

Abstract
The mechanisms of arsenic (As) hyperaccumulation in Pteris vittata, the first identified As hyperaccumulator, are unknown. We investigated the interactions of arsenate and phosphate on the uptake and distribution of As and phosphorus (P), and As speciation in P. vittata. In an 18-d hydroponic experiment with varying concentrations of arsenate and phosphate, P. vittata accumulated As in the fronds up to 27,000 mg As kg−1 dry weight, and the frond As to root As concentration ratio varied between 1.3 and 6.7. Increasing phosphate supply decreased As uptake markedly, with the effect being greater on root As concentration than on shoot As concentration. Increasing arsenate supply decreased the P concentration in the roots, but not in the fronds. Presence of phosphate in the uptake solution decreased arsenate influx markedly, whereas P starvation for 8 d increased the maximum net influx by 2.5-fold. The rate of arsenite uptake was 10% of that for arsenate in the absence of phosphate. Neither P starvation nor the presence of phosphate affected arsenite uptake. Within 8 h, 50% to 78% of the As taken up was distributed to the fronds, with a higher translocation efficiency for arsenite than for arsenate. In fronds, 49% to 94% of the As was extracted with a phosphate buffer (pH 5.6). Speciation analysis using high-performance liquid chromatography-inductively coupled plasma mass spectroscopy showed that >85% of the extracted As was in the form of arsenite, and the remaining mostly as arsenate. We conclude that arsenate is taken up by P. vittata via the phosphate transporters, reduced to arsenite, and sequestered in the fronds primarily as As(III).
Arsenic (As) is ubiquitous in the environment and is derived from both natural and anthropogenic sources. As is a nonessential element for plants, and inorganic As species are generally highly phytotoxic. Arsenate is the predominant As species in aerobic soils, whereas arsenite dominates under anaerobic conditions (Smith et al., 1998). Arsenate acts as a phosphate analog and can disrupt phosphate metabolism, whereas arsenite reacts with sulfhydryl groups of enzymes and tissue proteins, leading to inhibition of cellular function and death (Meharg and Hartley-Whitaker, 2002). In all plant species tested so far, it has been shown that arsenate is taken up via the phosphate transport systems (Asher and Reay, 1979; Lee, 1982; Ullrich-Eberius et al., 1989; Meharg and Macnair, 1992). Arsenate resistance has been identified in a range of plant species, which is generally achieved through a decreased uptake of arsenate because of suppression of the high-affinity phosphate uptake system (Meharg and Macnair, 1991, 1992; Meharg and Hartley-Whitaker, 2002). Inside plant cells, arsenate may be detoxified through reduction to arsenite, which is subsequently complexed with thiols, particularly phytochelatins (PCs; Pickering et al., 2000; Schmöger et al., 2000; Hartley-Whitaker et al., 2001). It is possible that As-PC complexes are sequestered in vacuoles, although direct evidence for this is still lacking (Meharg and Hartley-Whitaker, 2002).
The transfer of As from soil to plant is low for most plant species. This may be because of several reasons: (a) low bioavailability of As in soil, (b) restricted uptake by plant roots, (c) limited translocation of As from roots to shoots, and (d) As phytotoxicity at relatively low concentrations in plant tissues. Recently, the brake fern Pteris vittata has been found to be able to hyperaccumulate As from soil (Ma et al., 2001). It can accumulate up to 22,630 mg As kg−1 in the shoot (frond) dry weight. Furthermore, the bioconcentration factor, defined as the ratio of shoot As concentration to soil As concentration, was greater than 10. The fern possesses three key features that are typical of metal/metalloid hyperaccumulator plants: an efficient root uptake, an efficient root to shoot translocation, and a much-enhanced tolerance to As inside plant cells. After the discovery of this first As hyperaccumulator, several other fern species, including Pityrogramma calomelanos (Francesconi et al., 2002), Pteris cretica, Pteris longifolia, and Pteris umbrosa (Zhao et al., 2002), have recently been added to the list of As hyperaccumulators. The hyperaccumulation trait of these ferns may be potentially exploitable in phytoremediation of As contaminated soils.
Because As hyperaccumulation in P. vittata is a newly discovered phenomenon, the mechanisms involved have not been elucidated. We do not know if arsenate is taken up via the phosphate transporters in P. vittata, as has been shown for nonaccumulator species. If so, how do phosphorus (P) and As interact during uptake and root to shoot translocation? Specifically, how does P. vittata acquire and maintain sufficient P nutrition under high arsenate supply? Also, does P. vittata take up arsenate and arsenite similarly efficiently, as implied by Ma et al. (2001)? What are the As species in fronds? This is particularly relevant to the understanding of As detoxification inside plant cells. These questions are the focus of the present paper. To address the P × As interactions in plants, one has to use hydroponic culture rather than soil culture, because the latter also involves interactions of P and As in soil chemical processes.
RESULTS
The P. vittata plants from the University of Florida (Gainesville) were collected from As-contaminated soils (Ma et al., 2001), whereas the plants in the Oxford Botanical Garden (Oxford) grew on an uncontaminated soil. Our preliminary studies showed that both accessions hyperaccumulated As similarly, suggesting that As hyperaccumulation in P. vittata is a constitutive trait (Zhao et al., 2002).
Arsenate and Phosphate Interactions Experiment
Phytotoxicity was observed in the 416 and 2,080 μm arsenate treatments when P supply was low (20 μm). The symptoms of phytotoxicity were necrosis of pinnae around the edges and tips of fronds. Increasing P supply alleviated As phytotoxicity. However, shoot and root dry weights were not affected significantly by either As or P treatments (data not shown).
As hyperaccumulation was demonstrated clearly in this experiment, with As concentrations in the roots and shoots reaching up to 8,960 and 27,000 mg kg−1 dry weight, respectively (Fig. 1, a and b). In all +As treatments, the concentration of As in the shoots was always greater than that in the roots, with the shoot to root ratio of As concentration varying from 1.3 to 6.7. Increasing phosphate concentration in nutrient solution decreased both root and shoot concentrations of As significantly (P < 0.01), but the effect was proportionally larger for roots than for shoots. For example, in the 83 μm arsenate treatment, increasing phosphate from 20 to 100 μm decreased root and shoot As concentrations by 76% and 46%, respectively. In the 416 μm arsenate treatment, increasing phosphate from 20 to 500 μm decreased root and shoot As concentrations by 73% and 41%, respectively. The inhibitory effect of phosphate on root As concentration was larger than what may be expected from a simple “dilution” of arsenate by phosphate in the solution, implying that phosphate has a higher affinity to the uptake system in the roots than arsenate. In contrast, the inhibitory effect of phosphate on shoot As concentration was smaller than what may be expected from a simple “dilution” effect.
Figure 1.

Effects of varying concentrations of arsenate and phosphate in nutrient solution on the concentrations of As in roots (a) and shoots (b), and the concentrations of P in roots (c) and shoots (d) of P. vittata. P. vittata plants were precultured with full nutrient solution for 3 weeks, and then exposed to different concentrations of arsenate and phosphate for 18 d. Values are means ± ses (n = 4).
The concentrations of P in the roots and shoots of P. vittata are shown in Figure 1, c and d, respectively. Increasing arsenate concentration in the solution decreased the concentration of P in the roots significantly (P < 0.05), particularly when the P supply was low (20 μm). In contrast, the arsenate treatments had no significant effect on the P concentration in the shoots. The pattern of P distribution between roots and shoots was markedly different from that of As. In most cases, the concentration of P in the roots was greater than that in the shoots.
Kinetics of Arsenate Uptake
Depletion of arsenate in the uptake solution was monitored over 8 h, which reflects the net uptake of As by roots. Figure 2a shows the depletion curves of arsenate in one replicate of each of the three treatments. Because the rate of depletion depends on root weight, the depletion data of different replicates in the same treatment cannot be averaged. In Figure 2a, root weights were similar between the three treatments, allowing direct comparisons of the depletion rate between the treatments. It is clear that in the P-sufficient plants (control), the presence of 50 μm phosphate slowed the depletion of arsenate greatly. In the control without phosphate, the arsenate concentration in the uptake solution decreased to a minimal concentration after about 4 h, whereas in the +50 μm phosphate treatment, only about 20% of the arsenate was taken up during 8 h. Compared with the P-sufficient plants, the treatment where the P supply was withheld for 8 d before the depletion experiment showed markedly increased depletion rate (Fig. 2a).
Figure 2.

a, Depletion curves of arsenate in the uptake solution as affected by the presence of 50 μm phosphate, or by P starvation for 8 d. One replicate per treatment, with similar root fresh weights. For the Control −P and P-deficient −P treatments, a modified Michaelis-Menten equation was fitted to the data (see “Materials and Methods”). For the Control +50 μm phosphate treatment, a linear regression was fitted. b, Accumulation of As by P. vittata in the depletion experiment with arsenate, expressed on a basis of root fresh weight. Note that ses increased markedly when arsenate in the uptake solution was exhausted because of variation in the root fresh weight between replicates. A Gompertz model was fitted to the data of the Control −P and P-deficient −P treatments, and a linear model to the data of the Control +50 μm phosphate treatment.
Figure 2b shows the cumulative uptake of arsenate, expressed on a basis of root fresh weight, in the three treatments. The rate of arsenate accumulation by roots decreased markedly when phosphate was present in the uptake solution, and increased when the plants were starved of P for 8 d. The rate of As accumulation was approximately linear in the first 90 and 240 min for the P-deficient and -sufficient (control) treatments, respectively, without the presence of phosphate, but was linear for the entire uptake period (8 h) for the control +50 μm phosphate treatment. During the linear accumulation phase, the net uptake rate of arsenate was 90, 20, and 215 nmol g−1 root fresh weight h−1 for the control −P, control +50 μm phosphate, and P-deficient −P treatments, respectively.
Claassen and Barber (1974) and Drew et al. (1984) demonstrated that the depletion technique can be used to estimate kinetic parameters of ion influx, providing that a near complete depletion was achieved. Table I shows the kinetic parameters for arsenate uptake by P. vittata estimated from the depletion data of the control −P and P-deficient −P treatments. It was not possible to estimate the kinetic parameters for the control +50 μm phosphate treatment because the depletion of arsenate was far from complete. Compared with the P-sufficient plants (control), the P deficiency treatment increased Imax for arsenate by 2.5-fold (P < 0.001), but had no significant effect on Km (Table I). In addition, Cmin was significantly higher (P < 0.01) in the P-deficient plants.
Table I.
Kinetic parameters of arsenate influx in P. vittata
| Treatment | Maximum Net Influx (Imax) | Km | Minimal Ion Concentration below Which Net Influx Ceases (Cmin) |
|---|---|---|---|
| nmol g−1 root fresh wt h−1 | μm | ||
| Control −P | 129.6 ± 19.7 | 0.52 ± 0.21 | 0.096 ± 0.01 |
| P-deficient −P | 318.6 ± 25.8 | 0.97 ± 0.37 | 0.35 ± 0.06 |
Plants were precultured with a full nutrient solution for 3 weeks for the control treatments. In the P-deficient treatment, plants were starved of P for 8 d before the depletion experiment. The initial concentration of arsenate in the uptake solution was 5 μm. Values are means ± se (n = 5).
Kinetics of Arsenite Uptake
In the depletion experiment with arsenite, between 20% and 40% of the initial arsenite was depleted over the 8-h period (Fig. 3a). The rate of depletion was much slower than that for arsenate. The differences between the control −P, control +50 μm phosphate, and P-deficient treatments were small. When the depletion data were expressed as accumulation on a root fresh weight basis (Fig. 3b), the rate of arsenite accumulation by P. vittata roots was 8 to 10 nmol g−1 root fresh weight h−1. There were no significant differences between the three treatments. Because arsenite in the uptake solution was far from being depleted, kinetic parameters could not be obtained.
Figure 3.

a, Depletion curves of arsenite in the uptake solution as affected by the presence of 50 μm phosphate, or by P starvation for 8 d. One replicate per treatment, with similar root fresh weights, is shown. A linear regression was fitted to the data in different treatments. b, Accumulation of As by P. vittata in the depletion experiment with arsenite, expressed on the basis of root fresh weight. A linear regression was fitted to the data in different treatments.
Distribution of As between Roots and Shoots
At the end of the depletion experiments (8 h), on average 60% and 71% of the As taken up was distributed to the shoots (fronds) fed with arsenate and arsenite, respectively. A significantly (P < 0.05) higher proportion of As taken up was in the shoots when arsenite was supplied than when arsenate was supplied (Table II). Presence of 50 μm phosphate in the uptake solution had no significant effect on the distribution of As between roots and shoots. However, the pretreatment with P starvation for 8 d enhanced the distribution of As to the shoots significantly (P < 0.001), in both the arsenate and arsenite depletion experiments (Table II).
Table II.
Percentage of the As taken up during the 8-h depletion experiment that was distributed to the shoots
| As Species in the Uptake Solution | Treatment
|
||
|---|---|---|---|
| Control −P | Control + 50 μm phosphate | P-deficient −P | |
| Arsenate | 50.9 ± 4.2 | 53.6 ± 3.5 | 75.9 ± 3.7 |
| Arsenite | 67.0 ± 2.1 | 68.3 ± 4.6 | 78.2 ± 5.3 |
P. vittata plants were precultured with a full nutrient solution for 3 weeks for the control treatments. In the P-deficient treatment, plants were starved of P for 8 d before the depletion experiment. The initial concentration of arsenate or arsenite in the uptake solution was 5 μm. Values are means ± ses (n = 5).
As Speciation in P. vittata Fronds
Total uptake of As over 2 weeks differed for arsenite and arsenate, with As accumulation with arsenite being higher, though not significantly so, than for arsenate (Table III). However, the percentage of As extracted into a phosphate buffer at pH 5.6 was significantly higher for arsenate-treated fern (94%) than for arsenite (49%) (P < 0.05).
Table III.
Speciation of arsenic extracted by phosphate buffer (pH 5.6) from fronds of plants treated with either arsenate or arsenite
| Treatment | Total As | As Extractable with Phosphate Buffer | Speciation with Size Exclusion Chromatography (SEC)
|
Speciation with Anion-Exchange Chromatography (AEC)
|
Sum of As Species as % of Extracted As
|
|||
|---|---|---|---|---|---|---|---|---|
| As(III) | As(V) | As(III) | As(V) | SEC | AEC | |||
| mg kg−1 fresh wt | % | % | ||||||
| Arsenite | 1,068 ± 259 | 49 ± 9 | 95.7 ± 0.9 | 4.3 ± 0.9 | 94.0 ± 2 | 6 ± 1.52 | 65.3 ± 4.4 | 94.3 ± 11.2 |
| Arsenate | 440 ± 69 | 94 ± 2 | 88.5 ± 3.5 | 11.5 ± 3.5 | 85 ± 4.5 | 14.5 ± 10.2 | 49.5 ± 1.5 | 70 ± 2 |
P. vittata plants were precultured with full nutrient solution for 3 weeks, and then exposed to 50 μm arsenite or arsenate for 2 weeks. Values are means ± ses (n = 3).
Chromatography using inductively coupled plasma mass spectroscopy (ICP-MS)-specific detection for As of the phosphate extracts of fronds, using both anion-exchange and size exclusion HPLC columns, showed that only arsenite and arsenate were detected under the chromatography conditions, with arsenite predominating for both treatments. Chromatograms for size exclusion and anion exchange are presented in Figures 4 and 5, respectively, showing the separation of both the pure standards and of the fern extracts, and that good separations were achieved for the As species. There were no significant differences in the proportion of arsenite or arsenate detected between the arsenite- and arsenate-treated ferns.
Figure 4.

Chromatograms (HPLC-ICP-MS) using SEC (30 mm phosphate buffer pH 5.6) for As speciation (mass-to-charge ratio [m/z] 75). a, Standard mixture of pentavalent and trivalent inorganic As [As(III) and As(V)] and their methylated counterparts [monomethyarsonic acid (MMA) (III), MMA(V), dimethylarsinic acid (DMA) (III), and DMA(V)]. b, P. vittata cultured in hydroponic solution containing either 50 μm arsenate or 50 μm arsenite.
Figure 5.

Chromatograms (HPLC-ICP-MS) using anion-exchange chromatography (20 mm carbonate buffer, pH 8.08) for As speciation (m/z 75). a, Standard mixture of pentavalent and trivalent inorganic As [As(III) and As(V)] and their methylated counterparts [MMA(III), MMA(V), DMA(III), and DMA(V)]. b, P. vittata cultured in hydroponic solution containing either 50 μm arsenate or 50 μm arsenite.
When the proportion of the extracted As detected by size exclusion and anion exchange was investigated, it was found that more of the extracted As was detected by anion exchange, and also that more As in the extract was accounted for in the arsenite-treated plants compared with the arsenate-treated plants (Table III). Two-way analysis of variance showed that both the type of chromatography used (P < 0.01) and the As species with which the plants were treated (P < 0.05) were significant.
DISCUSSION
The ability of P. vittata to hyperaccumulate As in shoots was clearly demonstrated in the long-term (18 d) hydroponic experiment. The concentration of As in the shoots was up to 27,000 mg kg−1 dry weight, although phytotoxic symptoms started to appear once the concentration exceeded approximately 10,000 mg kg−1 dry weight. This threshold value is similar to that reported by Tu and Ma (2002), who grew the fern plants on a soil spiked with different sources of As. The tolerance of P. vittata to As is far greater than that observed for many non-hyperaccumulating plant species, which have a threshold concentration for phytotoxicity of between 5 and 100 mg kg−1 dry weight (Kabata-Pendias and Pendias, 1992). Internal detoxification of As must be an important feature of this hyperaccumulator species. In this respect, P. vittata is in sharp contrast to As resistant grass species, including Holcus lanatus, Deschampsia cespitosa, and Agrostis capillaris, which have a much lower uptake rate of arsenate than nonresistant populations of the same species (Meharg and Hartley-Whitaker, 2002).
In all plant species studied thus far, it has been shown that arsenate is taken up via the phosphate transport systems (Meharg and Hartley-Whitaker, 2002). Here, we have shown that the As hyperaccumulator P. vittata is no exception. The influx of arsenate was strongly depressed by the presence of phosphate in the uptake solution (Fig. 2). In the long-term hydroponic experiment, the concentrations of As in the plants were decreased by increasing the phosphate concentration in the nutrient solution (Fig. 1). The inhibitory effect was more apparent when the concentrations of phosphate and arsenate in nutrient solution were comparable, and affected the root As concentration more than the shoot As concentration.
It is well known that P deficiency increases the capacity of plant roots to take up phosphate (Lee, 1982; Drew et al., 1984). Expression of the genes encoding phosphate transporters is up-regulated in the roots of P-deficient plants (Muchhal et al., 1996; Smith et al., 1997; Liu et al., 1998), which also leads to a concurrent increase in the transporter protein (Muchhal and Raghothama, 1999). This means that plants increase their capacity for phosphate uptake in response to P deficiency by synthesis of additional transporter molecules. If phosphate transporters are responsible for arsenate uptake, then arsenate uptake should be enhanced in P-deficient plants. Such an effect has been reported in barley (Hordeum vulgare; Lee, 1982) and in the As-nonresistant population of H. lanatus (Meharg and Macnair, 1992), and was demonstrated clearly in this study for the As hyperaccumulator P. vittata (Fig. 2). P starvation for 8 d was found to increase Imax for arsenate by 2.5-fold, suggesting an increased density of phosphate/arsenate transporters on the plasma membranes in root cells. In contrast, the Km for arsenate was not affected significantly by P starvation.
Because the initial As concentration was low in the depletion experiments, As uptake was most likely attributed to high-affinity systems. Kinetic parameters for ion influx obtained under different experimental conditions are not strictly comparable. However, it is noteworthy that Imax for the P-sufficient plants of P. vittata was within a similar range of values reported for barley (Asher and Reay, 1979), rice (Oryza sativa; Abedin et al., 2002), and As-nonresistant H. lanatus, D. cespitosa, and A. capillaris (Meharg and Macnair, 1991; Meharg et al., 1994). The Km for P. vittata (0.5–1 μm, Table I) appears to be much lower than the values reported for the above plants species (6–25 μm). This difference may explain the highly efficient uptake of As by P. vittata from low-As soils (Ma et al., 2001). To elucidate whether P. vittata has an enhanced ability to take up arsenate requires a comparison of influx kinetics with an As non-hyperaccumulator fern that is taxonomically close to P. vittata, and has a similar root morphology and growth habit. So far, all accessions of P. vittata and three other Pteris spp. we have tested have proved to be As hyperaccumulators (Zhao et al., 2002).
Ma et al. (2001) reported that, apart from arsenate, P. vittata can also hyperaccumulate As when arsenite, MMA, or DMA was added to soil. However, we show in this study that arsenite was taken up by P. vittata very slowly, at a rate that was about one-tenth of the arsenate influx, when phosphate was not present in the uptake solution. Furthermore, phosphate in the medium did not compete with the uptake of arsenite, and nor did P starvation enhance arsenite influx, indicating that arsenite uptake does not share the same transport systems for phosphate. In the study of Ma et al. (2001), it is possible that arsenite was oxidized to arsenate in the soil before being taken up by the fern. Arsenate is the predominant species of As in aerobic soils (Smith et al., 1998). Mechanisms of arsenite uptake by higher plants remain unclear. Abedin et al. (2002) observed comparable rates of uptake for arsenate and arsenite by paddy rice, which grows in flooded environments where arsenite is the predominant As species. A recent study by Wysocki et al. (2001) showed that arsenite was transported across the plasma membrane of yeast (Saccharomyces cerevisiae) via a glycerol channel protein.
In the experiment with 2-week exposure of arsenate or arsenite, arsenite was taken up in as great a quantity as arsenate by the plants (Table III), yet the rate of arsenite transport was much lower (Fig. 3). The relatively high level of phosphate in the root incubating solution (100 μm) and in the pretreatment conditions (500 μm) will lead to suppression of high-affinity phosphate/arsenate transport (as observed in Fig. 2), and phosphate is highly efficient at competing with arsenate for this transporter (Fig. 2). Both of these factors will lead to suppression of arsenate, but not arsenite uptake.
The most striking feature associated with the As hyperaccumulation by P. vittata lies in the exceedingly efficient transport from roots to shoots. This trait is observed in other heavy metal hyperaccumulators (McGrath et al., 2002). In the depletion experiments (8 h), between 50% and 78% of As taken up had already been transported to the fronds. The frond to root ratio of As concentration was considerably larger than 1 in the long-term As × P experiment (and also Tu and Ma, 2002). In contrast, the shoot to root ratio of As concentration was <0.02 in tomato (Lycopersicum esculentum) (Burló et al., 1999), <0.1 in Brassica juncea (Pickering et al., 2000), and <0.2 in rice (Marin et al., 1992) when arsenate was supplied. The frond to root ratio of P concentration in P. vittata was smaller than 1. The fact that arsenate had little effect on the concentration of P in the fronds, and P had a much smaller effect on frond As concentration than on root As concentration (Fig. 1) suggests that phosphate does not compete with As in the long-distance transport from roots to shoots. Arsenate can be readily reduced to arsenite both enzymatically and nonenzymatically through, e.g. reactions with glutathione (Meharg and Hartley-Whitaker, 2002). If arsenite or its complexes were the main forms of As for xylem loading and transport, then it can be expected that As and P would not be competitive during the processes. The observation that arsenite was more mobile from roots to shoots than arsenate (Table II) supports this explanation. Pickering et al. (2000) showed that addition of the dithiol As chelator dimercaptosuccinate to the nutrient solution markedly enhanced As transport from roots to shoots in the As nonaccumulator B. juncea. Whether arsenite is complexed before xylem loading and xylem transport in P. vittata remains unclear.
As speciation in plant tissues by HPLC-ICP-MS is normally conducted on methanol/water extracts, including previous studies of As-hyperaccumulating ferns where only arsenate and arsenite have been detected (Ma et al., 2001; Francesconi et al., 2002). Such non-physiological extraction may disrupt As speciation in tissues. PC-As complexes, known to be important species of As in a range of plant species, dissociate under alkaline pH conditions to yield both arsenite and arsenate (Meharg and Hartley-Whitaker, 2002). Because of this pH sensitivity, it is thought that As-PC complexes are localized in the vacuole, and disrupting cells under nonphysiological conditions would rapidly lead to an increase in the pH environment of the vacuolar contents. However, in our study where pH was buffered to 5.6, only inorganic As as arsenate and arsenite was detected. SEC would have detected any PC-As complexes, as long as the complexes stayed intact or did not stick to the column. There is a possibility that the As-PC complexes were destroyed by our extraction procedure, but we know from other studies that the As-PC complexes can survive extraction and chromatography, though under different extraction and analytical conditions to those we used here (Sneller et al., 1999). A recent study using x-ray absorption near edge structure spectroscopy showed little evidence of coordination of As with S in a freeze-dried sample of P. vittata fronds (Lombi et al., 2002). Furthermore, the total concentrations of S in fronds were found to be much too small to allow for a significant complexation of arsenite by PCs in a 1:3 stoichiometry of As:thiol (-SH), once the concentration of As in fronds exceeded approximately 2,000 mg kg−1 dry weight (Zhao et al., 2002). Based on the results obtained in this study, we conclude that As is primarily present in inorganic forms in fronds of P. vittita, although we cannot exclude the possibility of complexation. However, it is certain that no simple organo-As compounds, which have been detected in other terrestrial plant species at low levels (i.e. arsenobetaine, arsenocholine, arsenosugars, and methylated forms; Geiszinger et al., 2002; Meharg and Hartley-Whitaker, 2002), were present in the frond extracts. If As is present predominantly as arsenite with some arsenate in P. vittata, this suggests effective storage of these species, either intracellularly (such as in vacuoles) or in specialized cells where disruption of biochemical processes or general plant function is either unimportant or limited because both species disrupt a wide range of metabolic activities (Meharg and Hartley-Whitaker, 2002). A recent study using energy-dispersive x-ray microanalysis showed that As was localized predominately in the vacuoles of the frond epidermal cells of P. vittata (Lombi et al., 2002).
It can be seen from both the efficiency of As extraction from the fronds, and from the As quantification by HPLC-ICP-MS, that other species of As are present in the ferns besides arsenite and arsenate, and that this speciation differs between the arsenate and arsenite exposure treatments. Almost one-half the quantity of As was extracted from the arsenite-treated plants compared with the arsenate-treated plants, where extraction was highly effective (94% extraction; Table III). Because arsenate, arsenite, and the common organo-arsenical species are water soluble, these differences in extractability are probably because of either As incorporation or complexation with structural cellular components. The formation of arsenolipids such as phospholipids or simply a binding to −SH groups or disulfide bridges in different kinds of macromolecules could result in a low extraction efficiency. Because arsenite shows a high affinity for sulfur, the latter scenario is more likely to take place for arsenite, which might be represented in the high abundance of the nonextractable As species in the arsenite-treated ferns. These interactions with cellular constituents are the primary causes of inorganic As toxicity. Because arsenate is readily reduced intracellularly to arsenite in plant cells (Meharg and Hartley-Whitaker, 2002), the differences in extraction efficiencies may reflect differences in toxicology. Differences between the toxicological stress imposed by the arsenate and arsenite treatments may have led to specific changes to the plants' biochemical processing of As, or to a nonspecific stress response that altered As biochemistry.
For the component extracted by phosphate buffer, there is also evidence that other species were present besides arsenate and arsenite, but that these species were not eluted, but rather absorbed to the column. About 20% to 30% more As was quantified from AEC compared with SEC (Table III). For the arsenate treatment, only 50% of the As was detected by the ICP-MS, with 50% remaining stuck to the column. Because the two columns behaved differently with respect to accounting for the As, yet only arsenate and arsenite were detected, this suggests differential breakdown of complexed inorganic As on the columns. The anion exchange was conducted at pH 8, where it is known that As-SH bonds are less stable than at lower pHs.
Although we have gained some indications as to how As is speciated in P. vittata, further work on the induction and regulation of PCs (and potentially metallothionines) should be conducted, intercellular and intracellular localization needs to be ascertained, and As interaction with macromolecules or structural polymers needs investigating.
MATERIALS AND METHODS
Plant Culture
Spores of Pteris vittata were kindly provided by Dr. Lena Ma (University of Florida, Gainesville) and were also obtained from the Oxford Botanical Garden. The spores were sprinkled on to moist potting compost in a seed tray. The tray was covered with a plastic cling film to maintain moisture. After spore germination, the prothalli developed and were fertilized, and grew into sporelings with true leaves (fronds). At the two-frond stage, sporelings were transplanted individually into 8-cm pots containing a general purpose potting compost, and allowed to grow to four to five fronds. Plant roots were then washed carefully with tap water to remove adhering compost, and transferred to 250-mL pots (one plant per pot) containing a nutrient solution, which was a modified Hoagland solution with one-half-strength major nutrients and full-strength micronutrients (except that Fe was supplied as Fe-EDDHA at 100 μm; Hewitt, 1966). The nutrient solution was aerated continuously and renewed weekly. Experiments were conducted inside a controlled environment growth chamber with the following conditions: 16-h light period with a light intensity of 350 μmol m−2 s−1, 25°C/20°C day/night temperature, and 60% relative humidity.
Arsenate × Phosphate Interactions Experiment
Fern plants raised from the spores from Florida were used in this experiment. After 3 weeks of preculture in hydroponics, plants were treated with different combinations of phosphate and arsenate concentrations. The phosphate concentrations were 20, 100, and 500 μm (supplied as NH4H2PO4), and the concentrations of arsenate (Na2HAsO4) were 0, 83, 416, and 2080 μm. The concentration of NH4+ was maintained at 1,000 μm in all treatments by varying the concentration of NH4Cl. All other nutrients were maintained at the same concentrations as in the preculture solution. Solution pH was adjusted to 7.0. Nutrient solutions were topped up every day and renewed once every 4 d. Each treatment was replicated 4-fold. Plants were harvested 18 d after the treatments were imposed. Roots and shoots were separated, and washed thoroughly with deionized water. Plant tissues were dried at 60°C for 48 h and dry weights recorded. Ground plant materials were digested with a mixture of HNO3:HClO4 (85:15 [v/v]), and the concentrations of As and P were determined using ICP-atomic emission spectroscopy (Fisons-ARL Accuris, Ecublens, Switzerland). The detection limit of the instrument for As in solution was 0.02 mg L−1. Blanks and internal standards were included for quality assurance.
Short-Term Uptake Kinetics of Arsenate and Arsenite
Fern plants raised from the spores obtained from the Oxford Botanical Garden were used in this experiment. The kinetics of arsenate and arsenite influx were studied using a solution depletion technique similar to that described by Claassen and Barber (1974) and by Drew et al. (1984). After 3 weeks of preculture in hydroponics, roots of intact plants were rinsed with deionized water and transferred to a pretreatment solution containing 0.5 mm CaCl2 and 5 mm MES with pH adjusted to 6.0. Twelve hours later and when the light period had already started for 2 h, the pretreatment solution was replaced with 240 mL of uptake solution containing either 5 μm arsenate (Na2HAsO4) or 5 μm arsenite (NaAsO2), together with 0.5 mm CaCl2 and 5 mm MES with pH adjusted to 6.0. The uptake solution was aerated vigorously and continuously. At 0, 15, and 30 min and thereafter every 30 min at intervals up to 8 h, 0.3 mL of uptake solution was removed for the determination of As concentration, and replaced with 0.3 mL of deionized water. Water losses through transpiration were compensated by additions of deionized water at hourly intervals. The temperature was maintained at 25°C ± 0.5°C. After 8 h, roots were separated from shoots, rinsed with deionized water, blotted dry, and weighed. The concentration of As in the uptake solution was determined using atomic absorption spectroscopy (4100ZL, Perkin-Elmer Applied Biosystems, Foster City, CA) equipped with a flow injection hydride generation unit (FIAS 400, Perkin-Elmer Applied Biosystems), after a prereduction step using KI and ascorbic acid. The instrument has a detection limit for As in solution of 0.1 μg L−1, equivalent to 0.05 μm As in the uptake solution. Dried plant tissues were ground and digested with HNO3/HClO4, and the concentration of As determined using ICP-atomic emission spectroscopy.
The depletion experiment was carried out for both arsenate and arsenite under three different treatments: (a) control, in which P-sufficient plants were used and the uptake solution contained no phosphate; (b) control + P, in which P-sufficient plants were used and the uptake solution contained 50 μm phosphate (Na2HPO4); and (c) P deficient, in which P-deficient plants were used by withholding P supply for 8 d before the uptake experiment, and the uptake solution contained no phosphate. Each treatment was replicated 5-fold.
Where possible, a modified Michaelis-Menten kinetic model (Eq. 1) was fitted to the depletion data, according to Claassen and Barber (1974) and Drew et al. (1984):
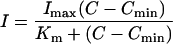 |
1 |
where I is the net influx, expressed as nmol g−1 root fresh weight h−1; Imax is the maximum net influx at saturating concentrations; Km is the ion concentration when I = ½ Imax; and Cmin is the minimal ion concentration below which net influx ceases.
As Speciation in P. vittata Fronds
After a 3-week preculture in hydroponics, P. vittata (from Oxford) plants were given either 50 μm arsenate (Na2HAsO4) or 50 μm arsenite (NaAsO2). The composition of other nutrients was the same as in the preculture, except that the phosphate concentration was decreased to 100 μm. The nutrient solution was topped up every day and renewed every week. Each treatment was replicated in three pots. After a 2-week exposure to As, plants were separated into roots and fronds, and washed with deionized water. For the As speciation, the fronds were crushed mechanically with a homogenizer in a suspension of 30 mm ammonium phosphate buffer at pH 5.6 (about 150 mg fresh weight mL−1). After 20 min, the suspension was centrifuged and the supernatant was stored at 4°C overnight. This solution was analyzed after being diluted with the buffer (1:70–1:100 [v/v]) and filtered through a 0.45-μm filter for its total As content by ICP-MS (Spectromass 2000, Spectro Analytical Instruments) directly, and for the As speciation by SEC and AEC. Quantitative determination of As in frond extracts and in the acid digests (microwave digestion for 10 min at 100% power with concentrated suprapur HNO3) was carried out with external calibration of the ICP-MS with arsenate standards. The As content in the standard reference material IAEA 140 (seaweed) was determined to be 42 ± 1 mg kg−1 (reference value of 42.2–46.4 mg kg−1).
The analytical setup for As speciation analysis consisted of two HPLC pumps (LKB, Bromma, Sweden), one for the separation and the other for the post-column addition of a cesium solution (20 μg L−1) as continuous internal standard. The outlet of the separation column was mixed with the standard solution via a “T” piece and then directly connected to a Meinhard nebulizer (GE Meinhard Assoc., Inc., Santa Ana, CA) and a water-jacketed cyclonic spray chamber of the ICP-MS. The ICP-MS was set up for the collection of transient signals for 75, 77, and 133 m/z with a dwell time of 500 ms to monitor As, a possible chloride interference (ArCl), and the internal standard cesium. The ICP-MS was optimized daily with an As standard solution (10 μg L−1). For the SEC, Asahipak GS 220 (250 × 7.5 mm, Showa, Denko K.K., Tokyo) with a sample loop of 20 μL was used. The same buffer as employed for extraction was used (30 mm ammonium phosphate buffer, pH 5.6) and pumped at 1 L min−1 through the system. For the anion-exchange chromatography, a PRPX 100 Hamilton (150 × 4.6 mm) + pre-column was used with 20 mm ammonium carbonate buffer (pH 8.08) as the mobile phase at 1 mL min−1. Comparisons of both retention times (SEC and AEC) with standards were used for peak identification and quantification was done by measuring peak areas of the individual species.
The buffer was prepared from suprapur phosphoric acid and ammonia (25% [w/v] suprapur) or ammonium carbonate. For speciation, As(III) was prepared from As2O3, As(V) was prepared from Na3AsO4, DMA(V) was prepared from Na-DMA, MMA(V) was prepared from Na2-MMA (a kind donation from Prof. William R. Cullen, University of British Columbia, Vancouver, Canada), and DMA(III) and MMA(III) were reduced from their pentavalent forms using thiosulfate and bisulfite according to the method of Reay and Asher (1977). All chemicals used were of the analytical grade or better, and were from BDH (Poole, UK), and deionized water was used throughout.
ACKNOWLEDGMENTS
We thank Dr. Lena Ma (University of Florida), Kate Pritchard (University of Oxford Botanic Garden), and Dr. Jackie Barker (IACR-Long Ashton) for providing spores of P. vittata, and Miss Sarah Dunham for technical assistance. J. Wang thanks China Scholarship Council for a one-year fellowship.
Footnotes
This work was supported by the Biotechnology and Biological Sciences Research Council of the UK (grant to Rothamsted Research).
Article, publication date, and citation information can be found at www.plantphysiol.org/cgi/doi/10.1104/pp.008185.
LITERATURE CITED
- Abedin MJ, Feldmann J, Meharg AA. Uptake kinetics of arsenic species in rice plants. Plant Physiol. 2002;128:1120–1128. doi: 10.1104/pp.010733. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Asher CJ, Reay PF. Arsenic uptake by barley seedlings. Aust J Plant Physiol. 1979;6:459–466. [Google Scholar]
- Burló F, Guijarro I, Carbonell-Barrachina AA, Valero D, Martinez-Sánchez F. Arsenic species: effects on and accumulation by tomato plants. J Agric Food Chem. 1999;47:1247–1253. doi: 10.1021/jf9806560. [DOI] [PubMed] [Google Scholar]
- Claassen N, Barber SA. A method for characterizing the relation between nutrient concentration and flux into roots of intact plants. Plant Physiol. 1974;54:564–568. doi: 10.1104/pp.54.4.564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drew MC, Saker LR, Barber SA, Jenkins W. Changes in the kinetics of phosphate and potassium absorption in nutrient-deficient barley roots measured by a solution-depletion technique. Planta. 1984;160:490–499. doi: 10.1007/BF00411136. [DOI] [PubMed] [Google Scholar]
- Francesconi K, Visoottiviseth P, Sridokchan W, Goessler W. Arsenic species in an arsenic hyperaccumulating fern, Pityrogramma calomelanos: a potential phytoremediator of arsenic-contaminated soils. Sci Total Environ. 2002;284:27–35. doi: 10.1016/s0048-9697(01)00854-3. [DOI] [PubMed] [Google Scholar]
- Geiszinger A, Goessler W, Kosmus W. Organoarsenic compounds in plants and soil on top of an ore vein. Appl Organomet Chem. 2002;16:245–249. [Google Scholar]
- Hartley-Whitaker J, Ainsworth G, Vooijs R, Ten Bookum W, Schat H, Meharg AA. Phytochelatins are involved in differential arsenate tolerance in Holcus lanatus. Plant Physiol. 2001;126:299–306. doi: 10.1104/pp.126.1.299. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hewitt EJ. Sand and Water Culture Methods Used in the Study of Plant Nutrition. Ed 2. Bucks, UK: Commonwealth Agricultural Bureaux, Farnham Royal; 1966. [Google Scholar]
- Kabata-Pendias A, Pendias H. Trace Elements in Soils and Plants. Ed 2. Boca Raton, FL: CRC Press; 1992. [Google Scholar]
- Lee RB. Selectivity and kinetics of ion uptake by barley plants following nutrient deficiency. Ann Bot. 1982;50:429–449. [Google Scholar]
- Liu CM, Muchhal US, Uthappa M, Kononowicz AK, Raghothama KG. Tomato phosphate transporter genes are differentially regulated in plant tissues by phosphorus. Plant Physiol. 1998;116:91–99. doi: 10.1104/pp.116.1.91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lombi E, Zhao FJ, Fuhrmann M, Ma LQ, McGrath SP (2002) Arsenic distribution and speciation in the fronds of the hyperaccumulator Pteris vittata. New Phytol (in press) [DOI] [PubMed]
- Ma LQ, Komar KM, Tu C, Zhang WH, Cai Y, Kennelley ED. A fern that hyperaccumulates arsenic: a hardy, versatile, fast-growing plant helps to remove arsenic from contaminated soils. Nature. 2001;409:579–579. doi: 10.1038/35054664. [DOI] [PubMed] [Google Scholar]
- Marin AR, Masscheleyn PH, Patrick WH. The influence of chemical form and concentration of arsenic on rice growth and tissue arsenic concentration. Plant Soil. 1992;139:175–183. [Google Scholar]
- McGrath SP, Zhao FJ, Lombi E. Phytoremediation of metals, metalloids, and radionuclides. Adv Agron. 2002;75:1–56. [Google Scholar]
- Meharg AA, Hartley-Whitaker J. Arsenic uptake and metabolism in arsenic resistant and nonresistant plant species. New Phytol. 2002;154:29–43. [Google Scholar]
- Meharg AA, Macnair MR. The mechanisms of arsenate tolerance in Deschampsia cespitosa (L.) Beauv and Agrostis capillaris L. New Phytol. 1991;119:291–297. doi: 10.1111/j.1469-8137.1991.tb01033.x. [DOI] [PubMed] [Google Scholar]
- Meharg AA, Macnair MR. Suppression of the high-affinity phosphate uptake system: a mechanism of arsenate tolerance in Holcus lanatus L. J Exp Bot. 1992;43:519–524. [Google Scholar]
- Meharg AA, Naylor J, Macnair MR. Phosphorus nutrition of arsenate tolerant and nontolerant phenotypes of velvetgrass. J Environ Qual. 1994;23:234–238. [Google Scholar]
- Muchhal US, Pardo JM, Raghothama KG. Phosphate transporters from the higher plant Arabidopsis thaliana. Proc Natl Acad Sci USA. 1996;93:10519–10523. doi: 10.1073/pnas.93.19.10519. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Muchhal US, Raghothama KG. Transcriptional regulation of plant phosphate transporters. Proc Natl Acad Sci USA. 1999;96:5868–5872. doi: 10.1073/pnas.96.10.5868. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pickering IJ, Prince RC, George MJ, Smith RD, George GN, Salt DE. Reduction and coordination of arsenic in Indian mustard. Plant Physiol. 2000;122:1171–1177. doi: 10.1104/pp.122.4.1171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reay PF, Asher CJ. Preparation and purification of 74As-labeled arsenate and arsenite for use in biological experiments. Anal Biochem. 1977;78:557–560. doi: 10.1016/0003-2697(77)90117-8. [DOI] [PubMed] [Google Scholar]
- Schmöger MEV, Oven M, Grill E. Detoxification of arsenic by phytochelatins in plants. Plant Physiol. 2000;122:793–801. doi: 10.1104/pp.122.3.793. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith E, Naidu R, Alston AM. Arsenic in the soil environment: a review. Adv Agron. 1998;64:149–195. [Google Scholar]
- Smith FW, Ealing PM, Dong B, Delhaize E. The cloning of two Arabidopsis genes belonging to a phosphate transporter family. Plant J. 1997;11:83–92. doi: 10.1046/j.1365-313x.1997.11010083.x. [DOI] [PubMed] [Google Scholar]
- Sneller FEC, Van Heerwaarden LM, Kraaijeveld-Smit FJL, Ten Bookum WM, Koevoets PLM, Schat H, Verkleij JAC. Toxicity of arsenate in Silene vulgaris, accumulation and degradation of arsenate-induced phytochelatins. New Phytol. 1999;144:223–232. [Google Scholar]
- Tu C, Ma LQ. Effects of arsenic concentrations and forms on arsenic uptake by the hyperaccumulator ladder brake. J Environ Qual. 2002;31:641–647. [PubMed] [Google Scholar]
- Ullrich-Eberius CI, Sanz A, Novacky AJ. Evaluation of arsenate- and vanadate-associated changes of electrical membrane potential and phosphate transport in Lemna gibba-G1. J Exp Bot. 1989;40:119–128. [Google Scholar]
- Wysocki R, Chery CC, Wawrzycka D, Van Hulle M, Cornelis R, Thevelein JM, Tamas MJ. The glycerol channel Fps1p mediates the uptake of arsenite and antimonite in Saccharomyces cerevisiae. Mol Microbiol. 2001;40:1391–1401. doi: 10.1046/j.1365-2958.2001.02485.x. [DOI] [PubMed] [Google Scholar]
- Zhao FJ, Dunham SJ, McGrath SP. Arsenic hyperaccumulation by different fern species. New Phytol. 2002;156:27–31. [Google Scholar]