

Abstract
Animal models suggest that experiencing high-stress levels induces changes in amygdalar circuitry and gene expression. In humans, combat exposure has been shown to alter amygdalar responsivity and connectivity, but abnormalities have been indicated to normalize at least partially upon the termination of stress exposure. In contrast, other evidence suggests that combat exposure continues to exert influence on exposed individuals well beyond deployment and homecoming, as indicated by longitudinal psychosocial evidence from veterans, and observation of greater health decline in veterans late in life. Accordingly, the experience of combat stress early in life may affect amygdalar responsivity late in life, a possibility requiring careful consideration of the confounding effects of aging, genetic factors, and symptoms of post-traumatic stress disorder. Here, we investigated amygdalar responsivity in a unique sample of 16 male monozygotic (MZ) twin pairs in their sixties, where one but not the other sibling had been exposed to combat stress in early adulthood. Forty years after combat experience, a generally blunted amygdalar response was observed in combat-exposed veterans compared to their non-exposed twin siblings. Spatial associations between these phenotypical changes and patterns of gene expression in the brain were found for genes involved in the synaptic organization and chromatin structure. Protein-protein interactions among the set of identified genes pointed to histone modification mechanisms. We conclude that exposure to combat stress early in life continues to impact brain function beyond the termination of acute stress and appears to exert prolonged effects on amygdalar function later in life via neurogenetic mechanisms.
Keywords: amygdala, fMRI, trauma, stress, twins, combat exposure
Introduction
Exposure to high levels of stress is thought to shape and rewire our brain through mechanisms involving the synaptic reorganization of limbic circuitry. Research findings in animal models suggest that exposure to traumatic stress induces changes in amygdalar circuitry and gene expression thought to result in remodeling of dendrites and synaptic connections in this brain region (Lori et al., 2018; McEwen et al., 2016). Still, the translation of these findings to humans is complicated by several factors. For instance, genetic factors are known to influence an individual’s response to stress (Ising and Holsboer, 2006; Stein et al., 2021), which is supported by molecular genetics studies of post-traumatic stress disorder (PTSD) (Stein et al., 2021) and by the observation that stress responses are more similar within monozygotic (MZ) as compared to dizygotic twin pairs (Federenko et al., 2004; Kirschbaum et al., 1992; Kupper et al., 2005). Controlling for the impact of genes is considerably more challenging to achieve in humans than in animal studies, where animals exposed and unexposed to traumatic events can be bred to be genetically identical. In addition to changes in DNA sequence, PTSD has also been associated with changes in epigenetics, that is, chemical changes to chromatin which control genomic transcription (Cao-Lei et al., 2022) and are thought to be sensitive to environmental influences, such as exposure to traumatic events. Notably, human and animal epigenetic studies in the context of PTSD show limited consistency in results (Cao-Lei et al., 2022). The translation of findings from animals to humans has been suggested to be constrained by differences in the heterogeneity of stressors, the complexity of stress responses, and the temporality of symptom onset, respectively (Cao-Lei et al., 2022).
Among human studies, the effects of exposure to traumatic events can be confused with the effects of PTSD, a condition marked by intrusive recollections and hyperarousal that can develop in some individuals exposed to traumatic events. Differences observed between individuals with PTSD and healthy controls unexposed to traumatic events can potentially be due to both the exposure and PTSD symptoms/diagnosis; hence, disentangling those two factors requires additional comparisons of individuals with PTSD to a control group that had been exposed to traumatic events (as defined, for example, in A1 criterion of the DSM-IV) without developing PTSD (Linnman et al., 2011; Marin et al., 2016; Shin et al., 2005). Such PTSD studies employing control groups of individuals with exposure to traumatic events without PTSD have provided strong evidence that PTSD involves a distinct neurobiological signature in its own right (Pitman et al., 2012). While allowing to distinguish between effects of exposure to traumatic events versus signs of PTSD, the above-mentioned comparisons typically involve genetic differences between the groups studied. Taken together, few studies have isolated the effect of exposure to traumatic events on brain function in humans without PTSD (Dannlowski et al., 2012; Ganzel et al., 2007; van Wingen et al., 2012a; van Wingen et al., 2011, 2012b), and even fewer studies have used a longitudinal design controlling for genetic factors to capture the evolution of changes in brain function associated with exposure to traumatic events in the absence of PTSD (van Wingen et al., 2012a; van Wingen et al., 2011, 2012b).
Psychosocial examination of military personnel suggests that the impact of combat stress on exposed individuals appears to change across the life span (Elder and Clipp, 1989; Marini et al., 2020). For example, a longitudinal study of veterans exposed to higher levels of combat stress indicated higher resilience at mid-age but significantly more negative impact of the combat stress experience (that is, stress symptoms still being present; the military experience being too painful to think about) when the same individuals were re-examined in their sixties (Elder and Clipp, 1989). Similarly, a health crossover has been described among veterans and nonveterans, with veterans showing better health around retirement age, followed by a greater age-related decline in health (Wilmoth et al., 2010). Accordingly, distinct but not mutually exclusive theories exist on the temporal dynamics of combat experience impact, predicting either immediate, adaptive, transient effects (stress evaporation hypothesis) or late, delayed, prolonged effects (stress intensification and stress resurfacing hypotheses) of combat stress on exposed individuals (Elder and Clipp, 1989). Notably, only the first, but not the latter perspective has yet been linked to a specific neural signature.
Consistent with a stress evaporation hypothesis of rapid war and postwar adaptation, van Wingen and colleagues (van Wingen et al., 2012a; van Wingen et al., 2011, 2012b) demonstrated an increase in amygdala activation in military personnel after exposure to combat (van Wingen et al., 2011), as well as a return to pre-combat baseline by approximately 16 months later (van Wingen et al., 2012a), suggesting that amygdalar responsivity adjusts to the presence versus absence of stress exposure. However, this research group also found evidence of persistent changes in functional amygdala-cingulate circuitry (van Wingen et al., 2012a) and in other brain systems (van Wingen et al., 2012b). As outlined above, it has also been posited that the combat experience may render exposed individuals vulnerable in life stages after homecoming, when facing critical life events such as social exclusion or loss of significant others later in life (Elder and Clipp, 1989). Whether such stress intensification and stress resurfacing hypotheses (Elder and Clipp, 1989) also involve a distinct neural signature remains to be tested. We sought to address this question by investigating combat-exposed individuals decades after homecoming, in order to test whether amygdalar responsivity would show signs of such exposure much later in life.
Research designs other than longitudinal ones can be used to determine the effect of trauma exposure on brain function in the absence of PTSD. Specifically, the MZ discordant twin design, which includes pairs in which one twin was exposed to traumatic events whereas the other was not (Pitman et al., 2012), may hold significant advantages over longitudinal designs when focusing on late-life consequences of the exposure. Longitudinal within-subjects designs would require following exposed individuals for decades, which would introduce problems dissociating the effects of combat stress exposure of the impact of aging and age-related decline. Although this could be addressed by comparisons to an age-matched control group also studied longitudinally, there would still be no control over genetic factors, as well as age x gene interaction effects. In contrast, age and genes are identical in MZ twin siblings studied at the same point in time, providing control for the effects of genes, age, age-related decline, and gene x age interaction.
In the present study, we included MZ twin pairs approximately 40 years after one but not the other co-twin had been deployed to combat in Vietnam. We aimed to determine late effects of combat exposure on amygdalar responsivity while controlling for potential confounders such as genetic factors, PTSD symptoms, and aging effects. Further, we intended to test for spatial similarity between phenotypic, combat exposure-induced changes in brain responsivity and patterns of gene expression (Diez and Sepulcre, 2018a; Sepulcre et al., 2018). We were particularly interested in testing whether the circumstances of identical genomes of MZ twin siblings may allow to relate putative stress-induced brain responsivity changes to epigenetic mechanisms.
Methods
Participants.
Male combat-exposed (Ex) veterans and their unexposed (Ux) monozygotic twin brothers served as participants in the study and were recruited through the Vietnam Era Twin (VET) Registry and pre-screening telephone interviews. Participants received a letter describing the study (including the MRI part), and completed and an MRI screening form on metal implants/items at the time of recruitment. Thirty-two twin pairs were initially included in the study; however, three pairs had to be excluded due to sleep apnea, incomplete data, and heterozygosity. An additional three pairs were excluded due to excessive head movement during scanning (see below). Each pair consisted of one combat veteran and co-twin who had not served in combat. As the purpose of the present study was to isolate the effect of trauma exposure on amygdalar reactivity, ten pairs were also excluded due to the presence of PTSD was indicated by the DSM-IV based Clinician-Administered PTSD-Scale (CAPS)(Weathers et al., 2001). The final sample consisted of 16 MZ non-PTSD pairs discordant for trauma exposure, each including an exposed Ex and his unexposed Ux co-twin. The study was approved by the Institutional Review Boards at the Massachusetts General Hospital and the VET Registry. Written informed consent was obtained from participants, who were reimbursed for their study participation.
Psychometric data.
No participant had a history of severe head injury, loss of consciousness for more than 10 minutes, tumor, epilepsy, cerebrovascular accidents, or neurological disorders. Michigan Alcoholism Screening Test (MAST)(Selzer, 1971) revealed alcohol abuse or at least risky drinking behavior in 7 of 32 participants (cutoff ≥ 5) (Ex: N = 4; Ux: N = 3). According to Structured Clinical Interview for DSM-IV Axis I Disorders (SCID-I; First et al., 1995), two participants fulfilled the criteria for the following conditions: Alcohol Abuse / Dependence (N = 1, Ex sibling), Substance Abuse / Dependence (Cannabis: N = 1, Ex sibling). Four of the 32 participants (N = 2 in Ex and N = 2 in Ux group) took psychopharmacological medication at the time of the study (Table 2). In addition to CAPS, MAST, and SCID, Beck Depression and Anxiety Inventories (BDI, BAI) were used to assess anxiety and depression symptoms.
Table 2.
Medication/drug use in unexposed (Ux) and exposed siblings (Ex).
| Type of medication/drug | Ex N = 16 | Ux N = 16 |
|---|---|---|
|
| ||
| Psychopharmalogical | 2 | 2 |
| Antidepressants: SSRIa/ SSNRIb | 2 | 2 |
| others | - | - |
| Neuroleptics | - | - |
| Cardiovascular medication | 10 | 12 |
| Benzodiazepinesc | 1 | - |
| Opiatesc | 1 | - |
| THCc | 2 | - |
Selective serotonin reuptake inhibitor,
Selective serotonin noradrenaline reuptake inhibitor,
as indicated by urine screen
Experimental procedure and stimuli.
Stimulus presentation has been described in detail previously (Shin et al., 2005). In brief, 28-second alternating blocks of grayscale fearful, happy, and neutral facial expressions were presented. Each face was presented for 200 milliseconds, followed by an interstimulus interval (300 milliseconds) (see Supplemental Methods for details).
fMRI data acquisition and analysis.
FMRI data were acquired on a 3.0 Tesla Siemens Magnetom Trio Tim MRI scanner (Siemens Medical Systems, Iselin, New Jersey, USA). Functional MRI data were analyzed using Statistical Parametric Mapping (SPM12; Wellcome Trust Centre for Neuroimaging, UCL Institute of Neurology, London; http://www.fil.ion.ucl.ac.uk/spm) (see Supplemental Methods). Functional images were AC-PC oriented, realigned, distortion-corrected using fieldmap scans, co-registered with structural scans, normalized to Montreal Neurological Institute (MNI) space based on segmentation of grey matter, white matter, and cerebrospinal fluid, and spatially smoothed using a full width at half maximum 6 mm 3D-Gaussian Kernel. Pairs were excluded if either sibling’s total rotational movement exceeded a critical value of 0,5° or the total translational movement exceeded 2 mm. Conditional effects were analyzed in the framework of the general linear model using SPM12, testing both for constant and adaptive responsivity (see Supplemental Methods). Contrast images were produced for all facial conditions combined (fearful, happy, and neutral) versus fixation (faces > fixation), for fearful and happy faces solely (versus fixation), as well as for the differential contrast fearful > happy used in our previous study (Shin et al., 2005), both for constant and adaptive responses.
At the group level, these whole-brain constant and adaptive response maps were analyzed separately using SPM12. Paired T-tests were used to analyze within-pair comparisons of contrast images from Ex and Ux siblings at the group level. Small volume corrections (svc) were conducted for the amygdala. The significance level was set at p < 0.05 (whole-brain family-wise error (FWE) corrected at the cluster level; FWE peak-level for amygdala small volume correction).
Associations between trauma-induced changes in brain phenotype and gene expression patterns for epigenetically active genes.
The Allen Human Brain Atlas (Hawrylycz et al., 2012) was used to test for associations/spatial similarity between brain activation changes induced by combat exposure (within-pair comparison of discordant MZ pairs) and gene expression in a previously described manner (Diez and Sepulcre, 2018b; Ortiz-Teran et al., 2017; Sepulcre et al., 2018) (see Supplemental Methods). A resulting list of genes showing high similarity scores between brain phenotypic change due to trauma and gene expression patterns were analyzed using Metascape (Zhou et al., 2019) (https://metascape.org) to determine functions and processes (Gene ontology (GO) Biological Processes, GO Molecular Functions, and GO Cellular Components) associated with positive and negative spatial similarity gene lists. Further, protein-protein interactions were tested using Metascape.
Results
Demographic and psychometric data
Participants (N=32; 16 MZ twin pairs) were aged 62.4 years (SD = 3.6) at the time of the study. No significant within-pair differences were observed regarding years of education level, marital status (Chi2(1, N=32) = 1.3, P = .24), non-clinical PTSD symptoms, anxiety symptoms, depressive symptoms, problematic drinking behavior, and traumatic experiences during childhood, whereas a significant difference was found for the total number of discrete critical life event occurrences (Table 1). There were also no significant differences regarding the use of medication or drugs (Table 2).
Table 1.
Demographic, clinical and life event data for monozygotic twin pairs exposed (Ex) and unexposed (Ux) to trauma.
| Exposed twin siblings M(SD) | Unexposed twin siblings M(SD) | Within-pair differences T Statistica | Within-pair differences P value | |
|---|---|---|---|---|
| age | 62.4 (3.6) | 62.4 (3.6) | ||
| years of education | 15.5 (3.5) | 14.6 (2.7) | 1.7 | 0.1 |
| times married | 1.3 (0.6) | 1.1 (0.8) | 0.7 | 0.5 |
| CAPS | 3.7 (5.5) | 2.3 (4.3) | .9 | 0.4 |
| MAST | 2.0 (2.6) | 2.4 (3.7) | −.5 | 0.6 |
| BAI | 2.3 (1.8) | 4.1 (6.7) | −1.1 | 0.3 |
| BDI | 4.3 (4.2) | 4.1 (3.6) | .1 | 0.9 |
| CTQ | 35.5 (8.4) | 37.1 (9.6) | −.9 | 0.4 |
| TLEQ – total number of different types of critical life events | 5.7 (2.3) | 4.9 (2.5) | 1.2 | 0.2 |
| TLEQ – total number of events evoking fear, helplessness or horror | 1.8 (2.0) | 1.9 (1.9) | −0.3 | 0.8 |
| TLEQ – total number of occurances/discrete events reported | 16.4 (7.7) | 10.4 (7.1) | 2.9 | 0.01 |
Abbreviations. SCID, Structured Clinical Interview for DSM-IV; CAPS, Clinician-Administered PTSD- Scale; CTQ, Childhood Trauma Questionnaire; MAST: Michigan Alcoholism Screening Test; BAI, Beck Anxiety Inventory; BDI, Beck Depression Inventory; TLEQ, Traumatic Life Events Questionnaire.
Paired T test testing for within-pair differences, df=15.
Constant brain responses across all facial expressions
Visual stimulation with facial expressions (fearful, happy, and neutral) revealed a significant activation cluster that included visual and fusiform areas in Ux siblings (Table 3). In their Ex siblings, a smaller significant cluster was observed (Table 3). When directly comparing Ex and Ux siblings’ constant responses across all facial expressions, a significant within-pair difference was found in the right amygdala (Table 3).
Table 3.
Amygdalar responses to fearful and happy faces in MZ twin siblings exposed (Ex) and unexposed (Ux) to trauma.
| Brain region | Exposed (Ux) | Unexposed (Ux) | Within-pair difference | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
|
|||||||||||||||||||
| MNI coordinates |
Statisticb |
MNI coordinates |
Statisticb |
MNI coordinates |
Statisticc |
||||||||||||||||
| Direction a | x | y | z | Z | ke | p FWE | Direction | x | y | z | Z | ke | p FWE | Direction | x | y | z | Z | ke | p FWE | |
|
| |||||||||||||||||||||
| faces > fixation | |||||||||||||||||||||
| Inf Occip Lobe | positive | −30 | −76 | −8 | 4.0 | 130 | .04 | ||||||||||||||
| Visual Cortex | positive | −16 | −100 | 12 | 4.8 | 1836 | .02 | ||||||||||||||
| Amygdala | Ux>Ex | 26 | −2 | −26 | 3.2 | 1 | .04 | ||||||||||||||
|
| |||||||||||||||||||||
| fearful > happy | |||||||||||||||||||||
| Amygdala | positive | −18 | −2 | −18 | 3.5 | 2 | .02 | ||||||||||||||
|
| |||||||||||||||||||||
| fearful > fixation | |||||||||||||||||||||
| Amygdala | positive | 26 | −4 | −26 | 4.3 | 4 | .002 | Ux>Ex | 26 | −4 | −26 | 4.5 | 3 | .001 | |||||||
| Visual Cortex | positive | −16 | −100 | 12 | 4.5 | 164 | .006 | ||||||||||||||
| Inf Occip Lobe | positive | −36 | −88 | −2 | 4.0 | 134 | .02 | ||||||||||||||
| Inf Occip Lobe | positive | 42 | −80 | −6 | 3.7 | 120 | .03 | ||||||||||||||
|
| |||||||||||||||||||||
| happy > fixation | |||||||||||||||||||||
| Visual Cortex | positive | −16 | −100 | 12 | 4.8 | 287 | .001 | ||||||||||||||
| Inf Occip Lobe | positive | 34 | −80 | −12 | 4.3 | 126 | .04 | ||||||||||||||
Abbreviations: Ex, exposed; Ux, unexposed; MNI, Montreal Neurological Institute; Inf, Inferior; Occip, Occipital; ke, cluster extent; FWE, family-wise error
. Direction of activation effect: positive indicates higher activity during fearful versus happy faces fearful > happy, and during facial presentation (fearful, happy) as compared to fixation.
. Significant within-group effects (corrected, pFWE < .05). Small volume corrections (svc) were performed using a bilateral anatomical amygdalar mask (WFU Pickatlas).
. Significant between-group differences (corrected, pFWE < .05). Svc performed using a bilateral anatomical amygdalar mask (WFU Pickatlas).
Constant brain responses to fearful and happy facial expressions
Ux siblings revealed significant activation to fearful faces > fixation (Figure 1 and Table 3) in the right amygdala and significant differential activation (fearful > happy) in the left amygdala (Table 3). No such activations were found in Ex siblings, and significant within-pair differences were found for fearful faces > fixation (Ux > Ex; Table 3, Figure 1). This difference remained significant (PFWE = .003) after exclusion of two pairs which included a sibling with a minor stroke and a sibling with transient ischemic attack, respectively. Trends were observed for within-pair differences comparing fearful > happy faces (Ux > Ex; MNIXYZ = −18, −2, −18; z = 2.97; Puncorrected = .001) and happy faces > fixation (Ux > Ex; MNIXYZ = −26, −2, −26; z = 2.88; Puncorrected = .002), respectively. Exploratory post hoc correlation analysis revealed no significant relationship between differences in amygdalar responsivity and differences in occurrences of critical life events (r = .15, P = .580).
FIG. 1. Within-pair differences in amygdalar responsivity.
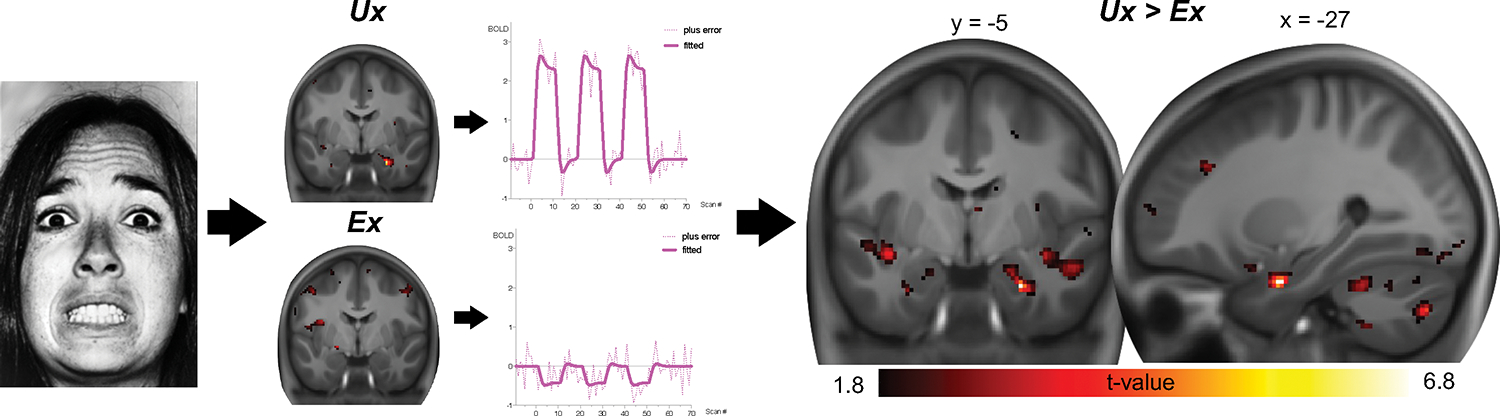
Amygdalar responsivity to fearful faces is presented for Ux and Ex siblings separately (left), revealing significant responsivity in Ux but not in Ex siblings. This difference is significant in the direct within-pair comparison (right, pFWE = 0.001, small-volume corrected). The minimum t value threshold of 1.8 used for display is equivalent to a threshold of p < 0.05, uncorrected.
Adaptive brain responses
Overall, no within-pair differences were observed for adaptive responses (Supplemental Table 1).
Self-rated emotional responsivity
Post hoc assessment of participants’ post-scan ratings of facial expressions presented during the fMRI session revealed no significant within-pair differences regarding subjective arousal and valence ratings of fearful, happy, or neutral facial expressions (all P > .22). However, when asked whether they generally responded emotionally to facial expressions, a smaller portion of Ex siblings (“Yes”: N=6; “No”: N=10) indicated to have emotionally responded to facial expressions as compared to Ux siblings (“Yes”: N=12; “No”= 4), a difference that reached significance (Chi2(1, N=32) = 4.8, P < .03). Exploratory post hoc correlational analyses indicated no direct relationship between amygdalar responsivity and subjective emotional responsivity (r = −.18, P = .52).
Brain phenotype – gene expression association analysis
Testing for the spatial similarity between changes in emotional brain activation induced by combat exposure (within-pair comparison, Ux>Ex contrast image from t-test for dependent samples) and brain patterns of gene expression resulted in one list of genes where brain expression patterns showed positive associations with the combat exposure-induced neural pattern, and one list of genes where brain gene expression was negatively associated with combat exposure-induced changes (Figure 2A). Next, positive and negative lists were analyzed using Metascape (Zhou et al., 2019), this revealed that the positive list included genes involved in synaptic organization, and postsynaptic processes in particular (Figure 2B), as well as negative associations with groups of genes with roles in reaction to toxic and arsenic substance (Supplemental Figure 1).
FIG. 2. Brain phenotype – gene expression association analysis.
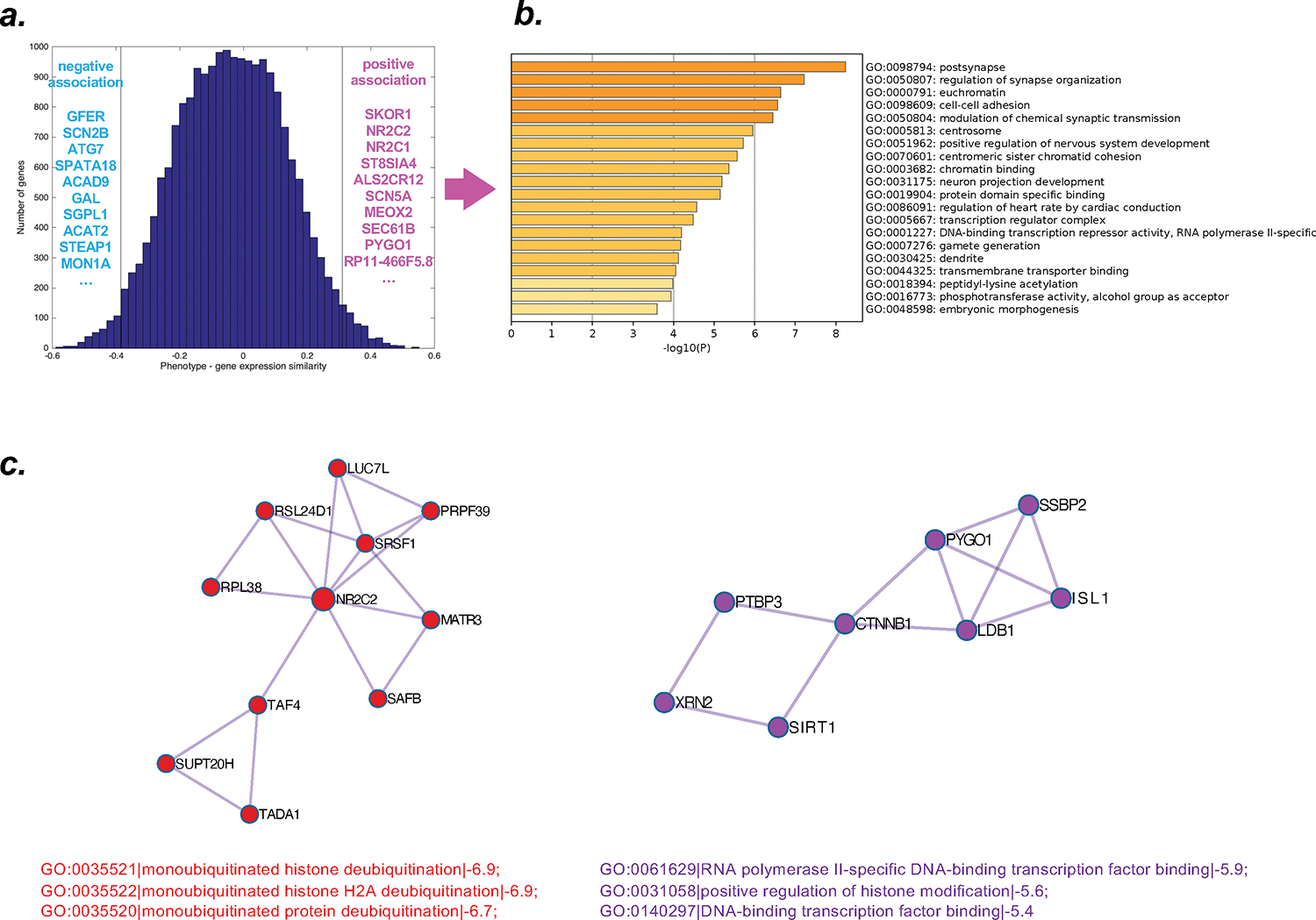
(a.) histogram displaying the extent of spatial similarity of stress-induced changes in brain responsivity (phenotype) and gene expression patterns. Both for positive and negative associations, the ten genes for which expression showed the highest similarity with the brain phenotype changes are provided (positive list/association: magenta; negative list/association: light blue). (b.) Gene ontology functions associated with the positive list of genes. (c.) two networks identified by protein-protein interaction analyses indicating histone modification/deubiquitination.
Finally, protein-protein interaction networks were formed based on GO enrichment analysis to extract “biological meanings”; the MCODE algorithm within Metascape (Zhou et al., 2019) was applied to identify neighborhoods where proteins are densely connected. For the positive list, this revealed two networks of proteins associated with neuronal communication (e.g., membrane depolarization during action potential, synaptic organization), and also two networks pointing to protein-protein interaction related to histone modification (Figure 2C), as well as additional neighborhoods of densely connected proteins (Supplemental Figure 2).
Discussion
The present study investigated whether combat stress exposure early in life influenced amygdalar responsivity late in life, using a unique sample of MZ twin pairs in their sixties where one but not the other sibling had been exposed to combat stress in early adulthood. Under strict control of key confounders such as genetic factors and PTSD symptoms, as well as age-related decline, we found a blunted amygdalar response in Ex individuals several decades after the traumatic event. The differential brain activation pattern was associated with spatial patterns of expression for genes known to be involved in synaptic transmission and chromatin structure. Analysis of the gene products of the list of identified genes (protein-by-protein interaction) pointed to histone modification, suggesting that epigenetic mechanisms may have influenced the observed changes in brain phenotype.
The observed changes were identified through within-pair comparisons of MZ twin pairs discordant for combat exposure. This study design is robust in discerning unique environmental effects, that is, responses to stressors only one but not the other sibling has experienced, while a myriad of confounders of shared environmental and genetic nature are fully controlled for. Thus, genetic variation or unmeasured factors at play when siblings grew up together (for example, household circumstances or diet) cannot be assumed to have impacted amygdalar responsivity during development. Also, we did not observe within-pair differences in retrospectively assessed childhood trauma, education, or clinical questionnaires (Table 1). Ex siblings reported significantly higher occurrence of critical life events, which could be interpreted as evidence of stress intensification or resurfacing hypotheses, but was however found to be unrelated to amygdalar responsivity. No significant differences in the use of psycho-active drugs or medications such as selective serotonin reuptake inhibitors (SSRI) were evident, which are known to dampen amygdalar responsivity to fearful faces (Murphy et al., 2009) (Table 2). This rules out possible explanations that the blunted amygdalar response observed could have been due to factors other than traumatic combat stress. Accordingly, even though we cannot ultimately exclude unmeasured non-shared factors, combat exposure is the most likely aspect that differentiates within each twin pair. Taken together, this leads us to conclude that the brain activation differences observed are most likely due to experiences or behaviors that are influenced by the fact that Ex siblings had been exposed to combat early in life.
Ux siblings showed significant amygdalar responsivity to facial stimuli, consistent with findings of numerous previous studies and meta-analysis (Fusar-Poli et al., 2009) (Table 3). Within-pair comparisons of amygdalar activation revealed hypo-responsiveness/blunted responsivity in Ex siblings, as compared to their Ux co-siblings, irrespective of face type (that is, faces > fixation), but most pronounced during the presentation of the fearful faces (Table 3). Even though no direct correlation was found between the pattern of reduced amygdalar responsivity and self-rated emotional responsivity, Ex siblings did report lower self-rated emotional responsivity to facial expressions. The blunted amygdalar responsivity observed may relate to the finding of lower cortisol levels associated with traumatic exposure (Yehuda et al., 1995), however it is worth noting that the relationship between adverse events and cortisol levels is influenced by several factors, such as features of the samples and the adversity exposure (Kalliokoski et al., 2019; Khoury et al., 2019).
Our finding of blunted amygdalar responsivity in trauma-exposed individuals without PTSD converges with results from a previous study demonstrating lower amygdalar metabolism in trauma-exposed individuals (Marin et al., 2016). However, a previous longitudinal study examining amygdalar responsivity before and after combat within a total timeframe of approximately two years observed hyperreactivity of the amygdala following trauma exposure (van Wingen et al., 2011), as well as normalization of amygdalar responsivity after combat stress. This previous and the present study are comparable regarding trauma type; individuals were exposed to combat in both studies, as well as their control of important confounders, such as genetic factors and PTSD symptoms. Subtle differences concern the experimental neuroimaging procedures of studies. Both studies used facial expressions to investigate amygdalar responsivity. However, Van Wingen et al. (van Wingen et al., 2011) included only negative facial expressions (fearful and angry) and low-level task demands. In contrast, the present study involved passive viewing of fearful, happy, and neutral facial expressions. Those differences in the experimental procedure may explain the diverging effects of combat exposure on amygdalar responsivity observed. A major difference between the studies is the amount of time that elapsed between trauma exposure and fMRI data acquisition, which may provide a much more compelling explanation. While the van Wingen et al. study investigated participants at baseline and 1.5 months after combat exposure (test-retest interval: 6.9 months)(van Wingen et al., 2011) and demonstrated normalization of amygdalar responsivity by 23 months after baseline, the present study assessed exposed individuals more than 40 years after trauma exposure. Taken together, we propose that the previous study (van Wingen et al., 2011) demonstrated normalization of amygdalar responsivity upon the termination of stress exposure in young adults, whereas the present study, which controlled for aging and aging x gene effects to identify a late neural signature of combat exposure experienced decades ago, is consistent with the notion that the stress exposure continues to exert influence on exposed individual’s later life stages.
Analysis of relationships between these late-life brain phenotypical changes and whole-brain gene expression patterns revealed significant positive associations with genes involved in synaptic (particularly postsynaptic) organization and chromatin structure (Figure 2b). Examination of the list of identified genes with respect to overlapping gene products (protein-protein interactions) suggested networks of genes that point to several processes crucial for neuronal communication (postsynapse, membrane depolarization during action potential, presynaptic active zone (Supplemental Figure 2), as well as histone modification mechanisms (Figure 2c). Accordingly, these results raise the possibility that epigenetic mechanisms may have influenced the brain changes observed as a late-life signature of early-life combat exposure.
The present study excluded twin pairs with PTSD. Our impression was that study of the specific effects of trauma exposure alone has received relatively little attention previously, and that the control of genetic factors is crucial and underappreciated in this context. In our case, additional comparison to twin pairs with PTSD would introduce the problem that between-pair differences (PTSD pairs versus non-PTSD pairs) in brain responsivity identified could be due to genetic differences (and not due to trauma exposure) as the two groups of pairs differ both regarding PTSD and genetic factors, respectively. In contrast, the effect of trauma in our within-pair comparisons of non-PTSD pairs is observed under strict control of genetic factors. Therefore, we followed the approach by van Wingen et al (van Wingen et al., 2012a; van Wingen et al., 2011, 2012b), where symptoms of PTSD were treated as a confounder and led to exclusion, and where control of genetic factors was provided by the longitudinal study design. Taken together, the Van Wingen et al studies and our data converge to suggest that traumatic combat exposure has a neural signature in its own right independent of PTSD symptoms and genetic factors. We hope that this observation and conclusion will encourage researchers to conduct more studies on the effects of traumatic exposure which are independent of PTSD symptoms.
The present study has several strengths and limitations. The advantages of the discordant MZ design in controlling the effects of genes, age, and their interaction to isolate non-shared environmental influence (combat exposure) have been mentioned in detail above. However, the study required twins to travel and undergo the MRI examination, which may have affected contacted pair’s decisions whether or not to participate. It is difficult to consider clinical implications of the findings as no within-pair differences emerged in clinical data. Future studies could examine whether the combination of resilience to PTSD and blunted neural responsivity later in life relates to the aforementioned health crossover observation in veterans (Wilmoth et al., 2010). Even though the predictor (combat exposure) preceded the outcome measurement (amygdalar responsivity) by several decades, the nature of the present study remains mainly cross-sectional. Our study design is not able to inform whether altered amygdalar responsivity was evident acutely after combat exposure and followed by normalization after stress termination as described previously (van Wingen et al., 2011). Thus, the present study is not suited to directly address the acute exposure effects predicted by the stress evaporation hypothesis. The lack of longitudinal psychosocial data precludes the identification of life circumstances and events that could have influenced amygdalar responsivity after homecoming. It remains unclear whether a stress resurfacing or stress intensification hypothesis might explain the observed pattern of results. Animal evidence indicating that chronic stress remodels dendrites and synaptic connections in the amygdala (McEwen et al., 2016) could serve as a mechanism potentially explaining the blunted amygdalar responsivity observed here in humans – however, the resolution and nature of the current data acquisition do not allow to directly address or support this assumption. Even so, the present study design was optimally suited to test whether combat exposure in young adulthood may be associated with altered amygdalar responsivity late in life. Evidence of such changes suggests that stress evaporation hypotheses should be extended concerning amygdalar responsivity and that prospective longitudinal studies are needed in order to discern additional factors that may contribute to blunted amygdalar responsivity late in life. Such factors could involve psychosocial factors (Elder and Clipp, 1989), critical life events, and/or a faster pace of biological aging (Belsky et al., 2015).
In conclusion, our findings reveal a late-life neurogenetic signature of combat stress exposure in early adulthood, which may be influenced by epigenetic mechanisms. Our finding of blunted amygdalar responsivity in Ex versus Ux twin siblings several decades after the experience of war suggests that the exposure continues to exert influence on brain function over the lifespan. This extends existing research findings concerning the effects of traumatic events on brain function (van Wingen et al., 2011).
Supplementary Material
ACKNOWLEDGEMENTS
The research reported in this publication was supported by the National Institute of Mental Health of the National Institutes of Health, award number R01MH054636. The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health. The Massachusetts General Hospital (MGH) Clinical Research Center was supported by Harvard Catalyst grants 1UL1RR025758, 8UL1TR000170, 1UL1TR001102. The MGH Athinoula A. Martinos Imaging Center was supported by the Center for Functional Neuroimaging Technologies grant P41RR14075. The creation and the ongoing development, management, and maintenance of the Vietnam- Era Twin (VET) Registry (CSP #256) is supported by the Cooperative Studies Program (CSP) of the United States Department of Veterans Affairs (VA) Office of Research & Development. All statements and opinions are solely of the authors and do not necessarily reflect the position or policy of the Vet Registry, the VA, or the United States Government. Most importantly, the authors gratefully acknowledge the continued cooperation and participation of the members of the VET Registry and their families. Without their contribution this research would not have been possible. This project was conducted, in part, by support from the University of Washington Twin Registry. We wish to thank the twins for taking part in the University of Washington Twin Registry and for their time and enthusiasm for this project. The authors thank Stacey Dubois, Andra M. Preda, Salman Y. Gany, Julia Felicione, Mary Foley, and Lawrence White for technical assistance.
Footnotes
CONFLICT OF INTEREST DISCLOSURE
The authors declare no conflict of interest.
References
- Belsky DW, Caspi A, Houts R, Cohen HJ, Corcoran DL, Danese A, Harrington H, Israel S, Levine ME, Schaefer JD, Sugden K, Williams B, Yashin AI, Poulton R, Moffitt TE, 2015. Quantification of biological aging in young adults. Proc Natl Acad Sci U S A 112(30), E4104–4110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Breiter HC, Etcoff NL, Whalen PJ, Kennedy WA, Rauch SL, Buckner RL, Strauss MM, Hyman SE, Rosen BR, 1996. Response and habituation of the human amygdala during visual processing of facial expression. Neuron 17(5), 875–887. [DOI] [PubMed] [Google Scholar]
- Cao-Lei L, Saumier D, Fortin J, Brunet A, 2022. A narrative review of the epigenetics of post-traumatic stress disorder and post-traumatic stress disorder treatment. Front Psychiatry 13, 857087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dannlowski U, Stuhrmann A, Beutelmann V, Zwanzger P, Lenzen T, Grotegerd D, Domschke K, Hohoff C, Ohrmann P, Bauer J, Lindner C, Postert C, Konrad C, Arolt V, Heindel W, Suslow T, Kugel H, 2012. Limbic scars: long-term consequences of childhood maltreatment revealed by functional and structural magnetic resonance imaging. Biol Psychiatry 71(4), 286–293. [DOI] [PubMed] [Google Scholar]
- Diez I, Sepulcre J, 2018a. Neurogenetic profiles delineate large-scale connectivity dynamics of the human brain. Nat Commun 9(1), 3876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diez I, Sepulcre J, 2018b. Neurogenetic profiles delineate large-scale connectivity dynamics of the human brain. Nature Communications 9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Elder GH Jr., Clipp EC, 1989. Combat experience and emotional health: impairment and resilience in later life. J Pers 57(2), 311–341. [DOI] [PubMed] [Google Scholar]
- Federenko IS, Nagamine M, Hellhammer DH, Wadhwa PD, Wust S, 2004. The heritability of hypothalamus pituitary adrenal axis responses to psychosocial stress is context dependent. J Clin Endocrinol Metab 89(12), 6244–6250. [DOI] [PubMed] [Google Scholar]
- First MB, Spitzer RL, Gibbon M, Williams JB, 1995. Structured Clinical Interview for DSM-IV Axis I Disorders, Patient Edition, January 1995 FINAL. SCID-I/P Version 2.0). New York, NY: Biometrics Research Department, New York State Psychiatric Institute. [Google Scholar]
- Fusar-Poli P, Placentino A, Carletti F, Landi P, Allen P, Surguladze S, Benedetti F, Abbamonte M, Gasparotti R, Barale F, Perez J, McGuire P, Politi P, 2009. Functional atlas of emotional faces processing: a voxel-based meta-analysis of 105 functional magnetic resonance imaging studies. J Psychiatry Neurosci 34(6), 418–432. [PMC free article] [PubMed] [Google Scholar]
- Ganzel B, Casey BJ, Glover G, Voss HU, Temple E, 2007. The aftermath of 9/11: effect of intensity and recency of trauma on outcome. Emotion 7(2), 227–238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hawrylycz MJ, Lein ES, Guillozet-Bongaarts AL, Shen EH, Ng L, Miller JA, van de Lagemaat LN, Smith KA, Ebbert A, Riley ZL, Abajian C, Beckmann CF, Bernard A, Bertagnolli D, Boe AF, Cartagena PM, Chakravarty MM, Chapin M, Chong J, Dalley RA, Daly BD, Dang C, Datta S, Dee N, Dolbeare TA, Faber V, Feng D, Fowler DR, Goldy J, Gregor BW, Haradon Z, Haynor DR, Hohmann JG, Horvath S, Howard RE, Jeromin A, Jochim JM, Kinnunen M, Lau C, Lazarz ET, Lee C, Lemon TA, Li L, Li Y, Morris JA, Overly CC, Parker PD, Parry SE, Reding M, Royall JJ, Schulkin J, Sequeira PA, Slaughterbeck CR, Smith SC, Sodt AJ, Sunkin SM, Swanson BE, Vawter MP, Williams D, Wohnoutka P, Zielke HR, Geschwind DH, Hof PR, Smith SM, Koch C, Grant SGN, Jones AR, 2012. An anatomically comprehensive atlas of the adult human brain transcriptome. Nature 489(7416), 391–399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ising M, Holsboer F, 2006. Genetics of stress response and stress-related disorders. Dialogues Clin Neurosci 8(4), 433–444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kalliokoski O, Jellestad FK, Murison R, 2019. A systematic review of studies utilizing hair glucocorticoids as a measure of stress suggests the marker is more appropriate for quantifying short-term stressors. Sci Rep 9(1), 11997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Khoury JE, Bosquet Enlow M, Plamondon A, Lyons-Ruth K, 2019. The association between adversity and hair cortisol levels in humans: A meta-analysis. Psychoneuroendocrinology 103, 104–117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kirschbaum C, Wust S, Faig HG, Hellhammer DH, 1992. Heritability of cortisol responses to human corticotropin-releasing hormone, ergometry, and psychological stress in humans. J Clin Endocrinol Metab 75(6), 1526–1530. [DOI] [PubMed] [Google Scholar]
- Kupper N, de Geus EJ, van den Berg M, Kirschbaum C, Boomsma DI, Willemsen G, 2005. Familial influences on basal salivary cortisol in an adult population. Psychoneuroendocrinology 30(9), 857–868. [DOI] [PubMed] [Google Scholar]
- Linnman C, Zeffiro TA, Pitman RK, Milad MR, 2011. An fMRI study of unconditioned responses in post-traumatic stress disorder. Biol Mood Anxiety Disord 1(1), 8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lori A, Maddox SA, Sharma S, Andero R, Ressler KJ, Smith AK, 2018. Dynamic Patterns of Threat-Associated Gene Expression in the Amygdala and Blood. Front Psychiatry 9, 778. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marin MF, Song H, VanElzakker MB, Staples-Bradley LK, Linnman C, Pace-Schott EF, Lasko NB, Shin LM, Milad MR, 2016. Association of Resting Metabolism in the Fear Neural Network With Extinction Recall Activations and Clinical Measures in Trauma-Exposed Individuals. Am J Psychiatry 173(9), 930–938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marini CM, Fiori KL, Wilmoth JM, Pless Kaiser A, Martire LM, 2020. Psychological Adjustment of Aging Vietnam Veterans: The Role of Social Network Ties in Reengaging with Wartime Memories. Gerontology 66(2), 138–148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McEwen BS, Nasca C, Gray JD, 2016. Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology 41(1), 3–23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy SE, Norbury R, O’Sullivan U, Cowen PJ, Harmer CJ, 2009. Effect of a single dose of citalopram on amygdala response to emotional faces. Br J Psychiatry 194(6), 535–540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ortiz-Teran L, Diez I, Ortiz T, Perez DL, Aragon JI, Costumero V, Pascual-Leone A, El Fakhri G, Sepulcre J, 2017. Brain circuit-gene expression relationships and neuroplasticity of multisensory cortices in blind children. Proceedings of the National Academy of Sciences of the United States of America 114(26), 6830–6835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pitman RK, Rasmusson AM, Koenen KC, Shin LM, Orr SP, Gilbertson MW, Milad MR, Liberzon I, 2012. Biological studies of post-traumatic stress disorder. Nat Rev Neurosci 13(11), 769–787. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Selzer ML, 1971. The Michigan Alcoholism Screening Test: The quest for a new diagnostic instrument. American journal of Psychiatry 127(12), 1653–1658. [DOI] [PubMed] [Google Scholar]
- Sepulcre J, Grothe MJ, d’Oleire Uquillas F, Ortiz-Teran L, Diez I, Yang HS, Jacobs HIL, Hanseeuw BJ, Li Q, El-Fakhri G, Sperling RA, Johnson KA, 2018. Neurogenetic contributions to amyloid beta and tau spreading in the human cortex. Nat Med 24(12), 1910–1918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shin LM, Wright CI, Cannistraro PA, Wedig MM, McMullin K, Martis B, Macklin ML, Lasko NB, Cavanagh SR, Krangel TS, Orr SP, Pitman RK, Whalen PJ, Rauch SL, 2005. A functional magnetic resonance imaging study of amygdala and medial prefrontal cortex responses to overtly presented fearful faces in posttraumatic stress disorder. Archives of general psychiatry 62(3), 273–281. [DOI] [PubMed] [Google Scholar]
- Stein MB, Levey DF, Cheng Z, Wendt FR, Harrington K, Pathak GA, Cho K, Quaden R, Radhakrishnan K, Girgenti MJ, Ho YA, Posner D, Aslan M, Duman RS, Zhao H, Department of Veterans Affairs Cooperative Studies, P., Program, V.A.M.V., Polimanti R, Concato J, Gelernter J, 2021. Genome-wide association analyses of post-traumatic stress disorder and its symptom subdomains in the Million Veteran Program. Nat Genet 53(2), 174–184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Wingen GA, Geuze E, Caan MW, Kozicz T, Olabarriaga SD, Denys D, Vermetten E, Fernandez G, 2012a. Persistent and reversible consequences of combat stress on the mesofrontal circuit and cognition. Proc Natl Acad Sci U S A 109(38), 15508–15513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Wingen GA, Geuze E, Vermetten E, Fernandez G, 2011. Perceived threat predicts the neural sequelae of combat stress. Mol Psychiatry 16(6), 664–671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Wingen GA, Geuze E, Vermetten E, Fernandez G, 2012b. The neural consequences of combat stress: long-term follow-up. Mol Psychiatry 17(2), 116–118. [DOI] [PubMed] [Google Scholar]
- Weathers FW, Keane TM, Davidson JR, 2001. Clinician-Administered PTSD Scale: A review of the first ten years of research. Depression and anxiety 13(3), 132–156. [DOI] [PubMed] [Google Scholar]
- Whalen PJ, Rauch SL, Etcoff NL, McInerney SC, Lee MB, Jenike MA, 1998. Masked presentations of emotional facial expressions modulate amygdala activity without explicit knowledge. The Journal of neuroscience : the official journal of the Society for Neuroscience 18(1), 411–418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilmoth JM, London AS, Parker WM, 2010. Military service and men’s health trajectories in later life. J Gerontol B Psychol Sci Soc Sci 65(6), 744–755. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yehuda R, Kahana B, Binder-Brynes K, Southwick SM, Mason JW, Giller EL, 1995. Low urinary cortisol excretion in Holocaust survivors with posttraumatic stress disorder. Am J Psychiatry 152(7), 982–986. [DOI] [PubMed] [Google Scholar]
- Zhou Y, Zhou B, Pache L, Chang M, Khodabakhshi AH, Tanaseichuk O, Benner C, Chanda SK, 2019. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat Commun 10(1), 1523. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.