

Abstract
Recombinant adeno-associated virus (rAAV) vectors are promising vehicles for achieving stable liver transduction in vivo. However, the mechanisms of liver transduction are not fully understood, and furthermore, the relationships between rAAV dose and levels of transgene expression, total number of hepatocytes transduced, and proportion of integrated vector genomes have not been well established. To begin to elucidate the liver transduction dose response with rAAV vectors, we injected mice with two different human factor IX or Escherichia coli lacZ-expressing AAV serotype 2-based vectors at doses ranging between 4.0 × 108 and 1.1 × 1013 vector genomes (vg)/mouse, in three- to sixfold increments. A 2-log-range linear dose-response curve of transgene expression was obtained from 3.7 × 109 to 3.0 × 1011 vg/mouse. Vector doses above 3.0 × 1011 vg/mouse resulted in disproportionately smaller increases in both the number of transduced hepatocytes and levels of transgene expression, followed by saturation at doses above 1.8 × 1012 vg/mouse. In contrast, a linear increase in the number of vector genomes per hepatocyte was observed up to 1.8 × 1012 vg/mouse concomitantly with enhanced vector genome concatemerization, while the proportion of integrated vector genomes was independent of the vector dose. Thus, the mechanisms that restrict a wide-range linear dose response at high doses likely involve decreased functionality of vector genomes and restriction of transduction to fewer than 10% of total hepatocytes. Such information may be useful to determine appropriate vector doses for in vivo administration and provides further insights into the mechanisms of rAAV transduction in the liver.
Adeno-associated virus (AAV) is a small, nonpathogenic replication-defective human parvovirus with a single-stranded (ss) DNA genome of approximately 4.7 kb. Recombinant viruses based on AAV serotype 2 are promising gene therapy vectors that deliver genes of interest into a variety of somatic tissues at therapeutic levels by direct administration into experimental animals. Recombinant AAV (rAAV) vectors are currently in use in several clinical trials of gene transfer for the treatment of cystic fibrosis, hemophilia B, and limb girdle muscular dystrophy (1, 13, 16, 32) and have been shown to be safe in preliminary trial reports (1, 2, 13, 16, 32).
Intraportal or intravenous injection of rAAV vectors into experimental animals results in stable transduction of hepatocytes with therapeutic levels of transgene expression in the circulation and/or the target tissue (3, 4, 8, 14, 16, 22, 29-31, 33, 34). Some of the mechanisms underlying stable hepatocyte transduction with rAAV have been elucidated, and these include the following: (i) rAAV vector genomes are found in most hepatocyte nuclei within 24 h after vector infusion, but only a small subset of these hepatocytes establish stable transduction (19); (ii) transduced hepatocytes represent a subset of cells, perhaps with a specific metabolic state (19); (iii) during the first 6 weeks after vector administration, ss rAAV vector genomes convert into a variety of different double-stranded (ds) forms including supercoiled ds circular monomers, ds linear monomers, circular and linear concatemers, and integrated proviral forms (21, 23, 26, 31); (iv) vector genome concatemerization occurs through intermolecular recombination of ds linear monomer intermediates, not through recombination between circular genomes or through self-amplification (25; H. Nakai et al., unpublished results); (v) ds linear monomer genomes appear to be the intermediate form most responsible for host chromosomal integration events (Nakai et al., unpublished results); and (vi) the majority of stable vector genomes exist as extrachromosomal forms, which are primarily responsible for stable hepatocyte transduction (26).
At doses of ∼1011 vector genomes (vg) per mouse, therapeutic levels of transgene expression were achieved in most of the experimental settings with various murine disease models (8, 14, 29-31, 34). However, the proportion of hepatocytes stably transduced at these doses is only about 5% (6, 24, 30, 31). Given that stable transduction is restricted to a small subpopulation of hepatocytes with many vector genomes (21, 30), even though all hepatocytes are initially infected (19), the relationship between vector dose and transgene expression in hepatocytes in vivo might not be straightforward. Whether the number of transduced hepatocytes and/or transgene expression, or the proportion of integrated rAAV vector genomes, increases in proportion to dose escalation has not been carefully examined.
The aim of the present study was to thoroughly characterize vector dose response in rAAV transduction in the liver and elucidate the underlying molecular mechanisms. By injecting mice with different doses of a liver-specific promoter-driven human coagulation factor IX (hF.IX) rAAV vector, a ubiquitous endogenous promoter-driven hF.IX vector, and a β-galactosidase (β-Gal)-expressing marker vector, we find a 2-log range of linearity between vector doses and levels of transgene expression, a good correlation between numbers of transduced hepatocytes and levels of transgene expression, and differential saturation of vector genomes and transgene expression at very high doses. These results may have important implications for defining effective vector doses for clinical applications and understanding the mechanisms of rAAV transduction in the liver.
MATERIALS AND METHODS
Construction of rAAV vectors.
All the rAAV vectors used in the present study were AAV serotype 2-based vectors. Construction and production of AAV-EF1α-F.IX and AAV-EF1α-nlsLacZ were described elsewhere (22, 24). AAV-hF.IX16, which is now being used for a human clinical trial, is the same as AAV-CM1 (26), except that AAV-hF.IX16 was produced with a vector plasmid with a different plasmid backbone sequence. Briefly, AAV-hF.IX16 carried a liver-specific promoter (the apolipoprotein E hepatic locus control region-human α1-antitrypsin gene promoter [20]), the hF.IX minigene (containing a 1.4-kb DNA fragment of the first intron from the hF.IX gene), and the bovine growth hormone polyadenylation signal [poly(A)]; AAV-EF1α-F.IX had the human elongation factor 1α (EF1α) enhancer-promoter, hF.IX cDNA, and the human growth hormone poly(A); and AAV-EF1α-nlsLacZ harbored the EF1α enhancer-promoter, the bacterial lacZ gene with a nuclear localizing signal, and the simian virus 40 poly(A). All the vectors were produced by the triple transfection method (18). AAV-EF1α-F.IX and AAV-EF1α-nlsLacZ were purified with a cesium chloride gradient as previously described (3), while AAV-hF.IX16 was purified by heparin column chromatography in the vector core facility at Avigen, Inc. The physical particle titers were determined by a quantitative dot blot assay (17).
Animal procedure.
Six- to eight-week-old female C57BL/6 and male C57BL/6 rag-1 mice were purchased from the Jackson Laboratory (Bar Harbor, Maine). The portal vein injection and surgical two-thirds partial hepatectomy were performed as previously described (15, 22). Blood samples were collected from the retro-orbital plexus. All the animal experiments were performed according to the guidelines for animal care at Stanford University.
Protein analysis.
hF.IX levels in mouse plasma were determined by an enzyme-linked immunosorbent assay (ELISA) specific for hF.IX (22). The detection limit of the ELISA was 2 ng/ml. Total liver proteins were extracted as previously described, and expression levels of β-Gal in rAAV-transduced mouse livers were determined with a β-Gal ELISA kit (Roche Molecular Biochemicals, Indianapolis, Ind.) (24). The β-Gal levels were normalized with the total protein concentration determined by the Lowry assay with a DC protein assay kit (Bio-Rad, Hercules, Calif.) and shown as picograms of β-Gal per milligram of protein.
Histological analysis.
Pieces of mouse liver lobes were embedded in Tissue-Tek Optimal Cutting Temperature compound (Sakura Finetek USA, Inc., Torrance, Calif.) and frozen on dry ice. Ten-micrometer-thick sections were cut, fixed with 1.25% glutaraldehyde, stained with 5-bromo-4-chloro-3-indolylphosphate (X-Gal) as described previously (15), and counterstained with light hematoxylin. To determine transduction efficiency, at least 2,000 nuclei per section were examined for β-Gal expression from each animal.
DNA analysis.
Extraction of total genomic DNA from mouse livers, Southern blot analysis, and densitometric analysis of the blots were performed as previously described (23), to determine vector genome number per diploid genomic equivalent (vg/dge) and analyze vector forms in transduced mouse livers. The vector genome copy number standards were prepared by adding an equivalent number of corresponding plasmid molecules to 20 μg of total DNA extracted from naive mouse liver. “Net” vector copy number per hepatocyte represents the number of vector genomes per transduced hepatocyte and is calculated based on a presumption that vector genomes in the liver are carried only by transduced hepatocytes (21, 30).
RESULTS
A linear vector dose response of transgene expression at doses up to 3.0 × 1011 vg/mouse.
To investigate the relationship between vector dose and transgene expression in rAAV transduction in mouse livers, AAV-hF.IX16 vector was administered via the portal vein at nine different doses ranging from 4.0 × 108 to 1.1 × 1013 vg/mouse, increasing in three- to sixfold increments, i.e., 4.0 × 108, 1.2 × 109, 3.7 × 109, 1.1 × 1010, 3.3 × 1010, 1.0 × 1011, 3.0 × 1011, 1.8 × 1012, and 1.1 × 1013 vg/mouse (n = 5 per dose, except for the highest dose [n = 4]). Plasma hF.IX levels were undetectable by ELISA over a 10-week period at doses below 3.7 × 109 vg/mouse. Above this dose and up to 3.0 × 1011 vg/mouse, log/log plotting of vector doses and hF.IX levels exhibited linearity with regression coefficients (r) of 0.996 (2 weeks postinjection) and 0.998 (10 weeks postinjection) (Fig. 1). Since the slopes (log y/log x) of the log/log fit vector dose-response curves between 3.7 × 109 and 3.0 × 1011 vg/mouse were close to 1.0, i.e., 1.159 (2 weeks postinjection) and 1.127 (10 weeks postinjection), hF.IX levels were linearly related to given vector doses within this dose range.
FIG. 1.

Plasma hF.IX levels after intraportal administration of various doses of AAV-hF.IX16 into C57BL/6 mice. Expression levels 2 and 10 weeks postinjection are shown as a function of given vector doses. The linear range of the vector dose response is indicated with brackets. Values are means ± standard errors.
Nonproportional rAAV vector-dose response at doses of greater than 3.0 × 1011 vg/mouse.
In the mice injected with AAV-hF.IX16, a small increase in hF.IX levels was observed between 3 × 1011 and 1.8 × 1012 vg/mouse, but this was not proportional to the sixfold difference between doses (Fig. 1 and Table 1). In addition, at doses above 1.8 × 1012 vg/mouse, no significant increase was observed for plasma hF.IX levels, which reached a plateau level at approximately 40 μg/ml (Table 1). To confirm these results, a second dose-response study was performed with an independent hF.IX vector, AAV-EF1α-F.IX. In addition, in order to be able to evaluate both the number of transduced hepatocytes as a function of dose and the absolute amount of transgene product, the AAV-EF1α-nlsLacZ vector was used. The dose ranges were between 5.0 × 1010 and 7.2 × 1012 and between 5.0 × 1010 and 3.9 × 1012 vg/mouse for the AAV-EF1α-F.IX and AAV-EF1α-nlsLacZ vectors, respectively. The results are summarized in Tables 2 and 3.
TABLE 1.
Liver transduction with AAV-hF.IX16 (14 weeks postinjection)a
| Dose (vg/mouse) | No. of mice | Pre-PHxb
|
Post-PHx
|
||
|---|---|---|---|---|---|
| hF.IX (ng/ml) | Vector copy no. (vg/dge) | hF.IX % dropc | Copy no. % dropd | ||
| 1.0 × 1011 | 5 | 4,524 ± 1,142 | 0.23 ± 0.03 | 80 ± 15 | 91 ± 11 |
| 3.0 × 1011 (3×)e | 5 | 16,625 ± 4,091 (3.7×) | 0.69 ± 0.16 (3.0×) | 87 ± 5 | 93 ± 2 |
| 1.8 × 1012 (6×) | 5 | 33,923 ± 9,230 (2.0×) | 2.87 ± 0.78 (4.2×) | 85 ± 3 | 90 ± 6 |
| 1.1 × 1013 (6×) | 4 | 39,945 ± 8,876 (1.2×) | 5.01 ± 1.71 (1.7×) | 82 ± 7 | 89 ± 5 |
All mean values are shown with standard deviations.
PHx, partial hepatectomy.
Averages of decreases in hF.IX levels 3 weeks after two-thirds partial hepatectomy (performed 14 weeks postinjection) relative to the levels at the time of hepatectomy (percent drop) in individual mice.
Averages of relative decreases in vector genome copy numbers per diploid genomic equivalent before and after hepatectomy in individual mice.
Numbers in parentheses indicate increments (for vector doses) or increases (for the values) compared to a dose and values at 1 lower dose.
TABLE 2.
Liver transduction with AAV-EF1α-F.IX (31 weeks postinjection)
| Dose (vg/mouse) | Mouse | hF.IX (ng/ml)
|
Vector copy no. (vg/dge)
|
Avg (mean ± SD)
|
|||||
|---|---|---|---|---|---|---|---|---|---|
| Pre-PHxa | Post-PHx | Pre-PHx | Post-PHx | Pre-PHx
|
Post-PHx
|
||||
| hF.IX (ng/ml) | Copy no. (vg/dge) | hF.IX drop (%)b | Copy no. drop (%)c | ||||||
| 5.0 × 1010 | 1 | 136 | <2 | 0.06 | <0.01 | 100 ± 32 | 0.08 ± 0.03 | >98 | NAd |
| 2 | 84 | <2 | 0.07 | <0.01 | |||||
| 3 | 79 | <2 | 0.11 | <0.01 | |||||
| 3.0 × 1011 (6×)e | 4 | 1,414 | 33 | 0.73 | 0.07 | 1,196 ± 306 (12.0×) | 0.38 ± 0.25 (4.8×) | 97 ± 1 | 85 ± 7 |
| 5 | 758 | 23 | 0.20 | 0.03 | |||||
| 6 | 1,212 | 34 | 0.20 | 0.05 | |||||
| 7 | 1,399 | 51 | 0.40 | 0.04 | |||||
| 1.8 × 1012 (6×) | 8 | 3,364 | 51 | 4.00 | 0.29 | 3,363 ± 694 (2.8×) | 4.69 ± 0.89 (12.3×) | 95 ± 5 | 84 ± 5 |
| 9 | 4,056 | 5.70 | |||||||
| 10 | 2,699 | 229 | 4.38 | 0.61 | |||||
| 7.2 × 1012 (4×) | 11 | 5,547 | 271 | 6.66 | 1.44 | 4,861 ± 970 (1.4×) | 5.66 ± 1.42 (1.2×) | 94 ± 1 | 75 ± 4 |
| 12 | 4,175 | 274 | 4.65 | 1.28 | |||||
PHx, partial hepatectomy.
Relative decrease in hF.IX levels 3 weeks after two-thirds partial hepatectomy performed 31 weeks postinjection.
Relative decrease in vector genome copy number per diploid genomic equivalent in individual mice.
NA, not applicable.
Numbers in parentheses indicate increments or increases compared to those at 1 lower dose.
TABLE 3.
Hepatocyte transduction with AAV-EF1α-nlsLacZ (28 weeks postinjection)
| Dose (vg/mouse) | Mouse | Transduction efficiency (%)a | β-Gal expression (pg/mg of protein)b | Vector copy no. (vg/dge)
|
Avg (mean ± SD)
|
||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre-PHxc | Post-PHx | Pre-PHx
|
Post-PHx copy no. % dropg | ||||||||
| Transduction efficiency (%) | β-Gal expression (pg/mg of protein) | Vector copy no. (vg/dge) | β-Gal expression rated | Net vector copy no./transduced celle (vg/dge) | |||||||
| 5.0 × 1010 | 1 | 0.6 | 88 | 0.13 | 0.01 | 0.6 ± 0.4 | 93 ± 55 | 0.19 ± 0.08 | 183 ± 137 | 40 ± 18 | 89 ± 4 |
| 2 | 0.3 | 20 | 0.15 | 0.02 | |||||||
| 3 | 0.3 | 115 | 0.18 | ||||||||
| 4 | 1.1 | 150 | 0.30 | 0.02 | |||||||
| 3.0 × 1011 (6×)f | 5 | 2.3 | 574 | 3.91 | 0.39 | 3.0 ± 0.5 (5.0×) | 544 ± 333 (5.8×) | 2.14 ± 1.26 (11.3×) | 187 ± 106 | 79 ± 62 | 77 ± 11 |
| 6 | 3.0 | 394 | 0.94 | 0.43 | |||||||
| 7 | 3.2 | 217 | 1.85 | 0.46 | |||||||
| 8 | 3.3 | 922 | 1.84 | 0.24 | |||||||
| 1.8 × 1012 (6×) | 9 | 7.8 | 2,152 | 13.61 | 8.1 ± 1.0 (2.7×) | 1,552 ± 676 (2.9×) | 15.57 ± 2.83 (7.4×) | 192 ± 88 | 196 ± 60 | 88 ± 3 | |
| 10 | 9.1 | 1,529 | 13.46 | 2.10 | |||||||
| 11 | 8.7 | 1,138 | 15.66 | 1.72 | |||||||
| 12 | 6.9 | 1,026 | 19.54 | 1.92 | |||||||
| 3.9 × 1012 (2.2×) | 13 | 8.9 | 1,237 | 16.22 | 1.50 | 8.9 (1.1×) | 1,237 (0.8×) | 16.22 (1.0×) | 138 | 182 | 91 |
X-Gal-positive nuclei/total hepatocyte nuclei counted (at least 2,000 hepatocyte nuclei were counted).
β-Gal antigen levels in liver extracts were normalized by total protein in samples.
PHx, partial hepatectomy.
β-Gal expression rate is defined as an amount of β-Gal protein per milligram of cellular protein (picograms per milligram of protein) produced from transduced hepatocytes corresponding to 1% of total hepatocytes, i.e., β-Gal expression (picograms per milligram of protein) divided by transduction efficiency (percent). The values reflect β-Gal protein production per transduced hepatocyte.
Net vector copy number is defined as the number of vector genomes per transduced hepatocyte, i.e., (vector copy number per cell [vg/dge]/transduction efficiency [%]) × 100.
Numbers in parentheses indicate increments or increases compared to 1 lower dose.
Levels of β-Gal expression after partial hepatectomy were not evaluated because of an unexpectedly long half-life of this protein in mouse hepatocytes (Nakai et al., unpublished).
As was observed with the AAV-hF.IX16-injected mice, the levels of transgene expression after AAV-EF1α-F.IX or AAV-EF1α-nlsLacZ injection were proportional (or slightly more than expected from a linear dose response) to doses between 5.0 × 1010 and 3.0 × 1011 vg/mouse, while at doses of 3.0 × 1011 vg/mouse or higher the increases in gene expression with dose were not proportional and were less than expected from a linear dose response (Tables 2 and 3). Importantly, the AAV-EF1α-nlsLacZ experiment demonstrated that the levels of transgene expression and the numbers of β-galactosidase-positive cells correlated with each other as we have seen before (24) and the number of transduced cells also increased in proportion to increasing dose between 5.0 × 1010 and 3.0 × 1011 vg/mouse, i.e., from 0.6 to 3.0% (Table 3). However, the proportion of stably transduced hepatocytes at 1.8 and 3.9 × 1012 vg/mouse (saturation doses based on the results from two independent experiments with hF.IX-expressing rAAV vectors [see Tables 1 and 2]), was only 7 to 9% (Table 3), indicating that there is likely an upper limit to the number of hepatocytes capable of being transduced.
The dose-response curves of rAAV transduction do not correlate with the number of rAAV genomes in the liver at doses above 3.0 × 1011 vg/mouse.
To investigate the molecular basis of vector dose response of rAAV transduction in the liver, we determined vector copy numbers per hepatocyte at each given vector dose by Southern blot analysis. The results are summarized in Tables 1, 2, and 3. Unless otherwise stated, vector copy number per cell or hepatocyte represents an average number of vector genomes in a cell among all the transduced and nontransduced cells in the liver. The net copy number reflects the presumed number of vector genomes in a transduced hepatocyte calculated based on the actual transduction efficiency (i.e., percentage of transduced hepatocytes). Unlike the profile of transgene expression, which started to vary off the linear dose-response curve at 3 × 1011 vg/mouse, vector genome copy numbers increased proportionally up to 1.8 × 1012 vg/mouse. At doses above 1.8 × 1012 vg/mouse, the increase in vector copy numbers per hepatocyte was no longer proportional to the increase in the vector dose. Thus, saturation of the capacity of a hepatocyte to take up or maintain vector could explain the saturation phenomenon at doses above 1.8 × 1012 vg/mouse, but it does not account for the nonlinear increase in transgene expression observed between 3 × 1011 and 1.8 × 1012 vg/mouse.
Southern blot analysis of vector genome DNA with an enzyme that does not cut within the vector genome revealed that the proportion of concatemers relative to supercoiled circular monomer genomes was increased at high doses compared to that at lower doses (Fig. 2). This observation is consistent with the restricted increases in the number of transduced hepatocytes at high doses. In addition, in the experiment using AAV-EF1α-nlsLacZ, presumed average net vector copy number per hepatocyte increased with dose from 40 vg/hepatocyte (5.0 × 1010 vg/mouse) to 196 vg/hepatocyte (1.8 × 1012 vg/mouse) while the average rate of β-Gal production per hepatocyte remained similar among the doses (Table 3). This suggests that, at higher doses of vector, the functionality of rAAV vector genomes was somehow decreased.
FIG. 2.

Southern blot analyses of rAAV vector genomes in the liver. Vector forms in AAV-hF.IX16-injected mouse liver harvested 14 weeks postinjection (A) and AAV-EF1α-nlsLacZ-injected mouse liver harvested 28 weeks postinjection (B) are shown. Twenty micrograms of total DNA was digested with SacI or NsiI (which does not cut the vector genome), separated on a 0.8% agarose gel, blotted, and probed with a vector sequence-specific probe. Lane 1.0 represents a 1.0-vg/dge standard (a 7.0-kb plasmid containing AAV-hF.IX16 vector sequence and a SacI recognition site for panel A and a 6.7-kb plasmid containing AAV-EF1α-nlsLacZ vector sequence and an NsiI recognition site for panel B), showing complete digestion of the DNA with the restriction enzyme used. Each lane represents an individual mouse. Open and filled arrowheads indicate supercoiled ds circular rAAV vector genomes and concatemers, respectively. (A) Lane 1, 1.0 × 1011; lane 2, 3.0 × 1011; lane 3, 1.8 × 1012; and lane 4, 1.1 × 1013 vg/mouse. (B) Lane 1, 5.0 × 1010; lane 2, 3.0 × 1011; lane 3, 1.8 × 1012; and lane 4, 3.9 × 1012 vg/mouse.
High-dose rAAV injection does not promote vector genome integration in the liver.
In vivo rAAV vector integration in the liver has been definitively established in a number of different studies (5, 21, 23, 26). Although the frequency of in vivo rAAV genome integration in the liver is low and a majority of vector genomes remain as extrachromosomal forms (26), rAAV integration in the liver has raised concern since the development of liver cancer was reported in one study in which rAAV vector was used to treat neonatal mucopolysaccharidosis type VII in mice (9). Although the mechanism of tumor formation in these mice is not known, the demonstration of low vector copy numbers in the tumors makes insertional mutagenesis unlikely in this case. Nonetheless, to investigate whether higher-dose vector injection promotes rAAV vector integration, we performed a two-thirds partial hepatectomy in the mice injected with AAV-hF.IX16, AAV-EF1α-F.IX, and AAV-EF1α-nlsLacZ at 14, 31, and 28 weeks postinjection, respectively. The decrease in the levels of transgene expression and vector genome copy numbers per cell was evaluated before and after hepatectomy. As summarized in Tables 1, 2, and 3, both hF.IX levels and vector copy numbers substantially dropped to levels previously observed (26), indicating that the majority of the vector genomes were extrachromosomal and that integration frequency relative to a given vector dose did not change even when higher doses of rAAV vector were administered.
In conclusion, our results demonstrated a 2-log-range linear dose response of rAAV transduction evaluated by both transgene expression and number of transduced hepatocytes. Efficiency of rAAV transduction correlated well with vector copy numbers per cell within this linear range. However, at very high doses, the increase in rAAV transduction and vector genome copy numbers were not concordant, there was an increased proportion of concatemers relative to circular monomer genomes, and liver transduction was still restricted to a small subset of hepatocytes. In addition, we also demonstrated that the relative integration frequency remained low even at high doses.
DISCUSSION
Our study defined a 2-log linear dose response of rAAV transduction in mouse liver. The highest dose in this linear range was 3.0 × 1011 vg/mouse, and assuming that the average weight of a 6- to 8-week-old animal is 20 g, the 2-log linear dose range is between 1.9 × 1011 and 1.5 × 1013 vg/kg of body weight. Interestingly, we demonstrated a nonlinear dose response at higher vector doses, indicating that saturation of rAAV transduction could occur.
Although transduction efficiency of mouse hepatocytes with rAAV vectors in vivo at doses of ∼1011 vg/mouse varies among studies, it is rarely more than ∼10%. Our study suggested that the number of hepatocytes capable of establishing stable transduction was limited to at most ∼10%, and the remaining ∼90% of hepatocytes, though capable of taking up vector genomes at these doses (19), were refractive to stable transduction even when extremely high doses of rAAV vector were administered. Given the large number of vector genomes contained within a small number of stably transduced hepatocytes (70 to 390 copies per cell) (21, 31), transduction may be limited to a small population of hepatocytes that have a specific metabolic state (19). If the protein production and secretion per hepatocyte reach capacity, further increases in gene expression may not be possible even if there are additional vector genomes per cell. This in part may limit the linear range of the dose response. Alternatively, since repeat-induced gene silencing has been shown previously for transgenic animals (12), a larger proportion of concatemers observed at high doses may cause a relative decrease in transgene transcription, resulting in a nonlinear dose response.
The extent of intermolecular recombination or joining of rAAV genomes to form concatemers is likely to depend upon the number of vector genomes in a hepatocyte; however, concatemerization might be actively promoted under certain cellular conditions. It has been proposed elsewhere that repeat-induced gene silencing may serve as a host defensive mechanism against a large number of incoming exogenous viruses (7, 28). Whether this is involved during the process of rAAV vector transduction is not known.
Nonetheless, when administration of a high dose of rAAV vector is considered, simply increasing vector dose is unlikely to linearly increase the number of stably transduced hepatocytes or transgene expression by present rAAV vector systems. One possible strategy to overcome the limited rAAV transduction in the liver is to inject multiple smaller doses at staggered intervals. We have previously found that administration of two different vectors several weeks apart results in the transduction of different populations of hepatocytes, suggesting that there is a changing subpopulation of hepatocytes permissive to transduction (19). This together with the data presented here suggests that splitting a vector dose into multiple subdoses might result in higher levels of gene expression than can be achieved from a single injection. The use of alternate serotyped vectors that may not cross-react with antibodies against AAV serotype 2 (27) may allow for repeated administration of rAAV vectors into the same subject.
If silencing of concatemers is a real event, a second strategy might be to find a means to drive the ds genomes into nonconcatemeric structures. Our recent study suggests that linear ds DNA forms are responsible for concatemer formation (Nakai et al., unpublished). Thus, if the ds molecules could be driven to form monomer circles, which are known not to form concatemers (Nakai et al., unpublished), perhaps higher levels of transgene expression could be achieved.
We have previously proposed that ds rAAV genomes responsible for stable transduction in vivo are primarily formed by annealing of complementary plus and minus ss rAAV vector genomes (25), rather than by second-strand synthesis (10, 11). Several of our previous studies supported this model for rAAV transduction in the liver (25). In our present study, we observed a linear dose response, which could be easily explained by the second-strand synthesis model. This led us to consider whether the annealing model is also viable with the observed 2-log-range linear dose response. To address this issue, we evaluated the model in mathematical terms. For simplicity, we made the assumption that all hepatocytes that establish stable transduction receive equal numbers of ss rAAV genomes and also that second-strand synthesis or annealing occurs with maximum efficiency. A ds monomer genome can be formed either by annealing of two complementary plus and minus parental ss genomes or by second-strand synthesis from one parental ss genome, depending on which model is under consideration. In the following equations, we defined a transduction unit as an amount of transgene product generated from one ds rAAV monomer genome in a cell.
In the second-strand synthesis model, a hepatocyte with n parental ss vector copies can generate n copies of ds rAAV genomes by leading-strand synthesis; therefore, it can produce n units of protein, i.e., the amount of protein per hepatocyte with n parental ss genomes = f1(n) = n (units).
In the annealing model, supposing that a hepatocyte takes up a total of n copies of plus and minus parental ss genomes randomly (n is an even number in this case), a hepatocyte with n parental ss vector genomes in theory can produce f2(n) units of protein, i.e.:
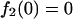 |
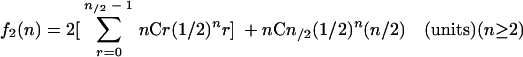 |
For instance, f2(2) = 0.5, f2(4) = 1.25, f2(10) = 3.77, f2(20) = 8.24, f2(40) = 17.49, and so forth. When these values are plotted as a function of parental ss vector copy number per cell, a theoretical dose-response curve in the annealing model also exhibits fair linearity (Fig. 3). Therefore, our observation of linear dose response in rAAV transduction is not inconsistent with our previous finding that annealing of vector genomes is responsible for the formation of stable ds rAAV vector genomes.
FIG. 3.

Theoretical dose-response curves in a second-strand synthesis model and an annealing model, in such a situation that all incoming ss genomes are utilized for stable transduction in a most efficient manner. Representative values (n = 0, 2, 4, 10, 20, and 40; n represents the number of parental ss vector genomes in a hepatocyte) are plotted. A transduction unit is defined as an amount of transgene product in a cell which can be produced from one ds rAAV genome. The regression curve for the annealing model in this graph is f2(n) = −0.049 + 0.413n (r = 0.999), and therefore f2(n) is increased in proportion to n.
In summary, our present study revealed a dose-response profile in rAAV-mediated liver transduction and confirmed low integration frequency and restricted transduction even after administration of high vector doses. We still do not know which episomal forms (supercoiled ds circular monomers versus episomal circular and/or linear concatemers) are primarily responsible for stable liver-directed gene expression; however, our present results are not inconsistent with our present hypothesis that circular monomer genomes, not concatemers, are the major source of transgene expression. Further analysis of transgene expression from different episomal vector forms should provide greater insights into the mechanisms underlying stable rAAV transduction in the liver.
Acknowledgments
H.N. and C.E.T. contributed equally to the work.
This study was supported by NIH grant HL 66948.
REFERENCES
- 1.Aitken, M. L., R. B. Moss, D. A. Waltz, M. E. Dovey, M. R. Tonelli, S. C. McNamara, R. L. Gibson, B. W. Ramsey, B. J. Carter, and T. C. Reynolds. 2001. A phase I study of aerosolized administration of tgAAVCF to cystic fibrosis subjects with mild lung disease. Hum. Gene Ther. 12:1907-1916. [DOI] [PubMed] [Google Scholar]
- 2.Arruda, V. R., P. A. Fields, R. Milner, L. Wainwright, M. P. De Miguel, P. J. Donovan, R. W. Herzog, T. C. Nichols, J. A. Biegel, M. Razavi, M. Dake, D. Huff, A. W. Flake, L. Couto, M. A. Kay, and K. A. High. 2001. Lack of germline transmission of vector sequences following systemic administration of recombinant AAV-2 vector in males. Mol. Ther. 4:586-592. [DOI] [PubMed] [Google Scholar]
- 3.Burton, M., H. Nakai, P. Colosi, J. Cunningham, R. Mitchell, and L. Couto. 1999. Coexpression of factor VIII heavy and light chain adeno-associated viral vectors produces biologically active protein. Proc. Natl. Acad. Sci. USA 96:12725-12730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chao, H., L. Mao, A. T. Bruce, and C. E. Walsh. 2000. Sustained expression of human factor VIII in mice using a parvovirus-based vector. Blood 95:1594-1599. [PubMed] [Google Scholar]
- 5.Chen, S. J., J. Tazelaar, A. D. Moscioni, and J. M. Wilson. 2000. In vivo selection of hepatocytes transduced with adeno-associated viral vectors. Mol. Ther. 1:414-422. [DOI] [PubMed] [Google Scholar]
- 6.Chen, S. J., J. Tazelaar, and J. M. Wilson. 2001. Selective repopulation of normal mouse liver by hepatocytes transduced in vivo with recombinant adeno-associated virus. Hum. Gene Ther. 12:45-50. [DOI] [PubMed] [Google Scholar]
- 7.Cogoni, C. 2001. Homology-dependent gene silencing mechanisms in fungi. Annu. Rev. Microbiol. 55:381-406. [DOI] [PubMed] [Google Scholar]
- 8.Daly, T. M., C. Vogler, B. Levy, M. E. Haskins, and M. S. Sands. 1999. Neonatal gene transfer leads to widespread correction of pathology in a murine model of lysosomal storage disease. Proc. Natl. Acad. Sci. USA 96:2296-2300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Donsante, A., C. Vogler, N. Muzyczka, J. M. Crawford, J. Barker, T. Flotte, M. Campbell-Thompson, T. Daly, and M. S. Sands. 2001. Observed incidence of tumorigenesis in long-term rodent studies of rAAV vectors. Gene Ther. 8:1343-1346. [DOI] [PubMed] [Google Scholar]
- 10.Ferrari, F. K., T. Samulski, T. Shenk, and R. J. Samulski. 1996. Second-strand synthesis is a rate-limiting step for efficient transduction by recombinant adeno-associated virus vectors. J. Virol. 70:3227-3234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Fisher, K. J., G. P. Gao, M. D. Weitzman, R. DeMatteo, J. F. Burda, and J. M. Wilson. 1996. Transduction with recombinant adeno-associated virus for gene therapy is limited by leading-strand synthesis. J. Virol. 70:520-532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Garrick, D., S. Fiering, D. I. Martin, and E. Whitelaw. 1998. Repeat-induced gene silencing in mammals. Nat. Genet. 18:56-59. [DOI] [PubMed] [Google Scholar]
- 13.Greelish, J. P., L. T. Su, E. B. Lankford, J. M. Burkman, H. Chen, S. K. Konig, I. M. Mercier, P. R. Desjardins, M. A. Mitchell, X. G. Zheng, J. Leferovich, G. P. Gao, R. J. Balice-Gordon, J. M. Wilson, and H. H. Stedman. 1999. Stable restoration of the sarcoglycan complex in dystrophic muscle perfused with histamine and a recombinant adeno-associated viral vector. Nat. Med. 5:439-443. [DOI] [PubMed] [Google Scholar]
- 14.Jung, S. C., I. P. Han, A. Limaye, R. Xu, M. P. Gelderman, P. Zerfas, K. Tirumalai, G. J. Murray, M. J. During, R. O. Brady, and P. Qasba. 2001. Adeno-associated viral vector-mediated gene transfer results in long-term enzymatic and functional correction in multiple organs of Fabry mice. Proc. Natl. Acad. Sci. USA 98:2676-2681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kay, M. A., Q. Li, T. J. Liu, F. Leland, C. Toman, M. Finegold, and S. L. Woo. 1992. Hepatic gene therapy: persistent expression of human α1-antitrypsin in mice after direct gene delivery in vivo. Hum. Gene Ther. 3:641-647. [DOI] [PubMed] [Google Scholar]
- 16.Kay, M. A., C. S. Manno, M. V. Ragni, P. J. Larson, L. B. Couto, A. McClelland, B. Glader, A. J. Chew, S. J. Tai, R. W. Herzog, V. Arruda, F. Johnson, C. Scallan, E. Skarsgard, A. W. Flake, and K. A. High. 2000. Evidence for gene transfer and expression of factor IX in haemophilia B patients treated with an AAV vector. Nat. Genet. 24:257-261. [DOI] [PubMed] [Google Scholar]
- 17.Kessler, P. D., G. M. Podsakoff, X. Chen, S. A. McQuiston, P. C. Colosi, L. A. Matelis, G. J. Kurtzman, and B. J. Byrne. 1996. Gene delivery to skeletal muscle results in sustained expression and systemic delivery of a therapeutic protein. Proc. Natl. Acad. Sci. USA 93:14082-14087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Matsushita, T., S. Elliger, C. Elliger, G. Podsakoff, L. Villarreal, G. J. Kurtzman, Y. Iwaki, and P. Colosi. 1998. Adeno-associated virus vectors can be efficiently produced without helper virus. Gene Ther. 5:938-945. [DOI] [PubMed] [Google Scholar]
- 19.Miao, C. H., H. Nakai, A. R. Thompson, T. A. Storm, W. Chiu, R. O. Snyder, and M. A. Kay. 2000. Nonrandom transduction of recombinant adeno-associated virus vectors in mouse hepatocytes in vivo: cell cycling does not influence hepatocyte transduction. J. Virol. 74:3793-3803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Miao, C. H., K. Ohashi, G. A. Patijn, L. Meuse, X. Ye, A. R. Thompson, and M. A. Kay. 2000. Inclusion of the hepatic locus control region, an intron, and untranslated region increases and stabilizes hepatic factor IX gene expression in vivo but not in vitro. Mol. Ther. 1:522-532. [DOI] [PubMed] [Google Scholar]
- 21.Miao, C. H., R. O. Snyder, D. B. Schowalter, G. A. Patijn, B. Donahue, B. Winther, and M. A. Kay. 1998. The kinetics of rAAV integration in the liver. Nat. Genet. 19:13-15. [DOI] [PubMed] [Google Scholar]
- 22.Nakai, H., R. W. Herzog, J. N. Hagstrom, J. Walter, S. H. Kung, E. Y. Yang, S. J. Tai, Y. Iwaki, G. J. Kurtzman, K. J. Fisher, P. Colosi, L. B. Couto, and K. A. High. 1998. Adeno-associated viral vector-mediated gene transfer of human blood coagulation factor IX into mouse liver. Blood 91:4600-4607. [PubMed] [Google Scholar]
- 23.Nakai, H., Y. Iwaki, M. A. Kay, and L. B. Couto. 1999. Isolation of recombinant adeno-associated virus vector-cellular DNA junctions from mouse liver. J. Virol. 73:5438-5447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nakai, H., T. A. Storm, and M. A. Kay. 2000. Increasing the size of rAAV-mediated expression cassettes in vivo by intermolecular joining of two complementary vectors. Nat. Biotechnol. 18:527-532. [DOI] [PubMed] [Google Scholar]
- 25.Nakai, H., T. A. Storm, and M. A. Kay. 2000. Recruitment of single-stranded recombinant adeno-associated virus vector genomes and intermolecular recombination are responsible for stable transduction of liver in vivo. J. Virol. 74:9451-9463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nakai, H., S. R. Yant, T. A. Storm, S. Fuess, L. Meuse, and M. A. Kay. 2001. Extrachromosomal recombinant adeno-associated virus vector genomes are primarily responsible for stable liver transduction in vivo. J. Virol. 75:6969-6976. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rabinowitz, J. E., F. Rolling, C. Li, H. Conrath, W. Xiao, X. Xiao, and R. J. Samulski. 2002. Cross-packaging of a single adeno-associated virus (AAV) type 2 vector genome into multiple AAV serotypes enables transduction with broad specificity. J. Virol. 76:791-801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Selker, E. U. 1999. Gene silencing: repeats that count. Cell 97:157-160. [DOI] [PubMed] [Google Scholar]
- 29.Snyder, R. O., C. Miao, L. Meuse, J. Tubb, B. A. Donahue, H. F. Lin, D. W. Stafford, S. Patel, A. R. Thompson, T. Nichols, M. S. Read, D. A. Bellinger, K. M. Brinkhous, and M. A. Kay. 1999. Correction of hemophilia B in canine and murine models using recombinant adeno-associated viral vectors. Nat. Med. 5:64-70. [DOI] [PubMed] [Google Scholar]
- 30.Snyder, R. O., C. H. Miao, G. A. Patijn, S. K. Spratt, O. Danos, D. Nagy, A. M. Gown, B. Winther, L. Meuse, L. K. Cohen, A. R. Thompson, and M. A. Kay. 1997. Persistent and therapeutic concentrations of human factor IX in mice after hepatic gene transfer of recombinant AAV vectors. Nat. Genet. 16:270-276. [DOI] [PubMed] [Google Scholar]
- 31.Song, S., J. Embury, P. J. Laipis, K. I. Berns, J. M. Crawford, and T. R. Flotte. 2001. Stable therapeutic serum levels of human α1 antitrypsin (AAT) after portal vein injection of recombinant adeno-associated virus (rAAV) vectors. Gene Ther. 8:1299-1306. [DOI] [PubMed] [Google Scholar]
- 32.Wagner, J. A., A. H. Messner, M. L. Moran, R. Daifuku, K. Kouyama, J. K. Desch, S. Manley, A. M. Norbash, C. K. Conrad, S. Friborg, T. Reynolds, W. B. Guggino, R. B. Moss, B. J. Carter, J. J. Wine, T. R. Flotte, and P. Gardner. 1999. Safety and biological efficacy of an adeno-associated virus vector-cystic fibrosis transmembrane regulator (AAV-CFTR) in the cystic fibrosis maxillary sinus. Laryngoscope 109:266-274. [DOI] [PubMed] [Google Scholar]
- 33.Xiao, W., S. C. Berta, M. M. Lu, A. D. Moscioni, J. Tazelaar, and J. M. Wilson. 1998. Adeno-associated virus as a vector for liver-directed gene therapy. J. Virol. 72:10222-10226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Xu, L., T. Daly, C. Gao, T. R. Flotte, S. Song, B. J. Byrne, M. S. Sands, and K. Parker Ponder. 2001. CMV-β-actin promoter directs higher expression from an adeno-associated viral vector in the liver than the cytomegalovirus or elongation factor 1α promoter and results in therapeutic levels of human factor X in mice. Hum. Gene Ther. 12:563-573. [DOI] [PubMed] [Google Scholar]