

Abstract
The transmission-blocking vaccine candidate Pfs48/45 from the human malaria parasite Plasmodium falciparum is known to be difficult to express in heterologous systems, either as full-length protein or as correctly folded protein fragments that retain conformational epitopes. In this study we express full-length Pfs48/45 in the rodent parasite P. berghei. Pfs48/45 is expressed as a transgene under control of the strong P. berghei schizont-specific msp1 gene promoter (Pfs48/45@PbMSP1). Pfs48/45@PbMSP1 schizont-infected red blood cells produced full-length Pfs48/45 and the structural integrity of Pfs48/45 was confirmed using a panel of conformation-specific monoclonal antibodies that bind to different Pfs48/45 epitopes. Sera from mice immunized with transgenic Pfs48/45@PbMSP1 schizonts showed strong transmission-reducing activity in mosquitoes infected with P. falciparum using standard membrane feeding. These results demonstrate that transgenic rodent malaria parasites expressing human malaria antigens may be used as means to evaluate immunogenicity and functionality of difficult to express malaria vaccine candidate antigens.
Keywords: Plasmodium, Pfs48/45, Transgene-expression, Immunization, Transmission blocking, Standard membrane feeding
1. Introduction
Efficient and conformationally-accurate expression of Plasmodium proteins in heterologous systems, such as yeast or bacteria, is frequently problematic resulting in misfolded or incorrectly modified proteins, which are often poorly expressed [1,2]. This hampers the screening of Plasmodium antigens in immunization studies for their suitability as vaccine candidate antigens. Preclinical evaluation of Plasmodium antigens often involves immunizing rodents with recombinant Plasmodium proteins followed by an examination of induced immune responses, either in vivo using rodent models of malaria or in vitro by performing functional assays with human malaria parasites incubated with immune sera [3]. Multiple factors contribute to inefficient expression of Plasmodium proteins, such as the high AT content of Plasmodium genes, large size and often unique protein structure (i.e. encoding repeated stretches of amino acids) and unique post-translational modifications [1,4]. This is particularly evident for cysteine-rich proteins where correct folding depends on accurate formation of disulfide bridges to form domains specific for Plasmodium proteins [5–7]. Transgenic rodent malaria parasites (RMP) expressing human malaria parasite (HMP) proteins are increasingly used to evaluate and rank order candidate malaria vaccines before investing in scalable manufacture to support advancement to clinical testing [3]. Such transgenic RMP have been used in preclinical assays to evaluate vaccine potential of HMP proteins, both in vivo where mice are immunized with HMP antigens and subsequently challenged with transgenic RMP expressing the cognate HMP or in in vitro assays where immune sera or antibodies are evaluated for inhibition of parasite growth or invasion. Both the functional complementation of RMP genes by the HMP orthologs [3] and analysis of HMP expression using antisera, provide evidence for correct expression of functional HMP proteins in transgenic RMP [8]. Based on these studies, we reasoned that transgenic RMP can be used as expression systems to more efficiently express, screen, validate and down-select HMP antigens as potential novel malaria vaccine candidates [2,9]. Further, the expression of conformationally-accurate Plasmodium proteins could be used to generate epitope-specific monoclonal antibodies, which in turn can be used to better characterize the vaccine antigen. The use of RMP would circumvent many of the above-mentioned problems associated with expression in heterologous expression systems including, but not limited to, peculiarities of post-translational modifications and Plasmodium-specific domains involved in protein trafficking and cellular location. As a proof of concept, we generated transgenic P. berghei (Pb) parasites that express full length Pfs48/45 from P. falciparum (Pf). The Pfs48/45 protein is expressed in Plasmodium gametocytes and gametes [10,11] and contains multiple cysteine-rich domains with multiple disulfide bonds [12–14]. These constitute distinct conformational B cell epitopes that can be recognized by several monoclonal antibodies some of which have transmission-blocking (TB) activity [15]. Pfs48/45 becomes exposed on the surface of gametes once the parasite is taken up in blood meal by a mosquito and here the antigen can be targeted by antibodies and other components of the blood meal [16]. Expression of Pfs48/45 for TB immunization studies has been problematic in most commonly used expression systems, mainly due to incorrect or insufficient protein folding, which is dependent on the correct formation of disulfide bridges in this cysteine-rich protein [17,18]. The limited reactivity of recombinant Pfs48/45 with monoclonal antibodies against conformational epitopes of Pfs48/45 has indicated this misfolding [19,20].
2. Results and discussion
In this study the coding sequence of the gene encoding Pfs48/45 (PF3D7_1346700) was introduced into the redundant P. berghei p230p gene locus (PBANKA_0306000) [3,21,22]. The Pfs48/45 gene was placed under control of 1.3 kb of the promoter region of the schizont-specific Pb msp1 gene (PBANKA_0831000). This promoter was chosen since msp1 is one of the highest transcribed genes in developing Pb schizonts [23] and the Pb schizont stage can be easily produced and purified in large quantities [24]. The transgenic parasite (Pfs48/45@ PbMSP1) was generated by the method of GIMO transfection and selection [21]. Using this method transgenes can be rapidly introduced into the p230p gene locus in a GIMOpbANKA parent line by replacing the positive-negative selectable marker expression cassette by the transgene expression cassette (Supplementary M&M and Fig. 1A). Correct replacement of the selectable marker cassette and insertion of the Pfs48/45 expression cassette in a cloned line of Pfs48/45@PbMSP1 (1807cl2) was confirmed by diagnostic PCR and Southern analysis of chromosomes separated by pulsed-field gel electrophoresis (Fig. 1B). Analysis of the growth rate of transgenic Pfs48/45@PbMSP1 parasites during the cloning period demonstrated normal growth of blood stages, comparable to wild type (WT) PbANKA parasites (i.e. all mice (n = 3) achieved a 0.5–2% parasitemia on day 8, after inoculation with a single infected red blood cell). To obtain transgenic schizonts, parasites were cultured overnight using standard methods to produce and purify Pb schizonts (Supplementary M&M).
Fig. 1.
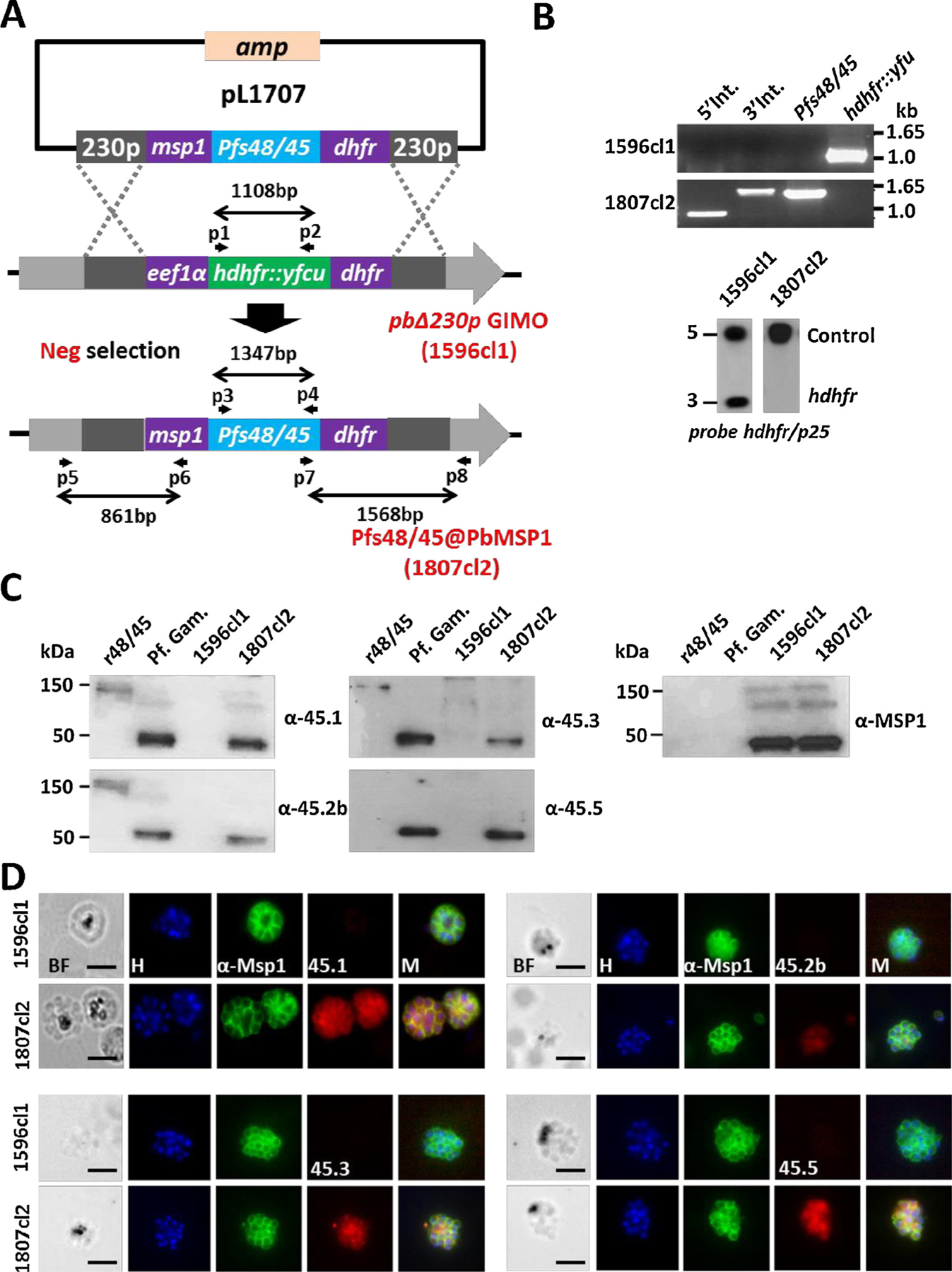
Generation, genotype and phenotype analyses of Pfs48/45@PbMSP1, a transgenic P. berghei parasite expressing P. falciparum P48/45 in schizonts.
(A) Schematic representation of the introduction of the Pfs48/45-expression cassette into the GIMOpbANKA parasite (line 1596cl1). Construct pL1707 contains the Pfs48/45 gene flanked by the msp1 promoter region and the 3′ pbdhfr UTR. This construct is integrated into the modified P. berghei 230p locus of GIMOpbANKA that contains the hdhfr::yfcu selectable marker (SM) cassette by double cross-over homologous recombination at the homology regions (230p; grey boxes). Negative selection with 5-FC selects for parasites that have the SM cassette replaced by the Pfs48/45 expression cassette. Location of primers used for PCR analysis and sizes of PCR products are shown.
(B) Diagnostic PCR (upper panel) and Southern analysis of PFG-separated chromosomes (lower panel) confirm correct integration of construct pL1707 in line 1807cl2 parasites. PCR shows the absence of the hdhfr::yfcu marker and the presence of the Pfs48/45. 5′ integration PCR (5′ int; primers p5/p6), 3′ integration PCR (3′ int; primers p7/p8), hdhfr::yfcu (primers p1/p2), Pfs48/45 (primers p3/p4). Primer locations and product sizes are shown in A and primer sequences in Table S1). Hybridization of PFG-separated chromosomes with a mixture of two probes (the hdhfr probe and a control probe recognizing p25 gene on chromosome 5) shows the removal of the SM cassette marker in the 230p locus on chromosome 3 in 1807cl2 parasites.
(C) Western analysis of Pfs48/45 expression in protein extracts of purified gametocytes of P. falciparum (Pf Gam), purified schizonts of wild type P. berghei (1596cl1) and purified schizonts of Pfs48/45@PbMSP1 (1807cl2). As a positive control, recombinant P. falciparum P48/45 fragment fused to GLURP R0 domain (R0.10C) was included (expected molecular size is 150 kDa). Blots were stained with 4 different anti-Pfs48/45 antibodies (45.1–3, 45.5) that recognize different epitopes. Anti-PyMSP1 antibody staining was used as a loading control.
(D) Immuno-fluorescence analyses of Pfs48/45 expression in purified schizonts of Pfs48/45@PbMSP1 (1807cl2), and the reference parent P. berghei GIMO line (i.e. WT; 1596cl1). Fixed parasites were stained with four different rat anti-Pfs48/45 mAbs (45.1–3, 45.5) and rabbit anti-PyMSP1 antibody followed by secondary conjugated antibodies anti-rabbit IgG Alexa Fluor ® 488 (green) or anti-rat IgG Alexa Fluor ® 594 (red). Nuclei stained with the DNA-specific dye Hoechst 33,342 (H). All pictures were recorded with the same exposure/gain times; anti-rabbit IgG Alexa Fluor ® 488 (green) 0.7 s; anti-rat IgG Alexa Fluor ® 594 (red) 0.6 s; Hoechst (blue) 0.136 s; bright field 0.62 s (1x gain). BF: bright field; M: merged. Scale bar: 2 μm.
We confirmed expression of Pfs48/45 in the transgenic schizonts by Western and immuno-fluorescence analysis using four anti-Pfs48/45 monoclonal antibodies 85RF45.1 (45.1), 85RF45.2b (45.2b), 85RF45.3 (45.3), and 85RF45.5 (45.5). Three of these (45.1, 45.2b and 45.3) recognize conformational epitopes (epitopes I, IIb and III respectively) in the C terminal region of Pfs48/45 [14].
In Western analysis all the antibodies recognize a protein of the expected size (48 kDa) in protein extracts from Pfs48/45@PbMSP1 schizonts and WT Pf gametocytes but not in extracts from a Pb line that does not express Pfs48/45 (i.e. GIMOpbANKA line 1596cl1). As a positive control, recombinant protein that contains a fragment of P. falciparum Pfs48/45 fused to the GLURP R0 domain (R0.10C) was included and as expected a 150 kDa band was present after probing with monoclonal antibodies 45.1, 45.2b and 45.3 but was not present after probing with 45.5 (Fig. 1C) [25]. We next examined the presence of Pfs48/45 epitopes using the anti-Pfs48/45 antibodies by immuno-fluorescence assay (IFA) (Fig. 1D). All antibodies recognized Pfs48/45 produced in the Pfs48/45@PbMSP1 schizonts and did not react with proteins of WT Pb schizonts (Fig. 1C and 1D). These results demonstrate that transgenic Pb schizonts can effectively express full length Pfs48/45, which retains a number of conformational epitopes. Pfs48/45, like MSP1, contains a GPI anchor and is present at the plasma membrane of Pf gametocytes/gametes [26]. The immuno-fluorescence analyses indicate that Pfs48/45 was located in the cytoplasm of the transgenic merozoites as the fluorescence signals did not completely overlap with fluorescence signals obtained with anti-MSP1 antibodies, which stain MSP1 at the merozoite plasma membrane in mature schizonts (Fig. 1D). Possible reasons for this observation is that GPI attachment may be different between rodent and human Plasmodium parasites, or that the attachment of Pfs48/45 onto P. berghei merozoites would require the presence of other Plasmodium proteins normally present in gametocytes/gametes.
In order to estimate the proportion of Pfs48/45 present in the protein lysates of Pfs48/45@PbMSP1 schizonts we performed a quantitative Western Blot analysis. Densitometry analysis of signals obtained after probing known amounts of Pfs48/45@PbMSP1 protein schizont lysates and a dilution series of recombinant Pfs48/45 with anti-Pfs48/45 monoclonal 45.1, revealed that the intensity of the schizont lysate signals corresponds to less than 1 ng of recombinant Pfs48/45 (Supplementary Fig. S1), indicating that is between 0.25 – 0.12% of the total schizont lysate is Pfs48/45 (Fig. 2A).
Fig. 2.
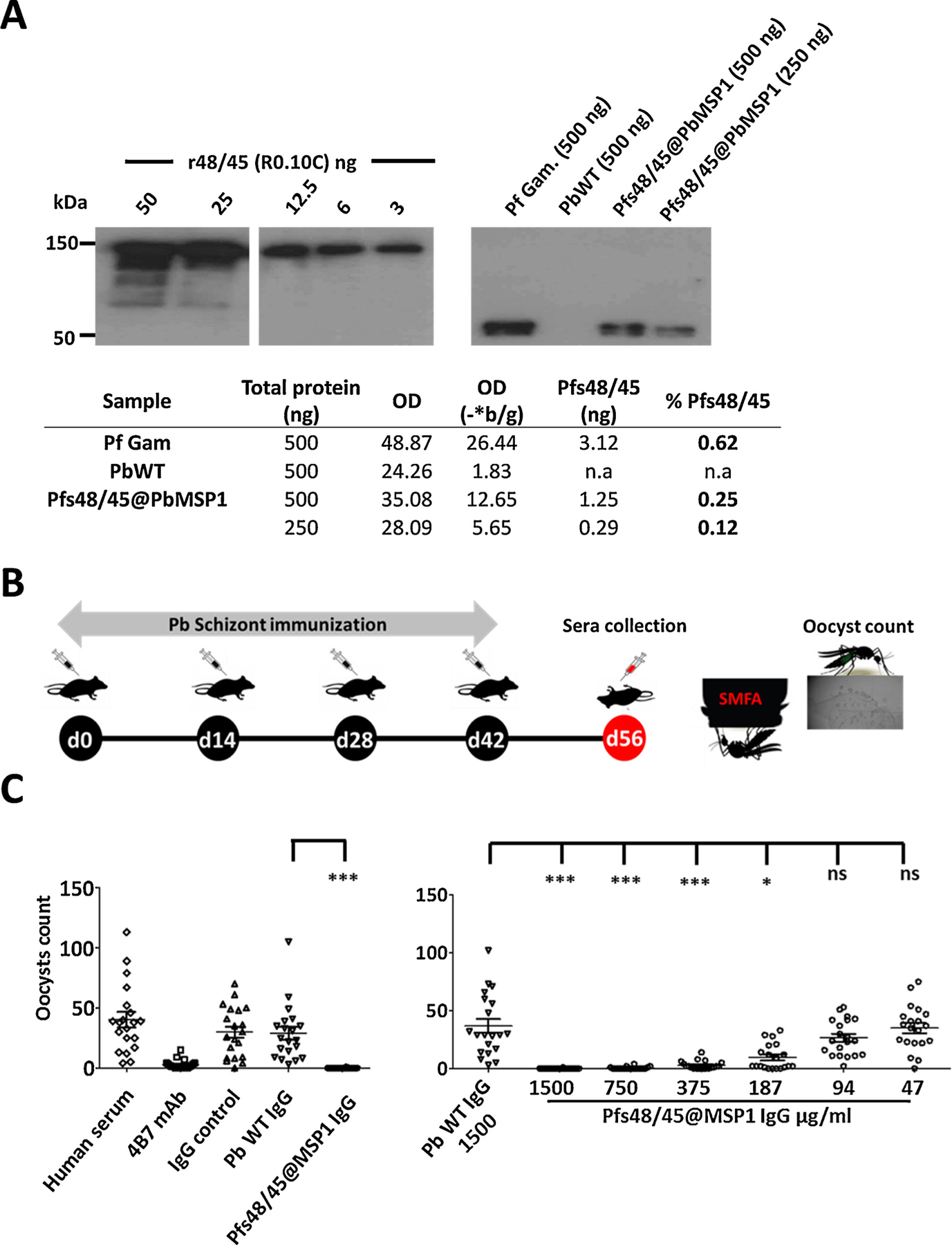
Quantification of Pfs48/45 protein in Pfs48/45@PbMSP1 schizont lysate and transmission reducing (TR) activity of IgG isolated from mice immunized with Pfs48/45@PbMSP1 schizont lysates.
A. Pfs48/45@PbMSP1 schizont lysates (500 & 250 ng), P. falciparum gametocytes (Pf Gam.; 500 ng) and P. berghei WT schizont lysate (PbWT; 500 ng) were analyzed in Western blot analysis using anti-Pfs48/45 monoclonal 85RF45.1 (1:2000). Densitometry analysis was performed on signals after probing Pfs48/45@PbMSP1 schizont lysate (500 ng) and a dilution series (50, 25, 12.5, 6 and 3 ng) of recombinant Pfs48/45 (r48/45; R0.10C) with antibody 45.1. The Table shows the calculated Pfs48/45 protein content (ng) and the percentage of Pfs48/45 protein in parasite samples; see Supplementary Fig. S1 for determination of Pfs48/45 in samples. *quantification performed after subtraction of background (b/g) Optical Densitometry (OD) values and **quantitation based on regression curve calculations (see Supplementary Fig. S1).
B. Timeline showing the immunization of mice with extracts of Pfs48/45@PbMSP1 and PbWT schizont lysates and collection of sera for isolation of IgG that is tested for TR activity in standard membrane feeding assays (SMFA) of P. falciparum gametocytes to Anopheles stephensi mosquitoes (see C).
C. Left panel: First SMFA with IgGs from mice immunized with purified schizonts of Pfs48/45@PbMSP1 and PbWT. TR activity was determined by the mean number of oocysts 8 days after feeding, and significance of inhibition was determined by the zero-inflated negative binomial model described previously [29]. Right panel: Second SMFA with serially diluted IgGs. IgG from mice immunized with purified schizonts of Pfs48/45@PbMSP1 was titrated resulting in the concentrations shown in the Figure. Significant TR activity was detected until a concentration of 187 μg/ml (*p = 0.014). Significant; *p < 0.05, ***p < 0.001.
Next, we examined if Pfs48/45@PbMSP1 transgenic schizont lysate could be used to raise sera that could block Pf transmission in mosquitoes, presumably by Pfs48/45 specific antibodies. Two groups of 10 C57BL/6 mice were immunized 4 times (2 week interval) with lysates of 1 × 108 schizonts of either Pfs48/45@PbMSP1 or WT (c15cy1) parasites (Fig. 2B). Purified schizonts were inactivated by three rounds of freezing on dry ice followed by thawing at room temperature before immunization and schizont lysates were injected intravenously. Two weeks after the final immunization, serum was collected from all animals and a serum pool made for each group. Total IgG was isolated from the pooled sera and tested for transmission-reducing activity (TR activity) in standard membrane feeding assays (SMFA) using Pf gametocytes (Supplementary M&M). Pf gametocytes were fed to A. stephensi mosquitoes in the presence of IgG obtained from mice immunized with schizonts of either Pfs48/45@PbMSP1 or WT. In the first experiment, IgG (1500 μg/ml) from Pfs48/45@PbMSP1-immunized mice showed 99.8% inhibition in oocyst density (p = 0.001) compared to the IgG obtained from WT immunized (Fig. 2C). Next, TR activity was determined in SMFA using a dilution series of the IgG obtained from Pfs48/45@PbMSP1-immunized mice. Significant TR activity with IgG from Pfs48/45@PbMSP1-immunized mice was still observed at a concentration of 187 μg/ml (p = 0.014) compared to the control IgG (Fig. 2C). The quantitative Western blot analysis (Fig. 2A) indicated that is between 0.25–0.12% of the total Pfs48/45@PbMSP1 schizont lysate was Pfs48/45 and therefore it is likely that the majority of the IgG from the immunized mice is not directed against Pfs48/45. The failure to induce TR activity of IgG of mice immunized with WT schizont lysate indicates that the small proportion of anti-Pfs48/45 antibodies are mediating the TR activity after Pfs48/45@PbMSP1 schizont lysate immunization. The strong TR activity mediated by the total IgG isolated from Pfs48/45@PbMSP1 immunized mice (Fig. 2C), indicates that Pfs48/45 expressed in P. berghei can induce antibodies with potent TR activity.
Combined, our proof-of concept studies demonstrate that transgenic Pb schizonts can be used as a system to produce a difficult to express HMP protein that is correctly folded and retains conformational epitopes of the native protein. This opens possibilities to use this expression system to evaluate the immunogenicity of other difficult to express antigens or specific domains of these parasites. Studies using sera obtained from mice immunized with Pf proteins expressed by transgenic Pb parasites could be used to rank-order novel vaccine candidate antigens, not only in TB studies but also for blood-stage antigens using blood stage growth inhibition assays (GIA) or sporozoite-antigens using inhibition of sporozoite invasion (ISI) assays [3]. Moreover, the expression in transgenic schizonts of HMP proteins with affinity tags will allow for the purification of these HMP proteins from whole parasite lysate preparations and immunization with purified protein will mean that all of the raised immune response is due to the target antigen and will permit a more detailed analyses of antigen immunogenicity, for example to examine and clone potent inhibitory and cross-reactive B-cells/antibodies after rodent immunization [27]. The creation of transgenic parasites that express antigens from multiple life-cycles that can induce potent immune responses is also of interest to the development of whole organism vaccines [28]. For example, genetically attenuated sporozoite vaccines could be further modified to induce immune responses against multiple life cycle stages by expression in sporozoites and liver stages antigens of blood- or transmission–stages to produce a multi stage-vaccine.
Supplementary Material
Acknowledgements
We would like to thank Dr. Michael Theisen and Dr. Susheel K. Singh, Statens Seum Institute, Copenhagen Denmark, for kindly providing us with the recombinant P. falciparum P48/45 protein fragment (R0.10C). A.S. Othman is supported by a Skim Latihan Akademik IPTA - SLAI (Ministry of Higher Education, Malaysia). C. M. Mogollon was supported by Colciencias pH.D. fellowship (Call 568 from 2012 Resolution 01,218 Bogotá, Colombia). The SMFA works were supported in part by the intramural program of the National Institute of Allergy and Infectious Diseases/NIH. This work was funded by PATH’s Malaria Vaccine Initiative under Grant No. OPP48782 from the Bill & Melinda Gates Foundation.
Footnotes
Appendix A. Supplementary data
Supplementary material related to this article can be found, in the online version, at doi:https://doi.org/10.1016/j.molbiopara.2018.07.009.
References
- [1].Flick K, et al. , Optimized expression of Plasmodium falciparum erythrocyte membrane protein I domains in Escherichia coli, Malar. J. (3) (2004). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [2].Tuju J, et al. , Vaccine candidate discovery for the next generation of malaria vaccines, Immunology 152 (2) (2017) 195–206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [3].Othman AS, et al. , The use of transgenic parasites in malaria vaccine research, Expert Rev. Vaccines 16 (7) (2017) 685–697. [DOI] [PubMed] [Google Scholar]
- [4].Ntege EH, et al. , Blood-stage malaria vaccines: post-genome strategies for the identification of novel vaccine candidates, Expert Rev. Vaccines 16 (8) (2017) 769–779. [DOI] [PubMed] [Google Scholar]
- [5].Klein MM, et al. , The cysteine-rich interdomain region from the highly variable Plasmodium falciparum erythrocyte membrane protein-1 exhibits a conserved structure, PLoS Pathog. 4 (9) (2008). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [6].Thompson J, et al. , Plasmodium cysteine repeat modular proteins 1–4: complex proteins with roles throughout the malaria parasite life cycle, Cell. Microbiol. 9 (6) (2007) 1466–1480. [DOI] [PubMed] [Google Scholar]
- [7].MacDonald NJ, et al. , Structural and immunological characterization of recombinant 6-Cysteine domains of the plasmodium falciparum sexual stage protein Pfs230, J. Biol. Chem. 291 (38) (2016) 19913–19922. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [8].Longley RJ, et al. , Comparative assessment of vaccine vectors encoding ten malaria antigens identifies two protective liver-stage candidates, Sci. Rep. 5 (2015). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [9].Miguel-Blanco C, et al. , Hundreds of dual-stage antimalarial molecules discovered by a functional gametocyte screen, Nat. Commun. 8 (2017). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [10].van Dijk MR, et al. , A central role for P48/45 in malaria parasite male gamete fertility, Cell 104 (1) (2001) 153–164. [DOI] [PubMed] [Google Scholar]
- [11].van Dijk MR, et al. , Three members of the 6-cys protein family of plasmodium play a role in gamete fertility, PLoS Pathog. 6 (4) (2010). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [12].Annoura T, et al. , Two Plasmodium 6-Cys family-related proteins have distinct and critical roles in liver-stage development, Faseb J. 28 (5) (2014) 2158–2170. [DOI] [PubMed] [Google Scholar]
- [13].Mistarz UH, et al. , Expression, purification and characterization of GMZ2’.10C, a complex disulphide-bonded fusion protein vaccine candidate against the asexual and sexual life-stages of the malaria-causing plasmodium falciparum parasite, Pharm. Res. 34 (9) (2017) 1970–1983. [DOI] [PubMed] [Google Scholar]
- [14].Theisen M, Jore MM, Sauerwein R, Towards clinical development of a Pfs48/45-based transmission blocking malaria vaccine, Expert Rev. Vaccines 16 (4) (2017) 329–336. [DOI] [PubMed] [Google Scholar]
- [15].Outchkourov N, et al. , Epitope analysis of the malaria surface antigen Pfs48/45 identifies a subdomain that elicits transmission blocking antibodies, J. Biol. Chem. 282 (23) (2007) 17148–17156. [DOI] [PubMed] [Google Scholar]
- [16].Vermeulen AN, et al. , Sequential expression of antigens on sexual stages of plasmodium-falciparum accessible to transmission-blocking antibodies in the mosquito, J. Exp. Med. 162 (5) (1985) 1460–1476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Milek RLB, et al. , Plasmodium falciparum: heterologous synthesis of the transmission-blocking vaccine candidate Pfs48/45 in recombinant vaccinia virus-infected cells, Exp. Parasitol. 90 (2) (1998) 165–174. [DOI] [PubMed] [Google Scholar]
- [18].Milek RLB, Stunnenberg HG, Konings RNH, Assembly and expression of a synthetic gene encoding the antigen Pfs48/45 of the human malaria parasite Plasmodium falciparum in yeast, Vaccine 18 (14) (2000) 1402–1411. [DOI] [PubMed] [Google Scholar]
- [19].Milek RLB, et al. , Immunological properties of recombinant proteins of the transmission blocking vaccine candidate, Pfs48/45, of the human malaria parasite Plasmodium falciparum produced in Escherichia coli, Parasite Immunol. 20 (8) (1998) 377–385. [DOI] [PubMed] [Google Scholar]
- [20].Outchkourov NS, et al. , Correctly folded Pfs48/45 protein of Plasmodium falciparum elicits malaria transmission-blocking immunity in mice, Proc. Natl. Acad. Sci. U.S.A. 105 (11) (2008) 4301–4305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].Lin JW, et al. , A novel’ Gene Insertion/Marker out’ (GIMO) method for transgene expression and gene complementation in rodent malaria parasites, Plos One 6 (12) (2011). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [22].van Dijk MR, et al. , Three members of the 6-cys protein family of Plasmodium play a role in gamete fertility, PLoS Pathog. 6 (4) (2010) e1000853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [23].Otto TD, et al. , A comprehensive evaluation of rodent malaria parasite genomes and gene expression, BMC Biol. 12 (2014). [DOI] [PMC free article] [PubMed] [Google Scholar]
- [24].Janse CJ, Waters AP, Plasmodium-berghei - the application of cultivation and purification techniques to molecular studies of malaria parasites, Parasitol. Today 11 (4) (1995) 138–143. [DOI] [PubMed] [Google Scholar]
- [25].Theisen M, et al. , A multi-stage malaria vaccine candidate targeting both transmission and asexual parasite life-cycle stages, Vaccine 32 (22) (2014) 2623–2630. [DOI] [PubMed] [Google Scholar]
- [26].Eksi S, et al. , Malaria transmission-blocking antigen, Pfs230, mediates human red blood cell binding to exflagellating male parasites and oocyst production, Mol. Microbiol. 61 (4) (2006) 991–998. [DOI] [PubMed] [Google Scholar]
- [27].Triller G, et al. , Natural parasite exposure induces protective human anti-malarial antibodies, Immunity 47 (6) (2017) 1197-+. [DOI] [PMC free article] [PubMed] [Google Scholar]
- [28].Bijker EM, et al. , Novel approaches to whole sporozoite vaccination against malaria, Vaccine 33 (52) (2015) 7462–7468. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.