

Short abstract
Using Release 3 of the euchromatic genomic sequence of Drosophila melanogaster, 85 known and eight novel families of transposable element have been identified, varying in copy number from one to 146. A total of 1,572 full and partial transposable elements were identified, comprising 3.86% of the sequence.
Abstract
Background
Transposable elements are found in the genomes of nearly all eukaryotes. The recent completion of the Release 3 euchromatic genomic sequence of Drosophila melanogaster by the Berkeley Drosophila Genome Project has provided precise sequence for the repetitive elements in the Drosophila euchromatin. We have used this genomic sequence to describe the euchromatic transposable elements in the sequenced strain of this species.
Results
We identified 85 known and eight novel families of transposable element varying in copy number from one to 146. A total of 1,572 full and partial transposable elements were identified, comprising 3.86% of the sequence. More than two-thirds of the transposable elements are partial. The density of transposable elements increases an average of 4.7 times in the centromere-proximal regions of each of the major chromosome arms. We found that transposable elements are preferentially found outside genes; only 436 of 1,572 transposable elements are contained within the 61.4 Mb of sequence that is annotated as being transcribed. A large proportion of transposable elements is found nested within other elements of the same or different classes. Lastly, an analysis of structural variation from different families reveals distinct patterns of deletion for elements belonging to different classes.
Conclusions
This analysis represents an initial characterization of the transposable elements in the Release 3 euchromatic genomic sequence of D. melanogaster for which comparison to the transposable elements of other organisms can begin to be made. These data have been made available on the Berkeley Drosophila Genome Project website for future analyses.
Background
Transposable element sequences are abundant yet poorly understood components of almost all eukaryotic genomes [1]. As a result, many biologists have an interest in the description of transposable elements in completely sequenced eukaryotic genomes. The evolutionary biologist wants to understand the origin of transposable elements, how they are lost and gained by a species and the role they play in the processes of genome evolution; the population geneticist wants to know the factors that determine the frequency and distribution of elements within and between populations; the developmental geneticist wants to know what roles these elements may play in either normal developmental processes or in the response of the organism to external conditions; finally, the molecular geneticist wants to know the mechanisms that regulate the transposition cycle of these elements and how they interact with the cellular machinery of the host. It is for all of these reasons and more that a description of the transposable elements in the recently completed Release 3 genomic sequence of D. melanogaster is desirable.
Our understanding of transposable elements owes much to research on Drosophila. Over 75 years ago, Milislav Demerec discovered highly mutable alleles of two genes in D. virilis, miniature and magenta ([2,3,4], reviewed in [5]). Both genes were mutable in soma and germline and, for the miniature-3α alleles, dominant enhancers of mutability were also isolated by Demerec. In retrospect, it seems clear that the mutability of these alleles was the result of transposition of mobile elements. The dominant enhancers may have been particularly active elements or mutations in host genes that affect transposability (see below). There matters stood until McClintock's analysis of the Ac and Ds factors in maize, which led to the discovery of transposition [6] and the discovery of insertion elements in the gal operon of Escherichia coli (see [7]).
Green [8] synthesized the available evidence to make a strong case for insertion as a mechanism of mutagenesis in Drosophila. Concurrently, Hogness' group had begun a molecular characterization of two elements in D. melanogaster, 412 and copia [9,10] and provided evidence that they were transposable [11,12,13]. Glover [14] unknowingly characterized the first eukaryotic transposable element at the molecular level, the insertion sequences of 28 S rRNA genes. The discovery of male recombination [15], and two systems of hybrid dysgenesis in D. melanogaster (see [16,17]) bridged the gap between genetic and molecular analyses. The discovery of the transposable elements that cause hybrid dysgenesis, the P element [18] and the I element [19], led to the first genomic analyses of transposable elements in a eukaryote.
The publication of the Release 1 genomic sequence in March 2000 [20] and the Release 2 genomic sequence in October 2000 encouraged several studies on the genomic distribution and abundance of transposable elements in D. melanogaster [21,22,23,24,25]. Unfortunately, neither release was suitable for rigorous analysis of its transposable elements. In the whole-genome shotgun assembly process, repetitive sequences (including transposable elements) were masked by the SCREENER algorithm and remained as gaps between unitigs [26]. During the repeat-resolution phase of the whole-genome assembly, an attempt was made to fill these gaps. However, comparisons of small regions sequenced by the clone-by-clone approach versus the whole-genome shotgun method show that this process did not produce accurate sequences for transposable elements [26,27]. These results demonstrate that rigorous analyses of the transposable elements, or any other repetitive sequence, requires a sequence of higher quality, now publicly available as Release 3 [28]. For the first time, the nature, number and location of the transposable elements can reliably be analyzed in the euchromatin of D. melanogaster.
Results and discussion
Identification of known and novel transposable elements
Eukaryotic transposable elements are divided into those that transpose via an RNA intermediate, the retrotransposons (class I elements), and those that transpose by DNA excision and repair, the transposons (class II elements [1]). Within the retrotransposons, the major division is between those that possess long terminal repeats (LTR elements) and those that do not (LINE and SINE elements [29]). Among the transposons, the majority transpose via a DNA intermediate, encode their own transposase and are flanked by relatively short terminally inverted repeat structures (TIR elements). Foldback (FB) elements, which are characterized by their property of reannealing after denaturation with zero-order kinetics, are quite distinct from prototypical class I or II elements, and have been included in our analyses [30]. Other classes of repetitive elements, such as DINE-1 [31,32,33], which are structurally distinct from all other classes, have not been included in this study.
We used a criterion of greater than 90% identity over more than 50 base-pairs (bp) of sequence to assign individual elements to families (see Materials and methods for details; a classification is shown in the additional data available (see Additional data files)). Subsequently, in order to ensure proper inclusion of elements in appropriate families, we generated multiple alignments for all families of transposable element represented by multiple copies. This allowed us to identify and remove spurious hits to highly repetitive regions of the genome, and it also enabled us to distinguish sequences of closely related families that share extensive regions of similarity.
A summary by class of the total number of complete and partial transposable elements in the Release 3 Drosophila euchromatic sequence is presented in Table 1, and detailed results for individual families of transposable element are listed in Table 2. Including those described here, there are 96 known families of transposable elements in D. melanogaster: 49 LTR families, 27 LINE-like families, 19 TIR families and the FB family. We have identified 1,572 full or partial elements from 93 of these 96 families (Table 1). In total, 3.86% (4.5 Mb) of the Release 3 sequence is composed of transposable elements. Previous analysis of both the euchromatic and heterochromatic sequences has suggested that 9% of the Drosophila genome is composed of repetitive elements [34]. One reason for this difference may be that the proportion of transposable element sequences in heterochromatic regions is higher than the genomic average [22,35].
Table 1.
An overview of the numbers of transposable elements in the euchromatic genome of D. melanogaster
| Class | Arm | Total transposable element sequence (in bp) | % of arm | Total number of transposable elements | Number full length | % Full length | Number of transposable elements perMb in genome | Number of transposable elements perMb in proximal 2 Mb |
| All families | X | 828,370 | 3.80 | 276 | 83 | 30.43 | 12.67 | 50 |
| 2L | 878,471 | 3.95 | 305 | 100 | 32.79 | 13.73 | 58.5 | |
| 2R | 870,914 | 4.29 | 313 | 84 | 26.84 | 15.42 | 89 | |
| 3L | 938,947 | 4.02 | 288 | 100 | 34.72 | 12.33 | 66.5 | |
| 3R | 866,971 | 3.11 | 288 | 102 | 35.76 | 10.33 | 24.5 | |
| 4 | 127,874 | 10.33 | 102 | 9 | 8.82 | 82.40 | - | |
| Total | 4,511,547 | 1,572 | 478 | |||||
| Average | 3.86 | 30.53 | 13.46 | 57.70 | ||||
| LTR | X | 628,924 | 2.89 | 134 | 54 | 41.04 | 6.15 | 19.00 |
| 2L | 603,536 | 2.72 | 127 | 67 | 52.76 | 5.72 | 20.00 | |
| 2R | 573,034 | 2.82 | 140 | 54 | 38.57 | 6.90 | 30.50 | |
| 3L | 618,441 | 2.65 | 117 | 58 | 49.57 | 5.01 | 24.00 | |
| 3R | 621,272 | 2.23 | 154 | 67 | 44.16 | 5.52 | 7.50 | |
| 4 | 44,121 | 3.56 | 10 | 4 | 40.00 | 8.08 | - | |
| Total | 3,089,328 | 682 | 304 | |||||
| Average | 2.65 | 44.87 | 5.84 | 20.20 | ||||
| LINE-like | X | 136,348 | 0.63 | 71 | 18 | 25.35 | 3.26 | 14.50 |
| 2L | 185,499 | 0.83 | 98 | 20 | 20.41 | 4.41 | 18.50 | |
| 2R | 225,984 | 1.11 | 109 | 18 | 16.51 | 5.37 | 37.50 | |
| 3L | 251,077 | 1.08 | 106 | 27 | 25.47 | 4.54 | 24.00 | |
| 3R | 176,355 | 0.63 | 70 | 19 | 27.14 | 2.51 | 4.50 | |
| 4 | 37,399 | 3.02 | 32 | 1 | 3.12 | 25.85 | - | |
| Total | 1,012,662 | 486 | 103 | |||||
| Average | 0.87 | 21.19 | 4.16 | 19.80 | ||||
| TIR | X | 45,324 | 0.21 | 59 | 7 | 11.86 | 2.71 | 14.50 |
| 2L | 82,761 | 0.37 | 76 | 11 | 14.47 | 3.42 | 18.00 | |
| 2R | 69,291 | 0.34 | 62 | 11 | 17.74 | 3.05 | 20.50 | |
| 3L | 52,743 | 0.23 | 57 | 12 | 21.05 | 2.44 | 16.50 | |
| 3R | 63,359 | 0.23 | 60 | 14 | 23.33 | 2.15 | 12.50 | |
| 4 | 44,195 | 3.57 | 58 | 3 | 5.17 | 46.85 | - | |
| Total | 357,673 | 372 | 58 | |||||
| Average | 0.31 | 15.59 | 3.19 | 16.40 | ||||
| FB | X | 17,774 | 0.08 | 12 | 4 | 33.33 | 0.55 | 2.00 |
| 2L | 6,675 | 0.03 | 4 | 2 | 50.00 | 0.18 | 2.00 | |
| 2R | 2,605 | 0.01 | 2 | 1 | 50.00 | 0.1 | 0.50 | |
| 3L | 16,686 | 0.07 | 8 | 3 | 37.50 | 0.34 | 2.00 | |
| 3R | 5,985 | 0.02 | 4 | 2 | 50.00 | 0.14 | 0.00 | |
| 4 | 2,159 | 0.17 | 2 | 1 | 50.00 | 1.62 | - | |
| Total | 51,884 | 32 | 13 | |||||
| Average | 0.04 | 40.62 | 0.27 | 1.30 |
For each class, the total numbers of each family of element, together with the numbers (and percentage of elements) that are full length is given for each chromosome arm. Column 3 gives the total base pairs contained within transposable elements, column 4 the percentage of each chromosome arm composed of transposable element sequences, column 8 the number of elements perMb, and column 9 the numbers of elements within the most proximal 2 Mb of each of the five major chromosome arms. Differences in density and amount of transposable element sequence were tested by binning major chromosomal arms into 50-kb windows and testing significance by Mann-Whitney U tests. The only significant difference (p < 0.05) observed, either including or excluding the proximal 2 Mb, was an increase in density and amount of sequence on the X chromosome relative to 3R.
Table 2.
The transposable elements of D. melanogaster
| Class | Family | Canonical length | X | 2L | 2R | 3L | 3R | 4 | Total number | Number full length | Number partial | Number in proximal 2 Mb | Average pairwise distance |
| LTR | 17.6 | 7439 | 2 | 0 | 3 | 5 | 2 | 0 | 12 | 7 | 5 | 4 | 0.006 |
| 1731 | 4648 | 1 | 0 | 0 | 0 | 1 | 0 | 2 | 1 | 1 | 1 | 0.000 | |
| 297 | 6995 | 22 | 12 | 6 | 7 | 10 | 0 | 57 | 18 | 39 | 12 | 0.032 | |
| 3S18 | 6126 | 4 | 0 | 1 | 1 | 0 | 0 | 6 | 4 | 2 | 3 | 0.075 | |
| 412 | 7566 | 8 | 0 | 7 | 11 | 5 | 0 | 31 | 24 | 7 | 6 | 0.024 | |
| accord | 7404 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | - | |
| aurora | 4263 | 0 | 0 | 2 | 1 | 0 | 0 | 3 | 1 | 2 | 3 | 0.074 | |
| blastopia | 5034 | 5 | 2 | 7 | 1 | 2 | 0 | 17 | 13 | 4 | 4 | 0.016 | |
| blood | 7410 | 1 | 11 | 2 | 3 | 5 | 0 | 22 | 22 | 0 | 6 | 0.001 | |
| Burdock | 6411 | 2 | 4 | 4 | 0 | 3 | 0 | 13 | 7 | 6 | 4 | 0.002 | |
| Circe | 6356 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 0 | 2 | 2 | 0.057 | |
| copia | 5143 | 4 | 13 | 4 | 5 | 4 | 0 | 30 | 26 | 4 | 3 | 0.002 | |
| diver | 6112 | 1 | 1 | 3 | 1 | 3 | 0 | 9 | 9 | 0 | 1 | 0.002 | |
| diver2 | 4917 | 0 | 4 | 3 | 2 | 0 | 0 | 9 | 0 | 9 | 9 | 0.032 | |
| Dm88 | 4558 | 0 | 0 | 2 | 0 | 30 | 0 | 32 | 0 | 32 | 2 | 0.015 | |
| frogger | 2483 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | - | |
| GATE | 8507 | 1 | 0 | 16 | 0 | 0 | 3 | 20 | 0 | 20 | 17 | 0.077 | |
| gtwin | 7411 | 2 | 2 | 0 | 2 | 0 | 0 | 6 | 2 | 4 | 3 | 0.038 | |
| gypsy | 7469 | 0 | 1 | 1 | 0 | 0 | 0 | 2 | 1 | 1 | 1 | 0.000 | |
| gypsy2 | 6841 | 1 | 0 | 0 | 2 | 0 | 0 | 3 | 1 | 2 | 2 | 0.067 | |
| gypsy3 | 6973 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 1 | 1 | 2 | 0.038 | |
| gypsy4 | 6852 | 0 | 1 | 0 | 0 | 1 | 0 | 2 | 1 | 1 | 2 | 0.041 | |
| gypsy5 | 7369 | 1 | 0 | 0 | 1 | 0 | 0 | 2 | 1 | 1 | 0 | 0.005 | |
| gypsy6 | 7826 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | - | |
| HMS-Beagle | 7062 | 4 | 5 | 1 | 0 | 3 | 0 | 13 | 9 | 4 | 3 | 0.054 | |
| Idefix | 7411 | 1 | 2 | 0 | 3 | 1 | 0 | 7 | 2 | 5 | 5 | 0.022 | |
| invader1 | 4032 | 0 | 0 | 4 | 3 | 18 | 1 | 26 | 1 | 25 | 7 | 0.023 | |
| invader2 | 5124 | 1 | 4 | 3 | 2 | 0 | 0 | 10 | 3 | 7 | 10 | 0.053 | |
| invader3 | 5484 | 2 | 5 | 2 | 2 | 5 | 0 | 16 | 3 | 13 | 5 | 0.044 | |
| invader4 | 3105 | 0 | 4 | 2 | 1 | 1 | 1 | 9 | 2 | 7 | 2 | 0.068 | |
| invader5 | 4038 | 1 | 4 | 0 | 1 | 0 | 0 | 6 | 0 | 6 | 6 | 0.068 | |
| McClintock | 6450 | 0 | 0 | 0 | 1 | 0 | 1 | 2 | 2 | 0 | 1 | 0.002 | |
| mdg1 | 7480 | 5 | 2 | 9 | 6 | 3 | 0 | 25 | 13 | 12 | 2 | 0.012 | |
| mdg3 | 5519 | 3 | 5 | 2 | 2 | 4 | 0 | 16 | 8 | 8 | 8 | 0.009 | |
| micropia | 5457 | 1 | 0 | 0 | 0 | 4 | 0 | 5 | 2 | 3 | 1 | 0.010 | |
| opus | 7521 | 3 | 6 | 6 | 6 | 3 | 0 | 24 | 16 | 8 | 5 | 0.003 | |
| qbert | 7650 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | - | |
| Quasimodo | 7387 | 2 | 7 | 0 | 4 | 1 | 0 | 14 | 5 | 9 | 5 | 0.016 | |
| roo | 9092 | 35 | 22 | 31 | 31 | 27 | 0 | 146 | 58 | 88 | 22 | 0.012 | |
| rooA | 7621 | 1 | 0 | 0 | 1 | 1 | 2 | 5 | 0 | 5 | 3 | 0.045 | |
| rover | 7318 | 3 | 0 | 1 | 0 | 2 | 0 | 6 | 3 | 3 | 3 | 0.035 | |
| springer | 7546 | 2 | 1 | 4 | 1 | 3 | 0 | 11 | 5 | 6 | 5 | 0.061 | |
| Stalker | 7256 | 3 | 1 | 0 | 5 | 3 | 0 | 12 | 3 | 9 | 7 | 0.014 | |
| Stalker2 | 8119 | 4 | 0 | 5 | 2 | 1 | 1 | 13 | 4 | 9 | 6 | 0.015 | |
| Stalker4 | 7379 | 1 | 0 | 0 | 0 | 1 | 0 | 2 | 2 | 0 | 0 | 0.001 | |
| Tabor | 7345 | 1 | 2 | 0 | 0 | 0 | 0 | 3 | 2 | 1 | 0 | 0.001 | |
| Tirant | 8526 | 4 | 3 | 3 | 4 | 5 | 1 | 20 | 15 | 5 | 5 | 0.001 | |
| Transpac | 5249 | 2 | 1 | 0 | 0 | 2 | 0 | 5 | 5 | 0 | 0 | 0.000 | |
| ZAM | 8435 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - | |
| LINE-like | baggins | 5453 | 1 | 2 | 10 | 0 | 0 | 1 | 14 | 0 | 14 | 12 | 0.076 |
| BS | 5142 | 2 | 6 | 6 | 7 | 8 | 0 | 29 | 6 | 23 | 6 | 0.028 | |
| Cr1a | 4470 | 1 | 5 | 17 | 21 | 2 | 10 | 56 | 1 | 55 | 42 | NC | |
| Doc | 4725 | 5 | 16 | 5 | 19 | 10 | 0 | 55 | 30 | 25 | 7 | 0.006 | |
| Doc2 | 4789 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | - | |
| Doc3 | 4740 | 0 | 1 | 6 | 1 | 0 | 1 | 9 | 0 | 9 | 8 | 0.065 | |
| F | 4708 | 2 | 7 | 10 | 10 | 11 | 2 | 42 | 16 | 26 | 11 | 0.019 | |
| G | 4346 | 1 | 2 | 0 | 0 | 0 | 0 | 3 | 0 | 3 | 1 | 0.059 | |
| G2 | 3102 | 1 | 9 | 1 | 2 | 1 | 0 | 14 | 2 | 12 | 2 | 0.036 | |
| G3 | 4605 | 1 | 1 | 2 | 0 | 0 | 0 | 4 | 0 | 4 | 3 | 0.029 | |
| G4 | 3856 | 0 | 5 | 0 | 2 | 3 | 1 | 11 | 0 | 11 | 6 | 0.100 | |
| G5 | 4856 | 0 | 5 | 0 | 1 | 2 | 1 | 9 | 0 | 9 | 6 | 0.063 | |
| G6 | 2042 | 1 | 1 | 0 | 1 | 0 | 0 | 3 | 1 | 2 | 3 | 0.006 | |
| Helena | 1317 | 2 | 1 | 1 | 1 | 2 | 0 | 7 | 0 | 7 | 5 | 0.097 | |
| HeT-A | 6083 | 0 | 0 | 0 | 0 | 0 | 3 | 3 | 0 | 3 | 0 | 0.018 | |
| I | 5371 | 7 | 5 | 6 | 3 | 5 | 2 | 28 | 8 | 20 | 9 | 0.037 | |
| Ivk | 5402 | 3 | 3 | 0 | 1 | 0 | 0 | 7 | 2 | 5 | 4 | 0.070 | |
| jockey | 5020 | 16 | 9 | 15 | 13 | 14 | 2 | 69 | 12 | 57 | 4 | 0.003 | |
| jockey2 | 3428 | 4 | 2 | 0 | 0 | 1 | 3 | 10 | 0 | 10 | 5 | 0.112 | |
| Juan | 4236 | 4 | 1 | 1 | 1 | 2 | 0 | 9 | 6 | 3 | 3 | 0.001 | |
| R1 | 5356 | 2 | 1 | 2 | 2 | 0 | 3 | 10 | 2 | 8 | 7 | 0.049 | |
| R2 | 3607 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - | |
| Rt1a | 5108 | 0 | 0 | 6 | 5 | 2 | 0 | 13 | 5 | 8 | 6 | 0.053 | |
| Rt1b | 5183 | 4 | 6 | 14 | 7 | 5 | 1 | 37 | 5 | 32 | 22 | 0.095 | |
| Rt1c | 5443 | 11 | 2 | 2 | 1 | 0 | 1 | 17 | 1 | 16 | 14 | 0.177 | |
| TART | 10654 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | - | |
| X | 4740 | 3 | 8 | 4 | 8 | 2 | 0 | 25 | 6 | 19 | 12 | 0.049 | |
| TIR | 1360 | 1177 | 10 | 18 | 16 | 20 | 11 | 30 | 105 | 10 | 95 | 48 | 0.087 |
| Bari1 | 1728 | 0 | 1 | 1 | 0 | 2 | 1 | 5 | 5 | 0 | 0 | 0.002 | |
| Bari2 | 1064 | 0 | 2 | 0 | 0 | 1 | 1 | 4 | 1 | 3 | 3 | 0.043 | |
| HB | 1653 | 5 | 7 | 6 | 3 | 7 | 4 | 32 | 5 | 27 | 21 | 0.114 | |
| H | 2959 | 5 | 11 | 2 | 1 | 5 | 0 | 24 | 1 | 23 | 3 | 0.069 | |
| hopper | 1435 | 5 | 2 | 3 | 2 | 3 | 0 | 15 | 11 | 4 | 6 | 0.048 | |
| hopper2 | 1680 | 0 | 3 | 0 | 2 | 0 | 0 | 5 | 1 | 4 | 0 | 0.042 | |
| looper1 | 1881 | 1 | 0 | 0 | 0 | 2 | 0 | 3 | 0 | 3 | 2 | 0.048 | |
| mariner2 | 912 | 5 | 5 | 2 | 2 | 0 | 3 | 17 | 4 | 13 | 13 | 0.144 | |
| NOF | 4347 | 2 | 0 | 2 | 2 | 1 | 0 | 7 | 0 | 7 | 4 | 0.013 | |
| P | 2907 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - | |
| pogo | 2121 | 9 | 12 | 5 | 7 | 11 | 0 | 44 | 5 | 39 | 5 | 0.020 | |
| S | 1736 | 9 | 4 | 11 | 11 | 12 | 4 | 51 | 14 | 37 | 27 | 0.079 | |
| S2 | 1735 | 1 | 3 | 3 | 3 | 2 | 1 | 13 | 0 | 13 | 8 | 0.268 | |
| Tc1 | 1666 | 2 | 1 | 4 | 1 | 2 | 11 | 21 | 1 | 20 | 7 | 0.085 | |
| transib1 | 2167 | 0 | 0 | 2 | 0 | 0 | 0 | 2 | 0 | 2 | 2 | 0.000 | |
| transib2 | 2844 | 3 | 5 | 2 | 2 | 0 | 0 | 12 | 0 | 12 | 6 | 0.082 | |
| transib3 | 2883 | 0 | 1 | 1 | 1 | 1 | 3 | 7 | 0 | 7 | 4 | 0.088 | |
| transib4 | 2656 | 2 | 1 | 2 | 0 | 0 | 0 | 5 | 1 | 4 | 4 | 0.152 | |
| FB | FB | 1492 | 12 | 4 | 2 | 8 | 4 | 2 | 32 | 13 | 19 | 13 | 0.066 |
The canonical length of each element (in bp) is shown in column 3, the total numbers of each family on each chromosome arm in columns 4-9, the grand totals for each family in column 10, and the numbers that are full length, partial and in the most proximal 2 Mb of the major chromosome arms, in columns 11-13. Partial elements are defined as those whose length is less than 97% of the canonical element. The average pairwise distance within each family is shown in column 14. The Cr1a family could not be reliably aligned and therefore average pairwise distance was not computed (NC).
As shown in Table 1 and Figure 1, the different classes vary in their contribution to the Drosophila euchromatin both in amount of sequence and number of elements. LTR elements make up the largest proportion of the euchromatin (2.65%), more sequence than the sum of all other classes of element (LINE-like elements 0.87%, TIR elements 0.31%, and FB elements 0.04%). LTR elements are also the most numerous class of transposable element in the euchromatic sequences (682) followed by LINE-like (486), TIR (372), and FB (32) elements. The largest family representing each of the three major classes is roo (146 copies; LTR), jockey (69 copies; LINE-like), and 1360 (105 copies; TIR) (Table 2). The average size of all transposable elements in our study is 2.9 kilobases (kb), smaller than the 5.6 kb average length of middle repetitive DNA, estimated from reassociation kinetics [36].
Figure 1.
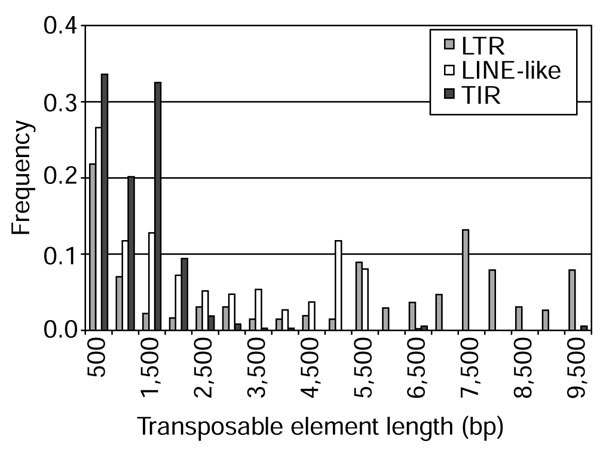
Frequency distribution of transposable element lengths in the Drosophila genome. Plotted are the lengths (in bp) of individual elements by functional class: LTR (gray), LINE-like (white), and TIR (black). Pairwise tests among all three classes (LTR versusLINE-like, LTRversusTIR, and LINE-like versus TIR) reveal that the distribution of individual element lengths differ significantly between functional classes (Mann-Whitney U test, p < 1 × 10-6).
Three of the 96 families are not described in this paper because they have not been found in the euchromatic portion of this sequence; these are the P element, R2 and ZAM. It is not surprising that we did not find any P elements, as the sequenced strain was selected to be free of them. While we did not find R2 and ZAM elements in the euchromatin, both of these elements were identified in unmapped scaffolds that derive from the heterochromatin [37]. The R2 element has previously been found only within the 28 S rDNA locus and in heterochromatin [38]. Strains of D. melanogaster are known to exist in which ZAM elements occur in low copy number in heterochromatic sequences [39]. The absence of the telomere-associated HeT-A and TART from the euchromatic portions of all chromosomes except chromosome 4 is not unexpected; the tandem arrays of these two elements are flanked by Taq microsatellite sequences [40,41] which are difficult to assemble and are under-represented in the current version of this sequence.
We discovered eight new families of transposable element within the Release 3 sequences. Two are members of the TIR class: Bari2 (EMBL: AF541951) and hopper2 (EMBL: AF541950). Six are members of the LTR class: frogger (EMBL: AF492763), rover (EMBL: AF492764), cruiser (a.k.a. Quasimodo) (EMBL: AF364550), McClintock (EMBL:AF541948), qbert (EMBL: AF541947), and Stalker4 (EMBL: AF541949). We identified Bari2 (four copies) by querying the D. melanogaster genome using a Bari1-like element isolated from D. erecta (EMBL: Y13853). The Bari2 element shares 52% amino acid identity with the Bari1 element; overall these elements share less than 50% nucleotide identity throughout their sequence. The hopper2 (five copies) and Stalker4 (two copies) families were identified by an analysis of the multiple alignment of the hopper and Stalker families, respectively. These alignments indicate distinct subfamilies on the basis of both nucleotide divergence and structural rearrangements over large regions of their alignment. The hopper2 and hopper elements share 70% amino-acid identity throughout their predicted open reading frames (ORFs). However, outside their ORFs, these elements are quite divergent and do not share significant nucleotide identity (< 30%). The Stalker4 element shares nearly 100% identity over the length of the predicted protein-coding domains, but these elements share only 50% nucleotide identity over the remainder of their sequence. A similar amount of conservation is seen between the previously defined [42] Stalker and Stalker2 sequences. The frogger element (one partial copy) was identified on the basis of its LTRs and a predicted protein-coding ORF that is 73% similar at the amino-acid level to that of the Dm88 family. The rover family (six copies) was identified in a BLAST search for repetitive elements in the genome; it is most closely related to the 17.6 element (71% amino acid identity). The cruiser family (14 copies) was identified during the finishing project by virtue of its LTRs, and is most closely related in sequence to the Idefix family, sharing 60% amino acid identity. The qbert family was identified by searching for regions of the genome that share similarity with protein-coding ORFs represented in our transposable element dataset. The qbert family (one copy) is most highly related to the accord family and shares regions of similarity that are 66% identical at the amino-acid level. The McClintock family (two copies), identified by its presence in a repeat region near the centromere of chromosome 4, is most closely related to the 17.6 family. McClintock shares 86% amino-acid identity with the protein-coding ORFs of 17.6; elsewhere these elements are quite divergent, sharing less than 50% identity over the first 5,000 bp of the McClintock element.
We have also discovered several other sequences with high sequence similarity to the protein-coding regions of transposable elements, but they are not associated with repeats (see also [24]). These elements cannot easily be classified into particular families. Although we have not included them in this analysis, they have been included in the Release 3 annotation of the genome [43]. While some may be examples of functional host genes derived from transposable elements, such as are known in humans (see, for example [44]) and ciliates (see, for example [45,46]), others may reflect remnants of elements that have become functionally constrained in the host genome (see, for example [47]).
Chromosomal distribution of elements
The percentage of each chromosome arm composed of transposable elements varies between 3.11% and 4.29%, except for chromosome 4, which is over 10% transposable elements (Table 1). The average transposable element density is 10-15 per million bases (Mb) for the major chromosome arms, and over 82 per Mb for chromosome 4. These densities are greater than the estimate of 5 per Mb derived from lower-resolution cytological methods [48], presumably because unclustered elements and partial elements may give weak in situ hybridization signals. In contrast to previous findings and theoretical expectations [22,49], we found no evidence for a reduction in density of transposable elements on the X chromosome relative to the major autosome arms (Table 1).
The densities of LINE-like elements and TIR elements on chromosome 4 are from five to ten times higher than their densities on the major chromosome arms, 25.85 per Mb and 46.85 per Mb, respectively, compared to 2.51-5.37 per Mb and 2.15-3.42 per Mb, respectively (Table 1, see also [22]). By contrast, the density of LTR elements on chromosome 4 is only slightly higher (8.08 per Mb) than on the five major chromosome arms (5.01-6.90 per Mb). Moreover, the percentage of chromosome 4 that is composed of LTR elements is only slightly higher than that of the major chromosome arms (3.56% versus 2.23-2.89%). Thus, the difference in density of transposable elements on chromosome 4 is predominantly due to an order-of-magnitude increase in the number of LINE-like and TIR elements.
Transposable element density is also known to vary along the major chromosome arms [20,21,22]. As shown in Figure 2 and discussed in Table 1, the density of transposable elements increases in the proximal euchromatin, here defined as the proximal 2 Mb of the assembly of each of the five major chromosome arms constituting about 10% of the euchromatic sequence analyzed. On the major chromosome arms, 36.7% (577/1,572) of the elements are located in proximal euchromatin, consistent with previous observations that the density of transposable elements is higher in heterochromatic regions of the genome [50,51,52,53,54]. These proximal sequences represent the transition between euchromatin and heterochromatin. Of 14 families located exclusively within the proximal 2 Mb, 12 are low copy number (defined here as less than 8 copies). Elements belonging to low copy number families show some tendency to be located in these regions; 78 of 142 elements that belong to low copy number families are located in the proximal 2 Mb of the chromosome arms.
Figure 2.
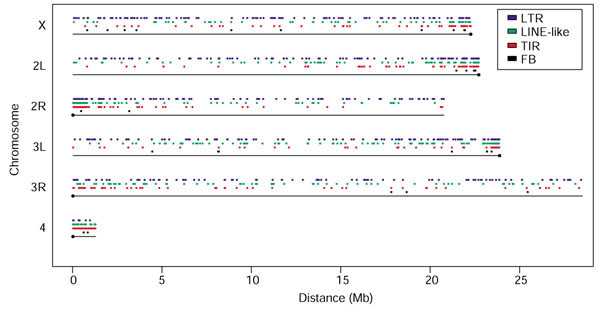
Distribution of transposable elements along chromosome arms. For each chromosome arm, the centromeres are indicated by circles. Each colored tick marks the start coordinate of an element belonging to one of the four classes of element (see key). Note the large number of LTR elements (blue) relative to the other classes on the major chromosome arms, and the higher number of LINE-like (green) and TIR (red) elements relative to the number of LTR elements seen for chromosome 4. While there is a relatively even distribution of transposable elements throughout the majority of each arm, there is a significant increase in the density of all classes of element in the proximal euchromatin (see also Table 1).
Finally, although the densities of transposable elements in the proximal euchromatin and chromosome 4 are both elevated with respect to the euchromatic average (58 and 82 elements per Mb, respectively), the composition of the elements in these regions is quite different. The increase in transposable element density in the proximal regions of the chromosome arms is due to increased numbers of elements belonging to all structural classes (Figure 2), while the increase in elements on chromosome 4 is due almost exclusively to LINE-like and TIR elements.
Analysis of structural variation
Transposable elements can be autonomous or defective with respect to transposition. Defective elements often exhibit deletions in ORFs or terminal repeats which are necessary for transposition. Assuming that canonical elements represent full-length active copies, we defined any element less than 97% of the length of the canonical member of their family as partial. On the basis of this criterion, more than two-thirds (1,094/1,572) of the elements in the Release 3 sequence are partial (Table 1). The proportion of partial elements is reasonably uniform among major chromosome arms (64-73%); 463 of 1,001 (46.3%) partial elements on the major chromosome arms lie within the proximal 2 Mb. In contrast, 91% of the transposable elements on chromosome 4 are partial. As LINE-like and TIR elements make up 88% of the elements on chromosome 4, these data indicate differences in proportions of partial elements between classes. In fact, 79% of LINE-like elements and 84% of TIR elements are partial, whereas only 55% of LTR elements are partial (Table 1).
Twenty-five of the 93 families (26.8%) represented in Release 3 are composed entirely of partial elements (8 LTR, 11 LINE-like, 6 TIR). An additional 17 families have only one full-length element. Fifteen of the 25 partial-only families are low copy number (less than 8 copies; 5 LTR, 6 LINE-like, 4 TIR) and 10 are high copy number (3 LTR, 5 LINE-like, 2 TIR). The majority of elements (133/196) in these 25 low copy number families are found in the proximal 2 Mb or on chromosome 4; all elements for 9 of these 25 families are found exclusively in these regions of the genome.
Analysis of the distribution of transposable element lengths scaled relative to the length of their canonical sequence shows that all three classes have bimodal distributions of scaled element lengths, but differ significantly from one another (Figure 3). The bimodal shape of these distributions presumably reflects the boundary states of the dynamic process of deletion, excision and transposition. Only a very small number of LINE-like (6) and TIR (16) elements exceed their canonical length, consistent with the fact that deletions occur more frequently than insertions in D. melanogaster [55]. A higher number of LTR elements (30) exceed the length of their canonical sequence, but on average these elements are less than 2% longer than their canonical length.
Figure 3.
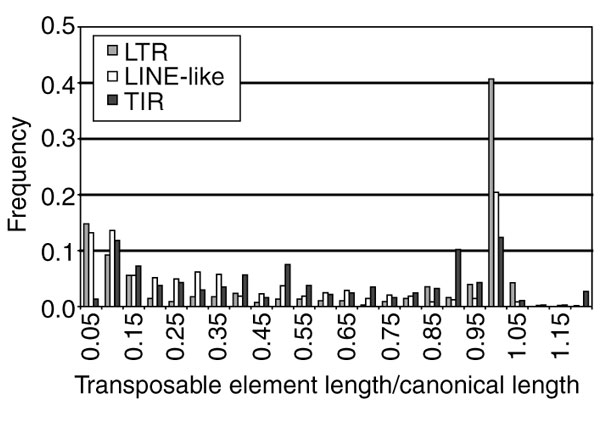
Frequency distribution of transposable element lengths scaled relative to their canonical lengths. Plotted are the scaled lengths of individual elements by functional class: LTR (gray), LINE-like (white), and TIR (black). Mann-Whitney U tests among all three classes (LTR versus LINE-like, LTRversusTIR, and LINE-like versus TIR) reveal that the distribution of scaled element lengths differ significantly between functional classes (Mann-Whitney U test, p < 1 × 10-4).
We characterized the distribution of structural variation for a representative element from each of the three major classes by determining the proportion of sequences represented in multiple alignments for a given nucleotide site (Figure 4). The resulting plot for the LINE-like jockey family approximates a negative exponential distribution starting from the 3' end (Figure 4a). LINE-like elements become deleted preferentially at their 5' ends, as a consequence of their mechanism of transposition [56]. The TIR element pogo shows a very different pattern; internal deletions predominate, leaving the inverted repeat termini intact [57] (Figure 4b). By analogy with patterns of deletion in P elements [58], these deleted elements will be non-autonomous with respect to transposition and presumably arise when double-stranded gap repair is interrupted [59,60]. By contrast, for the representative LTR element roo, there is a relatively uniform pattern of structural variation across the element, with the exception of two apparent deletion hotspots, at coordinates approximately 1 kb and approximately 8 kb, both of which occur in regions that are expected to be coding (Figure 4c).
Figure 4.
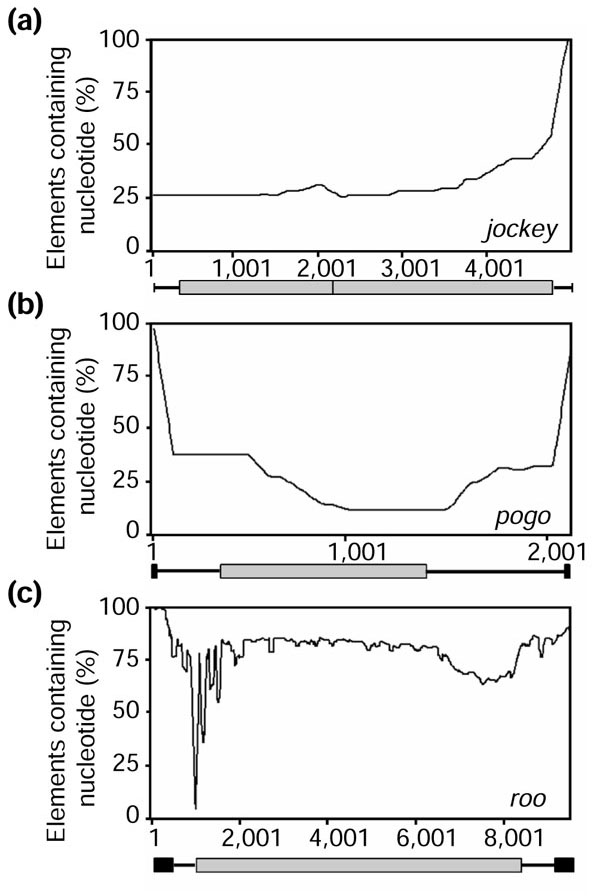
Structural variation within three common transposable elements: (a) jockey; (b) pogo; (c) roo. Multiple alignments generated from each respective family were used to approximate genomic variation. Each position along the length of the multiple alignment (x-axis) was measured for the presence of a nucleotide. The percentage of elements within an alignment that contained the nucleotide was determined and is indicated along the y-axis. A schematic drawing representing a jockey, pogo, or roo element is shown directly below each panel; coding regions are indicated by light-gray boxes and repeats (pogo and roo) are represented by black boxes.
One class of defective LTR elements, solo LTR sequences, has been known for some time in Drosophila [61] and other species [62,63,64]. These presumably arise by exchange between the two LTRs flanking an element, with the loss of the reciprocal product, a small circular molecule. In Saccharomyces cerevisiae, 85% of all LTR element insertions are solo LTRs [65]. We screened for solo LTRs of each family of element, using a criterion of 80% identity to the canonical LTR sequence of each family. Only 58 solo LTRs were identified, of which 14 are roo LTR elements.
Analysis of sequence variation within families
Point mutations in coding regions of the gypsy family of retrotransposons correlate with both transposition frequency and copy number [66]. We identified only one full-length gypsy class element (FBti0019898). Sequence comparison of this gypsy's ORF2 with that of the 'active' strain ORF2 shows these two ORFs to be identical and suggests that the single full-length gypsy element is 'active' in the sequenced strain. Other families of element have also been found to be polymorphic with respect to their coding potential in Drosophila. Kalmykova et al. [67] found that most 1731 elements have the +1 frameshift between their gag and pol gene regions typical of LTR elements; some do not, however, and instead express a Gag-Pol fusion protein. The single full-length 1731 element in Release 3 (FBti0020325) is of the latter type.
Sequence variation within families of element was estimated by analyzing the average pairwise distance within each family after multiple alignment (Table 2). These data show that intra-family variation ranges from complete identity to 26.8% average pairwise distance (see S2 family), but with only seven families having greater than 10% average pairwise distance. Analysis of the distribution of intra-family average pairwise distances by functional class shows that LTR families have lower levels of average pairwise distance relative to LINE-like or TIR families (Figure 5). The average sequence divergence for the LTR class of elements is only 2.6%; for the LINE-like and TIR classes it is 5.4% and 7.7%, respectively. These estimates of intra-family variation are remarkably similar to those of Wensink [68] who, by studying the kinetics of DNA reassociation of cloned middle repetitive sequences, showed that families of middle repetitive sequence exhibited on the order of 3-7% sequence divergence. These data illustrate differences in the amount of within-family variation seen between classes of element, which could be due to a variety of distinct mechanisms including differences in genealogical history or selective constraints.
Figure 5.
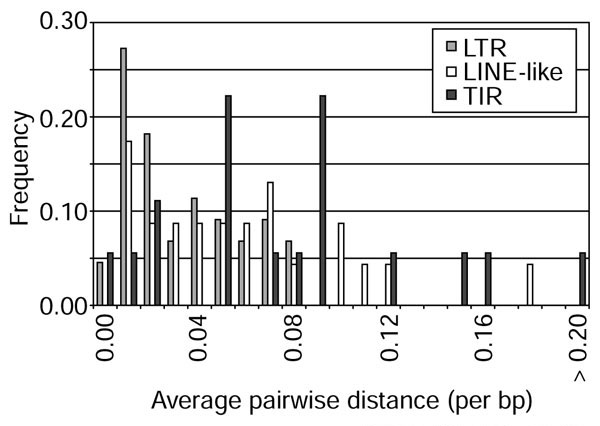
Frequency distribution of within-family average pairwise distances. Plotted are average pairwise distances (per bp) for individual transposable element families by functional class. Mann-Whitney U tests reveal that intra-family average pairwise distances differ significantly between LTR families and LINE-like families (p < 0.005), and between LTR families and TIR families (p < 0.0005), but not between LINE-like and TIR families (p < 0.311).
Nesting and clustering of transposable elements
The nesting of transposable elements is common in plant genomes [69,70,71,72]. For our analysis, transposable elements that have inserted within another element are termed nests; groups of transposable elements located within 10 kb of each other are defined as clusters. We found 62 nests or clusters of transposable elements containing 328 full or partial elements. This indicates that about 21% of transposable elements in this study are either inserted into another element or positioned adjacent to another element. The number of nested or clustered elements per arm ranges from 1.4 to 3.6 per Mb. The density of such elements is much higher in the proximal regions of the euchromatic arms; of the 62 nests or clusters, 25 are within the proximal 2 Mb regions of the major chromosome arms [73]. Eighty-nine percent of the elements belonging to nests or clusters are partial, in contrast to 69% of all elements. LTR elements are nested or clustered more often (29.3%) than either LINE-like elements (12.0%) or TIR elements (15.8%). This is presumably due to the larger proportion of LTR elements present in the Drosophila euchromatin.
Foldback (FB) elements often contain non-FB sequences [30,74]. Both NOF and HB elements have been found flanked by FB arms [30,75]. We identified two HB elements immediately adjacent to FB elements and four examples of NOF elements inserted into FB elements.
Patterns of element nesting can be very complex, as has been observed in other species [72], and may involve elements of the same class or elements of different classes. The insertion of a transposable element may trigger a runaway process, since it will provide a target into which other elements may insert without deleterious consequences [76]. In a sample of 31 simple nests each involving only two or three elements, we observed all nine possible combinations of nesting among the LTR, LINE-like and TIR classes. The largest euchromatic complex of elements is on chromosome arm 3R (coordinate approximately 8.3 Mb); it is a complex of 30 fragments of Dm88, 18 fragments of invader1 and three fragments of micropia elements, occupying 32.4 kb. Many of these fragments are identical; for example, of the 18 invader1 fragments, nine represent bases 1-424 of the canonical invader1 LTR sequence, three represent bases 143-424, two bases 80-424 and two bases 1-108. Losada et al. [77] have suggested that some novel transposable elements have evolved by nesting, in particular, that the Circe element arose as a consequence of the insertion of the Loa-like element of D. silvestris into the Ulysses-like element of D. virilis.
Several complex nests involve many different families of element. The nest near the base of 2L (coordinate approximately 20.1 Mb), for example, involves 11 different families of all three major classes of element. Large clusters containing only one family of element are also found. For example, there is a complex of seven GATE elements at coordinate approximately 14.2 Mb on chromosome arm 2R and a complex of six mdg1 elements at approximately 5.7 Mb on the same chromosome arm.
Some transposable elements are present as large tandem arrays. For example, the Tc1-like Bari1 is organized as a tandem array in the heterochromatin at the base of chromosome arm 2R [78]. Tandem LTR element pairs have also been found in the D. melanogaster genome (for example, FBti0019752 and FBti0019753); here, two roo elements share an internal LTR. A number of different mechanisms have been suggested to result in tandem Ty1 and Ty5 elements in S. cerevisiae [65,79]; all involve recombination between either linear cDNAs or circular DNA generated by LTR transposition and a chromosomal element. The mechanism(s) by which tandem elements arise in Drosophila is not known.
Insertion-site preferences of natural transposable elements
Transposable elements insert at a staggered cut in chromosomal DNA; after repair, this results in a duplication of the target sequence. For the R1 and R2 LINE-like elements, there is high insertion-site specificity for sites within the 28 S rDNA gene [38]. For some LTR retrotransposons, a preference for AT-rich sequences has been known for some time [80,81,82]. We estimated the physical characteristics of 500 bp of DNA flanking the insertion sites of three high-copy number elements, roo (LTR), jockey (LINE-like) and pogo (TIR). In our analysis, we included only elements for which the duplicated target sequence could be unambiguously identified. Within the individual element families, we found no recognizable motif in the nucleotide sequence of the repeats flanking insertions (data not shown). However, analysis of different physical characteristics of these sequences revealed distinct characteristics for each of the three families of element (Figure 6). Our data suggest that roo and pogo prefer to insert in sequences of either higher than average (roo) or lower than average (pogo) denaturation temperatures; this may reflect functional differences in the insertion mechanism of these elements. There is no obvious bias in the sequences into which jockey elements insert. Analysis of other high-copy families of element revealed distinct insertion site characteristics, suggesting that the characteristics shown in Figure 6 are not shared within classes of element (not shown).
Figure 6.
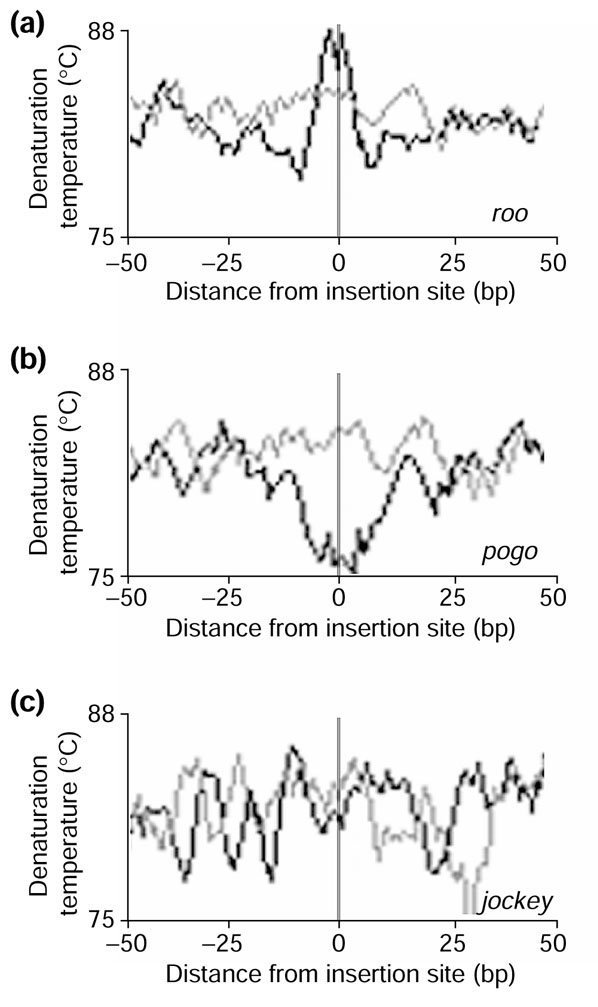
Structural characteristics of DNA insertion sites. Sites for (a) roo; (b) pogo; (c) jockey. Genomic sequence flanking the insertion site of each element was extracted from our dataset. Those elements for which duplicated target sequences could not be identified were discarded. The remaining sequences from each family were centered on the repeat (vertical gray line) and the average denaturation temperature across all sequences was determined using a 3-bp window size. In each panel, the light horizontal gray line represents the average denaturation temperature of random genomic sequence and the horizontal black line represents the average denaturation temperature of the experimental set of sequences. The x-axis represents the distance (in bp) from the insertion site and the y-axis represents the temperature (°C). The sequences flanking the roo (a) and pogo (b) elements have opposite characteristics; the roo sequences have a higher than average denaturation temperature whereas the pogo sequences have a lower than average denaturation temperature. The average denaturation temperature of the sequence flanking the jockey elements does not differ from that of the random sequence.
LTR retrotransposons use a tRNA primer for first-strand synthesis during transposition. In S. cerevisiae, 90% of LTR retrotransposons are within 750 bp of tRNA genes, and there are an average of 1.2 insertions per tRNA gene [65]. Our data suggest no relationship between the location of tRNAs and transposable elements in D. melanogaster. Of 313 elements on chromosome arm 2R, only five are within 10 kb of a tRNA gene, or tRNA gene cluster. However, Saigo [83] has described an association of a tRNA pseudogene and the 3' end of a copia element, possibly resulting from an aberrant reverse transcription; an initiating tRNA: Met pseudogene has also been described as being associated with repetitive sequences [84].
It has been known for many years that the P element shows a marked preference to insert immediately 5' to genes or within 5' exons [85]. This preference presumably reflects the chromatin environment at the time of P-element transposition. We analyzed the position of transposable elements with respect to the closest known or predicted gene from the Release 3 reannotation [86]. There are 551 elements located 5' to transcribed regions, 585 elements located 3' to transcribed regions, and 436 elements within transcribed regions. These ratios are consistent for all classes of element, suggesting that there is no insertion site bias with respect to genes. We also find no bias of insertion with respect to the transcribed strand.
The proportion of transposable elements is higher in intergenic regions than in transcribed regions. Only 27.7% (436/1,572) transposable elements map within regions that are annotated as transcribed, although over 50% of the major chromosome arms are predicted to be transcribed. This result suggests that a large proportion of transposable element insertions in transcribed regions have deleterious effects and are not incorporated into the genome of D. melanogaster. As with total numbers of transposable elements on each chromosome arm, we see no reduction in the number of transposable elements inserted within transcribed regions on the X chromosome. Of the 436 transposable elements inserted within genes, 79 are on the X chromosome, which is within the range seen on the other major chromosome (70-88). This is consistent with the percent of coding/non-coding sequence on the X chromosome (51.9%) relative to that of the other chromosome arms (53.8%).
All 436 transposable elements that map within transcribed regions are predicted to be within introns (see also [22]). However, during the reannotation of the genome [86], coding exons were not annotated in sequences with homology to transposable elements. Thus it is possible that a small number of transposons within transcribed regions actually are inserted into a coding exon. It is worth noting that, of the four mutations known to be carried by the sequenced strain one (bw1), and possibly a second (sp1), are mutated by the insertion of 412 elements.
In a recent study of five protein-coding genes located in the proximal regions of chromosome arms, Dimitri et al. [35] found that introns contain 50% transposable element sequence; this contrasts with euchromatic introns, which contain only 0.11% transposable element sequence.
Transposable elements in completely sequenced genomes
We can make a preliminary comparison of the transposable elements of D. melanogaster with those of the other fully sequenced eukaryote genomes: S. cerevisiae [87], Schizosaccharomyces pombe [64], Caenorhabditis elegans [88] and Arabidopsis thaliana [89].
In S. cerevisiae, all transposable elements are of the LTR class [62]; five different families are known and these comprise 3.1% of the entire genome in S. cerevisiae. The majority of these are solo LTRs (85%) [65]. This is in contrast to the few solo LTRs found in the Release 3 sequences of D. melanogaster. Transposable elements are quite rare in the sequenced strain of S. pombe; only 11 intact, and three defective, Tf2 LTR elements are known [64,90].
In C. elegans, all three major classes of transposable element are found. There are 19 families of LTR retrotransposons, with at most three full-length members [25,63]. There are three families of LINE-like retrotransposons, Rte-1, Sam and Frodo, with about 30 elements overall; 11 of Sam, three of Frodo and 10-15 of Rte-1 [91,92]. There are seven families of TIR (Tc) elements, with copy numbers between 61 and 294 [93], nine families of mariner-like element, with from one to 66 copies ([45], quoted in [46]), and five families of short DNA elements [94], with copy numbers from 81 to 1,204 [93]. Given the occurrence of retrotransposons in both C. elegans and D. melanogaster, it is interesting that the genomes of both species are characterized by very few retrotransposed pseudogenes [95,96,97]. These maybe generated at a low rate, or may be deleted quickly as suggested by studies of the lineages of Helena elements in D. virilis [98] and D. melanogaster [55].
Transposable elements are far more abundant in the genome of A. thaliana than in the euchromatic genomes of C. elegans or D. melanogaster. In Arabidopsis, over 5,500 transposable elements exist, representing 10% of the 'euchromatic' sequence [89]. The pericentromeric heterochromatin and the heterochromatic knob on chromosome 4 of Arabidopsis have a very high density of transposable elements and other repeats [89,99,100,101]. As in Drosophila, certain families of elements appear only in these heterochromatic regions, for example the Arabidopsis retrotransposon Athila. In contrast to Drosophila, class I and class II elements in Arabidopsis show very different chromosomal distributions, the former in the centromeric regions and the latter flanking these regions.
One class of element that is absent or so far unrecognized in the genome of D. melanogaster are the MITEs, miniature inverted repeat elements, characterized as short (under 500 bp) elements with inverted repeat termini and without a transposase necessary for autonomous transposition. Elements similar to MITEs have been described in D. subobscura and its relatives [102]. Whether or not MITEs are indeed a separate class of element, or simply represent internally deleted (and hence non-autonomous) TIR elements is unclear [103]. In C. elegans, these elements are abundant, with 5,000 elements in four sequence families; they show a non-random chromosomal distribution [104]. MITEs are characteristic of plant genomes; in A. thaliana, Surzycki and Belknap [105] have identified three families with a copy number of about 90. In maize there are an estimated 6,000 copies of the mPIF family of MITE elements alone, and there is evidence for autonomous family members [106].
An additional class of element not identified in the Drosophila genome are the SINEs (short interspersed elements) [29,107]. This class of element is often closely associated with LINE elements and it has been proposed that the transposition of SINEs utilizes proteins encoded by LINE elements [107]. The DINE-1 family of repetitive element, isolated in Drosophila [32], shares weak similarity to the SINEs, but this family lacks important structural features typical of other members of this class.
Comparison of sequence and cytological data
In this paper we have described the transposable elements of the euchromatin of D. melanogaster, as represented by the Release 3 sequences. Because Release 3 represents only a single sequence from the y1; cn1 bw1 sp1 isogenic strain first constructed in J. Kennison's laboratory in the early 1990s [108], it is important to determine whether the composition of transposable elements in this strain is typical of the species as a whole.
It is well established that Drosophila strains vary in the number and location of transposable elements; these differences are often taken as de facto evidence of transposability [12,13]. Large differences in the abundance of transposable elements have been observed between laboratory strains for families such as gypsy, Bari1, ZAM and Idefix [109,110,111,112]. Such variation in transposable element copy number may be associated with a mutation either of the element itself, or of host genes that would normally regulate copy number (see, for example, the role of the flamenco gene in regulating gypsy activity [113]; see [114,115] for a review of host-element interactions). Transposable elements also differ between laboratory strains and natural populations of D. melanogaster (Table 3, see [116] for review). Perhaps the most salient examples are those elements that cause hybrid dysgenesis - the P element, the I element and the H element (a.k.a. hobo) - which are either wholly absent from, or defective in, most laboratory strains, but abundant today in natural populations [58,117,118]. Only one of the H elements appears to be full length, and comparison of its coding sequence to that of the canonical suggests that this element is active in the sequenced strain. Further, eight of the I elements identified in the sequenced strain appear to be of similar length and sequence to the active canonical element. The ORFs of these elements are very similar to those of the canonical, suggesting that they too might be active.
Table 3.
A comparison of the numbers of euchromatic transposable elements in the Release 3 sequence with those estimated from natural populations and laboratory stocks by in situ hybridization
| Class | Family | Number in Release 3 | Range | Midpoint | Midpoint average | Source | Reference | |
| LTR | 17.6 | 12 | 8-17 | 12.5 | 12.5 | 7-10 isogenic lines, Beltsville, MD | [51] | |
| 1731 | 2 | 8-15 | 11.5 | 11.5 | 7-10 isogenic lines, Beltsville, MD | [51] | ||
| 297 | 57 | 18-35 | 26.5 | 26.6 | 7-10 isogenic lines, Beltsville, MD | [51] | ||
| 20-30 | 25 | 4 lab stocks | [13] | |||||
| 23 | 23 | 20 isogenic lines, Raleigh, NC | [49] | |||||
| 32 | 32 | 182 inbred lines | [145] | |||||
| 3S18 | 6 | 25 | 25 | 18.5 | 8 lab stocks | [146] | ||
| 12 | 12 | 6 lab stocks | [19] | |||||
| 412 | 31 | 18-38 | 28 | 25.8 | 7-10 isogenic lines, Beltsville, MD | [51] | ||
| 26-32 | 29 | 4 lab stocks | [13] | |||||
| 21 | 21 | 20 isogenic lines, Raleigh, NC | [49] | |||||
| 31 | 31 | 7 natural populations | [147] | |||||
| 20 | 20 | 182 inbred lines | [145] | |||||
| blood | 22 | 14 | 14 | 14.0 | 182 inbred lines | [145] | ||
| copia | 30 | 20-43 | 31.5 | 24.3 | 4 lab stocks | [13] | ||
| 12-25 | 18.5 | 18 inbred strains, Azerbaidjan | [148] | |||||
| 23 | 23 | 7 natural populations | [147] | |||||
| gypsy | 2 | 1 | 1 | 3.0 | 7 natural populations | [147] | ||
| 5 | 5 | 182 inbred lines | [145] | |||||
| HMS-Beagle 13 | 11 | 11 | 11.0 | Hey and Eanes (unpublished), [116] | ||||
| Idefix | 7 | 4-20 | 12 | 12.0 | 6 lab stocks | [110] | ||
| mdg1 | 25 | 11-23 | 17 | 20.3 | 7-10 isogenic lines, Beltsville, MD | [51] | ||
| 15-27 | 21 | 20 lab stocks | [149] | |||||
| 14-22 | 18 | 17 inbred stocks, Azerbaidjan | [150] | |||||
| 25 | 25 | 18 inbred strains, Azerbaidjan | [148] | |||||
| mdg3 | 16 | 5-18 | 11.5 | 11.5 | 20 lab stocks | [149] | ||
| opus | 24 | 12-23 | 17.5 | 15.0 | 7-10 isogenic lines, Beltsville, MD | [51] | ||
| 10-15 | 12.5 | 2 lab stocks | [151] | |||||
| roo | 147 | 55-75 | 65 | 63.0 | 7-10 isogenic lines, Beltsville, MD | [51] | ||
| 61 | 61 | 20 isogenic lines, Raleigh, NC | [49] | |||||
| Stalker | 12 | 2-6 | 4 | 4.0 | 8 lab stocks | [146] | ||
| Tirant | 20 | 3-13 | 8 | 9.5 | 10 wild-type stocks | [152] | ||
| 6-16 | 11 | 3 lab stocks | [153] | |||||
| ZAM | 0 | 0-15 | 7.5 | 7.8 | 4 lab stocks | [111] | ||
| 1-15 | 8 | 3 lab stocks | [39] | |||||
| LINE-like | Doc | 55 | 20-30 | 25 | 25.0 | 2 lab stocks | [154] | |
| F | 42 | 25-30 | 27.5 | 34.3 | 1 lab stock | [155] | ||
| 41 | 41 | Hey and Eanes (unpublished), [116] | ||||||
| I | 28 | 13-21 | 17 | 12.3 | 18 inbred strains, Azerbaidjan | [148] | ||
| 0-15 | 7.5 | 6 lab stocks | [19] | |||||
| jockey | 69 | 32-40 | 36 | 29.0 | 7-10 isogenic lines, Beltsville, MD | [51] | ||
| 22 | 22 | 182 inbred lines | [145] | |||||
| TIR | 1360 | 105 | 19-39 | 29 | 29 | 4 lab strains | [156] | |
| Bari1 | 5 | 4 | 4 | 9.8 | 4 lab stocks | [157] | ||
| 2-29 | 15.5 | 46 lab stocks and natural populations | [109] | |||||
| H | 24 | 8-60 | 34 | 27.0 | 17 inbred stocks, Azerbaidjan | [150] | ||
| 25-27 | 26 | natural populations, Greece | [158] | |||||
| 21 | 21 | 182 inbred lines | [145] | |||||
| NOF | 7 | 0-2 | 1 | 1.0 | 8 lab strains | [159] | ||
| S | 51 | 24-91 | 57.5 | 57.5 | 10 lab stocks and natural populations | [160] | ||
| FB | FB | 32 | 20-30 | 25 | 22.5 | 8 lab strains | [159] | |
| 20 | 20 | 182 inbred lines | [145] |
These data are illustrative of the published literature, not an exhaustive survey. The estimated range of copy number per family, range midpoint for each source, and midpoint averages across all sources are shown in columns 4, 5 and 6, respectively.
There has been extensive sampling of laboratory and natural strains of D. melanogaster for euchromatic transposable elements by the method of in situ hybridization [48,116]. These samples provide estimates of transposable element abundance that are relevant to compare with our data, as both types of studies sample euchromatic sequences. As shown in Table 3, the sequenced y1; cn1 bw1 sp1 isogenic strain is a typical D. melanogaster strain, at least with respect to the numbers of euchromatic elements. Overall, the Spearman rank order correlation coefficient between the number of elements of each family in Release 3 and the average midpoint of the ranges seen in other strains is 0.86 (p < 10-6). The correlation between these two types of data is imperfect as closely located elements (within 100 kb) of the same family and grossly deleted elements are not resolved by the method of in situ hybridization. Moreover, the copy number of any individual element may be very different in different strains (see above), and certain elements (for example, copia, Doc, roo) may dramatically increase in copy number in particular laboratory strains (see [119,120]). Nevertheless, this strong correlation suggests that results based on analysis of the sequenced strain may be representative of the species as a whole.
As previously noted, 25 of the 93 families of transposable element represented in the Release 3 euchromatic sequence have only partial elements. Full-length copies of these families may be discovered in other strains of D. melanogaster, in the heterochromatin, or in closely related species. Indeed, full-length copies of the aurora family are present in D. simulans (Dsim/ninja [121,122]), of the mariner clade in D. mauritiana [123], and of the Helena family in D. virilis [124]. Our understanding of the evolutionary dynamics of the transposable elements will also be immeasurably improved by comparative studies between D. melanogaster and other Diptera, such as Anopheles gambiae and D. pseudoobscura.
Materials and methods
The sequence and other datasets
The sequence releases
Release 1 of the 'complete' euchromatic sequence of the genome of D. melanogaster was made available in March 2000 [20,26]. As explained in Background, this sequence, by and large derived from an assembly of a 12.8× whole-genome shotgun sequence, is not suitable for the analysis of repeated sequences. Nor was the subsequent release made available from the Berkeley Drosophila Genome Project's (BDGP) website, Release 2 (October 2000), which filled 330 sequence gaps (but left some 1,300) and had improved the order and orientation data for scaffolds [125]. Release 3 is the first high-quality complete sequence of this genome; it is of Phase 3 quality [126]; that is, not only is the sequence quality itself high (an estimated sequence error rate of less than 1 in 30,000 bp), but there are very few gaps.
There are two caveats with respect to the assembly used in this analysis (see [28] for details). The first is that regions containing arrays of similar elements may suffer from assembly artifacts. Resolving these will require the use of a constrained assembly program. The second is that some parts of the distal X chromosome and of chromosome arm 3L are unfinished in Release 3 [28]. The transposable elements of these unfinished sequences were included in analyses of the abundance and distribution of elements, but were excluded from all analyses that required the alignment of element sequences. Seventy-three elements are included within Tables 1 and 2, but not included in alignments. These elements will be clearly indicated in our datasets (see below).
In Release 1 there were 3.8 Mb of sequence that could not be mapped to any chromosome arm [20]. These were assembled into unmapped scaffolds that included sequences from gaps in the euchromatic arms and sequences from the centric 'heterochromatin', including the entire Y chromosome [127]. The highly repetitive nature of these sequences makes them difficult to assemble, either from a whole genome shotgun or from a cloned-based sequencing strategy. The 'heterochromatin' also includes sequences that cannot be readily cloned, for instance the satellite DNA sequences and, perhaps, others. The distinct genetic and cytological nature of the pericentromeric regions of the Drosophila chromosomes, both metaphase and polytene interphase, has been known for many years [128,129] and its structure and properties clearly result from the nature of its sequences [130].
We defined euchromatin as all sequence that has been assembled into a chromosome arm scaffold, and heterochromatin as the rest (unmapped scaffolds, see [37]). We recognize, of course, that the transition from euchromatin to heterochromatin is not abrupt; indeed we show that the characteristics of the most basal regions of the chromosome arms differ in their sequence organization from the regions distal to them.
The dataset we used for unmapped scaffolds is from a new assembly provided by Celera Genomics ([28] and E. Myers and G. Sutton, personal communication). This assembly, WGS3, is the result of improvements to the Celera assembly algorithms [26]. WGS3 assembles 115.5 Mb into 14 mapped scaffolds, and leaves 22.2 Mb in 2,761 scaffolds, each less than 1 Mb in length, assigned to unmapped scaffolds. One reason for the increase in size of unmapped scaffolds between the first and third whole-genome shotgun assemblies is the inclusion in WGS3 of some 809,000 extra sequence reads not included in the two previous assemblies [127]. The 22.2 Mb of sequence in unmapped scaffolds includes sequences that properly belong to the euchromatin as well as heterochromatic sequences. For this reason we simply used this dataset as a subject sequence set for searching, by BLAST, for elements that we had been unable to discover in the euchromatic sequence. A description of the transposable elements of the heterochromatin, and of the telomeres, of D. melanogaster is the subject of a publication from the Drosophila Heterochromatin Genome Project [37].
Reference datasets
A reference dataset of 'canonical' sequences of transposable elements was built by M. Ashburner, P. Benos and G. Liao during the early stages of developing methods for Drosophila genome annotation. It was first used during the annotation of the 2.9 Mb 'Adh region' [131] and has been maintained subsequently, and made public, by the Cambridge and Berkeley groups [132]. As new sequences were published by others these were added to this file. Most of these were 'real' sequences, although some from Repbase [23,42] were consensus sequences. In addition, we made a determined effort to discover, from the evolving Release 3 sequence, 'complete' sequences of the many elements known only from small sequence fragments, for example of their LTR regions, as well as all new elements identified in this study. The following elements were available too late to be included in our analyses, but will be included in further updates of the data: Tc3-like [133], ninja (EMBL: AF520587) and the elements 'DREF', 'BG.DS00797' and 'CG.13775' of Robertson [46]. The sequences described by Robertson [46] may be examples of functional host genes derived from transposable elements, such as are known in humans (for example [44]) and ciliates (for example [45], and H. Robertson, personal communication).
Nomenclature
Many transposable elements of D. melanogaster have been described and named independently by several research groups. In this paper we use the names adopted by FlyBase, which attempts to reflect priority of publication (or sequence release). There are, in addition to those described here, many elements in FlyBase that have never been associated with a sequence or a restriction map. In the absence of further evidence, nothing more can be said about these, which will be marked as being of 'uncertain status' in their FlyBase records.
Available datasets
The following datasets are freely available for download [134] and are maintained by FlyBase. When using these resources please note, and publish, the Release numbers associated with the files.
File 1. A file containing a single ('canonical') sequence of each family of element; this is a frozen dataset of the sequences used to search the genome for transposable elements.
File 2. A file of annotated 'canonical' sequences, one for each identified family of transposable element. This file is, in effect, an update version of file 1. These sequences were chosen as the longest discovered in the genome with (where relevant and where possible) intact ORFs. There are a few families for which no intact element could be found. We have then attempted to construct an intact element from the available data. Such artifices are noted in the records. These data will be updated when new information becomes available, and will be further annotated by FlyBase. Each release will be archived.
File 3. A file, in FASTA format, of each individual element that has been discovered. The following data are to be found on the header line of each record:
>family_name,FBgn_id,FBti_id,chromosome_arm:Release 3_coordinates
FBgn_id is the FlyBase record for the family, FBti_id is the unique identifier of each occurrence of an element and the coordinates are from the Release 3 data. In addition to the sequence of each element, each record includes 500 bp of 5' and 3' flanking sequence. These data will be regularly updated, in step with each new Release of the assembled sequence. Each release will be archived.
File 4. The alignments of elements within a family used for the current analysis. This file is in MASE format [135] with each element identified by its FBti number. This is a frozen dataset that will not be updated by the BDGP.
File 5. The nested transposable elements and element complexes are available as an independent dataset. Included within each sequence is 500 bp of flanking sequence on each side of the element complex. Each nest or complex has a unique FBti identifier number in FlyBase; in addition each component of a nest or complex has its own FBti identifier number. In the FASTA header line for each sequence in this file the data included are:
>FBti_of_nest_or_complex,FBti_of_component,chromosome_arm: coordinates
Comparison with other datasets
To support our claim that the Release 1 sequence is an inadequate substrate for rigorous analysis we have compared the sequences of transposable elements in that release with those of Release 3. We determined the identity of elements in the two releases by a comparison of the 500 bp on their 5' flanks. Our results suggest that many, if not most, of the sequences from Release 1 are artifacts of that assembly. Of the 1,572 elements characterized in Release 3 only 381 (24%) were correctly determined in Release 1. Of the 1,191 (76%) sequences that were not correctly sequences in Release 1, 483 contained Ns, 45 were completely absent, and 663 contained an average of 34 incorrectly identified nucleotides per element. The complete data are available from [134].
Analytical methods
Identification of known transposable elements
WU-BLASTN 2.0 [136] was used to search all chromosome arms for regions of similarity to each element in the Release 3 dataset. The parameters for the BLAST search were M = 3, N = 3, Q = 3, R = 3, X = 3 and S = 3. BLAST searches were done on a 32-node dual PIII Linux-based compute farm supplied by Linux Network. Distribution of BLAST jobs to the cluster was managed by the Portable Batch System (PBS [137]). Individual BLAST jobs were submitted via pbsrsh, an rsh-like program (E.F., unpublished work). In addition, PBS was optimized and modified for the BDGP to handle a large number of queued jobs (E.F., unpublished work).
BLAST reports were generated by searching a single chromosome arm with each individual element. The results were then parsed to generate a list of the coordinates of all high-scoring pairs (HSPs) that were at least 50 bp long and whose query and subject sequences had a pairwise identity of at least 90%. All HSPs on this list that were within 10 kb of each other and summed to greater than 100 bp were pooled into a 'span'. Each span was bounded by two coordinates - a start coordinate that corresponds to the lowest coordinate of any HSP in a particular span, and an end coordinate that corresponds to the highest coordinate of any HSP in the same span. A master list was then generated that contained all spans for all elements on a particular arm. Any spans (for the same or different elements) that had overlapping coordinates were examined further by an analysis of the sequences of the HSPs. While this identified a small number of spurious spans that did not correspond to real elements, the majority of these instances correspond to the nested elements discussed below. Start and end coordinates for all spans belonging to each element were used to extract genomic sequences for multiple sequence alignment (see below). In some rare instances where it was not possible to differentiate the element to which the HSP belonged, overlapping coordinates were recorded. Spurious sequences that did not align with other family members were removed from both the list of spans and the multiple alignments. Other attempts to define transposable element families on the basis of sequence identity have used a 90% cutoff with reference to the protein sequence of the reverse transcriptase motif of LTR-elements [25,138]. For LINE-like transposons, Berezikov et al. [24] used a 70% nucleic acid sequence identity criterion over 200 bp.
Identification of new transposable elements through genome-genome comparison
The first approach to discovering new transposable elements was by an all-by-all BLAST using chromosome arms 2L, 2R, 3R, 4 and the proximal half of the X. The chromosome arms were divided into 20-kb segments, each segment overlapping the previous by 10 kb. We used the NCBI-BLAST 2.0 to compare each 20-kb section against the others. Hits with greater than 95% identity and 1,000 bp long were parsed and used as query sequences in a BLAST against the canonical element sequence dataset. Redundant results were removed. The coordinates of the repeats were parsed and known repeats were tagged. New repeats were reviewed in CONSED [139] for the presence of ORFs and repeat structure.
Identification of new transposable elements through isolation of LTR sequences
A second approach was taken to identify single-copy elements containing LTRs. Each chromosome arm was divided into 1,000-bp pieces with neighboring pieces overlapping each other by 500 bp. WU-BLASTN 2.0 was used to search each chromosome arm for all regions of similarity to each 1000-bp piece (parameters: M = 3, N = 3, Q = 3, R = 3, X = 3 and S = 3). The BLAST report from such a search was parsed to generate a list of all HSPs that were at least 100 bp long and whose query and subject sequences had a pairwise identity of at least 95%. Then, all HSPs on this list greater than 500 bp apart and less than 15 kb apart were pooled into a span. As above, each span was bounded by a start coordinate which corresponds to the lowest coordinate of any HSP in a particular pool and an end coordinate which corresponds to the highest coordinate of any HSP in the same pool. Each set of coordinates was compared to the list of coordinates of transposable elements identified in the screen for known elements and these were eliminated from this list. Then, the coordinates of the remaining spans were used to extract genomic sequence from the finished chromosome arms. Each piece of genomic sequence was then compared to the coding sequence of the known transposable elements using WU-TBLASTX 2.0 (with default parameters). Any span that produced a hit with a E < 10-8 was analyzed by searching through the non-redundant protein database at the NCBI using NCBI-BLASTX [126].
Alignment and calculation of evolutionary distances
Preliminary multiple alignments of elements within families were made using the default settings of DIALIGN v2-1 [140]. The resulting multiple alignments were visualized in the SEAVIEW alignment editor [135]. Subsequent realignment was done using the CLUSTALW (1.7.4) [141] implementation internal to SEAVIEW with manual refinement. Multiple alignments were used to calculate average pairwise distance within families using Kimura's 2-parameter substitution model (transition:transversion ratio = 2:1) [142] as implemented in the DNADIST program of the PHYLIP package [143].
Physical characteristics of element insertion sites
To analyze the physical properties of the insertion sites of transposable elements we used the programs developed by Liao et al. [144]. The flanking sequences of elements with canonical ends were aligned, centered on a single copy of the element's target site sequence (that duplicated on insertion). The sequences were then analyzed for A-philicity, propeller twist, duplex stability and denaturation temperature, as described in [144]. As a baseline we used a randomly generated 500-bp sequence set of the same base composition as the overall genome of D. melanogaster (G. Liao, personal communication). These analyses were performed with 49 roo element sequences, 12 jockey sequences and 28 pogo sequences. Additional analyses were carried out using elements from the following families: copia, blood, 412 and Doc.
Additional data files
A table showing the classification of transposable elements in the genus Drosophila is available.
Supplementary Material
A table showing the classification of transposable elements in the genus Drosophila
Acknowledgments
Acknowledgements
This work was supported by NIH grant HG00750 to G.M.R., by NIH grant HG00739 to FlyBase (W.M. Gelbart) and by programme grant G822559 from the Medical Research Council to M.A., D. Gubb and S. Russell. C.M.B is supported by NIH training grant T32 HL07279 to E. Rubin. Research was conducted at the Lawrence Berkeley National Laboratory under Department of Energy contract DE-AC0376SF00098, University of California. M.A. thanks Jean Wiborg for her help in the logistics of his visits to Berkeley. We thank Patrizio Dimitri, Bernardo Carvalho and Gary Karpen for allowing us to quote from their unpublished work, and Bob Levis, Mike Young, Stu Tsubota, Francois Payre, and Andy Flavell, for information on individual elements. We also thank Christian Biemont, Bernardo Carvalho, Patrizio Dimitri, Dan Hartl, Sergey Nuzhdin, Dmitri Petrov, Hugh Robertson, and Alfredo Ruiz, for their comments on the manuscript of this paper. We also thank the anonymous reviewers for their helpful suggestions and comments.
This article is part of a series of refereed research articles from Berkeley Drosophila Genome Project, FlyBase and colleagues, describing Release 3 of the Drosophila genome, which are freely available at http://genomebiology.com/drosophila/.
References
- Craig NL, Craigie R, Gellert M, Lambowitz AM, Eds Mobile DNA II Washington, DC: ASM Press; 2002.
- Green MM. Genetic instability in Drosophila melanogaster: Mutable miniature (mmu). Mutat Res. 1975;29:77–84. doi: 10.1016/0027-5107(75)90022-6. [DOI] [PubMed] [Google Scholar]
- Demerec M. Miniature-α - a second frequently mutating character in Drosophila virilis. Proc Natl Acad Sci USA. 1926;12:687–690. doi: 10.1073/pnas.12.12.687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demerec M. Magenta-α - a third frequently mutating character in Drosophila virilis. Proc Natl Acad Sci USA. 1927;13:249–253. doi: 10.1073/pnas.13.4.249. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Demerec M. Unstable genes. Bot Rev. 1935;1:233–248. [Google Scholar]
- McClintock B. The origin and behavior of mutable loci in maize. Proc Natl Acad Sci USA. 1950;36:344–355. doi: 10.1073/pnas.36.6.344. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Starlinger P. Mutations caused by the integration of IS1 and IS2 into the gal operon. In: Bukhari AI, Shapiro JA, Adhya SL, editor. In DNA Insertion Elements, Plasmids, and Episomes. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory; 1977. pp. 25–30. [Google Scholar]
- Green MM. A case for DNA insertion mutants in Drosophila melanogaster. In: Bukhari AI, Shapiro JA, Adhya SL, editor. In DNA Insertion Elements, Plasmids, and Episomes. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory; 1977. pp. 437–445. [Google Scholar]
- Rubin GM, Finnegan DJ, Hogness DS. The chromosomal arrangement of coding sequences in a family of repeated genes. Prog Nucleic Acid Res Mol Biol. 1976;19:221–226. doi: 10.1016/s0079-6603(08)60920-4. [DOI] [PubMed] [Google Scholar]
- Finnegan DJ, Rubin GM, Young MW, Hogness DS. Repeated gene families in Drosophila melanogaster. Cold Spring Harb Symp Quant Biol. 1978;42:1053–1063. doi: 10.1101/sqb.1978.042.01.106. [DOI] [PubMed] [Google Scholar]
- Ilyin YV, Tchurikov NA, Ananiev EV, Ryskov AP, Yenikolopov GN, Limborska SA, Maleeva NE, Gvozdev VA, Georgiev GP. Studies on the DNA fragments of mammals and Drosophila containing structural genes and adjacent sequences. Cold Spring Harb Symp Quant Biol. 1978;42:959–969. doi: 10.1101/sqb.1978.042.01.097. [DOI] [PubMed] [Google Scholar]
- Young MW. Middle repetitive DNA: a fluid component of the Drosophila genome. Proc Natl Acad Sci USA. 1979;76:6274–6278. doi: 10.1073/pnas.76.12.6274. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strobel E, Dunsmuir P, Rubin GM. Polymorphisms in the chromosomal locations of elements of the 412, copia and 297 dispersed repeated gene families in Drosophila. Cell. 1979;17:429–439. doi: 10.1016/0092-8674(79)90169-7. [DOI] [PubMed] [Google Scholar]
- Glover DM. Cloned segment of Drosophila melanogaster rDNA containing new types of sequence insertion. Proc Natl Acad Sci USA. 1977;74:4932–4936. doi: 10.1073/pnas.74.11.4932. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiraizumi Y. Spontaneous recombination in Drosophila melanogaster males. Proc Natl Acad Sci USA. 1971;68:268–270. doi: 10.1073/pnas.68.2.268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kidwell MG. Hybrid dysgenesis in Drosophila melanogaster: The relationship between the P-M and I-R interaction systems. Genet Res. 1979;33:205–217. [Google Scholar]
- Engels WR, Preston CR. Hybrid dysgenesis in Drosophila melanogaster: the biology of female and male sterility. Genetics. 1979;92:161–174. doi: 10.1093/genetics/92.1.161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bingham PM, Kidwell MG, Rubin GM. The molecular basis of P-M hybrid dysgenesis: the role of the P element, a P-strain-specific transposon family. Cell. 1982;29:995–1004. doi: 10.1016/0092-8674(82)90463-9. [DOI] [PubMed] [Google Scholar]
- Bucheton A, Paro R, Sang HM, Pelisson A, Finnegan DJ. The molecular basis of I-R hybrid dysgenesis in Drosophila melanogaster: identification, cloning, and properties of the I factor. Cell. 1984;38:153–163. doi: 10.1016/0092-8674(84)90536-1. [DOI] [PubMed] [Google Scholar]
- Adams MD, Celniker SE, Holt RA, Evans CA, Gocayne JD, Amanatides PG, Scherer SE, Li PW, Hoskins RA, Galle RF, et al. The genome sequence of Drosophila melanogaster. Science. 2000;287:2185–2195. doi: 10.1126/science.287.5461.2185. [DOI] [PubMed] [Google Scholar]
- Rizzon C, Marais G, Gouy M, Biemont C. Recombination rate and the distribution of transposable elements in the Drosophila melanogaster genome. Genome Res. 2002;12:400–407. doi: 10.1101/gr.210802. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartolome C, Maside X, Charlesworth B. On the abundance and distribution of transposable elements in the genome of Drosophila melanogaster. Mol Biol Evol. 2002;19:926–937. doi: 10.1093/oxfordjournals.molbev.a004150. [DOI] [PubMed] [Google Scholar]
- Jurka J. Repbase update: a database and an electronic journal of repetitive elements. Trends Genet. 2000;16:418–420. doi: 10.1016/s0168-9525(00)02093-x. [DOI] [PubMed] [Google Scholar]
- Berezikov E, Bucheton A, Busseau I. A search for reverse transcriptase-coding sequences reveals new non-LTR retrotransposons in the genome of Drosophila melanogaster. Genome Biol. 2000;1:research0012.1–0012.15. doi: 10.1186/gb-2000-1-6-research0012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bowen NJ, McDonald JF. Drosophila euchromatic LTR retrotransposons are much younger than the host species in which they reside. Genome Res. 2001;11:1527–1540. doi: 10.1101/gr.164201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Myers EW, Sutton GG, Delcher AL, Dew IM, Fasulo DP, Flanigan MJ, Kravitz SA, Mobarry CM, Reinert KH, Remington KA, et al. A whole-genome assembly of Drosophila. Science. 2000;287:2196–2204. doi: 10.1126/science.287.5461.2196. [DOI] [PubMed] [Google Scholar]
- Benos PV, Gatt MK, Murphy L, Harris D, Barrell B, Ferraz C, Vidal S, Brun C, Demaille J, Cadieu E, et al. From first base: the sequence of the tip of the X chromosome of Drosophila melanogaster, a comparison of two sequencing strategies. Genome Res. 2001;11:710–730. doi: 10.1101/gr.173801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Celniker SE, Wheeler DA, Kronmiller B, Carlson JW, Halpern A, Patel S, Adams M, Champe M, Dugan SP, Frise E, et al. Finishing a whole genome shotgun: Release 3 of the Drosophila melanogaster euchromatic genome sequence. Genome Biol. 2002;3:research0079.1–0079.14. doi: 10.1186/gb-2002-3-12-research0079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deininger PL. SINEs: Short interspersed repeated DNA elements in higher eukaryotes. In: Berg DE, Howe MM, editor. In Mobile DNA. Washington, DC: American Society of Microbiology; 1989. pp. 619–637. [Google Scholar]
- Truett MA, Jones RS, Potter SS. Unusual structure of the FB family of transposable elements in Drosophila. Cell. 1981;24:753–763. doi: 10.1016/0092-8674(81)90101-x. [DOI] [PubMed] [Google Scholar]
- Wilder J, Hollocher H. Mobile elements and the genesis of microsatellites in dipterans. Mol Biol Evol. 2001;18:384–392. doi: 10.1093/oxfordjournals.molbev.a003814. [DOI] [PubMed] [Google Scholar]
- Locke J, Howard LT, Aippersbach N, Podemski L, Hodgetts RB. The characterization of DINE-1, a short, interspersed repetitive element present on chromosome and in the centric heterochromatin of Drosophila melanogaster. Chromosoma. 1999;108:356–366. doi: 10.1007/s004120050387. [DOI] [PubMed] [Google Scholar]
- Locke J, Podemski L, Roy K, Pilgrim D, Hodgetts R. Analysis of two cosmid clones from chromosome 4 of Drosophila melanogaster reveals two new genes amid an unusual arrangement of repeated sequences. Genome Res. 1999;9:137–149. [PMC free article] [PubMed] [Google Scholar]
- Spradling AC, Rubin GM. Drosophila genome organization: conserved and dynamic aspects. Annu Rev Genet. 1981;15:219–264. doi: 10.1146/annurev.ge.15.120181.001251. [DOI] [PubMed] [Google Scholar]
- Dimitri P, Junakovic N, Arca B. Colonization of heterochromatic genes by transposable elements in Drosophila melanogaster. Mol Biol Evol. 2002. [DOI] [PubMed]
- Manning JE, Schmid CW, Davidson N. Interspersion of repetitive and nonrepetitive DNA sequences in the Drosophila melanogaster genome. Cell. 1975;4:141–155. doi: 10.1016/0092-8674(75)90121-x. [DOI] [PubMed] [Google Scholar]
- Hoskins RA, Smith CD, Carlson J, Carvalho AB, Halpern A, Kaminker JS, Kennedy C, Mungall CJ, Sullivan BA, Sutton GG, et al. Heterochromatic sequences in a Drosophila whole-genome shotgun assembly. Genome Biol. 2002;3:research0085.1–0085.16. doi: 10.1186/gb-2002-3-12-research0085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jakubczak JL, Burke WD, Eickbush TH. Retrotransposable elements R1 and R2 interrupt the rRNA genes of most insects. Proc Natl Acad Sci USA. 1991;88:3295–3299. doi: 10.1073/pnas.88.8.3295. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldrich E, Dimitri P, Desset S, Leblanc P, Codipietro D, Vaury C. Genomic distribution of the retrovirus-like element ZAM in Drosophila. Genetica. 1997;100:131–140. [PubMed] [Google Scholar]
- Beissmann H, Walter MF, Mason JM. Telomeres in Drosophila and other insects. In: Krupp G, Parwaresch R, editor. In Telomeres and Telomerases: Cancer and Biology. Georgetown, TX: Landes Biosciences; 2002. [Google Scholar]
- Pardue ML, DeBaryshe PG. Drosophila telomeres: two transposable elements with important roles in chromosomes. Genetica. 1999;107:189–196. [PubMed] [Google Scholar]
- Repbase Update http://www.girinst.org/Repbase_Update.html
- Berkeley Drosophila Genome Project: GadFly genome annotation database of Drosophila http://www.fruitfly.org/annot
- Robertson HM, Zumpano KL. Molecular evolution of an ancient mariner transposon, Hsmar1, in the human genome. Gene. 1997;205:203–217. doi: 10.1016/s0378-1119(97)00472-1. [DOI] [PubMed] [Google Scholar]
- Witherspoon DJ, Doak TG, Williams KR, Seegmiller A, Seger J, Herrick G. Selection on the protein-coding genes of the TBE1 family of transposable elements in the ciliates Oxytricha fallax and O. trifallax. Mol Biol Evol. 1997;14:696–706. doi: 10.1093/oxfordjournals.molbev.a025809. [DOI] [PubMed] [Google Scholar]
- Robertson HM. Evolution of DNA transposons in eukaryotes. In: Craig NL, Craigie R, Gellert M, Lambowitz AM, editor. In Mobile DNA II. Washington, DC: ASM Press; 2002. [Google Scholar]
- Maside X, Bartolome C, Charlesworth B. S-element insertions are associated with the evolution of the Hsp70 genes in Drosophila melanogaster. Curr Biol. 2002;12:1686–1691. doi: 10.1016/s0960-9822(02)01181-8. [DOI] [PubMed] [Google Scholar]
- Charlesworth B, Langley CH. The population genetics of Drosophila transposable elements. Annu Rev Genet. 1989;23:251–287. doi: 10.1146/annurev.ge.23.120189.001343. [DOI] [PubMed] [Google Scholar]
- Montgomery E, Charlesworth B, Langley CH. A test for the role of natural selection in the stabilisation of transposable element copy number in a population of Drosophila melanogaster. Genet Res. 1987;49:31–41. doi: 10.1017/s0016672300026707. [DOI] [PubMed] [Google Scholar]
- Carmena M, Gonzalez C. Transposable elements map in a conserved pattern of distribution extending from beta-heterochromatin to centromeres in Drosophila melanogaster. Chromosoma. 1995;103:676–684. doi: 10.1007/BF00344228. [DOI] [PubMed] [Google Scholar]
- Charlesworth B, Jarne P, Assimacopoulos S. The distribution of transposable elements within and between chromosomes in a population of Drosophila melanogaster. III. Element abundances in heterochromatin. Genet Res. 1994;64:183–197. doi: 10.1017/s0016672300032845. [DOI] [PubMed] [Google Scholar]
- Junakovic N, Terrinoni A, Di Franco C, Vieira C, Loevenbruck C. Accumulation of transposable elements in the heterochromatin and on the Y chromosome of Drosophila simulans and Drosophila melanogaster. J Mol Evol. 1998;46:661–668. doi: 10.1007/pl00006346. [DOI] [PubMed] [Google Scholar]
- Pimpinelli S, Berloco M, Fanti L, Dimitri P, Bonaccorsi S, Marchetti E, Caizzi R, Caggese C, Gatti M. Transposable elements are stable structural components of Drosophila melanogaster heterochromatin. Proc Natl Acad Sci USA. 1995;92:3804–3808. doi: 10.1073/pnas.92.9.3804. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dimitri P. Constitutive heterochromatin and transposable elements in Drosophilamelanogaster. Genetica. 1997;100:85–93. [PubMed] [Google Scholar]
- Petrov DA, Hartl DL. High rate of DNA loss in the Drosophila melanogaster and Drosophila virilis species groups. Mol Biol Evol. 1998;15:293–302. doi: 10.1093/oxfordjournals.molbev.a025926. [DOI] [PubMed] [Google Scholar]
- Finnegan DJ. Transposable elements: how non-LTR retrotransposons do it. Curr Biol. 1997;7:R245–R248. doi: 10.1016/s0960-9822(06)00112-6. [DOI] [PubMed] [Google Scholar]
- Tudor M, Lobocka M, Goodell M, Pettitt J, O'Hare K. The pogo transposable element family of Drosophila melanogaster. Mol Gen Genet. 1992;232:126–134. doi: 10.1007/BF00299145. [DOI] [PubMed] [Google Scholar]
- Engels WR. P elements in Drosophila melanogaster. In: Berg DE, Howe MM, editor. In Mobile DNA. Washington, DC: American Society of Microbiology; 1989. [Google Scholar]
- Engels WR, Johnson-Schlitz DM, Eggleston WB, Sved J. High-frequency P element loss in Drosophila is homolog dependent. Cell. 1990;62:515–525. doi: 10.1016/0092-8674(90)90016-8. [DOI] [PubMed] [Google Scholar]
- Hsia AP, Schnable PS. DNA sequence analyses support the role of interrupted gap repair in the origin of internal deletions of the maize transposon, MuDR. Genetics. 1996;142:603–618. doi: 10.1093/genetics/142.2.603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carbonare BD, Gehring WJ. Excision of copia element in a revertant of the white-apricot mutation of Drosophila melanogaster leaves behind one long-terminal repeat. Mol Gen Genet. 1985;199:1–6. doi: 10.1007/BF00327501. [DOI] [PubMed] [Google Scholar]
- Boeke JD. Transposable elements in Saccharomyces cerevisiae. In: Berg DE, Howe MM, editor. In Mobile DNA. Washington, DC: American Society of Microbiology; 1989. [Google Scholar]
- Ganko EW, Fielman KT, McDonald JF. Evolutionary history of Cer elements and their impact on the C. elegans genome. Genome Res. 2001;11:2066–2074. doi: 10.1101/gr.196201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wood V, Gwilliam R, Rajandream MA, Lyne M, Lyne R, Stewart A, Sgouros J, Peat N, Hayles J, Baker S, et al. The genome sequence of Schizosaccharomyces pombe. Nature. 2002;415:871–880. doi: 10.1038/nature724. [DOI] [PubMed] [Google Scholar]
- Kim JM, Vanguri S, Boeke JD, Gabriel A, Voytas DF. Transposable elements and genome organization: a comprehensive survey of retrotransposons revealed by the complete Saccharomyces cerevisiae genome sequence. Genome Res. 1998;8:464–478. doi: 10.1101/gr.8.5.464. [DOI] [PubMed] [Google Scholar]
- Lyubomirskaya NV, Smirnova JB, Razorenova OV, Karpova NN, Surkov SA, Avedisov SN, Kim AI, Ilyin YV. Two variants of the Drosophila melanogaster retrotransposon gypsy (mdg4): structural and functional differences, and distribution in fly stocks. Mol Genet Genomics. 2001;265:367–374. doi: 10.1007/s004380000427. [DOI] [PubMed] [Google Scholar]
- Kalmykova A, Maisonhaute C, Gvozdev V. Retrotransposon 1731 in Drosophila melanogaster changes retrovirus-like expression strategy in host genome. Genetica. 1999;107:73–77. [PubMed] [Google Scholar]
- Wensink PC. Sequence homology within families of Drosophila melanogaster middle repetitive DNA. Cold Spring Harb Symp Quant Biol. 1978;42:1033–1039. doi: 10.1101/sqb.1978.042.01.103. [DOI] [PubMed] [Google Scholar]
- Fu H, Park W, Yan X, Zheng Z, Shen B, Dooner HK. The highly recombinogenic bz locus lies in an unusually gene-rich region of the maize genome. Proc Natl Acad Sci USA. 2001;98:8903–8908. doi: 10.1073/pnas.141221898. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SanMiguel P, Gaut BS, Tikhonov A, Nakajima Y, Bennetzen JL. The paleontology of intergene retrotransposons of maize. Nat Genet. 1998;20:43–45. doi: 10.1038/1695. [DOI] [PubMed] [Google Scholar]
- SanMiguel P, Tikhonov A, Jin YK, Motchoulskaia N, Zakharov D, Melake-Berhan A, Springer PS, Edwards KJ, Lee M, Avramova Z, et al. Nested retrotransposons in the intergenic regions of the maize genome. Science. 1996;274:765–768. doi: 10.1126/science.274.5288.765. [DOI] [PubMed] [Google Scholar]
- Tikhonov AP, SanMiguel PJ, Nakajima Y, Gorenstein NM, Bennetzen JL, Avramova Z. Colinearity and its exceptions in orthologous adh regions of maize and sorghum. Proc Natl Acad Sci USA. 1999;96:7409–7414. doi: 10.1073/pnas.96.13.7409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O'Hare K, Chadwick BP, Constantinou A, Davis AJ, Mitchelson A, Tudor M. A 5.9-kb tandem repeat at the euchromatin-heterochromatin boundary of the X chromosome of Drosophila melanogaster. Mol Genet Genomics. 2002;267:647–655. doi: 10.1007/s00438-002-0698-x. [DOI] [PubMed] [Google Scholar]
- Caceres M, Puig M, Ruiz A. Molecular characterization of two natural hotspots in the Drosophila buzzatii genome induced by transposon insertions. Genome Res. 2001;11:1353–1364. doi: 10.1101/gr.174001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Harden N, Ashburner M. Characterization of the FB-NOF transposable element of Drosophila melanogaster. Genetics. 1990;126:387–400. doi: 10.1093/genetics/126.2.387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walbot V, Petrov DA. Gene galaxies in the maize genome. Proc Natl Acad Sci USA. 2001;98:8163–8164. doi: 10.1073/pnas.161278798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Losada A, Abad JP, Agudo M, Villasante A. The analysis of Circe, an LTR retrotransposon of Drosophila melanogaster, suggests that an insertion of non-LTR retrotransposons into LTR elements can create chimeric retroelements. Mol Biol Evol. 1999;16:1341–1346. doi: 10.1093/oxfordjournals.molbev.a026044. [DOI] [PubMed] [Google Scholar]
- Caizzi R, Caggese C, Pimpinelli S. Bari-1, a new transposon-like family in Drosophila melanogaster with a unique heterochromatic organization. Genetics. 1993;133:335–345. doi: 10.1093/genetics/133.2.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ke N, Voytas DF. High frequency cDNA recombination of the Saccharomyces retrotransposon Ty5: The LTR mediates formation of tandem elements. Genetics. 1997;147:545–556. doi: 10.1093/genetics/147.2.545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Inouye S, Yuki S, Saigo K. Sequence-specific insertion of the Drosophila transposable genetic element 17.6. Nature. 1984;310:332–333. doi: 10.1038/310332a0. [DOI] [PubMed] [Google Scholar]
- Freund R, Meselson M. Long terminal repeat nucleotide sequence and specific insertion of the gypsy transposon. Proc Natl Acad Sci USA. 1984;81:4462–4464. doi: 10.1073/pnas.81.14.4462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanda S, Shrimpton AE, Chueh LL, Itayama H, Matsubayashi H, Saigo K, Tobari YN, Langley CH. Retrovirus-like features and site specific insertions of a transposable element, tom, in Drosophila ananassae. Mol Gen Genet. 1988;214:405–411. doi: 10.1007/BF00330473. [DOI] [PubMed] [Google Scholar]
- Saigo K. A copia primer pseudogene possibly generated by an aberrant reverse transcription of a copia-related element in Drosophila. Nucleic Acids Res. 1986;14:7815. doi: 10.1093/nar/14.19.7815. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sharp S, DeFranco D, Silberklang M, Hosbach HA, Schmidt T, Kubli E, Gergen JP, Wensink PC, Soll D. The initiator tRNA genes of Drosophila melanogaster: evidence for a tRNA pseudogene. Nucleic Acids Res. 1981;9:5867–5882. doi: 10.1093/nar/9.22.5867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spradling AC, Stern DM, Kiss I, Roote J, Laverty T, Rubin GM. Gene disruptions using P transposable elements: an integral component of the Drosophila genome project. Proc Natl Acad Sci USA. 1995;92:10824–10830. doi: 10.1073/pnas.92.24.10824. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misra S, Crosby MA, Mungall CJ, Matthews BB, Campbell K, Hradecky P, Huang Y, Kaminker JS, Millburn GH, Prochnik SE, et al. Annotation of the Drosophila melanogaster euchromatic genome: a systematic review. Genome Biol. 2002;3:research0083.5–0083.22. doi: 10.1186/gb-2002-3-12-research0083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goffeau A, Aert R, Agostini-Carbone L, Ahmed A, Aigle M, Alberghina L, Albermann K, Albers M, Aldea M, Alexandraki D, et al. The yeast genome directory. Nature. 1997;387(Suppl):1–105. [Google Scholar]
- The C. elegans Sequencing Consortium Genome sequence of the nematode C. elegans: a platform for investigating biology. Science. 1998;282:2012–2018. doi: 10.1126/science.282.5396.2012. [DOI] [PubMed] [Google Scholar]
- The Arabidopsis Genome Initiative Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature. 2000;408:796–815. doi: 10.1038/35048692. [DOI] [PubMed] [Google Scholar]
- Weaver DC, Shpakovski GV, Caputo E, Levin HL, Boeke JD. Sequence analysis of closely related retrotransposon families from fission yeast. Gene. 1993;131:135–139. doi: 10.1016/0378-1119(93)90682-s. [DOI] [PubMed] [Google Scholar]
- Marin I, Plata-Rengifo P, Labrador M, Fontdevila A. Evolutionary relationships among the members of an ancient class of non-LTR retrotransposons found in the nematode Caenorhabditis elegans. Mol Biol Evol. 1998;15:1390–1402. doi: 10.1093/oxfordjournals.molbev.a025867. [DOI] [PubMed] [Google Scholar]
- Youngman S, van Luenen HG, Plasterk RH. Rte-1, a retrotransposon-like element in Caenorhabditis elegans. FEBS Lett. 1996;380:1–7. doi: 10.1016/0014-5793(95)01525-6. [DOI] [PubMed] [Google Scholar]
- Duret L, Marais G, Biemont C. Transposons but not retrotransposons are located preferentially in regions of high recombination rate in Caenorhabditis elegans. Genetics. 2000;156:1661–1669. doi: 10.1093/genetics/156.4.1661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Devine SE, Chissoe SL, Eby Y, Wilson RK, Boeke JD. A transposon-based strategy for sequencing repetitive DNA in eukaryotic genomes. Genome Res. 1997;7:551–563. doi: 10.1101/gr.7.5.551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jeffs P, Ashburner M. Processed pseudogenes in Drosophila. Proc R Soc Lond B Biol Sci. 1991;244:151–159. doi: 10.1098/rspb.1991.0064. [DOI] [PubMed] [Google Scholar]
- Harrison PM, Echols N, Gerstein MB. Digging for dead genes: an analysis of the characteristics of the pseudogene population in the Caenorhabditis elegans genome. Nucleic Acids Res. 2001;29:818–830. doi: 10.1093/nar/29.3.818. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang W, Brunet FG, Nevo E, Long M. Origin of sphinx, a young chimeric RNA gene in Drosophila melanogaster. Proc Natl Acad Sci USA. 2002;99:4448–4453. doi: 10.1073/pnas.072066399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrov DA, Lozovskaya ER, Hartl DL. High intrinsic rate of DNA loss in Drosophila. Nature. 1996;384:346–349. doi: 10.1038/384346a0. [DOI] [PubMed] [Google Scholar]
- Kapitonov VV, Jurka J. Molecular paleontology of transposable elements from Arabidopsis thaliana. Genetica. 1999;107:27–37. [PubMed] [Google Scholar]
- Lin X, Kaul S, Rounsley S, Shea TP, Benito MI, Town CD, Fujii CY, Mason T, Bowman CL, Barnstead M, et al. Sequence and analysis of chromosome 2 of the plant Arabidopsis thaliana. Nature. 1999;402:761–768. doi: 10.1038/45471. [DOI] [PubMed] [Google Scholar]
- Mayer K, Schuller C, Wambutt R, Murphy G, Volckaert G, Pohl T, Dusterhoft A, Stiekema W, Entian KD, Terryn N, et al. Sequence and analysis of chromosome 4 of the plant Arabidopsis thaliana. Nature. 1999;402:769–777. doi: 10.1038/47134. [DOI] [PubMed] [Google Scholar]
- Miller WJ, Nagel A, Bachmann J, Bachmann L. Evolutionary dynamics of the SGM transposon family in the Drosophila obscura species group. Mol Biol Evol. 2000;17:1597–1609. doi: 10.1093/oxfordjournals.molbev.a026259. [DOI] [PubMed] [Google Scholar]
- Kapitonov VV, Jurka J. POGON1, a 'bona fide' family of nonautonomous DNA transposons. Repbase Repts. 2002;2:7. [Google Scholar]
- Surzycki SA, Belknap WR. Repetitive-DNA elements are similarly distributed on Caenorhabditis elegans autosomes. Proc Natl Acad Sci USA. 2000;97:245–249. doi: 10.1073/pnas.97.1.245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Surzycki SA, Belknap WR. Characterization of repetitive DNA elements in Arabidopsis. J Mol Evol. 1999;48:684–691. doi: 10.1007/pl00006512. [DOI] [PubMed] [Google Scholar]
- Zhang X, Feschotte C, Zhang Q, Jiang N, Eggleston WB, Wessler SR. P instability factor: an active maize transposon system associated with the amplification of Tourist-like MITEs and a new superfamily of transposases. Proc Natl Acad Sci USA. 2001;98:12572–12577. doi: 10.1073/pnas.211442198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weiner AM. SINEs and LINEs: the art of biting the hand that feeds you. Curr Opin Cell Biol. 2002;14:343–350. doi: 10.1016/s0955-0674(02)00338-1. [DOI] [PubMed] [Google Scholar]
- Brizuela BJ, Elfring L, Ballard J, Tamkun JW, Kennison JA. Genetic analysis of the brahma gene of Drosophila melanogaster and polytene chromosome subdivisions 72AB. Genetics. 1994;137:803–813. doi: 10.1093/genetics/137.3.803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Caggese C, Pimpinelli S, Barsanti P, Caizzi R. The distribution of the transposable element Bari-1 in the Drosophila melanogaster and Drosophila simulans genomes. Genetica. 1995;96:269–283. doi: 10.1007/BF01439581. [DOI] [PubMed] [Google Scholar]
- Desset S, Conte C, Dimitri P, Calco V, Dastugue B, Vaury C. Mobilization of two retroelements, ZAM and Idefix, in a novel unstable line of Drosophila melanogaster. Mol Biol Evol. 1999;16:54–66. doi: 10.1093/oxfordjournals.molbev.a026038. [DOI] [PubMed] [Google Scholar]
- Leblanc P, Desset S, Dastugue B, Vaury C. Invertebrate retroviruses: ZAM a new candidate in D. melanogaster. EMBO J. 1997;16:7521–7531. doi: 10.1093/emboj/16.24.7521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim AI, Belyaeva ES, Aslanian MM. Autonomous transposition of gypsy mobile elements and genetic instability in Drosophila melanogaster. Mol Gen Genet. 1990;224:303–308. doi: 10.1007/BF00271566. [DOI] [PubMed] [Google Scholar]
- Prud'homme N, Gans M, Masson M, Terzian C, Bucheton A. Flamenco, a gene controlling the gypsy retrovirus of Drosophila melanogaster. Genetics. 1995;139:697–711. doi: 10.1093/genetics/139.2.697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Labrador M, Corces VG. Interactions between transposable elements and the host genome. In: Craig NL, Craigie R, Gellert M, Lambowitz AM, editor. In Mobile DNA II. Washington, DC: ASM Press; 2002. [Google Scholar]
- Labrador M, Corces VG. Transposable element-host interactions: regulation of insertion and excision. Annu Rev Genet. 1997;31:381–404. doi: 10.1146/annurev.genet.31.1.381. [DOI] [PubMed] [Google Scholar]
- Biemont C, Cizeron G. Distribution of transposable elements in Drosophila species. Genetica. 1999;105:43–62. doi: 10.1023/a:1003718520490. [DOI] [PubMed] [Google Scholar]
- Streck RD, MacGaffey JE, Beckendorf SK. The structure of hobo transposable elements and their insertion sites. EMBO J. 1986;5:3615–3623. doi: 10.1002/j.1460-2075.1986.tb04690.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crozatier M, Vaury C, Busseau I, Pelisson A, Bucheton A. Structure and genomic organization of I elements involved in I-R hybrid dysgenesis in Drosophila melanogaster. Nucleic Acids Res. 1988;16:9199–9213. doi: 10.1093/nar/16.19.9199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasyukova EG, Nuzhdin SV. Doc and copia instability in an isogenic Drosophila melanogaster stock. Mol Gen Genet. 1993;240:302–306. doi: 10.1007/BF00277071. [DOI] [PubMed] [Google Scholar]
- Nuzhdin SV, Mackay TF. The genomic rate of transposable element movement in Drosophila melanogaster. Mol Biol Evol. 1995;12:180–181. doi: 10.1093/oxfordjournals.molbev.a040188. [DOI] [PubMed] [Google Scholar]
- Shevelyov YY. Aurora, a non-mobile retrotransposon in Drosophila melanogaster heterochromatin. Mol Gen Genet. 1993;239:205–208. doi: 10.1007/BF00281619. [DOI] [PubMed] [Google Scholar]
- Kanamori Y, Hayashi H, Yamamoto MT. Molecular identification of the active ninja retrotransposon and the inactive aurora element in Drosophila simulans and D. melanogaster. Genes Genet Syst. 1998;73:385–396. doi: 10.1266/ggs.73.385. [DOI] [PubMed] [Google Scholar]
- Medhora MM, MacPeek AH, Hartl DL. Excision of the Drosophila transposable element mariner: identification and characterization of the Mos factor. EMBO J. 1988;7:2185–2189. doi: 10.1002/j.1460-2075.1988.tb03057.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petrov DA, Schutzman JL, Hartl DL, Lozovskaya ER. Diverse transposable elements are mobilized in hybrid dysgenesis in Drosophilavirilis. Proc Natl Acad Sci USA. 1995;92:8050–8054. doi: 10.1073/pnas.92.17.8050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berkeley Drosophila Genome Project: Release 2 Notes http://www.fruitfly.org/annot/release2.html
- National Center for Biotechnology Information http://ncbi.nlm.nih.gov
- Carvalho AB, Vibranovski MD, Carlson J, Celniker S, Hoskins R, Rubin GM, Sutton GG, Adams MD, Myers EW, Clark AG. Y chromosome and other heterochromatic sequences of the Drosophila melanogaster genome: how far can we go? Genetica. 2002. [DOI] [PubMed]
- Heitz E. Uber α- und β-Heterochromatin sowie Konstanz und Bau der chromomeren bei Drosophila. Biol Zentbl. 1934;54:588–609. [Google Scholar]
- Painter TS, Muller HJ. Parallel cytology and genetics of induced translocations and deletions in Drosophila. J Hered. 1929;20:287–298. [Google Scholar]
- Miklos GL, Yamamoto MT, Davies J, Pirrotta V. Microcloning reveals a high frequency of repetitive sequences characteristic of chromosome 4 and the β-heterochromatin of Drosophila melanogaster. Proc Natl Acad Sci USA. 1988;85:2051–2055. doi: 10.1073/pnas.85.7.2051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ashburner M, Misra S, Roote J, Lewis SE, Blazej R, Davis T, Doyle C, Galle R, George R, Harris N, et al. An exploration of the sequence of a 2.9-Mb region of the genome of Drosophila melanogaster: the Adh region. Genetics. 1999;153:179–219. doi: 10.1093/genetics/153.1.179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berkeley Drosophila Genome Project: Natural transposon elements in Release 3 sequence http://www.fruitfly.org/p_disrupt/TE.html
- Tu Z, Shao H. Intra- and inter-specific diversity of Tc3-like transposons in nematodes and insects and implications for their evolution and transposition. Gene. 2002;282:133–142. doi: 10.1016/s0378-1119(01)00841-1. [DOI] [PubMed] [Google Scholar]
- Berkeley Drosophila Genome Project http://www.fruitfly.org
- Galtier N, Gouy M, Gautier C. SEAVIEW and PHYLO_WIN: two graphic tools for sequence alignment and molecular phylogeny. Comput Appl Biosci. 1996;12:543–548. doi: 10.1093/bioinformatics/12.6.543. [DOI] [PubMed] [Google Scholar]
- WU BLAST 2.0 http://blast.wustl.edu/
- Portable Batch System http://www.openpbs.org
- Bowen NJ, McDonald JF. Genomic analysis of Caenorhabditis elegans reveals ancient families of retroviral-like elements. Genome Res. 1999;9:924–935. doi: 10.1101/gr.9.10.924. [DOI] [PubMed] [Google Scholar]
- Gordon D, Abajian C, Green P. Consed: a graphical tool for sequence finishing. Genome Res. 1998;8:195–202. doi: 10.1101/gr.8.3.195. [DOI] [PubMed] [Google Scholar]
- Morgenstern B. DIALIGN 2: improvement of the segment-to-segment approach to multiple sequence alignment. Bioinformatics. 1999;15:211–218. doi: 10.1093/bioinformatics/15.3.211. [DOI] [PubMed] [Google Scholar]
- Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994;22:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimura M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J Mol Evol. 1980;16:111–120. doi: 10.1007/BF01731581. [DOI] [PubMed] [Google Scholar]
- Felsenstein J. Phylip. Department of Genetics, University of Washington: Seattle, WA. 1993.
- Liao GC, Rehm EJ, Rubin GM. Insertion site preferences of the P transposable element in Drosophila melanogaster. Proc Natl Acad Sci USA. 2000;97:3347–3351. doi: 10.1073/pnas.050017397. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dominguez A, Albornoz J. Rates of movement of transposable elements in Drosophila melanogaster. Molec Gen Genet. 1996;251:130–138. doi: 10.1007/BF02172910. [DOI] [PubMed] [Google Scholar]
- Georgiev PG, Kiselev SL, Simonova OB, Gerasimova TI. A novel transposition system in Drosophila melanogaster depending on the Stalker mobile genetic element. EMBO J. 1990;9:2037–2044. doi: 10.1002/j.1460-2075.1990.tb07370.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vieira C, Biemont C. Selection against transposable elements in D.simulans and D.melanogaster. Genet Res. 1996;68:9–15. doi: 10.1017/s0016672300033838. [DOI] [PubMed] [Google Scholar]
- Biemont C, Gautier C. Localisation polymorphism of mdg-1, copia, I and P mobile elements in genomes of Drosophila melanogaster, from data of inbred lines. Heredity. 1988;1988:335–346. [Google Scholar]
- Belyaeva ES, Ananiev EV, Gvozdev V. Distribution of mobile dispersed genes (mdg-1 and mdg-3) in the chromosomes of Drosophila melanogaster. Chromosoma. 1984;90:16–19. [Google Scholar]
- Biemont C, Gautier C, Heizmann A. Independent regulation of mobile element copy number in Drosophila melanogaster inbred lines. Chromosoma. 1988;96:219–294. [Google Scholar]
- Whalen JH, Grigliatti TA. Molecular characterization of a retrotransposon in Drosophila melanogaster, nomad, and its relationship to other retrovirus-like mobile elements. Mol Gen Genet. 1998;260:401–409. doi: 10.1007/s004380050910. [DOI] [PubMed] [Google Scholar]
- Molto MD, Paricio N, Lopez-Preciado MA, Semeshin VF, Martinez-Sebastian MJ. Tirant: a new retrotransposon-like element in Drosophilamelanogaster. J Mol Evol. 1996;42:369–375. [PubMed] [Google Scholar]
- Viggiano L, Caggese C, Barsanti P, Caizzi R. Cloning and characterization of a copy of Tirant transposable element in Drosophila melanogaster. Gene. 1997;197:29–35. doi: 10.1016/s0378-1119(97)00217-5. [DOI] [PubMed] [Google Scholar]
- Vaury C, Chaboissier MC, Drake ME, Lajoinie O, Dastugue B, Pelisson A. The Doc transposable element in Drosophila melanogaster and Drosophila simulans: genomic distribution and transcription. Genetica. 1994;93:117–124. doi: 10.1007/BF01435244. [DOI] [PubMed] [Google Scholar]
- Di Nocera PP, Digan ME, Dawid IB. A family of oligo-adenylate-terminated transposable sequences in Drosophila melanogaster. J Mol Biol. 1983;168:715–727. doi: 10.1016/s0022-2836(83)80071-0. [DOI] [PubMed] [Google Scholar]
- Kholodilov NG, Bolshakov VN, Blinov VM, Solovyov VV, Zhimulev IF. Intercalary heterochromatin in Drosophila. III. Homology between DNA sequences from the Y chromosome, bases of polytene chromosome limbs, and chromosome 4 of D. melanogaster. Chromosoma. 1988;97:247–253. doi: 10.1007/BF00292968. [DOI] [PubMed] [Google Scholar]
- Berghella L, Dimitri P. The heterochromatic rolled gene of Drosophila melanogaster is extensively polytenized and transcriptionally active in the salivary gland chromocenter. Genetics. 1996;144:117–125. doi: 10.1093/genetics/144.1.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zabalou S, Alahiotis SN, Yannopoulos G. A three-season comparative analysis of the chromosomal distribution of P and hobo mobile elements in a natural population of Drosophila melanogaster. Hereditas. 1994;120:127–140. doi: 10.1111/j.1601-5223.1994.00127.x. [DOI] [PubMed] [Google Scholar]
- Harden N. Characterization of the transposable element FB-NOF in Drosophila melanogaster. In Department of Genetics Cambridge: University of Cambridge; 1989. p. 178.
- Merriman PJ, Grimes CD, Ambroziak J, Hackett DA, Skinner P, Simmons MJ. S elements: a family of Tc1-like transposons in the genome of Drosophila melanogaster. Genetics. 1995;141:1425–1438. doi: 10.1093/genetics/141.4.1425. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
A table showing the classification of transposable elements in the genus Drosophila