

Abstract
The present study tested a formal, or purely mathematical, theory of matching, and a modern account derived by McDowell (1986) that incorporates deviations from strict matching—bias and sensitivity. Six humans pressed a lever for monetary reinforcers on five concurrent variable interval (VI) schedules of reinforcement. All schedules were presented during each session. The magnitude on one alternative remained constant, and five magnitudes were presented across sessions on the other alternative. To test the formal account, two absolute response rate equations were fitted to the response and reinforcement rates at each alternative at each magnitude. Although the equations accounted for a high percentage of variance, there was a significant negative correlation between the standardized residuals and the predicted response rates. To test the modern account, an ensemble of four equations was fitted to the data. The equations predicted relative and absolute responding, and the independent variables in each equation were adjusted for bias and sensitivity. The equations accounted for a high percentage of variance, and the standardized residuals were not correlated with the predicted response rates. The values of the parameters were consistent with empirical findings and theoretical predictions, including the prediction that k should remain constant across changes in reinforcer magnitude. The results suggest that the formal theory of matching does not describe the data, and that the modern theory may provide an accurate and coherent description of concurrent and single-alternative responding.
Keywords: matching theory, concurrent schedules, reinforcer magnitude, variable-interval schedules, lever press, humans
Beyond the collection of uniform relationships lies the need for a formal representation of the data reduced to a minimal number of terms. A theoretical construction may yield greater generality than any assemblage of facts. . . . It will not stand in the way of our search for functional relations because it will arise only after relevant variables have been found and studied. Though it may be difficult to understand, it will not be easily misunderstood . . . (Skinner, 1950/1972, p. 100)
Skinner's assessment of theory was tempered by an emphatic recommendation that we must first establish an experimental analysis of how relevant variables affect behavior (Skinner, 1950/1972). In the case of a quantitative theory, for instance, the progression from experimental analysis to theory should increase the likelihood that the theory's parameters reflect the operation of definite variables and processes, rather than simply being “arbitrary constants.” Of course, a progression from theory to further experimental analysis may also reveal novel functional relations and behavioral processes. Regardless of the sequence, quantitative theory may increase the generality and precision of our understanding of environment–behavior relations. Just as the universal law of gravitation yields considerable predictive and practical advantages over the statement that objects fall when dropped, a quantitative theory of behavior can move us beyond the statement that operant responding increases when reinforced (or any similar “assemblage of facts”). In short, a quantitative theory should improve our ability to predict and influence behavior, which is a hallmark of behavior-analytic science.
In the 55 years since Skinner made his remarks about theory, operant psychologists have generated a wealth of orderly data from single and concurrent VI schedules of reinforcement, and several quantitative theories have emerged to provide a coherent account of performance on both types of schedules. Although matching theory has been the most investigated quantitative account of VI performance, a number of studies now suggest that Herrnstein's (1961, 1970; de Villiers & Herrnstein, 1976) original, or classic, theory of matching requires some revision (Dallery, McDowell, & Lancaster, 2000; Dallery, McDowell, & Soto, 2004; McDowell, 2005; McDowell & Dallery, 1999; McDowell & Wood, 1984, 1985; Soto, McDowell, & Dallery, 2005; Warren-Boulton, Silberberg, Gray, & Ollom, 1985; cf. Heyman & Monaghan, 1987, 1994; see Dallery & Soto, 2004, for a review). As we will discuss in more detail below, the purpose of the present study is to test two alternatives to Herrnstein's theory. First, we will review why such revisions are necessary.
The classic theory begins with the proposition that the proportion of responding at each of two choice alternatives matches the proportion of reinforcement obtained from those alternatives (Herrnstein, 1961), or:
 |
where R1 and r1 represent the rates of responding and reinforcement on Alternative 1, and R2 and r2 represent the rates of responding and reinforcement on Alternative 2, respectively. Herrnstein argued that even in situations in which one response alternative was arranged by the experimenter, extraneous alternatives exist (e.g., rearing, scratching, smelling, etc). In such a situation, Equation 1 can be rewritten as:
 |
where Re and re refer to the aggregate amount of responding and aggregate amount of reinforcement delivered on extraneous alternatives.
Then, by assuming that R1 and Re are exhaustive of the total amount of behavior possible in the environment and that this amount is constant across variations in reinforcer properties (e.g., reinforcer magnitude), Herrnstein let k = R1 + Re and then solved for R1, which produced:
 |
The subscript on R has been dropped because Equation 2 is fitted to only one response alternative. Thus, based on Equation 1, and with the assumption that the total amount of behavior, k, remains constant in a given environment, Herrnstein derived a hyperbolic equation and a theory to describe how reinforcement affects response strength (de Villiers & Herrnstein, 1976; Herrnstein, 1970, 1974).
Classic matching theory (Equation 2 and the assumptions that gave rise to it), however, is logically and empirically inconsistent with experimental findings. First, choice almost always deviates from strict, proportional matching (e.g., Baum, 1974, 1979; McDowell, 2005; Williams, 1988), which violates the mathematical propositions, Equations 1 and 1b that underlie Equation 2. Second, k varies with reinforcer magnitude (Bradshaw, Szabadi, & Bevan, 1978; Dallery, McDowell, & Lancaster, 2000; de Villiers, 1977; McDowell & Dallery, 1999; McDowell & Wood, 1984, 1985; Snyderman, 1983; cf. Heyman & Monaghan, 1987, 1994; see Dallery & Soto, 2004, for a review), which violates the assumption that the total amount of behavior is constant.
At least two alternatives to Herrnstein's classic theory are possible. First, McDowell (1986) has proposed a formal, purely mathematical theory of matching, but the formal theory has not been subjected to empirical test. Second, parameters can be incorporated into the concurrent and hyperbolic equations (Baum, 1974, 1979; McDowell, 1986) to account for deviations from strict matching. The addition of these parameters is not arbitrary; rather, they reflect the operation of particular variables on choice. As such, they may broaden the generality and precision of the matching account. To distinguish the equations with the additional parameters from the original, classic theory, and from the formal theory, we will refer to them collectively as the modern theory of matching. The purpose of the present experiment was to test the application of the formal and modern versions of matching to humans responding under concurrent schedules of monetary reinforcement.
Formal matching theory simply describes relations between rates of reinforcement and responding on concurrent- and single-alternative schedules. In contrast, Herrnstein's classic theory asserted that measures such as response rate, latency, duration, and force can represent response strength. The theory also asserted that response strength is a function of relative reinforcement, which can be manipulated by varying the rate, delay, magnitude, or some other dimension of reinforcement (de Villiers & Herrnstein, 1976). Despite these differences in the scope of the two theories, the formal theory begins in the same way as the classic theory: with proportional matching equations to describe choice in a concurrent schedule. The matching equations for the two arranged alternatives in a concurrent-schedule are:
 |
and
 |
where R1 and R2 refer to the response rate at each of two experimentally programmed alternatives, r1 and r2 refer to the reinforcement rate at each alternative, and re and Re represent the rate of extraneous reinforcement and responding, respectively. The organism can allocate behavior to one of two experimentally arranged alternatives or engage in extraneous responding (the equation for extraneous responding is not shown). The equations state that the proportion of responding at each alternative matches the proportion of reinforcement obtained at that alternative.
From these concurrent equations, the absolute response rate forms are obtained. These equations are obtained in a similar manner as the more familiar hyperbolic equation where only one schedule is explicitly arranged (Herrnstein, 1970). For the first alternative, let k = R1 + R2 + Re and then solve for R1:
 |
For the second alternative, again let k = R1 + R2 + Re and then solve for R2:
 |
It is important to note that the ks must be equal across both equations because both ks represent the same sum of response rates, R1 + R2 + Re.
The key test for formal theory, therefore, is whether Equations 4a and 4b accurately describe responding by using a single k. That is, within the same concurrent schedule, the value of k in both equations must be the same when each is applied to the respective alternative (i.e., 4a to Alternative 1 and 4b to Alternative 2). To illustrate this point, and for convenience, the general procedure of the current experiment will be outlined. First, we arranged a concurrent schedule with a different reinforcer magnitude at each alternative, and then varied reinforcement rates at each alternative. Then, we fitted Equations 4a and 4b to data from each alternative, where one value of k was shared by both equations. Having done so, the question was: Does each equation describe responding at each alternative (i.e., high percentage of variance accounted for and random residuals)? If the same k can be used in both equations, then formal theory would be supported.
To illustrate the difference between formal theory's equal-k requirement and classic theory's constant-k requirement, consider a situation in which a second concurrent schedule with different reinforcer magnitudes is arranged. For classic theory, k is constant at both alternatives across both magnitude pairs. Formal theory requires, within a magnitude pair, a single k because the k in each equation represents the same sum. However, the ks across the first and second magnitude pairs can vary because the sums of response rates (ks) in the first and second magnitude pairs do not represent the same sums. There is no theoretical constant-k requirement as in the classic theory. Thus formal theory's equal-k requirement is different than the classic theory's constant-k requirement.
The logic of the development of the formal theory is equivalent to the logic of the development of the classic theory. Because both theories begin with proportional matching equations, the formal theory is subject to the same criticism that McDowell (2005) leveled against the classic theory. That is, because departures from proportional matching are the rule rather than the exception, and because the absolute response rate equations are derived from the proportional matching equations, both the classic and formal theories of single variable-interval (VI) responding are inconsistent with empirical findings. The variables affecting behavior in a concurrent environment that cause deviations from strict matching also should affect behavior in a single VI environment. Similarly, these variables should cause deviations from strict matching when the absolute rate equations are applied to one response alternative in a concurrent arrangement. If all behavior is choice (Herrnstein, 1970), then the same variables should prevail regardless of the number of explicitly arranged alternatives. Despite this criticism, the test of the equal-k requirement still represents a valid test of a key feature of the formal theory.
The modern theory is a set of equations that incorporate deviations from strict matching into the concurrent and absolute response rate equations. The starting point for the modern version of matching is Baum's (1974, 1979) generalized matching equation, or
 |
where b represents bias and a represents the degree of sensitivity to changes in reinforcement rates. The bias parameter can account for asymmetries between the alternatives due to differences in response cost or reinforcer magnitude. We can obtain the absolute response rate forms for each component of a concurrent schedule from Equation 5 in the same manner outlined above for the classic theory (McDowell, 1986, 2005). The equations for each alternative are
 |
and
 |
There are three pairs of reinforcement rate ratios, with one bias parameter and one exponent applied to each of the three pairs. McDowell (1986) also showed that the modern theory entails another equation:
 |
Equation 7 includes the bias and sensitivity parameters that apply to the ratios of each arranged alternative and the extraneous alternative. This equation is a consequence of matching theory's requirement that all behavior be allocated among the available response alternatives (McDowell, 1986).
One difference between the formal and modern theories is that they make different assumptions concerning k. Specifically, modern theory retains Herrnstein's assumption that total behavior should remain constant in a given environment. Thus we start with the foundational equation (Equation 5), make the assumption that k remains constant, and then we simply derive the remaining equations algebraically (see McDowell, 1986, for more details on the derivations).
The present study evaluated the formal and modern versions of matching theory by varying reinforcer magnitude at one of two concurrently available response alternatives. The magnitude on the other alternative remained constant. We assessed five unique magnitude pairs. The same response manipulandum and range of VI schedules were used for each alternative (Findley, 1958). The key question for formal theory was whether one k per magnitude pair could describe the data (the equal-k requirement). For modern theory, an important question was whether one k overall (i.e., the constant-k requirement, or a single k for all fits of Equations 6a and 6b across all magnitude pairs) could describe the data. The fits were evaluated with respect to the quality of the fit (i.e., percentage of variance accounted for), randomness of the residuals, and reasonableness and theoretical consistency of the parameter estimates.
METHOD
Participants
Five females, ranging in age from 23 to 55, and one 20-year-old male agreed to participate. Applicants were recruited through advertisements in a local newspaper. All participants were experimentally naive, and none was a college student.
Apparatus
Experimental sessions were conducted in two windowless experimental rooms measuring 2.3 m by 1.2 m. Participants responded on a MED Associates Human Apparatus (ENV 600A). The apparatus measured 57.1 cm wide by 203.8 cm tall. A metal lever was centered horizontally and protruded from the front of the apparatus 106.7 cm above the floor. The lever required a minimum force of approximately 111.2 N to register a response.
A small pushbutton was located 137.8 cm above the floor and 17.8 cm to the right of center, and a blue stimulus light was mounted 5.7 cm above the pushbutton. A row of 10 white lights was centered 150.5 cm above the floor and signaled different reinforcement schedules. A row of 10 red lights was centered 155.6 cm above the floor and signaled different reinforcer magnitude conditions. A green light 166.4 cm above the floor and 17.8 cm to the right of center served as a session light. Reinforcers were signaled by a brief tone and a flash of an amber light, located 166.4 cm above the floor and 18.5 cm left of center. The monetary value of each reinforcer was added to a digital counter centered 168.3 cm above the floor.
A 15-W houselight situated behind the apparatus provided dim illumination. White noise was employed to mask extraneous sounds. A computer operating under MED-PC® software controlled programming of experimental events and recording of data.
Procedure
During each session, participants pressed the response lever to earn points exchangeable for money on a series of five concurrent VI VI schedules. The mean values were 17 s to 720 s, 25 s to 300 s, 51 s to 51 s, 300 s to 25 s, and 720 s to 17 s. The programmed VI values were determined by Fleshler and Hoffman's (1962) method. The VI schedule used for each alternative was independent of that used for the other alternative.
At the beginning of each session, the houselight, session light, a white light signaling the reinforcement schedule, and a red light signaling the reinforcer magnitude were illuminated. One VI VI pair was presented for 10 min, then a blackout period was presented for 5 min, and then the next pair was presented for 10 min, and so on (Bradshaw, Szabadi, & Bevan, 1976; Dallery et al., 2000), until all schedules were presented. Participants remained standing to make responses on the lever, but were permitted to sit during blackouts. The order of schedules within each session was arranged randomly without replacement. Each VI VI pair was signaled by a unique arrangement of the white and red stimulus lights, which were the same across participants.
Reinforcer magnitude was manipulated across sessions. The monetary values on one alternative (Schedule A) were $0.20, $0.12, $0.07, $0.004, and $0.0025. Reinforced lever presses on the other alternative (Schedule B) always resulted in a $0.07 reinforcer. The Schedule A value varied randomly without replacement across blocks of five sessions. Thus, across five sessions, participants were exposed to all five magnitude combinations.
Participants switched between the two concurrent alternatives by pressing the pushbutton (Findley, 1958). The blue light above the pushbutton signaled a changeover response by turning off if it was on, or turning on if it was off. A response at the changeover button changed the stimulus lights and associated schedules in effect. Following a response at the changeover button, a lever press could not be reinforced for X seconds, which is called a changeover delay (COD) (Herrnstein, 1961). COD values for the first 3 participants (H59, H60, and H64) were determined by exposing them to the five VI VI pairs and varying the COD between 0 s and 5 s. Reinforced responses on both alternatives resulted in $0.07. The changes in COD across 39, 42, and 101 sessions showed no systematic effect on the parameters of the generalized matching equation (i.e., Equation 5) for these 3 participants (H59, H60, and H64, respectively). Specifically, longer CODs did not produce sensitivity parameters closer to one, or perfect matching. Therefore, the COD was set at the final COD value for each of these 3 participants. The COD was 3 s for H59, and 2 s for H60 and H64. Because differences in the COD had little effect for the previous participants, the COD was set conservatively at 5 s for H67, H69, and H70.
Before beginning the experiment, participants were given an orientation session in which they viewed the experimental room, operated the apparatus, and received an explanation of what the experiment entailed. During the orientation, instructions about the experimental sessions were given (see Appendix A).
If the applicant expressed interest in participating, he or she read and signed a consent form that detailed the duration of the experiment, payment terms, and situations where monetary deductions could be assessed (e.g., withdrawal from the study before completion). The local Institutional Review Board approved all study procedures and the consent process.
RESULTS
A time-series analysis, Tryon's C-Statistic (α = 0.1; Tryon, 1982), was used to identify stable response rates across eight-session blocks in each condition. The first three sessions in each condition were excluded from the stability analysis. The analysis was performed on response rates for each alternative of the concurrent schedules. Table 1 lists the session at which the stability criterion was satisfied for each subject. Approximately half of the conditions were stable by Session 11, and the maximum number of sessions was 22. Reinforcement and response rates were averaged over the first stable eight-session block (see Appendix B). For example, if the data for a concurrent pair were stable by Session 11, then Sessions 4 to 11 for that pair were used in the analysis, and if the data were stable at Session 22 for another concurrent pair, then Sessions 15 to 22 were used in the analysis.
Table 1. The first session at which the stability criterion was satisfied for each participant at each concurrent VI VI pair. Response rates were deemed stable across the eight sessions before and including the session indicated below. For each concurrent pair, data were averaged across the eight sessions before and including the session indicated below.
| Participant | Schedule A magnitude | Concurrent VI VI schedules |
||||
| VI 17 – VI 720 | VI 25 – VI 300 | VI 51 – VI 51 | VI 300 – VI 25 | VI 720 – VI 17 | ||
| H59 | $0.2 | 15 | 15 | 11 | 15 | 12 |
| $0.12 | 12 | 13 | 11 | 11 | 13 | |
| $0.07 | 11 | 12 | 13 | 11 | 11 | |
| $0.004 | 11 | 11 | 19 | 19 | 11 | |
| $0.0025 | 11 | 11 | 14 | 20 | 20 | |
| H60 | $0.2 | 12 | 11 | 11 | 11 | 11 |
| $0.12 | 13 | 11 | 16 | 11 | 11 | |
| $0.07 | 11 | 11 | 11 | 16 | 11 | |
| $0.004 | 11 | 11 | 11 | 11 | 11 | |
| $0.0025 | 12 | 11 | 14 | 14 | 11 | |
| H64 | $0.2 | 11 | 13 | 11 | 13 | 12 |
| $0.12 | 11 | 11 | 16 | 11 | 11 | |
| $0.07 | 13 | 11 | 12 | 13 | 11 | |
| $0.004 | 18 | 12 | 14 | 12 | 11 | |
| $0.0025 | 12 | 13 | 11 | 21 | 12 | |
| H67 | $0.2 | 11 | 11 | 11 | 14 | 16 |
| $0.12 | 11 | 14 | 13 | 14 | 11 | |
| $0.07 | 11 | 11 | 11 | 11 | 11 | |
| $0.004 | 12 | 12 | 12 | 11 | 11 | |
| $0.0025 | 12 | 11 | 18 | 15 | 11 | |
| H69 | $0.2 | 11 | 12 | 11 | 16 | 11 |
| $0.12 | 13 | 11 | 11 | 16 | 16 | |
| $0.07 | 12 | 11 | 11 | 11 | 16 | |
| $0.004 | 13 | 12 | 11 | 11 | 13 | |
| $0.0025 | 11 | 12 | 11 | 18 | 12 | |
| H70 | $0.2 | 11 | 13 | 22 | 12 | 12 |
| $0.12 | 13 | 13 | 14 | 12 | 14 | |
| $0.07 | 11 | 11 | 13 | 13 | 11 | |
| $0.004 | 11 | 11 | 18 | 11 | 11 | |
| $0.0025 | 11 | 17 | 19 | 11 | 12 | |
First, the formal theory was tested. Recall that formal theory requires the same k within a magnitude pair. Thus a different k was allowed for each magnitude pair for each participant, which resulted in five ks per participant. In terms of re, the most conservative approach is to hold it constant across concurrent magnitude pairs. That is, insofar as formal theory only consists of rates and does not make predictions regarding how (or if) re should change with magnitude, extraneous reinforcement rate was assumed to remain constant across concurrent magnitude pairs.
Equations 4a and 4b were fitted simultaneously to the data from all magnitude pairs for each participant. The rationale for the simultaneous fitting method is that it permits different equations to share parameter values, which is ideal for testing the equal-k requirement. That is, one k was shared by both equations at each magnitude pair, and one re was shared for all equations at all magnitude pairs. The simultaneous fitting method proceeded as follows: Equation 4a produced a residual sum of squares (RSS) for each magnitude on Schedule A, and Equation 4b produced a RSS for each magnitude on Schedule B. The ratio of the RSS to the total sum of squares (i.e., RSS/SS), summed over all magnitudes, was minimized. In other words, given five magnitudes, five ks and one re can be found such that
 |
is a minimum. The numerical subscripts refer to each magnitude condition and the letters correspond to Schedules A and B. The analysis was performed using Microsoft® Excel's Solver routine. The same method was used for all subsequent fits.
Table 2 shows the results of the regressions. In most cases, the equation accounted for a high percentage of the variance in response rate. To fit the equations to H67's data, we constrained re to be greater than or equal to zero. Otherwise, the re became negative for this participant. The median percentage of variance accounted for (%VAC) and median parameter values for all participants, with the exception of H69, are shown at the bottom of the table. For H69, the mean of the response rates at each magnitude accounted for more variance than the equation, and negative %VACs were obtained for all but one fit ($0.0025; with an obtained %VAC of 7). As can be seen in Appendix B, response rates for this participant showed very little variability despite large changes in reinforcement rate and reinforcer magnitude. Data from this participant were therefore excluded from this analysis.
Table 2. Least square fits of the formal theory to the response rates for each participant. Both Equation 4a and Equation 4b were fitted simultaneously to the Schedule A and Schedule B data and all magnitudes for each participant. One k per magnitude pair and one re overall were obtained for each participant. The dashes under the Magnitude on Schedule A column are used to indicate that re was held constant across magnitudes. The percentages of variance accounted for (%VAC) by Equation 4a and 4b for each magnitude are provided in the right columns.
| Participant | Magnitude on Schedule A | k | re | %VAC |
|
| Equation 4a | Equation 4b | ||||
| H59 | – | 9.5 | |||
| $0.20 | 44.8 | 97 | 99 | ||
| $0.12 | 43.2 | 97 | 98 | ||
| $0.07 | 43.3 | 99 | 98 | ||
| $0.004 | 40.3 | 96 | 94 | ||
| $0.0025 | 40.9 | 99 | 83 | ||
| H60 | – | 13.4 | |||
| $0.20 | 57.5 | 89 | 97 | ||
| $0.12 | 56.4 | 92 | 94 | ||
| $0.07 | 57.7 | 95 | 98 | ||
| $0.004 | 54.3 | 88 | 62 | ||
| $0.0025 | 55.5 | 84 | 51 | ||
| H64 | – | 45.2 | |||
| $0.20 | 63.3 | 93 | 97 | ||
| $0.12 | 56.8 | 97 | 99 | ||
| $0.07 | 58.0 | 99 | 100 | ||
| $0.004 | 52.0 | 95 | 87 | ||
| $0.0025 | 49.2 | 99 | 96 | ||
| H67 | – | 0.0 | |||
| $0.20 | 61.8 | 65 | 85 | ||
| $0.12 | 61.2 | 66 | 79 | ||
| $0.07 | 62.9 | 79 | 81 | ||
| $0.004 | 59.1 | 70 | 55 | ||
| $0.0025 | 59.7 | 69 | 27 | ||
| H70 | – | 69.6 | |||
| $0.20 | 120.5 | 30 | 78 | ||
| $0.12 | 115.9 | 28 | 74 | ||
| $0.07 | 108.2 | 35 | 48 | ||
| $0.004 | 112.0 | 15 | −229a | ||
| $0.0025 | 104.5 | 40 | −101a | ||
| Median | – | 13.4 | |||
| $0.20 | 61.8 | 89 | 97 | ||
| $0.12 | 56.8 | 92 | 94 | ||
| $0.07 | 58.0 | 95 | 98 | ||
| $0.004 | 54.3 | 88 | 62 | ||
| $0.0025 | 55.5 | 84 | 51 | ||
A negative VAC occurs when the average of the data accounts for more variance than the fitted function.
Figure 1 shows the standardized residuals as a function of the response rates predicted by Equations 4a and 4b. Residuals are pooled across magnitudes and participants. The standardized residuals were significantly correlated with the predicted response rates for Schedule A (r = −.47, p < .05) and Schedule B (r = −.56, p < .05). The negative correlation indicates that the equations predicted lower response rates than obtained at low reinforcement rates, and higher response rates than obtained at high reinforcement rates.
Fig. 1. Standardized residuals as a function of the predicted response rates for Equation 4a, Schedule A (top panel) and Equation 4b, Schedule B (bottom panel).

Pearson's correlation coefficients are shown in each panel.
Testing the modern form of Herrnstein's theory requires simultaneously fitting Equations 5, 6a, 6b, and 7 to the data. For purposes of fitting, some of the parameters in Equations 6a, 6b, and 7 cannot be independently estimated. Thus composite parameters must be estimated as follows:
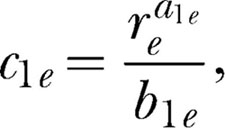 |
and
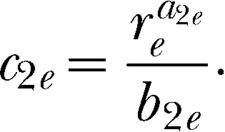 |
We, therefore, can write Equations 6a, 6b, and 7 as
 |
and
 |
Equations 5, 6a′, 6b′ and 7′ were fitted to the data.
The fitting procedure followed the same logic described for the formal theory. Equations 5 and 7′ produced a RSS for the ratio of response rates for each magnitude on Schedules A and B, Equation 6a′ produced a RSS for response rates for each magnitude on Schedule A, and Equation 6b′ produced a RSS for response rates for each magnitude on Schedule B. The ratio RSS/SS, summed over all magnitudes, was minimized. When minimizing this quantity, some parameters were free to vary across magnitude pairs, and some were held constant for a participant. We will discuss the reasons for these decisions about the parameters in more detail below.
Table 3 shows the results of the fits to the data. When a parameter was free to vary across magnitude pairs then several values are listed for a participant. When a parameter was shared by the appropriate equations, or held constant, one value is listed. For example, because all equations in which k appears shared the same k value, which includes fits of Equations 6a′ and 6b′ to each alternative across all magnitude pairs, one k was used. Because reinforcer magnitude varied on Schedule A, bias for this alternative should change across magnitude pairs (Baum, 1974, 1979), and therefore b12 and c1e were allowed to vary. Finally, the fitting procedure used three sensitivity parameters per participant, one for each of the three concurrent pairs (i.e., a12, a1e, and a2e), and one c2e per participant. As can be seen in Table 3, the fits were excellent. With few exceptions, the %VACs by the equations were above 90%.
Table 3. Least square fits of the modern version of Herrnstein's theory to the response rates for each participant. Equations 5, 6a′, 6b′ and 7′ were fitted simultaneously to the Schedule A and Schedule B data and all magnitudes for each participant. Shown are the values of each parameter—see the text for more details. The dashes indicate that the parameters listed in this row were held constant across magnitudes. The percentages of variance accounted for (%VAC) by each equation are provided in the right columns.
| Participant | Magnitude on Schedule A | k | a1e | a12 | a2e | c1e | c2e | b12 | %VAC |
|||
| Equation 5 | Equation 6a′ | Equation 6b′ | Equation 7′ | |||||||||
| H59 | – | 48.15 | 0.94 | 0.72 | 0.52 | 2.65 | ||||||
| $0.20 | 10.13 | 1.17 | 100 | 100 | 99 | 100 | ||||||
| $0.12 | 11.42 | 1.13 | 97 | 99 | 99 | 99 | ||||||
| $0.07 | 13.14 | 0.99 | 98 | 99 | 99 | 99 | ||||||
| $0.004 | 16.60 | 0.78 | 100 | 98 | 99 | 99 | ||||||
| $0.0025 | 20.56 | 0.64 | 99 | 99 | 98 | 99 | ||||||
| H60 | – | 59.52 | 0.68 | 0.61 | 0.56 | 1.13 | ||||||
| $0.20 | 1.14 | 1.46 | 99 | 99 | 99 | 99 | ||||||
| $0.12 | 1.48 | 1.14 | 100 | 100 | 99 | 100 | ||||||
| $0.07 | 1.74 | 1.05 | 99 | 100 | 99 | 99 | ||||||
| $0.004 | 3.35 | 0.50 | 100 | 100 | 100 | 99 | ||||||
| $0.0025 | 3.61 | 0.47 | 99 | 99 | 99 | 99 | ||||||
| H64 | – | 74.99 | 0.88 | 0.82 | 0.75 | 33.34 | ||||||
| $0.20 | 35.17 | 1.44 | 98 | 98 | 98 | 98 | ||||||
| $0.12 | 47.49 | 1.50 | 99 | 98 | 99 | 97 | ||||||
| $0.07 | 52.74 | 0.95 | 96 | 100 | 98 | 96 | ||||||
| $0.004 | 61.34 | 0.68 | 98 | 97 | 92 | 97 | ||||||
| $0.0025 | 74.20 | 0.69 | 98 | 99 | 97 | 98 | ||||||
| H67 | – | 72.63 | 0.40 | 0.47 | 0.53 | 0.49 | ||||||
| $0.20 | 0.24 | 1.29 | 98 | 98 | 98 | 98 | ||||||
| $0.12 | 0.24 | 1.26 | 95 | 97 | 94 | 94 | ||||||
| $0.07 | 0.28 | 1.08 | 99 | 99 | 99 | 99 | ||||||
| $0.004 | 0.38 | 0.77 | 96 | 96 | 97 | 97 | ||||||
| $0.0025 | 0.46 | 0.63 | 91 | 94 | 93 | 92 | ||||||
| H69 | – | 134.31 | 0.34 | 0.28 | 0.22 | 20.78 | ||||||
| $0.20 | 29.85 | 0.95 | 96 | 91 | 80 | 95 | ||||||
| $0.12 | 29.18 | 1.07 | 97 | 94 | 98 | 97 | ||||||
| $0.07 | 35.87 | 0.89 | 92 | 98 | 40 | 92 | ||||||
| $0.004 | 40.32 | 0.74 | 97 | 99 | 85 | 96 | ||||||
| $0.0025 | 34.48 | 0.96 | 98 | 89 | 69 | 97 | ||||||
| H70 | – | 104.48 | 0.48 | 0.39 | 0.32 | 1.28 | ||||||
| $0.20 | 1.30 | 1.62 | 91 | 96 | 96 | 92 | ||||||
| $0.12 | 1.67 | 1.40 | 98 | 99 | 98 | 98 | ||||||
| $0.07 | 2.07 | 1.16 | 98 | 97 | 99 | 98 | ||||||
| $0.004 | 4.61 | 0.46 | 93 | 94 | 86 | 94 | ||||||
| $0.0025 | 5.02 | 0.48 | 93 | 93 | 87 | 94 | ||||||
| Median | – | 73.81 | 0.58 | 0.54 | 0.52 | 1.97 | ||||||
| $0.20 | 5.71 | 1.37 | 98 | 98 | 98 | 98 | ||||||
| $0.12 | 6.55 | 1.20 | 98 | 99 | 99 | 98 | ||||||
| $0.07 | 7.61 | 1.02 | 98 | 99 | 99 | 98 | ||||||
| $0.004 | 10.61 | 0.71 | 97 | 98 | 94 | 97 | ||||||
| $0.0025 | 12.79 | 0.64 | 98 | 96 | 95 | 98 | ||||||
Figure 2 shows the standardized residuals plotted against the predicted response rates for Equations 6a′ and 6b′. Results are pooled across all participants. The correlations were not significant. Figure 3 shows the residual plots for Equation 5 and 7′. The correlations for both equations were not significant. In addition, none of the correlations for individual participants was significant with alpha set to .05.
Fig. 2. Standardized residuals as a function of the predicted response rates for Equations 6a′, Schedule A (top panel) and 6b′, Schedule B (bottom panel).

Pearson's correlation coefficients are shown in each panel.
Fig. 3. Standardized residuals as a function of the predicted response ratios for Equation 5 (top panel), and 7′ (bottom panel).

Pearson's correlation coefficients are shown in each panel.
Given that the fits and residuals were acceptable, we can now evaluate the parameters. The ks ranged from 48 responses per minute to 134 responses per minute, with a median of 74 responses per minute. The values for the sensitivity parameters, a12, a1e, and a2e, ranged from 0.22 to 0.94, with median values of 0.58, 0.54, and 0.52, respectively. The composite parameter c1e increased as magnitude decreased, from a median of 5.71 when $0.20 was the reinforcer on Schedule A, to 12.79 when $0.0025 was the reinforcer. The composite c2e parameter ranged from 0.49 to 33.34, with a median of 1.97. Finally, the bias parameter b12 decreased as the magnitude on Schedule A decreased, from a median of 1.37 when $0.20 was the reinforcer on Schedule A, to 0.64 when $0.0025 was the reinforcer.
Because joint fits of Equations 6a′ and 6b′ are probably unfamiliar, fits of these equations to the averaged reinforcement and response rates, across all participants, are shown in Figure 4. The left column shows the fits of Equation 6a′ to the Schedule A response rates, and the right column shows the fits of Equation 6b′ to the Schedule B response rates for three reinforcer magnitudes. Each plot shows how changes in reinforcement rate from two alternatives jointly govern response rate at one alternative of the concurrent schedule. Across plots, we also can see how changes in reinforcer magnitude affect responding. Only the curvature of the function changes with reinforcer magnitude.
Fig. 4. Relation between response rate at each alternative as a joint function of reinforcement rate on Schedule A (r1) and Schedule B (r2).

The smooth surface in each panel on the left represents the fit of Equation 6a′ to the data, and the smooth surface in each panel on the right represents the fit of Equation 6b′ to the data.
DISCUSSION
Six humans responded for points exchangeable for money on five concurrent VI VI schedules. Each concurrent schedule was presented during each daily session. Reinforcer magnitude on one alternative was varied systematically across sessions, while the magnitude at the other alternative remained constant. We tested the formal theory of matching by fitting Equations 4a and 4b to the data. One k was estimated per magnitude pair, as required by the theory. Formal theory requires a single k at each concurrent alternative because the ks in Equations 4a and 4b represent the same sum of response rates. One re per participant was estimated, which assumes that the rate of extraneous reinforcement remained constant across magnitude pairs. Although the equations accounted for a high percentage of variance in response rates, there was a significant negative correlation between the standardized residuals and the predicted response rates (Figure 1). The negative correlation indicates that the equations systematically under- and then over-predicted response rates as reinforcement rate increased. The results suggest that the formal theory of matching is not an adequate description of reinforced responding.
One may argue that our application of formal theory was too conservative because we used one re per participant. It may be tempting to assume that re must change across magnitude pairs, as the units for re are changing from magnitude pair to magnitude pair. However, in formal theory re is not a measure of amount per unit time; it is a measure of counts per unit time (McDowell, 1986, 2005). Furthermore, the re in both equations represents the same quantity. Just as k for each equation represents the same sum of response rates, re for each equation represents the same sum of reinforcement rates. Similarly, because magnitude differed between the two schedules (i.e., Schedule A and B), one may assume that the units for r1, r2, and re should be different, which would produce dimensional inconsistencies for each equation. However, all of the variables in the denominators are measured in terms of counts per unit time. Therefore, there are no dimensional inconsistencies in the denominators of Equations 4a and 4b.
Although it is reasonable to suppose that extraneous reinforcement rate should remain constant, formal theory does not prohibit variation in re. The rate of extraneous reinforcing events may vary across magnitude pairs, but the theory does not predict how or if re should vary with magnitude. In other words, variation in re could occur, and the variation could occur in any direction. To test formal theory with this more lenient constraint, we fitted Equations 4a and 4b and allowed one k and one re per magnitude pair, which resulted in five ks and five res per participant. The results were very similar to those discussed above. Specifically, although the %VAC by the equations remained high, the residuals were correlated with the predicted response rate for Equation 4a (r = −.45) and Equation 4b (r = −.57). Both were statistically significant with alpha set at .05. Thus, regardless of the constraints on re, the theory does not account for the data.
For theoretical reasons, no further restrictions can be lifted on whether k or re varies with reinforcer magnitude. It would be possible, however, to estimate two ks per magnitude pair, and then examine whether the two values were equal. This is not a more lenient test; it is merely a different way to test the same theoretical restriction that the ks obtained at each alternative of a concurrent schedule must be equal. The drawback to this method is that, in the absence of large differences between the ks, the results are more difficult to interpret relative to the simultaneous fitting procedure. For example, assume we measure two ks, and both have a standard error of five responses per minute. At what point would we consider the ks “equal”: when they differ by 10, 5, or only 2.5 responses per minute? The current method is simply a more sensitive and robust test of the equal-k requirement. The negative correlation in the residuals leaves little doubt that the formal model does not describe the data.
The modern version of matching provided an excellent description of reinforced responding. The median %VAC by the four modern matching equations ranged from 95% to 99%. The residual plots appeared random (Figures 2 and 3), and none of the correlations between the standardized residuals and the predicted response rates was statistically significant. The fits of Equations 6a′ and 6b′ shown in Figure 4 imply that changes in response rate on each alternative of a concurrent schedule can be accounted for by changes in reinforcement rate and bias for one or the other alternative, and that behavior was somewhat insensitive to changes in reinforcement rate.
In addition, the fits were consistent with theoretical predictions and requirements. For example, because the development of modern theory followed Herrnstein's (1970, 1974; de Villiers & Herrnstein, 1976) assumption that total behavior should remain constant, k was held constant. With just one k, the fits were excellent. McDowell (2005) also found that the modern version of matching accurately described responding on single and concurrent schedules of reinforcement in rats, and that it could account for the violation of classic theory's constant-k requirement. Furthermore, because reinforcer magnitude varied on Schedule A, bias for this alternative should change across magnitude pairs (Baum, 1974, 1979). This meant that b12 and c1e were allowed to vary across magnitude pairs. As expected, b12 decreased as the magnitude on Schedule A decreased. Similarly, c1e increased as magnitude decreased, which is reasonable given that the denominator—bias for the Schedule A alternative—should have decreased as magnitude on that alternative decreased. The numerator of c1e, the rate of extraneous reinforcement, rea1e, was assumed to remain constant. The other composite parameter, c2e, also consists of a bias parameter, b2e, and an extraneous reinforcement parameter, re. Bias should not have changed between Schedule B and extraneous alternatives, as the magnitude on Schedule B was always $0.07, and we assumed that extraneous reinforcement remained constant. Therefore, c2e was held constant. Of course, we cannot determine whether the numerator or denominator changed in either composite parameter, but their values were consistent with these assumptions.
The assertion that re should have remained constant, despite changes in reinforcer magnitude, may still come as a surprise. The idea that re represents counts per unit time was introduced in the context of formal theory, as it is a requirement of formal theory. In the context of modern theory, re also represents counts per unit time. Here, the bias parameter accommodates any change in reinforcer magnitude (or changes in response properties for that matter). This is consistent with the foundational equation for modern theory, the generalized matching equation. If different reinforcer magnitudes are arranged on two concurrent schedules, then b in Equation 5 accounts for the difference in magnitude. Therefore, to be consistent with common usage in matching theory, the same would hold if the constituents of the concurrent pair were a programmed and an unprogrammed (extraneous) alternative (McDowell, 2005). The units for both reinforcer events would be counts per unit time, and any change in magnitude would be accommodated by changes in bias, b. Therefore, re was assumed to remain constant across magnitude pairs (see Soto et al., 2005, for further discussion of the constant-re assumption).
Finally, the fitting procedure also used three sensitivity parameters per participant, one for each of the three concurrent pairs (i.e., a12, a1e, and a2e). The decision to use one a per concurrent pair, rather than letting them vary with magnitude, was based on parsimony and the lack of evidence that sensitivity to reinforcement rate should change with magnitude (Baum, 1979; Landon, Davison, & Elliffe, 2003). The obtained values appear to be consistent with existing data (e.g., Baum, 1979), although they are somewhat lower than expected. In addition, the sensitivity parameters for Participant H69 are quite low, which reflects this subject's relatively constant response rate despite large changes in reinforcement rate (H69's mean response rate was 12 responses per minute, with a range of 5 to 20; see Appendix B).
In a recent test of modern matching theory, McDowell (2005) used one a for all concurrent pairs. This is a more conservative approach than the one taken here, which assumed that sensitivity varied depending on the particular alternatives that comprised each concurrent pair. Arguably, a human may discriminate better between a programmed and an unprogrammed source of reinforcement (i.e., a1e and a2e) compared to two programmed sources of reinforcement (i.e., a12). In the former case, the responses (pressing a lever versus engaging in any other behavior) and reinforcers are qualitatively different, whereas in the latter case, the responses are identical (pressing the same lever) and the reinforcers are similar (different amounts of money). Although the highest as were obtained consistently for a1e, there was very little difference between a12 and a2e. Indeed, the median as for each concurrent pair suggest that the as were roughly equivalent regardless of the constituents of the concurrent pair (Table 3). Such data lend some support to the notion that the source of reinforcement may not affect the organism's sensitivity to reinforcement rates derived from these sources, which accords with McDowell's single-a approach.
Other features of the present results are consistent with McDowell's (2005) test of modern matching theory's description of responding under concurrent schedules. McDowell fitted Equations 5, 6a′, and 6b′ to Dallery et al's (2004) data from 4 rats responding on concurrent VI VI schedules. A 75% sucrose pellet was used as the reinforcer in one component and a 50% sucrose pellet was used as the reinforcer in the other component. The results of the reanalysis by McDowell were consistent with the results of the current study in that the median %VAC by the equations was 96.5%, and the standardized residuals were not correlated with the predicted response rates. Furthermore, the estimates of the bias parameter, b12, were 1.05, 1.08, 1.05, and 0.88, which indicates a slight preference for the 75% sucrose pellet over the 50% pellet for 3 of the 4 rats. Finally, the estimates of the single exponent, a, were 0.52, 0.64, 0.80, and 0.76.
Previous tests of matching theory's description of single VI responding were tests of classic matching theory (e.g., Bradshaw et al., 1978; Dallery et al., 2000; de Villiers, 1977; Heyman & Monaghan, 1987, 1994; McDowell & Dallery, 1999; McDowell & Wood, 1984, 1985; Snyderman, 1983; see Dallery & Soto, 2004, and Williams, 1988, for reviews). That is, these studies used a hyperbolic equation, Equation 2, which did not include parameters for bias and sensitivity, and they tested the assumption that k remains constant across reinforcer magnitude. Because k varied with magnitude, several authors suggested that total behavior does not remain constant, and therefore that Herrnstein's classic theory of response rate is false. In contrast, the present results suggest that modern theory's constant-k requirement is tenable. After incorporating bias and sensitivity into the relevant equations, using a single k across magnitude pairs provided an excellent description of the relation between reinforcement and responding. The modern theory of matching may represent a theoretically coherent and accurate description of responding on concurrent and single-alternative schedules of reinforcement.
Acknowledgments
We thank Leann Boni for considerable help collecting and analyzing the data, and Matthew Locey and Bethany Raiff for editorial comments. This research was supported by NSF Grant BNS-8908921 to J. J McDowell.
APPENDIX A
Instructions Given to Participants During the Orientation Session
During a session, this green light will turn on. Also, one of these white lights and one of these red lights will turn on. While the green light, one of the white lights, one of the red lights, and the small lamp-light behind the machine are on, you can earn money by pressing the lever.
The lever must be pushed all the way down and then let all the way back up in order for it to count. You can tell that you have pushed far enough when you hear a click from the machine. Remember, the lever has to go all the way down and all the way back up again. This is not a test of strength or speed. However, the amount of money that you earn does depend on your pressing this lever.
This counter will keep track of the money that you have earned. The amount that you earn will be shown here in dollars. Sometimes when you press the lever, a beep will sound and this amber light will flash. When you hear the beep and see the amber light flash, you have earned more money. Each time you earn more money, it will be added to the previous amount, and will be shown on the counter.
These lights will be on for ten minutes (the green, white, and red lights, and the light behind the machine). During that time you can press the lever and earn money. After ten minutes, all lights except the small green one will turn off for five minutes. During this time you are to rest here in the room. You cannot earn money during the rest. After the five-minute rest, the light behind the machine, a white light, and a red light will turn on for ten minutes, and you can again press to earn money. These ten-minute work periods followed by five-minute rest periods will continue for the entire session, which is about 70 minutes long. You will have three of these 70-minute sessions each day, with a ten-minute break between each. At the end of each 70-minute session, the green, white, and red lights will all turn off. Then I will come in and record the amount of money that you earned.
At any time during a work period, you can press this button and switch to work with a different combination of red and white lights. Pressing this button will turn on the small blue light above the button. Most of the time, it will also turn on a different white light, or a different white light and a different red light. Pressing the button again will turn off the light beside the button, and will return you to the previous combination of lights.
Before you leave for the day, I will give you a receipt for your earnings. You will receive payment according to the terms of the contract that you sign. On average, you can earn up to $6.50 per hour, although your exact pay will depend on your performance pressing the lever. This is an average over the entire project. Some days you will earn more money than others. However, at the conclusion of the project the average amount that you earn could be as much as $6.50 per hour.
APPENDIX B
Averaged reinforcement (r, measured in reinforcers per hour) and response (R, measured in responses per minute) rates over each stable eight-session block for all schedules and participants.
| Participant | VI–VI | $0.20 |
$0.12 |
$0.07 |
$0.004 |
$0.00250 |
|||||||||||||||
| Schedule A |
Schedule B |
Schedule A |
Schedule B |
Schedule A |
Schedule B |
Schedule A |
Schedule B |
Schedule A |
Schedule B |
||||||||||||
| r | R | r | R | r | R | R | R | r | R | r | R | r | R | r | R | r | R | r | R | ||
| H59 | 17–720 | 188.3 | 43.2 | 4.5 | 2.5 | 177.0 | 40.2 | 3.0 | 3.3 | 180.0 | 39.9 | 3.0 | 3.1 | 158.3 | 37.0 | 9.8 | 6.1 | 159.0 | 35.3 | 7.5 | 7.6 |
| 25–300 | 124.5 | 39.2 | 9.8 | 5.2 | 135.0 | 39.2 | 12.8 | 4.7 | 129.0 | 37.9 | 13.5 | 5.5 | 110.3 | 31.6 | 12.8 | 8.3 | 109.5 | 32.8 | 12.0 | 8.8 | |
| 51–51 | 51.0 | 23.8 | 45.0 | 17.4 | 54.0 | 24.5 | 40.5 | 16.2 | 54.0 | 21.7 | 51.8 | 19.7 | 53.3 | 16.2 | 39.0 | 18.8 | 55.5 | 17.2 | 53.3 | 26.0 | |
| 300–25 | 11.3 | 7.6 | 123.0 | 36.5 | 15.8 | 9.8 | 103.5 | 31.5 | 13.5 | 7.3 | 102.8 | 32.7 | 10.5 | 4.5 | 124.5 | 34.3 | 16.5 | 5.0 | 119.3 | 32.9 | |
| 720–17 | 10.5 | 6.7 | 156.0 | 38.2 | 9.0 | 4.7 | 172.5 | 38.5 | 11.3 | 4.5 | 172.5 | 39.2 | 12.0 | 5.5 | 142.5 | 36.0 | 11.3 | 2.7 | 180.0 | 35.8 | |
| H60 | 17–720 | 183.8 | 55.5 | 1.5 | 2.0 | 189.8 | 55.3 | 2.3 | 2.9 | 182.3 | 54.4 | 0.8 | 2.3 | 133.5 | 40.8 | 6.8 | 14.2 | 150.0 | 42.1 | 7.5 | 14.8 |
| 25–300 | 137.3 | 54.2 | 1.5 | 2.0 | 117.0 | 49.0 | 7.5 | 7.8 | 126.0 | 48.5 | 5.3 | 7.3 | 105.8 | 34.3 | 12.8 | 20.2 | 81.0 | 30.0 | 15.0 | 25.0 | |
| 51–51 | 63.8 | 39.6 | 35.3 | 15.7 | 51.0 | 28.4 | 48.8 | 26.9 | 56.3 | 33.1 | 43.5 | 22.2 | 28.5 | 13.5 | 63.0 | 41.3 | 30.8 | 13.6 | 66.0 | 41.5 | |
| 300–25 | 11.3 | 14.4 | 116.3 | 41.3 | 21.8 | 17.2 | 102.8 | 39.3 | 16.5 | 13.8 | 120.8 | 42.6 | 9.0 | 5.7 | 131.3 | 50.3 | 12.0 | 7.1 | 125.3 | 49.1 | |
| 720–17 | 12.0 | 10.1 | 167.3 | 46.2 | 9.8 | 9.9 | 141.8 | 42.5 | 6.0 | 4.7 | 187.5 | 52.6 | 7.5 | 3.8 | 186.8 | 52.9 | 6.0 | 2.4 | 195.8 | 55.6 | |
| H64 | 17–720 | 174.0 | 54.6 | 6.0 | 2.4 | 151.5 | 44.5 | 1.5 | 0.6 | 138.0 | 43.7 | 3.0 | 0.6 | 138.8 | 43.5 | 10.5 | 9.8 | 102.0 | 28.8 | 9.0 | 6.5 |
| 25–300 | 117.8 | 46.5 | 2.3 | 2.0 | 84.8 | 35.0 | 5.3 | 2.0 | 94.5 | 38.0 | 3.0 | 1.8 | 50.3 | 20.0 | 9.8 | 7.9 | 74.3 | 28.0 | 9.0 | 4.8 | |
| 51–51 | 45.0 | 26.9 | 29.3 | 13.9 | 52.5 | 28.7 | 21.0 | 9.9 | 44.3 | 19.4 | 24.0 | 12.0 | 7.5 | 3.1 | 44.3 | 26.4 | 7.5 | 3.6 | 50.3 | 23.1 | |
| 300–25 | 12.8 | 13.4 | 93.0 | 34.8 | 6.8 | 4.3 | 98.3 | 36.5 | 5.3 | 1.7 | 106.5 | 40.8 | 2.3 | 1.9 | 90.0 | 34.6 | 6.8 | 3.3 | 88.5 | 35.5 | |
| 720–17 | 6.0 | 6.5 | 144.8 | 42.7 | 6.0 | 5.8 | 134.3 | 38.5 | 1.5 | 0.6 | 159.8 | 45.0 | 1.5 | 0.8 | 129.0 | 33.6 | 0.8 | 0.2 | 125.3 | 35.8 | |
| H67 | 17–720 | 167.3 | 56.4 | 10.5 | 13.4 | 163.5 | 55.1 | 11.3 | 15.7 | 166.5 | 57.9 | 8.3 | 15.5 | 149.3 | 49.9 | 11.3 | 19.1 | 152.3 | 49.0 | 10.5 | 21.7 |
| 25–300 | 123.0 | 55.4 | 15.8 | 16.1 | 121.5 | 56.2 | 16.5 | 16.2 | 117.8 | 54.8 | 17.3 | 17.9 | 103.5 | 45.4 | 16.5 | 20.8 | 110.3 | 44.8 | 14.3 | 22.5 | |
| 51–51 | 67.5 | 45.6 | 54.0 | 24.7 | 63.0 | 44.4 | 49.5 | 22.9 | 64.5 | 38.1 | 59.3 | 33.2 | 54.8 | 23.7 | 61.5 | 44.6 | 50.3 | 18.0 | 66.8 | 53.6 | |
| 300–25 | 12.8 | 20.5 | 111.0 | 50.3 | 17.3 | 19.4 | 111.0 | 51.0 | 10.5 | 18.1 | 115.5 | 53.4 | 14.3 | 16.8 | 118.5 | 53.3 | 15.8 | 16.2 | 114.0 | 55.2 | |
| 720–17 | 8.3 | 16.7 | 152.3 | 55.1 | 9.8 | 18.9 | 155.3 | 51.1 | 9.0 | 15.1 | 168.0 | 57.9 | 13.5 | 15.8 | 163.5 | 55.7 | 11.3 | 14.2 | 164.3 | 56.4 | |
| H69 | 17–720 | 100.5 | 17.4 | 9.0 | 8.5 | 117.0 | 17.5 | 9.0 | 8.4 | 105.8 | 15.5 | 9.0 | 10.2 | 92.3 | 13.0 | 10.5 | 8.4 | 108.8 | 16.7 | 8.3 | 8.3 |
| 25–300 | 78.0 | 16.1 | 18.0 | 10.9 | 84.0 | 16.4 | 17.3 | 10.3 | 81.8 | 13.3 | 16.5 | 8.8 | 78.0 | 12.4 | 14.3 | 9.5 | 80.3 | 13.6 | 15.0 | 8.8 | |
| 51–51 | 42.0 | 12.1 | 43.5 | 12.6 | 48.0 | 14.3 | 45.8 | 12.9 | 42.0 | 10.1 | 51.8 | 12.9 | 37.5 | 9.7 | 50.3 | 13.1 | 37.5 | 9.5 | 52.5 | 12.8 | |
| 300–25 | 18.0 | 11.8 | 70.5 | 16.0 | 17.3 | 9.2 | 77.3 | 13.9 | 10.5 | 6.7 | 76.5 | 15.5 | 18.0 | 6.9 | 84.0 | 15.1 | 16.5 | 6.7 | 84.8 | 11.1 | |
| 720–17 | 9.8 | 7.2 | 107.3 | 17.9 | 7.5 | 9.1 | 109.5 | 15.6 | 9.0 | 6.7 | 105.0 | 12.0 | 8.3 | 5.4 | 131.3 | 19.6 | 9.0 | 8.0 | 114.0 | 16.5 | |
| H70 | 17–720 | 180.0 | 90.9 | 5.3 | 6.5 | 163.5 | 75.5 | 12.8 | 16.1 | 156.8 | 70.5 | 9.8 | 18.0 | 87.8 | 41.2 | 12.0 | 46.8 | 81.8 | 35.7 | 11.3 | 46.5 |
| 25–300 | 120.8 | 75.9 | 11.3 | 18.3 | 120.0 | 69.4 | 12.8 | 19.2 | 120.0 | 63.8 | 10.5 | 23.4 | 59.3 | 33.5 | 12.8 | 48.6 | 84.0 | 41.2 | 13.5 | 36.2 | |
| 51–51 | 56.3 | 54.9 | 45.0 | 35.1 | 57.0 | 54.9 | 40.5 | 34.7 | 39.0 | 43.6 | 42.8 | 41.7 | 29.3 | 27.4 | 56.3 | 54.0 | 37.5 | 29.7 | 56.3 | 49.6 | |
| 300–25 | 17.3 | 42.1 | 88.5 | 50.3 | 14.3 | 38.1 | 70.5 | 49.0 | 17.3 | 28.3 | 101.3 | 53.4 | 6.0 | 12.3 | 118.5 | 75.1 | 8.3 | 10.0 | 132.8 | 73.9 | |
| 720–17 | 8.3 | 28.6 | 143.3 | 63.7 | 8.3 | 23.2 | 157.5 | 65.4 | 7.5 | 25.7 | 147.8 | 61.6 | 3.8 | 9.3 | 170.3 | 80.4 | 7.5 | 9.3 | 168.0 | 77.2 | |
REFERENCES
- Baum W.M. On two types of deviation from the matching law: Bias and undermatching. Journal of the Experimental Analysis of Behavior. 1974;22:231–242. doi: 10.1901/jeab.1974.22-231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baum W.M. Matching, undermatching, and overmatching in studies of choice. Journal of the Experimental Analysis of Behavior. 1979;32:269–281. doi: 10.1901/jeab.1979.32-269. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradshaw C.M, Szabadi E, Bevan P. Behavior of humans in variable-interval schedules of reinforcement. Journal of the Experimental Analysis of Behavior. 1976;26:135–141. doi: 10.1901/jeab.1976.26-135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bradshaw C.M, Szabadi E, Bevan P. Relationship between response rate and reinforcement frequency in variable-interval schedules: The effect of concentration of sucrose reinforcement. Journal of the Experimental Analysis of Behavior. 1978;29:447–452. doi: 10.1901/jeab.1978.29-447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dallery J, McDowell J.J, Lancaster J.S. Falsification of matching theory's account of single-alternative responding: Herrnstein's k varies with sucrose concentration. Journal of the Experimental Analysis of Behavior. 2000;73:23–43. doi: 10.1901/jeab.2000.73-23. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dallery J, McDowell J.J, Soto P.L. The measurement and functional properties of reinforcer value in single-alternative responding: A test of linear system theory. The Psychological Record. 2004;54:45–65. [Google Scholar]
- Dallery J, Soto P.L. Herrnstein's hyperbola and behavioral pharmacology: Review and critique. Behavioural Pharmacology. 2004;15:443–459. doi: 10.1097/00008877-200411000-00001. [DOI] [PubMed] [Google Scholar]
- de Villiers P. Choice in concurrent schedules and a quantitative formulation of the law of effect. In: Honig W.K, Staddon J.E.R, editors. Handbook of operant behavior. Englewood Cliffs, NJ: Prentice Hall; 1977. pp. 233–287. [Google Scholar]
- de Villiers P.A, Herrnstein R.J. Toward a law of response strength. Psychological Bulletin. 1976;83:1131–1153. [Google Scholar]
- Findley J.D. Preference and switching under concurrent scheduling. Journal of the Experimental Analysis of Behavior. 1958;1:123–144. doi: 10.1901/jeab.1958.1-123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fleshler M, Hoffman H.S. A progression for generating variable interval schedules. Journal of the Experimental Analysis of Behavior. 1962;5:529–530. doi: 10.1901/jeab.1962.5-529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrnstein R.J. Relative and absolute strength of response as a function of frequency of reinforcement. Journal of the Experimental Analysis of Behavior. 1961;4:267–272. doi: 10.1901/jeab.1961.4-267. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrnstein R.J. On the law of effect. Journal of the Experimental Analysis of Behavior. 1970;13:243–266. doi: 10.1901/jeab.1970.13-243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herrnstein R.J. Formal properties of the matching law. Journal of the Experimental Analysis of Behavior. 1974;21:159–164. doi: 10.1901/jeab.1974.21-159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heyman G.M, Monaghan M.M. Effects of changes in response requirement and deprivation on the parameters of the matching law equation: New data and review. Journal of Experimental Psychology: Animal Behavior Processes. 1987;13:384–394. [Google Scholar]
- Heyman G.M, Monaghan M.M. Reinforcer magnitude (sucrose concentration) and the matching law theory of response strength. Journal of the Experimental Analysis of Behavior. 1994;61:505–516. doi: 10.1901/jeab.1994.61-505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Landon J, Davison M, Elliffe D. Concurrent schedules: Reinforcer magnitude effects. Journal of the Experimental Analysis of Behavior. 2003;79:351–365. doi: 10.1901/jeab.2003.79-351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDowell J.J. On the falsifiability of matching theory. Journal of the Experimental Analysis of Behavior. 1986;45:63–74. doi: 10.1901/jeab.1986.45-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDowell J.J. On the classic and modern theories of matching. Journal of the Experimental Analysis of Behavior. 2005;84:111–127. doi: 10.1901/jeab.2005.59-04. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDowell J.J, Dallery J. Falsification of matching theory: Changes in the asymptote of Herrnstein's hyperbola as a function of water deprivation. Journal of the Experimental Analysis of Behavior. 1999;72:251–268. doi: 10.1901/jeab.1999.72-251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDowell J.J, Wood H.M. Confirmation of linear system theory prediction: Changes in Herrnstein's k as a function of changes in reinforcer magnitude. Journal of the Experimental Analysis of Behavior. 1984;41:183–192. doi: 10.1901/jeab.1984.41-183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDowell J.J, Wood H.M. Confirmation of linear system theory prediction: Rate of change of Herrnstein's k as a function of response-force requirement. Journal of the Experimental Analysis of Behavior. 1985;43:61–73. doi: 10.1901/jeab.1985.43-61. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Skinner B.F. Are theories of learning necessary? In: Skinner B.F, editor. Cumulative Record: Third Edition. New York: Appleton Century Crofts; 1972. (Reprinted from Psychological Review, 57, 1950, 193–216) [Google Scholar]
- Snyderman M. Body weight and response strength. Behavior Analysis Letters. 1983;3:255–265. [Google Scholar]
- Soto P.L, McDowell J.J, Dallery J. Effects of adding a second reinforcement alternative: Implications for Herrnstein's interpretation of re. Journal of the Experimental Analysis of Behavior. 2005;84:185–225. doi: 10.1901/jeab.2005.09-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tryon W.W. A simplified time-series analysis for evaluating treatment interventions. Journal of Applied Behavior Analysis. 1982;15:423–429. doi: 10.1901/jaba.1982.15-423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Warren-Boulton F.R, Silberberg A, Gray M, Ollom R. Reanalysis of the equation for simple action. Journal of the Experimental Analysis of Behavior. 1985;43:265–277. doi: 10.1901/jeab.1985.43-265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams B.A. Reinforcement, choice, and response strength. In: Atkinson R.C, Herrnstein R.J, Lindzey G, Luce R.D, editors. Stevens' Handbook of Experimental Psychology. New York: Wiley; 1988. (2nd ed., Vol. 2, pp. 167–244) [Google Scholar]