


Abstract
Replication Protein A (RPA) is required for DNA recombination, repair and replication in all eukaryotes. RPA participation in these pathways is mediated by single-stranded DNA binding and protein interactions. We herein identify a novel protein, Replication Protein Binding Trans-Activator (RBT1), in a yeast two-hybrid assay employing the second subunit of human RPA (RPA32) as bait. RBT1–RPA32 binding was confirmed by glutathione S-transferase pull-down and co-immunoprecipitation. Fluorescence microscopy indicates that green fluorescence protein-tagged RBT1 is localized to the nucleus in vivo. RBT1 mRNA expression, determined by semi-quantitative RT–PCR, is significantly higher in cancer cell lines MCF-7, ZR-75, SaOS-2 and H661, compared to the cell lines normal non-immortalized human mammary epithelial cells and normal non-immortalized human bronchial epithelial cells. Further, yeast and mammalian one-hybrid analysis shows that RBT1 is a strong transcriptional co-activator. Interestingly, mammalian transactivation data is indicative of significant variance between cell lines; the GAL4–RBT1 fusion protein has significantly higher transcriptional activity in human cancer cells compared to human normal primary non-immortalized epithelial cells. We propose that RBT1 is a novel transcriptional co-activator that interacts with RPA, and has significantly higher activity in transformed cells.
INTRODUCTION
Replication Protein A (RPA) is a ubiquitous and abundant heterotrimeric protein composed of 70, 32 and 11 kDa subunits and is required for DNA replication, repair and recombination (1–6). In yeast, RPA is found to bind specific regulatory sequences in the promoters of DNA repair and metabolism genes (7) suggesting that RPA may have a role in the regulation of transcription (8). Previous studies have demonstrated that RPA function requires an ability to bind and destabilize dsDNA and that its specific activity is mediated by protein interaction and/or RPA phosphorylation (9). RPA–protein interactions appear to be largely mediated by the large 70 kDa subunit (RPA70). Of note, RPA70 has been reported to interact with p53, GAL4, VP16, EBNA1, SV40 large T-antigen and DNA polymerase α (9–14). RPA binding is also important for the proper function of DNA repair proteins involved in damage recognition and excision—interaction with Xeroderma Pigmentosum Complementation Group F (XPF) stimulates its 5′ junction-specific endonuclease activity and interaction with XPG targets this endonuclease to damaged DNA (15,16).
The possibility of interaction by the aforementioned repair proteins with the second subunit of RPA (RPA32) has not been clearly elucidated. However, interactions with some DNA repair proteins appear to be mediated by RPA32, such as interaction with XPA and uracil-DNA glycosylase (UDG) (17,18). A region of homology between XPA and UDG is suggestive of a common binding motif to RPA32. Furthermore, some important protein interactions, such as with RAD52, appear to involve all three subunits of RPA (19).
The physiological role of RPA phosphorylation is less clearly understood. RPA phosphorylation occurs on amino acid residues proximal to the N-terminal of RPA32 resulting in retardation in the electrophoretic mobility of this subunit (20–22). Although RPA is phosphorylated throughout the S phase, in response to radiation and during apoptosis, mutations of the major phosphorylation sites do not produce a clear, detectable phenotype in the model eukaryote, Saccharomyces cerevisiae, nor do they affect DNA binding, SV40 DNA replication in vitro or nucleotide excision repair (9,20,23–30). It is plausible that phosphorylation, rather than being an absolute requirement for physiological function, affects the conformation of RPA modulating its affinity for DNA and its protein interactors, and altering the balance between DNA replication and repair (9). It is interesting to note that the interaction between RPA and p53 is disrupted by UV damage suggesting at least one mechanism by which DNA replication and repair are co-ordinated with checkpoint controls (31).
In the present study, protein–protein interactors of RPA32 were screened by the yeast two-hybrid assay. We describe a novel protein interactor of RPA32 which we refer to as Replication Protein Binding Trans-Activator 1 (RBT1). RBT1, a 196 amino acid protein, is able to transactivate reporter genes in both yeast and mammalian one-hybrid experiments.
MATERIALS AND METHODS
Plasmids
RPA32 nucleotide sequence was PCR amplified from cDNA derived from cell line MCF-7 and cloned into the yeast two-hybrid plasmids pBTM116 and pACT2 in frame to LexA (1–202) and GAL4-TA, respectively. Similarly, both XPA and UDG nucleotide coding sequences were PCR amplified from cDNA derived from cell line normal non-immortalized human mammary epithelial cells (NHMEC) and cloned in frame into pBTM116 and pACT2.
RBT1, obtained from the yeast two-hybrid screen, was subcloned from pACT2 into pBTM116. For purposes of cloning truncation plasmids, RBT1 was PCR amplified from IMAGE hEST clone, locus AI003615, and cloned into both pBTM116 and pACT2. Plasmids RBT1-ΔC16, RBT1-ΔC36, RBT1-ΔC57 and RBT1-ΔC79 refer to the amino acid deletion of each RBT1 construct truncated from the C-terminal end.
Similarly, mammalian one-hybrid constructs of RBT1 were cloned into pSG424 subsequent to PCR using the IMAGE hEST clone as template. Plasmids RBT1-ΔC16, RBT1-ΔC36, RBT1-ΔC57 and RBT1-ΔC84 refer to truncations from the C-terminal of RBT1. The RBT1 clone obtained initially from the yeast two-hybrid screen was also cloned into pSG424 by digestion with BamHI and ligation; orientation was confirmed by restriction digest, and this construct, referred to as RBT1-ΔN22, has a deletion of 22 amino acids from the RBT1 N-terminus. The clone representing the entire open reading frame of RBT1 fused to GAL4 in pSG424 is referred to as RBT1. The sequence for the 22 amino acids proximal to the C-terminus of RBT1 was cloned from the PCR product generated using primers RBT-5-TD (5′-CAACGAATTCTGTGCCCCAGGTTCTTGGGA-3′) and RBT-MAM-3-F (5′-CCAGAGTTGCATTCAGGGATCCAGG-3′) and is called RBT1-TD22.
The GAL4-LUC and GAL4-TLS (positive control) plasmids were obtained from Dr Rongtuan Lin (Lady Davis Institute, Montreal, Canada) (32). The full RBT1 coding sequence was cloned in frame into pEGFP-C1 (Clontech, Palo Alto, CA) to yield RBT1–GFP.
The RBT1 and RPA14 clones obtained from the yeast two-hybrid screen were subcloned by digesting with SmaI and XhoI and ligating into the respective sites in plasmid pGEX-4T-2 to yield plasmids pRBT1-GST and pRPA14-GST, respectively.
RPA32 was cloned into Bluescript SK+ (Stratagene, La Jolla, CA) for subsequent use as template for in vitro translation. The hEST clone AI003615 was digested with EcoRI and BamHI, ligated into Bluescript SK+ also for purposes of in vitro translation.
Primer sequences for semi-quantitative RT–PCR are: RBT-SQ-5A (5′-TCCTCATGTCATCTTCGGTGG-3′), RBT-SQ-3B (5′-GGTTTCCACCCTATTGCACG-3′), GAPDH-SQ-5A (5′-CCATGGAGAAGGCTGGGG-3′) and GAPDH-SQ-3B (5′-CAAAGTTGTCATGGATGACC-3′).
All cDNA clones were sequenced using a dye primer cycle sequencing kit (Perkin-Elmer, Norwalk, CT) at Guelph Molecular Supercentre (University of Guelph, Guelph, Canada) using a DNA sequencer (Perkin-Elmer ABI 377).
cDNA library screening
The pLexA-RPA32 yeast two-hybrid plasmid construct was transformed into yeast strain L40 [MATa trp1 leu2 his3 URA3::(lexAop)8-lacZ LYS2::(lexAop)4-HIS3 lys2 ura3 ade2 gal80 gal4] prior to library transformation. A human osteosarcoma cDNA library was obtained as a gift from Dr Karaplis (Lady Davis Institute, Montreal, Canada) and amplified according to the Clontech recommended protocol. All yeast transformations were done using lithium acetate as previously described (33).
Cell line and cell culture
Cell lines NHMEC, NHBEC (normal non-immortalized human bronchial epithelial cells) and PrEC (normal non-immortalized human prostate epithelial cell line) were maintained in medium supplemented by the manufacturer, Biowhittaker (Walkersville, MD). B2BN (normal bronchial epithelial cell line immortalized with SV40 large T-antigen) obtained from Dr B. I. Gerwin (NCI, Bethesda, MD), PC3 (human prostate adenocarcinoma cell line), SaOS-2 (human osteosarcoma cell line), ZR-75, MCF-7, MDA-231 and Hs578T (human breast adenocarcinoma cell lines), H460, H661 and H322 (human non-small cell lung cancer cell lines), HEK293 (human normal embryo kidney cells), Hep3B (human hepatoblastoma cell line) and NIH3T3 (mouse fibroblasts) were cultured in American Tissue Culture Collection (ATCC, Rockville, MD) recommended media containing 5 or 10% fetal bovine serum (FBS) and supplemented with 100 U/ml penicillin and 100 µg/ml streptomycin. All cell lines were maintained in culture at 37°C in an atmosphere of 5% CO2.
Semi-quantitative RT–PCR
Cell lines were grown in appropriate media to 80% confluence, collected and RNA was isolated using the High Pure RNA Isolation Kit (Boehringer Mannheim, now Roche Diagnostics, Laval, Canada). RNA from each sample (5 µg of total) was used for synthesis of cDNA using the First Strand cDNA Synthesis Kit (MBI Fermentas, Amherst, NY) where the reverse transcriptase reaction was primed with the included oligo (dT)18 primer. PCR was performed using 1 µl of a 1000-fold dilution of each first strand cDNA reaction mix and included primers for both RBT1 (RBT-SQ-5A and RBT-SQ-3B) and GAPDH (GAPDH-SQ-5A and GAPDH-SQ-3B). [α-32P]dCTP was added to each PCR reaction tube and 20 cycles performed (94°C for 15 s, 55°C for 30 s, 72°C for 40 s). Aliquots of 2 µl from each PCR reaction mix were loaded on a 6% acrylamide sequencing gel, electrophoresed for 2 h and exposed to film. Band intensity was quantitated using the software program, NIH Image.
Localization
To analyze the subcellular localization of RBT1, a green fluorescence protein (GFP) fusion-based method was employed. MDA-231 cells were transfected using LipofectAMINE reagent according to the manufacturer’s recommendations (Gibco BRL, Rockville, MD) with an expression plasmid coding for EGFP alone (pEGFP-C1) or for an EGFP–RBT1 (pRBT1-GFP) fusion protein. After 48 h, transfected cells were washed with phosphate buffered saline (PBS) and fixed in 3% paraformaldehyde/PBS. Subsequent to fixation, cells were incubated for 1 h with PBS containing 2% BSA, 2% normal goat serum and 0.2% gelatine at room temperature to reduce non-specific binding of the RPA32 antibody. All washing and incubations with both primary (RPA32) and secondary (Texas Red conjugated) antibodies were done in PBS containing 0.2% BSA. After labeling, the coverslips were mounted in Airvol (Air Products and Chemicals Inc., Allentown, PA) and photographed with a Zeiss Axiophot fluorescent microscope. Monoclonal antibody against RPA32 was purchased from Oncogene Research Products (Cambridge, MA).
Transactivation assays
Yeast. L40 cells were transformed with plasmid(s) and assayed for LacZ reporter activity by using the liquid culture β-galactosidase assay using o-nitrophenyl B-d-galactopyranoside (ONPG) as substrate as described in Clontech Matchmaker protocols. All experiments were done in triplicate.
Mammalian. Mammalian cell transfections were performed using the LipofectAMINE (Gibco BRL) reagent according to the manufacturer’s recommendations; 0.36 µg of reporter plasmid (p5XUPS-GAL4-LUC), 0.36 µg of a given RBT1-pSG424 fusion plasmid or pSG424 empty vector or pGAL4-TLS positive control and 0.08 µg pRL-TK (Renilla luciferase reporter under the control of a TK promoter) internal control plasmid for a total of 0.8 µg DNA in 12 well plates. Transfections were done in triplicate and repeated 3–5 times. Twenty-four hours after transfection, cells were washed with PBS without Ca2+ or Mg2+ and harvested with Passive Lysis Buffer (Promega, Madison, WI). Luciferase assays were performed using reagents from the Dual-Luciferase Reporter Assay System (Promega) and with an EG&G Berthold model Lumat LB9507 luminometer. Relative luciferase activity was calculated and reported as a ratio between firefly luciferase and Renilla luciferase activity.
GST pull-down assay and in vitro translation
Glutathione S-transferase (GST), GST–RBT1 and GST–RPA14 were expressed and isolated from Escherichia coli DH5α following a 3 h induction with 1 mM isopropyl-β-d-thiogalactopyranoside (Pharmacia) at 37°C according to the Pharmacia recommended protocol; subsequent to lysis, GST fusion proteins were incubated for 1 h with glutathione–Sepharose beads (Pharmacia), the beads were washed three times with PBS/1% Triton X-100 and once with PBS. Captured GST fusion proteins were used for the GST pull-down assay.
RPA32 was in vitro translated in a reaction mixture containing [35S]methionine (Amersham) using a TnT coupled reticulocyte lysate system (Promega) according to the manufacturer’s instructions. The translated product was incubated with the GST fusion proteins and GST alone and subsequently washed. Binding was analyzed by 10–20% SDS–PAGE and subsequent exposure to film. RBT1 was in vitro translated in a similar manner to RPA32.
Protein extraction, immunoprecipitation
Mammalian cells were lysed with 10 vol of lysis buffer [50 mM Tris–HCl (pH 8.0), 150 mM NaCl, 0.1 mM EDTA, 0.5% Nonidet P-40, 10% glycerol, 1 mM dithiothreitol, 0.2 mM phenylmethylsulfonyl fluoride, aprotinin (3 µg/ml), pepstatin (1 µg/ml), leupeptin (1 µg/ml), 1 mM Na3VO4 and 10 mM NaF] and the supernatant was collected after centrifugation.
For immunoprecipitation, 400 µg of protein was mixed with 30 µl of Protein G-Sepharose and either anti-RPA32 mouse monoclonal antibody, anti-RBT1 rabbit polyclonal antibody (generated against purified GST–RBT1 protein at the McGill Animal Resource Centre, Montreal, Canada) or non-specific rabbit IgG antibody overnight at 4°C. Beads were pelleted, boiled in SDS sample buffer and the immunoprecipitates were subjected to 15% SDS–PAGE. Subsequent to transfer onto a polyvinylidene difluoride membrane (Millipore, Bedford, MA), the blot was incubated with anti-RPA32 monoclonal antibody for 2 h at room temperature, washed and visualized by enhanced chemiluminescence (Amersham).
RESULTS
A yeast two-hybrid screen of a human osteosarcoma GAL4 cDNA library using LexA-RPA32 as bait yielded 15 putative positive colonies from a total of 400 000 colonies assayed. Eight of the interactors were identified as the 14 kDa subunit of RPA (RPA14) by PCR and/or cycle sequencing. RPA32–RPA14 binding represents the highest affinity of interaction in this system based on the vigorous growth of the colonies on dropout media lacking histidine (containing 25 mM 3-amino-1,2,4-triazole) and strong β-gal activity; the isolation of RPA14 suggested that RPA32 is faithfully translated in yeast and that its conformation is not compromised.
RBT1 sequence
A putative positive identified as a probable RPA32 interacting protein was sequenced and found to have no strong homology to any other previously characterized protein and is subsequently referred to as RBT1. The original clone obtained by yeast two-hybrid was fully sequenced and used in a BLAST search of human ESTs (expressed sequence tags). Sequence overlap was used to pick AI003615 as representing the full coding sequence of RBT1 and part of the 5′-UTR and 3′-UTR. The full RBT1 sequence is derived from automated sequencing of human EST clone locus AI003615. The nucleotide sequence of clone AI003615 corresponds to the clone obtained by yeast two-hybrid except that this latter clone is truncated at the N-terminal and putatively reads from amino acid 22 (P-A-G-L-Q-S....). The RBT1 cDNA has an open reading frame of 196 amino acids; the nucleotide and amino acid sequence is shown in Figure 1A. In vitro translated RBT1 migration corresponded to the theoretical protein size of ∼22 kDa and this corresponded with western blot analysis using the generated polyclonal anti-RBT1 antibody (data not shown).
Figure 1.

Structure of RBT1 cDNA. (A) The nucleotide sequence of RBT1 cDNA and the deduced amino acid sequence (GenBank accession no. AF192529). (B) SH3-like consensus binding sites, where X tends to be alipathic (G, A, V, L, I), the two prolines (P) are necessary for high affinity binding and the scaffolding residue (p) is often proline.
Although RBT1 does not share significant homology with previously characterized proteins, it does contain four motifs with proline residues arranged in a manner similar to the core consensus sequence for SH3 binding proteins or the PY motif of WW domain-binding proteins (Fig. 1B). These motifs are arranged pseudo-symmetrically with the first and fourth SH3-like binding motifs sharing the common sequence P-P-E-P, and the second and third P-P-Q. Also, a W-E-W motif is evident 19 amino acids from the N-terminal and 15 amino acids from the C-terminal end of RBT1. In concert with the SH3-like binding motifs, RBT1 may be a protein with ‘mirrored symmetry’ possibly important for protein–protein interactions.
GST pull-down assay and co-immunoprecipitation
The binding of RPA32 to RBT1 was analyzed by performing a GST pull-down assay. As shown in Figure 2, the GST–RBT1 and GST–RPA14 bind to RPA32 in vitro, whereas GST alone does not bind. RPA32 pulled down by GST–RPA14 appears significantly stronger than that of GST–RBT1; this was as expected since the affinity of interaction between RPA32 and RPA14 is extremely strong. Further, we verified that RPA32 had specificity in this binding assay by including in one experiment GST-tagged Basal Transcription Factor 3 (BTF3) and IKKγ both of which did not pull down the in vitro translated RPA32 (data not shown).
Figure 2.
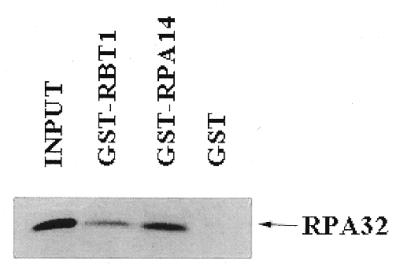
GST pull-down. Input of labeled RPA32 is shown in the first lane. The fraction of labeled RPA32 pulled down by GST–RBT1, GST–RPA14 and GST alone, respectively, are shown in the following lanes.
Co-immunoprecipitation using anti-RBT1 antibody and revealing with anti-RPA32 antibody also showed RBT1 binding to RPA32 (Fig. 3); the presence of RBT1 in the anti-RPA32 immunoprecipitate was not demonstrated because of significant background on the immunoblot. While the proportion of RPA32 brought down by the anti-RBT1 polyclonal antibody is less than that of RPA14, it was able to bind more RPA32 than the same amount of anti-XPA polyclonal antibody, a known interactor of RPA32 (data not shown). RPA32 binding to RPA14 has extremely high affinity, but this does not preclude RPA32 binding to other proteins with moderate affinity.
Figure 3.

Co-immunoprecipitation. 293 kidney cell extracts were incubated with anti-RPA32, anti-RBT1 and non-specific polyclonal IgG (negative control) antibody, respectively; these were done in duplicate. Lanes labeled ‘None’ contain 100 µg of protein extract as a positive control for the anti-RPA32 antibody used to immunoblot this membrane.
RBT1 mRNA size and semi-quantitative RT–PCR
A single transcript of ∼1.6 kb was detected by northern blot analysis in cell line H322 (data not shown). Semi-quantitative PCR and analysis of the clonal origins of the hESTs of RBT1 in GenBank suggest that RBT1 is expressed in various human tissues, but with variance in abundance. We noted a high frequency of EST matches from cancer cell lines and sought to ascertain whether there was a difference in expression between cancer and normal cell lines. Indeed, RBT1 expression levels appear to be significantly higher in all cancer cell lines examined by semi-quantitative RT–PCR in comparison to non-transformed cell lines (Fig. 4). Levels of RBT1 mRNA appear to be at least 5–10 times more abundant in cancer cell lines MCF-7, ZR-75, SaOS-2 and H661 as compared to both NHMEC and NHBEC normalizing to GAPDH mRNA.
Figure 4.

Semi-quantitative RT–PCR. RBT1 mRNA expression normalized to GAPDH mRNA in cell lines NHMEC, MCF-7 and ZR-75, SaOS-2, NHBEC and H661.
Localization of RBT1
Cells transfected with GFP alone displayed diffuse green fluorescence throughout the cells. On the other hand, a distinct nuclear fluorescent pattern was observed in cells expressing GFP–RBT1 (Fig. 5). RPA32 immunofluorescence overlapped GFP–RBT1 demonstrating that the fluorescence is localized to the nucleus. A putative nuclear localization sequence, K-R-K-H, is also found near the N-terminal end of RBT1.
Figure 5.

Nuclear localization of RBT1. (Top row) Diffuse expression of GFP in MDA231 cell line, RPA32 localization defining the nucleus and phase contrast photograph of same cells. (Bottom row) Expression of RBT1–GFP and RPA32 localization to the nucleus and phase contrast photograph of same cells.
Yeast one-hybrid and two-hybrid data
Interactions between RPA32 and RPA14, RPA32 and XPA, and RPA32 with UDG were confirmed by yeast two-hybrid (Table 1 and Fig. 6). The protein product of pACT2-RBT1 did not show interaction with the protein products of pBTM116, pLexA-UDG or pLexA-RPA14 suggesting that it is not a spurious positive interactor with RPA32.
Table 1. Yeast two-hybrid assays.
| DNA-binding domain | Activation domain plasmid | Growth |
|---|---|---|
| (pBTM116) | (pACT2) | (colony formation) |
| RPA32 | – | |
| RPA32 | – | |
| RPA32 | RPA14 | ++++ |
| RPA14 | RPA32 | ++++ |
| XPA | ++++ | |
| XPA | – | |
| XPA | RPA32 | ++++ |
| RPA32 | XPA | +++ |
| UDG | – | |
| UDG | – | |
| UDG | RPA32 | +++ |
| RPA32 | UDG | +++ |
| UDG | RBT1 | – |
| RBT1 | +++++ | |
| RBT1-ΔC16 | ++++ | |
| RBT1-ΔC36 | +++ | |
| RBT1-ΔC57 | ++ | |
| RBT1-ΔC79 | – | |
| RBT1 (pAS2-1; strain Y190) | +++++ | |
| RBT1 | – | |
| RBT1-ΔC16 | – | |
| RBT1-ΔC36 | – | |
| RBT1-ΔC57 | – | |
| RBT1-ΔC79 | – | |
| RBT1 | RPA32 | ++++ |
| RPA14 | RBT1 | – |
| RPA32 | RBT1 | +++ |
| RPA32 | RBT1-ΔC16 | +++ |
| RPA32 | RBT1-ΔC36 | +++ |
| RPA32 | RBT1-ΔC57 | +++ |
| RPA32 | RBT1-ΔC79 | +++ |
Growth on histidine deficient media containing 25 mM 3-amino-1,2,4-triazole determined qualitatively by vigour of colony formation. Where two proteins are assayed for interaction, the selection media is also deficient for tryptophan, leucine and histidine. Where a protein is assayed for activation of the HIS3 reporter gene by itself, the media is deficient for either tryptophan or leucine and histidine. Plus signs refer to how quickly and the vigour with which the resultant colonies grow.
Figure 6.

β-gal activity in yeast one-hybrid and two-hybrid assays. (Top graph) Comparison of β-gal activity of RPA32 as bait: alone, with RBT1, with UDG and with XPA. (Second graph) Comparison of β-gal activity of RBT1 in the bait plasmid with RPA32 and RPA32 as bait with RPA14. (Third graph) Comparison of β-gal activity of RPA32 in pACT2: alone, with UDG and with XPA. Also shown are activities of UDG and XPA in the bait plasmid alone. Note the strong transactivation of XPA by itself. (Bottom graph) Comparison of β-gal activity of full-length RBT1 with truncations of RBT1 from the putative C-terminal domain. Also included are RBT1–RPA32 and RPA32–RBT1 β-gal activities as comparative reference.
RBT1 and RPA32 were shuffled into pBTM116 and pACT2, respectively, to further verify the interaction. However, we were unable to do so using pLexA-RBT1 because of extremely strong transactivation of the yeast reporter genes by itself. This observation suggested the possibility that RBT1 has characteristics of a transcriptional co-activator (Table 1 and Fig. 6). Truncations of the RBT1 sequence suggest that the transactivation domain of RBT1 lies proximal to the C-terminal end of the putative protein. Furthermore, RPA32 interacts with RBT1-ΔC79 which suggests that the binding domain lies somewhere in the N-terminal half of RBT1; RBT1-ΔC79 by itself does not activate the HIS3 reporter gene and has minimal β-gal expression.
Of note is that the fusion product of LexA-XPA alone transactivates both reporter genes approximately twice as strongly as that shown by the XPA and RPA32 interaction in yeast. The pLexA-XPA transformed yeast cells were plated on media containing leucine, but this difference does not necessarily explain the significant difference. In the context of GAL4 (BD), another group was able to show by yeast two-hybrid the physiological interaction of XPA with RPA32; to perform the yeast two-hybrid screen, XPA is not likely to have had significant reporter gene activity in the context of GAL4 (BD) by itself. Similarly, RBT1 transactivates much more strongly alone than when co-transformed with RPA32—binding appears in these cases to repress the strong transactivation by both XPA and RBT1 possibly because the GAL4 transactivation domain is weak relative to these genes or because there is conformational interference between the transactivation domains of the interacting proteins.
The choice of DNA-binding domains in the yeast two-hybrid may affect the conformation of the gene of interest and therefore impact on observed interactions or physiological effects. It is, however, unlikely that RBT1 is affected by the choice of LexA or GAL4 DNA-binding domain since both such fusions gave transcriptional promoting activities in yeast and mammalian cells, respectively; we cloned into a yeast vector (pAS2-1), RBT1 downstream from GAL4 (DB) and transformation into yeast demonstrated transactivation by the protein product of this construct (Table 1).
Mammalian one-hybrid
RBT1 in the context of the mammalian one-hybrid system demonstrated strong trans-activity in support of our yeast data. A clone representing the C-terminal 22 amino acids showed extremely high luciferase activity suggesting that much of the transactivation potential of this protein lies within this small domain (Fig. 7A). The initial clone obtained via yeast two-hybrid which lacks 22 amino acids from the putative N-terminal (RBT1-FULL) also gives relatively high activity within the context of the mammalian one-hybrid system.
Figure 7.

Relative luciferase activity of RBT1 in mammalian one-hybrid assay. (A) Activity in MCF-7 cell line. Comparison of relative luciferase activity of full-length RBT1, the C-terminal 22 amino acids of RBT1 (RBT1-TD), the initial clone obtained via yeast two-hybrid which lacks 22 amino acids from the putative N-terminal (RBT1-ΔN22) and RBT1 truncations (RBT1-ΔC16, RBT1-ΔC36, RBT1-ΔC57 and RBT1-ΔC84). (B) Activity of RBT1 in cancerous and non-transformed cell lines in comparison to TLS. Transactivation by RBT1 in cell line MCF-7 is greater than the scale provided but is the same as that shown in Figure 5A.
The activity of RBT1 was examined in several cell lines, both cancerous and non-transformed, and compared to the activity produced by GAL4-TLS. As can be seen in Figure 7B, relative luciferase activity is significantly higher in cancer cell lines in comparison to non-transformed lines. RBT1-FULL showed highest activity in MCF-7 cells. These results are not a function of transfection efficiency since the data are reported as ratios of firefly to Renilla luciferase activity and the raw data was indicative of transfection efficacy.
DISCUSSION
RPA plays a pivotal role in DNA metabolism. Its physiological role is associated with DNA destabilization, binding and recognition; it is not surprising then that it has numerous functional interactions with different proteins involved in one or more aspects of DNA replication, repair or recombination. While conventional biochemical methods have described the major interactors of RPA, our library screen suggests that there may be other physiologically relevant interactions that remain uncharacterized. RBT1 may be one of these.
We attempted to determine the protein interactors of RBT1 by shuffling the clone obtained in the screen into the bait plasmid and subsequently screening our library, but were unable because of the extremely high trans-activity produced by the LexA–RBT1 fusion protein alone. We therefore examined whether RBT1 has a functional transactivation domain. Indeed, several proteins have been serendipitously characterized as transcriptional co-activators prior to the initial yeast two-hybrid screen for protein–protein interactors; proteins such as BRCA1 and Npw38 were found to contain transcription-promoting activities in yeast and were subsequently found to have similar activities in mammalian cells (34,35).
RBT1 does not have significant homology to any previously characterized protein. In silico analysis (TBLASTN) using the RBT1 amino acid sequence did not show any EST homologs in other species with the exceptions of human and mouse. However, we note the presence of SH3-like (or WW domain-like) consensus binding sites arranged in mirrored symmetry. Interestingly, the PY motif is found in the activation domains of many transcriptions factors, including c-Jun, AP-2 and NF-E2 (36). One hypothesis for the relevance of the PY motif suggests that transcription stimulation is partially conferred by interaction with WW domain-containing proteins which themselves contain strong transactivation domains. The significance of the proline rich sites in RBT1, which are similar to the PY motif, remains to be determined, but they may be important for interaction with enhancers or repressors of its transactivation function.
The minimal transactivation domain at the C-terminal end of RBT1 has no significant homology to previously described domains that impart transcription-promoting activities. However, this domain is not particularly acidic suggesting that it is not a spurious trans-activator as can often be the case with acidic amino acid regions. Furthermore, the relative luciferase activity by this domain as well as for full-length RBT1 is at least comparable and often much higher than that observed with our positive control for transactivation, Translocated in Liposarcoma (TLS). TLS was previously described within the context of the mammalian one-hybrid system to have trans-activity similar to levels observed with GAL4 fused to VP16 (37). We used TLS as a positive control, and while trans-activity was similar in most cell lines, this was not the case in MCF-7 and Hs578T cells where RBT1 had significantly stronger activity. This suggests the possibility that, at least in some cellular contexts, RBT1 trans-activity is affected by more than one component or pathway of transcription.
While some random amino acid sequences can produce non-physiological trans-activity in the context of one-hybrid systems, the extremely high level of activity observed for RBT1 in cancer cells indicate that it is likely that the activity observed is physiologically relevant. We suggest that the short, minimal transactivation domain of RBT1, as well as the full-length RBT1, is functionally strong. Mutational analysis of this region will allow us to determine which amino acids are essential for basal transcriptional activity. While binding of co-factors may be necessary for optimal RBT1 trans-activity, the strong transactivation data obtained using the C-terminal of RBT1 suggests that it may itself interact with the basal transcription machinery.
The observed RBT1 transactivation, within the context of the mammalian one-hybrid system, is high in cancer cell lines in comparison to non-transformed cell lines; transcript level is correlated to this upregulation and this was found to be well correlated to protein levels—we were able to detect RBT1 protein in extracts from cancer cell lines but not from normal cell lines (data not shown). We suggest the possibility that RBT1 may play a role in the regulation of cell proliferation or may contribute to the maintenance or promotion of oncogenesis. Overexpression of the GAL4–RBT1 construct does not show similar levels of luciferase reporter gene activity in cancerous and non-transformed cell lines suggesting that a co-factor present in abundance in cancerous cells is necessary for optimal transactivation and/or that an inhibitor is not present or is non-functional. Since cancer cell lines usually proliferate with greater vigour, they likely have an abundance of proteins involved in replication and would be conducive to strong RBT1 transactivation.
One candidate for inhibition of RBT1 transactivation is p53. Preliminary data in this laboratory show significant inhibition of RBT1 transactivation by wild-type (wt) p53. Expression of wt p53 in several p53 null cells [SaOS-2, Hep3B and NIH-3T3(null)] results in a >50% decrease in transactivation by RBT1 (data not shown). While the mechanism of this inhibition is yet to be clearly defined, it is plausible that binding of p53 by RPA is correlated to a reduction in the affinity of binding between RBT1 and RPA or that inhibition of RPA by p53 leads to a concomitant inhibition of RBT1. It is also possible that p53 binds RBT1. These possibilities are currently being investigated in conjunction with the determination of the specificity of the inhibition of RBT1 by wt p53 versus p53 mutant forms.
RBT1 is a novel cDNA encoding a nuclear protein capable of binding RPA32 within the context of the yeast two-hybrid system, as well as GST pull-down and co-immunoprecipitation. We plan to address other significant questions such as: whether the phosphorylation state of RPA32 affects the affinity of binding to RBT1, whether RBT1 binds to the other subunits of RPA and in which aspects of cellular function including DNA metabolism and DNA repair does RBT1 play a role.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Dr A. Karaplis, Dr R. Lin, Dr H. Kwon, Dr C. DeLuca, Dr M. Katabi, Dr A. Wong, Dr M. Whiteway and Dr J. Hiscott for materials and technical assistance. This work was supported by a grant from the Canadian Institutes of Health Research and in part by the Canadian Breast Cancer Initiative of the National Cancer Institute of Canada (M.A.A.J.). J.M.C. is supported by a studentship from the Canadian Institutes of Health Research, and M.A.A.J. is a senior scientist of the ‘Fonds de Recherches en Sante du Quebec’.
DDBJ/EMBL/GenBank accession no. AF192529
REFERENCES
- 1.Wobbe C.R., Weissbach,L., Borowiec,J.A., Dean,F.B., Murakami,Y., Bullock,P. and Hurwitz,J. (1987) Proc. Natl Acad. Sci. USA, 84, 1834–1838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Wold M.S. and Kelly,T. (1988) Proc. Natl Acad. Sci. USA, 85, 2523–2527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Coverley D., Kenny,M.K., Munn,M., Rupp,W.D., Lane,D.P. and Wood,R.D. (1991) Nature, 349, 538–541. [DOI] [PubMed] [Google Scholar]
- 4.Coverley D., Kenny,M.K., Lane,D.P. and Wood,R.D. (1992) Nucleic Acids Res., 20, 3873–3880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Heyer W.D., Rao,M.R., Erdile,L.F., Kelly,T.J. and Kolodner,R.D. (1990) EMBO J., 9, 2321–2329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Moore S.P., Erdile,L., Kelly,T. and Fishel,R. (1991) Proc. Natl Acad. Sci. USA, 88, 9067–9071. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Singh K. and Samson,K. (1995) Proc. Natl Acad. Sci. USA, 92, 4907–4911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gailus-Durner V., Chintamaneni,C., Wilon,R., Brill,S. and Vershon,A. (1997) Mol. Cell. Biol., 17, 3536–3546. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Wold M.S. (1997) Annu. Rev. Biochem., 66, 61–92. [DOI] [PubMed] [Google Scholar]
- 10.He Z., Brinton,B.T., Greenblatt,J., Hassell,J.A. and Ingles,C.J. (1993) Cell, 73, 1223–1232. [DOI] [PubMed] [Google Scholar]
- 11.Li R. and Botchan,M.R. (1993) Cell, 73, 1207–1221. [DOI] [PubMed] [Google Scholar]
- 12.Dutta A., Ruppert,J.M., Aster,J.C. and Winchester,E. (1993) Nature, 365, 79–82. [DOI] [PubMed] [Google Scholar]
- 13.Lin Y.L., Chen,C., Keshav,K.F., Winchester,E. and Dutta,A. (1996) J. Biol. Chem., 271, 17190–17198. [DOI] [PubMed] [Google Scholar]
- 14.Braun K., Lao,Y., He,Z., Ingles,C. and Wold,M. (1997) Biochemistry, 36, 8443–8454. [DOI] [PubMed] [Google Scholar]
- 15.Bessho T., Sancar,A., Thompson,L. and Thelen,M. (1997) J. Biol. Chem., 272, 3833–3837. [DOI] [PubMed] [Google Scholar]
- 16.He Z., Henricksen,L.A., Wold,M.S. and Ingles,C.J. (1995) Nature, 374, 566–569. [DOI] [PubMed] [Google Scholar]
- 17.Matsuda T., Saijo,M., Kuraoka,I., Kobayashi,T., Nakatsu,Y., Nagai,A., Enjoji,T., Masutani,C., Sugasawa,K., Hanaoka,F., Yasui,A. and Tanaka,K. (1995) J. Biol. Chem., 270, 4152–4157. [DOI] [PubMed] [Google Scholar]
- 18.Nagelhus T.A., Haug,T., Singh,K.K., Keshav,K.F., Skorpen,F., Otterlei,M., Bharati,S., Lindmo,T., Benichou,S., Benarous,R. and Krokan,H.E. (1997) J. Biol. Chem., 270, 6561–6566. [DOI] [PubMed] [Google Scholar]
- 19.Hays S., Firmenich,A., Massey,P., Banerjee,R. and Berg,P. (1998) Mol. Cell. Biol., 18, 4400–4406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Henricksen L.A., Carter,T., Dutta,A. and Wold,M.S. (1996) Nucleic Acids Res., 24, 3107–3112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lee S.H. and Kim,D.K. (1995) J. Biol. Chem., 270, 12801–12807. [DOI] [PubMed] [Google Scholar]
- 22.Niu H., Erdjument-Bromage,H., Pan,Z.Q., Lee,S.H., Tempst,P. and Hurwitz,J. (1997) J. Biol. Chem., 272, 12634–12641. [DOI] [PubMed] [Google Scholar]
- 23.Din S., Brill,S.J., Fairman,M.P. and Stillman,B. (1990) Genes Dev., 4, 968–977. [DOI] [PubMed] [Google Scholar]
- 24.Fang F. and Newport,J.W. (1993) J. Cell Sci., 106, 983–994. [DOI] [PubMed] [Google Scholar]
- 25.Liu V.F. and Weaver,D.T. (1993) Mol. Cell. Biol., 13, 7222–7231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Carty M.P., Zernik-Kobak,M., McGrath,S. and Dixon,K. (1994) EMBO J., 13, 2114–2123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Boubnov N.V. and Weaver,D.T. (1995) Mol. Cell. Biol., 15, 5700–5706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fried L.M., Koumenis,C., Peterson,S.R., Green,S.L., van Zijl,P., Allalunis-Turner,J., Chen,D.J., Fishel,R., Giaccia,A.J., Brown,J.M. and Kirchgessner,C.U. (1996) Proc. Natl Acad. Sci. USA, 93, 13825–13830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Treuner K., Okuyama,A., Knippers,R. and Fackelmayer,F.O. (1999) Nucleic Acids Res., 27, 1499–1504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pan Z.Q., Amin,A.A., Gibbs,E., Niu,H. and Hurwitz,J. (1994) Proc. Natl Acad. Sci. USA, 91, 8343–8347. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Abramova N., Russel,J., Botchan,M. and Li,R. (1997) Proc. Natl Acad. Sci. USA, 94, 7186–7191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Lin R., Mamane,Y. and Hiscott,J. (1999) Mol. Cell. Biol., 19, 2465–2474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Schiestl R.H. and Gietz,D.R. (1989) Curr. Genet., 16, 339–346. [DOI] [PubMed] [Google Scholar]
- 34.Chapman M.S. and Verma,I.M. (1996) Nature, 382, 678–679. [DOI] [PubMed] [Google Scholar]
- 35.Komuro A., Saeki,M. and Kato,S. (1999) Nucleic Acids Res., 27, 1957–1965. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Yagi R., Chen,L.F., Shigesada,K., Murakami,Y. and Ito,Y. (1999) EMBO J., 18, 2551–2562. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Zinszner H., Albalat,R. and Ron,D. (1994) Genes Dev., 8, 2513–2526. [DOI] [PubMed] [Google Scholar]