

Abstract
The genomic rate and distribution of effects of deleterious mutations are important parameters in evolutionary theory. The most detailed information comes from the work of Mukai and Ohnishi, who allowed mutations to accumulate on Drosophila melanogaster second chromosomes, shielded from selection and recombination by being maintained heterozygous in males. Averaged over studies, the estimated rate of nonlethal viability mutations per second chromosome per generation under an equal-effects model, UBM, was 0.12, suggesting a high genomic mutation rate. We have performed a mutation-accumulation experiment similar to those of Mukai and Ohnishi, except that three large homozygous control populations were maintained. Egg-to-adult viability of 72 nonlethal mutation-accumulation (MA) lines and the controls was assayed after 27–33 generations of mutation accumulation. The rate of decline in mean viability was significantly lower than observed by Mukai, and the rate of increase in among-line variance was significantly higher. Our UBM estimate of 0.02 is much lower than the previous estimates. Our results suggest that the rate of mutations that detectably reduce viability may not be much greater than the lethal mutation rate (0.01 in these lines), but the results also are consistent with models that include many mutations with very small effects.
The genomic rate and distribution of effects of deleterious mutations are critical parameters in models of the evolution of sex and recombination (1, 2), of the long-term viability of small populations (3, 4), and of molecular and phenotypic variation (5, 6). The classic experiments of Mukai (7, 8) and Ohnishi (9) on Drosophila melanogaster, in which mutations were allowed to accumulate on chromosomes shielded from selection and recombination, gave evidence that the rate of mutations with mild negative effects (<10%) on pre-adult viability is on the order of one per zygote per generation. Among other important consequences, such a high rate of mildly deleterious mutations could doom populations with effective sizes of a few hundred to extinction (3, 10).
More recent work, however, has given reason to question whether rates of mutations with detectable effects on fitness traits are typically as high as estimated by Mukai and Ohnishi. Experiments on Drosophila (11) and Caenorhabditis elegans (12) showed much slower declines in fitness traits on relaxation of selection against new mutations than observed by Mukai and Ohnishi. Although methodological differences could explain this discrepancy (13), another possibility is that the mean decline was overestimated in the earlier studies. This could have happened because the studies lacked concurrent controls for viability changes in the mutation-accumulation (MA) lines; viability instead was compared with base generation values that were obtained months earlier. If some change in experimental conditions occurred in the interim that depressed actual or apparent viability, both the mean decline and the genomic mutation rate would have been overestimated. Statistical arguments in favor of this hypothesis have been presented (14, 15).
We have performed a mutation-accumulation experiment using similar methods as Mukai and Ohnishi, with the difference that in addition to MA lines, we maintained three large control populations homozygous for the progenitor chromosome. Our evidence indicates that the control populations changed little or not at all in viability during the course of the experiment. To estimate the decline in mean viability of the MA lines, viability of the MA lines and control populations was measured simultaneously. In addition, because there is evidence that estimates of rates and effects of fitness mutations are sensitive to the conditions under which fitness assays are performed (16), we measured viability in treatments differing in temperature, degree of larval competition, and dietary composition. Estimates of genomic rates and effects of viability mutations were obtained by using traditional methods (8, 17) and maximum-likelihood analysis (14, 18).
MATERIALS AND METHODS
Culture Conditions.
Flies were reared in 2.5-cm diameter shell vials with 9 ml of cornmeal-molasses-agar medium (recipe modified from ref. 19) seeded with a few grains of live yeast. The temperature was 25°C unless otherwise indicated. Adults were handled under CO2 anesthesia.
MA and Control Lines.
The MA and control lines were made by using the balancer stock In(2LR)O, Cy dplvI Roi cn2 bw/In(2LR)bwVI; ve (20); the Cy Roi and bwVI chromosomes will be referred to as Cy and Pm, respectively. The X and third chromosomes of this stock, with the exception of the region around veinlet, were derived from an outbred population founded by pooling 33 isofemale lines that were collected in Raleigh, NC, in 1994. To initiate the experiment, a single Cy/+ male was crossed to balancer stock females; this male had a wild-type second chromosome derived from one of the isofemale lines and first and third chromosomes derived from the balancer stock. Cy/+ and Pm/+ offspring of the male (n = 107) were crossed singly to balancer stock females to establish the MA lines. Subsequently, each MA line was maintained by crossing two Cy/+ or Pm/+ males to five balancer stock females every 2- to 3-week generation (Fig. 1). The use of two males as opposed to one was unlikely to have affected estimates of mutational parameters (8). To establish the initial relative viability (see below) of the progenitor chromosome, additional Cy/+ male and female offspring of the original 107 males were crossed to each other in 33 vials at a density of 8 pairs per vial, and emerging Curly and wild-type offspring were counted. In addition, wild-type females and males collected from the same crosses were used to found three wild-type control populations. Each control population was maintained on 4-week generations at 18°C in 15 vials at a density of 10 pairs per vial with complete mixing of flies from the different vials each generation. The greater population size of the controls is expected to make selection against deleterious mutations more effective than in the MA lines, and their longer generation time also slows the rate of mutation accumulation compared with the MA lines.
Figure 1.

Crosses used to generate Cy/+ flies for the relative viability assays illustrated for one MA line (Left) and one control population (Right). In all crosses, females are listed first; all Cy/Pm females come from the balancer stock.
Viability Assays.
Viability of the MA lines was measured by the classic relative viability index (21), relative viability = 2 (Nwt)/(NCy + 1), where NCy and Nwt are the number of Curly and wild-type flies emerging from a cross between Cy/+ flies from a given MA line. An initial viability screen of all 99 surviving MA lines was conducted after 18–21 generations of mutation accumulation using methods similar to those for the “standard” treatment described below, with three crosses per line. Fourteen lines were identified as lethal and one as semilethal at this time (relative viabilities <5% and <20%, respectively); these lines were not tested further. By focusing on lines with relative viability >20%, we followed a similar procedure as the previous authors (7–9). More detailed examination of viability of the remaining 84 lines was conducted in two sets, with 40 lines assayed at generation (G) 27 and the remaining 44 assayed at G33. At G27, viability was assayed in three treatments: a standard treatment, with a density of six Cy/+ pairs per vial at 25°C; a low-density treatment, also at 25°C but with only two pairs per vial; and a low-temperature treatment, with six pairs per vial at 18°C. The six-pair and two-pair densities are roughly equivalent to those used by Mukai (7, 8) and Ohnishi (9), respectively. At G33, viability was measured in the standard treatment and an ethanol treatment, which was similar to the standard treatment except that the medium was supplemented with 10% ethanol (19). In all treatments, parent flies were transferred to new vials after 5 days and allowed to lay eggs for an additional 5 days before being discarded. Counts of flies emerging from both vials were pooled. In the 25°C (18°C) treatments, emerging flies were counted up to the 19th (35th) day after initiation of the vials. Expression of Curly was weak at 18°C and in some late-emerging flies at 25°C; all counts were therefore made under a stereomicroscope so that the presence or absence of Roi (Rough eye) could be determined in flies not showing an obvious Curly phenotype. At both G27 and G33, lines with <5% relative viability based on the first two crosses in the standard treatment were classified as lethals and not tested further (no semilethals were found).
The G27 and G33 assays were conducted in blocks, with all crosses in a given block set up on the same day. At G27 (G33), there were six (four) blocks consisting of one (two) crosses per treatment per MA line. The three control populations, and all six possible crosses between them (e.g., Cy/+A females × Cy/+B males, where +A and +B are second chromosomes from control populations A and B), were simultaneously assayed for viability, with two (four) replicate crosses per block and treatment for the pure control populations, and one (two) for each between-population cross, at G27 (G33).
Because the first and third chromosomes of the balancer stock were not isogenic, we used a crossing scheme to produce Cy/+ parental flies for the viability assays that ensured that most variation in genetic backgrounds was within, rather than among, lines (Fig. 1). For each MA line, Cy/+ flies were collected from a minimum of eight crosses of five balancer stock females to Cy/+ or Pm/+ males (Fig. 1). Cy/+ flies from different crosses necessarily had low coancestries for the X and third chromosomes; therefore the primary effect of variation in genetic backgrounds, if any, would be to inflate the within-line variance in viability. Within-line variances in this study (data not shown) were similar to those reported by Mukai (7), who used an isogenic balancer stock. Additional evidence that variation in genetic backgrounds did not affect the results is presented below.
Contamination Checks.
After the G27 and G33 viability assays, homozygous stocks were produced from each viable, fertile MA line by collecting wild-type progeny from intercrosses of Cy/+ flies. A sample of 17 of the homozygous MA lines, as well as all 3 control populations, were examined for insertion sites of roo transposable elements (22, 23). All lines shared a common set of 19 roo sites spanning the length of the second chromosome (cytological sites 22A to 59A). These results give strong evidence that none of the examined lines was contaminated (24) and indicate that the balancer was effective in suppressing recombination. The veinlet marker present in all lines provided an additional check for contamination; no ve+ flies were observed.
Statistical Analysis.
Mutational parameters were estimated by the traditional Bateman–Mukai formulae (8, 17) and by maximum-likelihood analysis (14, 18).
Given the assumption of equal mutational effects, the Bateman–Mukai formulae allow estimation of UBM (the rate of mutations affecting viability per haploid second chromosome per generation) and SBM (the effect of the mutations):
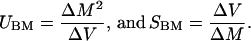 |
Here, ΔM is the decline in average viability of the MA lines per generation, and ΔV is the increase in among-line variance in viability per generation. ΔM was estimated from the difference between control and MA-line viabilities and ΔV from the variance component among MA lines in an ANOVA. Lethal lines were excluded from the calculations, and both quantities are expressed on a scale in which the mean viability of the controls equals one. Bootstrap 95% confidence intervals for ΔM, ΔV, UBM, and SBM were calculated by using the percentile method (25) based on 20,000 bootstrap samples. Details of the bootstrap procedure are available on request.
As an alternative method for estimating ΔV, we used the covariance between mean viability of MA lines in the initial screen at G18–G21 and mean viability observed later (G27 or G33). For a set of lines initially tested at generation i (i = 18–21), the covariance between the initial mean and later mean divided by i estimates ΔV. Means from G27 and G33 were based on pooled data from the 25°C treatments (see below for rationale). ΔV was calculated separately for each set of lines tested in the same generation (G18–G21, n = 15–20 lines per generation), and the four estimates were averaged to produce a single estimate. Bootstrap confidence intervals were calculated by resampling the lines from each of generations 18–21.
Maximum-Likelihood Analysis.
To relax the assumption of equal mutation effects, estimates of the mutation rate U were made by using maximum likelihood analysis under the assumption that mutation effects come from a γ distribution, g(a|α, β), for which the parameters α and β specify the scale and shape of the distribution, respectively. By using maximum likelihood, the fit to the data of different kinds of distributions of mutation effects can be compared. β → ∞ is the case of equal effects, and as β → 0 the distribution becomes increasingly leptokurtic. Residual values were assumed to be normally distributed with mean M and variance σE2. In the basic analysis, U, α, and β, along with M and σE2 were estimated. The likelihood equation is similar to Eq. 2 in ref. 18 except that replicate information is included:
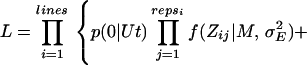 |
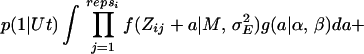 |
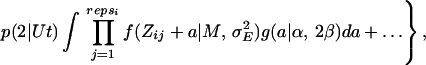 |
1 |
where repsi is the number of replicates within line i, t is the generation number, p(x| Ut) is the probability of x mutations if the distribution of mutation numbers is Poisson with parameter Ut, Zij is the viability for replicate j from line i, and f (z| M, σE2) is the normal density function. Likelihood was evaluated numerically and maximized as described elsewhere (18).
Two other models with additional parameters were investigated: (i) γ plus nonmutational change (γ + k), used to model the situation where the MA and control line means differ by a nonmutational effect (14), is equivalent to evaluating Eq. 1 with different means for the control and MA lines. If data from independent sets of lines maintained for different numbers of generations are simultaneously analyzed, the equation is similar to Eq. 1, except that the term f(Zij + a| M, σE2) becomes f(Zij + a + kt| M, σE2), and f(Zij| M, σE2) becomes f(Zij + kt| M, σE2). The parameter k is therefore the nonmutational effect per generation and is estimated along with the parameters described above. (ii) Mixed γ distribution + equal effects. A proportion p of mutations was assumed to have a constant deleterious effect ɛ, and the remaining proportion was assumed to have a γ distribution as above (cf. ref. 14). Terms in Eq. 1 are expanded to account for the binomial probabilities of equal and γ-distributed effects, and the parameters p and ɛ are estimated simultaneously with the other parameters.
RESULTS
Lethal Mutations.
In the initial viability assays at G18–G21, 14 of the 99 MA lines were classified as lethal and 1 as semilethal. Of the remaining lines, 2 of 40 retested at G27 and 10 of 44 retested at G33 had accumulated a lethal mutation since G18–G21. Based on these figures, the rate of lethal plus semilethal mutations per haploid second chromosome per generation can be estimated by using maximum likelihood analysis (26), assuming a constant rate per generation. The resulting estimate of 0.010, with two log-likelihood support limits (analogous to 95% confidence limits) of 0.007–0.015, is significantly greater than the typically reported values of 0.004–0.006 for the second chromosome (7, 8, 27, 28).
Control Populations.
The three control populations and hybrids between them were measured for relative viability at the same time as the G27 and G33 MA lines. If the viability of some or all of the control populations had declined appreciably because of mutation accumulation, some variation among them in viability would likely result. At both G27 and G33, there was no significant variation among the control populations or their hybrids in viability (Table 1). Additional evidence that viability of the control populations did not change appreciably comes from comparing base-generation viability with viability measured in the control populations at G27 and G33. Mean relative viability of all control crosses in the standard treatment (six Cy/+ pairs per vial, 25°C) at G27 and G33 were 0.82 and 0.81, respectively. Mean viability in the base generation assay (eight pairs per vial, 25°C) was 0.84. These figures are not significantly different by ANOVA (F2,194 = 0.78, P > 0.45). To determine whether the slightly higher density used in the base generation might have depressed viability, viability of the three control populations was measured at each density 11 months after the G33 MA line assays, with ≈22 crosses per population per density. No significant effect of density was found (P > 0.3), and mean viability (0.85) was similar to the earlier values.
Table 1.
Analysis of variance of relative viability of control populations at MA line G27 and G33
| Source | Denominator mean square | MA line G27
|
MA line G33
|
||
|---|---|---|---|---|---|
| df | P | df | P | ||
| Group† | G × B | 5 | >0.4 | 5 | >0.9 |
| Treatment | T × B | 2 | >0.3 | 1 | >0.2 |
| Block | Synthetic‡ | 5 | >0.9 | 3 | >0.9 |
| G × T | G × T × B | 10 | >0.3 | 1 | >0.5 |
| T × B | G × T × B | 10 | ∗∗ | 3 | >0.3 |
| G × B | G × T × B | 25 | >0.6 | 15 | >0.5 |
| G × T × B | residual | 50 | >0.6 | 15 | >0.3 |
| Residual | 104 | 138 | |||
Block and interactions involving block were considered random, and other effects fixed.
∗∗, 0.001 < P < 0.01.
Three control populations, plus three sets of hybrids between the populations. Reciprocal crosses were pooled.
Satterthwaite approximate F-test used (29).
Genotype–Environment Interactions.
The G27 and G33 viability data sets (MA lines + controls) were analyzed by using ANOVA (Table 2). Because there was no significant variation among the control populations in viability at either time, all control crosses were treated as belonging to a single line. There was significant treatment-by-line interaction only at G27, caused primarily by two MA lines that had much lower viability at 18°C than at 25°C (Fig. 2). With the 18°C treatment excluded, the interaction was no longer significant, indicating that mutations affected viability similarly in the two remaining treatments (two and six pairs at 25°C). At G33, addition of 10% ethanol to the medium significantly reduced viability compared with unsupplemented medium (Table 2); mean viability was about 9% lower in the ethanol treatment. Nonetheless, there was no significant treatment-by-line interaction, indicating that mutations again affected viability similarly in the two treatments, both of which were at 25°C.
Table 2.
ANOVA of relative viability of MA lines and the pooled controls at G27 and G33 of mutation accumulation
| Source | Generation
|
|||||
|---|---|---|---|---|---|---|
| G27
|
G27, without 18°C treatment
|
G33
|
||||
| df | P | df | P | df | P | |
| Treatment | 2 | >0.6 | 1 | >0.1 | 1 | ∗ |
| Line | 38 | ∗∗∗ | 38 | ∗∗∗ | 34 | ∗∗∗ |
| Block | 5 | >0.5 | 5 | >0.2 | 3 | ∗ |
| T × L | 76 | ∗∗∗ | 38 | >0.3 | 34 | >0.5 |
| T × B | 10 | ∗ | 5 | >0.5 | 3 | ∗ |
| L × B | 190 | >0.9 | 190 | >0.3 | 102 | >0.05 |
| Residual | 563 | 311 | 541 | |||
All effects except treatment were considered random. Main effects were tested using Satterthwaite approximate F-tests (29); interactions were tested over the residual mean squares.
∗, 0.05 < P < 0.01; ∗∗∗, P < 0.001.
Figure 2.

Distributions of relative viabilities of the MA lines.
Estimates of Mutational Parameters.
Because of the lack of genotype–environment interaction involving the treatments at 25°C, data from these treatments were pooled. Based on the combined G27 and G33 data, relative viability at 25°C declined an average of 0.24% per generation (Table 3), and similar estimates were obtained when data from G27 and G33 were analyzed separately. The mean decline is significantly lower than the point estimates obtained by Mukai and coworkers (7, 8). In contrast, the mutational variances estimated from the pooled 25°C data and from the G33 data alone were significantly higher than the point estimates from all three previous studies. Reflecting these differences, UBM estimates from the 25°C data are significantly lower than the previous point estimates, and SBM estimates are significantly higher. Estimates of mutational parameters from the G27 18°C dataset (data not shown) were similar to those at 25°C but had broader confidence limits reflecting the smaller sample sizes.
Table 3.
Estimates of the decline in mean viability per generation among nonlethal lines (ΔM), mutational variance for viability (ΔV), rate of mutations affecting viability per haploid second chromosome per generation under an equal-effects model (UBM), and effect of the mutations (SBM)
| Data set | ΔM, % | ΔV × 104 | UBM | SBM |
|---|---|---|---|---|
| 25°C, G27 + G33 | 0.24 (0.16–0.31) | 2.7 (1.6–3.6) | 0.021 (0.011–0.038) | 0.113 (0.073–0.162) |
| 25°C, G27 | 0.24 (0.13–0.34) | 1.9 (0.80–3.0) | 0.029 (0.012–0.066) | 0.081 (0.040–0.134) |
| 25°C, G33 | 0.23 (0.12–0.35) | 3.4 (1.8–4.9) | 0.016 (0.0051–0.036) | 0.147 (0.081–0.262) |
| Mukai 1964 (7) | 0.38 | 1.0 | 0.14 | 0.027 |
| Mukai et al. 1972 (8) | 0.40 | 0.94 | 0.17 | 0.023 |
| Ohnishi 1977 (9) | 0.17 | 0.51 | 0.06 | 0.030 |
Bootstrap 95% confidence intervals are given in parentheses. 25°C estimates are based on data pooled across treatments. Estimates from the other studies are those based on excluding “severely deleterious” lines (usually, those with viability <50% of the controls); this is the most relevant comparison, because there were no lines with <65% viability of the controls at 25°C (Fig. 2).
Because the balancer stock used was not isogenic, variation in genetic backgrounds of the MA lines could conceivably have inflated the ΔV estimates. As a check on whether background effects might have influenced the results, we estimated ΔV from the covariance between mean viability of MA lines in the initial screen at G18–G21 and mean viability in the 25°C treatments at G27 and G33. Because at least six generations of backcrossing to the balancer stock occurred between the two measurements, the genetic backgrounds of the lines at the two times should have been essentially independent. ΔV estimated by this method was 3.2 × 10−4, with 95% confidence interval 1.5–4.7 × 10−4, similar to the ANOVA-based estimate in Table 3.
Maximum-Likelihood Estimates.
Mutation rates were estimated by maximum likelihood under the assumptions that mutation effects conform to a γ distribution, and that residual variances are normally distributed. We report estimates for the combined G27 + G33 25°C data set; estimates from the separate G27 and G33 25°C data sets and from the G27 18°C data set gave essentially similar conclusions. The distribution of control-line viability values did not differ significantly from normal (Shapiro–Wilk test, P = 0.38, n = 327), supporting the assumption of normal residual variances.
With the shape parameter, β, set to infinity (corresponding to equal mutational effects) likelihood estimates of mutation rates and effects were similar to those obtained by the Bateman–Mukai method (Table 4). Finite values of β gave significantly higher likelihoods, however, meaning that the equal-effects model can be rejected. Likelihoods changed little for values of β ≤ 1, so it is not possible to distinguish between the case of moderate variation in mutational effects (β = 1, corresponding to a coefficient of variation of 100%), and an extremely leptokurtic distribution of effects (β → 0). Because estimated U → ∞ as β → 0, it is not possible to put an upper bound on the mutation rate without making assumptions about β. For example, if 0.25 is taken as the lower limit for β (upper limit on coefficient of variation of effects = 200%), then the upper support limit for U is 0.20. The true mutation rate could be much higher, however.
Table 4.
Maximum likelihood estimates of mutational parameters for the combined G27 + G33 25°C data set, under a model assuming a γ distribution of mutational effects
| β | U | E(a) | Fit | P |
|---|---|---|---|---|
| →∞ | 0.024 (0.010–0.047) | 0.096 (0.061–0.140) | −2.2 | <0.05 |
| 2 | 0.029 (0.012–0.060) | 0.078 (0.045–0.140) | −0.2 | >0.5 |
| 1 | 0.039 (0.018–0.080) | 0.059 (0.034–0.110) | 0.0 | >0.9 |
| 0.5 | 0.060 (0.027–0.120) | 0.040 (0.023–0.076) | 0.0 | >0.9 |
| 0.25 | 0.100 (0.045–0.200) | 0.024 (0.013–0.049) | 0.0 | >0.9 |
β, shape parameter; U, mutation rate per haploid second chromosome per generation; E(a), average effect of mutations. Maximum-likelihood estimates of U and E(a) are given for the fixed values of β indicated, with two log-likelihood support limits (analogous to 95% confidence intervals) in parentheses. Fit is measured as the difference in log-likelihood from the best fitting model. Probabilities <0.05 indicate poor fit of the given β.
The likelihood analysis gives a lower bound for U of about 0.01 (Table 4), similar to that found by bootstrapping UBM. The analysis also suggests an upper bound for the average effect of a mutation, E(a), of 0.14.
In his reanalysis of data of Mukai et al. (8) and Ohnishi (9), Keightley (14) found that allowing a nonmutational change of MA-line viabilities resulted in significantly improved fit relative to the γ-distribution model. This was not the case in this study (P > 0.6, likelihood ratio test). In contrast, allowing an additional class of mutations with equal effects (γ + equal model) resulted in a significant improvement in fit (P = 0.03). The improvement was much smaller than for the Mukai and Ohnishi data sets (14), however.
DISCUSSION
Our estimate of the decline in mean viability caused by second-chromosome mutations based on the 72 MA lines tested at 25°C is 0.24% per generation. This ΔM estimate, although significantly lower than the estimates of Mukai (7, 8), is not significantly different from that of Ohnishi (9) and does not differ dramatically from the mean of the previous estimates (=0.32). In contrast, our estimate of the mutational variance, ΔV, is about three times as high as the mean of the Mukai–Ohnishi estimates (Table 3). This could be the result of a higher rate of moderately deleterious mutations (effects ≈10–20%) in our study than in the previous studies, an explanation that would be consistent with the 2-fold higher rate of lethal mutations that we observed. The most striking difference between our results and the previous results, however, are in our estimates of UBM and SBM, the rate and effects of viability mutations under an equal-effects model. Our UBM estimate is ≈12–35% those obtained by Mukai and Ohnishi, whereas our SBM estimate is ≈4- to 5-fold higher than theirs.
Our UBM estimate of 0.02 is likely to be a considerable underestimate of the overall rate of deleterious mutations on the second chromosome for at least two reasons. First, it is well known that UBM underestimates the true mutation rate when the equal-effects assumption is violated, and we were able to reject this assumption by using a likelihood approach. Second, the estimate does not include the effects of mutations that affect components of fitness other than egg-to-adult viability or the effects of mutations that affect viability but act maternally. Nonetheless, the same biases apply to the Mukai and Ohnishi estimates. Furthermore, it is of some consequence whether one’s underestimate of the genomic deleterious mutation rate is 0.1 (our estimate, scaled to the entire diploid genome) or almost one (the Mukai estimates, similarly scaled). Only if we accept the latter figure can we be confident that the true mutation rate is well over one, and it is such a high rate that is necessary for many of the hypothesized consequences of mutations for populations such as the evolution of sexual reproduction (1). Therefore, it is important to consider possible explanations for the difference between our results and the previous results.
Environmental Differences.
Kondrashov and Houle (16) argued that fitness assays performed under relatively harsh conditions are likely to produce higher estimates of the genomic mutation rate than those performed under benign conditions. Although this may be correct, it does not seem to be able to explain the difference between our results and those of Mukai and Ohnishi. The viability assay we used was similar to theirs, and our estimates were not affected by moderate variation in density and temperature or by addition of ethanol to the medium, even though the latter manipulation significantly reduced viability. In addition, relative viability of the progenitor chromosome (0.81–0.85) was somewhat lower than in the Mukai and Ohnishi studies (usually 0.90 or higher), suggesting that our conditions might have been more rather than less harsh.
Decline in Viability of the Control Populations.
If our control populations had declined in viability during the experiment, we would have underestimated ΔM, the rate of mean decline in the MA lines, and hence underestimated the genomic mutation rate. There is reason to expect that some viability decline of the control populations might have occurred. In a large, initially homozygous population, the frequencies of mutant alleles are expected to rise slowly until the equilibrium frequencies of u/hs are reached, where u is the mutation rate and hs is the selection coefficient against heterozygotes. We have therefore calculated UBM under the assumption that the control populations declined in viability by various amounts (Table 5). If the control populations had declined in viability at half the rate of the MA lines, the corrected UBM estimate would still be lower than the previous estimates. The corrected UBM estimate would approach those of Mukai only if the control populations declined in viability 90–95% as fast as the MA lines. Such a rapid decline seems unlikely for two reasons. First, it would imply that selection was almost completely ineffective in the control populations even though they consisted of 300 flies each, and probably had effective sizes on the order of 100 (30). Second, it would require that the control populations had declined in viability by at least 11% by MA line G33 (Table 5). For this decline to have gone undetected, some environmental change would have necessarily raised the apparent viability of the controls by approximately the same amount, but our results suggest that the relative viability measure is not highly sensitive to minor variations in conditions. Block and treatment effects generally were weak or absent (Tables 1 and 2), and the maximum difference in viability between any pair of treatments (0% vs. 10% ethanol) was 9%. We emphasize that control-population viabilities measured at the beginning of the experiment, at G27 and G33 of the MA lines, and 11 months later did not differ significantly, with all means between 0.81 and 0.85. The most parsimonious interpretation of these results is that control viabilities changed little during the course of the experiment.
Table 5.
Effect of control population viability declines on the parameter estimates
| ΔMCONT/ ΔMMA* | ΔM,† % | UBM† | SBM† | G33, % decline‡ |
|---|---|---|---|---|
| 0.25 | 0.28 | 0.030 | 0.095 | 1.5 |
| 0.50 | 0.35 | 0.045 | 0.077 | 3.8 |
| 0.75 | 0.46 | 0.079 | 0.058 | 7.6 |
| 0.90 | 0.57 | 0.120 | 0.047 | 11.2 |
| 0.95 | 0.61 | 0.141 | 0.044 | 12.8 |
Per-generation viability decline of control populations, relative to that of the MA lines.
Estimates after correcting for control decline. For example, at MA G27, 17 control generations had elapsed. Assuming ΔMCONT/ΔMMA = 0.5, the corrected ΔM estimate is given by 0.24/[1 − (17/27)(0.5)]; 0.24 is the uncorrected estimate from Table 3, and the second term in the brackets corrects for the underestimation of base generation viability caused by the control decline. Performing a similar calculation for the G33 data and averaging the results gives the value shown, 0.35. The corrected UBM and SBM estimates are calculated using the corrected ΔM and ΔV from Table 3; the latter depends on data from the MA lines only.
Percent by which the viability of the control populations would have declined by MA G33, calculated as 22 (number of control generations) times the product of the first two columns.
Overestimation of ΔM in the Previous Studies.
If Mukai and Ohnishi had overestimated ΔM, they also would have overestimated UBM. Keightley (14) suggested that such overestimation could have been caused by evolution of higher viability of the Cy balancer chromosome. Our results give no support for this hypothesis; balancer evolution would have caused the control mean viabilities to decline, which we did not observe. An alternative possibility is errors in distinguishing Cy and wild-type individuals. In our MA lines, a fraction of individuals with the Cy, Roi balancer showed such weak expression of Cy that they were identifiable only by the presence of Roi, and Cy expression is known to be affected by a variety of genetic and environmental influences (20). An investigator scoring many flies over the course of months might gradually become better at recognizing individuals with subtle Cy expression, thus leading to an apparent decrease in the relative viability of wild-type flies.
Among-Strain Differences in the Rates and Distributions of Effects of Mutations.
Among-strain differences in the rates and distributions of effects of mutations is the simplest explanation for the difference between our results and those of the previous studies, and the only one for which some direct evidence exists, in that we observed a higher lethal mutation rate and higher mutational variance than Mukai and Ohnishi. Unusually high activity of one or more families of transposable elements could be responsible for both the high lethal rate and the high mutational variance; preliminary results (S.V.N. and J.D.F., unpublished data) indicate that copia transposed at a high frequency in the MA lines. High activity of transposable elements, however, could not explain the lower mean decline we observed compared with the two Mukai studies. If among-strain differences entirely account for the difference between our results and those from previous studies, one would have to postulate that there was simultaneously a higher frequency of mutations with relatively large effects (which disproportionally affect ΔV) and a lower frequency of small-effect mutations in our study compared with the previous studies.
Conclusions.
Although we could put a lower bound of 0.01 on the rate of second chromosome mutations with sublethal effects on viability, it was not possible to put an upper bound on the rate. Likelihoods under the γ-distribution model became flat as β → 0 (increasingly leptokurtic distribution of effects) and U → ∞ (Table 4). Although we could reject an equal-effects model, little other information about the distribution of mutational effects could be gleaned from the likelihood analysis. U and β are likely to become confounded unless sample sizes are very large, the number of mutations per MA line are low, and the distribution of effects is not too leptokurtic, making it difficult to make inferences about the distribution of mutational effects (31). Because the distribution of effects in our study could have been highly leptokurtic, with many mutations of very small effect, our results do not rule out the possibility that the rate of second-chromosome viability mutations is orders of magnitude higher than 0.01.
Nonetheless, our results and those of other recent studies and reanalyses (11, 12, 14, 15, 32) give reason to question whether the genomic mutation rate for fitness detectable in experiments with invertebrates is typically as high as the Mukai–Ohnishi estimates. This issue will require additional careful studies to resolve.
Acknowledgments
We thank T. F. C. Mackay for providing laboratory space and helpful discussions and B. Charlesworth for helpful comments on an earlier version of the manuscript. This project was supported by National Science Foundation Grants DEB-9317754 and DEB-9707470 (J.D.F. and S.L.H.), the Royal Society (P.D.K.), and University of California, Davis start-up funds (S.V.N.).
ABBREVIATIONS
- G
generation
- MA
mutation accumulation
References
- 1.Kondrashov A S. Nature (London) 1988;336:435–441. doi: 10.1038/336435a0. [DOI] [PubMed] [Google Scholar]
- 2.Charlesworth B. Genet Res. 1990;55:199–221. doi: 10.1017/s0016672300025532. [DOI] [PubMed] [Google Scholar]
- 3.Lande R. Conserv Biol. 1995;9:782–791. [Google Scholar]
- 4.Lynch M, Conery J, Burger R. Am Nat. 1995;146:489–518. [Google Scholar]
- 5.Charlesworth B, Morgan M T, Charlesworth D. Genetics. 1993;134:1289–1303. doi: 10.1093/genetics/134.4.1289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Barton N H, Turelli M. Annu Rev Genet. 1989;23:337–370. doi: 10.1146/annurev.ge.23.120189.002005. [DOI] [PubMed] [Google Scholar]
- 7.Mukai T. Genetics. 1964;50:1–19. doi: 10.1093/genetics/50.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Mukai T, Chigusa S I, Mettler L E, Crow J F. Genetics. 1972;72:335–355. doi: 10.1093/genetics/72.2.335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ohnishi O. Genetics. 1977;87:529–545. doi: 10.1093/genetics/87.3.529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Schultz S T, Lynch M. Evolution. 1997;51:1363–1371. doi: 10.1111/j.1558-5646.1997.tb01459.x. [DOI] [PubMed] [Google Scholar]
- 11.Fernández J, López-Fanjul C. Genetics. 1996;143:829–837. doi: 10.1093/genetics/143.2.829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Keightley P D, Caballero A. Proc Natl Acad Sci USA. 1997;94:3823–3827. doi: 10.1073/pnas.94.8.3823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Shabalina S A, Yampolsky L Y, Kondrashov A S. Proc Natl Acad Sci USA. 1997;94:13034–13039. doi: 10.1073/pnas.94.24.13034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Keightley P D. Genetics. 1996;144:1993–1999. doi: 10.1093/genetics/144.4.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.García-Dorado A. Evolution. 1997;51:1130–1139. doi: 10.1111/j.1558-5646.1997.tb03960.x. [DOI] [PubMed] [Google Scholar]
- 16.Kondrashov A S, Houle D. Proc R Soc Lond B. 1994;258:221–227. doi: 10.1098/rspb.1994.0166. [DOI] [PubMed] [Google Scholar]
- 17.Bateman A J. Int J Radiat Biol. 1959;1:170–180. [Google Scholar]
- 18.Keightley P D, Ohnishi O. Genetics. 1998;148:753–766. doi: 10.1093/genetics/148.2.753. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Fry J D, Heinsohn S L, Mackay T F C. Evolution. 1996;50:2316–2327. doi: 10.1111/j.1558-5646.1996.tb03619.x. [DOI] [PubMed] [Google Scholar]
- 20.Lindsley D L, Zimm G G. The Genome of Drosophila melanogaster. San Diego: Academic; 1992. [Google Scholar]
- 21.Haldane J B S. J Genet. 1956;54:294–296. [Google Scholar]
- 22.Shrimpton A E, Montgomery E A, Langley C H. Genetics. 1986;114:125–135. doi: 10.1093/genetics/114.1.125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Nuzhdin S V, Pasyukova E G, Dilda C A, Zeng Z-B, Mackay T F C. Proc Natl Acad Sci USA. 1997;94:9734–9739. doi: 10.1073/pnas.94.18.9734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Charlesworth B, Lapid A, Canada D. Genet Res. 1992;60:103–114. doi: 10.1017/s0016672300030792. [DOI] [PubMed] [Google Scholar]
- 25.Mooney C Z, Duval R D. Bootstrapping: A Nonparametric Approach to Statistical Inference. Newbury Park, CA: Sage Publications; 1993. [Google Scholar]
- 26.Weir B S. Genetic Data Analysis II. Sunderland, MA: Sinauer; 1996. [Google Scholar]
- 27.Crow J F, Simmons M J. In: The Genetics and Biology of Drosophila. Ashburner M, Carson H L, Thompson J N, editors. 3c. London: Academic; 1983. pp. 1–35. [Google Scholar]
- 28.Ohnishi O. Genetics. 1977;87:519–527. doi: 10.1093/genetics/87.3.519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.SAS Institute, Inc. SAS/STAT User’s Guide, Version 6. Cary, NC: SAS Inst.; 1989. [Google Scholar]
- 30.Crow J F, Morton N E. Evolution. 1955;9:202–214. [Google Scholar]
- 31.Keightley P D. Genetics. 1998;150:1283–1293. doi: 10.1093/genetics/150.3.1283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Keightley P D, Caballero A, García-Dorado A. Curr Biol. 1998;8:R235–R237. doi: 10.1016/s0960-9822(98)70148-4. [DOI] [PubMed] [Google Scholar]