

Abstract
Drosophila Mini spindles (Msps) protein belongs to a conserved family of microtubule-associated proteins (MAPs). Intriguingly, this family of MAPs, including Xenopus XMAP215, was reported to have both microtubule stabilising and destabilising activities. While they are shown to regulate various aspects of microtubules, the role in regulating interphase microtubules in animal cells has yet to be established. Here, we show that the depletion or mutation of Msps prevents interphase microtubules from extending to the cell periphery and leads to the formation of stable microtubule bundles. The effect is independent of known Msps regulator or effector proteins, kinesin-13/KinI homologues or D-TACC. Real-time analysis revealed that the depletion of Msps results in a dramatic increase of microtubule pausing with little or no growth. Our study provides the first direct evidence to support a hypothesis that this family of MAPs acts as an antipausing factor to exhibit both microtubule stabilising and destabilising activities.
Keywords: bundle, interphase, microtubule, pause, RNAi
Introduction
The microtubule network of eukaryotic cells plays important roles in a variety of cellular processes, including cell division, morphogenesis, polarity, migration and intracellular transport. Microtubules are polymers consisting of subunits of α- and β-tubulin, and display behaviour, described as dynamic instability, both in vivo and in vitro (Mitchison and Kirschner, 1984; Cassimeris et al, 1988; Shelden and Wadsworth, 1993). Individual microtubules independently undergo polymerisation or depolymerisation with infrequent changes between the two states. In addition, the third state, pausing, in which microtubules undergo no significant growth or shrinkage is often observed in cells (Shelden and Wadsworth, 1993; Tirnauer et al, 1999; Rusan et al, 2001), but its functional significance is yet to be understood.
In order for microtubules to fulfil their various functions within the cell, microtubules need to be organised into an array of different structures. While microtubules form only one basic structure, the bipolar spindle, in dividing cells, microtubules form a vast array of different arrangements in interphase cells depending on the cell type. Compared to spindle formation, little information is available on how interphase microtubules are organised.
A number of microtubule-associated proteins (MAPs), both motor or nonmotor proteins, collectively regulate microtubule dynamics and organisation in the cell. The Dis1/TOG family is a family of nonmotor MAPs highly conserved from yeast to plants and humans (Ohkura et al, 2001; Kinoshita et al, 2002). The Xenopus homologue, XMAP215, has been characterised in vitro as a major regulator of microtubule plus end growth (Gard and Kirschner, 1987; Vasquez et al, 1994). In addition, in Xenopus egg extracts, the protein acts as a suppressor of microtubule catastrophes by antagonising the activity of XKCM1, a member of the microtubule destabilising kinesin-13/KinI family (Tournebize et al, 2000). These two proteins are proposed to be the principal regulators of microtubule dynamics, as addition of the two proteins to pure tubulin results in microtubule dynamics similar to those seen in vivo (Kinoshita et al, 2001). Dis1/TOG proteins have long been regarded as microtubule stabilising proteins; however, recent reports indicate that, at least under some conditions, XMAP215 and the budding yeast homologue Stu2 can act as microtubule destabilising proteins (van Breugel et al, 2003; Shirasu-Hiza et al, 2003). While it is clear that the Dis1/TOG family are important regulators of microtubule dynamics, detailed understanding of the activities of the protein remains to be elucidated.
Functional studies of Dis1/TOG proteins in vivo have revealed a role for the proteins in the organisation and function of the spindle. Most of the Dis1/TOG proteins localise to the spindle poles as well as the microtubules (Wang and Huffaker, 1997; Charrasse et al, 1998; Matthews et al, 1998; Cullen et al, 1999; Graf et al, 2000; Tournebize et al, 2000; Garcia et al, 2001, Nakaseko et al, 2001). Dramatic spindle defects are seen in mutants of the Drosophila homologue, mini spindles (msps) (Cullen et al, 1999; Cullen and Ohkura, 2001). Very similar defects were subsequently described in human cells following depletion of the human homologue, TOGp (Gergely et al, 2003; Holmfeldt et al, 2004). Additionally, defects in astral microtubules are also seen in mutants of budding yeast stu2 and the worm homologue zyg-9 (Matthews et al, 1998; Usui et al, 2003). Kinetochore function has been described for the fission yeast, Dis1 and Alp14, and in budding yeast Stu2 (Garcia et al, 2001; Kosco et al, 2001; Nakaseko et al, 2001; Severin et al, 2001).
There are some indications that the Dis1/TOG family also plays a role in interphase microtubule regulation. Two temperature-sensitive alleles of the plant homologue, mor-1, have revealed a function for the protein in cortical microtubule organisation (Whittington et al, 2001). Defects in interphase microtubules have also been described in fission yeast and budding yeast (Radcliffe et al, 1998; Kosco et al, 2001; van Breugel et al, 2003). In contrast to clear evidence in yeasts and plants, the roles in regulating interphase microtubules are not well established in animal cells. Reports describing depletion of the human TOG protein state that no interphase microtubule defects were observed in cells, although microtubule dynamics were not examined (Gergely et al, 2003; Holmfeldt et al, 2004). Very recently, defects in cytoplasmic microtubules and the localisation of bicoid mRNA in Drosophila oocytes were described in msps mutants (Moon and Hazelrigg, 2004).
To understand the role of Msps protein in interphase microtubule regulation, we decided to use Drosophila culture cells, which allow the visualisation of individual microtubules and are also amenable to RNA interference (RNAi). Here we demonstrate that Msps is a major regulator of interphase microtubule organisation, and that this activity is independent of known regulators and effectors, the kinesin-13/KinI homologues and D-TACC. Further analysis of microtubule dynamics indicates that Msps acts as a microtubule antipause factor in interphase cells.
Results
Msps is associated with interphase microtubules
As the first step to understand the cellular roles of Msps during interphase, we examined Msps protein localisation in Drosophila S2 culture cells. When these cells were grown on a concanavalin A (con A)-coated surface, they spread and the interphase microtubules extend outwards to the cell periphery, enabling visualisation of individual microtubules in the flattened region of the cell (Rogers et al, 2002). S2 cells were subjected to immunostaining using antibodies against Msps (Cullen et al, 1999) and α-tubulin. During mitosis, Msps concentrated on the centrosomes and the spindle microtubules, as seen in Drosophila embryos. During interphase, a high level of Msps staining is seen in the central region of the cell where microtubule density is high (Figure 1A and Supplementary Figure 1). Punctate staining was also seen along individual microtubules that extended out toward the cell cortex. Additionally, Msps staining was seen strongly on the plus ends of microtubules at the edge of the cell (Figure 1A, arrowheads). These stainings were greatly reduced after RNAi of Msps (Supplementary Figure 2), indicating that the stainings represent Msps protein. The association of Msps protein with microtubules in interphase prompted us to study the roles of Msps in the regulation of interphase microtubules.
Figure 1.
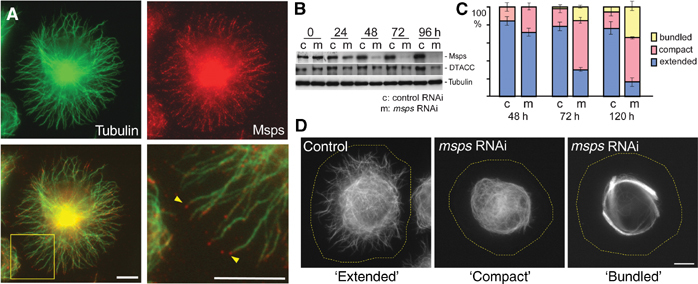
Msps depletion disrupts the organisation of interphase microtubules. (A) Localisation of Msps along interphase microtubules with concentration in plus ends in S2 cells. Drosophila S2 cells were plated out on the con A-coated surface and stained with antibodies against α-tubulin and Msps. The yellow box indicates the area magnified in the right bottom panel. The arrowheads indicate Msps staining, which coincides with microtubule plus ends. (B) Depletion of Msps by RNAi. S2 cells were incubated with dsRNA corresponding to a part of the msps gene (m), or bacteria β-lactamase as control (c). Immunoblots indicated that Msps protein was well depleted by msps RNAi. In addition, the amount of D-TACC, the binding partner of Msps, was decreased to a lesser extent. (C) Interphase microtubule organisation of cells treated with Msps dsRNA or control dsRNA. Most of cells treated with control dsRNA showed normal ‘extended' microtubule organisation in which interphase microtubules extend towards the periphery of the cell from a microtubule-dense region in the centre of the cells. The depletion of Msps induced abnormal microtubule organisation in which microtubules are not extended from the central region (‘compact') or form thick bundles in the central region (‘bundled'). Standard deviations are represented as lines at the top of corresponding bars. (D) Typical microtubule organisation in control and Msps-depleted cells. The yellow dotted lines indicate the outlines of the cell. Bar=10 μm.
Depletion of Msps by RNAi disrupts interphase microtubule organisation
To study the role of Msps protein in interphase, we depleted Msps from S2 cells by RNAi. Incubations with double-stranded RNAs (dsRNAs) corresponding to different nonoverlapping parts of msps gave the same results, while incubation with unrelated control dsRNA (bacteria β-lactamase) showed no effects (Figures 1C and 3D). A reduction in the level of Msps was seen at 24 h and more than 70% of the protein was depleted after 48 h (Figure 1B), while at 120 h greater than 95% was depleted (Supplementary Figure 2). No change in Msps protein level was detected in the cells treated with the control dsRNA (Figure 1B).
Figure 3.
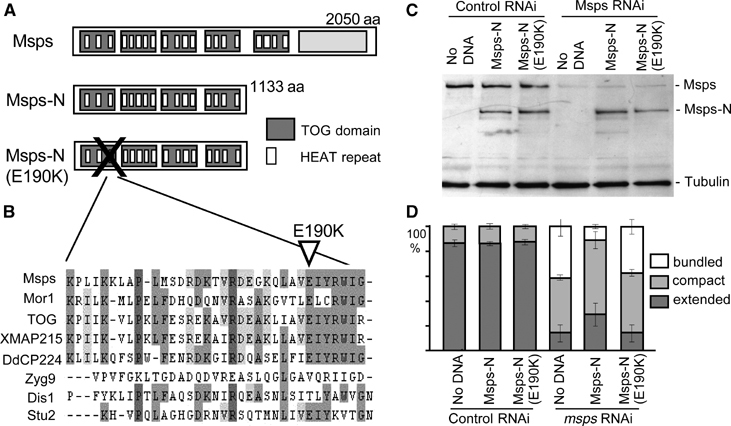
N-terminal region of Msps partially rescues depletion of the endogenous Msps. (A) Domain structures of Msps protein and mutants used in this study. A full-length Msps protein consists of five TOG domains and the C-terminal conserved region. Each TOG domain is made of up to five HEAT repeats. Constructs were made to express the N-terminal 1133 amino acids (Msps-N) and the same region with one mutation (Msps-N[E190K]) that results in a change (glutaminate to lysine) of the conserved 190th amino acid. (B) Sequence comparison of the last HEAT repeats of the first TOG domain among Msps homologues. E190 in Msps is well conserved. (C) Expression of Msps-N and depletion of endogenous full-length Msps. Endogenous full-length Msps was depleted (Msps RNAi) using dsRNA that corresponds to the C-terminal region. dsRNA corresponding to the β-lactamase gene was used as a control. Msps-N and Msps-N[E190K] were expressed under the native msps promoter by transfections of plasmids. Msps proteins were detected by an antibody that recognises the N-terminal half of Msps. An α-tubulin antibody was used as a loading and blotting control. (D) Partial rescue of Msps depletion by the N-terminal region of Msps. Interphase microtubule phenotypes were examined 4 days after the addition of dsRNA. Defects caused by Msps depletion are partially rescued by expression of Msps-N but not Msps-N[E190K].
Msps protein forms a complex with D-TACC protein (Cullen and Ohkura, 2001; Lee et al, 2001), and this interaction is conserved in other organisms (Bellanger and Gönczy, 2003; Gergely et al, 2003; Srayko et al, 2003; Sato et al, 2004). In the case of Caenorhabditis elegans, depletion of the Msps homologue Zyg-9 reduces the amount of D-TACC homologue TAC-1 (Bellanger and Gönczy, 2003). Immunoblotting indicated that the amount of D-TACC protein was indeed reduced in Msps-depleted cells, but at earlier time points, the level of D-TACC depletion was significantly lower than that of Msps depletion (Figure 1B). Nevertheless, as the depletion of D-TACC does not result in the defects seen in Msps RNAi (see below), it alone cannot account for the defects caused by Msps depletion.
Microtubule organisation in Msps-depleted cells was examined by α-tubulin immunostaining. Abnormal spindles were observed in mitotic cells, accompanied by an elevated mitotic index. Abnormalities included broad or split spindle poles and multiple bipolar spindles with some shared poles (Supplementary Figure 3), similar to those seen in cells of the larval central nervous systems in msps mutants (Cullen et al, 1999) and also to the phenotype seen by RNAi in human cultured cells (Gergely et al, 2003; Holmfeldt et al, 2004).
In interphase cells, Msps depletion dramatically altered microtubule organisation. In the control cells, microtubules are typically extended towards the cell periphery from a mass of microtubules in the centre of the cell (Figure 1D). Following Msps depletion, in many cells, the microtubules were concentrated only in the central region of the cell and did not extend out to the periphery (Figure 1D). At 72 h, normal extended microtubules were observed in only 28% of cells. In addition, thick bundles of microtubules formed around the nucleus situated in the cell centre in some cells. At 120 h, extensive microtubule bundling was found in 34% of cells.
In our typical protocol, cells were fixed after culturing them for 2 h on a con A-coated surface (Materials and methods). It was therefore possible that Msps depletion was affecting the speed of microtubule rearrangement and extension towards the cell periphery after cell spreading. To exclude this possibility, S2 cells were continuously cultured on a con A-coated surface from the start of Msps depletion. The same microtubule defects were observed as those seen with 2 h spreading (Supplementary Figure 4), indicating that Msps is important for normal microtubule organisation.
The dramatic affect of microtubule organisation could have secondary effects on cell architecture. Staining with rhodamine-labelled phalloidin indicated that Msps depletion did not affect spreading on the con A-coated surface or actin distribution (Supplementary Figure 5).
Effects of depletion is independent of kinesin-13/KinI proteins or D-TACC
Vertebrate homologues of Msps are known to antagonise the activity of a catastrophe factor, MCAK/XKCM1, which belongs to the kinesin-13/KinI family of kinesin-like proteins (Tournebize et al, 2000). The Drosophila genome has three members of the kinesin-13/KinI family, KLP10A, KLP59C and KLP59D (Rogers et al, 2004). To test if the kinesin-13 proteins were involved in generation of the interphase microtubule phenotype seen upon Msps depletion, RNAi was carried out on the three kinesin-13 proteins in combination with Msps.
Single RNAi of the three kinesin-13 proteins produced no obvious interphase microtubule phenotype (data not shown). Depletion of KLP10A and KLP59C was confirmed by the appearance of mitotic phenotypes as previously reported (Rogers et al, 2004). Consistent with the previous report, RNAi of KLP59D produced no mitotic phenotype. Considering the robustness of RNAi in S2 cells (Goshima and Vale, 2003), successful depletion of KLP59D is also likely.
Individual depletion of each of the three kinesin-13 proteins in combination with Msps did not rescue the interphase microtubule defects caused by Msps depletion (Figure 2A). Interestingly, codepletion of KLP10A with Msps resulted in more severe microtubule defects. The number of cells with extensive microtubule bundling rose from 24% (in cells singly depleted of Msps) to 58% (in cells doubly depleted of Msps and KLP10A; Figure 2A). In contrast, depletion of KLP10A partially rescued mitotic spindle defects caused by Msps depletion (Supplementary Figure 6).
Figure 2.
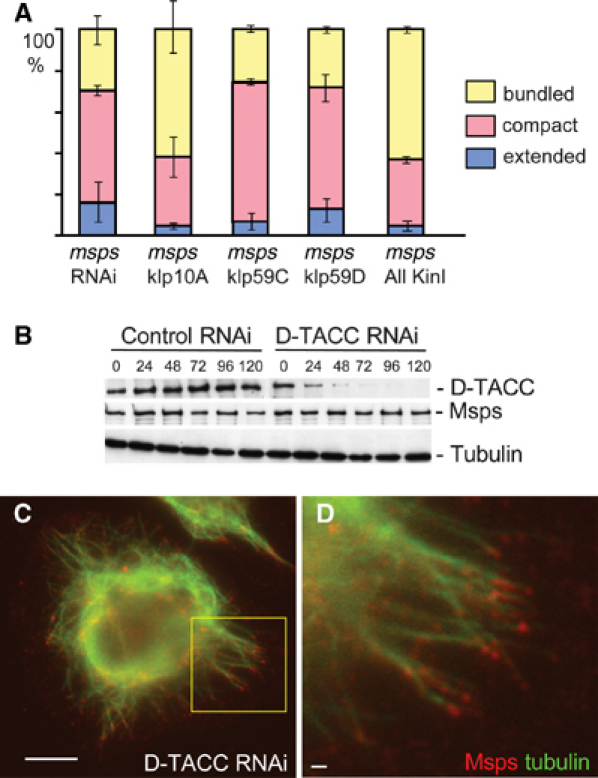
Interphase function of Msps is independent of known effectors or regulators, kinesin-13/KinI and D-TACC. (A) Interphase microtubule organisation of cells that were depleted of the kinesin-13/KinI homologues and Msps (120 h after dsRNA addition). Msps was depleted by RNAi alone (msps RNAi), together with single kinesin-13/KinI homologues (msps klp), and all three kinesin-13 (msps All KinI). KLP10A depletion enhanced the effect of Msps depletion. Simultaneous depletions of kinesin-13/KinI homologues did not rescue interphase microtubule defects of Msps depletion. (B) Depletion of the Msps binding partner D-TACC by RNAi. Immunoblots using Msps and D-TACC antibodies show that D-TACC was effectively depleted, while Msps was not affected. An α-tubulin antibody was used as a loading and blotting control. (C, D) Msps localisation to microtubules in D-TACC-depleted cells. D corresponds to the area marked by the yellow box in panel C. The preferential localisation to microtubule plus ends was observed as in untreated cells. Bar=10 μm.
To exclude the possibility that effects of kinesin-13 depletion were masked by functional redundancy between the kinesin-13 proteins, all three proteins were simultaneously depleted with Msps. The triple depletion did not rescue the interphase microtubule defects resulting from Msps depletion (Figure 2A), indicating that the defect is caused by a mechanism independent of kinesin-13/KinI proteins. Therefore, Msps function in regulating interphase microtubule organisation is not mediated by an antagonising kinesin-13 activity.
Msps protein forms a complex with D-TACC protein and the localisation and function of Msps to spindle poles depend on D-TACC in female meiosis and early embryos (Cullen and Ohkura, 2001; Lee et al, 2001). Because Msps depletion resulted in the reduction of D-TACC in S2 cells (Figure 1B), we examined whether D-TACC may be involved in interphase microtubule organisation. In D-TACC RNAi experiments in S2 cells, we found that D-TACC protein was well depleted by 120 h, but the amount of Msps protein was unaffected in these cells (Figure 2B). Immunostaining indicated that depletion of D-TACC had no significant effects on interphase microtubule organisation (Figure 2C). Additionally, Msps protein was able to localise to interphase microtubules in D-TACC-depleted cells (Figure 2D). These results indicate that Msps protein function in interphase microtubule organisation is independent of D-TACC.
The N-terminal half of the Msps protein partially rescues Msps depletion
To gain a further insight into interphase Msps function, we located an active region within Msps. The N-terminal region of Msps is made of five TOG domains, each consisting of five HEAT repeats (Figure 3A). We expressed the N-terminal 1133 amino acids of Msps (Msps-N; Figure 3A) containing four repeats of the TOG domains in S2 cells to test whether the fragment can rescue the microtubule defects caused by Msps depletion. The equivalent region in XMAP215 has only a weak affinity to microtubules, but has been shown to have both microtubule stabilising and destabilising activities (Popov et al, 2001; Shirasu-Hiza et al, 2003). As a control, we created a mutant that contains an alteration of a highly conserved amino acid (E190K) in the first TOG domain (Figure 3A and B). This mutation is equivalent to a temperature-sensitive mutation in the Arabidopsis homologue (mor1-2), which disrupts cortical microtubules (Whittington et al, 2001).
To examine if Msps-N can carry out interphase function, the endogenous full-length Msps protein was depleted by RNAi and then Msps-N or Msps-N[E190K] was ectopically expressed by transfection of a corresponding plasmid. To avoid dsRNA destroying the RNA encoding Msps-N, dsRNA corresponding to the C-terminal region was used for depletion of endogenous Msps protein. Immunoblotting using an antibody against the N-terminal region of Msps confirmed that full-length Msps was successfully depleted and the Msps-N protein expressed (Figure 3C).
The Msps-N and Msps-N[E190K] proteins were found to localise throughout the cytoplasm with some granular concentrations (Supplementary Figure 7). Consistent with results from the Xenopus homologue (Popov et al, 2001), this fragment does not localise to the centrosomes in mitosis (data not shown). Expression of Msps-N protein in cells treated with control dsRNA did not affect microtubule organisation (Figure 3D). When Msps-N was expressed in Msps-depleted cells, the proportion of cells with abnormal microtubule organisation was reduced. In particular, the proportion of cells with extensive microtubule bundling dropped from 43 to 13% in Msps-depleted cells. The mutation, Msps-N[E190K], abolished the ability of the protein to rescue the defects of Msps depletion (Figure 3D), although it may be partly due to slightly lower protein level.
These results indicate that the Msps-N can, at least partially, rescue the defects caused by Msps depletion. This N-terminal region is likely to contain an activity that is important for Msps regulation of interphase microtubules and the conserved residue in the first TOG domain is crucial for the activity.
Extensive microtubule pausing in Msps-depleted cells
In order to achieve a better understanding of interphase microtubule defects, we examined dynamics of individual microtubules in the Msps-depleted cells. For real-time analysis of microtubule dynamics, we used an S2 cell that stably expresses α-tubulin fused with GFP (Goshima and Vale, 2003). The depletion of Msps by RNAi in this cell line showed the same microtubule organisation defects as those observed in S2 cells (data not shown). Since individual microtubules cannot be resolved in most cells after 72 h, we observed cells incubated with dsRNA for 48 h at which time microtubule organisation is mildly affected (Figure 1C).
We followed the behaviour of individual microtubules both in control cells and Msps partially depleted cells by live-cell fluorescent microscopy. Microtubules in control cells exhibited dynamic instability (Figure 4A and C), alternating between growth phase and shrinkage phase. In addition, microtubule ends spent time in a paused state with little growth or shrinkage. We noticed that microtubule behaviour is different at the cortex than when microtubule ends are internal. Internal microtubules tend to grow fairly continuously with few catastrophes (0.22 events per minute). When microtubules reach the cortex, they tend to pause (time spent in a paused state rises from 29.7 to 48.4%; Figure 4E) and catastrophe frequency more than doubles (0.52 events per minute).
Figure 4.
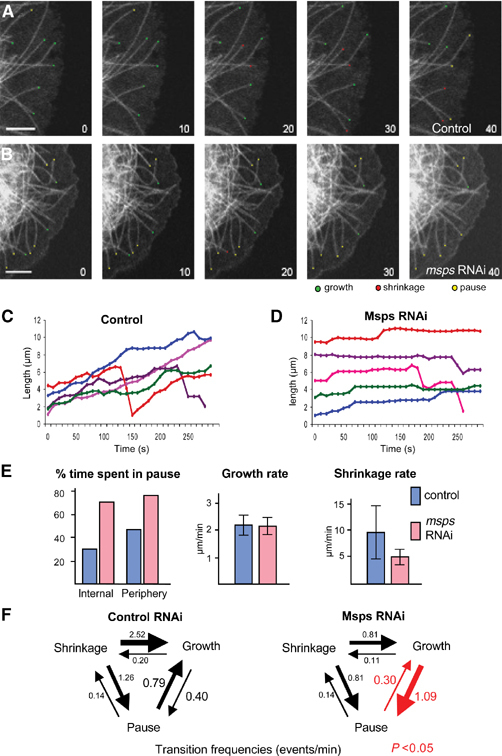
Extensive microtubule pausing is induced by Msps depletion. (A) Time-lapse images of microtubule plus ends in control GFP-tubulin cells. Growing, shrinking and pausing ends were marked with green, red and yellow dots. Numbers at the bottom left represent time in seconds. Microtubules show dynamic instability with occasional pauses. Bar=10 μm. (B) The dynamics of microtubule plus ends in cells partially depleted of Msps. Cells were observed 48 h after the addition of dsRNA. Microtubule plus ends exhibit a prolonged pausing. (C) Life history plots of microtubules in control cells. Microtubule plus ends show persistent growth and occasional shrinkages with some pausing. (D) Life history plots of microtubules in cells partially depleted of Msps. Microtubules spent most of their time pausing with occasional growth and shrinkage. (E) Parameters of microtubule dynamics. Times spent in pause are shown in percentages. In control cells, microtubule plus ends near the cell periphery (within 0.2 μm) spent more time in pause than those inside of the cell (internal). In both cases, Msps partial depletion increased time spent in pause. Average growth and shrinkage rates were shown with standard deviations (lines). (F) Transition frequencies between three microtubule states: growth, shrinkage and pause. Only the transition frequencies marked with red arrows and numbers were significantly different in Msps-depleted cells from those of control cells (P<0.05). The transition from pause to growth significantly decreased and the transition from growth to pause increased. Transition frequencies from shrinkage to growth or pause may be increased, but they were not proved to be statistically significant due to small sample size of time microtubules spent in shrinkage.
Microtubule behaviour was very different in Msps-depleted cells. The most significant change was the increase in the amount of time spent pausing (Figure 4E). For internal microtubules in control cells, on average, 29.7% of their time is spent pausing, while internal microtubules in Msps-depleted cells are paused for 71.4% of their lives. Microtubules that reach the cortex also spend more time in paused state than control cells (76.1 versus 48.4%).
Life history plots of microtubules from Msps-depleted cells show that long periods of pause are interspersed with short grow periods or shrinkage events, but unlike control cells, it was rare to see consistent periods of growth followed by shrinkage (Figure 4D). The rate of growth in Msps-depleted cells (2.1 μm/min) is not significantly different from control cells (2.2 μm/min) while the shrinkage rate is reduced in Msps-depleted cells from 9.5 to 4.8 μm/min (Figure 4E).
For further understanding of the increased paused state, transition frequencies between the three states of growth, pause and shrinkage were determined. For internal microtubule ends, a significant increase in the transition from growth to pause, and a significant decrease in transition from pause to growth were observed (Figure 4F). Overall, this will result in a shift towards the paused state from a growth state.
It is possible that Msps depletion eliminated a population of dynamic microtubules rather than changed dynamics of individual microtubules. We do not think this is the case for the following reasons. Firstly, in control RNAi cells, we did not find microtubules behaving like those in Msps RNAi cells. Secondly, we did not find any significant differences in the amount of tubulin in polymers between control and Msps RNAi cells (Supplementary Figure 8).
In summary, partial depletion experiments indicate that Msps protein suppresses pause state by promoting the transition from pause to growth and reducing the transition from growth to pause.
Microtubule dynamics are greatly reduced in Msps-depleted cells
Individual microtubule dynamics could not be observed in cells well depleted of Msps. Therefore, two alternative strategies—use of a depolymerising drug and fluorescent recovery after photo bleaching (FRAP) analysis—were employed to gain insights into microtubule dynamics in cells with over 90% Msps depletion.
Firstly, we compared the stability of microtubules between control and Msps-depleted cells by monitoring resistance to the microtubule depolymerising drug colchicine. In control cells, nearly all microtubules were depolymerised when exposed to 200 μM colchicine for 16 h (Figure 5A and B). In contrast, more than half of Msps-depleted cells still contained bundles of microtubules after the same treatment (Figure 5A and B). This result indicates that microtubules are stable in Msps-depleted cells in terms of resistance to a microtubule destabilising drug.
Figure 5.
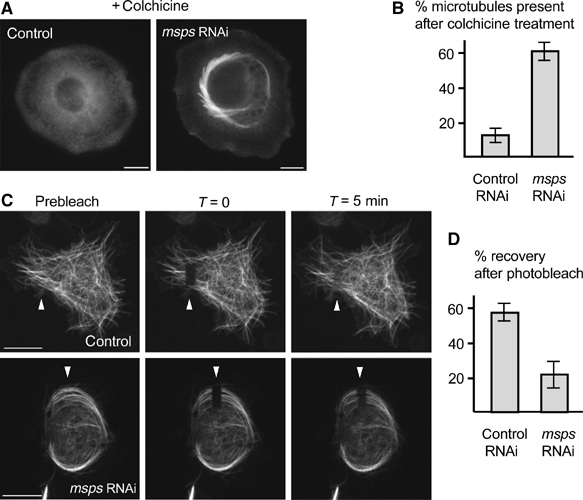
Msps depletion results in stable interphase microtubules. (A) Microtubule organisation after addition of colchicine, a microtubule destabilising drug. Control and Msps-depleted cells (5 days after dsRNA addition) were observed after 16 h incubation with colchicine. Some interphase microtubules remained in Msps-depleted cells, while nearly all microtubules were depolymerised in control cells. (B) Frequencies of cells that have residual microtubules after colchicine treatment. Lines indicate standard deviations from three independent experiments. (C) FRAP of GFP -tubulin. Small areas (arrowheads) of cells expressing GFP-tubulin were bleached and the recovery of fluorescence monitored for 5 min. Cells were treated with dsRNA for 5 days before the photobleach. (D) Percentages of the fluorescent recovery 5 min after photobleaching. Lines indicate standard deviations. Decreased turnover of tubulin dimers into microtubules was observed in Msps-depleted cells, indicating decreased microtubule dynamics. The estimated maximum recovery and t1/2 are 74±5% and 54±6 s in control, and 39±9% and 63±18 s in Msps RNAi.
A second approach for assaying microtubule dynamics was to examine turnover of tubulin dimers in microtubule bundles by FRAP. The GFP-tubulin-expressing line was again used in these experiments. An area of microtubules was bleached in the cell and the recovery followed by time-lapse photography (Figure 5C). The percentage of recovery after 5 min was measured for control and Msps dsRNA-treated cells. After 5 min, control cells had recovered, on average, to 57% of the prebleach level, while after the same time period Msps-depleted cells had recovered on average to 23% of the prebleach level (Figure 5D). Msps microtubule bundles have a lower turnover of tubulin dimmers, indicating an attenuation of microtubule dynamics.
These results from drug and FRAP experiments are consistent with real-time observations of individual microtubule dynamics, which indicate that microtubules are stabilised by depletion of Msps.
Defective organisation of interphase microtubules in haemocytes from msps mutant larvae
Our results demonstrated that Msps protein is a major regulator of interphase microtubule organisation in S2 cultured cells. To confirm the role of Msps in interphase microtubule regulation, we looked for evidence of interphase microtubule defects in msps mutant flies.
We isolated haemocytes from wild-type and msps mutant third-instar larvae, and incubated them on the con A-coated surface. Microtubule organisation of these cells was examined by immunostaining. In wild-type cells, the microtubule organisation was similar to that of S2 cells, with an extended microtubule network reaching the cell periphery (Figure 6A). Haemocytes were obtained from a number of msps mutants. The null alleles produced very few haemocytes, probably due to defects in mitosis and were therefore not analysed (unpublished results). However, two hypomorphic alleles, mspsP and msps146 (Materials and methods), were found to produce enough haemocytes to investigate the microtubule phenotype. Both mutant alleles showed a reduction in the proportion of cells with an extended microtubule array and a rise in the proportion of cells with a compact circle of microtubules in the centre of the cell (Figure 6A). The more severe phenotype was demonstrated by the mspsP mutant, in which 60% of haemocytes had compact microtubule organisation, compared to less than 15% in wild-type flies (Figure 6B). Additionally, 15% of mspsP cells showed bundling of microtubules, while none were seen in wild-type cells. These defects are similar to those in Msps-depleted S2 cells.
Figure 6.
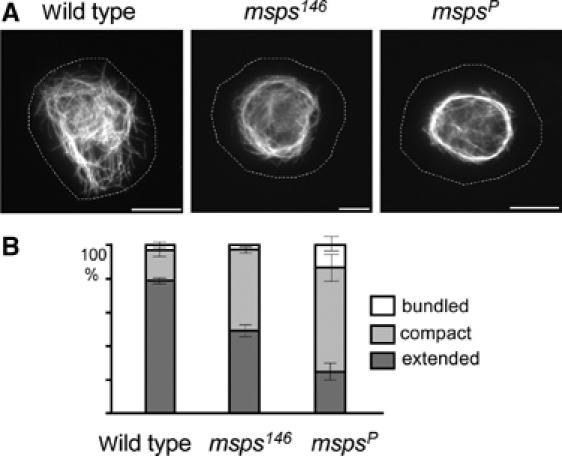
Abnormal interphase microtubule organisation in haemocytes from msps mutants. (A) Interphase microtubule organisation of haemocytes from wild-type larvae and msps mutants (allele 146 and P) after being plated out on a con A-coated surface. Most haemocytes from wild type have ‘extended' microtubule organisation (left), while those from msps mutants contain ‘compact' (middle) or ‘bundled' microtubule organisation as seen in msps-depleted S2 cells. (B) Quantitative analysis of interphase microtubule organisation in msps mutants.
These results from msps mutants indicate that Msps is a major regulator of interphase microtubule organisation in flies as well as in cultured cells.
Discussion
The Dis1/TOG proteins are one of the most conserved families of MAPs among eukaryotes. Studies in various systems revealed that this family of MAPs regulates a variety of microtubule functions in the cell (Ohkura et al, 2001). However, the focus of most studies has been on functions in mitosis, while interphase microtubule regulation has featured less prominently. Crucially, in vivo roles of this family of MAPs in interphase animal cells need to be established. Here, we have reported the first detailed functional study of the role of a member of the Dis1/TOG family in interphase microtubule dynamics in animal cells.
Our study revealed that the depletion of Msps results in a dramatic increase in the amount of time spent in a paused state. Strikingly, even the partial depletion of Msps has stronger effects than nearly complete depletion of EB1, the only protein previously shown to have antipause activity (Rogers et al, 2002). Previous in vitro experiments revealed activities of the Msps homologues to increase growth and shrinkage rates, and to suppress catastrophe frequency (Gard and Kirschner, 1987; Vasquez et al, 1994; Tournebize et al, 2000; Kinoshita et al, 2001). However, antipause activity has not been reported in other Msps homologues in vivo or in vitro except the budding yeast homologue stu2, the deletion of which modestly increases pausing of cytoplasmic microtubules (Kosco et al, 2001). It would be interesting to see whether microtubule dynamics are altered by the depletion of TOG in mammalian cells in the light of our finding.
Dynamic instability is an intrinsic property of microtubules. Microtubules assembled in vitro from purified tubulin either grow or shrink, with infrequent alternation between the two states (Mitchison and Kirschner, 1984). In cells, however, microtubule dynamics are strongly influenced by additional proteins, and microtubule plus ends often pause without significant growth or shrinkage. This pausing is more frequent at the cortex (this study and others; Komarova et al, 2002), indicating that it is regulated by cellular factors. This regulation is potentially critical for linking microtubule ends to the cell cortex. Despite being a common feature of microtubules in cells, relatively little attention has been paid to the pause state, and consequently only limited information is available on the nature and regulation of this state.
The most enigmatic aspect of this family of MAPs is to have apparently opposing activities, both stabilising and destabilising microtubules (Tournebize et al, 2000; van Breugel et al, 2003; Shirasu-Hiza et al, 2003). In vitro functional analysis of XMAP215 indicates that the protein has the intrinsic ability to promote both the polymerisation and depolymerisation of microtubules (Tournebize et al, 2000; Shirasu-Hiza et al, 2003). Shirasu-Hiza et al (2003) proposed that the two opposing activities of XMAP215 could be reconciled if it functions as an antipause factor. This model is based on the hypothesis (Tran et al, 1997) that the pause state may be the obligate intermediate between growth and shrinkage. It is also proposed (Arnal et al, 2000) that a pause corresponds to blunt-ended ultrastructures distinct from those of growth (sheet-like) and shrinkage (curled). Destabilising the pause state will promote the transitions from pause to both growth and shrinkage, therefore it can act as a microtubule stabiliser as well as a destabiliser.
Our results in Drosophila cells provide the first direct experimental evidence to support the hypothesis proposed by Shirasu-Hiza et al (2003) that Msps acts as an antipause factor. Consistent with the hypothesis, our further analysis shows that Msps promotes transitions from pause to growth and inhibits transitions from growth to pause. On the other hand, we did not see that Msps significantly promoted the transition from pause to shrinkage. However, this may be due to partial depletion of Msps, which allows direct observation of microtubule dynamics. In cells that are well depleted of Msps, microtubules are more resistant to the depolymerising drug colchicine, suggesting that catastrophe or depolymerisation is inhibited.
We showed that Msps associates with microtubule plus ends in the cell. Therefore, it could directly destabilise the paused state by altering polymer conformation or it could act indirectly by preventing a pause factor from accessing the polymer. In Xenopus egg extracts both from mitosis and interphase, XMAP215 is shown to stabilise microtubules mainly by antagonising the microtubule depolymerising activity of the kinesin-13/KinI family kinesin, XKCM1 (Tournebize et al, 2000). In contrast, our study shows that Msps regulation of interphase microtubules in S2 cells is unlikely to be through the kinesin-13 proteins. We also found that Msps activity and localisation to interphase microtubules are independent of its binding partner D-TACC, suggesting this interphase function is independent of known regulators or effectors. As our knowledge on microtubule pausing is very limited, we cannot speculate on the precise mechanism of the antipause activity. However, our finding represents a significant advance towards the molecular understanding of this third state of microtubule dynamics.
Msps depletion dramatically altered interphase microtubule organisation in Drosophila S2 cells. Microtubules are unable to extend to the periphery of the cell and occasionally form extensive bundles. The importance of Msps for microtubule organisation in developing flies was confirmed by the presence of similar defects in haemocytes from msps mutant larvae. In addition, a very recent report (Moon and Hazelrigg, 2004) has described defects in microtubule organisation in msps mutant oocytes. Therefore, Msps is likely to be a general regulator of interphase microtubules in Drosophila. Mutations in msps homologues have a dramatic impact on the organisation of interphase microtubules in yeasts, Dictyostelium amoebae and plants (Garcia et al, 2001; Whittington et al, 2001; Graf et al, 2003). In contrast, the depletion of the mammalian homologue, TOG, by RNAi does not alter interphase microtubule organisation (Gergely et al, 2003; Holmfeldt et al, 2004), although none of these studies have examined microtubule dynamics.
Extensive pausing can explain the failure of interphase microtubules to extend out to the cell cortex seen in Msps-depleted cells. It is not clear whether the microtubule bundling phenotype is a secondary consequence of extensive pausing or whether Msps somehow acts directly as an antibundling factor. It is also unclear whether microtubule stabilisation caused bundling or vice versa. Interestingly, although microtubule bundling is commonly observed during overexpression of MAPs (Bu and Su, 2001; Haren and Merdes, 2002; Raemaekers et al, 2003), Msps is the only protein that, as far as we are aware, induces microtubule bundling when it is depleted.
In conclusion, our functional study of Msps demonstrated an essential role in the organisation and dynamics of interphase microtubules in vivo, and uncovered the antipause activity, which provides a crucial insight into this family of MAPs.
Materials and methods
Cell culture and RNAi
Drosophila S2 cells were cultured and RNAi was performed as described (Clemens et al, 2000; Rogers et al, 2002). dsRNAs were produced by in vitro transcription using T7 polymerase (Ambion) and templates made as follows. About 0.6 kb within an exon of the target gene was first amplified from genomic DNA or cDNA using gene-specific primers containing half of the T7 promoter sequence, and further amplified using primers with a full T7 sequence. dsRNAs corresponding to the bacterial β-lactamase gene were used for control RNAi throughout.
For expression of the N-terminal 1133 amino acids (Msps-N), the 6.2 kb SpeI–KpnI genomic DNA containing the promoter and 5′ half of msps gene were cloned into pBluescript (Stratagene). A point mutation, E190K, was made using the site-directed mutagenesis kit (Stratagene). These constructs were transfected into S2 cells that had been treated with control or Msps dsRNA for 2 days. Cells were transfected using Effectene Transfection Reagent (Qiagen). After transfection, 15 μg of dsRNA was added to the cells to ensure depletion of the protein.
Molecular and immunological techniques
Standard molecular techniques were followed throughout (Harlow and Lane, 1988; Sambrook et al, 1989). For immunoblotting, 5 × 104 S2 cells were loaded per lane. An α-tubulin antibody (DM1A; Sigma) was used throughout this study to examine tubulin distribution and as a loading control. In addition to an antibody against the C-terminal of Msps (264; Cullen et al, 1999), rabbit antibodies were made against the following bacterially produced proteins: middle region (746–1225 amino acids) of Msps fused with GST (268), MBP-D-TACC containing the N-terminal 2–433 amino acids or the C-terminal 250 amino acids (Gergely et al, 2000; gifts from J Raff). Antibodies were affinity purified as described (Smith and Fisher, 1984). Peroxidase-conjugated secondary antibodies (Jackson Lab) and the ECL kit (Amersham) were used for detection.
Immunofluorescence microscopy
For immunostaining, S2 cells were cultured on a con A-coated coverslip for 2 h and fixed with 90% methanol, 3% formaldehyde and 5 mM sodium carbonate at −80°C as described (Rogers et al, 2002). Haemocytes were collected from third-instar larvae by tearing the cuticle in 100 μl Schneiders media (Tirouvanziam et al, 2004), and were then left to attach onto a con A-coated coverslip for 2 h before fixation as above. Secondary antibodies conjugated with Alexa488 or Cy3 (Jackson Lab or Molecular Probes) and DAPI (Sigma) were used for visualisation. For quantification, a total of at least 300 cells were routinely counted in three separate experiments. Images were captured using an Axioplan2 microscope (Zeiss) attached with a CCD camera (Hamamatsu) controlled by OpenLab (Improvision) or an Axiovert microscope attached with LSM510Meta confocal scan head (Zeiss), and processed using Photoshop (Adobe).
Microtubule dynamics in live S2 cells
For observation of individual microtubule dynamics, cells expressing GFP-tubulin (a gift from R Vale; Goshima and Vale, 2003) were treated with 15 μg of control or msps dsRNA for 2 days. The cells were then grown on con A-coated coverslips for 1–2 h before being observed on the confocal microscope described above. Images were acquired every 5 s for 5 min. Microtubule length was measured from an arbitrary chosen point on the microtubule over time using ImageJ (http://rsb.info.nih.gov/ij). Only microtubules that could be followed for at least 3 min were included in the analysis. Pauses were defined in this study as events without significant change in the microtubule length (<0.5 μm) for 30 s or more. The transition frequencies between growth, pause and shrinkage were calculated as the total number of transitions (from A to B) divided by the total time spent in the pretransition state (A). Microtubule ends at the cell periphery (within 0.2 μm from the cell edge) and in the internal region were analysed separately.
FRAP
S2 cells expressing GFP-tubulin treated with either control or Msps dsRNA for 5–6 days were plated on con A-coated coverslips. The FRAP analysis was carried out using the confocal microscope described above. An area 2.5 μm × 6 μm was bleached on 60% laser power for 50 iterations. Images were recorded prebleach, immediately after bleaching and subsequently images were captured every 20 s for at least 5 min. Signals in a nonbleached area were also measured to compensate a reduction of fluorescence caused by image captures. For a comparison, per cent fluorescence recovery was calculated as the amount of recovery (the difference between images immediately after photobleach and 5 min later) relative to photobleach (the difference before and immediately after photobleach).
Fly stocks
Standard techniques for fly manipulation were followed (Ashburner, 1989). All stocks were grown at 25°C in standard cornmeal media. w1118 flies were used as wild type. msps alleles were kept over TM6C (Lindsley and Zimm, 1992). mspsP is a P-element insertion allele, which greatly reduced Msps protein level (Cullen et al, 1999), and msps146 contains a mutation in a splicing junction (unpublished). mspsP dies as early pupae and msps146 dies as pharate, while null alleles die as early third-instar larvae. Their lethality is rescued by the wild-type msps transgene.
Supplementary Material
Supplementary Figure 1
Supplementary Figure 2
Supplementary Figure 3
Supplementary Figure 4
Supplementary Figure 5
Supplementary Figure 6
Supplementary Figure 7
Supplementary Figure 8
Acknowledgments
We are grateful to S Rogers and R Vale (UCSF) for making GFP-tubulin cell line available, to J Raff for D-TACC constructs and antibody and to Marissa Brown and Duncan Sproul for helping with antibody production and EMS screening. The work is supported by The Wellcome Trust and BBSRC.
References
- Arnal I, Karsenti E, Hyman AA (2000) Structural transitions at microtubule ends correlate with their dynamic properties in Xenopus egg extracts. J Cell Biol 149: 767–774 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashburner M (1989) Drosophila. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press [Google Scholar]
- Bellanger JM, Gönczy P (2003) TAC-1 and Zyg-9 form a complex that promotes microtubule assembly in C. elegans embryos. Curr Biol 13: 1488–1498 [DOI] [PubMed] [Google Scholar]
- Bu W, Su LK (2001) Regulation of microtubule assembly by human EB1 family proteins. Oncogene 20: 3185–3192 [DOI] [PubMed] [Google Scholar]
- Cassimeris L, Pryer NK, Salmon ED (1988) Real-time observations of microtubule dynamic instability in living cells. J Cell Biol 107: 2223–2231 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charrasse S, Schroeder M, Gauthier-Rouviere C, Ango F, Cassimeris L, Gard DL, Larroque C (1998) The TOGp protein is a new human microtubule-associated protein homologous to the Xenopus XMAP215. J Cell Sci 111: 1371–1383 [DOI] [PubMed] [Google Scholar]
- Clemens JC, Worby CA, Simonson-Leff N, Muda M, Maehama T, Hemmings BA, Dixon JE (2000) Use of double-stranded RNA interference in Drosophila cell lines to dissect signal transduction pathways. Proc Natl Acad Sci USA 97: 6499–6503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cullen CF, Deak P, Glover DM, Ohkura H (1999) mini spindles: a gene encoding a conserved microtubule-associated protein required for the integrity of the mitotic spindle in Drosophila. J Cell Biol 146: 1005–1018 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cullen CF, Ohkura H (2001) Msps protein is localized to acentrosomal poles to ensure bipolarity of Drosophila meiotic spindles. Nat Cell Biol 3: 637–642 [DOI] [PubMed] [Google Scholar]
- Garcia MA, Vardy L, Koonrugsa N, Toda T (2001) Fission yeast ch-TOG/XMAP215 homologue Alp14 connects mitotic spindles with the kinetochore and is a component of the Mad2-dependent spindle checkpoint. EMBO J 20: 3389–3401 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gard DL, Kirschner MW (1987) A microtubule-associated protein from Xenopus eggs that specifically promotes assembly at the plus-end. J Cell Biol 105: 2203–2215 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gergely F, Kidd D, Jeffers K, Wakefield JG, Raff JW (2000) D-TACC: a novel centrosomal protein required for normal spindle function in the early Drosophila embryo. EMBO J 19: 241–252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gergely F, Draviam VM, Raff JW (2003) The ch-TOG/XMAP215 protein is essential for spindle pole organization in human somatic cells. Genes Dev 17: 336–341 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goshima G, Vale RD (2003) The roles of microtubule-based motor proteins in mitosis: comprehensive RNAi analysis in the Drosophila S2 cell line. J Cell Biol 162: 1003–1016 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graf R, Daunderer C, Schliwa M (2000) Dictyostelium DdCP224 is a microtubule-associated protein and a permanent centrosomal resident involved in centrosome duplication. J Cell Sci 113: 1747–1758 [DOI] [PubMed] [Google Scholar]
- Graf R, Euteneuer U, Ho TH, Rehberg M (2003) Regulated expression of the centrosomal protein DdCP224 affects microtubule dynamics and reveals mechanisms for the control of supernumerary centrosome number. Mol Biol Cell 14: 4067–4074 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haren L, Merdes A (2002) Direct binding of NuMA to tubulin is mediated by a novel sequence motif in the tail domain that bundles and stabilizes microtubules. J Cell Sci 115: 1815–1824 [DOI] [PubMed] [Google Scholar]
- Harlow E, Lane D (1988) Antibodies: A Laboratory Manuel. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press [Google Scholar]
- Holmfeldt P, Stenmark S, Gullberg M (2004) Differential functional interplay of TOGp/XMAP215 and the KinI kinesin MCAK during interphase and mitosis. EMBO J 23: 627–637 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kinoshita K, Arnal I, Desai A, Drechsel DN, Hyman AA (2001) Reconstitution of physiological microtubule dynamics using purified components. Science 294: 1340–1343 [DOI] [PubMed] [Google Scholar]
- Kinoshita K, Habermann B, Hyman AA (2002) XMAP215: a key component of the dynamic microtubule cytoskeleton. Trends Cell Biol 12: 267–273 [DOI] [PubMed] [Google Scholar]
- Komarova YA, Vorobjev IA, Borisy GG (2002) Life cycle of MTs: persistent growth in the cell interior, asymmetric transition frequencies and effects of the cell boundary. J Cell Sci 115: 3527–3539 [DOI] [PubMed] [Google Scholar]
- Kosco KA, Pearson CG, Maddox PS, Wang PJ, Adams IR, Salmon ED, Bloom K, Huffaker TC (2001) Control of microtubule dynamics by Stu2p is essential for spindle orientation and metaphase chromosome alignment in yeast. Mol Biol Cell 12: 2870–2880 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee MJ, Gergely F, Jeffers K, Peak-Chew SY, Raff JW (2001) Msps/XMAP215 interacts with the centrosomal protein D-TACC to regulate microtubule behavior. Nat Cell Biol 3: 643–649 [DOI] [PubMed] [Google Scholar]
- Lindsley DL, Zimm GG (1992) The Genome of Drosophila Melanogaster. New York: Academic Press [Google Scholar]
- Matthews LR, Carter P, Thierry-Mieg D, Kemphues K (1998) ZYG-9, a Caenorhabditis elegans protein required for microtubule organization and function, is a component of meiotic and mitotic spindle poles. J Cell Biol 141: 1159–1168 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitchison T, Kirschner M (1984) Dynamic instability of microtubule growth. Nature 312: 237–242 [DOI] [PubMed] [Google Scholar]
- Moon W, Hazelrigg T (2004) The Drosophila microtubule-associated protein Mini spindles is required for cytoplasmic microtubules in oogenesis. Curr Biol 14: 1957–1961 [DOI] [PubMed] [Google Scholar]
- Nakaseko Y, Goshima G, Morishita J, Yanagida M (2001) M phase-specific kinetochore proteins in fission yeast: microtubule-associating Dis1 and Mtc1 display rapid separation and segregation during anaphase. Curr Biol 11: 537–549 [DOI] [PubMed] [Google Scholar]
- Ohkura H, Garcia MA, Toda T (2001) Dis1/TOG universal microtubule adaptors – one MAP for all? J Cell Sci 114: 3805–3812 [DOI] [PubMed] [Google Scholar]
- Popov AV, Pozniakovsky A, Arnal I, Antony C, Ashford AJ, Kinoshita K, Tournebize R, Hyman AA, Karsenti E (2001) XMAP215 regulates microtubule dynamics through two distinct domains. EMBO J 20: 397–410 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Radcliffe P, Hirata D, Childs D, Vardy L, Toda T (1998) Identification of novel temperature-sensitive lethal alleles in essential β-tubulin and nonessential α2-tubulin genes as fission yeast polarity mutants. Mol Biol Cell 9: 1757–1771 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raemaekers T, Ribbeck K, Beaudouin J, Annaert W, Van Camp M, Stockmans I, Smets N, Bouillon R, Ellenberg J, Carmeliet G (2003) NuSAP, a novel microtubule-associated protein involved in mitotic spindle organization. J Cell Biol 162: 1017–1029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers GC, Rogers SL, Schwimmer TA, Ems-McClung SC, Walczak CE, Vale RD, Scholey JM, Sharp DJ (2004) Two mitotic kinesins cooperate to drive sister chromatid separation during anaphase. Nature 427: 364–370 [DOI] [PubMed] [Google Scholar]
- Rogers SL, Rogers GC, Sharp DJ, Vale RD (2002) Drosophila EB1 is important for proper assembly, dynamics, and positioning of the mitotic spindle. J Cell Biol 158: 873–884 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rusan NM, Fagerstrom CJ, Yvon AM, Wadsworth P (2001) Cell cycle-dependent changes in microtubule dynamics in living cells expressing green fluorescent protein-α tubulin. Mol Biol Cell 12: 971–980 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sambrook J, Fritsch EF, Maniatis T (1989) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press [Google Scholar]
- Sato M, Vardy L, Angel GM, Koonrugsa N, Toda T (2004) Interdependency of fission yeast Alp14/TOG and coiled coil protein Alp7 in microtubule localization and bipolar spindle formation. Mol Biol Cell 15: 1609–1622 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Severin F, Habermann B, Huffaker T, Hyman T (2001) Stu2 promotes mitotic spindle elongation in anaphase. J Cell Biol 153: 435–442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shelden E, Wadsworth P (1993) Observation and quantification of individual microtubule behavior in vivo: microtubule dynamics are cell-type specific. J Cell Biol 120: 935–945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shirasu-Hiza M, Coughlin P, Mitchison T (2003) Identification of XMAP215 as a microtubule-destabilizing factor in Xenopus egg extract by biochemical purification. J Cell Biol 161:349–358 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith DE, Fisher PA (1984) Identification, developmental regulation, and response to heat shock of two antigenically related forms of a major nuclear envelope protein in Drosophila embryos: application of an improved method for affinity purification of antibodies using polypeptides immobilized on nitrocellulose blots. J Cell Biol 99: 20–28 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Srayko M, Quintin S, Schwager A, Hyman AA (2003) Caenorhabditis elegans TAC-1 and Zyg-9 form a complex that is essential for long astral and spindle microtubules. Curr Biol 13: 1506–1511 [DOI] [PubMed] [Google Scholar]
- Tirnauer JS, O'Toole E, Berrueta L, Bierer BE, Pellman D (1999) Yeast Bim1p promotes the G1-specific dynamics of microtubules. J Cell Biol 145: 993–1007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tirouvanziam R, Davidson CJ, Lipsick JS, Herzenberg LA (2004) Fluorescence-activated cell sorting (FACS) of Drosophila hemocytes reveals important functional similarities to mammalian leukocytes. Proc Natl Acad Sci USA 101: 2912–2917 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tournebize R, Popov A, Kinoshita K, Ashford AJ, Rybina S, Pozniakovsky A, Mayer TU, Walczak CE, Karsenti E, Hyman AA (2000) Control of microtubule dynamics by the antagonistic activities of XMAP215 and XKCM1 in Xenopus egg extracts. Nat Cell Biol 2: 13–19 [DOI] [PubMed] [Google Scholar]
- Tran PT, Walker RA, Salmon ED (1997) A metastable intermediate state of microtubule dynamic instability that differs significantly between plus and minus ends. J Cell Biol 138: 105–117 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Usui T, Maekawa H, Pereira G, Schiebel E (2003) The XMAP215 homologue Stu2 at yeast spindle pole bodies regulates microtubule dynamics and anchorage. EMBO J 22: 4779–4793 [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Breugel M, Drechsel D, Hyman A (2003) Stu2p, the budding yeast member of the conserved Dis1/XMAP215 family of microtubule-associated proteins is a plus end-binding microtubule destabilizer. J Cell Biol 161: 359–369 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vasquez RJ, Gard DL, Cassimeris L (1994) XMAP from Xenopus eggs promotes rapid plus end assembly of microtubules and rapid microtubule polymer turnover. J Cell Biol 127: 985–993 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang PJ, Huffaker TC (1997) Stu2p: a microtubule-binding protein that is an essential component of the yeast spindle pole body. J Cell Biol 139: 1271–1280 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whittington AT, Vugrek O, Wei KJ, Hasenbein NG, Sugimoto K, Rashbrooke MC, Wasteneys GO (2001) MOR1 is essential for organizing cortical microtubules in plants. Nature 411: 610–613 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Figure 1
Supplementary Figure 2
Supplementary Figure 3
Supplementary Figure 4
Supplementary Figure 5
Supplementary Figure 6
Supplementary Figure 7
Supplementary Figure 8