the effect of 12 ILs on the structure and activity of trypsin from a toxicity point of view.356 The order of ILs for trypsin inhibition was presented as [C10mim]Br > [C8mim]Br ≈ [C6mim]Br > [C4mim]Br; [C10mim]Cl > [C8mim]Cl ≈ [C6mim]Cl > [C4mim]Cl; and [C4mim]Br ≈ [C4mim][NO3] = [C4mim]Cl ≈ [C4mim][BF4] ≈ [C4mim][TfMs] > [C4mim][OAc].
From the thermodynamic analysis, H-bonding supported by hydrophobic interactions was given as the key driving force for structure and consequent activity inhibition of trypsin. Based on these studies, Fan et al.355,356 prepared a regression-based model, which produced satisfactory results to describe the relationship between the inhibitory ability and hydrophobicity or H-bonding ability of ILs toward proteins. This model could be useful and decrease the analysis time when investigating the interactions of ILs with proteins. Ventura et al.357 reported that ILs with higher β and π values are more effective in decreasing the activity of Candida Antarctica lipase B due to direct H-bonding and dispersion interactions with proteins around the active site. Support to the various experimental molecular interaction results also came from molecular dynamics studies, wherein Klähn et al.73 studied thermally induced unfolding of Candida antarctica lipase B (CAL-B) in the presence of a series of ILs, namely, [C4mim][NO3], [C4mim][BF4], [C4mim][PF6], [CH3OC2mim][BF4], acyclic [BAGUA][BF4], cyclic [BCGUA][BF4], cyclic nitrate [MCGUA][NO3], and cyclic [DCGUA][NO3]. They found that the destabilization of the protein surface was mostly facilitated by Coulombic interactions established between the IL anion that exhibits localized charge and strong polarization with the cationic residues, whereas the destabilization of the protein core was facilitated by direct hydrophobic interactions with core and alkyl chains of ILs, further inducing major conformational changes that enabled the access of ILs to the protein core. The surface instability resulted in the unraveling of α-helices, and an increase of surface area and radius of gyration of proteins, whereas core instability resulted in the disintegration of β-sheets due to the diffusion of ions into CAL-B and hence increasing protein-IL van der Waals interactions. Jaeger et al.358 simulated the effect of [C2mim][OAc] and [C2mim][C2OSO3] on the structure and activity of 11 xylanase from Trichoderma longibrachiatum, concluding that the enzyme solvated in higher concentrations of ILs generally remains more stable with respect to its crystal structure than when solvated in water. On the other hand, the decrease in activity arises due to the strong binding of [C2mim]+ to the active site of the enzyme by competitive inhibition. The mechanism of the IL’s interaction with protein surface via favorable interactions is illustrated in Figure 22.
Figure 22.

Mechanism of IL interaction with protein surface via favorable interactions.
From the described studies, it is evident that in the case of protein destabilization by ILs, the process begins with Coulombic interactions with the protein surface, which opens the protein hydrophobic core to facilitate hydrophobic associations. Further unfolding, refolding, or compaction of proteins depends on the IL concentration and its effect on the microenvironment of the protein. For example, Mangialardo et al.359 reported that [C2NH3][NO3] induced significant conformational transition of lysozyme, from fibril state to natural state, observed by a decrease in β-interchain conformation. The [C2NH3][NO3] reversed change in the microenvironment around Phe, Tyr, and Trp residues of lysozyme occurred via fibrillation. Similarly, Sankaranarayanan et al.360 reported the β → helix → β sheet conformational transition of myoglobin induced by an amino acid anion-based IL, i.e., [C1mim][Phe], at different concentrations by altering the polarity around the protein. Sankaranarayanan et al.361 also studied the effect of pH on nonspecific interactions of ILs to induce helix ↔ β conformation transition of bovine β-lactoglobulin at pH 4.0 and 7.5 in the presence of [C2mim][C2OSO3]. At pH 4.0, the protein initially changed to an intermediate β-turn structure, followed by a change to a more stable native β-sheet structure; at pH 7.5, the initial native β state traversed to a helical structure and then returned to the native β state due to the change in the microenvironment around the protein due to the IL concentration. Ghaedizadeh et al.362 reported a decrease in the activity of Renilla Luciferase due to [C4mim][BF4] and [C4mim][PF6] induced transition to a shrunken conformation and a coil-shaped structure due to collapse in α/β fold and reduction in α-helices. Rawat et al.363 reported dissociation of the aggregates of BSA, β-lactoglobulin, and immunoglobulin (IgG) into oligomers by ILs, namely, [C2mim]Cl, [C4mim]Cl, [C6mim]Cl, and [C8mim]Cl. The cations bind selectively to the negatively charged patches on the protein to induce dissociation, whereas the Cl– anion stabilized oligomers via hydration effects. Bisht et al.364 reported the complete refolding of unfolded lysozyme by [C4(C1)3N][NTf2] and [C1(C2)2C1OC2N][NTf2], with a 13% increase in its functional activity by altering the solution properties.
Besides giving valuable information on the molecular mechanism of IL-protein interactions, the electrolytic nature of ILs as adjuvants has also been exploited for applications in oxygen sensing and IL-PAGE for the separation of proteins.365,366 Ding et al.365 reported that a direct electrochemical response of myoglobin could be observed for oxygen reduction from basal plane graphite (BPG) when [OHC2mim][BF4] was used as a supporting electrolyte. The myoglobin adsorbs on the surface of BPG forming a stable monolayer. The biosensor developed could directly detect the concentration of oxygen in an aqueous solution with a detection limit of 2.3 × 10–8 M. Hasan et al.366 exploited pyridinium-based ILs, viz. [CnPyrBr] (n = 4, 8), as buffer additive for high-resolution separation of low and high molecular weight proteins, including catalase, transferrin, BSA, ovalbumin, and α-lactalbumin, in IL-polyacrylamide gel electrophoresis as an alternative to the SDS-PAGE technique.
3.4. Ionic Liquids as Surfactants
Apart from the above-mentioned applications, formulations comprising surface-active ionic liquids (SAILs) and proteins are an emerging area of IL-protein research. SAILs are ILs that behave as ionic surfactants.145 SAILs show aggregation behavior similar to ionic surfactants when dissolved in any medium (polar/nonpolar), but they have different adsorption characteristics and may display superior surface-active properties if properly designed.145 The properties sometimes mimic the ones induced by the addition of bulky organic cations/anions to conventional ionic surfactants.367 Na[DBS], a common laundry detergent, has been compared with the SAIL [C8mim][C12OSO3] for the stabilization of a laundry enzyme—cellulase.120 Comparing the adsorption isotherms, Na[DBS] reduces the surface tension at CAC (2.9 mmol L–1) to 32.6 mN·m–1 and forms micelles,120 whereas [C8mim][C12OSO3] reduces the surface tension at CAC (0.42 mmol L–1) to 26 mN·m–1 and forms vesicles.121
Therefore, [C8mim][C12OSO3] has better surface-active properties than Na[DBS] and is a promising candidate to replace it, if not bound by the constraints of biodegradation. The superior surface-active properties of SAILs could be exploited to formulate more efficient SAIL-protein systems for several applications.143,144,223−226 Most of the reported studies are focused on interfacial and spectroscopic analysis of the SAILs binding to proteins in different concentration regimes, and the consequent effect on their structure/function, coacervation, and storage in confined domains of microemulsions, as shown in Figure 23.
Figure 23.

SAIL-protein systems: (a) SAIL-protein interaction studies; (b) SAIL-protein colloidal complexes; and (c) SAIL-protein microemulsions.
Geng et al.116,117 reported endothermic binding of [C14mim]Br to BSA, leading to (i) the protein secondary structure stabilization due to H-bonding below the CMC and (ii) destabilization due to hydrophobic interactions above the CMC. They concluded that compared to traditional cationic surfactants, [C14mim]Br is superior both in the protection and destabilization of BSA. Kumar and Kang’s research groups118−123 have been actively involved in understanding the effect of the SAILs chain length, headgroup functionality, and amphiphilicity on the structure and function of various proteins to materialize practically feasible formulations. For example, they have reported that by increasing the chain length of both the IL cation and anion, from [C4mim][C8OSO3]118 to [C8mim][C12OSO3],119 the stability of BSA is significantly enhanced.118,119 Increasing the chain length decreases the CAC from 26 mM to 0.39 mM, which reduces the denaturing monomeric binding regime of the SAIL to BSA and hence stabilizes it.118,119 They generalized this idea for colloidal stabilization of BSA in the cat-anionic vesicles of the ILs [C8mim]Br and the laundry detergent [Na][DBS] for 1 week of incubation, showing the relevance of IL-based molecular assemblies for kinetic stabilization of proteins.120 The authors further exploited the higher surface activity and protein stabilizing tendency of [C8mim][C12OSO3] to formulate a stable liquid colloidal formulation with a laundry enzyme cellulase as a candidate for detergent applications.121 For enhanced practical relevance, they synthesized a biodegradable SAIL, [Cho][DBS], and formulated a stable colloidal formulation with cellulase for potential liquid detergent application.368 In an interesting work,122 they further reported that the structure of the SAIL can be tuned to make assemblies that can mimic biological membranes. For this, they synthesized a SAIL, [Cho][AOT], having structural similarity to phosphatidylcholine, which is part of the inner membrane of mitochondria. The bilayers of [Cho][AOT] biomimicked the inner membrane of mitochondria in inducing all-α to α+β conformational transition of cytochrome c, supporting the relevance of this SAIL for studying in vitro membrane-protein interactions.122 Further, they altered the headgroup functionality of the imidazolium cation and reported ordered self-assemblies (long rods and helical fibers) of BSA in a specific concentration regime of amide, [C12Amim]Cl, and ester, [C12Emim]Cl, functionalized SAILs.123 Interestingly, fibers dissolved below and above this concentration regime, which gives an advantage to these SAILs, particularly for long-term stable kinetic packaging of BSA in the form of fibers.123 They also exploited colloidal complexes of the SAILs [C12C1im]Cl, [C12Amim]Cl, and [C12C2mim]Cl with BSA for pH-dependent controlled transport of a lipophilic dye—Rhodamine 6G (R6G).369
Several other research groups have also studied the effect of the cationic headgroup functionality on the structure of proteins. For example, Wang et al.124 reported that ester-functionalized SAILs, [C1COOC2C1im][C12OSO3] and [C1COOC2C1Py][C12OSO3], stabilized the secondary structure of BSA below 1 × 10–3 mol·L–1 and destabilized it above this concentration. The cationic imidazolium was found more destabilizing compared to the pyrrolidinium headgroup.124 Yan et al.125 reported that [C10mim]Br induced marked changes in the secondary structure of BSA, driven by strong hydrophobic interactions, compared to [Cnmim]Br (n = 4, 6, 8).125 Pinto et al.126 reported that the pharmaceutical SAILs cetylpyridinium salicylate ([CetPy][Sal]) and benzethonium salicylate ([Be][Sal]) bind strongly to the hydrophobic sites of HSA and quench tryptophan fluorescence.126 Zhou et al.127 reported higher unfolding of BSA by imidazolium-based gemini surfactants ([Cn-s-Cnim]Br2, n = 10, 12, 14, s = 2, 4, 6) compared to quaternary ammonium surfactants (C12C2C12) and their corresponding monomers ([C12mim]Br and DTAB). This effect is due to stronger π–π interactions between the imidazolium rings on gemini surfactants with aromatic residues (Trp, Tyr, and Phe) of BSA, in addition to the electrostatic and hydrophobic interactions. Gospodarczyk et al.128 reported the stabilization of BSA by dicationic imidazolium (Gemini) surfactants, namely, [3, 3′[1,8(2,7-dioxooctane)]bis(1-dodecylimidazolium)chloride ([C12-oxyC4–C12im]Cl2) and [3, 3′[1,12(2,11-dioxadodecane)]bis(1-dodecylimidazolium)chloride ([C12-oxyC8–C12im]Cl2), at low concentration, followed by partial unfolding at higher concentration with the retrieval of unfolded structure post-CMC.128 Maurya et al.129 reported quenching of HSA fluorescence and a decrease in α-helical content, from 62.97% to 26.61%, in the concentration window of 0.99 × 10–5 to 13.0 × 10–5 M of a cationic imidazolium-based Gemini surfactant [C12-4-C12im]Br2 due to hydrophobic interactions.129 The IL [C12-4-C12im]Br2 was reported to act similarly toward lysozyme by quenching the fluorescence and decreasing the α-helix (from 34% to 29%) and β-sheet (from 28% to 9%).130
Comparing the SAIL-protein interaction mechanism with ionic surfactant-protein systems, the mechanism is not very dissimilar.224 The overall analysis of the mechanism of interaction requires several techniques.223,224 Two key techniques to identify these interaction regimes are isothermal titration calorimetry (ITC) and tensiometry (Figure 24a,b).
Figure 24.

SAIL-protein interactions in aqueous solution at various concentration regimes, specified as I (0 → C1), II (C1 → C2), III (C2 → C3), and IV (>C3). These regimes can be identified with various techniques such as (a) ITC curves indicating various interaction regimes and (b) surface tension vs concentration plots indicating various interaction regimes. (c) Graphical illustration of SAIL-protein structures in various interaction regimes. The terms are C1, critical aggregation concentration; C2, protein saturation concentration; and C3, critical micelle/vesicular concentration.
In ITC, the binding curve of SAIL to protein can be obtained by subtracting the enthalpogram of the SAIL titration to the buffer solution from the SAIL titration to the protein solution (Figure 24a). Additionally, the adsorption isotherm of SAIL in protein solution is generally different from the one from the buffer solution until C2. This is because proteins, being differentially charged macromolecules, also possess inherent surface-active properties. Due to this they also adsorb at the air–liquid interface, thus leading to lower surface tension as compared to that of pure water. When SAIL is added to the protein solution it interacts with proteins both at the air–liquid interface and in bulk solution, leading to the formation of different SAIL-protein monomers and SAIL-protein aggregate complexes until C2. However, when the protein gets saturated with SAIL, further addition of SAIL displaces the SAIL-protein complexes from the air–liquid interface into the bulk and starts adsorbing at the interface until the formation of free micelles or vesicles in the solution (Figure 24c). That is why the adsorption isotherm of SAIL in pure water and SAIL in protein solution beyond C2 match exactly (Figure 24b). The feasibility of SAIL-protein interaction can be calculated using the following equation:120,224
| 1 |
where ΔGoPS is the standard free energy of protein-surfactant interaction; ΔGob is the standard free energy of surfactant aggregation on polymer; ΔGoCMC is the standard free energy of aggregation of surfactant in the protein solution; R is the universal gas constant; T is the temperature in Kelvin; and XCAC and XCMC are the mole fractions of SAIL at the CAC and CMC in the protein solution, respectively. The number of surfactant molecules binding per protein can be calculated from the slope of protein concentration vs the surfactant CMC in protein solution using the following equation:120
| 2 |
where [S]CMC is the surfactant concentration at CMC, [S]Free is the free surfactant concentration, [P] is the concentration of protein under study, and N is the number of surfactant molecules attached to the protein.
These studies are important when considering interfacial and bulk complexation of SAILs with proteins to gain insights into their potential applications in food products. For example, Singh et al.131 reported interfacial complexation of [C4mim][C8OSO3] and [C8mim]Cl with a food protein—gelatin—at low concentration, whereas coacervation in the bulk solution was observed near the CMC. Interestingly, the [C8mim]Cl-gelatin coacervates remained stable up to a very high concentration beyond the IL CMC. Liu et al.132 investigated the detailed mechanism of the interaction of [C12mim]Br with the milk protein β-casein micelles (β-CM) in different concentration regimes. Below C1, the individual [C12mim]Br monomers bind to the β-CM shell close to the hydrophobic core to form a β-CM-[C12mim]Br (monomer) complex, further leading to a decrease in the environmental polarity of β-CM. Just over C1, [C12mim]Br molecules aggregate into micelle-like aggregates on the micellar shell, which led to the collapse of the N-terminal of β-casein and strengthened the hydrophobicity of the protein molecules, resulting in a more compact structure of β-CM. With a continuous increase in [C12mim]Br concentration, β-CMs are associated with each other in a network-like structure. Beyond C3, the net positive charges on the complexes, owing to the binding of more cationic surfactant molecules, lead to redissociation of the complexes, corresponding to the formation of the new nanosized β-CM-[C12mim]Br complexes. All the β-casein molecules are saturated by SAIL aggregates above Cs, and free SAIL micelle-like aggregates appear in the bulk phase above the CMC.132,133
Cao et al.134 investigated the interfacial behavior of the protein β-casein in the presence of [C16mim]Br and found that at high concentrations of surfactant the [C16mim]Br and β-casein coadsorb at the air/water interface. However, when compared with β-casein/DTAB mixture at the air/water interface, the β-casein/[C16mim]Br solutions have a lower interfacial activity, which results from the stronger attraction with aromatic rings through π–π interaction. Cao et al.135 also studied the effect of [C16mim]Br on the interfacial properties of various forms (normal N form as well as the fast F and aged A forms) of BSA at the decane/water interface. The addition of [C16mim]Br did not influence the structure of BSA below the isoelectric point; however, a significant influence on the dynamic interfacial properties of BSA was observed above the isoelectric point due to the electrostatic interactions established between BSA and [C16mim]Br.135 Haung et al.370 investigated the interfacial and bulk properties of pepsin (PEP) in the presence of [C16mim]Br, wherein the globular structure of pepsin was proposed to be the decisive factor controlling the nature of the interfacial film. The authors reported negligible change in the conformation of pepsin both at the interface and in the bulk phase at low IL concentrations (1 × 10–8 to 1 × 10–6 mol L–1), whereas preferential unfolding of pepsin was observed primarily at the decane-water interface at moderate IL concentrations (1 × 10–6 to 5 × 10–5 mol L–1).370 Mandal et al.136 reported the formation of lysozyme-[C4mim][C8OSO3] coacervates between C1 and C2 controlled by protein concentration, secondary structural alterations, and solution pH. Singh et al.371 reported enhanced antimicrobial activity of lysozyme upon complexation with SAILs in water, namely, [Cho][Sar] and [Cho][Doc]. While these works have given useful insights about SAIL-protein coacervation and have shown promise in terms of enhanced emulsification, due to the toxicity of some imidazolium-based ILs, reservations persist for their practical applications in food industries.
Beyond interactions and coacervation studies, SAILs have been also used to stabilize the protein in confined domains of microemulsions for kinetic packaging and interfacial catalysis.137−140,372−375 Debnath et al.372 reported preservation of the secondary structure of trypsin with 4-fold higher activity in oil in water (o/w) microemulsion of [C2mim]Br confined in the water pool, using CTAB as an emulsifier. Pavlidis et al.138 reported a 4.4-fold rise in the activity and a 25-fold increase in the shelf life of lipase due to the adoption of a more rigid form of the protein in the confined domain of the water in IL ([C4mim][PF6] and [C4mim][BF4]) microemulsion, emulsified with Tween 80.138 Kundu et al.373 reported the stabilization of BSA in the confined water domain of water in oil microemulsion comprising a SAIL, i.e., [ProC3][LS], as the emulsifier and cyclohexane as the dispersion medium.373 Mao et al.140 selectively isolated hemoglobin at pH 5.0 in a water in oil microemulsion comprising [C10mim]Br as the emulsifier and [C4mim][PF6] as the dispersion medium. The isolated hemoglobin could be back-extracted with 55.6% efficiency in Britton-Robinson buffer at pH 12.0. Kaur et al.374 reported a high thermal stability of lysozyme, up to 120 °C, at the nanointerfaces of ethylene glycol in the [C2mim][NTf2] microemulsion with a dialkylimidazolium-based SAIL as emulsifier.374
Few other works have reported positive results on the encapsulation of proteins in SAILs micelles, their fibril inhibition, and refolding ability. For example, Bento et al.375 reported enhanced degradation of the dye indigo carmine B by laccase encapsulated in aqueous micelles of [N10111]Br and [C10mim]Cl. Alves et al.376 reported the stable encapsulation of lysozyme in micelles of a fluorinated anion-based SAIL, [C1mim][C4F9SO3], for protein drug delivery. Kundu et al.377 demonstrated the chain length effect in decreasing order of BSA fibril inhibition tendency of SAILs, as follows: [C16mim]Cl > [C12mim]Cl > [C8mim]Cl. Singh et al.378 reported higher refolding and stabilization of molten globule state of urea/GndHCl and alkali denatured Cyt c by [C10mim]Cl compared to [C8mim]Cl.378
3.4.1. IL Surfactant-Protein vs Ionic Surfactant-Protein Systems
The comparative analysis of IL surfactant-protein vs ionic surfactant-protein systems is depicted in Figure 25. The available SAIL-protein systems, although few to date, are superior in most cases when compared to the conventional surfactant-protein systems. For example, the SAILs are generally less denaturing toward proteins when compared to the conventional ionic surfactants.118−130 This behavior is due to the presence of a large organic cation in the IL which counters the direct interactions of the long-chain anion with proteins. Furthermore, ILs increase the aggregation tendency at low concentrations and decrease the denaturing monomeric binding region with the protein.118−130
Figure 25.

Comparative analysis of IL surfactant-protein vs ionic surfactant-protein systems.
Due to the low interfacial tension of SAIL-protein complexes, and protein stabilization in the monomeric regime, these systems have practical implications for practical in vitro stabilization of TPs during transportation. Moreover, they form stable colloidal complexes with food-grade proteins, like gelatin131 and β-casein132−134 at much lower concentrations; however, the possible toxicity of imidazolium-based SAILs used in such studies preclude their applications in food industries in the current scenario. The high emulsifying tendency of SAILs has been exploited to isolate, store, and enhance the activity of proteins in the confined domains or nanointerfaces by formulating them into microemulsions.140,372−374 Bharmoria et al.149 reported the thermal stabilization of cytochrome c in vesicular assemblies of the SAIL [Cho][AOT] in [C2mim][C2OSO3], up to 180 °C. Such assemblies could be useful in the stable packaging of TPs and other proteins translated to confined domains of microemulsions. Considering the stability of proteins observed in the quaternary ammonium family of ILs,86,110 these proteins can be confined in the polar phase of IL microemulsions for long-term packaging along with TPs. For example, Kaur et al.374 showed the stabilization of lysozyme at 120 °C in the confined nanointerface of the microemulsion containing ethylene glycol as the confined polar phase, dialkylimidazolium-based SAIL as an emulsifier, and [C2mim][Tf2N] as the nonpolar phase. These results lead to future possibilities for long-term functional packaging of proteins in all IL-based solvent media. SAILs can be a potential competitor to ionic surfactants in detergent industries due to their superior surface activity; however, efforts are needed to address their toxicity issues in addition to the stability of laundry enzymes (lipase, protease, and cellulases) in the formulation.
3.5. Proteins in Ionic-Liquid-Based Aqueous Biphasic Systems
The processing/extraction of pure proteins from aqueous media involves, most of the time, the use of highly viscous hydrophobic (nonwater-soluble) ILs with a low protein-friendly character. To overcome these concerns, hydrophilic ILs have been investigated in liquid–liquid extraction in the form of aqueous biphasic systems (ABSs). ABSs are ternary systems composed of water and two other phase-forming components, in which above a certain concentration occurs phase separation. Each phase is aqueous and enriched in one of the other remaining components. Conventional ABSs are formed by polymer–polymer, polymer–salt, or salt–salt combinations.379 In 2003, Rogers and co-workers380 demonstrated the potential of ILs to prepare ABSs by mixing them with inorganic salts. Thereafter, several studies available in the literature clarified that IL-based ABSs can be generated with a wide variety of organic/inorganic salts, polymers, amino acids, and carbohydrates, which have been reviewed in detail, including applications with proteins, in our earlier reviews.17,381
More recently a large number of works published to date have focused on the extraction performance of ABSs for proteins, with few addressing however the protein’s stability and mechanisms of interaction with ILs. Although we could think on the IL co-solvent effect, as discussed before, ABSs are more intricate since a third component is also present at the IL-rich phase. An illustration of steps carried out for the extraction and purification of proteins using IL-ABSs is shown in Figure 26.
Figure 26.

Extraction and purification of proteins using IL-based ABSs.
Due to their natural affinity for water, proteins are expected to partition into the aqueous phase of IL-ABS, but both phases are water-rich. Therefore, there is not a universal phenomenon dictating protein partitioning; still, in ABSs comprising ILs most proteins tend to partition to the IL-rich phase, a phenomenon that is independent of having or not a strong salting-out species as the third phase-forming component. For example, Pereira et al.378 reported the single-step extraction of BSA into the IL-rich phase of ABS composed of phosphonium- and quaternary ammonium-based ILs combined with potassium citrate/citric acid at pH 7.0. The authors used different ILs ([P4444]Br, [P4444]Cl, [Pi(444)1][Tos], [P4441][C1OSO3], and [N4444]Cl) for the separation of BSA. SE-HPLC and FT-IR studies confirmed the stability of BSA extracted into the IL-rich phase.
Furthermore, BSA was recovered from the IL-rich phase via dialysis, followed by the IL reuse. On the other hand, Quental et al.157 reported 100% extraction efficiency of a stable BSA from the fetal bovine serum in the IL-rich phase of ABSs formed by cholinium-based ILs ([Cho][OAc], [Cho][Bit], and [Cho][Prop]) and polypropylene glycol with a molecular weight of 400 g·mol–1 (PPG 400). Both works382,157 show that proteins prefer the IL-rich phase, independent of having a second phase enriched in a salt or in a polymer. Therefore, protein’s portioning in IL-based ABSs is majorly governed by specific interactions occurring with the ILs instead of being ruled by a salting-out phenomenon.
Taha et al.383 explored the use of ABSs comprising cholinium-based ILs with Good’s buffer anions for the extraction of BSA. The advantages of the studied ILs are their self-buffering properties in the biological pH range and their easily tunable polarity and hydrophobicity. Complete extraction of BSA was achieved into the IL-rich phase, without compromising the structural stability of BSA as confirmed from CD and FT-IR analysis. The authors also compared the stability of BSA in aqueous solutions of the ILs with the respective Good’s buffer precursors, a more conventional IL ([Cho]Cl), and a well-known protein stabilizer (sucrose). The α-helicity of BSA was shown to follow the order [Cho][TES] > [Cho][Tricine] > [Cho][HEPES] > sucrose > TES > [Cho][Cl] > HEPES > Tricine. These results show that the studied ILs are better stabilizers of BSA, which is due to the dual hydrogen bonding established between the IL ions and BSA as revealed from molecular docking studies. Gupta et al.384 also reported 100% extraction efficiency of α-chymotrypsin using IL-ABSs comprising Good’s buffer ILs. They concluded that the studied ILs provide a better stabilizing effect toward α-chymotrypsin, which was confirmed from tryptophan fluorescence studies. The same type of ILs was used for the extraction of TPs, namely, immunoglobulin Y (IgY) from egg yolk, with extraction efficiencies ranging from 79 to 94%.385 Ramalho et al.386 reported the single-step extraction of IgG from rabbit serum using IL-ABS, as confirmed by SE-HPLC.386 Mondal et al.387 also reported the single-step extraction of IgG from rabbit serum into the IL-rich phase using ABSs formed by cholinium-based ILs, with 85% extraction efficiency. The structural integrity of the extracted protein was confirmed by SE-HPLC, SDS-PAGE, and FT-IR. The purity of the extracted IgG was enhanced by 58% when compared with its purity in serum samples.387
Beyond molecular interactions and salting-out effects, other factors like molecular recognition and temperature have been reported to affect the stability of proteins in IL-ABSs. For example, Tseng et al.388 explored the importance of biomolecular recognition for the extraction of arginine and lysine-based peptides and proteins from the aqueous phase to crowned 1,2,3-triazolium cation and [NTf2]-based IL phase of IL-ABS. Both peptides and proteins retained their structural stability upon extraction into the IL-rich phase, as confirmed from NMR and CD analysis.388 Ikeda et al.389 investigated the effect of the lower critical solution temperature of the IL [P4,4,4,4][TMBS] for the partitioning of Cyt c. The authors found the selective partitioning of the oxidized and reduced form of Cytochrome c into the IL or buffer phase. The oxidized cytochrome c gets transferred to the [P4,4,4,4][TMBS] phase, whereas reduced Cyt c remained in the buffer phase in the functionally active forms (Figure 27a).
Figure 27.

Illustration of the use of thermo-reversible IL-based systems for the separation of proteins: (a) 50% [P4,4,4,4][TMBS] + 50% H2O and (b) 6% [N11[2(N11)]0][C1CO2]+ 30 PPG 400 + 64% H2O.
Passos et al.158 also investigated the low critical solution temperature behavior of IL-ABS comprising protic ILs and PPG 400 for the separation of proteins, namely, Cytochrome c and azocasein. Both proteins maintained their native structure in the IL-rich phase at 45 °C, as confirmed from FT-IR results (Figure 27b). The described temperature-sensitive IL-ABSs should be explored at the practical scale for the extraction and purification of high-value proteins, especially TPs such as immunoglobulins and other recombinant proteins. Additionally, challenges associated with the economic viability and the stable extraction with high purity from the real protein matrices must be resolved, aiming at establishing sustainable IL-ABSs for protein purification and stabilization.390
3.6. Poly(ionic liquid)-Protein Conjugates
PEGylation is extensively used to shield proteins from enzymatic degradation, improving their long-term storage and pharmacokinetic efficiency.153 However, sometimes the success of such protein–polymer conjugations is largely constrained, namely by (i) low protein conjugation efficiency; (ii) restricted choice of solvents, such as water, methanol, and DMSO; (iii) laborious and time-consuming polymerization, workup, and storage processes; (iv) denaturation of proteins during harsh conditions applied for polymer conjugation; and (v) competitive hydrolysis of polymer side-end chain having amide functionalities.153 Taking these issues altogether, ILs can be employed as potential alternatives for the synthesis of protein/peptide–polymer conjugates and as media for packaging proteins. Table 2 summarizes the reported works in both fields. These works can be divided into three categories: (1) ILs or A-ILCSs as medium for stable protein-polymer conjugation; (2) stable protein-IL polymer conjugates; and (3) ILs as medium for solubilization and thermal stability of protein-polymer conjugates. An illustration of these systems is shown in Figure 28.
Table 2. Summary of the IL-Based Protein/Peptide Polymer Conjugates.
| P–P Conjugate | Ionic Liquid as Solvent/Conjugate |
|---|---|
| Lysozyme-polyMPC153 | [C2mim][TfO] |
| BM2391 | [C2mim][OAc] |
| Imidazolium salt-peptide conjugates392 | [(CO2H)4C4C1im]Br– and [(CO2H)15C15C1im]Br– |
| Cytochrome P450-PEO389 | [C2mim][Tf2N] |
| Lipase A-PAcMO394 | [C4mim][PF6] |
| Myoglobin-glycolic acid ethoxylate lauryl ether150 | [bmpy][TfO] and [bmpy][Tf2N] |
| [C-IgG][S] liquid ionic conjugate395 | [C-IgG][S] |
Figure 28.

Strategies of IL-based protein–polymer conjugates: (a) ILs or A-ILCS as medium for stable protein–polymer conjugation; (b) ILs as medium for solubilization and stability of protein–polymer conjugates; and (3) stable protein-IL polymer conjugates.
Chen et al.153 reported one-pot synthesis of lysozyme-poly(methacryloyloxyethyl phosphorylcholine) (polyMPC) conjugate in [C2mim][TfO] with sodium borate buffer at pH 9.0. The authors observed less than 5% of aggregation of the conjugate from SEC-HPLC analysis, thus demonstrating the significant stability of the prepared conjugate. Baumruck et al.391 reported the native chemical ligation of highly hydrophobic membrane-associated peptides in [C2mim][OAc]/buffer mixture at pH 7–7.5, with 80–95% yield, outperforming the existing gold standard ligation method. The secondary structural integrity of these peptides was confirmed from CD spectroscopy.391 Reinhardt et al.392 reported a structurally stable imidazolium salt-antimicrobial peptide conjugate showing enhanced antimicrobial activity compared to the nonconjugated peptide.392 Ohno and co-workers393 showed solubility and enhanced thermal stability up to 120 °C of a chemically modified Cytochrome P450 with poly(ethylene oxide) in [C2mim][Tf2N].393 Chado et al.394 reported 19-fold enhancement in the activity of covalent functionalized lipase A with polymer poly(4-acryloylmorpholine) (PAcMO) dissolved in [C4mim][PF6].394 Brogan et al.150 conjugated myoglobin with surfactant glycolic acid ethoxylate lauryl ether, leading to 84 surfactant molecules per protein. The PP-conjugate showed significant solubility in dry ILs, with the retention of the secondary structure up to 55 °C.150 Overall, this is an innovative approach to overcome the limitations of the robustness of proteins to get solubilized in dry ILs and should be implemented for thermal stabilization of other proteins and TPs. In this direction, Slocik et al.395 investigated the polycationic nature of modified antibodies and their ability to form ion pairs for the conversion of primary Immunoglobulin G antibodies into stable protein liquids that retained more than 60% binding activity after repeated heating up to 125 °C. The IgG cantonized with 1-ethyl-3-(3-dimethyl aminopropyl) carbodiimide (C-IgG) was paired with Poly(ethylene glycol) 4-nonyl phenyl 3-sulfopropyl ether anion (S) to form a room-temperature protein ionic liquid (P-IL).395 This is an important advancement on grounds of the poor temperature-dependent stability of therapeutic proteins for long-term storage and must be more deeply investigated. Using atomistic MD simulations, Balasubramanian et al.396 reported that the polymer surfactant functionalized lipase A, lysozyme, and myoglobin dissolved in [C2mim][NTf2] exhibits higher intraprotein hydrogen bonding, in addition to the greater thermal stability and IL tolerance due to the screening of protein-IL interactions.396
IL-based protein-polymer conjugates are mainly comprised of imidazolium-based ILs combined with chloride, bromide, acetate, Dicyandiamide, and tetrafluoroborate anions. Very few studies emphasized the potential of biocompatible and biodegradable cholinium-based ILs as prospective alternatives to imidazolium ones. Nowadays, there is pervasive availability of more benign ILs, such as those composed of cholinium-based cations combined with anions derived from carboxylic acids, biological buffers, among others, which should be explored for the synthesis of therapeutic protein-polymer conjugates and application in the biopharmaceutical field.
4. Conclusions and Future Prospects
Herein, we have comprehensively reviewed the works published to date on IL-protein systems, excluding however works dealing with biocatalysis. These systems comprise proteins dissolved in (i) neat ILs; (ii) ILs as co-solvents; (iii) ILs as adjuvants; (iv) ILs as surfactants; (v) ILs as phase-forming components of aqueous biphasic systems; and (vi) IL-polymer-protein/peptide conjugates. The reviewed works allowed us to make inferences regarding the main molecular mechanisms and IL-protein interactions affecting the stability/conformational alteration/unfolding/misfolding/refolding of proteins.
The expectancy of high-temperature biocatalysis has been the driving force in studies involving enzymes and NILs. However, few NILs have successfully cracked the hard nut of solubilizing proteins of different secondary structural conformations such as BSA, HSA, α-chymotrypsin, ovalbumin, myoglobin, lactoferrin, silk fibroin, zein, and keratin. The formation or disruption of hydrogen bonding between IL ions and proteins has been found to be the major force for the stable solubilization of the investigated proteins. Therefore, from a protein packaging perspective, ILs possessing H-bonding ability in both the cation and anion, such as [CnOHmim][HOCnSO3], have been identified as the most efficient. Few other ILs, such as [C2mim][C2OSO3] and [C4mim][OAc], have been found to solubilize proteins of all kinds of secondary structures. This possibility is due to their nanoheterogeneous structure and should be taken as standard when designing new NILs for protein solubilization. Although the published results to date are promising and optimistic, the practical feasibility of such systems for protein packaging is rather gloomy. This is due to various factors, namely, (i) lack of systematic research in one IL or protein direction (too many scattered ILs and proteins have been investigated), (ii) biocompatibility issues of promising ILs reported, and (iii) lack of processes for suitable re-extraction of proteins after solubilization in NILs.
On the co-solvent front, results reported for protein stabilization are more promising than in NILs. The paradigm of introducing water or an aqueous medium as the environment around proteins is indeed an advantage from several points of view. The stability of proteins has been explained by hydrophobic solvation, H-bonding, and preferential exclusion phenomena. The most promising IL-water systems identified are the ones containing quaternary ammonium cations, which have shown stabilization (thermal, structural, and functional) toward several proteins, operating via either preferential exclusion or H-bonding mechanisms. However, the stabilization effect in most cases depends on the concentration of IL and type of IL ions. Unlike with the fewer works available on NIL-protein systems, concerted efforts have been made by various research groups in A-ILCS-protein systems. A-ILCSs of the quaternary ammonium family have been particularly investigated for protein stability, refolding, and cryoprotection. [Cho][dhp] has been identified as the best candidate to stabilize Cytochrome c for long periods, providing evidence on the use of A-ILCS of [Cho][dhp] for protein packaging purposes. The [Cho][dhp] was also investigated for the stabilization of relevant TPs, namely, Interleukin 2 and IgG. The stabilization of other TPs, such as insulin, against thermal and aggregation-induced denaturation was demonstrated with the use of A-ILCS comprising [(C2)3NH][dhp]. Concerning imidazolium-based ILs, some of them have shown protein stabilization via hydrophobic solvation and clustering of the imidazolium cation at the protein surface, both via electrostatic and hydrophobic interactions. However, their precipitation effect on proteins came as blessings in disguise, followed by their use to develop strategies for the crystallization of proteins.
As far as IL adjuvants are concerned, valuable information regarding molecular-level interactions with proteins has been reported from both experimental and computational studies. Intrinsic fluorescence of BSA by Trp residue was used to locate the binding regions based on fluorescence quenching. However, the structural alterations in proteins were mainly documented from secondary structural behavior, assessed by circular dichroism and FT-IR. All kinds of noncovalent interactions (electrostatic, H-bonding, hydrophobic, and van der Waals) were used to support the stabilization/destabilization/conformational transition effects depending upon the cation and anions of ILs used. The Hofmeister effect based on preferential binding or exclusion phenomena has been applied to explain the IL-induced effect on proteins, however with large inconsistency and with rare consensus among different research groups. The understanding of the molecular mechanism of IL-protein interactions using spectroscopic techniques allowed the report of a regression-based model that produces satisfactory results to describe the relationship between the inhibitory ability, hydrophobicity, and H-bonding ability of ILs toward proteins. This type of model could be useful and can limit the analysis time when investigating the interactions of ILs with proteins. Moreover, the application of ILs as adjuvants to resolve bands in SDS-PAGE is another promising application reported, having relevant implications for biochemists and biotechnologists, and must be exercised with other ILs seeking further improvements in resolution.
In the case of surfactant ILs, the most important advantage over conventional ionic surfactants is that they are most of the time observed with higher surface activity, with reduced CAC. This behavior is useful for application in food industries and cleaning purposes. The mechanism of interactions is not very dissimilar to conventional surfactant-protein systems. On a positive note, their denaturing tendency toward proteins has been found to be lesser than their conventional counterparts. This behavior can be attributed to the presence of larger organic cations. Moreover, SAIL-protein combinations lead to lower interfacial tensions and stabilize the protein in a monomeric regime, which is important for TP stabilization and from a toxicity point of view. One of the key advantages that SAILs further exhibit is their thermal stability, which could be utilized to store proteins in assemblies like microemulsions. In this field, the works reporting the isolation of hemoglobin in the polar part of the water in IL microemulsion and thermal stabilization of cytochrome c in vesicular assemblies up to 180 °C hold high relevance. The key limitation of SAILs is that most of them belong to the imidazolium cation family that has been raising toxicity and biocompatibility issues.
As far as IL-ABS-protein and IL-protein-polymer conjugates are concerned, they are promising systems with wide implications. IL-ABSs are particularly useful on account of providing a water-rich medium during extraction or purification steps. However, the stability of the protein has been poorly addressed in IL-ABSs after the extraction step. Promising results in this direction are works on the use of IL-ABSs to purify high-value proteins, such as biopharmaceuticals, and the use of stimuli-responsive IL-ABSs that provide technical advantages. IL-protein-polymer conjugates have been scarcely investigated. Yet, promising results have been published on their use toward the stable packaging of proteins at room or high temperature. In the available works, proteins were functionalized with polymers to be either soluble in ILs or to behave as an IL by themselves.
Since the inception of the IL-protein journey, significant contributions have been made by several authors, on both the fundamental and applied fronts. The field has moved beyond enzyme biocatalysis, and relevant and high-value proteins, such as TPs, have been considered with ILs. All contributions have certainly shown the relevance of ILs in the field of proteins, but it is still in its infancy concerning mature and concrete technological applications. On the optimistic front, the promising contributions available do support the IL potential in the field of proteins, and yes, it can be stated that “Ionic liquids, if properly designed, do exhibit suitable characteristics to dissolve, extract, stabilize and purify proteins”. The IL-protein interactions with different forms of ILs and applications are summarized in Figure 29. The list of promising systems having both fundamental and applicative implications produced to date is summarized in Table 3.
Figure 29.

Summary of IL-protein interactions wherein (1) ILs as adjuvants in most cases pull out water from the protein surface due to favorable interactions with the protein surface; (2) ILs as co-solvents co-cluster in the protein hydration layer; and (3) ILs as native solvents solvate the protein.
Table 3. List of Most Promising Protein-IL Systems Using ILs in Various Forms.
| Protein | Ionic Liquids | Category | Goal |
|---|---|---|---|
| CAL B29 | [Me(OEt)3-Et3N][OAc], [Me(OEt)3-Et-Im][OAc] | NIL | Stable Packaging |
| Cytochrome c31,32 | [C2mim][C2OSO3], [Amim]Cl | NIL | Stable Packaging |
| CALB275 | [C2OHmim][Tf2N], [C3OHmim][Tf2N] [C3OHTEA][HOC2SO3] | NIL | Stable packaging |
| Insulin282 | [Cho][gerenate] | NIL | Stable packaging |
| Monellin59 | [C4mpy][Tf2N] | A-ILCS | Stable packaging |
| Cytochrome61,63,66,296 | [Cho][dhp] | A-ILCS | Stable packaging |
| Interleukin 2, IgG299,300 | [Cho][dhp] | A-ILCS | Stable packaging |
| α-chymotrypsin68,291 | [(C2H5)3NH][OAc], [(C2H5)3NH][PO4] | A-ILCS | Stable packaging |
| Insulin301 | [(C2)3NH][dhp], | A-ILCS | Aggregation suppression |
| Lysozyme312 | [CH3CH2NH3][NO3] | A-ILCS | Stable packaging |
| Lysozyme333 | [C4mim]Cl | A-ILCS | Crystallization |
| Lysozyme331 | [OHC2NH3][HCOO], [(C1)2OHC2NH][OHC2COO] | A-ILA | Crystallization |
| BSA, β-lactoglobulin (IgG)363 | [C2mim]Cl, [C4mim]Cl, [C6mim]Cl, [C8mim]Cl | A-ILA | Protein fibril dissolution |
| BSA, ovalbumin, α-lactalbumin366 | [C4Pyr]Br, [C8Pyr]Br, [C11Pyr]Br | A-ILA | Protein’s separation |
| Insulin317 | [PAN][NO3] | A-ILA | Aggregation suppression |
| pepsin and papain354 | [NH2C2C4im]Br | A-ILA | Activity enhancer |
| Trypsin372 | [C2min]Br/CTAB | IL-ME | Activity enhancer |
| Lipase138 | [C4mim][PF6], [C4mim][BF4] | IL-ME | Activity enhancer |
| Hemoglobin140 | [C10mim][Br]+[C4mim][PF6] | IL-ME | Stable extraction |
| Cellulase121,368 | [C8mim][C12OSO3], [Cho][DBS] | SAIL | Detergency |
| Cytochrome c149 | [Cho][AOT]+[C2mim][C2OSO3] | SAIL | Stable packaging |
| BSA123 | [C12Amim]Cl, [C12Emim]Cl | SAIL | Protein fibril dissolution |
| BSA383 | [Cho][TES], [Cho][Tricine], [Cho][HEPES], | IL-ABS | Stable extraction |
| IgY383 | [Cho][MES], [Cho][Tricine], [Cho][CHES], [Cho][HEPES], [Cho][TES] | IL-ABS | Stable extraction |
| Cytochrome c and Azocasein158 | [N11[2(N11)]0][C1CO2] | IL-ABS | Stable extraction |
| IgG387 | [Cho][Asc] | IL-ABS | Stable extraction |
| Myoglobin150 | [bmpy][NTf2], [bmpy][OTf] | P–P–Conjugate | Stable Packaging |
| IgG395 | [C-IgG][S] | P-IL | Stable Packaging |
Apart from the discussed possibilities and based on the listed promising systems making use of ILs, IL-protein systems should be investigated for other promising applications. Some identified opportunities are summarized in Figure 30. For thermodynamic packaging of proteins or TPs in dry ILs, the strategy of Brogan et al.150 is very useful. Its combination with thermoreversible IL-ABS developed by Passos et al.158 would provide a suitable pathway for protein re-extraction. The proposed process can avoid the power consumed during protein storage at low temperatures.
Figure 30.

Future prospects of IL-protein systems. (a) Thermodynamic storage and extraction of proteins. (b) Air stable light-harvesting via fluorescent ILs embedded in the protein matrix. (c) Utilization of IL-protein chemistry to prepare fibrils from ECM and further use to create cytocompatible hydrogels for 3D-cell growth. (d) Anticancer membrane peptide–ionic liquids for cancer treatment.
The tuning nature of ILs can further be investigated to introduce fluorescence. The solubilization of fluorescent ILs in the matrix of structural proteins could provide a suitable platform for air-stable exciton energy transfer, like the one observed in the thylakoid membrane during photosynthesis. ILs have been used earlier in light-harvesting by photon upconversion via the triplet–triplet annihilation process, which can give direction for the design of a suitable system.397−399 The hybrid protein hydrogels prepared using protein fibrils of extracellular matrices should be considered as promising cytocompatible 3D platforms for cell culture. Generally, the synthetic acrylate hydrogels used for 3D cell growth have been found to be toxic;400 therefore, developing hydrogels from biological raw material could overcome such issues. Most studies on IL-protein systems have been carried out on single IL-single protein systems, which is not the case for biological systems where proteins are never found alone in the cell, except chlorella. Therefore, future studies, if targeted at the protein of a specific location of the cell, must be done in the presence of other proteins of that cell, i.e., protein crowded medium for biomimicking biological fluids.401−403 This would reveal the actual effect of IL-protein and protein-protein interactions when we think of applying them for biochemistry, cell biology, and biotechnology purposes. Furthermore, when studying the effect of ILs and proteins (individual or combined), the effect of the IL cation and anion and their use at different ratios should also be investigated to see the actual relevance of using them in the IL form. We think that these protocols would foster the field with high scientific credibility rather than notoriety.
The biomembrane plays an important role in the signaling growth activities in cells. However, a mutation in the gene responsible for cell growth can result in amplification of cell growth signals due to fast heterodimerization of epidermal growth factor receptors (EGFR), which can cause enhanced cell growth and lead to cancer.404−406 This trend can be controlled by stopping the growth signal through transfection of the cell with a competitive synthetic EGFR membrane-active peptide (EGFR-MAB) for fake heterodimerization. Here, membrane peptide labeled surface-active ILs can play a key role because their self-assembled structures can alter the cancer cell membrane either via altering the membrane structure through hydrophobic interactions or by forming fake heterodimers with membrane peptides to stop the cell signaling for malignant growth. Fundamental studies in this direction can lead to significant advances in the treatment of cancer.
Although not considered in this review, the use of ILs to allow the stable immobilization of enzymes on nanomaterials is an emerging field.407−409 Enzyme immobilization on nanoscale materials offers valuable advantages in enhancing enzyme stability during challenging reaction conditions. However, current immobilization methods suffer from a significant drawback: irreversible damage caused by the intense interaction between the enzyme and the carrier. To address this issue, the use of ILs becomes crucial as they promote softer interactions, reducing the damage caused during the immobilization process in a solid matrix. By combining enzyme surface modification through nanomaterials and solvent manipulation using ILs, an environmentally friendly approach can be created in the form of protein nanoconstructs for resilient biocatalysis.
Acknowledgments
This work was developed within the scope of the project CICECO-Aveiro Institute of Materials, UIDB/50011/2020 (DOI 10.54499/UIDB/50011/2020), UIDP/50011/2020 (DOI 10.54499/UIDP/50011/2020), and LA/P/0006/2020 (DOI 10.54499/LA/P/0006/2020), financed by national funds through the FCT/MCTES (PIDDAC). This work was funded by the PRR - Recovery and Resilience Plan and by the NextGenerationEU funds at Universidade de Aveiro, through the scope of the Agenda for Business Innovation “InsectERA” (Project no. 20 with the application C644917393-00000032). P.B. acknowledges La-Caixa junior research leadership-post doctoral program (ID: 100010434, fellowship code: LCF/BQ/P122/11910023); A.A.T. acknowledges the Knut and Alice Wallenberg Foundation via the Wallenberg Centre for Molecular and Translational Medicine (AT), Swedish Research Council (Project no. 2020-04299) and Cancerfonden, Sweden (Project no. 222409Pj); D.M. acknowledges DST-SERB, India (EEQ/2021/000059), and NANOPLANT project, which received funding from the European Union’s Horizon 2020 research and innovation program under grant agreement no. 856961. T.S.K. acknowledges DST-SERB, India (CRG/2021/005897), and A.K. acknowledges CSIR-CSMCRI under in-house Project MLP0029 for financial supports. Authors acknowledge Ana Sofia Marques from CICECO-Aveiro Institute of Materials for assistance during drawing of some figures.
Glossary
Abbreviations
- IL
Ionic Liquid
- TPs
Therapeutic proteins
- FDA
Food and Drug Administration
- NILs
Neat ionic liquids
- A-ILCS
Aqueous concentrated-IL solution
- A-ILAS
Aqueous adjuvant-IL solution
- SAIL
Surface active ionic liquid
- IL-ME
Ionic liquid-microemulsion
- IL-ABS
IL-aqueous biphasic system
- P-P-Conjugate
Poly(ionic liquid)-protein conjugate
- P-IL
Protein-ionic liquid
- CAL B
Candida antarctica lipase B
- BSA
Bovine serum albumin
- HSA
Human serum albumin
- CRL
Candida rugosa lipase
- [Cnmim]
Alkyl imidazolium
- [CnOHmim]
Hydroxy alkylimidazolium
- [CnPy]
Alkyl pyrolidinium
- [CnPr]
Alkylpyridinium
- [CnP]
Alkyl phosphonium
- [Cho]
Choline
- [CnN]
Tetraalkyl ammonium
- [CnNH3]
Alkyl ammonium
- [DEA]
Diethanolammonium
- [BAGUA]
Butylpentamethylguanidinium
- [BCGUA]
Butyltrimethylguanidinium
- [MCGUA]
Tetramethylguanidinium
- [DCGUA]
Decyltrimethylguanidinium
- [Gdn]
Guanidinium
- [Mor]
Morpholinium
- [TMG]
Tetramethylguanidinium
- [C3OHTEA]
Hydroxypropyl triethylammonium
- [DMEA]
N-N-dimethylethanolammonium
- [OAc]
Acetate
- [HCOO]
Formate
- [Tf2N]
Bis(trifluoromethylsulfonyl)imide
- [Sar]
Sarcosinate
- [Doc]
Deoxycholate
- [TGA]
Thioglycolate
- [Pn]
Propionate
- [CnOPO3]
Alkylphosphate
- [SCN]
Thiocynate
- [HSO4]
Hydrogen sulfate
- [MDEGSO4]
2(2-Methoxyethoxy)ethylsulfate
- [Gly]
Glycolate
- [Lac]
Lactate
- [OH]
Hydroxy
- [BF4]
Tetrafluoroborate
- [Bit]
Bitartarate
- [DHCit]
Dihydrogencitrate
- [Prop]
Propionate
- [But]
Butyrate
- [dca]
Dicyanamide
- [dhp]
Dihydrogen phosphate
- [NO3]
Nitrate
- [PF6]
Hexafluorophosphate
- [CnOSO3]
Alkyl sulfate
- [TfO]
Trifluoromethanesulfonate
- [Asc]
L-Ascorbate
- I
Iodide
- Cl
Chloride
- Br
Bromide
- [DBS]
Dodecylbenzenesulfonate
- SDS
Sodium dodecyl sulfate
- CTAB
Cetyltriethylammonium bromide
- HEPES
2-[4-(2-Hydroxyethyl) piperazin-1-yl]ethanesulfonic acid
- TES
2-[(2-Hydroxy-1,1-bis(hydroxymethyl) ethyl) amino] ethanesulfonic acid
- Tricene
N-Tris(hydroxymethyl)methyl] glycine
- CHES
2-(cyclohexylamino)ethanesulfonic acid
- MES
2-(N-Morpholino) ethanesulfonic acid
- PEO
Poly(ethylene oxide)
- PEG
Polyethylene glycol
Biographies
Pankaj Bharmoria, La-Caixa Junior Research Leader at Institute of Materials Science of Barcelona (ICMAB-CSIC), Barcelona, Spain. He obtained his Ph.D. degree in biophysical chemistry from AcSIR-Central Salt and Marine Chemicals Research Institute, Bhavnagar, India, under the supervision of Dr. Arvind Kumar. Since 2016, he has been conducting postdoctoral research across Portugal (CICECO-Aveiro University), Japan (Kyushu University), Sweden (Chalmers University of Technology & Gothenburg University), and Spain (ICMAB-CSIC) with independent grants like JSPS, MSCA-IF, and La-Caixa. His current research is focused on exploiting triplet state photochemistry for photon upconversion and photopharmacology, and ionic liquid-protein chemistry for targeted cancer treatment.
Alesia A. Tietze, Associate Professor in Medicinal Chemistry at the Department of Chemistry and Molecular Biology at the University of Göteborg. Her research team is a part of the Wallenberg Centre for Molecular and Translational Medicine (WCMTM) and Centre for Antibiotic Resistance Research (CARe). Her research focuses on bioinspired chemical engineering of peptides and peptidomimetics towards diagnosis and drug delivery. Main areas of research include the development of bioinspired nanopores for the detection of biomarkers in body fluids, targeted treatment of cancer, and discovery of nature-inspired antimicrobial and antivirulence peptides.
Dibyendu Mondal is the ERA chair holder for the NANOPLANT project at IPG PAS in Poznan, Poland. He earned his Ph.D. in Chemical Science from AcSIR-CSMCRI, India, in 2015, followed by a postdoctoral position at CICECO, University of Aveiro, Portugal, from 2015 to 2017. In 2017, he became an assistant professor at CNMS, JAIN (Deemed-to-be University), India. His primary research interests include the value addition of bioresources with neoteric solvents, protein engineering and biocatalysis, bioinspired and active nanoconstructs, and nanotechnology for sustainable agriculture.
Tejwant Singh Kang, Associate professor in physical chemistry at the Department of Chemistry, Guru Nanak Dev University, Amritsar, Punjab, India. He received his Ph.D. in 2011 from the Central Salt & Marine Chemicals Research Institute, Bhavnagar, India. In 2011, he worked as a JSPS postdoctoral researcher at Kyushu University, Japan in collaboration with Professor Nobuo Kimizuka. He has been a visiting researcher at Tohoku University and Kyushu University, Japan. His research focuses on developing various ionic liquid-based systems for their use in photocatalysis, enzyme stabilization, and biomass valorization. He has contributed to about 95 research papers in journals of international repute.
Arvind Kumar is the Chief Scientist and Honorary Professor at AcSIR, CSIR-Central Salt and Marine Chemicals Research Institute, Bhavnagar, India, where he heads a division working on salt and marine chemicals. He did a Ph.D. in Chemistry at Kurukshetra University. He has also worked at the University of Karlsruhe, Germany, under a DAAD Fellowship, and the Centre for Green Chemistry, Alabama University, Alabama, USA, under a CSIR-Raman Research Fellowship. His research interests cover design and development of greener solvents such as ionic liquids and deep eutectic solvents and their use in biomass processing and materials development for sustainable technologies.
Mara G. Freire holds the position of Coordinating Researcher at CICECO-Aveiro Institute of Materials, University of Aveiro, Portugal. She is the Coordinator of the Biomimetic, Biological, and Living Materials group within CICECO and Deputy Chair of the Scientific Council of the University of Aveiro. Her research interests primarily involve the development of cost-effective production and separation platforms for high-value compounds with therapeutic applications, with a primary focus on biopharmaceuticals. The developed processes often include the use of ionic liquids and deep eutectic solvents as neoteric solvents.
Author Contributions
P.B., A.K., and M.G.F. conceptualized the idea of the review. P.B. and M.G.F. wrote the first draft of the manuscript. A.K., A.A.T., D.M., and T.S.K. assisted in revising the manuscript.
The authors declare no competing financial interest.
Special Issue
Published as part of Chemical Reviewsvirtual special issue “Ionic Liquids for Diverse Applications”.
References
- Borman S. Proteins from birth to death. Chem. Eng. News. 2007, 85, 34. 10.1021/cen-v085n035.p034. [DOI] [Google Scholar]
- Feeney R. E. Chemical Modification of Food Proteins, Chapter 1, Food proteins. Adv. Chem. 1977, 160, 3–36. 10.1021/ba-1977-0160.ch001. [DOI] [Google Scholar]
- Shang N.; Chaplot S.; Wu J.. Chapter 12, Food proteins for health and nutrition. In Proteins in Food Processing, 2nd ed.; 2018; pp 301–336. 10.1016/B978-0-08-100722-8.00013-9 [DOI] [Google Scholar]
- Roy S.; Desrochers M.; Jurasek L. Proteins as Potential Retention Aids in the Pulp and Paper Industry. J. Wood Chem. Technol. 1989, 9, 407–420. 10.1080/02773818908050307. [DOI] [Google Scholar]
- Dettmer A.; Cavalli E.; Ayub M. A.; Gutterres M. Optimization of the Unhairing Leather Processing With Enzymes And The Evaluation of Inter-Fibrillary Proteins Removal: An Environment-Friendly Alternative. Bioprocess. Biosyst. Eng. 2012, 35, 1317–1324. 10.1007/s00449-012-0719-z. [DOI] [PubMed] [Google Scholar]
- Sheldon R. A.; Woodley J. M. Role Of Biocatalysis in Sustainable Chemistry. Chem. Rev. 2018, 118, 801–838. 10.1021/acs.chemrev.7b00203. [DOI] [PubMed] [Google Scholar]
- Jurado E.; García-Román M.; Luzón G.; Altmajer-Vaz D.; Jiménez-Pérez J. L. Optimization of Lipase Performance in Detergent Formulations For Hard Surfaces. Ind. Eng. Chem. Res. 2011, 50, 11502–11510. 10.1021/ie201486a. [DOI] [Google Scholar]
- de la Rica R.; Matsui H. Applications of Peptide and Protein-Based Materials in Bionanotechnology. Chem. Soc. Rev. 2010, 39, 3499–3509. 10.1039/b917574c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manning M. C.; Chou D. K.; Murphy B. M.; Payne R. W.; Katayama D. S. Stability of Protein Pharmaceuticals: An Update. Pharm. Res. 2010, 27, 544–575. 10.1007/s11095-009-0045-6. [DOI] [PubMed] [Google Scholar]
- Nisthal A.; Wang C. Y.; Ary M. L.; Mayo S. L. Protein Stability Engineering Insights Revealed by Domain-Wide Comprehensive Mutagenesis. Proc. Natl. Acad. Sci. U. S. A. 2019, 116, 16367–16377. 10.1073/pnas.1903888116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chandler P. G.; Broendum S. S.; Riley B. T.; Spence M. A.; Jackson C. J.; McGowan S.; Buckle A. M. Strategies for Increasing Protein Stability. Methods Mol. Biol. 2020, 2073, 163–181. 10.1007/978-1-4939-9869-2_10. [DOI] [PubMed] [Google Scholar]
- e Silva F. A.; Pereira J. F. B.; Kurnia K. A.; Ventura S. P. M.; Silva A. M. S.; Rogers R. D.; Coutinho J. A. P.; Freire M. G. Temperature Dependency of Aqueous Biphasic Systems: an Alternative Approach for Exploring the Differences Between Coulombic-Dominated Salts and Ionic Liquids. Chem. Commun. 2017, 53, 7298–7301. 10.1039/C7CC02294H. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers R. D. Materials Science: Reflections on ionic liquids. Nature 2007, 447, 917–918. 10.1038/447917a. [DOI] [PubMed] [Google Scholar]
- Shiflett M. B.Commercial Applications of Ionic Liquids. Springer International Publishing, 2020; Series ISSN: 2196-6982. [Google Scholar]
- Yang Z. Hofmeister Effects: An Explanation for the Impact of Ionic Liquids on Biocatalysis. J. Biotechnol. 2009, 144, 12–22. 10.1016/j.jbiotec.2009.04.011. [DOI] [PubMed] [Google Scholar]
- Weingärtner H.; Cabrele C.; Herrmann C. How Ionic Liquids Can Help to Stabilize Native Proteins. Phys. Chem. Chem. Phys. 2012, 14, 415–426. 10.1039/C1CP21947B. [DOI] [PubMed] [Google Scholar]
- Freire M. G.; Cláudio A. F. M.; Araújo J. M. M.; Coutinho J. A. P.; Marrucho I. M.; Lopes J. N. C.; Rebelo L. P. N. Aqueous Biphasic Systems: A Boost Brought About by Using Ionic Liquids. Chem. Soc. Rev. 2012, 41, 4966–4995. 10.1039/c2cs35151j. [DOI] [PubMed] [Google Scholar]
- Zhao H. Protein Stabilization and Enzyme Activation in Ionic Liquids: Specific Ion Effects. J. Chem. Technol. Biotechnol. 2016, 91, 25–50. 10.1002/jctb.4837. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Canongia Lopes J. N. A.; Padua A. A. H. Nanostructural Organization in Ionic Liquids. J. Phys. Chem. B 2006, 110, 3330–3335. 10.1021/jp056006y. [DOI] [PubMed] [Google Scholar]
- Russina O.; Triolo A.; Gontrani L.; Caminiti R.; Xiao D.; Hines L. G.; Bartsch R. A. Jr; Quitevis E. L.; Pleckhova N.; Seddon K. R. Morphology, and Intermolecular dynamics of 1-Alkyl-3-methylimidazolium bis{(trifluoromethane)sulfonyl}amide Ionic Liquids: Structural and Dynamic Evidence of Nanoscale Segregation. J. Phys.: Condens. Matter 2009, 21, 424121–424129. 10.1088/0953-8984/21/42/424121. [DOI] [Google Scholar]
- Fumino K.; Wulf A.; Ludwig R. Hydrogen Bonding in Protic Ionic Liquids: Reminiscent of Water. Angew. Chem. Int. 2009, 48, 3184–3186. 10.1002/anie.200806224. [DOI] [PubMed] [Google Scholar]
- Zhu X.; Cui P.; Zhang D.; Liu C. Theoretical Study for Pyridinium-Based Ionic Liquid 1-Ethylpyridinium Trifluoroacetate: Synthesis Mechanism, Electronic Structure, and Catalytic Reactivity. J. Phys. Chem. A 2011, 115, 8255–8263. 10.1021/jp201246j. [DOI] [PubMed] [Google Scholar]
- Liu X.; Zhao Y.; Zhang X.; Zhou G.; Zhang S. Microstructures, and Interaction Analyses of Phosphonium-Based Ionic Liquids: A Simulation Study. J. Phys. Chem. B 2012, 116, 4934–4942. 10.1021/jp210696r. [DOI] [PubMed] [Google Scholar]
- Stassen H. K.; Ludwig R.; Wulf A.; Dupont J. Imidazolium Salt Ion-Pairs in Solution. Chem.—Eur. J. 2015, 21, 8324. 10.1002/chem.201500239. [DOI] [PubMed] [Google Scholar]
- Campetella M.; Bodo E.; Montagna M.; De Santis S.; Gontrani L. Theoretical Study of Ionic Liquids Based on the Cholinium Cation. Ab Initio Simulations of Their Condensed Phases. J. Chem. Phys. 2016, 144, 104504. 10.1063/1.4943197. [DOI] [PubMed] [Google Scholar]
- Kang T. S.; Ishiba K.; Morikawa M.; Kimizuka N. Self-Assembly of Azobenzene Bilayer Membranes in Binary Ionic Liquid-Water Nanostructured Media. Langmuir 2014, 30, 2376–2384. 10.1021/la405010f. [DOI] [PubMed] [Google Scholar]
- Hayes R.; Warr G. G.; Atkin R. Structure and Nanostructure in Ionic Liquids. Chem. Rev. 2015, 115, 6357–6426. 10.1021/cr500411q. [DOI] [PubMed] [Google Scholar]
- Madeira Lau R.; Sorgedrager M. J.; Carrea G.; Rantwijk F. v.; Secundo F.; Sheldon R. A. Dissolution of Candida antarctica lipase B in Ionic Liquids: Effects on Structure and Activity. Green Chem. 2004, 6, 483–487. 10.1039/b405693k. [DOI] [Google Scholar]
- Zhao H.; Jones C. L.; Cowins J. V. Lipase Dissolution and Stabilization in Ether-Functionalized Ionic Liquids. Green Chem. 2009, 11, 1128–1138. 10.1039/b905388c. [DOI] [Google Scholar]
- Bose S.; Armstrong D. W.; Petrich J. W. Enzyme-Catalyzed Hydrolysis of Cellulose in Ionic Liquids: A Green Approach Toward the Production of Biofuels. J. Phys. Chem. B 2010, 114, 8221–8227. 10.1021/jp9120518. [DOI] [PubMed] [Google Scholar]
- Bihari M.; Russell T. P.; Hoagland D. A. Dissolution and Dissolved State of Cytochrome c in a Neat, Hydrophilic Ionic Liquid. Biomacromolecules 2010, 11, 2944–2978. 10.1021/bm100735z. [DOI] [PubMed] [Google Scholar]
- Tamura K.; Nakamura N.; Ohno H. Cytochrome c Dissolved in 1-Allyl-3-Methylimidazolium Chloride Type Ionic Liquid Undergoes a Quasi-Reversible Redox Reaction up to 140°C. Biotechnol. Bioeng. 2012, 109, 729–735. 10.1002/bit.24357. [DOI] [PubMed] [Google Scholar]
- Idris A.; Vijayaraghavan R.; Patti A. F.; MacFarlane D. R. Distillable Protic Ionic Liquids for Keratin Dissolution and Recovery. ACS Sustainable Chem. Eng. 2014, 2, 1888–1894. 10.1021/sc500229a. [DOI] [Google Scholar]
- Tomlinson S. R.; Kehr C. W.; Lopez M. S.; Schlup J. R.; Anthony J. L. Solubility of the Corn Protein Zein in Imidazolium-Based Ionic Liquids. Ind. Eng. Chem. Res. 2014, 53, 2293–2298. 10.1021/ie403659x. [DOI] [Google Scholar]
- Madeira Lau R.; Van Rantwijk F. v.; Seddon K. R.; Sheldon R. A. Lipase-Catalyzed Reactions in Ionic Liquids. Org. Lett. 2000, 2, 4189–4191. 10.1021/ol006732d. [DOI] [PubMed] [Google Scholar]
- Schöfer S. H.; Kaftzik N.; Wasserscheid P.; Kragl U. Enzyme Catalysis in Ionic Liquids: Lipase Catalysed Kinetic Resolution of 1-Phenylethanol with Improved Enantioselectivity. Chem. Commun. 2001, 425–426. 10.1039/b009389k. [DOI] [Google Scholar]
- Sheldon R. A.; Lau R. M.; Sorgedrager M. J.; van Rantwijk F.; Seddon K. R. Biocatalysis in Ionic Liquids. Green Chem. 2002, 4, 147–151. 10.1039/b110008b. [DOI] [Google Scholar]
- Kragl U.; Eckstein M.; Kaftzik N. Enzyme Catalysis in Ionic Liquids. Curr. Opin. Biotechnol. 2002, 13, 565–571. 10.1016/S0958-1669(02)00353-1. [DOI] [PubMed] [Google Scholar]
- Park S.; Kazlauskas R. J. Biocatalysis in Ionic Liquids-Advantages Beyond Green Technology. Curr. Opin. Biotechnol. 2003, 14, 432–437. 10.1016/S0958-1669(03)00100-9. [DOI] [PubMed] [Google Scholar]
- van Rantwijk F.; Madeira Lau R.; Sheldon R. A. Biocatalytic Transformations in Ionic Liquids. Trend. Biotechnol. 2003, 21, 131–138. 10.1016/S0167-7799(03)00008-8. [DOI] [PubMed] [Google Scholar]
- Yang Z.; Pan W. Ionic Liquids: Green Solvents for Non-aqueous Bio-catalysis. Enz. Microb. Techn. 2005, 37, 19–28. 10.1016/j.enzmictec.2005.02.014. [DOI] [Google Scholar]
- Moon Y. H.; Lee S.; Ha M. S. H.; Koo Y.-M. Enzyme-Catalyzed Reactions in Ionic Liquids. Korean J. Chem. Eng. 2006, 23, 247–263. 10.1007/BF02705724. [DOI] [Google Scholar]
- van Rantwijk F.; Sheldon R. A. Biocatalysis in Ionic Liquids. Chem. Rev. 2007, 107, 2757–2785. 10.1021/cr050946x. [DOI] [PubMed] [Google Scholar]
- Roosen C.; Müller P.; Greiner L. Ionic Liquids in Biotechnology: Applications and Perspectives for Biotransformations. Appl. Microbiol. Biotechnol. 2008, 81, 607–614. 10.1007/s00253-008-1730-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pinto P. C. A. G.; Saraiva M. L. M. F. S.; Lima J. L. F. C. Oxidoreductase Behavior in Ionic Liquids: A Review. Anal. Sci. 2008, 24, 1231–1238. 10.2116/analsci.24.1231. [DOI] [PubMed] [Google Scholar]
- Goldfeder M.; Fishman A. Modulating Enzyme Activity using Ionic Liquids or Surfactant. Appl. Microbiol. Biotechnol. 2014, 98, 545–554. 10.1007/s00253-013-5395-7. [DOI] [PubMed] [Google Scholar]
- Wahlström R. M.; Suurnäkki A. Enzymatic Hydrolysis of Lignocellulosic Polysaccharides in the Presence of Ionic Liquids. Green Chem. 2015, 17, 694–714. 10.1039/C4GC01649A. [DOI] [Google Scholar]
- Potdar M. K.; Kelso G. F.; Schwarz L.; Zhang C.; Hearn M. T. W. Recent Developments in Chemical Synthesis with Biocatalysts in Ionic Liquids. Molecules 2015, 20, 16788–16816. 10.3390/molecules200916788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gao W.-W.; Zhang F.-X.; Zhang G.-X.; Zhou C.-H. Key Factors Affecting the Activity and Stability of Enzymes in Ionic Liquids and Novel Applications in Biocatalysis. Biochem. Bioeng. J. 2015, 99, 67–84. 10.1016/j.bej.2015.03.005. [DOI] [Google Scholar]
- Itoh T. Ionic Liquids as Tool to Improve Enzymatic Organic Synthesis. Chem. Rev. 2017, 117, 10567–10607. 10.1021/acs.chemrev.7b00158. [DOI] [PubMed] [Google Scholar]
- Meyer L.-E.; von Langermann J.; Kragl U. Recent developments in Biocatalysis in Multiphasic Ionic Liquid Reaction Systems. Biophys Rev. 2018, 10, 901–910. 10.1007/s12551-018-0423-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xu P.; Liang S.; Zong M.-H.; Lou W.-Y. Ionic Liquids for Regulating Biocatalytic Process: Achievements and Perspectives. Biotechnol. Adv. 2021, 51, 107702. 10.1016/j.biotechadv.2021.107702. [DOI] [PubMed] [Google Scholar]
- Kunz W.; Lo Nostro P.; Ninham B. W. The Present State of Affairs with Hofmeister Effects. Curr. Opin. Colloid Interface Sci. 2004, 9, 1–18. 10.1016/j.cocis.2004.05.004. [DOI] [Google Scholar]
- Dupont J. On the Solid, Liquid and Solution Structural Organization of Imidazolium Ionic Liquids. J. Braz. Chem. Soc. 2004, 15, 341–350. 10.1590/S0103-50532004000300002. [DOI] [Google Scholar]
- Schröder U.; Wadhawan J.; Compton R.; Marken F.; Suarez P.; Consorti C.; Dupont S. R. Water-Induced Accelerated Ion Diffusion: Voltammetric Studies in 1-Methyl-3-[2,6-(S)-dimethylocten-2-yl] imidazolium tetrafluoroborate, 1-Butyl-3-Methylimidazolium Tetrafluoroborate and Hexafluorophosphate Ionic Liquids. New J. Chem. 2000, 24, 1009–1015. 10.1039/b007172m. [DOI] [Google Scholar]
- Seddon K. R.; Stark A.; Torres M.-J. Influence of Chloride, Water, and Organic Solvents on the Physical Properties of Ionic Liquids. Pure Appl. Chem. 2000, 72, 2275–2287. 10.1351/pac200072122275. [DOI] [Google Scholar]
- Erbeldinger M.; Mesiano A. J.; Russell A. J. Enzymatic Catalysis of Formation of Z-Aspartame in Ionic Liquid-An Alternative to Enzymatic Catalysis in Organic Solvents. Biotechnol. Prog. 2000, 16, 1129–1131. 10.1021/bp000094g. [DOI] [PubMed] [Google Scholar]
- Lozano P.; Diego T. D.; Carrié D.; Vaultier M.; Iborra J. L. Over-Stabilization of Candida antarctica lipase B by Ionic Liquids in Ester Synthesis. Biotechnol. Lett. 2001, 23, 1529–1533. 10.1023/A:1011697609756. [DOI] [Google Scholar]
- Baker S. N.; McCleskey T. M.; Pandey S.; Baker G. A. Fluorescence Studies of Protein Thermostability in Ionic Liquids. Chem. Commun. 2004, 940–941. 10.1039/b401304m. [DOI] [PubMed] [Google Scholar]
- De Diego T.; Lozano P.; Gmouh S.; Vaultier M.; Iborra J. L. Fluorescence and CD Spectroscopic Analysis of the Α-Chymotrypsin Stabilization by the Ionic Liquid, 1-Ethyl-3-Methylimidazolium Bis[(Trifluoromethyl)Sulfonyl]Amide. Biotechnol. Bioeng. 2004, 88, 916–924. 10.1002/bit.20330. [DOI] [PubMed] [Google Scholar]
- Fujita K.; MacFarlane D. R.; Forsyth M. Protein Solubilising and Stabilising Ionic Liquids. Chem. Commun. 2005, 4804–4806. 10.1039/b508238b. [DOI] [PubMed] [Google Scholar]
- Lozano P.; Diego T. D.; Gmouth S.; Vaultier M.; Iborra J. L. Dynamic Structure/Function Relationships in Enzyme Stabilization by Ionic Liquids. Biocatal. Biotransform. 2005, 23, 169–176. 10.1080/10242420500198657. [DOI] [Google Scholar]
- Fujita K.; MacFarlane D. R.; Forsyth M.; Fujita M. Y.; Murata K.; Nakamura N.; Ohno H. Solubility and Stability of Cytochrome c in Hydrated Ionic Liquids: Effect of Oxo Acid Residues and Kosmotropicity. Biomacromolecules 2007, 8, 2080–2086. 10.1021/bm070041o. [DOI] [PubMed] [Google Scholar]
- McCarty T. A.; Page P. M.; Baker G. A.; Bright F. V. Behavior of Acrylodan-Labeled Human Serum Albumin Dissolved in Ionic Liquids. Ind. Eng. Chem. Res. 2008, 47, 560–569. 10.1021/ie071165k. [DOI] [Google Scholar]
- Byrne N.; Angell C. A. The Solubility of Hen Lysozyme in Ethylammonium Nitrate/H2O Mixtures and a Novel Approach to Protein Crystallization. Molecules 2010, 15, 793–803. 10.3390/molecules15020793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fujita K.; Ohno H. Enzymatic Activity and Thermal Stability of Metallo Proteins in Hydrated Ionic Liquids. Biopolymers 2010, 93, 1093–1099. 10.1002/bip.21526. [DOI] [PubMed] [Google Scholar]
- Attri P.; Venkatesu P.; Kumar A. Activity and Stability of α-Chymotrypsin in Biocompatible Ionic Liquids: Enzyme Refolding by Triethyl Ammonium Acetate. Phys. Chem. Chem. Phys. 2011, 13, 2788–2796. 10.1039/C0CP01291B. [DOI] [PubMed] [Google Scholar]
- Jha I.; Attri P.; Venkatesu P. Unexpected Effects of the Alteration of Structure and Stability of Myoglobin and Hemoglobin in Ammonium-based Ionic Liquids. Phys. Chem. Chem. Phys. 2014, 16, 5514–5526. 10.1039/c3cp54398f. [DOI] [PubMed] [Google Scholar]
- Bekhouche M.; Blum L. J.; Doumèche B. Contribution of Dynamic and Static Quenchers for the Study of Protein Conformation in Ionic Liquids by Steady-State Fluorescence Spectroscopy. J. Phys. Chem. B 2012, 116, 413–423. 10.1021/jp205094c. [DOI] [PubMed] [Google Scholar]
- Baker G. A.; Heller W. T. Small-Angle Neutron Scattering Studies of Model Protein Denaturation in Aqueous Solutions of the Ionic Liquid 1-butyl-3-methylimidazolium Chloride. Chem. Eng. J. 2009, 147, 6–12. 10.1016/j.cej.2008.11.033. [DOI] [Google Scholar]
- Heller W. T.; O’Neill H. M.; Zhang Q.; Baker G. A. Characterization of the Influence of the Ionic Liquid 1-Butyl-3-methylimidazolium Chloride on the Structure and Thermal Stability of Green Fluorescent Protein. J. Phys. Chem. B 2010, 114, 13866–13871. 10.1021/jp105611b. [DOI] [PubMed] [Google Scholar]
- Page T. A.; Kraut N. D.; Page P. M.; Baker G. A.; Bright F. V. Dynamics of Loop 1 of Domain I in Human Serum Albumin When Dissolved in Ionic Liquids. J. Phys. Chem. B 2009, 113, 12825–12830. 10.1021/jp904475v. [DOI] [PubMed] [Google Scholar]
- Klähn M.; Lim G. S.; Seduraman A.; Wu P. On the Different Roles of Anions and Cations in the Solvation of Enzymes in Ionic Liquids. Phys. Chem. Chem. Phys. 2011, 13, 1649–1662. 10.1039/C0CP01509A. [DOI] [PubMed] [Google Scholar]
- Micaêlo N. M.; Soares C. M. Protein Structure and Dynamics in Ionic Liquids. Insights from Molecular Dynamics Simulation Studies. J. Phys. Chem. B 2008, 112, 2566–2572. 10.1021/jp0766050. [DOI] [PubMed] [Google Scholar]
- Wehofsky N.; Wespe C.; Cerovsky V.; Pech A.; Hoess E.; Rudolph R.; Bordusa F. Ionic Liquids and Proteases: A Clean Alliance for Semisynthesis. Chem. Bio. Chem. 2008, 9, 1493–1499. 10.1002/cbic.200800025. [DOI] [PubMed] [Google Scholar]
- Cláudio A. F. M.; Neves M. C.; Shimizu K.; Lopes J. N. C.; Freire M. G.; Coutinho J. A. P. The Magic of Aqueous Solutions of Ionic Liquids: Ionic Liquids as a Powerful Class of Catanionic Hydrotropes. Green Chem. 2015, 17, 3948–3963. 10.1039/C5GC00712G. [DOI] [PMC free article] [PubMed] [Google Scholar]
- de Diego T.; Lozano P.; Gmouh M.; Voltier M.; Iborra I. M. Understanding Structure-Stability Relationships of Candida Antartica Lipase B in Ionic Liquids. Biomacromolecules 2005, 6, 1457–1464. 10.1021/bm049259q. [DOI] [PubMed] [Google Scholar]
- Perkin S.; Salanne M.; Madden P.; Lynden-Bell R. Is a Stern and Diffuse Layer Model Appropriate to Ionic Liquids at Surfaces?. Pro. Nat. Aca. Sci. 2013, 110, E4121 10.1073/pnas.1314188110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yee P.; Shah J. K.; Maginn E. J. State of Hydrophobic and Hydrophilic Ionic Liquids in Aqueous Solutions: Are the Ions Fully Dissociated?. J. Phys. Chem. B 2013, 117, 12556–12566. 10.1021/jp405341m. [DOI] [PubMed] [Google Scholar]
- Bernardes C. E. S.; Minas da Piedade M. E.; Canongia Lopes J. N. The Structure of Aqueous Solutions of a Hydrophilic Ionic Liquid: The Full Concentration Range of 1-Ethyl-3-methylimidazolium Ethylsulfate and Water. J. Phys. Chem. B 2011, 115, 2067–2074. 10.1021/jp1113202. [DOI] [PubMed] [Google Scholar]
- Jiang W.; Wang Y.; Voth G. A. Molecular Dynamics Simulation of Nanostructural Organization in Ionic Liquid/Water Mixtures. J. Phys. Chem. B 2007, 111, 4812–4818. 10.1021/jp067142l. [DOI] [PubMed] [Google Scholar]
- Zhu X.; Zhang H.; Li H. The Structure of Water in Dilute Aqueous Solutions of Ionic Liquids: IR and NMR Study. J. Mol. Liq. 2014, 197, 48–51. 10.1016/j.molliq.2014.04.032. [DOI] [Google Scholar]
- Saihara K.; Yoshimura Y.; Ohta S.; Shimizu A. Properties of Water Confined in Ionic Liquids. Sci. Rep. 2015, 5, 10619. 10.1038/srep10619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y.; Cremer P. S. The Inverse and Direct Hofmeister Series for Lysozyme. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 15249–15253. 10.1073/pnas.0907616106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldwin R. L. How Hofmeister Ion Interactions Affect Protein Stability. Biophys. J. 1996, 71, 2056–2063. 10.1016/S0006-3495(96)79404-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jha I.; Venkatesu P. Endeavour to Simplify the Frustrated Concept of Protein-Ammonium Family Ionic Liquid Interactions. Phys. Chem. Chem. Phys. 2015, 17, 20466–20484. 10.1039/C5CP01735A. [DOI] [PubMed] [Google Scholar]
- Hofmeister F. About Regularities in the Protein Precipitating Effects of Salts and the Relation of These Effects with the Physiological Behavior of Salts. Arch. Exp. Pathol. Pharmacol. 1888, 24, 247–260. 10.1007/BF01918191. [DOI] [Google Scholar]
- Hofmeister F. About the Water Withdrawing Effect of the Salts. Arch. Exp. Pathol. Pharmacol. 1888, 25, 1–30. 10.1007/BF01838161. [DOI] [Google Scholar]
- Cox W. M.; Wolfenden J. H. The Viscosity of Strong Electrolytes Measured by a Differential Method. Proc. R. Soc. London 1934, 145, 475–488. 10.1098/rspa.1934.0113. [DOI] [Google Scholar]
- Gurney R. W.Ionic Processes in Solution; McGraw-Hill: Chicago, 1953. [Google Scholar]
- Marcus Y. Effect of Ions on the Structure of Water: Structure Making and Breaking. Chem. Rev. 2009, 109, 1346–1370. 10.1021/cr8003828. [DOI] [PubMed] [Google Scholar]
- Zhang Y.; Cremer P. S. The Inverse and Direct Hofmeister Series for Lysozyme. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 15249–15253. 10.1073/pnas.0907616106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldwin R. L. How Hofmeister Ion Interactions Affect Protein Stability. Biophys. J. 1996, 71, 2056–2063. 10.1016/S0006-3495(96)79404-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins K. D.; Washabaugh M. W. Q. The Hofmeister Effect and the Behavior of Water at Interfaces. Rev. Biophys. 1985, 18, 323–422. 10.1017/S0033583500005369. [DOI] [PubMed] [Google Scholar]
- Jungwirth P.; Cremer P. S. Beyond Hofmeister. Nat. Chem. 2014, 6, 261–263. 10.1038/nchem.1899. [DOI] [PubMed] [Google Scholar]
- Gregory K. P.; Wanless E. J.; Webber G. B.; Craig V. S. J.; Page A. J. The Electrostatic Origins of Specific Ion Effects: Quantifying the Hofmeister Series for Anions. Chem. Sci. 2021, 12, 15007–15015. 10.1039/D1SC03568A. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tadeo X.; López-Méndez B.; Castanó D.; Trigueros T.; Millet O. Protein Stabilization, and the Hofmeister Effect: The Role of Hydrophobic Solvation. Biophys. J. 2009, 97, 2595–2603. 10.1016/j.bpj.2009.08.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Y.; Cremer P. S. Interactions between Macromolecules, and Ions: the Hofmeister Series. Curr. Opin. Chem. Bio. 2006, 10, 658–663. 10.1016/j.cbpa.2006.09.020. [DOI] [PubMed] [Google Scholar]
- Ball P. Water as a Biomolecule. ChemPhysChem 2008, 9, 2677–2685. 10.1002/cphc.200800515. [DOI] [PubMed] [Google Scholar]
- Tobias D. J.; Hemminger J. C. Getting Specific about Specific Ion Effects. Science 2008, 319, 1197–1198. 10.1126/science.1152799. [DOI] [PubMed] [Google Scholar]
- Hribar B.; Southall N. T.; Vlachy V.; Dill K. A. How Ions Affect the Structure of Water. J. Am. Chem. Soc. 2002, 124, 12302–12311. 10.1021/ja026014h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thomas A. S.; Elcock A. H. Molecular Dynamics Simulations of Hydrophobic Associations in Aqueous Salt Solutions Indicate a Connection Between Water Hydrogen Bonding and the Hofmeister Effect. J. Am. Chem. Soc. 2007, 129, 14887–14898. 10.1021/ja073097z. [DOI] [PubMed] [Google Scholar]
- Freire M. G.; Neves C. M. S. S.; Silva A. M. S.; Santos L. M. N. B. F.; Marrucho I. M.; Rebelo L. P. N.; Shah J. K.; Maginn E. J.; Coutinho J. A. P. 1H NMR and Molecular Dynamics Evidence for an Unexpected Interaction on the Origin of Salting-In/Salting-Out Phenomena. J. Phys. Chem. B 2010, 114, 2004–2014. 10.1021/jp9095634. [DOI] [PubMed] [Google Scholar]
- Govindarajan S.; Goldstein R. A. On the Thermodynamic Hypothesis of Protein Folding. Proc. Natl. Acad. Sci. U.S.A. 1998, 95, 5545–5549. 10.1073/pnas.95.10.5545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hvidsten T. R.; Lægreid A.; Kryshtafovych A.; Andersson G.; Fidelis K.; Komorowski J. A Comprehensive Analysis of The Structure-Function Relationship in Proteins Based on Local Structure Similarity. PLoS One 2009, 4, e6266 10.1371/journal.pone.0006266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Anfinsen C. Principles that Govern the Folding of Protein Chains. Science 1973, 181, 223–230. 10.1126/science.181.4096.223. [DOI] [PubMed] [Google Scholar]
- Dill K. A. Dominant Forces in Protein Folding. Biochemistry 1990, 29, 7133–7155. 10.1021/bi00483a001. [DOI] [PubMed] [Google Scholar]
- Kim P. S.; Baldwin R. L. Intermediates in the Folding Reactions of Small Proteins. Annu. Rev. Biochem. 1990, 59, 631–660. 10.1146/annurev.bi.59.070190.003215. [DOI] [PubMed] [Google Scholar]
- Baker D.; Agard D. A. Kinetics versus Thermodynamics in Protein Folding. Biochemistry 1994, 33, 7505–7509. 10.1021/bi00190a002. [DOI] [PubMed] [Google Scholar]
- Kumar A.; Venkatesu P. Overview of the Stability of α-Chymotrypsin in Different Solvent Media. Chem. Rev. 2012, 112, 4283–4307. 10.1021/cr2003773. [DOI] [PubMed] [Google Scholar]
- Benedetto A.; Ballone P. Room Temperature Ionic Liquids Meet Biomolecules: A Microscopic View of Structure and Dynamics. ACS Sustainable Chem. Eng. 2016, 4, 392–412. 10.1021/acssuschemeng.5b01385. [DOI] [Google Scholar]
- Chowdhury R.; Mojumdar S. S.; Chattoraj S.; Bhattacharyya K. Effect of Ionic Liquid on the Native and Denatured State of a Protein Covalently Attached to a Probe: Solvation Dynamics Study. J. Chem. Phys. 2012, 137, 055104. 10.1063/1.4739922. [DOI] [PubMed] [Google Scholar]
- Kumar Das D.; Kumar Das A.; Kumar Mandal A.; Mondal T.; Bhattacharyya K. Effect of an Ionic liquid on the Unfolding of Human Serum Albumin: A Fluorescence Correlation Spectroscopy Study. Chem.Phys.Chem. 2012, 13, 1949–1955. 10.1002/cphc.201100421. [DOI] [PubMed] [Google Scholar]
- Sen Mojumdar S.; Chowdhury R.; Chattoraj S.; Bhattacharyya K. Role of Ionic Liquid on the Conformational Dynamics in the Native, Molten Globule, and Unfolded States of Cytochrome C: A Fluorescence Correlation Spectroscopy Study. J. Phys. Chem. B 2012, 116, 12189–12198. 10.1021/jp307297s. [DOI] [PubMed] [Google Scholar]
- Pabbathi A.; Ghosh S.; Samanta A. FCS Study of the Structural Stability of Lysozyme in the Presence of Morpholinium Salts. J. Phys. Chem. B 2013, 117, 16587–16593. 10.1021/jp409842d. [DOI] [PubMed] [Google Scholar]
- Geng F.; Zheng L.; Liu J.; Yu L.; Tung C. Interactions Between a Surface-Active Imidazolium Ionic Liquid and BSA. Colloid Polym. Sci. 2009, 287, 1253–1259. 10.1007/s00396-009-2085-1. [DOI] [Google Scholar]
- Geng F.; Zheng L.; Yu L.; Li G.; Tung C. Interaction of Bovine Serum Albumin and Long-Chain Imidazolium Ionic Liquid Measured by Fluorescence Spectra and Surface Tension. Process Biochem. 2010, 45, 306–311. 10.1016/j.procbio.2009.10.001. [DOI] [Google Scholar]
- Singh T.; Bharmoria P.; Morikawa M.; Kimizuka N.; Kumar A. Ionic Liquids Induced Structural Changes of Bovine Serum Albumin in Aqueous Media: A Detailed Physicochemical and Spectroscopic Study. J. Phys. Chem. B 2012, 116, 11924–11935. 10.1021/jp303609h. [DOI] [PubMed] [Google Scholar]
- Bharmoria P.; Rao K. S.; Trivedi T. J.; Kumar A. Biamphiphilic Ionic Liquid Induced Folding Alterations in the Structure of Bovine Serum Albumin in Aqueous Medium. J. Phys. Chem. B 2014, 118, 115–124. 10.1021/jp4102042. [DOI] [PubMed] [Google Scholar]
- Gehlot P. S.; Rao K. S.; Bharmoria P.; Damarla K.; Gupta H.; Drechsler M.; Kumar A. Spontaneous Formation of Multiarchitecture Vesicles of [C8mim]Br + [Na]DBS in Aqueous Medium: Synergic Interplay of Electrostatic, Hydrophobic, and π-π Stacking Interactions. J. Phys. Chem. B 2015, 119, 15300–15309. 10.1021/acs.jpcb.5b09850. [DOI] [PubMed] [Google Scholar]
- Bharmoria P.; Mehta M. J.; Pancha I.; Kumar A. Structural and Functional Stability of Cellulase in Aqueous-Biamphiphilic Ionic Liquid Surfactant Solution. J. Phys. Chem. B 2014, 118, 9890–9899. 10.1021/jp506211b. [DOI] [PubMed] [Google Scholar]
- Bharmoria P.; Trivedi T. J.; Kumar A. Ionic Liquid Induced All-α to α+β Conformational Transition in Cytochrome c with Improved Peroxidase Activity in Aqueous Medium. Phys. Chem. Chem. Phys. 2015, 17, 10189–10199. 10.1039/C4CP06044J. [DOI] [PubMed] [Google Scholar]
- Singh G.; Kang T. S. Ionic Liquid Surfactant Mediated Structural Transitions and Self-Assembly of Bovine Serum Albumin in Aqueous Media: Effect of Functionalization of Ionic Liquid Surfactants. J. Phys. Chem. B 2015, 119, 10573–10585. 10.1021/acs.jpcb.5b04854. [DOI] [PubMed] [Google Scholar]
- Wang X.; Liu J.; Sun L.; Yu L.; Jiao J.; Wang R. Interaction of Bovine Serum Albumin with Ester-Functionalized Anionic Surface-Active Ionic Liquids in Aqueous Solution: A Detailed Physicochemical and Conformational Study. J. Phys. Chem. B 2012, 116, 12479–12488. 10.1021/jp307516a. [DOI] [PubMed] [Google Scholar]
- Yan H.; Wu J.; Dai G.; Zhong A.; Chen H.; Yang J.; Han D. Interaction Mechanism of Ionic Liquids [Cnmim][Br] (n = 4, 6, 8, 10) with Bovine Serum Albumin. J. Lumin. 2012, 132, 622–628. 10.1016/j.jlumin.2011.10.026. [DOI] [Google Scholar]
- Pinto P. C. A. G.; Ribeiro D. M. G. P.; Azevedo A. M. O.; Justina V. D.; Cunha E.; Bica K.; Vasiloiu M.; Reisa S.; Saraiva M. L. M. F. S. Active Pharmaceutical Ingredients Based on Salicylate Ionic Liquids: Insights into the Evaluation of Pharmaceutical Profiles. New J. Chem. 2013, 37, 4095–4102. 10.1039/c3nj00731f. [DOI] [Google Scholar]
- Zhou T.; Ao M.; Xu G.; Liu T.; Zhang J. Interactions of Bovine Serum Albumin with Cationic Imidazolium and Quaternary Ammonium Gemini Surfactants: Effects of Surfactant Architecture. J. Colloids Interface Sci. 2013, 389, 175–181. 10.1016/j.jcis.2012.08.067. [DOI] [PubMed] [Google Scholar]
- Gospodarczyk W.; Kozak M. Interaction of two Imidazolium Gemini Surfactants with two Model Proteins BSA and HEWL. Colloid Polym. Sci. 2015, 293, 2855–2866. 10.1007/s00396-015-3671-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maurya J. K.; Mir M. U. H.; Maurya N.; Dohare N.; Ali A.; Patel R. A Spectroscopic and Molecular Dynamic Approach on the Interaction between Ionic Liquid type Gemini Surfactant and Human Serum Albumin. J. Biomol. Str. Dyn. 2016, 34, 2130. 10.1080/07391102.2015.1109552. [DOI] [PubMed] [Google Scholar]
- Maurya J. K.; Mir M. U. H.; Singh U. K.; Maurya N.; Dohare N.; Patel S.; Ali A.; Patel R. Molecular Investigation of the Interaction between Ionic Liquid Type Gemini Surfactant and Lysozyme: A Spectroscopic and Computational Approach. Biopolymers 2015, 103, 406–415. 10.1002/bip.22647. [DOI] [PubMed] [Google Scholar]
- Singh T.; Boral S.; Bohidar H. B.; Kumar A. Interaction of Gelatin with Room Temperature Ionic Liquids: A Detailed Physicochemical Study. J. Phys. Chem. B 2010, 114, 8441–8448. 10.1021/jp102419f. [DOI] [PubMed] [Google Scholar]
- Liu Y.; Yang L.; Guo R. Interaction between β-Casein Micelles and Imidazolium Based Ionic Liquid Surfactant. Soft Matter 2013, 9, 3671–3680. 10.1039/c2sm27134f. [DOI] [Google Scholar]
- Liu Y.; Yang L.; Mao H.; Guo R. Comparative Studies on the Interaction of [C4mim]Br, and [C8mim]Br the β-casein Micelles. Colloids Surf., A 2014, 441, 581–588. 10.1016/j.colsurfa.2013.10.012. [DOI] [Google Scholar]
- Cao C.; Lei J.; Zhang L.; Du F.-P. Equilibrium and Dynamic Interfacial Properties of Protein/Ionic-Liquid-Type Surfactant Solutions at the Decane/Water Interface. Langmuir 2014, 30, 13744–13753. 10.1021/la502890w. [DOI] [PubMed] [Google Scholar]
- Cao C.; Lei J.; Huang T.; Du F.-P. Impact of Ionic Liquid-Type Imidazolium Surfactant Addition on Dynamic Properties of BSA Adsorption Layers at Different pH. Soft Matter 2014, 10, 8896–8904. 10.1039/C4SM01666A. [DOI] [PubMed] [Google Scholar]
- Mandal B.; Mondal S.; Pan A.; Moulik S. P.; Ghosh S. Physicochemical Study of the Interaction of Lysozyme with Surface Active Ionic Liquid 1-butyl-3-methylimidazolium Octylsulfate [C4mim][C8OSO3] in Aqueous and Buffer Media. Colloids Surf., A 2015, 484, 345–353. 10.1016/j.colsurfa.2015.07.052. [DOI] [Google Scholar]
- Kuchlyan J.; Kundu N.; Sarkar N. Ionic Liquids in Microemulsions: Formulation and Characterization. Curr. Opin. Colloid Interface Sci. 2016, 25, 27–38. 10.1016/j.cocis.2016.05.011. [DOI] [Google Scholar]
- Pavlidis I. V.; Tzafestas K.; Stamatis H. Water-in-Ionic Liquid Microemulsion-Based Organogels as Novel Matrices for Enzyme Immobilization. Biotechnol. J. 2010, 5, 805–812. 10.1002/biot.201000052. [DOI] [PubMed] [Google Scholar]
- Ventura S. P. M.; Santos L. D. F.; Saraiva J. A.; Coutinho J. A. P. Ionic liquids Microemulsions: The Key to Candida antarctica lipase B Superactivity. Green Chem. 2012, 14, 1620–1625. 10.1039/c2gc35197h. [DOI] [Google Scholar]
- Mao Q.-X.; Wang H.; Shu Y.; Chen X.-W.; Wang J.-H. A Dual-Ionic Liquid Microemulsion System for the Selective Isolation of Hemoglobin. RSC Adv. 2014, 4, 8177–8182. 10.1039/c3ra46736h. [DOI] [Google Scholar]
- Zhu L.-Y.; Li G.-Q.; Zheng F.-Y. Interaction of Bovine Serum Albumin, with two Alkylimidazolium based Ionic Liquids Investigated by Microcalorimetry and Circular Dichroism. J. Biophys. Chem. 2011, 02, 147–152. 10.4236/jbpc.2011.22018. [DOI] [Google Scholar]
- Paul B. K.; Ganguly A.; Guchhait N. Deciphering the Interaction of a Model Transport Protein with a Prototypical Imidazolium Room Temperature Ionic Liquid: Effect on the Conformation and Activity of the Protein. J. Photochem. Photobiol., B 2014, 133, 99–107. 10.1016/j.jphotobiol.2014.03.007. [DOI] [PubMed] [Google Scholar]
- Fan Y.; Dong X.; Zhong Y.; Li J.; Miao J.; Hua S.; Li Y.; Cheng B.; Chen W. Effects of Ionic Liquids on the Hydrolysis of Casein by Lumbrokinase. Biochem. Eng. J. 2016, 109, 35. 10.1016/j.bej.2015.12.020. [DOI] [Google Scholar]
- Banerjee C.; Roy A.; Kundu N.; Banik D.; Sarkar N. A New Strategy to Prepare Giant Vesicles from Surface Active Ionic Liquids (SAILs): A Study of Protein Dynamics in a Crowded Environment using a Fluorescence Correlation Spectroscopic Technique. Phys. Chem. Chem. Phys. 2016, 18, 14520–14530. 10.1039/C5CP07225E. [DOI] [PubMed] [Google Scholar]
- Rao K. S.; Bharmoria P.; Trivedi T. J.; Arvind K.. Self-assembly of Surface-Active Ionic Liquids in Aqueous Medium. In Ionic Liquid-Based Surfactant Science: Formulation, Characterization, and Applications; John Wiley & Sons, Inc., 2015; Chapter 8, ISBN: 978-1-118-83419-0. [Google Scholar]
- Bhadani A.; Misono T.; Singh S.; Sakai K.; Sakai H.; Abe M. Structural Diversity, Physicochemical Properties and Application of Imidazolium Surfactants: Recent Advances. Adv. Colloid Interface Sci. 2016, 231, 36–58. 10.1016/j.cis.2016.03.005. [DOI] [PubMed] [Google Scholar]
- Ananthapadmanabhan K. P.Interactions of Surfactants with Polymers and Proteins; Goddard E. D., Ananthapadmanabhan K. P., Eds.; CRC Press, Inc: London, U.K., 1993; Chapter 8. [Google Scholar]
- Dalgleish D. G. In Emulsions and Emulsion Stability; Sjoblom J., Ed.; Marcel Dekker: New York, 1996; Chapter 5. [Google Scholar]
- Bharmoria P.; Kumar A. Unusually High Thermal Stability and Peroxidase Activity of Cytochrome c in Ionic Liquid Colloidal Formulation. Chem. Commun. 2016, 52, 497–500. 10.1039/C5CC05722A. [DOI] [PubMed] [Google Scholar]
- Brogan A. P. S.; Hallett J. P. Solubilizing and Stabilizing Proteins in Anhydrous Ionic Liquids through Formation of Protein-Polymer Surfactant Nanoconstructs. J. Am. Chem. Soc. 2016, 138, 4494–4501. 10.1021/jacs.5b13425. [DOI] [PubMed] [Google Scholar]
- Brogan A. P. S.; Sharma K. P.; Perriman A. W.; Mann S. Enzyme Activity in Liquid Lipase Melts as a Step Towards Solvent-Free Biology at 150°C. Nat. Commun. 2014, 5, 5058. 10.1038/ncomms6058. [DOI] [PubMed] [Google Scholar]
- Byrne N.; Angell C. A. Formation and Dissolution of Hen Egg White Lysozyme Amyloid Fibrils in Protic Ionic Liquids. Chem.Commun. 2009, 1046–1048. 10.1039/b817590j. [DOI] [PubMed] [Google Scholar]
- Chen X.; McRae S.; Samanta D.; Emrick T. Polymer-Protein Conjugation in Ionic Liquids. Macromolecules 2010, 43, 6261–6263. 10.1021/ma101156e. [DOI] [Google Scholar]
- Tseng M.-C.; Tseng M.-J.; Chu Y.-H. Affinity Ionic Liquid. Chem. Commun. 2009, 7503–7505. 10.1039/b916411a. [DOI] [PubMed] [Google Scholar]
- Ventura S. P. M.; Sousa S. G.; Serafim L. S.; Lima Á. S.; Freire M. G.; Coutinho J. A. P. Ionic-Liquid-Based Aqueous Biphasic Systems with Controlled pH: The Ionic Liquid Anion Effect. J. Chem. Eng. Data 2012, 57, 507–512. 10.1021/je2010787. [DOI] [Google Scholar]
- Passos H.; Freire M. G.; Coutinho J. A. P. Ionic Liquid Solutions as Extractive Solvents for Value-Added Compounds from Biomass. Green. Chem. 2014, 16, 4786–4815. 10.1039/C4GC00236A. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Quental M. V.; Caban M.; Pereira M. M.; Stepnowski P.; Coutinho J. A. P.; Freire M. G. Enhanced Extraction of Proteins using Cholinium-Based Ionic Liquids as Phase-Forming Components of Aqueous Biphasic Systems. Biotechnol. J. 2015, 10, 1457–1466. 10.1002/biot.201500003. [DOI] [PubMed] [Google Scholar]
- Passos H.; Luís A.; Coutinho J. A. P.; Freire M. G. Thermoreversible (Ionic-Liquid- Based) Aqueous Biphasic Systems. Sci. Rep. 2016, 6, 20276. 10.1038/srep20276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cooper G. M.; Sunderland M. A.. The Molecular Composition of Cells. In Cell: A Molecular Approach, 2nd ed.; 2000. [Google Scholar]
- Nandi N.; Bhattacharyya K.; Bagchi B. Dielectric Relaxation and Solvation Dynamics of Water in Complex Chemical and Biological Systems. Chem. Rev. 2000, 100, 2013–2045. 10.1021/cr980127v. [DOI] [PubMed] [Google Scholar]
- Jeffrey G. A.; Saenger W.. Hydrogen Bonding in Biological Systems; Springer-Verlag: Berlin, 1991; Chapter 23. [Google Scholar]
- Oncley J. L.Proteins, Amino Acids and Peptides; John E. J., Edsall J. T., Eds.; Reinhold: New York, 1943; Chapter 22, p 543. [Google Scholar]
- Ball P. Water as a Biomolecule. ChemPhysChem 2008, 9, 2677–2685. 10.1002/cphc.200800515. [DOI] [PubMed] [Google Scholar]
- Wang J. H.; Anfinsen C. B.; Polestra F. M. Theory of the Self-diffusion of Water in Protein Solutions. A New Method for Studying the Hydration and Shape of Protein Molecules. J. Am. Chem. Soc. 1954, 76, 4755–4763. 10.1021/ja01648a001. [DOI] [Google Scholar]
- Pal S. K.; Zewail A. H. Dynamics of Water in Biological Recognition. Chem. Rev. 2004, 104, 2099–2123. 10.1021/cr020689l. [DOI] [PubMed] [Google Scholar]
- Lesser G. J.; Rose G. D. Hydrophobicity of Amino Acid Subgroups in Proteins. Proteins Struct. Funct. Genet. 1990, 8, 6–13. 10.1002/prot.340080104. [DOI] [PubMed] [Google Scholar]
- Phillips G. N. Jr.; Montgomery Pettitt B. Structure and Dynamics of the Water around Myoglobin. Protein Sci. 1995, 4, 149–158. 10.1002/pro.5560040202. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nick Pace C.; Treviño S.; Prabhakaran E.; Scholtz J. M. Protein Structure, Stability and Solubility in Water and Other Solvents. Philos. Trans. R. Soc. London B 2004, 359, 1225–1235. 10.1098/rstb.2004.1500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith J. C.; Merzel F.; Bondar A.-N.; Tournier A.; Fischer S. Structure, Dynamics, and Reactions of Protein Hydration Water. Philos. Trans. R. Soc. London B 2004, 359, 1181–1190. 10.1098/rstb.2004.1497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bhattacharyya K. Nature of biological Water: A Femtosecond Study. Chem. Commun. 2008, 2848–2857. 10.1039/b800278a. [DOI] [PubMed] [Google Scholar]
- Zhou R.; Huang X.; Margulis C. J.; Berne B. J. Hydrophobic Collapse in Multi-domain Protein Folding. Science 2004, 305, 1605–1609. 10.1126/science.1101176. [DOI] [PubMed] [Google Scholar]
- Hong S.; Kim D. Interaction between Bound Water Molecules and Local Protein Structures: A Statistical Analysis of the Hydrogen Bond Structures Around Bound Water Molecules. Proteins 2016, 84, 43–51. 10.1002/prot.24953. [DOI] [PubMed] [Google Scholar]
- D’Aquino J. A.; Gomez J.; Hilser V. J.; Lee K. H.; Amzel L. M.; Freire E. The Magnitude of the Backbone Conformational Entropy Change in Protein Folding. Proteins Struct. Funct. Genet. 1996, 25, 143–156. 10.1002/prot.1. [DOI] [PubMed] [Google Scholar]
- Chong S.-H.; Ham S. Distinct Role of Hydration Water in Protein Misfolding and Aggregation Revealed by Fluctuating Thermodynamics Analysis. Acc. Chem. Res. 2015, 48, 956–965. 10.1021/acs.accounts.5b00032. [DOI] [PubMed] [Google Scholar]
- Gupta M. N. Enzyme Function in Organic Solvents. Eur. J. Biochem. 1992, 203, 25–32. 10.1111/j.1432-1033.1992.tb19823.x. [DOI] [PubMed] [Google Scholar]
- Klibanov A. M. Improving Enzymes by using them in Organic Solvents. Nature 2001, 409, 241–246. 10.1038/35051719. [DOI] [PubMed] [Google Scholar]
- Rupley J. A.; Gratton E.; Careri G. Water and Globular Proteins. Trends Biochem. Sci. 1983, 8, 18–22. 10.1016/0968-0004(83)90063-4. [DOI] [Google Scholar]
- Zaks A.; Klibanov A. M. Enzymatic Catalysis in Nonaqueous Solvent. J. Biol. Chem. 1988, 263, 3194–3201. [PubMed] [Google Scholar]
- Braco L.; Dabulis K.; Klibanov A. M. Production of Abiotic Receptors by Molecular Imprinting of Proteins. Proc. Natl. Acad. Sci. U.S.A. 1990, 87, 274–277. 10.1073/pnas.87.1.274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stahl M.; Mansson M.; Mosbach K. The Synthesis of a D-amino Acid Eester in an Organic Media with α-Chymotrypsin Modified by a Bio-Imprinting Procedure. Biotechnol. Lett. 1990, 12, 161–166. 10.1007/BF01026792. [DOI] [Google Scholar]
- Serdakowski A. L.; Dordick J. S. Enzyme Activation for Organic Solvents Made Easy. Trends Biotechnol. 2008, 26, 48–54. 10.1016/j.tibtech.2007.10.007. [DOI] [PubMed] [Google Scholar]
- Khmelnitsky Y. L.; Welch S. H.; Clark D. S.; Dordick J. S. Salts Dramatically Enhance Activity of Enzymes Suspended in Organic Solvents. J. Am. Chem. Soc. 1994, 116, 2647–2648. 10.1021/ja00085a066. [DOI] [Google Scholar]
- Lindsay J. P.; Clark D. S.; Dordick J. S. Combinatorial Formulation of Biocatalyst Preparations for Increased Activity in Organic Solvents: Salt Activation of Penicillin Aamidase. Biotechnol. Bioeng. 2004, 85, 553–560. 10.1002/bit.20002. [DOI] [PubMed] [Google Scholar]
- Doukyu N.; Ogino H. Organic Solvent-Tolerant Enzymes. Biochem. Eng. J. 2010, 48, 270–282. 10.1016/j.bej.2009.09.009. [DOI] [Google Scholar]
- Stepankova V.; Bidmanova S.; Koudelakova T.; Prokop Z.; Chaloupkova R.; Damborsky J. Strategies for Stabilization of Enzymes in Organic Solvents. ACS Catal. 2013, 3, 2823–2836. 10.1021/cs400684x. [DOI] [Google Scholar]
- Zhang J.Protein-Protein Interactions in Salt Solutions. In Protein-Protein Interactions - Computational and Experimental Tools, Cai W., Ed.; InTech, 2012, ISBN: 978-953-51-0397-4, . [Google Scholar]
- Kunz W., Ed.; Specific Ion Effects; World Scientific Publishing Co.: Singapore, 2010. [Google Scholar]
- Zangi R. Can Salting-In/Salting-Out Ions Be Classified as Chaotropes/Kosmotropes?. J. Phys. Chem. B 2010, 114, 643–650. 10.1021/jp909034c. [DOI] [PubMed] [Google Scholar]
- Stradner A.; Schurtenberger P. Potential and Limits of a Colloid Approach to Protein Solutions. Soft Matter 2020, 16, 307–323. 10.1039/C9SM01953G. [DOI] [PubMed] [Google Scholar]
- Lu H.; Zhou Q.; He J.; Jiang Z.; Peng C.; Tong R.; Shi J. Recent Advances in The Development of Protein-Protein Interactions Modulators: Mechanisms and Clinical Trials. Signal Transduct. Target Ther. 2020, 5, 213. 10.1038/s41392-020-00315-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rembert K. B.; Paterová J.; Heyda J.; Hilty C.; Jungwirth P.; Cremer P. S. Molecular Mechanisms of Ion-Specific Effects on Proteins. J. Am. Chem. Soc. 2012, 134, 10039–10046. 10.1021/ja301297g. [DOI] [PubMed] [Google Scholar]
- Janc T.; Lukšič M.; Vlachy V.; Rigaud B.; Rollet A.-L.; Korb J.-P.; Mériguet G.; Malikova N. Ion-Specificity and Surface Water Dynamics in Protein Solutions. Phys. Chem. Chem. Phys. 2018, 20, 30340–30350. 10.1039/C8CP06061D. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Finet S.; Vivares D.; Bonnete F.; Tardieu A. Controlling Biomolecular Crystallization by Understanding the Distinct Effects of PEGs and Salts on Solubility. Methods Enzymol. 2003, 368, 105–129. 10.1016/S0076-6879(03)68007-9. [DOI] [PubMed] [Google Scholar]
- Nihonyanagi S.; Yamaguchi S.; Tahara T. Counterion Effect on Interfacial Water at Charged Interfaces and Its Relevance to the Hofmeister Series. J. Am. Chem. Soc. 2014, 136, 6155–6158. 10.1021/ja412952y. [DOI] [PubMed] [Google Scholar]
- Jungwirth P.; Winter B. Ions at aqueous interfaces: From Water Surface to Hydrated Proteins. Annu. Rev. Phys. Chem. 2008, 59, 343–366. 10.1146/annurev.physchem.59.032607.093749. [DOI] [PubMed] [Google Scholar]
- Sarangapani P. S.; Hudson S. D.; Jones R. L.; Douglas J. F.; Pathak J. A. Critical Examination of the Colloidal Particle Model of Globular Proteins. Biophys. J. 2015, 108, 724–737. 10.1016/j.bpj.2014.11.3483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan Y.-H.; Luo R. Protein Stability Prediction: A Poisson-Boltzmann Approach. J. Phys. Chem. B 2008, 112, 1875–1883. 10.1021/jp709660v. [DOI] [PubMed] [Google Scholar]
- de los Rios M. A.; Plaxco K. W. Apparent Debye-Hückel Electrostatic Effects in The Folding of a Simple, Single Domain Protein. Biochemistry 2005, 44, 1243–1250. 10.1021/bi048444l. [DOI] [PubMed] [Google Scholar]
- Zhou H.-X. Interactions of Macromolecules with Salt Ions: An Electrostatic Theory for the Hofmeister Effect. Proteins 2005, 61, 69–78. 10.1002/prot.20500. [DOI] [PubMed] [Google Scholar]
- Molina-Bolívar J. A.; Ortega-Vinuesa J. L. How Proteins Stabilize Colloidal Particles by Means of Hydration Forces. Langmuir 1999, 15, 2644–2653. 10.1021/la981445s. [DOI] [Google Scholar]
- Bostrom M.; Williams D. R. M.; Ninham B. W. Specific Ion Effects: Why DLVO Theory Fails for Biology and Colloid Systems. Phys. Rev. Lett. 2001, 87, 168103. 10.1103/PhysRevLett.87.168103. [DOI] [PubMed] [Google Scholar]
- Ianeselli L.; Zhang F.; Skoda M. W. A.; Jacobs R. M. J.; Martin R. A.; Callow S.; Prevost S.; Schreiber F. Protein-Protein Interactions in Ovalbumin Solutions Studied by Small-Angle Scattering: Effect of Ionic Strength and the Chemical Nature of Cations. J. Phys. Chem. B 2010, 114, 3776–3783. 10.1021/jp9112156. [DOI] [PubMed] [Google Scholar]
- Zhang I. L. F.; Skoda M. W. A.; Jacobs R. M. J.; Zorn S.; Martin R. A.; Martin C. M.; Clark G. F.; Weggler S.; Hildebrandt A.; Kohlbacher O.; Schreiber F. Reentrant Condensation of Proteins in Solution Induced by Multivalent Counterions. Phys. Rev. Lett. 2008, 101, 148101. 10.1103/PhysRevLett.101.148101. [DOI] [PubMed] [Google Scholar]
- Petsev D. N.; Thomas B. R.; Yau S. T.; Vekilov P. G. Interactions and Aggregation of Apoferritin Molecules in Solution: Effects of Added Electrolytes. Biophys. J. 2000, 78, 2060–2069. 10.1016/S0006-3495(00)76753-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Retailleau P.; Ries-Kautt M.; Ducruix A. 1997. No Salting-In of Lysozyme Chloride Observed at Low Ionic Strength Over a Large Range of pH. Biophys. J. 1997, 73, 2156–2163. 10.1016/S0006-3495(97)78246-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grigsby J. J.; Blanch H. W.; Prausnitz J. M. Cloud-Point Temperatures for Lysozyme in Electrolyte Solutions: Effect of Salt Type, Salt Concentration, and pH. Biophys. Chem. 2001, 91, 231–243. 10.1016/S0301-4622(01)00173-9. [DOI] [PubMed] [Google Scholar]
- Zbacnik T. J.; Holcomb R. E.; Katayama D. S.; Murphy B. M.; Payne R. W.; Coccaro R. C.; Evans G. J.; Matsuura J. E.; Henry C. S.; Manning M. C. Role of Buffers in Protein Formulations. J. Pharm. Sci. 2017, 106, 713–733. 10.1016/j.xphs.2016.11.014. [DOI] [PubMed] [Google Scholar]
- Brudar S.; Hribar-Lee B. Effect of Buffer on Protein Stability in Aqueous Solutions: A Simple Protein Aggregation Model. J. Phys. Chem. B 2021, 125, 2504–2512. 10.1021/acs.jpcb.0c10339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yancey P. H.; Clark M. E.; Hand S. C.; Bowlus R. D.; Somero G. N. Living with Water Stress: Evolution of Osmolyte Systems. Science 1982, 217, 1214–1222. 10.1126/science.7112124. [DOI] [PubMed] [Google Scholar]
- Burg M. B.; Peters E. M.; Bohren K. M.; Gabbay K. H. Factors Affecting Counteraction by Methylamines of Urea Effects. Proc. Natl. Acad. Sci. U. S. A. 1999, 96, 6517–6522. 10.1073/pnas.96.11.6517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lanyi J. K. Salt-Dependent Properties of Proteins from Extremely Halophilic Bacteria. Bacteriol. Rev. 1974, 38, 272–290. 10.1128/br.38.3.272-290.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takano K.; Saito M.; Morikawa M.; Kanaya S. Mutational and Structural-Based Analyses of the Osmolyte Effect on Protein Stability. J. Biochem. 2004, 135, 701–708. 10.1093/jb/mvh085. [DOI] [PubMed] [Google Scholar]
- Bolen D. W.; Baskakov I. V. The Osmophobic effect: Natural Selection of a Thermodynamic Force in Protein Folding. J. Mol. Biol. 2001, 310, 955–963. 10.1006/jmbi.2001.4819. [DOI] [PubMed] [Google Scholar]
- Bolen D. W. Effects of Naturally Occurring Osmolytes on Protein Stability and Solubility: Issues Important in Protein Crystallization. Methods 2004, 34, 312–322. 10.1016/j.ymeth.2004.03.022. [DOI] [PubMed] [Google Scholar]
- Street T. O.; Bolen D. W.; Rose G. D. A Molecular Mechanism for Osmolyte-Induced Protein Stability. Proc. Nat. Aca. Sci. USA 2006, 103, 13997–14002. 10.1073/pnas.0606236103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Auton M.; Rösgen J.; Sinev M.; Holthauzen L. M. F.; Bolen D. W. Osmolyte Effects on Protein Stability and Solubility: A Balancing act between Backbone and Sidechains. Biophys. Chem. 2011, 159, 90–99. 10.1016/j.bpc.2011.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barnett G. V.; Razinkov V. I.; Kerwin B. A.; Blake S.; Qi W.; Curtis R. A.; Roberts C. J. Osmolyte Effects on Monoclonal Antibody Stability and Concentration-Dependent Protein Interactions with Water and Common Osmolytes. J. Phys. Chem. B 2016, 120, 3318–3330. 10.1021/acs.jpcb.6b00621. [DOI] [PubMed] [Google Scholar]
- Choudhary S.; Kishore N.; Hosur R. V. Inhibition of Insulin Fibrillation by Osmolytes: Mechanistic Insights. Sci. Rep. 2015, 5, 17599. 10.1038/srep17599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruździak P.; Panuszko A.; Jourdan M.; Stangret J. Protein Thermal Stabilization in Aqueous Solutions of Osmolytes. Acta Biochim. Polym. 2016, 63, 65–70. 10.18388/abp.2014_950. [DOI] [PubMed] [Google Scholar]
- Dukhin S. S.; Kretzschmar G.; Miller R.. Dynamics of Adsorption at Liquid Interfaces; Elsevier: Amsterdam, 1995. [Google Scholar]
- Rao K. S.; So S.; Kumar A. Vesicle and Reverse Vesicles of Ionic Liquids in Ionic Liquids. Chem. Commun. 2013, 49, 8111–8113. 10.1039/c3cc44462g. [DOI] [PubMed] [Google Scholar]
- Tian M.; Zhu L.; Yu D.; Wang Y.; Sun S.; Wang Y. Aggregate Transitions in Mixtures of Anionic Sulfonate Gemini, Surfactant with Cationic Ammonium Single-Chain Surfactant. J. Phys. Chem. B 2013, 117, 433–440. 10.1021/jp310065y. [DOI] [PubMed] [Google Scholar]
- Jones M. N. Surfactant Interactions with Biomembranes and Proteins. Chem. Soc. Rev. 1992, 21, 127–136. 10.1039/cs9922100127. [DOI] [Google Scholar]
- Otzen D. Protein-Surfactant Interactions: A Tale of Many States. Biochim. Biophys. Acta 2011, 1814, 562–591. 10.1016/j.bbapap.2011.03.003. [DOI] [PubMed] [Google Scholar]
- Tanford C.; Nozaki Y.; Reynolds J. A.; Makino S. Molecular Characterization of Proteins in Detergent Solutions. Biochemistry 1974, 13, 2369–2376. 10.1021/bi00708a021. [DOI] [PubMed] [Google Scholar]
- Lye G. J.; Asenjo J. A.; Pyle D. L. Protein Extraction Using Reverse Micelles: Kinetics of Protein Partitioning. Chem. Eng. Sci. 1994, 49, 3195–3204. 10.1016/0009-2509(94)00147-2. [DOI] [Google Scholar]
- Saitoh T.; Tani H.; Kamidate T.; Watanabe H. Phase Separation in Aqueous Micellar Solutions of Nonionic Surfactants for Protein Separation. Trends Anal. Chem. 1995, 14, 213–217. 10.1016/0165-9936(95)91372-Y. [DOI] [Google Scholar]
- Liu C. L.; Kamei D. T.; King J. A.; Wang D. I. C.; Blankschtein D. J. Separation of Proteins and Viruses Using Two-Phase Aqueous Micellar Systems. Chromatogr. B Biomed. Sci. Appl. 1998, 711, 127–138. 10.1016/S0378-4347(98)00013-9. [DOI] [PubMed] [Google Scholar]
- McClements D. J.Food Emulsions: Principles, Practice and Techniques, 2nd ed.; CRC Press: Boca Raton, FL, 2004. [Google Scholar]
- Anson M. L. The Denaturation of Proteins by Synthetic Detergents and Bile Salts. J. Gen. Physiol. 1939, 23, 239. 10.1085/jgp.23.2.239. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Putnam F. W. The Interactions of Proteins and Synthetic Detergents. Adv. Prot. Chem. 1948, 4, 79–122. 10.1016/S0065-3233(08)60005-4. [DOI] [PubMed] [Google Scholar]
- Steinhardt J.; Reynolds J. A.. Multiple Equilibria in Proteins; Academic Press: New York, 1969. [Google Scholar]
- Tanford C. Protein denaturation. Part C. Theoretical Models for the Mechanism of Denaturation. Adv. Prot. Chem. 1970, 24, 1–95. 10.1016/S0065-3233(08)60241-7. [DOI] [PubMed] [Google Scholar]
- Yonath A.; Podjarny A.; Honig B.; Sielecki A.; Traub W. Crystallographic Studies of Protein Denaturation and Renaturation. 2. Sodium Dodecyl Sulfate Induced Structural Changes in Triclinic Lysozyme. Biochemistry 1977, 16, 1418–1424. 10.1021/bi00626a028. [DOI] [PubMed] [Google Scholar]
- Jones M. N.Protein-Surfactant Interactions. In Surface Activity of Proteins; Magdassi S., Ed.; Marcel Dekker: NY, 1996; pp 237–284. [Google Scholar]
- Andersen K. K.; Oliveira C. L. P.; Larsen K. L.; Poulsen F. M.; Callisen T. H.; Westh P.; Pedersen J. S.; Otzen D. E. The Role of Decorated SDS Micelles in Sub-CMC Protein Denaturation and Association. J. Mol. Biol. 2009, 391, 207–226. 10.1016/j.jmb.2009.06.019. [DOI] [PubMed] [Google Scholar]
- Svergun D. I. Small-angle X-ray and Neutron Scattering as a Tool for Structural Systems Biology. Biol. Chem. 2010, 391, 737–743. 10.1515/bc.2010.093. [DOI] [PubMed] [Google Scholar]
- Mehan S.; Aswal V. K.; Kohlbrecher J. Tuning of Protein-Surfactant Interactions to Modify Resultant Structure. Phys. Rev. E 2015, 92, 032713. 10.1103/PhysRevE.92.032713. [DOI] [PubMed] [Google Scholar]
- Gull N.; Sen P.; Khan R. H.; Din K. Spectroscopic Studies on the Comparative Interaction of Cationic Single-Chain and Gemini Surfactants with Human Serum Albumin. J. Biochem. 2008, 145, 67–77. 10.1093/jb/mvn141. [DOI] [PubMed] [Google Scholar]
- Gelamo E. L.; Tabak M. Spectroscopic Studies on the Interaction of Bovine (BSA) and Human (HSA) Serum Albumins with Ionic Surfactants. Spectrochim. Acta Part A 2000, 56, 2255–2271. 10.1016/S1386-1425(00)00313-9. [DOI] [PubMed] [Google Scholar]
- Mogensen J. E.; Sehgal P.; Otzen D. E. Activation, Inhibition and Destabilization of Thermomyces lanuginosus lipase by Detergents. Biochemistry 2005, 44, 1719–1730. 10.1021/bi0479757. [DOI] [PubMed] [Google Scholar]
- Sehgal P.; Nielsen S. B.; Pedersen S.; Wimmer R.; Otzen D. E. Modulation of Cutinase Structure and Stability by Phospholipid Detergents. Biochim. Biophys. Acta 2007, 1774, 1544–1554. 10.1016/j.bbapap.2007.09.011. [DOI] [PubMed] [Google Scholar]
- Faustino C. M. C.; Calado A. R. T.; Garcia-Rio L. Gemini Surfactants Protein interactions, Effect of Temperature, pH, and Surfactant Stereochemistry. Biomacromolecules 2009, 10, 2508–2514. 10.1021/bm9004723. [DOI] [PubMed] [Google Scholar]
- Branco M. A.; Pinheiro L.; Faustino C. Amino Acid based Cationic Gemini Surfactant-Protein Interactions. Colloids, Surf. A Physico. Eng. Asp. 2015, 480, 105–112. 10.1016/j.colsurfa.2014.12.022. [DOI] [Google Scholar]
- Mahler H.-C.; Mueller R.; et al. Induction and Analysis of Aggregates in a Liquid IgG1-Antibody Formulation. Eur. J. Pharma. Biopharm. 2005, 59, 407–417. 10.1016/j.ejpb.2004.12.004. [DOI] [PubMed] [Google Scholar]
- Frøkjaer S.; Otzen D. Protein Drug Stability: A Formulation Challenge. Nat. Rev. Drug Discovery 2005, 4, 298–306. 10.1038/nrd1695. [DOI] [PubMed] [Google Scholar]
- Levine H. L.; Ransohoff T. C. The Use of Surface Tension Measurements in the Design of Antibody-Based Product Formulations. J. Parent. Sci. Techn. 1991, 45, 160–165. [PubMed] [Google Scholar]
- Wang W. Instability, Stabilization, and Formulation of Liquid Protein Pharmaceuticals. Int. J. Pharm. 1999, 185, 129–188. 10.1016/S0378-5173(99)00152-0. [DOI] [PubMed] [Google Scholar]
- Wang W.; Singh S.; et al. Antibody Structure, Instability, and Formulation. J. Pharm. Sci. 2007, 96, 1–26. 10.1002/jps.20727. [DOI] [PubMed] [Google Scholar]
- Wang W.; Wang Y. J.; et al. Dual Effects of Tween 80 on Protein Stability. Int. J. Pharm. 2008, 347, 31–38. 10.1016/j.ijpharm.2007.06.042. [DOI] [PubMed] [Google Scholar]
- Chang B. S.; Kendrick B. S.; et al. Surface-Induced Denaturation of Proteins during Freezing, and Its Inhibition by Surfactants. J. Pharma. Sci. 1996, 85, 1325–1330. 10.1021/js960080y. [DOI] [PubMed] [Google Scholar]
- Chou D. K.; Krishnamurthy R.; et al. Effects of Tween 20, and Tween 80 on the Stability of Albutropin during Agitation. J. Pharma. Sci. 2005, 94, 1368–1381. 10.1002/jps.20365. [DOI] [PubMed] [Google Scholar]
- Katakam M.; Bell L. N.; et al. Effect of Surfactants on the Physical Stability of Recombinant Human Growth Hormone. J. Pharm. Sci. 1995, 84, 713–716. 10.1002/jps.2600840609. [DOI] [PubMed] [Google Scholar]
- Lee H. J.; McAuley A.; Schilke K. F.; McGuire J. Molecular Origins of Surfactant-Mediated Stabilization of Protein Drugs. Adv. Drug Delivery Rev. 2011, 63, 1160–1171. 10.1016/j.addr.2011.06.015. [DOI] [PubMed] [Google Scholar]
- Khan T. A.; Mahler H.-C.; Kishore R. S. K. Key Interactions of Surfactants in Therapeutic Protein Formulations: A Review. Eur. J. Pharma. Biopharma. 2015, 97, 60–67. 10.1016/j.ejpb.2015.09.016. [DOI] [PubMed] [Google Scholar]
- Duncan R. The Dawning era of Polymer Therapeutics. Nat. Rev. Drug Discovery 2003, 2, 347–360. 10.1038/nrd1088. [DOI] [PubMed] [Google Scholar]
- Canalle L. A.; Lowik D. W. P. M.; van Hest J. C. M. Polypeptide-Polymer Bioconjugates. Chem. Soc. Rev. 2010, 39, 329–353. 10.1039/B807871H. [DOI] [PubMed] [Google Scholar]
- Abuchowski A.; Vanes T.; Palczuk N. C.; Davis F. F. Alteration of Immunological Properties of Bovine Serum Albumin by Covalent Attachment of Polyethylene Glycol. J. Biol. Chem. 1977, 252, 3578–3581. 10.1016/S0021-9258(17)40291-2. [DOI] [PubMed] [Google Scholar]
- Abuchowski A.; McCoy J. R.; Palczuk N. C.; Vanes T.; Davis F. F. Effect of Covalent Attachment of Polyethylene Glycol on Immunogenicity and Circulating Life of Bovine Liver Catalase. J. Biol. Chem. 1977, 252, 3582–3586. 10.1016/S0021-9258(17)40292-4. [DOI] [PubMed] [Google Scholar]
- Alconcel S. N. S.; Baas A. S.; Maynard H. D. FDA-Approved Poly(ethylene glycol)-Protein Conjugate Drugs. Polym. Chem. 2011, 2, 1442–1448. 10.1039/c1py00034a. [DOI] [Google Scholar]
- FDA Approved PEGylated Drugs 2020, Biochempeg, https://www.biochempeg.com/article/58.html.
- Norman M. E.; Williams P.; Illum L. Human serum Albumin as a Probe for Surface Conditioning (Opsonization) of Block Copolymer-Coated Microspheres. Biomaterials 1992, 13, 841–849. 10.1016/0142-9612(92)90177-P. [DOI] [PubMed] [Google Scholar]
- Carrstensen H.; Muller R. H.; Muller B. W. Particle Size, Surface Hydrophobicity and Interaction with Serum of Parenteral Fat Emulsions and Model Drug Carriers as Parameters Related to RES Uptake. Clin. Nutr. 1992, 11, 289–297. 10.1016/0261-5614(92)90006-C. [DOI] [PubMed] [Google Scholar]
- Roser M.; Fischer D.; Kissel T. Surface-Modified Biodegradable Albumin Nano- and Microspheres. II: Effect of Surface Charges on in Vitro Phagocytosis and Biodistribution in Rats. Eur. J. Pharm. Biopharm. 1998, 46, 255–263. 10.1016/S0939-6411(98)00038-1. [DOI] [PubMed] [Google Scholar]
- Pelegri-O’Day E. M.; Lin E.-W.; Maynard H. D. Therapeutic Protein-Polymer Conjugates: Advancing Beyond PEGylation. J. Am. Chem. Soc. 2014, 136, 14323–14332. 10.1021/ja504390x. [DOI] [PubMed] [Google Scholar]
- Richter A. W.; Akerblom E. Polyethylene Glycol Reactive Antibodies in Man: Titer Distribution in Allergic Patients Treated with Monomethoxy Polyethylene Glycol Modified Allergens or Placebo, and in Healthy Blood Donors. Int. Arch. Allergy Appl. Immunol. 2004, 74, 36–39. 10.1159/000233512. [DOI] [PubMed] [Google Scholar]
- Liu Y.; Reidler H.; Pan J.; Milunic D.; Qin D.; Chen D.; Vallejo Y. R.; Yin R. A Double Antigen Bridging Immunogenicity ELISA for the Detection of Antibodies to Polyethylene Glycol Polymers. J. Pharmacol. Toxicol. Methods 2011, 64, 238–245. 10.1016/j.vascn.2011.07.003. [DOI] [PubMed] [Google Scholar]
- Fisher T. C.; Armstrong J. K.; Wenby R. B.; Meiselman H. J.; Leger R.; Garratty G. Isolation and Identification of a Human Antibody to Poly(Ethylene Glycol). Blood 2003, 102, 559A. [Google Scholar]
- Schellekens H.; Hennink W. E.; Brinks V. The Immunogenicity of Polyethylene Glycol: Facts and Fiction. Pharm. Res. 2013, 30, 1729–1734. 10.1007/s11095-013-1067-7. [DOI] [PubMed] [Google Scholar]
- Zhao W.; Liu F.; Chen Y.; Bai J.; Gao W. Synthesis of Well-Defined Protein- Polymer Conjugates for Biomedicine. Polymer 2015, 66, A1–A10. 10.1016/j.polymer.2015.03.054. [DOI] [Google Scholar]
- Mecerreyes D. Polymeric Ionic Liquids: Broadening the Properties and Applications of Polyelectrolytes. Prog. Polym. Sci. 2011, 36, 1629–1648. 10.1016/j.progpolymsci.2011.05.007. [DOI] [Google Scholar]
- van Rantwijk F.; Secundo F.; Sheldon R. A. Structure and Activity of Candida antarctica lipase B in Ionic Liquids. Green Chem. 2006, 8, 282–286. 10.1039/B513062J. [DOI] [Google Scholar]
- Shim Y.; Kim H. J.; Jung Y. J. Solvation of a Small Metal-Binding Peptide in Room-Temperature Ionic Liquids. Bull. Korean Chem. Soc. 2012, 33, 3601–3606. 10.5012/bkcs.2012.33.11.3601. [DOI] [Google Scholar]
- Bermejo M. D.; Kotlewska A. J.; Florusse L. J.; Cocero M. J.; van Rantwijk F.; Peters C. J. Influence of the Enzyme Concentration on the Phase Behavior for Developing a Homogeneous Enzymatic Reaction in Ionic Liquid-CO2 Media. Green Chem. 2008, 10, 1049–1054. 10.1039/b805011b. [DOI] [Google Scholar]
- Ou G.; He B.; Halling P. Ionization Basis for Activation of Enzymes Soluble in Ionic Liquids. Biochim. Biophys. Acta 2016, 1860, 1404–1408. 10.1016/j.bbagen.2016.04.004. [DOI] [PubMed] [Google Scholar]
- Shimojo K.; Kamiya N.; Tani F.; Naganawa H.; Naruta Y.; Goto M. Extractive Solubilization, Structural Change, and Functional Conversion of Cytochrome c in Ionic Liquids via Crown Ether Complexation. Anal. Chem. 2006, 78, 7735–7742. 10.1021/ac0612877. [DOI] [PubMed] [Google Scholar]
- Phillips D. M.; Drummy L. F.; Conrady D. G.; Fox D. M.; Naik R. R.; Stone M. O.; Trulove P. C.; De Long H. C.; Mantz R. A. Dissolution and Regeneration of Bombyx mori Silk Fibroin Using Ionic Liquids. J. Am. Chem. Soc. 2004, 126, 14350–14351. 10.1021/ja046079f. [DOI] [PubMed] [Google Scholar]
- Chen J.; Vongsanga K.; Wang X.; Byrne N. What Happens During Natural Protein Fiber Dissolution in Ionic Liquids?. Materials 2014, 7, 6158–6168. 10.3390/ma7096158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biswas A.; Shogren R. L.; Stevenson D. G.; Willett J. L.; Bhowmik P. K. Ionic Liquids as Solvents for Biopolymers: Acylation of Starch and Zein Protein. Carbohy. Polym. 2006, 66, 546–550. 10.1016/j.carbpol.2006.04.005. [DOI] [Google Scholar]
- Strassburg S.; Bermudez H.; Hoagland D. Lysozyme Solubility and Conformation in Neat Ionic Liquids and Their Mixtures with Water. Biomacromolecules 2016, 17, 2233–2239. 10.1021/acs.biomac.6b00468. [DOI] [PubMed] [Google Scholar]
- J Mehta M.; Bharmoria P.; Bhayani K.; Kumar A. Gelatin Solubility and Processing in Ionic Liquids: An Approach Towards Waste to Utilization. Chem. Select 2017, 2, 9895–9900. 10.1002/slct.201702015. [DOI] [Google Scholar]
- Banerjee A.; Ibsen K.; Brown T.; Chen R.; Agatemor C.; Mitragotri S. Ionic Liquids for Oral Insulin Delivery. Proc. Natl. Acad. Sci. U. S. A. 2018, 115, 7296–7301. 10.1073/pnas.1722338115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spange S.; Lungwitz R.; Schade A. Correlation of Molecular Structure and Polarity of Ionic Liquids. J. Mol. Liq. 2014, 192, 137–143. 10.1016/j.molliq.2013.06.016. [DOI] [Google Scholar]
- Zeindlhofer V.; Schroder C. Computational Solvation Analysis of Biomolecules in Aqueous Ionic Liquid Mixtures. Biophysical Reviews 2018, 10, 825–840. 10.1007/s12551-018-0416-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burney P. R.; Pfaendtner J. Structural and Dynamic Features of Candida rugosa lipase 1 in Water, Octane, Toluene, and Ionic Liquids BMIM-PF6 and BMIM-NO3. J. Phys. Chem. B 2013, 117, 2662–2670. 10.1021/jp312299d. [DOI] [PubMed] [Google Scholar]
- Klähn M.; Lim S. G.; Wu P. How Ion Properties Determine the Stability of a Lipase Enzyme in Ionic Liquids: A Molecular Dynamics Study. Phys. Chem. Chem. Phys. 2011, 13, 18647–18660. 10.1039/c1cp22056j. [DOI] [PubMed] [Google Scholar]
- Doherty T. V.; Mora-Pale M.; Foley S. E.; Linhardt R. J.; Dordick J. S. Ionic Liquid Solvent Properties as Predictors of Lignocellulose Pretreatment Efficacy. Green Chem. 2010, 12, 1967–1975. 10.1039/c0gc00206b. [DOI] [Google Scholar]
- Palgunadi J.; Hong S. Y.; Lee J. K.; Lee H.; Lee S. D.; Cheong M.; Kim H. S. Correlation Between Hydrogen Bond Basicity and Acetylene Solubility in Room Temperature Ionic Liquids. J. Phys. Chem. B 2011, 115, 1067–1074. 10.1021/jp108351f. [DOI] [PubMed] [Google Scholar]
- Dhumal N. R.; Kim H. J.; Kiefer J. Electronic Structure and Normal Vibrations of the 1-Ethyl-3-methylimidazolium Ethyl Sulfate Ion Pair. J. Phys. Chem. A 2011, 115, 3551–3558. 10.1021/jp1122322. [DOI] [PubMed] [Google Scholar]
- Falcioni F.; Housden H. R.; Ling Z.; Shimizu S.; Walker A. J.; Bruce N. C. Soluble, Folded and Active Subtilisin in a Protic Ionic Liquid. Chem. Commun. 2010, 46, 749–751. 10.1039/B916497A. [DOI] [PubMed] [Google Scholar]
- Attri P.; Venkatesu P.; Kumar A.; Byrne N. A Protic Ionic Liquid Attenuates the Deleterious Actions of Urea on α-chymotrypsin. Phys. Chem. Chem. Phys. 2011, 13, 17023–17026. 10.1039/c1cp22195g. [DOI] [PubMed] [Google Scholar]
- Attri P.; Venkatesu P. Thermodynamic Characterization of the Biocompatible Ionic Liquid Effects on Protein Model Compounds and Their Functional Groups. Phys. Chem. Chem. Phys. 2011, 13, 6566–6575. 10.1039/c0cp02768e. [DOI] [PubMed] [Google Scholar]
- Vasantha T.; Attri P.; Venkatesu P.; Devi R. S. R. Structural Basis for the Enhanced Stability of Protein Model Compounds and Peptide Backbone Unit in Ammonium Ionic Liquids. J. Phys. Chem. B 2012, 116, 11968–11978. 10.1021/jp308443f. [DOI] [PubMed] [Google Scholar]
- Attri P.; Venkatesu P. Ammonium Ionic Liquids as Convenient Co-solvents for the Structure and Stability of Succinylated Con A. J. Chem. Thermodynamics 2012, 52, 78–88. 10.1016/j.jct.2012.02.013. [DOI] [Google Scholar]
- Attri P.; Venkatesu P. Influence of Protic ionic Liquids on the Structure and Stability of Succinylated Con A. Int. J. Biol. Macromol. 2012, 51, 119–128. 10.1016/j.ijbiomac.2012.04.017. [DOI] [PubMed] [Google Scholar]
- Fujita K.; Forsyth M.; MacFarlane D. R.; Reid R. W.; Elliott G. D. Unexpected Improvement in Stability and Utility of Cytochrome c by Solution in Biocompatible Ionic Liquids. Biotechnol. Bioeng. 2006, 94, 1209–1213. 10.1002/bit.20928. [DOI] [PubMed] [Google Scholar]
- Mazid R. R.; Vijayaraghavan R.; MacFarlane D. R.; Cortez-Jugo C.; Cheng W. Inhibited Fragmentation of mAbs in Buffered Ionic Liquids. Chem. Commun. 2015, 51, 8089–8092. 10.1039/C5CC01877C. [DOI] [PubMed] [Google Scholar]
- Fujita K.; Nakamura N.; Igarashi K.; Samejima M.; Ohno H. Biocatalytic Oxidation of Cellobiose in a Hydrated Ionic Liquid. Green Chem. 2009, 11, 351–354. 10.1039/B813529K. [DOI] [Google Scholar]
- Weaver K. D.; Vrikkis R. M.; Vorst M. P. V.; Trullinger J.; Vijayaraghavan R.; Foureau D. M.; McKillop I. H.; MacFarlane D. R.; Krueger J. K.; Elliott G. D. Structure and Function of Proteins in Hydrated Choline Dihydrogen Phosphate Ionic Liquid. Phys. Chem. Chem. Phys. 2012, 14, 790–801. 10.1039/C1CP22965F. [DOI] [PubMed] [Google Scholar]
- Dhiman D.; Bisht M.; Tavares A. P. M.; Freire M. G.; Venkatesu P. Cholinium-Based Ionic Liquids as Efficient Media for Improving the Structural and Thermal Stability of Immunoglobulin G Antibodies. ACS Sustain. Chem. Eng. 2022, 10, 5404–5420. 10.1021/acssuschemeng.1c07979. [DOI] [Google Scholar]
- Kumar A.; Venkatesu P. Prevention of Insulin Self-aggregation by a Protic Ionic Liquid. RSC Adv. 2013, 3, 362–367. 10.1039/C2RA22277A. [DOI] [Google Scholar]
- Papadopoulou A. A.; Tzani A.; Alivertis D.; Katsoura M. H.; Polydera A. C.; Detsi A.; Stamatis H. Hydroxyl Ammonium Ionic Liquids as Media for Biocatalytic Oxidations. Green Chem. 2016, 18, 1147–1158. 10.1039/C5GC02381E. [DOI] [Google Scholar]
- Bhakuni K.; Bisht M.; Venkatesu P.; Mondal D. Designing Biological Fluid Inspired Molecularly Crowded Ionic Liquid Media as A Sustainable Packaging Platform for Cytochrome C. Chem. Commun. 2019, 55, 5747–5750. 10.1039/C9CC02340B. [DOI] [PubMed] [Google Scholar]
- Matias S. C.; Lourenço N. M. T.; Fonseca L. J. P.; Cordas C. M. Comparative Electrochemical Behavior of Cytochrome c on Aqueous Solutions Containing Choline-Based Room Temperature Ionic Liquids. Chem. Select 2017, 2, 8701–8705. 10.1002/slct.201702052. [DOI] [Google Scholar]
- Bisht M.; Mondal D.; Pereira M. M.; Freire M. G.; Venkatesu P.; Coutinho J. A. P. Long-Term Protein Packaging in Cholinium-Based Ionic Liquids: Improved Catalytic Activity and Enhanced Stability of Cytochrome c Against Multiple Stresses. Green Chem. 2017, 19, 4900–4911. 10.1039/C7GC02011B. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takekiyo T.; Ishikawa Y.; Yoshimura Y. Cryopreservation of Proteins Using Ionic Liquids: A Case Study of Cytochrome c. J. Phys. Chem. B 2017, 121, 7614–7620. 10.1021/acs.jpcb.7b05158. [DOI] [PubMed] [Google Scholar]
- Bisht M.; Jha I.; Venkatesu P. Does Choline-Based Amino Acid Ionic Liquid Behave as a Biocompatible Solvent for Stem Bromelain Structure?. Process Biochem. 2018, 74, 77–85. 10.1016/j.procbio.2018.07.005. [DOI] [Google Scholar]
- Bisht M.; Venkatesu P. Influence of Cholinium-Based Ionic Liquids on the Structural Stability and Activity of α-Chymotrypsin. New J. Chem. 2017, 41, 13902–13911. 10.1039/C7NJ03023A. [DOI] [Google Scholar]
- Fujita K.; Kajiyama M.; Liu Y.; Nakamura N.; Ohno H. Hydrated Ionic Liquids as a Liquid Chaperon for Refolding of Aggregated Recombinant Protein Expressed in Escherichia Coli. Chem. Commun. 2016, 52, 13491–13494. 10.1039/C6CC06999A. [DOI] [PubMed] [Google Scholar]
- Fujita K.; Nakano R.; Nakaba R.; Nakamura N.; Ohno H. Hydrated Ionic Liquids Enable Both Solubilization and Refolding of Aggregated Concanavalin A. Chem. Commun. 2019, 55, 3578–3581. 10.1039/C8CC10102G. [DOI] [PubMed] [Google Scholar]
- Constatinescu D.; Herrmann C.; Weingärtner H. Patterns of Protein Unfolding and Protein Aggregation in Ionic Liquids. Phys. Chem. Chem. Phys. 2010, 12, 1756–1763. 10.1039/b921037g. [DOI] [PubMed] [Google Scholar]
- Byrne N.; Wang L.-M.; Belieres J.-P.; Angell C. A. Reversible Folding-Unfolding, Aggregation Protection, and Multi-Year Stabilization, in High Concentration Protein Solutions, Using Ionic Liquids. Chem. Commun. 2007, 2714–2716. 10.1039/B618943A. [DOI] [PubMed] [Google Scholar]
- Byrne N.; Angell C. A. Protein Unfolding, and the “Tuning In” of Reversible Intermediate States, in Protic Ionic Liquid Media. J. Mol. Biol. 2008, 378, 707–714. 10.1016/j.jmb.2008.02.050. [DOI] [PubMed] [Google Scholar]
- Byrne N.; Belieres J.-P.; Angell C. A. The ‘Refoldability’ of Selected Proteins in Ionic Liquid as a Stabilization Criterion, Leading to a Conjecture on Biogenesis. Aust. J. Chem. 2009, 62, 328–333. 10.1071/CH08441. [DOI] [Google Scholar]
- Wijaya E. C.; Separovic F.; Drummond C. J.; Greaves T. L. Stability and Activity of Lysozyme in Stoichiometric and Non-Stoichiometric Protic Ionic Liquid (PIL)-Water Systems. J. Chem. Phys. 2018, 148, 193838. 10.1063/1.5010055. [DOI] [PubMed] [Google Scholar]
- Mann J. P.; McCluskey A.; Atkin R. Activity and Thermal Stability of Lysozyme in Alkylammonium Formate Ionic Liquids-Influence of Cation Modification. Green Chem. 2009, 11, 785–792. 10.1039/b900021f. [DOI] [Google Scholar]
- Takekiyo T.; Yamaguchi E.; Abe H.; Yoshimura Y. Suppression Effect on the Formation of Insulin Amyloid by the Use of Ionic Liquids. ACS Sustanable. Chem. Eng. 2016, 4, 422–428. 10.1021/acssuschemeng.5b00936. [DOI] [Google Scholar]
- Bharmoria P.; Mondal D.; Pereira M. M.; Neves M. C.; Almeida M. R.; Gomes M. C.; Mano J. F.; Bdikin I.; Ferreira R. A. S.; Coutinho J. A. P.; Freire M. G. Instantaneous Fibrillation of Egg White Proteome with Ionic Liquid and Macromolecular Crowding. Commun. Mater. 2020, 1, 34. 10.1038/s43246-020-0035-0. [DOI] [Google Scholar]
- Takekiyo T.; Yamazaki K.; Yamaguchi E.; Abe H.; Yoshimura Y. High Ionic Liquid Concentration-Induced Structural Change of Protein in Aqueous Solution: A Case Study of Lysozyme. J. Phys. Chem. B 2012, 116, 11092–11097. 10.1021/jp3057064. [DOI] [PubMed] [Google Scholar]
- Yoshimura Y.; Takekiyo T.; Mori T. Structural Study of Lysozyme in Two Ionic Liquids at Cryogenic Temperature. Chem. Phys. Lett. 2016, 664, 44–49. 10.1016/j.cplett.2016.10.013. [DOI] [Google Scholar]
- Tavares A. P. M.; Rodriguez O.; Macedo E. A. Ionic Liquids as Alternative Co-Solvents for Laccase: Study of Enzyme Activity and Stability. Biotechnol. Bioeng. 2008, 101, 201–207. 10.1002/bit.21866. [DOI] [PubMed] [Google Scholar]
- Ferdjani S.; Ionita M.; Roy B.; Dion M.; Djeghaba Z.; Rabiller C.; Tellier C. Correlation Between Thermostability and Stability of Glycosidases in Ionic liquid. Biotechnol. Lett. 2011, 33, 1215–1219. 10.1007/s10529-011-0560-5. [DOI] [PubMed] [Google Scholar]
- Kim H. S.; Ha S. H.; Sethaphong L.; Koo Y.-M.; Yingling Y. G. The Relationship between Enhanced Enzyme Activity and Structural Dynamics in Ionic Liquids: A Combined Computational and Experimental Study. Phys. Chem. Chem. Phys. 2014, 16, 2944–2953. 10.1039/c3cp52516c. [DOI] [PubMed] [Google Scholar]
- Nordwald E. M.; Armstrong G. S.; Kaar J. L. NMR-Guided Rational Engineering of an Ionic-Liquid-Tolerant Lipase. ACS Catal. 2014, 4, 4057–4064. 10.1021/cs500978x. [DOI] [Google Scholar]
- Bui-Le L.; Clarke C. J.; Bröhl A.; Brogan A. P. S.; Arpino J. A. J.; Polizzi K. M.; Hallett J. P. Revealing the Complexity of Ionic Liquid-Protein Interactions Through a Multi-Technique Investigation. Commun. Chem. 2020, 3, 55. 10.1038/s42004-020-0302-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lou W.-Y.; Zong M.-H.; Smith T. J.; Wu H.; Wang J.-F. Impact of Ionic Liquids on Papain: An Investigation of Structure-Function Relationships. Green Chem. 2006, 8, 509–512. 10.1039/B600930A. [DOI] [Google Scholar]
- Jha I.; Bisht M.; Mogha N. K.; Venkatesu P. The Effect of Imidazolium-Based Ionic Liquids on Structure and Stability of Stem Bromelain: Concentration and Alkyl Chain Length Effect. J. Phys. Chem. B 2018, 122, 7522–7529. 10.1021/acs.jpcb.8b04661. [DOI] [PubMed] [Google Scholar]
- Figueiredo A. M.; Sardinha J.; Moore G. R.; Cabrita E. J. Protein Destabilisation in Ionic Liquids: The Role of Preferential Interactions in Denaturation. Phys. Chem. Chem. Phys. 2013, 15, 19632–19643. 10.1039/c3cp53395f. [DOI] [PubMed] [Google Scholar]
- Sasmal D. K.; Mondal T.; Sen Mojumdar S.; Choudhury A.; Banerjee R.; Bhattacharyya K. An FCS Study of Unfolding and Refolding of CPM-Labeled Human Serum Albumin: Role of Ionic Liquid. J. Phys. Chem. B 2011, 115, 13075–13083. 10.1021/jp207829y. [DOI] [PubMed] [Google Scholar]
- Pusey M. L.; Paley M. S.; Turner M. B.; Rogers R. D. Protein Crystallization Using Room Temperature Ionic Liquids. Cryst. Growth Des. 2007, 7, 787–793. 10.1021/cg060696t. [DOI] [Google Scholar]
- Hekmat D.; Hebel D.; Joswig S.; Schmidt M.; Weuster-Botz D. Advanced Protein Crystallization Using Water-Soluble Ionic Liquids as Crystallization Additives. Biotechnol. Lett. 2007, 29, 1703–1711. 10.1007/s10529-007-9456-9. [DOI] [PubMed] [Google Scholar]
- Wang Z.; Fang W.; Li L.; Li Y.; Yu X.; Gu Q.; Zhou F. Effects of an Ionic Liquid as an Additive on the Thermal Denaturation/Degradation of Lysozyme Crystals. Asia-Pac. J. Chem. Eng. 2015, 10, 163–169. 10.1002/apj.1855. [DOI] [Google Scholar]
- Chen X.; Ji Y.; Wang J. Improvement on the Crystallization of Lysozyme in the Presence of Hydrophilic Ionic Liquid. Analyst 2010, 135, 2241–2248. 10.1039/c0an00244e. [DOI] [PubMed] [Google Scholar]
- Kowacz M.; Mukhopadhyay A.; Carvalho A. L.; Esperança J. M. S. S.; Romão M. J.; Rebelo L. P. N. Hofmeister Effects of Ionic Liquids in Protein Crystallization: Direct and Water-Mediated Interactions. CrystEngComm 2012, 14, 4912–4921. 10.1039/c2ce25129a. [DOI] [Google Scholar]
- Jaganathan M.; Ramakrishnan C.; Velmurugan D.; Dhathathreyan A. Understanding Ethylammonium Nitrate Stabilized Cytochrome c-Molecular Dynamics and Experimental Approach. J. Mol. Struct. 2015, 1081, 334–341. 10.1016/j.molstruc.2014.10.049. [DOI] [Google Scholar]
- Burney P. R.; Nordwald E. M.; Hickman K.; Kaar J. L.; Pfaendtner J. Molecular Dynamics Investigation of the Ionic Liquid/Enzyme Interface: Application to Engineering Enzyme Surface Charge. Proteins 2015, 83, 670–680. 10.1002/prot.24757. [DOI] [PubMed] [Google Scholar]
- Mohammad Latif M. A.; Micaelo N. M.; Abdul Rahman M. B. Influence of Anion-Water Interactions on the Behavior of Lipases in Room Temperature Ionic Liquids. RSC Adv. 2014, 4, 48202–48211. 10.1039/C4RA07460B. [DOI] [Google Scholar]
- Li W.; Wang L.; Zhou R.; Mu Y. Ionic Liquid Induced Inactivation of Cellobiohydrolase I from Trichoderma reesei. Green Chem. 2015, 17, 1618–1625. 10.1039/C4GC02136C. [DOI] [Google Scholar]
- Manna B.; Ghosh A. Structure and Dynamics of Ionic Liquid Tolerant Hyperthermophilic Endoglucanase Cel12A from Rhodothermus marinus. RSC Adv. 2020, 10, 7933–7947. 10.1039/C9RA09612D. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jaeger V. W.; Pfaendtner J. Destabilization of Human Serum Albumin by Ionic Liquids Studied Using Enhanced Molecular Dynamics Simulations. J. Phys. Chem. B 2016, 120, 12079–12087. 10.1021/acs.jpcb.6b09410. [DOI] [PubMed] [Google Scholar]
- Constantinescu D.; Weingärtner H.; Herrmann C. Protein Denaturation by Ionic Liquids, and the Hofmeister Series: A Case Study of Aqueous Solutions of Ribonuclease A. Angew. Chem., Int. Ed. 2007, 46, 8887–8889. 10.1002/anie.200702295. [DOI] [PubMed] [Google Scholar]
- Weibels S.; Syguda A.; Herrmann C.; Weingärtner H. Steering the Enzymatic Activity of Proteins by Ionic Liquids. A Case Study of the Enzyme Kinetics of Yeast Alcohol Dehydrogenase. Phys. Chem. Chem. Phys. 2012, 14, 4635–4639. 10.1039/c2cp24041f. [DOI] [PubMed] [Google Scholar]
- Kumar A.; Rani A.; Venkatesu P. A Comparative Study of the Hofmeister series of Anions of the Ionic Salts and Ionic Liquids on the Stability of α-Chymotrypsin. New J. Chem. 2015, 39, 938–952. 10.1039/C4NJ01596G. [DOI] [Google Scholar]
- Kumar A.; Venkatesu P. Does the Stability of Proteins in Ionic Liquids Obey the Hofmeister Series?. Int. J. Biol. Macromol. 2014, 63, 244–253. 10.1016/j.ijbiomac.2013.10.031. [DOI] [PubMed] [Google Scholar]
- Nordwald E. M.; Kaar J. L. Mediating Electrostatic Binding of 1-Butyl-3-methylimidazolium Chloride to Enzyme Surfaces Improves Conformational Stability. J. Phys. Chem. B 2013, 117, 8977–8986. 10.1021/jp404760w. [DOI] [PubMed] [Google Scholar]
- Jha I.; Venkatesu P. Unprecedented Improvement in the Stability of Haemoglobin in the Presence of Promising Green Solvent 1-Allyl-3-methylimidazolium Chloride. ACS Sustainable Chem. Eng. 2016, 4, 413–421. 10.1021/acssuschemeng.5b00939. [DOI] [Google Scholar]
- Rawat K.; Bohidar H. B. Universal Charge Quenching and Stability of Proteins in 1-Methyl-3-alkyl (Hexyl/Octyl) Imidazolium Chloride Ionic Liquid Solutions. J. Phys. Chem. B 2012, 116, 11065–11074. 10.1021/jp3049108. [DOI] [PubMed] [Google Scholar]
- Heller W. T. Comparison of the Thermal Denaturing of Human Serum Albumin in the Presence of Guanidine Hydrochloride and 1-Butyl-3-methylimidazolium Ionic Liquids. J. Phys. Chem. B 2013, 117, 2378–2383. 10.1021/jp400079p. [DOI] [PubMed] [Google Scholar]
- Shu Y.; Liu M.; Chen S.; Chen X.; Wang J. New Insight into Molecular Interactions of Imidazolium Ionic Liquids with Bovine Serum Albumin. J. Phys. Chem. B 2011, 115, 12306–12314. 10.1021/jp2071925. [DOI] [PubMed] [Google Scholar]
- Huang R.; Zhang S.; Pan L.; Li J.; Liu F.; Liu H. Spectroscopic Studies on the Interactions Between Imidazolium Chloride Ionic Liquids and Bovine Serum Albumin. Spectrochim.an Acta Part A: Mol. Biomol. Spectr. 2013, 104, 377–382. 10.1016/j.saa.2012.11.087. [DOI] [PubMed] [Google Scholar]
- Fan Y.; Zhang S.; Kong J.; Miao J. Study on the Interaction Between an Ionic Liquid and L-tryptophan by Fluorescence Spectroscopic Technique. Microchem. J. 2011, 99, 439–442. 10.1016/j.microc.2011.06.019. [DOI] [Google Scholar]
- Fan Y.; Yan J.; Zhang S.; Li J.; Chen D.; Duan P. Fluorescence Spectroscopic Analysis of the Interaction of Papain with Ionic Liquids. Appl. Biochem. Biotechnol. 2012, 168, 592–603. 10.1007/s12010-012-9801-x. [DOI] [PubMed] [Google Scholar]
- Fan Y.; Zhang S.; Wang Q.; Li J.; Fan H.; Shan D. Investigation of the Interaction of Pepsin with Ionic Liquids by Using Fluorescence Spectroscopy. Appl. Spectrosc. 2013, 67, 648–655. 10.1366/12-06793. [DOI] [PubMed] [Google Scholar]
- Fan Y.; Zhang S.; Wang Q.; Li J.; Fan H.; Shan D. Interaction of an Amino-Functionalized Ionic Liquid with Enzymes: A Fluorescence Spectroscopy Study. Spectrochim. Acta Part A: Mol. Biomol. Spectr. 2013, 105, 297–303. 10.1016/j.saa.2012.12.038. [DOI] [PubMed] [Google Scholar]
- Fan Y.; Dong X.; Li X.; Zhong Y.; Kong J.; Hua S.; Miao J.; Li Y. Spectroscopic Studies on the Inhibitory Effects of Ionic Liquids on Lipase Activity. Spectrochim. Acta Part A: Mol. Biomol. Spectr. 2016, 159, 128–133. 10.1016/j.saa.2016.01.047. [DOI] [PubMed] [Google Scholar]
- Fan Y.; Dong X.; Yan L.; Li D.; Hua S.; Hu C.; Pan C. Evaluation of the Toxicity of Ionic Liquids on Trypsin: A Mechanism Study. Chemosphere 2016, 148, 241–247. 10.1016/j.chemosphere.2016.01.033. [DOI] [PubMed] [Google Scholar]
- Ventura S. P. M.; Santos L. D. F.; Saraiva J. A.; Coutinho J. A. P. Concentration Effect of Hydrophilic Ionic Liquids on the Enzymatic Activity of Candida antarctica lipase B. World J. Microb. Biotechnol. 2012, 28, 2303–2310. 10.1007/s11274-012-1037-y. [DOI] [PubMed] [Google Scholar]
- Jaeger V. W.; Pfaendtner J. Structure, Dynamics, and Activity of Xylanase Solvated in Binary Mixtures of Ionic Liquid and Water. ACS Chem. Biol. 2013, 8, 1179–1186. 10.1021/cb3006837. [DOI] [PubMed] [Google Scholar]
- Mangialardo S.; Gontrani L.; Leonelli F.; Caminiti R.; Postorino P. Role of Ionic Liquids in Protein Refolding: Native/Fibrillar versus Treated Lysozyme. RSC Adv. 2012, 2, 12329–12336. 10.1039/c2ra21593d. [DOI] [Google Scholar]
- Sankaranarayanan K.; Sathyaraj G.; Nair B. U.; Dhathathreyan A. Reversible and Irreversible Conformational Transitions in Myoglobin: Role of Hydrated Amino Acid Ionic Liquid. J. Phys. Chem. B 2012, 116, 4175–4180. 10.1021/jp300596z. [DOI] [PubMed] [Google Scholar]
- Sankaranarayanan K.; Sreedhar B.; Nair B. U.; Dhathathreyan A. Microviscosity-Induced Conformational Transition in β-Lactoglobulin in the Presence of an Ionic Liquids. J. Phys. Chem. B 2013, 117, 1234–1240. 10.1021/jp310198f. [DOI] [PubMed] [Google Scholar]
- Ghaedizadeh S.; Emamzadeh R.; Nazari M.; Rasa S. M. M.; Zarkesh-Esfahani S. H.; Yousefi M. Understanding the Molecular Behavior of Renilla Luciferase in Imidazolium-based Ionic Liquids, a New Model for the α/β Fold Collapse. Biochem. Eng. J. 2016, 105, 505–513. 10.1016/j.bej.2015.10.024. [DOI] [Google Scholar]
- Rawat K.; Bohidar H. B. Heparin-like Native Protein Aggregate Dissociation by1-Alkyl-3-methyl imidazolium Chloride Ionic Liquids. Int. J. Biol. Macromol. 2015, 73, 23–30. 10.1016/j.ijbiomac.2014.10.057. [DOI] [PubMed] [Google Scholar]
- Bisht M.; Kumar A.; Venkatesu P. Refolding Effects of Partially Immiscible Ammonium-Based Ionic Liquids on the Urea-Induced Unfolded Lysozyme Structure. Phys. Chem. Chem. Phys. 2016, 18, 12419–12422. 10.1039/C6CP01022A. [DOI] [PubMed] [Google Scholar]
- Ding S.-F.; Xu M.-Q.; Zhao G.-C.; Wei X.-W. Direct Electrochemical Response of Myoglobin Using a Room Temperature Ionic Liquid, 1-(2-hydroxyethyl)-3-methyl imidazolium Tetrafluoroborate, as Supporting Electrolyte. Electrochem. Commun. 2007, 9, 216–220. 10.1016/j.elecom.2006.09.007. [DOI] [Google Scholar]
- Hasan F.; Vidanapathirana P.; Das S.; Fernand V. E.; Siraj N.; Losso J. N.; Warner I. M. Ionic Liquids as Buffer Additives in Ionic Liquid Polyacrylamide Gel Electrophoresis Separation of Mixtures of Low and High Molecular Weight Proteins. RSC Adv. 2015, 5, 69229–69237. 10.1039/C5RA11559K. [DOI] [Google Scholar]
- Yu D.; Huang X.; Deng M.; Lin Y.; Jiang L.; Huang J.; Wang Y. Effects of Inorganic and Organic Salts on Aggregation Behavior of Cationic Gemini Surfactants. J. Phys. Chem. B 2010, 114, 14955–14964. 10.1021/jp106031d. [DOI] [PubMed] [Google Scholar]
- Gehlot P. S.; Kulshrestha A.; Bharmoria P.; Damarla K.; Chokshi K.; Kumar A. Surface-Active Ionic Liquid Cholinium Dodecylbenzenesulfonate: Self-Assembling Behavior and Interaction with Cellulase. ACS Omega 2017, 2, 7451–7460. 10.1021/acsomega.7b01291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh G.; Kaur M.; Aswal V. K.; Kang T. S. Aqueous Colloidal Systems of Bovine Serum Albumin and Functionalized Surface-Active Ionic Liquids for Material Transport. RSC Adv. 2020, 10, 7073–7082. 10.1039/C9RA05549E. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang T.; Cao C.; Liu Z.; Li Y.; Du F. Interaction of Pepsin-[C16mim][Br] System: Interfacial Dilational Rheology and Conformational Studies. Soft Matter 2014, 10, 6810–6819. 10.1039/C4SM00950A. [DOI] [PubMed] [Google Scholar]
- Singh G.; Kaur M.; Singh D.; Kesavan A. K.; Kang T. S. Antimicrobial Colloidal Complexes of Lysozyme with Bio-Based Surface-Active Ionic Liquids in Aqueous Medium. J. Phys. Chem. B 2020, 124, 3791–3800. 10.1021/acs.jpcb.0c00339. [DOI] [PubMed] [Google Scholar]
- Debnath S.; Das D.; Dutta S.; Das P. K. Imidazolium Bromide-Based Ionic Liquid Assisted Improved Activity of Trypsin in Cationic Reverse Micelles. Langmuir 2010, 26, 4080–4086. 10.1021/la9040419. [DOI] [PubMed] [Google Scholar]
- Kundu K.; Singh A. P.; Panda S.; Singh V.; Gardas R. L.; Senapati S. Study on the Conformation of Entrapped Protein inside the Reverse Micellar Confinement Based on the Amino Acid Derived Ionic Liquid. Chem. Select 2018, 3, 4768–4776. 10.1002/slct.201800918. [DOI] [Google Scholar]
- Kaur M.; Singh G.; Kaur A.; Sharma P. K.; Kang T. S. Thermally Stable Ionic Liquid-Based Microemulsions for High- Temperature Stabilization of Lysozyme at Nanointerfaces. Langmuir 2019, 35, 4085–4093. 10.1021/acs.langmuir.9b00106. [DOI] [PubMed] [Google Scholar]
- Bento R. M. F.; Almeida M. R.; Bharmoria P.; Freire M. G.; Tavares A. P. M. Improvements in the Enzymatic Degradation of Textile Dyes Using Ionic Liquid-Based Surfactants. Sep. Purif. Technol. 2020, 235, 116191. 10.1016/j.seppur.2019.116191. [DOI] [Google Scholar]
- Alves M.; Vieira N. S.M.; Rebelo L. P. N.; Araújo J. M. M.; Pereiro A. B.; Archer M. Fluorinated Ionic Liquids for Protein Drug Delivery Systems: Investigating Their Impact on the Structure and Function of Lysozyme. Int. J. Pharm. 2017, 526, 309–320. 10.1016/j.ijpharm.2017.05.002. [DOI] [PubMed] [Google Scholar]
- Kundu S.; Banerjee C.; Sarkar N. Inhibiting the Fibrillation of Serum Albumin Proteins in Presence of Surface-Active Ionic Liquids (SAILs) at Low pH: A Spectroscopic and Microscopic Study. J. Phys. Chem. B 2017, 121, 7550–7560. 10.1021/acs.jpcb.7b03457. [DOI] [PubMed] [Google Scholar]
- Singh U. K.; Patel R. Dynamics of Ionic Liquids Assisted Refolding of Denatured Cytochrome c: A Study of Preferential Interactions towards Renaturation. Mol. Pharmaceutics 2018, 15, 2684–2697. 10.1021/acs.molpharmaceut.8b00212. [DOI] [PubMed] [Google Scholar]
- Albertsson P.-A.Partition of Cell Particles and Macromolecules: Separation and Purification of Biomolecules, Cell Organelles, Membranes and Cells in Aqueous Polymer two Phase Systems and Their use in Biochemical Analysis and Biotechnology; John Wiley and Sons: Chichester, 1986. [Google Scholar]
- Gutowski K. E.; Broker G. A.; Willauer H. D.; Huddleston J. G.; Swatloski R. P.; Holbrey J. D.; Rogers R. D. Controlling the Aqueous Miscibility of Ionic Liquids: Aqueous Biphasic Systems of Water-Miscible Ionic Liquids and Water-Structuring Salts for Recycle, Metathesis, and Separations. J. Am. Chem. Soc. 2003, 125, 6632–6633. 10.1021/ja0351802. [DOI] [PubMed] [Google Scholar]
- Ventura S. P. M.; e Silva F. A.; Quental M. V.; Mondal D.; Freire M. G.; Coutinho J. A. P. Ionic-Liquid-Mediated Extraction and Separation Processes for Bioactive Compounds: Past, Present, and Future Trends. Chem. Rev. 2017, 117, 6984–7052. 10.1021/acs.chemrev.6b00550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pereira M. M.; Pedro S. N.; Quental M. V.; Lima Á S.; Coutinho J. A. P.; Freire M. G. Enhanced Extraction of Bovine Serum Albumin with Aqueous Biphasic Systems of Phosphonium- and Ammonium-based Ionic Liquids. J. Biotechnol. 2015, 206, 17–25. 10.1016/j.jbiotec.2015.03.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taha M.; Quental M. V.; Correia I.; Freire M. G.; Coutinho J. A. P. Extraction and Stability of Bovine Serum Albumin (BSA) using Cholinium-Based Good’s Buffers Ionic Liquids. Proc. Biochem. 2015, 50, 1158–1166. 10.1016/j.procbio.2015.03.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gupta B. S.; Taha M.; Lee M. J. Extraction of an Active Enzyme by Self-Buffering Ionic Liquids: A Green Medium for Enzymatic Research. RSC Adv. 2016, 6, 18567–18576. 10.1039/C6RA00607H. [DOI] [Google Scholar]
- Taha M.; Almeida M. R.; Silva F. A.; Domingues P.; Ventura S. P. M.; Coutinho J. A. P.; Freire M. G. Novel Biocompatible and Self-buffering Ionic Liquids for Biopharmaceutical Applications. Chem.—Eur. J. 2015, 21, 4781. 10.1002/chem.201405693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramalho C. C.; Neves C.M. S. S.; Quental M. V.; Coutinho J. A. P.; Freire M. G. Separation of Immunoglobulin G using Aqueous Biphasic Systems Composed of Cholinium-based Ionic Liquids and Poly(propylene glycol). J. Chem. Technol. Biotechnol. 2018, 93, 1931–1939. 10.1002/jctb.5594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mondal D.; Sharma M.; Quental M. V.; Tavares A. P. M.; Prasad K.; Freire M. G. Suitability of Bio-based Ionic Liquids for the Extraction and Purification of IgG Antibodies. Green Chem. 2016, 18, 6071–6081. 10.1039/C6GC01482H. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tseng M.-C.; Yuan T.-C.; Li Z.; Chu Y.-H. Crowned Ionic Liquids for Biomolecular Interaction Analysis. Anal. Chem. 2016, 88, 10811–10815. 10.1021/acs.analchem.6b03323. [DOI] [PubMed] [Google Scholar]
- Ikeda K.; Ikari R.; Nakamura N.; Ohno H.; Fujita K. Distribution of Cytochrome c in LCST-Type Ionic Liquid/Water Mixtures Controlled by Applied Potential and Temperature. J. Electrochem. Soc. 2018, 165, G96–G100. 10.1149/2.1071807jes. [DOI] [Google Scholar]
- Lee S. Y.; Khoiroh I.; Ooi C. W.; Ling T. C.; Show P. L. Recent Advances in Protein Extraction Using Ionic Liquid-based Aqueous Two-phase Systems. Sep. Purif. Rev. 2017, 46, 291–304. 10.1080/15422119.2017.1279628. [DOI] [Google Scholar]
- Baumruck A. C.; Yang J.; Thomas G.-F.; Beyer L. I.; Tietze D.; Tietze A. A. Native Chemical Ligation of Highly Hydrophobic Peptides in Ionic Liquid-Containing Media. J. Org. Chem. 2021, 86, 1659–1666. 10.1021/acs.joc.0c02498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reinhardt A.; Horn M.; Schmauck J. P.; Bröhl A.; Giernoth R.; Oelkrug C.; Schubert A.; Neundorf I. Novel Imidazolium Salt-Peptide Conjugates and Their Antimicrobial Activity. Bioconjugate Chem. 2014, 25, 2166–2174. 10.1021/bc500510c. [DOI] [PubMed] [Google Scholar]
- Wiwatchaiwong S.; Matsumura H.; Nakamura N.; Yohda M.; Ohno H. Stability of Thermophilic Cytochrome P450 Modified with Poly(ethylene oxide) in Ionic Liquid. Chem. Lett. 2006, 35, 798–799. 10.1246/cl.2006.798. [DOI] [Google Scholar]
- Chado G. R.; Holland E. N.; Tice A. K.; Stoykovich M. P.; Kaar J. L. Modification of Lipase with Poly(4-acryloylmorpholine) Enhances Solubility and Transesterification Activity in Anhydrous Ionic Liquids. Biomacromolecules 2018, 19 (4), 1324–1332. 10.1021/acs.biomac.8b00176. [DOI] [PubMed] [Google Scholar]
- Slocik J. M.; Dennis P. B.; Kuang Z.; Pelton A.; Naik R. R. Creation of Stable Water-Free Antibody Based Protein Liquids. Comm. Mater. 2021, 2, 118. 10.1038/s43246-021-00222-2. [DOI] [Google Scholar]
- Behera S.; Balasubramanian S. Molecular Simulations Explain the Exceptional Thermal Stability, Solvent Tolerance and Solubility of Protein-Polymer Surfactant Bioconjugates in Ionic Liquids. Phys. Chem. Chem. Phys. 2022, 24, 21904–21915. 10.1039/D2CP02636H. [DOI] [PubMed] [Google Scholar]
- Hisamitsu S.; Yanai N.; Kimizuka N. Photon-Upconverting Ionic Liquids: Effective Triplet Energy Migration in Contiguous Ionic Chromophore Arrays. Angew. Chem., Int. Ed. 2015, 54, 11550–11554. 10.1002/anie.201505168. [DOI] [PubMed] [Google Scholar]
- Hisamitsu S.; Yanai N.; Kouno H.; Magome E.; Matsuki M.; Yamada T.; Monguzzi A.; Kimizuka N. Two-Dimensional Structural Ordering in a Chromophoric Ionic Liquid for Triplet Energy Migration-Based Photon Upconversion. Phys. Chem. Chem. Phys. 2018, 20, 3233–3240. 10.1039/C7CP06266D. [DOI] [PubMed] [Google Scholar]
- Bharmoria P.; Hisamitsu S.; Nagatomi H.; Ogawa T.; Morikawa M.; Yanai N.; Kimizuka N. Simple and Versatile Platform for Air-Tolerant Photon Upconverting Hydrogels by Biopolymer-Surfactant-Chromophore Co-assembly. J. Am. Chem. Soc. 2018, 140, 10848–10855. 10.1021/jacs.8b05821. [DOI] [PubMed] [Google Scholar]
- Prince E.; Kumacheva E. Design and Applications of Man-made Biomimetic Fibrillar Hydrogels. Nat. Rev. Mater. 2019, 4, 99–115. 10.1038/s41578-018-0077-9. [DOI] [Google Scholar]
- Richter K.; Nessling M.; Lichter P. Experimental Evidence for the Influence of Molecular Crowding on Nuclear Architecture. Cell Sci. 2007, 120, 1673–1680. 10.1242/jcs.03440. [DOI] [PubMed] [Google Scholar]
- Kim Y. C.; Best R. B.; Mittal J. Macromolecular Crowding effects on Protein-Protein Binding Affinity and Specificity. J. Chem. Phys. 2010, 133, 205101. 10.1063/1.3516589. [DOI] [PubMed] [Google Scholar]
- Tan C.; Saurabh S.; Bruchez M. P.; Schwartz R.; LeDuc P. Molecular Crowding Shapes Gene Expression in Synthetic Cellular Nanosystems. Nat. Nanotechnol. 2013, 8, 602–608. 10.1038/nnano.2013.132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coussens L.; Yang-Feng T. L.; Liao C.; Chen E.; Gray A.; McGrath J.; Seeburg P. H.; Libermann T. A.; Schlessinger J.; Francke U.; et al. Tyrosine Kinase Receptor With Extensive Homology to EGF Receptor Shares Chromosomal Location With Neu Oncogene. Science. 1985, 230, 1132–1139. 10.1126/science.2999974. [DOI] [PubMed] [Google Scholar]
- Li B. T.; Ross D. S.; Aisner D. L.; Chaft J. E.; Hsu M.; Kako S. L.; Kris M. G.; Varella-Garcia M.; Arcila M. E. HER2 Amplification and HER2Mutation Are Distinct Molecular Targets in Lung Cancers. J. Thorac Oncol. 2016, 11, 414–419. 10.1016/j.jtho.2015.10.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mineev K. S.; Bocharov E. V.; Pustovalova Y. E.; Bocharova O. V.; Chupin V. V.; Arseniev A. S. Spatial Structure of the Transmembrane Domain Heterodimer of ErbB1 and ErbB2 Receptor Tyrosine Kinases. J. Mol. Biol. 2010, 400, 231–243. 10.1016/j.jmb.2010.05.016. [DOI] [PubMed] [Google Scholar]
- Thayallath S. K.; Shet S. M.; Bisht M.; Bharadwaj P.; Pereira M. M.; Franklin G.; Nataraj S. K.; Mondal D. Designing Protein Nano-Construct in Ionic Liquid: A Boost in Efficacy of Cytochrome C Under Stresses. Chem. Commun. 2023, 59, 5894–5897. 10.1039/D3CC00644A. [DOI] [PubMed] [Google Scholar]
- Shet S. M.; Thayallath S. K.; Bisht M.; Pereira M. M.; Coutinho J. A. P.; Sanna Kotrappanavar N.; Mondal D. Engineering Cytochrome C with Quantum Dots and Ionic Liquids: A Win-Win Strategy for Protein Packaging against Multiple Stresses. ACS Sustainable Chem. Eng. 2021, 9, 8327–8335. 10.1021/acssuschemeng.1c03173. [DOI] [Google Scholar]
- Shet S. M.; Bisht M.; Pramanik S.; Roy S.; Kumar T. S.; Nataraj S. K.; Mondal D.; Bhandari S. Engineering Quantum Dots with Ionic Liquid: A Multifunctional White Light Emitting Hydrogel for Enzyme Packaging. Adv. Opt. Mater. 2020, 8, 1902022. 10.1002/adom.201902022. [DOI] [Google Scholar]