

Abstract
Plants are constantly exposed to environmental changes and need to integrate multiple external stress cues. Calcium-dependent protein kinases (CDPKs) are implicated as major primary Ca2+ sensors in plants. CDPK activation, like activation of mitogen-activated protein kinases (MAPKs), is triggered by biotic and abiotic stresses, although distinct stimulus-specific stress responses are induced. To investigate whether CDPKs are part of an underlying mechanism to guarantee response specificity, we identified CDPK-controlled signaling pathways. A truncated form of Nicotiana tabacum CDPK2 lacking its regulatory autoinhibitor and calcium-binding domains was ectopically expressed in Nicotiana benthamiana. Infiltrated leaves responded to an abiotic stress stimulus with the activation of biotic stress reactions. These responses included synthesis of reactive oxygen species, defense gene induction, and SGT1-dependent cell death. Furthermore, N-terminal CDPK2 signaling triggered enhanced levels of the phytohormones jasmonic acid, 12-oxo-phytodienoic acid, and ethylene but not salicylic acid. These responses, commonly only observed after challenge with a strong biotic stimulus, were prevented when the CDPK's intrinsic autoinhibitory peptide was coexpressed. Remarkably, elevated CDPK signaling compromised stress-induced MAPK activation, and this inhibition required ethylene synthesis and perception. These data indicate that CDPK and MAPK pathways do not function independently and that a concerted activation of both pathways controls response specificity to biotic and abiotic stress.
Plants respond with a rapid activation of intracellular signaling to unfavorable changes in their environment caused by biotic or abiotic factors, such as pathogen attack, wounding, or cold. As in animals, protein kinase cascades are involved in the induction of plant immunity and for more general stress responses (1, 2). Interactions between signal transduction pathways have, however, often been ignored. In plants, two classes of stress-activated protein kinases, mitogen-activated protein kinases (MAPKs) and calcium-dependent protein kinases (CDPKs), have so far been reported to integrate multiple environmental stresses and undergo rapid biochemical activation upon exposure to biotic and abiotic stimulation. In particular, two MAPKs, tobacco salicylic acid (SA)-induced protein kinase (SIPK) and wound-induced protein kinase (WIPK), and their respective orthologs in other plant species were proposed to function as central convergence points in stress signaling (3). In gain-of-function experiments, the expression of constitutively active MAPK kinases NtMEK2DD (tobacco) or LeMKK2DD (tomato) resulted in pathogen-independent activation of respective MAPKs and triggered plant defense responses, including synthesis of reactive oxygen species, phytohormone production, or induction of cell death (4-6).
In contrast, loss-of-function studies based on silencing of SIPK, WIPK, or their orthologs only partially compromised respective defense responses, causing an attenuation of N-mediated tobacco mosaic virus resistance or reduced resistance to a bacterial pathogen (7, 8). Likewise, suppression of MAPK signaling in Arabidopsis yielded only a partial reduction of flagellin-triggered defense responses or resistance to fungal pathogen Peronospora parasitica (9-11). These data suggest that an additional signaling branch is required to mount a full defense response. Here, we report that a stress-induced CDPK contributes to early signal transduction processes and is connected to MAPK activation and phytohormone signaling.
We previously studied early gene-for-gene-dependent defense responses in tobacco plants expressing the disease resistance gene Cf-9 from tomato. Upon elicitation with the corresponding Avr9 peptide derived from the tomato pathogen Cladosporium fulvum, intracellular responses, including changes in ion fluxes, production of reactive oxygen species, activation of MAPKs SIPK and WIPK, and changes in gene transcription were observed (12). In addition, a CDPK from tobacco, Nicotiana tabacum CDPK2 (NtCDPK2), became rapidly activated biochemically and transcriptionally after Avr9/Cf-9 interaction (described synonymously as biotic stress) and abiotic hypoosmotic stress. Thereby, the biotic stress triggered a stronger and prolonged activation (13). Suppression of the NtCDPK2 and its closely related homologs by using virus-induced gene silencing in Nicotiana benthamiana resulted in plants that were attenuated in the induction of the Avr9/Cf-9-dependent hypersensitive cell death response (13). These data imply a major role for NtCDPK2 in plant defense signaling but does not exclude involvement in other signal transduction processes.
CDPKs are serine/threonine protein kinases that have a conserved modular structure of four domains (VKJC): an N-terminal variable domain (V), a protein kinase domain (K), an autoinhibitory junction domain (J), and a C-terminal calmodulin-like domain (C) containing EF-hand Ca2+-binding motifs (Fig. 1A) (14-16). Here, we report that plants expressing a truncated VK but not full-length NtCDPK2 variant respond to a mild abiotic stress stimulus with the induction of biotic defense reactions, including cell death. Our data not only identify NtCDPK2 as a point of convergence in abiotic and biotic signal responses but also provide first evidence for ethylene as a link mediating cross-talk between CDPK and MAPK signaling pathways.
Fig. 1.

A truncated variant of NtCDPK2 activates plant defense responses. (A) Schematic structure of full-length NtCDPK2 and NtCDPK3 with V, K, J, and C protein domains (VKJC) and C-terminal triple-myc or HA tag. Also shown are the schematic structures of a truncated VK variant lacking the J and C domains and the translational fusions of the NtCDPK2 J domain to a tandem GFP protein molecule (J-GG) and the double GFP fusion protein (GG) alone. Black boxes indicate a EF-hand calcium-binding motif; *, a putative N-terminal myristoylation and palmitoylation site. (B) Transient expression of full-length (VKJC), truncated (VK), and truncated kinase-inactive [VK(D/A)] NtCDPK2 (2) and NtCDPK3 (3) variants or coexpression of VK (2) and J domain GFP fusions in N. benthamiana leaves. Three days after infiltration of Agrobacterium strains, an abiotic stress stimulus was applied locally by injecting water into one leaf panel (red arrow). Photos were taken after 24 h. (C) Protein expression determined by Western blot analysis of solubilized membrane extracts (Left) or total protein extracts (Right). The molecular mass difference between VK (2) and VK (3) is due to the different protein tags. (D) Synthesis of hydrogen peroxide in leaves expressing NtCDPK2 variants and/or the J domain GFP fusions after infiltration of a 3,3′-diaminobenzidine solution and forceps wounding.
Materials and Methods
Agrobacterium-Mediated Transient Expression in Plant Leaves. Agrobacterium-mediated transient expression experiments were performed in 4- to 6-week-old N. benthamiana plants as described in ref. 13. Three days after infiltration of strains, leaves were subjected to a secondary stress stimulus by injecting water (hypoosmotic shock; abiotic stress) or by elicitation of Cf-9-expressing leaves with intercellular fluid with (Avr9+) or without (Avr9-) the Avr9 peptide (Avr9/Cf-9 gene-for-gene interaction; biotic stress). Leaf discs or tissue was harvested over a time course, immediately frozen in liquid nitrogen, and stored at -80°C. In experiments interfering with ethylene perception or synthesis, a 40 μM silver thiosulphate solution [freshly prepared before each application by mixing silver nitrate and sodium thiosulphate (17)] or 40 μM aminoethoxyvinylglycine (AVG) solution was infiltrated 1 day after infection with Agrobacterium tumefaciens and again together with intercellular fluid (Avr9+).
Tagged full-length (VKJC) and truncated (VK) variants of NtCDPK2-myc and NtCDPK3-hemagglutinin (-HA) were generated by PCR and cloned in pBIN19 (myc tag) and pBIN61 (HA tag) vector derivatives (13, 18, 19). Amino acid substitutions D241A (NtCDPK2) and D237A (NtCDPK3) were introduced by PCR-based, site-specific mutagenesis. The tandem GFP fusion genes are based on 35S-GFP (20) to which the PCR-generated J-moiety of NtCDPK2 was fused, and the resulting genes were cloned in pBIN19. A detailed description of plasmids and of the construction of GFP fusions can be found in Supporting Materials and Methods, which is published as supporting information on the PNAS web site. All constructs were sequenced by using ThermoSequenase (Amersham Biosciences) on an automated sequencer (Li-Cor, Lincoln, NE). In VK, a truncated variant of NtCDPK2, Lys-59 was replaced by Glu, representing the amino acid of NtCDPK3 at this position. Experiments reported in this manuscripts concerning VK were repeated also with strain VK59K representing the NtCDPK2 wild-type sequence (Fig. 5, which is published as supporting information on the PNAS web site). Therefore, we could rule out that this amino acid substitution was responsible for the observed responses. Binary vector derivatives were electroporated into A. tumefaciens GV3101 (Table 1, which is published as supporting information on the PNAS web site).
Immunoblotting and Protein Kinase Assays. The preparation of protein extracts from leaf discs and immunoblotting was performed as described in refs. 13 and 21. The following antibodies were used: monoclonal anti-c-myc (9E10, Santa Cruz Biotechnology), polyclonal anti-c-myc (A14, Santa Cruz Biotechnology), monoclonal anti-HA (3F10, Roche), and monoclonal anti-GFP (B34, Covance Research Products, Denver). MAPK activity was determined by in-gel kinase assays with myelin basic protein (Sigma) as substrate, as described in ref. 21. Equal protein expression of NtCDPK2 variants and of SIPK and WIPK was confirmed by Western analysis and SIPK and WIPK activity, respectively.
Northern Blot Analysis and Detection of Hydrogen Peroxide. Total RNA from two 1-cm diameter leaf discs was isolated by using the Tri-Reagent method according to the procedure described by the manufacturer (Sigma). Ten micrograms of total RNA was subjected to 1.1% formaldehyde-containing-agarose gel electrophoresis and transferred to nylon membranes (Hybond-N+, Amersham Biosciences) (22). RNA was crosslinked by baking the membrane for 1 h at 80°C. Equal loading of RNA was confirmed by staining of the membrane with methylene blue before hybridization. Hybridizations were performed at 65°C in high-SDS buffer (23). Hybridization probes were radiolabeled with [α-32P]dCTP by using an oligolabeling kit (Amersham Biosciences). As probes, cDNA fragments were amplified from N. benthamiana cDNA by PCR with corresponding primers (Table 2, which is published as supporting information on the PNAS web site). Hydrogen peroxide accumulation was detected by endogenous peroxidase-dependent in situ histochemical staining using 3,3′-diaminobenzidine as described in ref. 24.
Determination of 1-Aminocyclopropane-1-Carboxylic Acid (ACC), Jasmonic Acid (JA), 12-Oxo-Phytodienoic Acid (OPDA), SA, and Ethylene. Leaf discs from leaves of three independent plants (70 mg of fresh weight in total) per sample were harvested before and after the abiotic stress stimulus, pooled, and stored at -80°C. For determining ACC levels, the frozen leaf tissues were placed in 1 ml of 50% (vol/vol) methanol and incubated under shaking for 30 min at 55°C. The ACC content in the methanol extract was determined by chemical oxidation to ethylene (25) and quantified by gas chromatography. For JA and OPDA determination, leaf tissue from three independent plants (≈0.5 g of fresh weight in total) per sample was harvested before and after the flooding stimulus, pooled, and stored at -80°C. JA and OPDA levels were determined upon derivatization to JA- and dn-OPDA pentafluoro-benzyl esters as described in ref. 26. For SA determination, leaf tissue from three independent plants (0.5 g of fresh weight each) per sample was harvested before and after the flooding stimulus. Free and total SA values were determined as described in ref. 27. For ethylene determination, leaf discs from four independent plants (100 mg of fresh weight in total) per sample were harvested before and 1 h after elicitation into a glass vial that was sealed air-tight and incubated at room temperature for 30 min before storage at -80°C. Ethylene concentration was determined in duplicate air samples of 500 μl by gas chromatography.
Results and Discussion
To dissect CDPK-mediated transduction of external stress, a full-length (VKJC) or a truncated variant (VK) of NtCDPK2 were ectopically expressed in N. benthamiana leaves by using Agrobacterium-based transient transformation (Fig. 1 A). The truncated variant lacked its regulatory J and C domains and was therefore expected to act as a dominant, constitutively active kinase (28, 29). Expression of NtCDPK2 or its truncated variant did not cause any visual phenotypic difference up to 5 days after infection with the respective Agrobacterium strain (data not shown). However, in contrast to leaves expressing full-length NtCDPK2, the leaves expressing VK responded even to a mild abiotic stress stimulus, such as a single local injection of water (hypoosmotic stress) (Fig. 1B) or wounding (data not shown) with hypersensitive reaction (HR)-like cell death. This necrosis, once initiated by the stress stimulus, spread from the initial site of infiltration and propagated through the entire leaf. The same response was observed when a HA-tagged VK variant of NtCDPK2 was expressed (data not shown). No propagating cell death occurred with the full-length or kinase-inactive truncated variant VK(D/A) or in controls containing an empty vector or a GFP transgene (Fig. 1B). Also, no secondary stress-activated cell death was observed when a HA-tagged VK variant of NtCDPK3 (Fig. 1B) (13) or a NtCDPK2 variant consisting only of the kinase domain was expressed (data not shown). Western blot analysis confirmed that all proteins were expressed to comparable levels (Fig. 1C and data not shown). Thus, the VK variant of NtCDPK2 still retains cellular regulation, localization, and isoform specificity, and the observed cell death phenotype is not simply due to overexpression of a truncated CDPK. In contrast to gain-of-function studies in MAPK signaling based on the expression of constitutively active MAPK kinase variants (6, 10, 30), the VK variant of NtCDPK2 was not constitutively active with regard to the induction of plant cell death. Rather, the VK-triggered cell death response remained stimulus-dependent, suggesting that further posttranslational modifications are involved in CDPK regulation in addition to calcium binding. This stimulus dependency allowed us to dissect specific NtCDPK2-mediated signaling events.
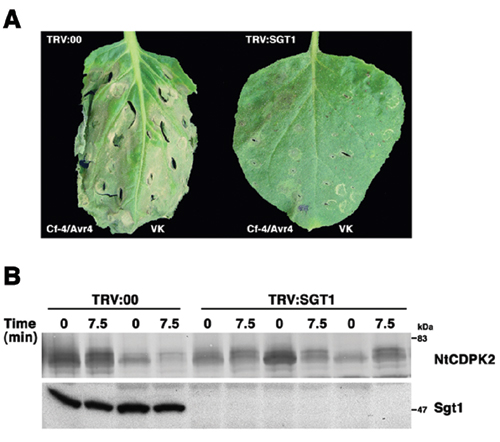
The application of a mild abiotic stress, such as the infiltration of water, would usually not cause cell death symptoms in N. benthamiana. To address whether the VK-dependent necrosis is mechanistically related to the HR activated in biotic gene-for-gene interactions, we compared both responses in plants silenced for the SGT1 gene. SGT1 was first characterized in yeast as a component of the SCF E3 ubiquitin ligase complex. SGT1 homologs in plants were recently shown to be specifically required for various plant defense response pathways (31). Leaf halves of SGT1-silenced and nonsilenced control plants were subjected to Avr4/Cf-4-induced biotic stress or to the expression of VK followed by an abiotic flooding stimulus. Whereas cell death occurred at day 4 in nonsilenced control leaves, no necrosis developed on either leaf half when the SGT1 gene was silenced (Fig. 6, which is published as supporting information on the PNAS web site). Thus, SGT1 may function downstream of NtCDPK2, and negative regulators of cell death and the defense response may become phosphorylated and subjected to SGT1- dependent degradation.
Furthermore, the VK-dependent propagating HR was preceded by an increased production of hydrogen peroxide at sites of wounding or infiltration (Fig. 1D). No accumulation of hydrogen peroxide visualized as brown diaminobenzidine precipitates was observed in leaves expressing the truncated kinase-inactive VK(D/A) or the full-length variant of NtCDPK2.
VK-mediated HR and accumulation of hydrogen peroxide were compromised in the presence of the NtCDPK2's intrinsic autoinhibitory J peptide delivered in trans (Fig. 1 B and D). Fused to a tandem GFP protein, the junction peptide (J-GG) or, as control, the tandem-GFP (GG) alone (Fig. 1 A) was coexpressed with the VK variant (Fig. 1C). Coexpression of J-GG but not GG suppressed VK-dependent responses after an abiotic stress stimulus in vivo, indicating that the VK protein can still be regulated by the autoinhibitory J-domain. This result provides independent proof for the VK variant retaining its CDPK-specific regulation of downstream responses and validates our experimental system for dissecting NtCDPK2-mediated signaling events.
In leaves expressing the VK variant, a local abiotic stress stimulus caused a HR-like response commonly only observed after application of a strong biotic stress. To further corroborate this overlap between NtCDPK2-mediated abiotic and biotic stress responses, we examined the transcriptional activation of pathogenesis-related (PR) genes by determining mRNA levels after the infiltration stimulus (Fig. 2A). VK-expressing leaves showed an enhanced and prolonged induction of HinI, a HR marker gene in tobacco (32), and tpoxC1, a gene differentially expressed in N-gene-mediated resistance to tobacco mosaic virus in tobacco (33); conversely, a delayed and reduced transient HinI transcript accumulation peaking at 1-2 h was observed in control leaves or leaves expressing full-length NtCDPK2 or the kinase-inactive VK(D/A) variant. Acidic PR genes PR1a and PR2a, which are usually under the control of SA-regulated pathways, remained at low transcript levels in VK-expressing leaves, whereas transcript accumulation occurred in the control or in leaves containing VK(D/A) (Fig. 2A). In contrast, in leaves synthesizing the VK variant but not in control leaves, a strong constitutive transcript accumulation was observed for the basic PR genes PR1b and PR2b and the wound-responsive gene PI-II, whose transcriptional induction was reported to be triggered by JA and ethylene-mediated signaling pathways. An intermediate activation for PR2b could be observed when full-length NtCDPK2 was expressed, and these PR2b transcript levels were consistent with intermediate or high accumulation of PR2b protein found in protein extracts from full-length or VK expressing leaves, respectively (data not shown). Elevated PR gene mRNA accumulation observed before application of a secondary stress stimulus likely was caused by constant weak stimulation of the leaves because of the presence of agrobacteria.
Fig. 2.

NtCDPK2 signaling activates JA and ethylene signal transduction pathways. (A) Northern blot analysis of total RNA isolated from leaves expressing NtCDPK2 variants over a time course after an abiotic infiltration stimulus. Duplicate membranes were hybridized with probes for the HR markers HinI and tpoxC1, the acidic and basic PR1 and PR2 genes, and the wound-responsive gene PI-II, respectively. Equal loading of RNA was confirmed by methylene-blue (MB) staining before hybridization. (B) Phytohormone levels. Pooled tissue from three to four independent leaves per sample expressing NtCDPK2 full-length (VKJC; circles) or VK (diamonds) or from control leaves (squares) was harvested over a time course after an abiotic stress stimulus and the levels of JA, OPDA, ACC, and SA were determined.
To test whether NtCDPK2-mediated signal transduction triggers the synthesis of distinct stress hormones or their precursors, we determined the levels of SA, JA, OPDA, and ACC, the precursor of ethylene (Fig. 2B). Leaves expressing VK contained elevated basal levels of JA and OPDA, and a further 5- to 10-fold increase was observed after the abiotic infiltration stimulus, compared with control leaves or leaves synthesizing full-length NtCDPK2. Also, VK-expressing leaves showed significantly (25-fold) higher basal levels of ACC and a further rapid and transient increase could be detected peaking at 30 min after the abiotic stimulus (Fig. 2B). Expression of full-length NtCDPK2 resulted in intermediate ACC levels, and only minor ACC production was provoked by the infiltration stimulus in control leaves. In accordance with the SA-regulated PR1a transcription pattern, no NtCDPK2-dependent elevation of SA levels occurred, whereas high amounts accumulated in control leaves (Fig. 2B). When endogenous levels of SA were reduced by concomitant expression of the NahG gene coding for a salicylate-hydroxylase (34), the VK-dependent HR still developed (data not shown). Consistently, although SA levels were observed in tomato cotyledons in a Cf gene-dependent interaction (35), SA was not required for the Cf-9-mediated resistance in tomato (27). In addition, elicitation of leaves expressing Cf-9 with Avr9 resulted in ACC production (Fig. 5E). Furthermore, our results obtained with the VK variant correlated with expression profiling data for Avr9/Cf-9 rapidly expressed genes in tobacco, where components of ethylene and JA biosynthesis and respective downstream signaling pathways were identified (36). This finding demonstrates that NtCDPK2 is a point of convergence in plant stress signal transduction that controls overlapping abiotic and biotic stress responses by activating ethylene and JA pathways. In Arabidopsis, a synergism between these two hormones is required for the induction of ethylene response factor 1, a transcription factor that regulates differential gene expression in response to pathogen or herbivore attack (37).
Whether NtCDPK2 is directly involved in phytohormone biosynthesis by controlling key regulatory enzymes is unclear. The spreading behavior of the VK-dependent HR beyond the initial wounding sites indicates the presence of a systemic signal. The VK-dependent accumulation of hydrogen peroxide occurred predominantly adjacent to the vasculature, where, interestingly, the machinery for JA synthesis has also been localized (26). This correlation suggests that, during pathogen defense or stress responses, NtCDPK2 may indirectly be involved in the generation of a systemic signal, which could be found among the classes of enzymatically and nonenzymatically derived oxygenated fatty acid molecules, such as oxylipins or phytoprostanes (38).
The rate-limiting step in ethylene production is catalyzed by ACC synthase (ACS). Different subgroups of ACS have been identified and were shown to require protein phosphorylation for regulation. Tomato LeACS2 was reported to be phosphorylated by a CDPK and the identified phosphorylation site in LeACS2 is conserved in some other ACS members (39, 40). Activation of Arabidopsis ACS5 required the dissociation of ETO1 protein, a process that likely involves phosphorylation at a site differing from a MAPK phosphorylation motif (41, 42). Conversely, a MAPK cascade activating MPK6 was shown to regulate ACS6 activity in Arabidopsis, and the expression of gain-of-function MAPK kinase mutants causing constitutive activation of respective MAPKs resulted in constitutive ethylene biosynthesis (10).
To investigate a potential connection or overlap between CDPK and MAPK pathways during induction of plant stress responses, we analyzed MAPK activation after application of both abiotic and biotic stress conditions. In accordance with our previous results in stably transformed Cf-9 tobacco plants (21), in N. benthamiana leaves expressing a TAP-tagged version of Cf-9 (43) elicitation with Avr9 caused a transient activation of the SIPK and WIPK peaking at 30 min after elicitation (Fig. 3A). Concomitantly, strong accumulation of ethylene occurred within 1 h after elicitation in the gene-for-gene interaction, whereas substantially lower ethylene production was observed in the controls lacking Avr9 or Cf-9 (Fig. 3E). When applying a mild abiotic stress by infiltration of water to N. benthamiana leaves, transient activation of SIPK and WIPK was observed peaking 10 min after the stress stimulus (Fig. 3B). Remarkably, in leaves that expressed a truncated VK variant of NtCDPK2, this stimulus-induced activation of both MAPKs was compromised, compared with leaves expressing either full-length VKJC or kinase-inactive VK(D/A) variants (Fig. 3B). The same interference between NtCDPK2 signaling and SIPK and WIPK activation was observed during the biotic Avr9/Cf-9 interaction, where the strong MAPK activation was abrogated in leaves expressing the VK variant (Fig. 3C). Expression of full-length NtCDPK2 resulted in an intermediate inhibitory effect on Avr9/Cf-9-dependent MAPK activation. Leaves expressing the VK variant contained elevated basal levels of ethylene at time point 0 and a further accumulation occurred upon Avr9/Cf-9 interaction (Fig. 3E). Otherwise, no difference could be observed in Avr9/Cf-9-dependent ethylene production before and 1 h after elicitation, when NtCDPK2 variants were coexpressed. Therefore, full activation of SIPK and WIPK is dispensable for the stress-induced synthesis of ethylene and, as seen in Fig. 1, for the onset of hydrogen peroxide production and the HR-like cell death response. This result indicates that at least one additional signaling pathway exists that is sufficient to induce ethylene production and activate further downstream defense responses. This hypothesis is supported by the fact that Arabidopsis plants silenced for the SIPK ortholog MPK6 still responded to pathogen infection with (although reduced) ethylene biosynthesis and developed HR symptoms (10). To investigate whether ethylene itself may be required for connecting CDPK and MAPK signaling, leaves expressing NtCDPK2 variants were pretreated with pharmacological inhibitors of either ethylene synthesis (AVG) or ethylene perception (silver thiosulphate) (Fig. 3E), and MAPK activation was detected before and 30 min after elicitation with Avr9. Remarkably, upon treatment with either inhibitor and, thus, under conditions of blocked ethylene signal transduction, the Avr9/Cf-9-induced activation of SIPK and WIPK was entirely restored, even in the presence of VK or full-length NtCDPK2 (Fig. 3D and data not shown). The application of the respective inhibitors alone or together with intercellular fluid (Avr9-) did not cause MAPK activation (data not shown). Our data demonstrate that high levels of ethylene due to enhanced NtCDPK2 signaling during VK expression are responsible for the observed inhibition of stimulus-dependent MAPK activation. These data may seem contradictory to experiments for which MAPK-dependent induction of ethylene production was shown upon expression of gain-of-function MAPK kinase mutants (10, 30, 44). However, the target of CDPK-triggered MAPK inhibition through ethylene may not necessarily be the MAPK itself but may reside at a component of the signaling cascade further upstream, for example the MAPK kinase. Upon expression of constitutively active MAPK kinase mutants, such a feedback mechanism would have been missed.
Fig. 3.

NtCDPK2 signaling interferes with MAPK activation via the ethylene perception pathway. (A) MAPK activation of WIPK and SIPK after biotic Avr9/Cf-9 gene-for-gene interaction. N. benthamiana leaves expressing TAP-tagged Cf-9 or control leaves were infiltrated with intercellular fluid containing the Avr9 peptide (+) or a control fluid (-), and samples were taken at the time points indicated. Cf-9 protein was determined by Western blot analysis with anti-c-myc antibody (Upper); SIPK and WIPK activation was analyzed by an in-gel protein kinase assay with myelin basic protein as substrate (Lower). (B) Activation of WIPK and SIPK before and 10 and 30 min after abiotic infiltration stimulus in leaves expressing NtCDPK2 variants. Protein expression of NtCDPK2 variants was determined by Western blot. (Upper) NtCDPK2 modification by phosphorylation can be observed as a shift in mobility. (Lower) MAPK activation was determined by in-gel kinase assay. (C) MAPK activation of WIPK and SIPK in the Avr9/Cf-9 gene-for-gene interaction in leaves coexpressing Cf-9 and NtCDPK2 variants. (D) MAPK activation before and 30 min after elicitation in the Avr9/Cf-9 gene-for-gene interaction as in C except that leaves were pretreated with 40 μM AVG or 40 μM silver thiosulphate (AgTS) before infiltrating Avr9. (E) Ethylene production before (white bars) and 60 min after (black bars) elicitation in the Avr9/Cf-9 gene-for-gene interaction as described in A or after elicitation of leaves coexpressing Cf-9 and VK pretreated with AVG or AgTS as indicated.
Our data suggest that a balanced interplay between parallel signaling branches including MAPK- and CDPK-dependent cascades controls the induction of specific stress and defense responses (Fig. 4). In response to environmental stress, CDPK and MAPK signal transduction cascades become activated in parallel. Parallel signaling branches not only provide a back-up system to guarantee multiple activation events in reaction to one stimulus but also allow fine-tuning of responses by regulating different sets of partially overlapping reactions. Thereby, different isoforms and, in case of ethylene biosynthesis, for example, different ACS members (45) may become activated depending on the signaling branch. One may further assume that once downstream stress and pathogen responses are induced and high levels of ethylene are generated, a feedback mechanism comes into place to reset the system and switch off various reactions. The CDPK-mediated inhibition of SIPK and WIPK activation through ethylene may be part of such a resetting system. Negative communication between these distinct pathways may enable the plant to attenuate and switch off responses once the original triggering stimulus has been removed. Finally, the existence of parallel signaling branches to stress tolerance may explain why no MAPK or CDPK has so far been identified by forward genetic screens, because functional redundancy may not only take place within one class of signal mediators, such as MAPK cascades, but may also encompass different branches, for example, CDPKs.
Fig. 4.

Model for cross-talk between CDPK and MAPK signaling pathways in tobacco. Biotic and abiotic stress stimuli trigger inside a cell parallel MAPK- and CDPK-dependent signal transduction pathways. Partially overlapping subsets of early stress responses become activated and control the induction of various stress and defense responses. The expression of the truncated NtCDPK2-VK variant caused prolonged signaling after stress (CDPK*) and resulted in enhanced activation of NtCDPK2-dependent downstream responses (bold). These responses include H2O2 production after activation of an NADPH oxidase, synthesis of ethylene and oxylipins, induction of basic PR gene expression and initiation of cell death. NtCDPK2 may also control negative regulators of the hypersensitive cell death that, upon phosphorylation, would be destroyed by the SGT1-dependent proteolysis machinery. NtCDPK2-mediated inhibition of SIPK and WIPK activation through ethylene may be part of a resetting system and provides a link between the two signaling pathways.
Supplementary Material
Acknowledgments
We thank J. Peart and P. Moffet (The Sainsbury Laboratory) for the SGT1-silenced plants and K. Shirasu (The Sainsbury Laboratory) for providing the anti-SGT1 antibody. We thank G. Boison and H. Bothe for sharing gas chromatography facilities (Cologne University, Cologne, Germany). This research was supported by the Gatsby Charitable Foundation, Cross-Talk in Signaling in Plants Project European Commission Grant HRPN/CT-2000-00093, and the Max Planck Society. A.A.L. was supported by a European Community Marie Curie Fellowship and European Commission Grant HPMF/CT-2001-01288. T.R. was funded by the Program for Investment in the Future (ZIP) from the German Ministry of Education and Science and the Alexander von Humboldt Foundation.
This paper was submitted directly (Track II) to the PNAS office.
Abbreviations: CDPK, calcium-dependent protein kinase; NtCDPK, Nicotiana tabacum CDPK; MAPK, mitogen-activated protein kinase; SA, salicylic acid; SIPK, SA-induced protein kinase; WIPK, wound-induced protein kinase; OPDA, 12-oxo-phytodienoic acid; JA, jasmonic acid; ACC, 1-aminocyclopropane-1-carboxylate; PR, pathogenesis-related; HR, hypersensitive reaction; HA, hemagglutinin; AVG, aminoethoxyvinylglycine; ACS, ACC synthase; J, junction domain; C, C-terminal calmodulin-like domain; K, protein kinase domain; V, variable domain.
References
- 1.Romeis, T. (2001) Curr. Opin. Plant Biol. 4, 407-414. [DOI] [PubMed] [Google Scholar]
- 2.Peck, S. C. (2003) Curr. Opin. Plant Biol. 6, 334-338. [DOI] [PubMed] [Google Scholar]
- 3.Jonak, C., Okresz, L., Bogre, L. & Hirt, H. (2002) Curr. Opin. Plant Biol. 5, 415-424. [DOI] [PubMed] [Google Scholar]
- 4.Ren, D. T., Yang, H. P. & Zhang, S. Q. (2002) J. Biol. Chem. 277, 559-565. [DOI] [PubMed] [Google Scholar]
- 5.Kim, C. Y., Liu, Y., Thorne, E. T., Yang, H., Fukushige, H., Gassmann, W., Hildebrand, D., Sharp, R. E. & Zhang, S. (2003) Plant Cell 15, 2707-2718. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.del Pozo, O., Pedley, K. F. & Martin, G. B. (2004) EMBO J. 23, 3072-3082. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jin, H., Liu, Y., Yang, K. Y., Kim, C. Y., Baker, B. & Zhang, S. (2003) Plant J. 33, 719-731. [DOI] [PubMed] [Google Scholar]
- 8.Sharma, P. C., Ito, A., Shimizu, T., Terauchi, R., Kamoun, S. & Saitoh, H. (2003) Mol. Gen. Genomics 269, 583-591. [DOI] [PubMed] [Google Scholar]
- 9.Asai, T., Tena, G., Plotnikova, J., Willmann, M. R., Chiu, W. L., Gomez-Gomez, L., Boller, T., Ausubel, F. M. & Sheen, J. (2002) Nature 415, 977-983. [DOI] [PubMed] [Google Scholar]
- 10.Liu, Y. & Zhang, S. (2004) Plant Cell 16, 3386-3399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Menke, F. L. H., van Pelt, J. A., Pieterse, C. M. J. & Klessig, D. F. (2004) Plant Cell 16, 897-907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Rivas, S. & Thomas, C. M. (2002) Mol. Plant Pathol. 3, 277-282. [DOI] [PubMed] [Google Scholar]
- 13.Romeis, T., Ludwig, A. A., Martin, R. & Jones, J. D. G. (2001) EMBO J. 20, 5556-5567. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Scrase-Field, S. A. & Knight, M. R. (2003) Curr. Opin. Plant Biol. 6, 500-506. [DOI] [PubMed] [Google Scholar]
- 15.Ludwig, A. A., Romeis, T. & Jones, J. D. G. (2004) J. Exp. Bot. 55, 181-188. [DOI] [PubMed] [Google Scholar]
- 16.Harper, J. F., Breton, G. & Harmon, A. (2004) Annu. Rev. Plant Physiol. Plant Mol. Biol. 55, 263-288. [DOI] [PubMed] [Google Scholar]
- 17.Davies, K. M., Hobson, G. E. & Grierson, D. (1988) Plant Cell Environ. 11, 729-738. [Google Scholar]
- 18.Bevan, M. (1984) Nucl. Acid Res. 12, 8711-8721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bendahmane, A., Kanyuka, K. & Baulcombe, D. C. (1999) Plant Cell 11, 781-791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Jones, L., Hamilton, A. J., Voinnet, O., Thomas, C. L., Maule, A. J. & Baulcombe, D. C. (1999) Plant Cell 11, 2291-2301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Romeis, T., Piedras, P., Zhang, S. Q., Klessig, D. F., Hirt, H. & Jones, J. D. G. (1999) Plant Cell 11, 273-287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Sambrook, F., Fritsch, E. F. & Maniatis, T. (1989) in Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Lab. Press, Plainview, NY), 2nd Ed.
- 23.Church, G. M. & Gilbert, W. (1984) Proc. Natl. Acad. Sci. USA 81, 1991-1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Rivas, S., Rougon-Cardoso, A., Smoker, M., Schauser, L., Yoshioka, H. & Jones, J. D. G. (2004) EMBO J. 23, 2156-2165. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 25.Lizada, M. C. & Yang, S. F. (1979) Anal. Biochem. 100, 140-145. [DOI] [PubMed] [Google Scholar]
- 26.Stenzel, I., Hause, B., Miersch, O., Kurz, T., Maucher, H., Weichert, H., Ziegler, J., Feussner, I. & Wasternack, C. (2003) Plant J. 33, 577-589. [DOI] [PubMed] [Google Scholar]
- 27.Brading, P.A., Hammond-Kosack, K. E., Parr, A. & Jones, J. D. G. (2000) Plant J. 23, 305-318. [DOI] [PubMed] [Google Scholar]
- 28.Harper, J. F., Huang, J. F. & Lloyd, S. J. (1994) Biochemistry 33, 7267-7277. [DOI] [PubMed] [Google Scholar]
- 29.Sheen, J. (1996) Science 274, 1900-1902. [DOI] [PubMed] [Google Scholar]
- 30.Yang, K.Y., Liu, Y. & Zhang, S. (2001)) Proc. Natl. Acad. Sci. USA 98, 741-746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Peart, J. R., Lu, R., Sadanandom, A., Malcuit, I., Moffett, P., Brice, D. C., Schauser, L., Jaggard, D. A., Xiao, S., Coleman, M. J., et al. (2002) Proc. Natl. Acad. Sci. USA 99, 10865-10869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Gopalan, S., Wei, W. & He, S. Y. (1996) Plant J. 10, 591-600. [DOI] [PubMed] [Google Scholar]
- 33.Hiraga, S., Ito, H., Yamakawa, H., Ohtsubo, N., Seo, S., Mitsuhara, I., Matsui, H., Honma, M. & Ohashi, Y. (2000) Mol. Plant Microbe Interact. 13, 210-216. [DOI] [PubMed] [Google Scholar]
- 34.Gaffney, T., Friedrich, L., Vernooij, B., Negrotto, D., Nye, G., Uknes, S., Ward, E. & Ryals, J. (1993) Science 261, 754-756. [DOI] [PubMed] [Google Scholar]
- 35.Hammond-Kosack, K. E., Silverman, P., Raskin, I. & Jones, J. D. G. (1996) Plant Physiol. 110, 1381-1394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Durrant, W. E., Rowland, O., Piedras, P., Hammond-Kosack, K. E. & Jones, J. D. G. (2000) Plant Cell 12, 963-977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lorenzo, O., Piqueras, R., Sanchez-Serrano, J. J. & Solano, R. (2003) Plant Cell 15, 165-178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Farmer, E. E., Almeras, E. & Krishnamurthy, V. (2003) Curr. Opin. Plant Biol. 6, 372-378. [DOI] [PubMed] [Google Scholar]
- 39.Sebastia, C. H., Hardin, S. C., Clouse, S. D., Kieber, J. J. & Huber, S. C. (2004) Arch. Biochem. Biophys. 428, 81-91. [DOI] [PubMed] [Google Scholar]
- 40.Tatsuki, M. & Mori, H. (2001) J. Biol. Chem. 276, 28051-28057. [DOI] [PubMed] [Google Scholar]
- 41.Ecker, J. R. (2004) Plant Cell 16, 3169-3173. [DOI] [PMC free article] [Google Scholar]
- 42.Wang, K. L. C., Yoshida, H., Lurin, C. & Ecker, J. R. (2004) Nature 428, 945-950. [DOI] [PubMed] [Google Scholar]
- 43.Rivas, S., Romeis, T. & Jones, J. D. G. (2002) Plant Cell 14, 689-702. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 44.Ouaked, F., Rozhon, W., Lecourieux, D. & Hirt, H. (2003) EMBO J. 22, 1282-1288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yamagami, T., Tsuchisaka, A., Yamada, K., Haddon, W. F., Harden, L. A. & Theologis, A. (2003) J. Biol. Chem. 278, 49102-49112. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}