

Abstract
Although sodium channel blockers are effective analgesics in neuropathic pain, their effectiveness in inflammatory pain has been little studied. Sodium channels are substantially up-regulated in inflamed tissue, which suggests they play a role in maintenance of chronic inflammatory pain. We have examined the effects of sodium channel blockers on mobility, joint hyperalgesia and inflammation induced by complete Freund's adjuvant injected in one ankle joint of adult rats. The clinically effective sodium channel blocker, mexiletine, was compared with crobenetine (BIII 890 CL), a new, highly use-dependent sodium channel blocker.
Rats were treated for 5 days, starting on the day of induction of arthritis and were tested daily for joint hyperalgesia, hind limb posture and mobility. At post-mortem, joint stiffness and oedema were assessed. Dose response curves were constructed for each test compound (3 – 30 mg kg day−1). Control groups were treated with vehicle or with the non-steroidal anti-inflammatory drug, meloxicam (4 mg kg day−1 i.p.).
Both sodium channel blockers produced dose dependent and significant reversal of mechanical joint hyperalgesia and impaired mobility with an ID50 of 15.5±1.1 mg kg day−1 for crobenetine and 18.1±1.2 mg kg day−1 for mexiletine. Neither compound affected the responses of the contralateral non-inflamed joint, nor had any effect on swelling and stiffness of the inflamed joint.
We conclude that sodium channel blockers are analgesic and anti-hyperalgesic in this model of arthritis. These data suggest that up regulation of sodium channel expression in primary afferent neurones may play an important role in the pain and hyperalgesia induced by joint inflammation.
Keywords: Hyperalgesia, pain, arthritis, mexiletine, compete Freund's adjuvant
Introduction
Sodium channel blockers are widely used clinically to treat neuropathic pain (e.g. Tanelian & Brose, 1991; Sindrup & Jensen, 1999). The regulation of sodium channel subunit expression and properties of sodium channels in dorsal root ganglion neurones in animals with peripheral nerve damage has been extensively studied (for reviews see Waxman et al., 1999; Baker & Wood, 2001). However, sub-acute or chronic inflammation also produces profound changes in the excitability of primary afferent neurones innervating the inflamed tissue (Schaible & Schmidt, 1996). These changes underline the initiation and maintenance of chronic inflammatory pain states. Recent evidence suggests that post-translational modifications or abnormal expression of sodium channels in dorsal root ganglion neurones occurs after tissue inflammation (see Waxman et al., 1999; Baker & Wood, 2001).
Inflammatory mediators such as prostaglandins or serotonin can modulate the properties of tetrodotoxin(TTX)-resistant sodium channels (e.g. England et al., 1996). Hindpaw inflammation with carrageenan or complete Freund's adjuvant results in an increased expression of mRNA coding for TTX-resistant sodium channels (Tanaka et al., 1998; Tate et al., 1998) in dorsal root ganglion neurones innervating the inflamed tissue. Consistent with these changes, the TTX-resistant sodium current in these neurones also increases (Tanaka et al., 1998). A marked increase in immunoreactivity for sodium channels of all types in dorsal root ganglion neurones is observed when complete Freund's adjuvant was injected into their projection field (Gould et al., 1998; 1999). The up-regulation of sodium channels reaches a maximum at 24 h post injection and lasts for more than 2 months (Gould et al., 1998; 1999). The intensity of the response parallels the mechanical hyperalgesia expressed in the inflamed paw.
The effect of sodium channel blockade on inflammatory pain has been little studied. However, genetic manipulation of the TTX-resistant SNS/PN3 sodium channel reduces inflammatory pain behaviour. Knockdown of the channel with antisense oligonucleotides has been shown to reduce hyperalgesia in rats injected with complete Freund's adjuvant (Porreca et al., 1999), and mice with a disruption of the gene encoding the SNS/PN3 channel show delayed onset of hyperalgesia after intraplantar carrageenan (Akopian et al., 1999). The present studies were designed to explore the possible analgesic activity of sodium channel blockers in a model of arthritis in the rat. The effects of two sodium channel blockers on joint hyperalgesia, on the spontaneous pain-related behaviour and on joint inflammation of rats with adjuvant-induced monoarthritis were compared. The compounds tested were mexiletine, a clinically effective sodium channel blocker which has a 26-fold higher affinity for the inactivated over the resting form of sodium channels (Weiser et al., 1999) and crobenetine (BIII 890 CL), a novel compound which has a 230-fold higher affinity (Carter et al., 2000). These compounds block both TTX-sensitive and TTX-insensitive sodium channels (Weiser et al., 1999; Carter et al., 2000; Krause et al., 2000).
Methods
Adult female Wistar rats (from the colony maintained in the University of Alcalá Animal House) with weights of 185 – 270 g were used. The animals were housed in groups of six per cage for at least 2 weeks before the start of the experiment. Each cage contained one rat per treatment group. Monoarthritis was induced by intraarticular injection of 0.05 ml of complete Freund's adjuvant (CFA) into the right ankle (Butler et al., 1992) under brief halothane anaesthesia (4% in 100% O2). Dose-response curves for the two test compounds (crobenetine and mexiletine) were constructed in separate experiments. In the first experiment, crobenetine (3, 10 or 30 mg kg day−1) or vehicle (5% xylitol solution) was administered s.c. (dose volume=15 ml kg−1) daily for 5 days starting on the day of induction of the arthritis (n=6 per group). As a positive control, a group of six rats received intraarticular CFA and daily injections of 4 mg kg−1 i.p. meloxicam (suspended in 1% methylcellulose). This dose and route was chosen from a previous study of the effects of meloxicam in this model of arthritis (Laird et al., 1997) as a maximally effective dose so as to provide a clear positive control group. A further control group of six rats received an intraarticular injection of mineral oil (CFA vehicle), and daily s.c. injections of 15 ml kg−1 xylitol solution. Behavioural tests were performed immediately before induction of the arthritis, and on the 2nd, 4th and 5th days, 15 min after drug administration. In the second experiment, mexiletine (3, 10 or 30 mg kg day−1) or vehicle (distilled water) was administered s.c. (dose volume=1 ml kg−1) daily for 5 days starting on the day of induction of the arthritis (n=6 per group) and an identical protocol was followed, including both the positive and negative control groups.
The experimental protocol was based on the pharmacokinetic profile of the test compounds. Maximal plasma levels of crobenetine and mexiletine are achieved less than 15 min after s.c. administration in rats, and maintained at these levels for around 2 h after dosing. The half-life of crobenetine in plasma after i.v. dosing is 2.6 h (V. Häselbarth and A.J. Carter, unpublished observations).
The rats were tested daily for responses to movement of each ankle joint (‘ankle-bend test'; Danziger et al., 1999). The rat was held by the experimenter, and five successive extensions and five successive flexions of the contralateral, non-injected joint were applied manually, and the number of vocalizations evoked by these 10 movements noted. The test was then repeated extending and flexing the injected joint. The stance and mobility of the rats whilst they moved freely in a large empty cage were also assessed daily according to adaptations of the scales of Butler et al. (1992). The stance scale assigned scores as follows; 1=paw lifted continuously, 2=paw touching but with no weight-bearing, 3=some weight bearing on paw, 4=normal, weight-bearing on all 4 paws equally. The mobility scale assigned scores as follows; 1=walks with difficulty, 2=walks and runs with difficulty, 3=some limping, 4=normal.
Sodium channel blockers may produce motor effects at high doses (e.g. Hirotsu et al., 1988). Crobenetine has no effect on rotorod performance in mice at doses of 30 mg kg−1 s.c. (Carter et al., 2000). Further, for the purposes of the present study, an additional control study was performed. Normal (non-arthritic) rats were injected s.c. with vehicle, 5, 20 or 80 mg kg−1 of crobenetine and their locomotor behaviour monitored for 1 h prior to dosing and 15 h after. There was no significant effect of crobenetine at any dose on locomotor behaviour.
The rats were killed with an overdose of sodium pentobarbitone i.p. on the 5th day after injection of the ankle. In a post-mortem examination, the circumference of both ankles was measured, and the resistance to flexion and extension through the normal range of movement of both ankles tested. An experimenter blind to the treatment the rats received performed all behavioural and post-mortem observations.
Drugs
Crobenetine (BIII 890 CL) (2R-[2a,3(S*),6a]]-1,2,3,4,5,6-Hexahydro-6,11,11-trimethyl-3-[2-(phenyl-methoxy)propyl]-2,6-methano-3-benzazocin-10-ol hydrochloride), mexiletine (2-(2,6-dimethyl-phenoxy)-1-methyl-ethylamine hydrochloride) and meloxicam (4-hydroxy-2-methyl-N-(5-methyl-2-thiazolyl)-2H-1,2-benzothiazine-3-carboxamide-1,1-dioxide) were synthesized in the Department of Medicinal Chemistry, Boehringer Ingelheim Pharma KG. Xylitol was purchased as a 5% solution for infusion from Braun Melsungen, Germany. Complete Freund's adjuvant, mineral oil and methylcellulose were purchased from Sigma, Spain.
Data analysis
Statistical analysis was performed on the raw data obtained from behavioural experiments using Kruskal-Wallis one-way analysis of variance (ANOVA), followed by post-hoc Dunn's tests when a significant main effect was seen. The ankle inflammation data were analysed using parametric ANOVA followed by Tukey's tests. The data on joint fixation were analysed using Fisher's exact test. Values of P<0.05 were taken as statistically significant in all cases. Results are quoted as mean±s.e.mean. When significant inhibitory effects of the test compounds were seen, the dose required to produce a half maximal inhibition (ID50±s.e.mean) was calculated from a four parameter inhibition curve fitted to the raw data points using an iterative procedure and commercial software (GraphPad Prism, GraphPad Software Inc. U.S.A.).
All experiments were performed according to European Union and Spanish regulations on animal experimentation, and the experimental protocols were approved and supervised by the veterinary staff of the University Animal Care Facility.
Results
Behaviour in rats with adjuvant-induced mono-arthritis
Intraarticular injection of CFA in vehicle treated animals (n=12) resulted in a mono-arthritis with associated spontaneous pain, joint hyperalgesia and joint inflammation which were significantly different from responses in the control group of vehicle-related animals injected with mineral oil in the joint (n=12; Figures 1, 2 and 3, Table 1) as previously described (Laird et al., 1997). Changes were restricted to the injected joint, since there were neither significant changes in responses to stimulation of the contralateral joint (Figure 1) nor changes in its appearance or circumference (data not shown).
Figure 1.
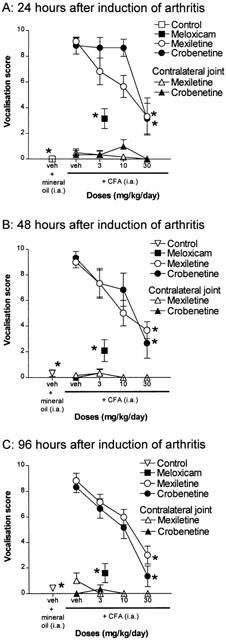
Effects of mexiletine or crobenetine on the vocalisation responses evoked by flexion and extension of the CFA-injected ankles and the contralateral non-injected ankle, n=6 for each group. Rats injected with mineral oil and those treated with 4 mg kg day−1 meloxicam are also shown for comparison, n=12 for each group. (A) The responses obtained 24 h after induction of arthritis. (B) The responses obtained 48 h after induction of arthritis. (C) The responses obtained 96 h after induction of arthritis. Results are expressed as mean±s.e.mean. *Denote those groups in which the response was significantly different from the vehicle-treated CFA group (P<0.05).
Figure 2.
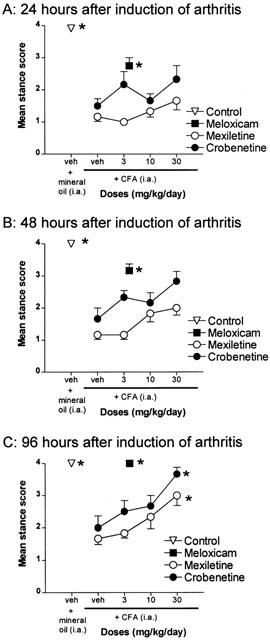
Effects of treatment with mexiletine or crobenetine on stance of the rats injected in the ankle joint with CFA (n=6 per group). Rats injected with mineral oil and those treated with 4 mg kg day−1 meloxicam are also shown for comparison, n=12 for each group. (A) The responses obtained 24 h after induction of arthritis. (B) The responses obtained 48 h after induction of arthritis. (C) The responses obtained 96 h after induction of arthritis. Results are expressed as mean±s.e.mean. *Denote those groups in which the response was significantly different from the vehicle-treated CFA group (P<0.05).
Figure 3.
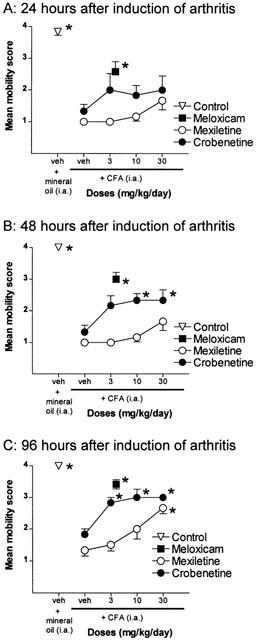
Effects of treatment with mexiletine or crobenetine on mobility of the rats injected in the ankle joint with CFA (n=6 per group). Rats injected with mineral oil and those treated with 4 mg kg day−1 meloxicam are also shown for comparison, n=12 for each group. (A) The responses obtained 24 h after induction of arthritis. (B) The responses obtained 48 h after induction of arthritis. (C) The responses obtained 96 h after induction of arthritis. Results are expressed as mean±s.e.mean. *Denote those groups in which the response was significantly different from the vehicle-treated CFA group (P<0.05).
Table 1.
Effect of test compounds on oedema of the ankle joint

A group of six rats treated with meloxicam (4 mg kg day−1 i.p.) was included as a positive control group in each experiment. In both experiments, the meloxicam-treated group showed a statistically-significant reversal of the CFA evoked effects in all measures (Figures 1, 2 and 3, Tables 1 and 2). There were no statistically significant differences in the results obtained in the different control groups in the two experiments, hence the data have been pooled for graphical representation.
Table 2.
Effect of test compounds on fixation of the ankle joint

Hyperalgesia of the inflamed joint was significantly reduced by treatment with crobenetine with a minimum effective dose (MED) of 30 mg kg day−1 at all time points tested compared to the vehicle-treated group (Figure 1). When tested at 24 h after induction of arthritis, only the group treated with the highest dose of 30 mg kg−1 s.c. showed clear signs of reduction in hyperalgesia (Figure 1A). At later time points (48 and 96 h after induction of arthritis), the groups of animals treated with 3 or 10 mg kg day−1 showed signs of reduced joint hyperalgesia, although the responses of these groups were not statistically significant (Figure 1B,C). Thus there was some indication of an increasing effectiveness of crobenetine treatment on joint hyperalgesia over the course of the 5 day study.
Mexiletine treatment was also effective at reducing the hyperalgesia of the inflamed joint at all time points tested (Figure 1). The effect of mexiletine was dose-dependent with a MED of 30 mg kg day−1 at all time points (Figure 1). In contrast to the effects observed with crobenetine, there was no apparent increase in the effectiveness of mexiletine treatment on joint hyperalgesia over the time course of the study (Figure 1).
Rats with CFA-induced mono-arthritis showed an impaired stance and mobility during unrestricted movement on all days examined, whereas rats injected with mineral oil showed almost no impairment as a result of the ankle injection (Figures 2 and 3). This difference was significant throughout the period of the study (Figures 2 and 3).
Crobenetine treatment had no significant effect on the impaired stance of the arthritic rats at 24 and 48 h after induction of arthritis (Figure 2A,B), although there was some indication of a dose-related improvement at the 48 h time point (Figure 2B). By 96 h after induction of arthritis, there was a clear dose-related and statistically significant effect of crobenetine treatment on the impaired stance of the arthritic rats, with a MED of 30 mg day−1 (Figure 2C). At this time point, the group treated with 30 mg kg−1 crobenetine showed an almost complete reversal of the impaired stance (Figure 2C).
Mexiletine treatment had no significant effect on the impaired stance of the arthritic rats at 24 or 48 h after induction of arthritis (Figure 2A,B), although there was an apparent dose-related improvement of the impairment at both time points. However, by the 96 h time point, mexiletine treatment produced a statistically significant improvement of the impaired stance with a MED of 30 mg kg day−1, although the maximal effect observed represented only a partial reversal of the CFA-induced effect (Figure 2C).
Treatment with crobenetine had no significant effect on impaired mobility 24 h after induction of arthritis (Figure 3A). However, at 48 and 96 h after arthritis induction, crobenetine had a significant and dose dependent antinociceptive effect (Figure 3B,C). The MEDs for this reversal of impaired mobility were 10 mg kg day−1 at 48 h and 3 mg kg day−1 at 96 h respectively (Figure 3B,C). However, the maximal effect of crobenetine on mobility represented only a partial reversal of impairment, in contrast to the almost complete recovery of impaired stance observed in the same animals (Figure 2C).
Mexiletine had no significant effects on impaired mobility in arthritic rats either at 24 or 48 h after induction of arthritis (Figure 3A,B), although there were some signs of a dose related improvement. At 96 h post-arthritis induction, mexiletine treatment had a dose-related and statistically significant effect on the impaired mobility, with a MED of 30 mg kg day−1 (Figure 3C).
Rats injected with CFA showed a small decrease in body weight (mean change=−4±1%) over the 5 days of the experiment. However, the change in body weight was not significantly different from that observed in rats injected in one ankle joint with mineral oil (mean change=1±1%). Neither mexiletine nor crobenetine treatment had a significant effect on body weight changes over the period of study.
Post-mortem examination of inflammatory signs
In vehicle-treated animals the ankle joint injected with CFA (n=12) had a mean circumference of 38±1 mm on post-mortem which was significantly greater than the mean circumference of 26±1 mm measured in rats with ankle joints injected with mineral oil (n=12; Table 1). Treatment with mexiletine or crobenetine had no significant effect on the circumference of the inflamed joint (Table 1). In contrast, meloxicam treatment significantly reduced the circumference of the injected joint (Table 1). The circumference of the contralateral, non-injected ankle joints was not significantly different between CFA and mineral oil treated groups (24±0.4 and 23±0.4 mm respectively). Drug treatment had no effect on the circumference in these contralateral joints (data not shown).
All vehicle treated animals injected with CFA (n=12) showed ‘fixation' of the injected joint i.e., on post-mortem there was resistance to flexion and extension through the normal range of movement (Table 2). Resistance to flexion and extension of the injected joint was not observed in any of the rats injected with mineral oil (Table 2). Neither mexiletine nor crobenetine treated rats showed any significant reduction in fixation of the inflamed joint compared to mono-arthritic vehicle-treated rats using Fisher's exact test (Table 2). Meloxicam treatment did produce a significant reduction (P<0.05) in the number of animals showing resistance to flexion and extension compared to mono-arthritic vehicle-treated rats (Table 2). None of the rats in the study showed any evidence of fixation of the contralateral, non-injected ankle joint (data not shown).
Discussion
In this study we found that the two sodium channel blockers tested were effective in reversing the joint hyperalgesia and signs of ongoing pain in rats with an inflamed ankle joint. Neither compound had a significant effect on the inflammatory signs in the joint (oedema and joint fixation). Crobenetine had a similar potency to mexiletine when considered in terms of doses in mg kg−1. However, crobenetine has a molecular weight approximately 3 fold that of mexiletine, and thus is more potent expressed in terms of mol kg−1. This difference would be expected because crobenetine has a considerably higher affinity for sodium channels than mexiletine. The affinity of crobenetine for the inactive state of the channel is 77 nM (Carter et al., 2000), whereas the affinity of mexiletine is 21 μM (Weiser et al., 1999), a difference of more than 200 fold. Thus, the difference in vivo between the two compounds was not as great as would be predicted from the affinity data. This can be partly explained by the differences in pharmacokinetics of the compounds after s.c. administration. Injection of 30 mg kg−1 crobenetine s.c. in the rat produces a plasma level of 469±149 ng m−1 (mean±s.d.) at 15 min. Under equivalent conditions, plasma levels of mexiletine are 3 fold higher (V. Häselbarth and A. Carter, unpublished observations). The highest dose of crobenetine tested in the present study was as efficacious in reversing joint hyperalgesia as meloxicam was found to be in a previous study using the same animal model of mono-arthritis (Laird et al., 1997). Meloxicam has been shown to be effective in reducing pain associated with rheumatoid arthritis in clinical trials in man (e.g. Lemmel et al., 1997).
Crobenetine showed an increasing antihyperalgesic and analgesic effect over the course of the 5 day study. A similar although less pronounced effect was observed with mexiletine in the observations of stance and mobility during unrestricted movement. This apparent increase in the analgesic effects of crobenetine is unlikely to be due to drug accumulation. Crobenetine has a half life in plasma of less than 3 h (see Methods), and so complete elimination in a 24 h period would be expected. Furthermore studies have shown no evidence of accumulation with chronic dosing in rats over 4 weeks at up to 40 mg kg day−1 (V. Häselbarth and A. Carter, unpublished observations). One possible explanation is a change in the nature or number of sodium channels expressed over the duration of the CFA-induced inflammation, since studies of expression in rats injected with CFA show a relatively slow time course of up-regulation with changes occurring over several days (Gould et al., 1998; 1999).
This is the first report of the effects of sodium channel blockers on pain and hyperalgesia evoked by joint inflammation. However, several previous studies in rats have shown analgesic effects of sodium channel blockers, including mexiletine, in behavioural tests of allodynia and hyperalgesia evoked by peripheral nerve damage (Chaplan et al., 1995; Hedley et al., 1995; Jett et al., 1997) and also inhibitory effects on responses of dorsal horn neurons in rats with peripheral neuropathy (Chapman et al., 1997; Omana-Zapata et al., 1997; Sotgiu et al., 1994).
Sodium channel blockers have also been shown to be effective in blocking the acute inflammatory pain responses to formalin injection (Hitosugi et al., 1999; Jett et al., 1997) and reducing electrically-evoked responses of dorsal horn neurones after carrageenan injection into the hindpaw (Chapman & Dickenson, 1997). However, mexiletine has no effect on nociceptive reflex responses to mechanical and thermal stimuli in the dose range used in the present study and which is effective in models of neuropathic pain (Jett et al., 1997; Hedley et al., 1995; Xu et al., 1992).
Motor deficits may be observed in experimental animals treated with high doses of sodium channel blockers (e.g. Hirotsu et al., 1988). However, crobenetine has no effect on rotorod performance (Carter et al., 2000) or on spontaneous locomotor activity (see Methods) at doses greater than those tested for analgesic effects in the present study and thus shows a separation of analgesic and anti-hyperalgesic effects from motor effects. Mexiletine has been found to have mild motor side effects at doses similar to those found effective here in some studies (Xu et al., 1992), but not in others (Jett et al., 1997). In any case, such motor effects may influence the measures of spontaneous pain (mobility and stance), but are unlikely to affect vocalization, the end-point used in the test of joint hyperalgesia. Cardiovascular effects may also affect the utility of sodium channel blockers as analgesics. Mexiletine has effects on heart rate but not cardiac output in rat in the dose range used in the present study (Kume et al., 1992), thus its cardiovascular effects are unlikely to be responsible for the analgesic effects observed in the current study. Crobentine is highly use-dependent and therefore in principle unlikely to affect cardiovascular sodium channels in physiological conditions. This assumption is borne out by control experiments which indicate that at doses of up to 80 mg kg−1 s.c. in awake rats there is no effect on cardiovascular parameters (E. Lehr and A.K. Carter, unpublished observations).
Mechanism of action
Both test compounds penetrate the blood – brain barrier after systemic administration, since they are effective in animal models of cerebral ischaemia after systemic administration (Stys & Lesiuk, 1996; Carter et al., 2000). Thus their site of analgesic action may be on the peripheral or central components of the nociceptive pathway. Use dependent sodium channel blockers, including crobenetine, have been shown to reduce the number of TTX-insensitive action potentials generated by sustained depolarisations of dorsal root ganglion at low concentrations which do not block single action potentials in neurones (Scholz & Vogel, 2000; Krause et al., 2000). This is an attractive possibility for the analgesic action of the test compounds in the present study, as ∼75 – 80% of Aδ- and C-fibre joint afferents start to fire spontaneously after joint inflammation (Schaible & Schmidt, 1996). This population of fine, mainly nociceptive afferents, expresses TTX-sensitive channels. An action on sodium channels in primary afferents is also suggested by the observations of selective effects in animals with inflammation, since under these conditions, primary afferent sodium channels are upregulated (Gould et al., 1998; 1999), compared to control animals.
However, central actions may also contribute to the analgesic effects seen in the present study. For example, systemic mexiletine reduces the signs of allodynia and mechanical hyperalgesia evoked by ischaemic spinal cord injury in rats (Xu et al., 1992) and also inhibits pain responses in mice induced by intrathecal injection of algogens (Hitosugi et al., 1999).
Our results show that the sodium channel blockers, crobenetine and mexiletine, are effective at reversing joint hyperalgesia in rats with adjuvant-induced mono-arthritis and also in decreasing spontaneous pain-related behavioural changes evoked by mono-arthritis. The analgesic actions were more potent than the anti-inflammatory actions, an observation that suggests a direct effect on the nociceptive system. These data indicate that upregulation of sodium channel expression in primary afferent neurones may play an important role in the pain and hyperalgesia induced by joint inflammation.
Acknowledgments
This work was supported by a Contrato Programa from the Madrid Regional Government, Spain (to Prof F. Cervero) and by Boehringer Ingelheim, Germany. We are grateful to Prof Erich Lehr and Dr Volkmar Häselbarth for providing additional data on crobenetine.
Abbreviations
- ANOVA
analysis of variance
- CFA
complete Freund's adjuvant
- MED
minimum effective dose
- TTX
tetrodotoxin
References
- AKOPIAN A.N., SOUSLOVA V., ENGLAND S., OKUSE K., OGATA N., URE J., SMITH A., KERR B.J., MCMAHON S.B., BOYCE S., HILL R.G., STANFA L., DICKENSON A.H., WOOD J.N. The tetrodotoxin-resistant sodium channel SNS has a specialized function in pain pathways. Nat. Neurosci. 1999;2:541–548. doi: 10.1038/9195. [DOI] [PubMed] [Google Scholar]
- BAKER M.D., WOOD J.N. Involvement of Na+ channels in pain pathways. TIPS. 2001;22:27–31. doi: 10.1016/s0165-6147(00)01585-6. [DOI] [PubMed] [Google Scholar]
- BUTLER S.H., GODEFROY F., BESSON J.-M., WEIL-FUGAZZA J. A limited arthritic model for chronic pain studies in the rat. Pain. 1992;48:73–81. doi: 10.1016/0304-3959(92)90133-V. [DOI] [PubMed] [Google Scholar]
- CARTER A.J., GRAUERT M., PSCHORN U., BECHTEL W.D., BARTMANN-LINDHOLM C., QU Y., SCHEUER T., CATTERALL W.A., WEISER T. Potent blockade of sodium channels and protection of brain tissue from ischemia by BIII 890 CL. Proc. Natl. Acad. Sci. U.S.A. 2000;97:4944–4949. doi: 10.1073/pnas.040577097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- CHAPLAN S.R., BACH F.W., SHAFER S.L., YAKSH T.L. Prolonged alleviation of tactile allodynia by intravenous lidocaine in neuropathic rats. Anesthesiology. 1995;83:775–785. doi: 10.1097/00000542-199510000-00017. [DOI] [PubMed] [Google Scholar]
- CHAPMAN V., DICKENSON A.H. Inflammation reveals inhibition of noxious responses of rat spinal neurones by carbamazepine. Neuroreport. 1997;8:1399–1404. doi: 10.1097/00001756-199704140-00016. [DOI] [PubMed] [Google Scholar]
- CHAPMAN V., WILDMAN M.A., DICKENSON A.H. Distinct electrophysiological effects of two spinally administered membrane stabilising drugs, bupivacaine and lamotrigine. Pain. 1997;71:285–295. doi: 10.1016/s0304-3959(97)03365-4. [DOI] [PubMed] [Google Scholar]
- DANZIGER N., WEIL-FUGAZZA J., LE BARS D., BOUHASSIRA D. Alterations of descending modulation of nociception during the course of monoarthritis in the rat. J. Neurosci. 1999;19:2394–2400. doi: 10.1523/JNEUROSCI.19-06-02394.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- ENGLAND J.D., BEVAN S., DOCHERTY R. PGE2 modulates the tetrodotoxin-sensitive sodium current in neonatal rat dorsal root ganglion neurones via the cyclic AMP-protein kinase A cascade. J. Physiol. (Lond.) 1996;495:429–440. doi: 10.1113/jphysiol.1996.sp021604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- GOULD H.J., ENGLAND J.D., LIU Z.P., LEVINSON S.R. Rapid sodium channel augmentation in response to inflammation induced by complete Freund's adjuvant. Brain Res. 1998;802:69–74. doi: 10.1016/s0006-8993(98)00568-x. [DOI] [PubMed] [Google Scholar]
- GOULD H.J., GOULD T.N., PAUL D., ENGLAND J.D., LIU Z.P., REEB S.C., LEVINSON S.R. Development of inflammatory hypersensitivity and augmentation of sodium channels in rat dorsal root ganglia. Brain Res. 1999;824:296–299. doi: 10.1016/s0006-8993(99)01218-4. [DOI] [PubMed] [Google Scholar]
- HEDLEY L., MARTIN B., WATERBURY L.D., CLARKE D.E., HUNTER J.C. A comparison of the action of mexiletine and morphine in rodent models of acute and chronic pain. Proc. West. Pharmacol. Soc. 1995;38:103–104. [PubMed] [Google Scholar]
- HIROTSU I., KIHARA T., NAKAMURA S., HATTORI Y., HATTA M., KITAKAZE Y., TAKAHAMA K., HASHIMOTO Y., MIYATA T., ISHIHARA T. General pharmacological studies on N-2,6-dimethylphenyl)-8-pyrrolizidineacetamide hydrochloride hemihydrate. 1st communication: on the central nervous system. Arzneimittelforschung. 1988;38:1398–1409. [PubMed] [Google Scholar]
- HITOSUGI H., KASHIWAZAKI T., OHSAWA M., KAMEI J. Effects of mexiletine on algogenic mediator-induced nociceptive responses in mice. Methods Find. Exp. Clin. Pharmacol. 1999;21:409–413. doi: 10.1358/mf.1999.21.6.541921. [DOI] [PubMed] [Google Scholar]
- JETT M.F., MCGURK J., WALIGORA D., HUNTER J.C. The effects of mexiletine, desipramine and fluoxetine in rat models involving central sensitisation. Pain. 1997;69:161–169. doi: 10.1016/s0304-3959(96)03231-9. [DOI] [PubMed] [Google Scholar]
- KRAUSE U., WEISER T., CARTER A.J., GRAUERT M., VOGEL W., SCHOLZ A. BIII 890 CL is a potent, use dependent blocker of TTX-resistant sodium channels in dorsal root ganglion neurons. Soc. Neurosci. Abstr. 2000;26:1218. [Google Scholar]
- KUME M., KASHIMOTO S., KUMAZAWA T. Functional and metabolic effects of mexiletine in the rat heart-lung preparation. Eur. J. Anaesthesiol. 1992;9:223–228. [PubMed] [Google Scholar]
- LAIRD J.M.A., HERRERO J.F., GARCIA DE LA RUBIA P., CERVERO F. Analgesic activity of the novel COX-2 selective NSAID, meloxicam, in experimental monoarthritis in the rat: Central and peripheral components. Inflammation Res. 1997;46:203–210. doi: 10.1007/s000110050174. [DOI] [PubMed] [Google Scholar]
- LEMMEL E.M., BOLTEN W., BURGOSVARGAS R., PLATT P., NISSILA M., SAHLBERG D., BJORNEBOE O., BAUMGARTNER H., VALAT J., FRANCHIMONT P., BLUHMKI E., HANFT G., DISTEL M. Efficacy and safety of meloxicam in patients with rheumatoid arthritis. J. Rheumatol. 1997;24:282–290. [PubMed] [Google Scholar]
- OMANA-ZAPATA I., KHABBAZ M.A., HUNTER J.C., BLEY K.R. QX-314 inhibits ectopic nerve activity associated with neuropathic pain. Brain Res. 1997;771:228–237. doi: 10.1016/s0006-8993(97)00770-1. [DOI] [PubMed] [Google Scholar]
- PORRECA F., LAI J., BIAN D., WEGERT S., OSSIPOV M.H., EGLEN R.M., KASSOTAKIS L., NOVAKOVIC S., RABERT D.K., SANGAMESWARAN L., HUNTER J.C. A comparison of the potential role of the tetrodotoxin-insensitive sodium channels PN3/SNS and NaN/SNS2, in rat models of chronic pain. Proc. Natl. Acad. Sci. U.S.A. 1999;96:7640–7644. doi: 10.1073/pnas.96.14.7640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- SCHAIBLE H.-G., SCHMIDT R.F.Neurobiology of articular nociceptors Neurobiology of nociceptors 1996Oxford, Oxford University Press; 202–219.Edited by Bemonte, C., and Cervero, F. pp [Google Scholar]
- SCHOLZ A., VOGEL W. Tetrodotoxin-resistant action potentials in dorsal root ganglion neurons are blocked by local anesthetics. Pain. 2000;89:47–52. doi: 10.1016/S0304-3959(00)00345-6. [DOI] [PubMed] [Google Scholar]
- SINDRUP S.H., JENSEN T.S. Efficacy of pharmacological treatments of neuropathic pain: an update and review of effect related to mechanism of action. Pain. 1999;83:389–400. doi: 10.1016/S0304-3959(99)00154-2. [DOI] [PubMed] [Google Scholar]
- SOTGIU M.L., BIELLA G., CASTAGNA A., LACERENZA M., MARCHETTINI P. Different time courses of i.v. lidocaine effect on ganglionic and spinal units in neuropathic rats. Neuroreport. 1994;5:873–876. doi: 10.1097/00001756-199404000-00005. [DOI] [PubMed] [Google Scholar]
- STYS P.K., LESIUK H. Correlation between electrophysiological effects of mexiletine and ischemic protection in central nervous system white matter. Neuroscience. 1996;71:27–36. doi: 10.1016/0306-4522(95)00430-0. [DOI] [PubMed] [Google Scholar]
- TANAKA M., CUMMINS T.R., ISHIKAWA K., DIBHAJJ S.D., BLACK J., WAXMAN S.G. SNS Na+ channel expression increases in dorsal root ganglion neurons in the carrageenan inflammatory pain model. Neuroreport. 1998;9:967–972. doi: 10.1097/00001756-199804200-00003. [DOI] [PubMed] [Google Scholar]
- TANELIAN D.L., BROSE W.G. Neuropathic pain can be relieved by drugs that are use-dependent sodium channel blockers: Lidocain, carbamazepine, and mexiletine. Anaesthesiology. 1991;74:949–951. doi: 10.1097/00000542-199105000-00026. [DOI] [PubMed] [Google Scholar]
- TATE S., BENN S., HICK C., TREZISE D., JOHN V., MANNION R.J., COSTIGAN M., PLUMPTON C., GROSE D., GLADWELL Z., KENDALL G., DALE K., BOUNTRA C., WOOLF C.J. Two sodium channels contribute to the TTX-R sodium current in primary sensory neurons. Nat. Neurosci. 1998;1:653–655. doi: 10.1038/3652. [DOI] [PubMed] [Google Scholar]
- WAXMAN S.G., CUMMINS T.R., DIBHAJJ S.D., FJELL J., BLACK J.A. Sodium channels, excitability of primary sensory neurons and the molecular basis of pain. Muscle Nerve. 1999;22:1177–1187. doi: 10.1002/(sici)1097-4598(199909)22:9<1177::aid-mus3>3.0.co;2-p. [DOI] [PubMed] [Google Scholar]
- WEISER T., QU T., CATTERALL W.A., SCHEUER T. Differential site of action of R mexiletine with the local anesthetic receptor site on brain and heart sodium channel alpha-subunits. Mol. Pharmacol. 1999;56:1238–1244. doi: 10.1124/mol.56.6.1238. [DOI] [PubMed] [Google Scholar]
- XU X.-J., HAO J.-X., SEIGER A., ARNER S., LINDBLOM U., WIESENFELD-HALLIN Z. Systemic mexiletine relieves chronic allodynia-like symptoms in rats with ischemic spinal cord injury. Anesthe. Analg. 1992;74:649–652. doi: 10.1213/00000539-199205000-00005. [DOI] [PubMed] [Google Scholar]