

Short abstract
The importin-β-like family of nuclear transport receptors mediates the transport of a large number of substrates between the cytoplasm and the nucleus. The family is made up of importins and exportins which shuttle between the nucleus and the cytoplasm and are regulated by the small GTPase Ran.
Abstract
In recent years, our understanding of macromolecular transport processes across the nuclear envelope has grown dramatically, and a large number of soluble transport receptors mediating either nuclear import or nuclear export have been identified. Most of these receptors belong to one large family of proteins, all of which share homology with the protein import receptor importin β (also named karyopherin β). Members of this family have been classified as importins or exportins on the basis of the direction they carry their cargo. To date, the family includes 14 members in the yeast Saccharomyces cerevisiae and at least 22 members in humans. Importins and exportins are regulated by the small GTPase Ran, which is thought to be highly enriched in the nucleus in its GTP-bound form. Importins recognize their substrates in the cytoplasm and transport them through nuclear pores into the nucleus. In the nucleoplasm, RanGTP binds to importins, inducing the release of import cargoes. In contrast, exportins interact with their substrates only in the nucleus in the presence of RanGTP and release them after GTP hydrolysis in the cytoplasm, causing disassembly of the export complex. Thus, common features of all importin-β-like transport factors are their ability to shuttle between the nucleus and the cytoplasm, their interaction with RanGTP as well as their ability to recognize specific transport substrates.
Gene organization and evolutionary history
A major effort in the nucleocytoplasmic transport field has been directed towards the analysis of all members of the importin β family, in particular with the aim of identifying specific transport cargoes. This has led to the characterization of a large number of related proteins (both importins and exportins) in all eukaryotic species analyzed. New members have primarily been identified by sequence homology or biochemically via their interaction with the small GTPase Ran. Strikingly, the roles of many of these receptors is conserved from yeast to humans. There are 14 putative members of the importin β family in the completed Saccharomyces cerevisiae genome, nine of which have been shown to function as importins and four as exportins [1]. The genes encoding yeast transport receptors are dispersed throughout the genome and none of them contains introns. Higher eukaryotes contain an even larger number of importin-β-like proteins. It has been proposed that there are more than 22 putative members in mammals [1,2,3]. Little is known about the gross structure of the genes encoding these nuclear transport receptors, and our knowledge of the chromosomal localization or the organization of the individual genes encoding members of this family is very poor.
Characteristic structural features
The relative molecular masses of members of importin-β-like proteins vary between 90 kDa and 130 kDa, but all are characterized by an acidic isoelectric point. The overall sequence similarity between various transport receptors is low (less than 20% amino acid identity) and, in many cases, is restricted to the amino-terminal domain. Work mainly on importin β has demonstrated that these receptors bind RanGTP via the amino-terminal domain and cargo via the carboxy-terminal domain [4,5,6,7,8]. To permit shuttling through the nuclear pore complex (NPC), transport receptors also contain one or multiple binding domains for components of the NPCs, called nucleoporins. Truncation studies using importin β indicate that the binding site for nucleoporins containing FxFG repeats (in the single-letter amino acid code, where x is, in many cases, a small polar residue or glycine) is located in an amino-terminal/central region of importin β (residues 152-352) [4,5]. This was recently confirmed in the crystal structure of an amino-terminal fragment of importin β in a complex with five FxFG nucleoporin repeats [9]. Additional information comes from the crystal structures of importin β and of transportin 1 (also known as karyopherin β2), which were solved either in a complex with RanGTP [7,8], with a cargo [6], or in the free form [10].
Overall, the structures of importin-β-like receptors are characterized by a very similar series of helical HEAT repeats (19 in importin β and 18 in transportin 1; Figure 1). HEAT repeats are approximately 40 residues in length and are found in many eukaryotic proteins such as the PR65/A subunit of protein phosphatase 2A [11]. The fundamental repeat unit is a right-handed superhelical structure consisting of a hairpin made up of two β helices, named A and B, separated by a sharp turn. Each hairpin is connected to the next by a linker region. In transportin 1, almost all linkers contain a third helix, but there are very few linker helices in importin β. In both receptors, one turn is extended into a long acidic loop, which has been suggested to be important for RanGTP-mediated cargo release. Full-length importin β complexed with the importin-β-binding domain of importin α (IBB) forms a snail-like superhelical structure wrapping tightly around the IBB domain. The structure of the uncomplexed amino terminus of importin β reveals a different superhelical architecture with a much steeper helical pitch than the cargo-bound or RanGTP-bound forms [10]. This suggests that importin β undergoes twisted conformational changes in its HEAT-repeat helix stacking, which could be essential for the regulation of cargo binding and release and/or for protein interactions during the translocation through the NPC. No structure of an exportin has yet been reported. Although the sequence homology is limited, it is expected that exportins will fold in a similar way to the reported importin structures. It still remains unclear, however, why RanGTP is required for binding of cargo to exportins but causes cargo dissociation from importins.
Figure 1.
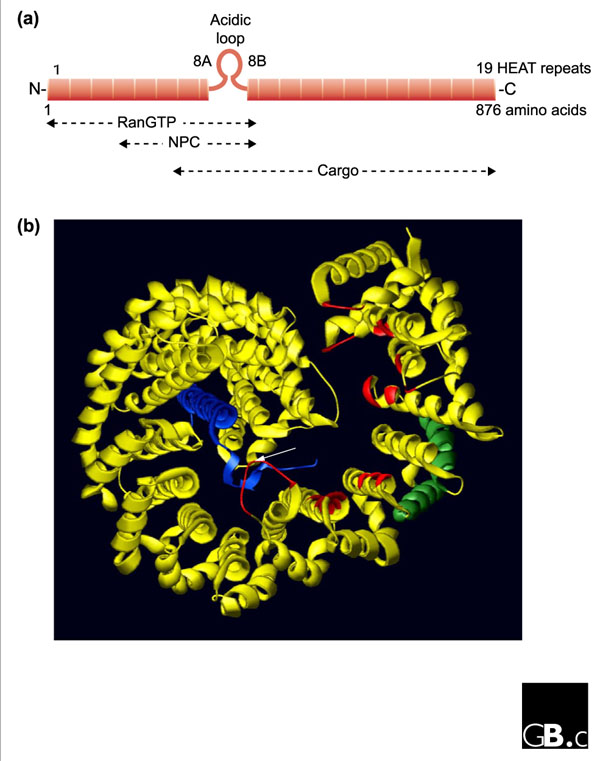
Structure of importin β. (a) Importin β is composed of 19 helical-repeat motifs (HEAT repeats). Each consists of an A and a B helix connected by a short turn, which in HEAT-8 is replaced by an acidic loop critical for the regulation of substrate binding and release. The HEAT repeats 1-8 are required for high-affinity binding to RanGTP [4,5]. The importin-β-binding (IBB) domain of importin α interacts mainly with residues located in repeats 7-19 of importin β [6]. The binding site for nucleoporins of the NPC is located between residues 152 and 352, corresponding to repeats 4-8 [4,5]. On the basis of the crystal structure, the A helices of HEAT repeats 5 and 6 and a region between HEAT repeats 6 and 7 are thought to be critical for recognition of the FxFG motif [9]. N, amino terminus; C, carboxyl terminus. (b) Structure of importin β bound to the IBB domain of importin α (adapted from [6]). Importin β (yellow) forms a superhelical structure that wraps like a snail around the IBB domain (blue). The 19 HEAT repeats share a common core of 21 residues, comprising the A helix with about three turns and the B helix with about four turns. The helices critical for the interaction with FxFG-repeat nucleoporins [9] are in green. Important residues for interaction with RanGTP [8] are in red. Note the acidic loop, which contacts both RanGTP and the IBB domain (white arrow).
Localization and function
A major function of transport factors of the importin β family is to mediate the transport between the nucleus and cytoplasm of macromolecules that contain nuclear import or export signals [1,12]. To this end, all transport factors constantly shuttle between the nucleus and the cytoplasm. At steady state, they can be found in the nucleus, at the NPC or in the cytoplasm. At present, very little is known about the tissue distribution of this family of proteins. All members have the ability to recognize and bind specific cargoes, either directly or via adaptor molecules, to bind RanGTP and to interact with nucleoporins at the NPC. Interactions between the proteins of the importin β family and nucleoporin repeats have been shown both in vitro [4,5,13,14,15,16,17] and in vivo [18]. These interactions contribute to the import or export of importin β family members and their cargoes through the central transporter of the NPC.
Import and export are multistep processes that are initiated by the recognition of nuclear localization signals (NLSs) and nuclear export signals (NESs). The most thoroughly studied import signals are the 'classical' and the bipartite NLSs, first identified in SV40 large T antigen and nucleoplasmin, respectively [19]. Their transport is mediated by importin β, the first-characterized member of this protein family. Importin β indirectly associates with these NLS motifs via the adaptor molecule importin α [20,21,22,23,24]. Additional importin-β-dependent adaptors in vertebrates include snurportin 1 (involved in import of m3G-capped small nuclear ribonucleoproteins, snRNPs [25]) and XRIPα, (involved in the import of replication protein A, RPA [26]). Importin β can also form a complex with another importin-β-like factor, importin 7, in order to transport the linker histone H1 into the nucleus [27]. In addition, importin β is able to interact directly with a large variety of different cargoes, including the T-cell protein tyrosine phosphatase [28], the human immunodeficency virus (HIV) TAT and Rev proteins [29], human T-cell leukemia virus Rex protein [30], ribosomal proteins L23a, S7, and L5 [31], cyclin B1 [32,33], Smad [34] and the parathyroid-hormone-related protein [35]. Table 1 details known importins, exportins and their cargoes and adaptors.
Table 1.
Proteins in the importin-β-like family and their cargoes
| Vertebrate | Yeast | Cargoes |
| Importins | ||
| Importin β (Kap β1, p97, PTACβ) [L39793] | Kap95p [AAB67265] | Ribosomal proteins (H.s.) [31], viral proteins [29], cyclin B1 (X.l. and |
| H.s.) [32,33], Smad proteins (H.s.) [34], T-cell protein tyrosine phos- | ||
| phatase (H.s.) [28]. Adaptors: importin α: cNLS-containing proteins | ||
| (reviewed in [1]); importin α/4E-T: eukaryotic translation initiation | ||
| factor 4E (H.s.) [78]; snurportin: m3G-capped U snRNPs (H.s.) [25] | ||
| XRIP α: RPA (X.l.) [26] | ||
| Transportin 1 (importin β2, karyopherin β2) | hnRNP protiens (A1, F) (H.s.) [39,42]; ribosomal proteins (H.s.) [31] | |
| [U70322] | ||
| Kap104p [CAA84959] | Nab2p (S.c.) and Hrp1p (S.c.) [38] | |
| Transportin-SR (transportin-SR2 & Mtr10a) | SR domain proteins (H.s.) [43,44] | |
| [AJ133769] | ||
| Importin 5 (RanBP5, karyopherin β3) | Ribosomal proteins (H.s.) [31] | |
| [U72761] | ||
| Kap108p/SXM1 [AAB64837] | Lhp1p (S.c.) & ribosomal proteins (S.c.) [79] | |
| Kap111p/Mtr10p [CAA99366] | Npl3p (S.c.) [80] | |
| Kap114p/YGL241W [CAA96960] | TBP (S.c.) [45] | |
| Importin 7 (RanBP7) [AF098799] | Ribosomal proteins (H.s) [31]; mediates import of histone H1 (H.s.) | |
| [27] when bound to importin β | ||
| Kap119p/Nmd5p [CAA89663] | Hog1p (S.c.) [58], TFIIS (S.c.) [81] | |
| Kap121p/Pse1p [CAA89141] | Pho4p (S.c.) [82], ribosomal proteins (S.c.) [36], Spo12p (S.c.) [83] | |
| Kap122p/Pdr6p [CAA96716] | Toa1p and Toa2p (S.c.) [84] | |
| Kap123p/Yrb4 [AAC03208] | Ribosomal protein (S.c.) [36,37] | |
| Kap142p/Msn5p [AAB64771] | Replication protein A (S.c.) [65] | |
| Importin 11 [from ESTAA082435] | UbcM2 (murine) [3] | |
| Exportins | ||
| Exportin 1 (CRM1, XPO1) [Y08614] | Crm1p/Xpo1p [CAA97246] | NES-containing proteins [46,47,48,49,52,54,55], m7G-capped U snRNAs |
| (X.l.) [53], snurportin 1 [H.s] [51], IκBα (avian) [50], Ssb1p (S.c.) [59], | ||
| Ace2p (S.c.) [57], Yap1p (S.c.) [56], Hog1p (S.c.) [58] | ||
| CAS [U33286] | Cse1p/Kap109p [CAA96957] | Importin α (H.s. and S.c.) [60,61,62,63] |
| Exportin-t [AF039022 ] | Los1p [CAA82050] | tRNA (H.s. and S.c.) [66,67,68] |
| Exportin 4 [AF145021] | eIF-5A (H.s.) [85] | |
| Kap142p/Msn5p [AAB64771] | Pho4p (S.c.) [64], Mig1p (S.c.) [86], Far1p (S.c.) [87], Ste5p (S.c.) [88] | |
| Examples of orphan importins/exportins | ||
| Transportin 2 (karyopherin β2b) [AF019039] | Unknown | |
| RanBP6 [AF039023] | Unknown | |
| RanBP8 [U77494] | Found in a complex with importin β, unknown function [89] | |
| RanBP13 (Mtr10b) [AF267987] | Unknown | |
| RanBP16 [AF064729] | Unknown | |
| RanBP17 [AF222747] | Unknown | |
| Kap120p/Lph2p [AAB68237] | Unknown |
Abbreviations: H.s., Homo sapiens; S.c., Saccharomyces cerevisiae; X.l., Xenopus laevis. GenBank accession numbers are given in square brackets for each protein, and alternative names are in parenthesis.
The import of ribosomal proteins seems to rely on at least partially redundant mechanisms. In yeast, the importin Kap123p/Yrb4p has been shown to be an important mediator of ribosomal-protein import [36,37], but the related protein Kap121p/Pse1p can functionally substitute for Kap123p in vivo [36]. In mammalian cells, at least four importin-β-like transport factors are able to mediate import of ribosomal proteins [31]. Interestingly, both the β-like importin receptor binding (BIB) domain and some ribosomal proteins can be imported by any of the four receptors importin β, transportin 1, importin 5 or importin 7 [31].
The yeast importin Kap104p mediates nuclear import of the mRNA-binding proteins Nab2p and Nab4p [38]. Its vertebrate homolog, transportin 1 (Kapβ2), also mediates import of the RNA-binding proteins hnRNP A1 and hnRNP F, but also of ribosomal proteins [31,39,40,41,42]. Mammalian transportin-SR mediates nuclear import of a group of abundant arginine/serine-rich proteins, which are essential pre-mRNA splicing factors [43,44]. In yeast, the TATA-binding protein (TBP) is imported into the nucleus by Kap114p [45].
Other members of the importin-β family have been found to mediate nuclear export events. Exportin 1 (Crm1p, Xpo1p) has been identified in both yeast and higher eukaryotes as an export receptor for leucine-rich NES-containing proteins [46,47,48,49]. Human exportin 1 has been found to export protein kinase inhibitor α (PKIα) and HIV Rev [46], IκBα [50], snurportin 1 [51], HTLV Rex [52], the small nuclear RNA [46,53], cyclin B1 [54] and the transcription factor NF-AT4 [55]. Targets for exportin in S. cerevisiae include the transcription factors Yap1p [56], and Ace2p [57], the mitogen-activated protein kinase Hog1p [58], and the heat-shock protein Ssb1p [59]. CAS (yeast Cse1p) functions in the export and recycling of importin α [60,61,62,63]. Msn5p was first identified as a yeast exportin that exports the phosphorylated form of the transcription factor Pho4p [64]. Interestingly, Msn5p was recently shown to function as an import receptor for the trimeric RPA [65], suggesting that individual members of the importin-β-like protein family can function both as import and export receptors. The first cellular RNA export receptor to be discovered, named exportin-t in higher eukaryotes or Los1p in yeast, mediates nuclear export of tRNAs [66,67,68].
Many of the nuclear transport factors identified in S. cerevisiae are not essential for viability, even though they transport essential cargoes. This phenomenon can be explained by the fact that cargoes can use alternative transport pathways. This is probably best exemplified in the import pathway of ribosomal proteins, which is mediated by at least four different import receptors (see above).
Mechanism and regulation
Nuclear transport mediated by importins and exportins is strongly directional in vivo: importins bind their cargo in the cytoplasm and transfer it to the nucleus, whereas exportins interact with their substrates in the nucleus and mediate their export to the cytoplasm (Figure 2). Protein translocation through the NPC is thought to occur by an essentially similar mechanism for all importin-β-related receptors, except for the fact that, in some situations, additional adaptors are required to bridge the cargo-receptor interaction. The most-studied pathway is the import of classical NLS-containing proteins. This is mediated by importin β together with its adaptor importin α, which binds both the NLS-containing cargo and importin β in the cytoplasm. After a trimeric importin α-importin β-NLS complex is formed, importin β mediates docking at the NPC. In the presence of RanGDP and free GTP, this trimeric complex translocates through the NPC. Translocation is terminated by binding of RanGTP to importin β, which releases the complex from the NPC and dissociates importin α from importin β (reviewed in [1,12,69]). Free importin α has a lower affinity for the NLS cargo [60], and release from importin β is therefore believed to trigger release of the NLS cargo as well. Thereafter, the importin β-RanGTP complex, and importin α bound to its exportin (CAS) and RanGTP, are re-exported to the cytoplasm for another round of import [1,12,69].
Figure 2.
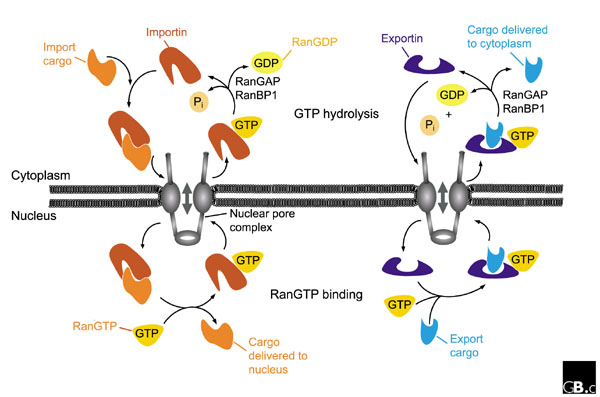
A schematic representation of nuclear import and export cycles through the NPC. Typically, an import cargo is first recognized by its importin in the cytoplasm. The cargo-loaded importin translocates through the NPC into the nucleus, where the cargo is dissociated from the importin by binding of importin to RanGTP. The importin-RanGTP complex recycles back to the cytoplasm, where RanGTP hydrolysis is stimulated by RanGAP and RanBP1; this frees the importin for the next round of import. Binding of cargoes to exportins is regulated in a converse manner. Exportins bind their export substrates in the nucleus, forming a trimeric cargo-exportin-RanGTP complex. This complex is exported from the nucleus and dissociated in the cytoplasm by hydrolysis of RanGTP to RanGDP and inorganic phosphate (Pi). This releases the export substrate, and the exportin is recycled back into the nucleus. For details, see text.
Transport in the reverse direction, mediated by exportins, is regulated in a converse manner (Figure 2) [1,12,69]. A paradigm for transport out of the nucleus is the export of leucine-rich NES-containing proteins. Exportin 1 binds to substrates containing a leucine-rich NES in the nucleus, forming a trimeric complex with RanGTP. This complex is then transferred to the cytoplasm by a mechanism involving binding of exportin 1 to the NPC. Once in the cytoplasm, GTP hydrolysis results in dissociation of Ran from the complex, allowing exportin 1 to release its cargo. Free exportin 1 re-enters the nucleus to bind and export additional cargo molecules.
As illustrated in these examples, the RanGTP cycle plays a key role in conferring directionality to nucleocytoplasmic transport events [70,71] and RanGTP acts as a marker of the nuclear compartment for both nuclear import and export (Figure 2; reviewed in [1,12,69]). Remarkably, this model predicts that only a single molecule of GTP is hydrolyzed per import/export cycle; it strictly requires that RanGTP is highly enriched in the nucleus. It is thought that a steep RanGTP-RanGDP gradient is generated by the cellular compartmentalization of regulators of the Ran cycle. Specifically, the guanine-nucleotide exchange factor of Ran (RanGEF or RCC1), which regenerates RanGTP is nuclear and bound to chromatin [72]. In contrast, the main GTPase-activating protein (RanGAP), and the Ran-binding proteins, RanBP1 and RanBP2, which stimulate GTP hydrolysis by Ran, are found in the cytoplasm [73,74,75,76]. This asymmetric distribution predicts that Ran is present mainly in the GTP-bound form in the nucleus, whereas Ran is immediately converted to a GDP-bound state in the cytoplasm.
Recently, it was suggested that in addition to the Ran cycle, the NPC itself could provide an additional mechanism to ensure transport directionality [77]. Given that several nucleoporins implicated in binding to importins and exportins have distinctive locations in the structure of the NPC, the asymmetric design of the NPC may also be important to efficiently drive nuclear import and export.
Frontiers
Despite the large amount of progress that has been made in the nucleocytoplasmic transport field in recent years, many important questions remain unsolved. Many import and export receptors have now been characterized and their first cargoes have been identified. The further characterization of new transport receptors and adaptors, and the identification of new import and export substrates, will lead to a more complete picture of nucleocytoplasmic transport. A big challenge for the future will be to understand how translocation through the NPC occurs, and how nucleocytoplasmic transport is regulated. How does the NPC achieve its tremendous amount of selectivity? What type of changes in NPC conformation are required for passage through the NPC? To gain insight into these questions, a quantitative analysis of interactions between transport receptors and nucleoporins will be required. It will be also interesting to see whether an increasing affinity gradient of receptors for nucleoporins along the NPC exists and, if so, whether it makes an important contribution towards the direction of transport.
Other issues that remain to be solved include the structural differences between members of the importin and exportin family. How does RanGTP dissociate import complexes in the nucleus but promote binding of export cargoes to exportins? A key mechanistic topic that is poorly understood is the export of RNAs and RNPs. Although a tRNA-export factor has been identified, the mechanism of rRNA or mRNA export is still poorly understood. Several proteins in S. cerevisiae, such as Mex67p and Yra1p, and their metazoan counterparts TAP and Aly, have been indicated to play an important role in mRNA export. Mex67p/TAP does not belong to the family of importin-β-like proteins, suggesting that there are alternative translocation pathways through the NPC. Future studies will provide insights into these important questions.
References
- Görlich D, Kutay U. Transport between the cell nucleus and the cytoplasm. Annu Rev Cell Dev Biol. 1999;15:607–660. doi: 10.1146/annurev.cellbio.15.1.607. A comprehensive review on nucleocytoplasmic transport, which describes the role of importins, exportins and Ran in nuclear transport and contains many references to the original work and other reviews. [DOI] [PubMed] [Google Scholar]
- Kutay U, Hartmann E, Treichel N, Calado A, Carmo-Fonseca M, Prehn S, Kraft R, Görlich D, Bischoff FR. Identification of two novel RanGTP-binding proteins belonging to the importin {beta} superfamily. J Biol Chem. 2000;275:40163–40168. doi: 10.1074/jbc.M006242200. Using affinity chromatography with immobilized RanGTP the authors isolate RanBP16. Another close homolog, RanBP17, is also identified on the basis of RanBP16 sequence. [DOI] [PubMed] [Google Scholar]
- Plafker SM, Macara IG. Importin-11, a nuclear import receptor for the ubiquitin-conjugating enzyme, UbcM2. EMBO J. 2000;19:5502–5513. doi: 10.1093/emboj/19.20.5502. This paper identifies importin 11, a human member of the importin-β family that mediates the nuclear import of UbcM2, an E2 ubiquitin-conjugating enzyme. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutay U, Izaurralde E, Bischoff FR, Mattaj IW, Görlich D. Dominant-negative mutants of importin-beta block multiple pathways of import and export through the nuclear pore complex. EMBO J. 1997;16:1153–1163. doi: 10.1093/emboj/16.6.1153. The authors analyze the interaction sites of importin β with its multiple partners. These data suggest that termination of import involves binding of RanGTP to importin β. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chi NC, Adam SA. Functional domains in nuclear import factor p97 for binding the nuclear localization sequence receptor and the nuclear pore. Mol Biol Cell. 1997;8:945–956. doi: 10.1091/mbc.8.6.945. The authors map the binding domains of importin β with the pore and the NLS receptor using deletion analysis and site-directed mutagenesis. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cingolani G, Petosa C, Weis K, Muller CW. Structure of importin-beta bound to the IBB domain of importin-alpha. Nature. 1999;399:221–229. doi: 10.1038/20367. The authors describe the crystal structure of full-length human importin β bound to the IBB domain of importin α. [DOI] [PubMed] [Google Scholar]
- Chook YM, Blobel G. Structure of the nuclear transport complex karyopherin-beta2-Ran x GppNHp. Nature. 1999;399:230–237. doi: 10.1038/20375. This paper describes the crystal structure of full-length karyopherin β2 (transportin) bound to RanGTP. In the complex, RanGTP shows extensive structural rearrangements compared with the GDP-bound form. [DOI] [PubMed] [Google Scholar]
- Vetter IR, Arndt A, Kutay U, Görlich D, Wittinghofer A. Structural view of the Ran-importin beta interaction at 2.3 A resolution. Cell. 1999;97:635–646. doi: 10.1016/s0092-8674(00)80774-6. This paper reports the three-dimensional structure of a complex between Ran bound to GTP and an amino-terminal 462-residue fragment of importin β. [DOI] [PubMed] [Google Scholar]
- Bayliss R, Littlewood T, Stewart M. Structural basis for the interaction between FxFG nucleoporin repeats and importin-beta in nuclear trafficking. Cell. 2000;102:99–108. doi: 10.1016/s0092-8674(00)00014-3. The authors describe the crystal structure of a complex formed between residues 1-442 of importin β and the FxFG repeats from the nucleoporin Nsp1. From this information they engineer a point mutation in importin β that impairs binding to FxFG nucleoporins and decreases nuclear import in vitro. [DOI] [PubMed] [Google Scholar]
- Lee SJ, Imamoto N, Sakai H, Nakagawa A, Kose S, Koike M, Yamamoto M, Kumasaka T, Yoneda Y, Tsukihara T. The adoption of a twisted structure of importin-beta is essential for the protein-protein interaction required for nuclear transport. J Mol Biol. 2000;302:251–264. doi: 10.1006/jmbi.2000.4055. The authors report the crystal structure of the free amino-terminal fragment of mouse importin β. Structural analysis reveals flexible movement and rearrangements of importin β. [DOI] [PubMed] [Google Scholar]
- Groves MR, Hanlon N, Turowski P, Hemmings BA, Barford D. The structure of the protein phosphatase 2A PR65/A subunit reveals the conformation of its 15 tandemly repeated HEAT motifs. Cell. 1999;96:99–110. doi: 10.1016/s0092-8674(00)80963-0. The authors report the crystal structure of the human PR65/A subunit, revealing a left-handed superhelical conformation distinct from the right-handed superhelical fold found in importin β. [DOI] [PubMed] [Google Scholar]
- Nakielny S, Dreyfuss G. Transport of proteins and RNAs in and out of the nucleus. Cell. 1999;99:677–690. doi: 10.1016/s0092-8674(00)81666-9. This comprehensive review covers a large body of information, including details on cargoes, transporters and models of translocation through the NPC. [DOI] [PubMed] [Google Scholar]
- Radu A, Moore MS, Blobel G. The peptide repeat domain of nucleoporin Nup98 functions as a docking site in transport across the nuclear pore complex. Cell. 1995;81:215–222. doi: 10.1016/0092-8674(95)90331-3. The authors report the characterization of the nucleoporin Nup98, which is located at the nucleoplasmic side of the NPC, and map an importin-β-docking site at the amino-terminal half of Nup98. [DOI] [PubMed] [Google Scholar]
- Rexach M, Blobel G. Protein import into nuclei: association and dissociation reactions involving transport substrate, transport factors, and nucleoporins. Cell. 1995;83:683–692. doi: 10.1016/0092-8674(95)90181-7. Using a solution binding assay the authors characterize the RanGTP-induced dissociation of importin α and β and analyze the dynamics of interactions between nucleoporins and soluble transport factors bound to their cargoes. [DOI] [PubMed] [Google Scholar]
- Shah S, Tugendreich S, Forbes D. Major binding sites for the nuclear import receptor are the internal nucleoporin Nup153 and the adjacent nuclear filament protein Tpr. J Cell Biol. 1998;141:31–49. doi: 10.1083/jcb.141.1.31. This work shows that importin β co-purifies with Nup153 and Tpr, but not with p62, Nup93, Nup98 or Nup214. Importin β is shown to bind Nup153 directly. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kehlenbach RH, Dickmanns A, Kehlenbach A, Guan T, Gerace L. A role for RanBP1 in the release of CRM1 from the nuclear pore complex in a terminal step of nuclear export. J Cell Biol. 1999;145:645–657. doi: 10.1083/jcb.145.4.645. An in vitro assay is used to identify cytosolic factors required for Crm1-mediated protein export. Reconstitution experiments demonstrate that RanBP1 and the homologous domains of Nup358/RanBP2 are capable of dissociating Crm1 from the NPC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seedorf M, Damelin M, Kahana J, Taura T, Silver PA. Interactions between a nuclear transporter and a subset of nuclear pore complex proteins depend on Ran GTPase. Mol Cell Biol. 1999;19:1547–1557. doi: 10.1128/mcb.19.2.1547. Immunoprecipitation of several nuclear receptor-GFP fusions from yeast extracts show an overlapping but not identical pattern of interactions with different Nups. Pse1p-Nup interactions are shown to be dependent on the nucleotide-bound state of Ran. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Damelin M, Silver PA. Mapping interactions between nuclear transport factors in living cells reveals pathways through the nuclear pore complex. Mol Cell. 2000;5:133–140. doi: 10.1016/s1097-2765(00)80409-8. Interactions between Nups and importins are investigated in vivo using fluorescence resonance energy transfer (FRET) in S. cerevisiae. Different variants of GFP fused to Nups and the transport factors Pse1p/Kap121p or Msn5p reveal differences in the translocation pathways. [DOI] [PubMed] [Google Scholar]
- Dingwall C, Laskey RA. Nuclear targeting sequences - a consensus? Trends Biochem Sci. 1991;16:478–481. doi: 10.1016/0968-0004(91)90184-w. An overview of nuclear targeting sequences. This review provides the first description of a consensus for a bipartite NLS motif. [DOI] [PubMed] [Google Scholar]
- Adam EJ, Adam SA. Identification of cytosolic factors required for nuclear location sequence-mediated binding to the nuclear envelope. J Cell Biol. 1994;125:547–555. doi: 10.1083/jcb.125.3.547. The authors describe the purification of a factor that is required to reconstitute the first step in nuclear transport, binding to the pore complex. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Adam SA, Gerace L. Cytosolic proteins that specifically bind nuclear location signals are receptors for nuclear import. Cell. 1991;66:837–847. doi: 10.1016/0092-8674(91)90431-w. Two polypeptides of 54 and 56 kDa were purified from bovine erythrocytes and found to bind specifically to the NLS of SV40 large T antigen. [DOI] [PubMed] [Google Scholar]
- Görlich D, Vogel F, Mills AD, Hartmann E, Laskey RA. Distinct functions for the two importin subunits in nuclear protein import. Nature. 1995;377:246–248. doi: 10.1038/377246a0. An importin α-importin β-import substrate complex is shown to dock to the NPC via importin β. Then, Ran-mediated translocation through the pore results in the accumulation of import substrate and importin α in the nucleus. In contrast, importin β was found to accumulate at the nuclear envelope, not in the nucleoplasm. [DOI] [PubMed] [Google Scholar]
- Imamoto N, Shimamoto T, Kose S, Takao T, Tachibana T, Matsubae M, Sekimoto T, Shimonishi Y, Yoneda Y. The nuclear pore-targeting complex binds to nuclear pores after association with a karyophile. FEBS Lett. 1995;368:415–419. doi: 10.1016/0014-5793(95)00699-a. In this study, the authors cloned a cDNA encoding a 97 kDa protein of the nuclear pore-targeting complex (PTAC; identical to importin β). PTAC97 was found to reconstitute the nuclear-binding step in conjunction with a 58 kDa component of PTAC (PTAC58/importin α). [DOI] [PubMed] [Google Scholar]
- Weis K, Mattaj IW, Lamond AI. Identification of hSRP1 alpha as a functional receptor for nuclear localization sequences. Science. 1995;268:1049–1053. doi: 10.1126/science.7754385. This work describes hSRP1 alpha (importin α), which binds specifically to proteins containing either a simple or bipartite NLS motif. The hSRP1 alpha protein is shown to promote docking of import substrates to the pore and, together with Ran, to reconstitute nuclear protein import. [DOI] [PubMed] [Google Scholar]
- Huber J, Cronshagen U, Kadokura M, Marshallsay C, Wada T, Sekine M, Luhrmann R. Snurportin1, an m3G-cap-specific nuclear import receptor with a novel domain structure. EMBO J. 1998;17:4114–4126. doi: 10.1093/emboj/17.14.4114. Snurportin1 was identifed and found to function as an snRNP import receptor by binding m3G-cap-containing RNAs. Snurportin1 enhances cap-dependent nuclear import of U snRNPs and functions as an adaptor-like protein that interacts with importin β through an IBB domain. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jullien D, Görlich D, Laemmli UK, Adachi Y. Nuclear import of RPA in Xenopus egg extracts requires a novel protein XRI-Palpha but not importin alpha. EMBO J. 1999;18:4348–4358. doi: 10.1093/emboj/18.15.4348. By using a yeast two-hybrid screen, the authors identified XRIPα, which interacts with the largest subunit RPA and is required for its nuclear import. This work suggests that XRIPα serves as an adaptor to link RPA to importin β. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakel S, Albig W, Kutay U, Bischoff FR, Schwamborn K, Doenecke D, Görlich D. The importin beta/importin 7 heterodimer is a functional nuclear import receptor for histone H1. EMBO J. 1999;18:2411–2423. doi: 10.1093/emboj/18.9.2411. Importin β and importin 7 (Imp7, RanBP7) are shown to play a critical role in nuclear import of the linker histone H1. Individually, the import receptors bind H1 weakly, but binding is strong for the heterodimer. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiganis T, Flint AJ, Adam SA, Tonks NK. Association of the T-cell protein tyrosine phosphatase with nuclear import factor p97. J Biol Chem. 1997;272:21548–21557. doi: 10.1074/jbc.272.34.21548. The nuclear-import factor p97 (importin β) is shown to bind directly to T-cell protein tyrosine phosphatase (TCPTP) and, in a permeabilized cell assay, was found to be necessary for its nuclear import. [DOI] [PubMed] [Google Scholar]
- Truant R, Cullen BR. The arginine-rich domains present in human immunodeficiency virus type 1 Tat and Rev function as direct importin beta-dependent nuclear localization signals. Mol Cell Biol. 1999;19:1210–1217. doi: 10.1128/mcb.19.2.1210. This study defines a novel class of arginine-rich NLSs that are direct targets for importin β and function independently of importin α. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palmeri D, Malim MH. Importin beta can mediate the nuclear import of an arginine-rich nuclear localization signal in the absence of importin alpha. Mol Cell Biol. 1999;19:1218–1225. doi: 10.1128/mcb.19.2.1218. The arginine-rich NLS of the Rex protein of human T-cell leukemia virus type 1 is shown to use importin β for import but does so via a mechanism which is importin-α-independent. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakel S, Görlich D. Importin beta, transportin, RanBP5 and RanBP7 mediate nuclear import of ribosomal proteins in mammalian cells. EMBO J. 1998;17:4491–4502. doi: 10.1093/emboj/17.15.4491. Nuclear import of ribosomal proteins is mediated by several importin-β-like transport factors, which function in the absence of importin α. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moore JD, Yang J, Truant R, Kornbluth S. Nuclear import of Cdk/cyclin complexes: identification of distinct mechanisms for import of Cdk2/cyclin E and Cdc2/cyclin B1. J Cell Biol. 1999;144:213–224. doi: 10.1083/jcb.144.2.213. Cyclin E and cyclin B1 are demonstrated to be imported into nuclei via distinct mechanisms. Cyclin E behaves like a classical NLS-containing protein and binds to importin α, wheras cyclin B1 is imported into the nucleus via a direct interaction with importin β. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takizawa CG, Weis K, Morgan DO. Ran-independent nuclear import of cyclin B1-Cdc2 by importin beta. Proc Natl Acad Sci USA. 1999;96:7938–7943. doi: 10.1073/pnas.96.14.7938. Importin β promotes cyclin B1 import in the absence of cytosol or Ran and in the presence of a dominant-negative Ran mutant. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Xiao Z, Liu X, Lodish HF. Importin beta mediates nuclear translocation of Smad 3. J Biol Chem. 2000;275:23425–23428. doi: 10.1074/jbc.C000345200. Smad 3 binds directly to importin β. [DOI] [PubMed] [Google Scholar]
- Lam MH, Briggs LJ, Hu W, Martin TJ, Gillespie MT, Jans DA. Importin beta recognizes parathyroid hormone-related protein with high affinity and mediates its nuclear import in the absence of importin alpha. J Biol Chem. 1999;274:7391–7398. doi: 10.1074/jbc.274.11.7391. Importin β mediates nuclear import of parathyroid-hormone-related protein in the absence of importin α. [DOI] [PubMed] [Google Scholar]
- Rout MP, Blobel G, Aitchison JD. A distinct nuclear import pathway used by ribosomal proteins. Cell. 1997;89:715–725. doi: 10.1016/s0092-8674(00)80254-8. The previously uncharacterized yeast beta karyopherin Kap123p is shown to be involved in the import of ribosomal proteins into the nucleus, and the related protein Pse1p has a partially redundant function in that pathway. [DOI] [PubMed] [Google Scholar]
- Schlenstedt G, Smirnova E, Deane R, Solsbacher J, Kutay U, Görlich D, Ponstingl H, Bischoff FR. Yrb4p, a yeast ran-GTP-binding protein involved in import of ribosomal protein L25 into the nucleus. EMBO J. 1997;16:6237–6249. doi: 10.1093/emboj/16.20.6237. The identification of Yrb4p (Kap123p), a protein related to importin β. Cells disrupted for YRB4 were found to be defective in nuclear import of ribosomal protein L25. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Aitchison JD, Blobel G, Rout MP. Kap104p: a karyopherin involved in the nuclear transport of messenger RNA binding proteins. Science. 1996;274:624–627. doi: 10.1126/science.274.5287.624. The yeast karyopherin Kap104p was found to mediate nuclear import of two mRNA-binding proteins, Nab2p and Nab4p. [DOI] [PubMed] [Google Scholar]
- Pollard VW, Michael WM, Nakielny S, Siomi MC, Wang F, Dreyfuss G. A novel receptor-mediated nuclear protein import pathway. Cell. 1996;86:985–994. doi: 10.1016/s0092-8674(00)80173-7. The authors identified a 90 kDa protein, transportin, which is related to importin β. Transportin was found to mediate nuclear import of proteins containing an M9-type NLS. [DOI] [PubMed] [Google Scholar]
- Fridell RA, Truant R, Thorne L, Benson RE, Cullen BR. Nuclear import of hnRNP A1 is mediated by a novel cellular cofactor related to karyopherin-beta. J Cell Sci. 1997;110:1325–1331. doi: 10.1242/jcs.110.11.1325. The identification of a human protein related to importin β, termed MIP (transportin), which binds the M9 sequence of hnRNP A1. [DOI] [PubMed] [Google Scholar]
- Bonifaci N, Moroianu J, Radu A, Blobel G. Karyopherin beta2 mediates nuclear import of a mRNA binding protein. Proc Natl Acad Sci USA. 1997;94:5055–5060. doi: 10.1073/pnas.94.10.5055. The authors cloned and sequenced the cDNA for human karyopherin β2 (transportin). Karyopherin β2/transportin binds directly to hnRNP A1 and is able to dock A1 at the pore as well as importing it into the nucleus. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siomi MC, Eder PS, Kataoka N, Wan L, Liu Q, Dreyfuss G. Transportin-mediated nuclear import of heterogeneous nuclear RNP proteins. J Cell Biol. 1997;138:1181–1192. doi: 10.1083/jcb.138.6.1181. Transportin 1 is shown to interact with, and mediate import of, hnRNPs. The authors also identify transportin 2, a transportin 1 homolog. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kataoka N, Bachorik JL, Dreyfuss G. Transportin-SR, a nuclear import receptor for SR proteins. J Cell Biol. 1999;145:1145–1152. doi: 10.1083/jcb.145.6.1145. Describes a novel import receptor, transportin-SR, which binds directly to SR proteins, a group of abundant arginine/serine-rich proteins that include ASF/SF2 and SC35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lai MC, Lin RI, Huang SY, Tsai CW, Tarn WY. A human importin-beta family protein, transportin-SR2, interacts with the phosphorylated RS domain of SR proteins. J Biol Chem. 2000;275:7950–7957. doi: 10.1074/jbc.275.11.7950. By using the human papillomavirus E2 activator, which contains an arginine/serine-rich domain, as a bait in a yeast two-hybrid screen the authors identified transportin-SR2, a new importin β family member. [DOI] [PubMed] [Google Scholar]
- Pemberton LF, Rosenblum JS, Blobel G. Nuclear import of the TATA-binding protein: mediation by the karyopherin Kap114p and a possible mechanism for intranuclear targeting. J Cell Biol. 1999;145:1407–1417. doi: 10.1083/jcb.145.7.1407. Kap114p was identified as the import receptor for TATA-binding protein. The authors show that the RanGTP-mediated dissociation of TATA-binding protein from Kap114p is stimulated by TATA-containing DNA and TFIIA. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fornerod M, Ohno M, Yoshida M, Mattaj IW. CRM1 is an export receptor for leucine-rich nuclear export signals. Cell. 1997;90:1051–1060. doi: 10.1016/s0092-8674(00)80371-2. Xenopus Crm1 binds cooperatively to an NES and RanGTP, and Crm1 mediates the nuclear export of NES-containing proteins. [DOI] [PubMed] [Google Scholar]
- Ossareh-Nazari B, Bachelerie F, Dargemont C. Evidence for a role of CRM1 in signal-mediated nuclear protein export. Science. 1997;278:141–144. doi: 10.1126/science.278.5335.141. Interactions between human Crm1 and an NES-containing protein, were studied in reticulocyte lysates. An export assay using permeabilized cells was used to characterize the export of NES-containing proteins. [DOI] [PubMed] [Google Scholar]
- Stade K, Ford CS, Guthrie C, Weis K. Exportin 1 (Crm1p) is an essential nuclear export factor. Cell. 1997;90:1041–1050. doi: 10.1016/s0092-8674(00)80370-0. Identification of yeast Crm1p/Xpo1p as an NES export receptor using an in vivo export assay with a shuttling NES/NLS-green fluorescence protein reporter. [DOI] [PubMed] [Google Scholar]
- Fukuda M, Asano S, Nakamura T, Adachi M, Yoshida M, Yanagida M, Nishida E. CRM1 is responsible for intracellular transport mediated by the nuclear export signal. Nature. 1997;390:308–311. doi: 10.1038/36894. The cytotoxin leptomycin Bis shown to be an inhibitor of NES-dependent nuclear export of proteins. In Xenopus oocyte extracts, a protein of 110 kDa that binds to the NES was found and characterized as Crm1. [DOI] [PubMed] [Google Scholar]
- Sachdev S, Bagchi S, Zhang DD, Mings AC, Hannink M. Nuclear import of IkappaBalpha is accomplished by a ran-independent transport pathway. Mol Cell Biol. 2000;20:1571–1582. doi: 10.1128/mcb.20.5.1571-1582.2000. The authors provide mechanistic insight into nuclear shuttling of IκBα. Nuclear import of IκBα occurs via a Ran-independent mechanism, while nuclear export was found to require the Ran-dependent Crm1 nuclear export receptor. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paraskeva E, Izaurralde E, Bischoff FR, Huber J, Kutay U, Hartmann E, Luhrmann R, Görlich D. CRM1-mediated recycling of snurportin 1 to the cytoplasm. J Cell Biol. 1999;145:255–264. doi: 10.1083/jcb.145.2.255. Re-export of snurportin 1 is mediated via Crm1. Snurportin 1 was found to be a potent competitor of other Crm1-dependent export pathways and is shown to bind to Crm1 with a high affinity. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hakata Y, Umemoto T, Matsushita S, Shida H. Involvement of human CRM1 (exportin 1) in the export and multimerization of the Rex protein of human T-cell leukemia virus type 1. J Virol. 1998;72:6602–6607. doi: 10.1128/jvi.72.8.6602-6607.1998. The role of human exportin 1 in the export of Rex encoded by human T-cell leukemia virus type 1 is described. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohno M, Segref A, Bachi A, Wilm M, Mattaj IW. PHAX, a mediator of U snRNA nuclear export whose activity is regulated by phosphorylation. Cell. 2000;101:187–198. doi: 10.1016/S0092-8674(00)80829-6. U snRNA export requires an additional protein, named PHAX, which acts as an adaptor between the CBC-RNA complex and the CRM1-RanGTP proteins. [DOI] [PubMed] [Google Scholar]
- Yang J, Bardes ES, Moore JD, Brennan J, Powers MA, Kornbluth S. Control of cyclin B1 localization through regulated binding of the nuclear export factor CRM1. Genes Dev. 1998;12:2131–2143. doi: 10.1101/gad.12.14.2131. The binding of Crm1 to the NES of cyclin B1 is demonstrated; phos-phorylation within the NES inhibits the interaction with Crm1. This suggests that cyclin B1 localization is controlled by phosphorylation. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhu J, McKeon F. NF-AT activation requires suppression of Crm1-dependent export by calcineurin. Nature. 1999;398:256–260. doi: 10.1038/18473. The authors show that nuclear import of nuclear factor of activated T cells (NF-AT) is not sufficient to activate NF-AT target genes, and that NF-AT is exported to the cytoplasm using the Crm1 export pathway. [DOI] [PubMed] [Google Scholar]
- Yan C, Lee LH, Davis LI. Crm1p mediates regulated nuclear export of a yeast AP-1-like transcription factor. EMBO J. 1998;17:7416–7429. doi: 10.1093/emboj/17.24.7416. Characterization of Yap1p as a target of Crm1-mediated export. Recognition of Yap1p by Crm1p is inhibited by oxidation, and this inhibition requires cysteine residues flanking the NES. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jensen TH, Neville M, Rain JC, McCarthy T, Legrain P, Rosbash M. Identification of novel Saccharomyces cerevisiae proteins with nuclear export activity: cell cycle-regulated transcription factor Ace2p shows cell cycle-independent nucleocytoplasmic shuttling. Mol Cell Biol. 2000;20:8047–8058. doi: 10.1128/mcb.20.21.8047-8058.2000. The results of a yeast two-hybrid screen using S. cerevisiae Crm1p as a bait are described. New proteins with export activity (named Cip1p to Cip3p) were found. Cip3p is the cell-cycle-regulated transcription factor Ace2p, which contains both an NES and an NLS activity and enters the nucleus at all stages of the cell cycle. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrigno P, Posas F, Koepp D, Saito H, Silver PA. Regulated nucleo/cytoplasmic exchange of HOG1 MAPK requires the importin beta homologs NMD5 and XPO1. EMBO J. 1998;17:5606–5614. doi: 10.1093/emboj/17.19.5606. The localization of the osmotic stress-response MAP kinase Hog1p is controlled by Nmd5p and Crm1p. Hog1p phosphorylation triggers its nuclear import, whereas dephosphorylation is likely to promote its export via Crm1p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shulga N, James P, Craig EA, Goldfarb DS. A nuclear export signal prevents Saccharomyces cerevisiae Hsp70 Ssb1p from stimulating nuclear localization signal-directed nuclear transport. J Biol Chem. 1999;274:16501–16507. doi: 10.1074/jbc.274.23.16501. Demonstrates a leucine-rich NES in the carboxy-terminal domain of Ssb1p and shows that it is responsible for both the different subcellular localizations of Ssa1p and Ssb1p and for their differential function in NLS-directed nuclear transport. [DOI] [PubMed] [Google Scholar]
- Kutay U, Bischoff FR, Kostka S, Kraft R, Görlich D. Export of importin alpha from the nucleus is mediated by a specific nuclear transport factor. Cell. 1997;90:1061–1071. doi: 10.1016/s0092-8674(00)80372-4. CAS binds to importin α in a RanGTP-dependent manner and functions as an exportin for importin α. [DOI] [PubMed] [Google Scholar]
- Hood JK, Silver PA. Cse1p is required for export of Srp1p/importin-alpha from the nucleus in Saccharomyces cerevisiae. J Biol Chem. 1998;273:35142–35146. doi: 10.1074/jbc.273.52.35142. Cse1p is required for export of Srp1p (importin α) from the nucleus. [DOI] [PubMed] [Google Scholar]
- Kunzler M, Hurt EC. Cse1p functions as the nuclear export receptor for importin alpha in yeast. FEBS Lett. 1998;433:185–190. doi: 10.1016/s0014-5793(98)00892-8. Cse1p, Ran and importin a (Srp1p) interact in a yeast two-hybrid system and the recombinant proteins form a trimeric complex in vitro. [DOI] [PubMed] [Google Scholar]
- Solsbacher J, Maurer P, Bischoff FR, Schlenstedt G. Cse1p is involved in export of yeast importin alpha from the nucleus. Mol Cell Biol. 1998;18:6805–6815. doi: 10.1128/mcb.18.11.6805. This paper characterizes the yeast homolog of CAS, which was previously identified as Cse1p. Cse1p forms a trimeric complex with Srp1p and RanGTP. Complex formation was prevented by NLS peptides, suggesting that importin α is exported only after cargo release. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaffman A, Rank NM, O'Neill EM, Huang LS, O'Shea EK. The receptor Msn5 exports the phosphorylated transcription factor Pho4 out of the nucleus. Nature. 1998;396:482–486. doi: 10.1038/24898. Nuclear export of the yeast transcription factor Pho4p is mediated by the exportin Msn5p. This is the first example of an NES regulated directly by phosphorylation. [DOI] [PubMed] [Google Scholar]
- Yoshida K, Blobel G. The karyopherin Kap142p/Msn5p mediates nuclear import and nuclear export of different cargo proteins. J Cell Biol. 2001;152:729–739. doi: 10.1083/jcb.152.4.729. Kap142p/Msn5p mediates nuclear export as well as nuclear import. The ssDNA-binding protein complex RPA was identified as an import cargo for Kap142p/Msn5p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutay U, Lipowsky G, Izaurralde E, Bischoff FR, Schwarzmaier P, Hartmann E, Görlich D. Identification of a tRNA-specific nuclear export receptor. Mol Cell. 1998;1:359–369. doi: 10.1016/s1097-2765(00)80036-2. Together with [67], this paper characterizes the exportin that is involved in export of t-RNA. See annotation to [67]. [DOI] [PubMed] [Google Scholar]
- Arts GJ, Fornerod M, Mattaj IW. Identification of a nuclear export receptor for tRNA. Curr Biol. 1998;8:305–314. doi: 10.1016/s0960-9822(98)70130-7. This paper identifies the function of exportin-t, which was previously identified as an uncharacterized member of the importin-β family. Exportin-t was found to stimulate the export of tRNA from microinjected Xenopus nuclei, to shuttle between the nucleus and cytoplasm and to bind tRNA in a RanGTP-dependent manner. Exportin-t is the first exportin to be shown to bind directly to a nucleic acid. [DOI] [PubMed] [Google Scholar]
- Hellmuth K, Lau DM, Bischoff FR, Kunzler M, Hurt E, Simos G. Yeast Los1p has properties of an exportin-like nucleocytoplasmic transport factor for tRNA. Mol Cell Biol. 1998;18:6374–6386. doi: 10.1128/mcb.18.11.6374. Los1p binds to nucleoporins and to RanGTP. Formation of the trimeric Los1p-Ran-GTP complex was stimulated in the presence of tRNA. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weis K. Importins and exportins: how to get in and out of the nucleus. Trends Biochem Sci. 1998;23:185–189. doi: 10.1016/s0968-0004(98)01204-3. A detailed review of nucleocytoplasmic transport and the individual transport pathways. [DOI] [PubMed] [Google Scholar]
- Nachury MV, Weis K. The direction of transport through the nuclear pore can be inverted. Proc Natl Acad Sci USA. 1999;96:9622–9627. doi: 10.1073/pnas.96.17.9622. The directionality of nuclear transport is dependent on the compartmentalized distribution of RanGTP and that the direction of transport can be inverted in vitro by cytoplasmic addition of a GTP-bound mutant Ran, RanQ69LGTP. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Izaurralde E, Kutay U, von Kobbe C, Mattaj IW, Görlich D. The asymmetric distribution of the constituents of the Ran system is essential for transport into and out of the nucleus. EMBO J. 1997;16:6535–6547. doi: 10.1093/emboj/16.21.6535. Using microinjection experiments, the authors demonstrate that the asymmetric distribution of RanGTP is crucial for many nucleocytoplasmic transport pathways. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bischoff FR, Ponstingl H. Catalysis of guanine nucleotide exchange on Ran by the mitotic regulator RCC1. Nature. 1991;354:80–82. doi: 10.1038/354080a0. RCC1 is indentified as the guanine-nucleotide exchange factor of Ran. [DOI] [PubMed] [Google Scholar]
- Bischoff FR, Klebe C, Kretschmer J, Wittinghofer A, Ponstingl H. RanGAP1 induces GTPase activity of nuclear Ras-related Ran. Proc Natl Acad Sci USA. 1994;91:2587–2591. doi: 10.1073/pnas.91.7.2587. The purification and identification of the Ran GTPase-activating protein, RanGAP1, a nuclear homodimeric 65 kDa protein. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bischoff FR, Krebber H, Smirnova E, Dong W, Ponstingl H. Co-activation of RanGTPase and inhibition of GTP dissociation by Ran-GTP binding protein RanBP1. EMBO J. 1995;14:705–715. doi: 10.1002/j.1460-2075.1995.tb07049.x. The authors characterize RanBP1, which binds tightly to RanGTP. RanBP1 does not activate the RanGTPase alone, but cooperates with RanGAP1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu J, Matunis MJ, Kraemer D, Blobel G, Coutavas E. Nup358, a cytoplasmically exposed nucleoporin with peptide repeats, Ran-GTP binding sites, zinc fingers, a cyclophilin A homologous domain, and a leucine-rich region. J Biol Chem. 1995;270:14209–14213. doi: 10.1074/jbc.270.23.14209. The authors report the characterization of a NPC-localized 358 kDa Ran-binding protein, Nup358 (RanBP2). [DOI] [PubMed] [Google Scholar]
- Yokoyama N, Hayashi N, Seki N, Pante N, Ohba T, Nishii K, Kuma K, Hayashida T, Miyata T, Aebi U, et al. A giant nucleopore protein that binds Ran/TC4. Nature. 1995;376:184–188. doi: 10.1038/376184a0. The identification and characterization of a 3224-residue nuclear pore protein, RanBP2 (Nup358), that interacts with RanGTP. [DOI] [PubMed] [Google Scholar]
- Ben-Efraima I, Gerace L. Gradient of increasing affinity of importin β for nucleoporins along the pathway of nuclear import. J Cell Biol. 2001;152:411–417. doi: 10.1083/jcb.152.2.411. Using a solid-phase binding analysis, the affinity of an importin-β-cargo complex for different nucleoporins was determined. The results support a model in which importin β binds to nucleoporins with progressively increasing affinity as the import complex moves from the cytoplasmic to the central and the nucleoplasmic regions of the NPC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dostie J, Ferraiuolo M, Pause A, Adam SA, Sonenberg N. A novel shuttling protein, 4E-T, mediates the nuclear import of the mRNA 5' cap-binding protein, eIF4E. EMBO J. 2000;19:3142–3156. doi: 10.1093/emboj/19.12.3142. The cloning and the characterization of an eIF4E-binding protein, 4E-T (eIF4E-transporter). 4E-T was found to be a nucleocytoplasmic shuttling protein that targets eIF4E for nuclear import. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rosenblum JS, Pemberton LF, Blobel G. A nuclear import pathway for a protein involved in tRNA maturation. J Cell Biol. 1997;139:1655–1661. doi: 10.1083/jcb.139.7.1655. Demonstrates that the yeast La protein Lhp1p is specifically imported by the importin-β-like protein Kap108p/Sxm1p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Senger B, Simos G, Bischoff FR, Podtelejnikov A, Mann M, Hurt E. Mtr10p functions as a nuclear import receptor for the mRNA-binding protein Npl3p. EMBO J. 1998;17:2196–2207. doi: 10.1093/emboj/17.8.2196. MTR10 genetically interacts with NUP85 and is responsible for the nuclear import of Npl3p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albertini M, Pemberton LF, Rosenblum JS, Blobel G. A novel nuclear import pathway for the transcription factor TFIIS. J Cell Biol. 1998;143:1447–1455. doi: 10.1083/jcb.143.6.1447. The yeast protein Nmd5p/Kap119p functions as an importin for the transcription elongation factor TFIIS. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaffman A, Rank NM, O'Shea EK. Phosphorylation regulates association of the transcription factor Pho4 with its import receptor Pse1/Kap121. Genes Dev. 1998;12:2673–2683. doi: 10.1101/gad.12.17.2673. Pho4p is imported into the nucleus via Pse1p/Kap121p. The interaction between Pho4p and Pse1p is inhibited by phosphorylation, suggesting that phosphorylation of Pho4p regulates its import. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaves SR, Blobel G. Nuclear import of Spo12p, a protein essential for meiosis. J Biol Chem. 2001;276:17712–17717. doi: 10.1074/jbc.M010760200. Demonstrates that Spo12p is imported into the nucleus by the karyopherin Kap121p/Pse1p. [DOI] [PubMed] [Google Scholar]
- Titov AA, Blobel G. The karyopherin Kap122p/Pdr6p imports both subunits of the transcription factor IIA into the nucleus. J Cell Biol. 1999;147:235–246. doi: 10.1083/jcb.147.2.235. Characterization of Pdr6p/Kap122p. A complex of the large subunit (Toa1p) and the small subunit (Toa2p) of TFIIA was identified as an import substrate for Kap122p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lipowsky G, Bischoff FR, Schwarzmaier P, Kraft R, Kostka S, Hartmann E, Kutay U, Görlich D. Exportin 4: a mediator of a novel nuclear export pathway in higher eukaryotes. EMBO J. 2000;19:4362–4371. doi: 10.1093/emboj/19.16.4362. The identification of exportin 4 as an exportin for eIF-5A. The export signal in eIF-5A involves its unique hypusine modification. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DeVit MJ, Johnston M. The nuclear exportin Msn5p is required for nuclear export of the Mig1p glucose repressor of Saccharomyces cerevisiae. Curr Biol. 1999;9:1231–1241. doi: 10.1016/s0960-9822(99)80503-x. Demonstrates that Msn5, a member of the importin β family, is required to mediate the regulated export of Mig1 from the nucleus. [DOI] [PubMed] [Google Scholar]
- Blondel M, Alepuz PM, Huang LS, Shaham S, Ammerer G, Peter M. Nuclear export of Far1p in response to pheromones requires the export receptor Msn5p/Ste21p. Genes Dev. 1999;13:2284–2300. doi: 10.1101/gad.13.17.2284. Far1p is exported in an Msn5p-dependent manner in response to pheromone treatment. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mahanty SK, Wang Y, Farley FW, Elion EA. Nuclear shuttling of yeast scaffold Ste5 is required for its recruitment to the plasma membrane and activation of the mating MAPK cascade. Cell. 1999;98:501–512. doi: 10.1016/s0092-8674(00)81978-9. The authors show that Ste5p is a shuttling protein whose nuclear export is stimulated by pheromone. Ste5p export seems to be dependent on MSN5. [DOI] [PubMed] [Google Scholar]
- Görlich D, Dabrowski M, Bischoff FR, Kutay U, Bork P, Hartmann E, Prehn S, Izaurralde E. A novel class of RanGTP binding proteins. J Cell Biol. 1997;138:65–80. doi: 10.1083/jcb.138.1.65. This paper describes the novel superfamily of Ran-binding proteins that includes importin β. Two proteins are characterized in more detail, namely RanBP7 and RanBP8. Both resemble importin β in their interaction with Ran, and both bind directly to nuclear pore complexes. [DOI] [PMC free article] [PubMed] [Google Scholar]