

Abstract
We used DNA microarrays of the Escherichia coli genome to trace the progression of chromosomal replication forks in synchronized cells. We found that both DNA gyrase and topoisomerase IV (topo IV) promote replication fork progression. When both enzymes were inhibited, the replication fork stopped rapidly. The elongation rate with topo IV alone was 1/3 of normal. Genetic data confirmed and extended these results. Inactivation of gyrase alone caused a slow stop of replication. Topo IV activity was sufficient to prevent accumulation of (+) supercoils in plasmid DNA in vivo, suggesting that topo IV can promote replication by removing (+) supercoils in front of the chromosomal fork.
Keywords: topo IV, coumarins, positive supercoiling
Topoisomerases are needed in all three phases of bacterial DNA replication: initiation at the origin of replication, elongation with progressive outward movement of the forks until they meet halfway around the chromosome, and termination, including the final disentanglement of the catenated daughter chromosomes. Bacterial cells have two indispensable topoisomerases: gyrase and topoisomerase IV (topo IV) (1). (−) supercoiling by gyrase is involved in all three phases of replication: (−) supercoiling facilitates unwinding in initiation, reduces links in front of the fork during elongation, termed the swivel function (2), and compacts DNA in termination.
It was widely believed that gyrase was the only topoisomerase needed during elongation (3, 4). This was inferred from the observation that replication stops in the absence of gyrase (5), whereas inhibition of topo IV has only a small effect on DNA synthesis (6). However, the interpretation of past results is complicated by the multiple roles of gyrase in replication and the failure until quite recently to recognize that drugs used to inhibit gyrase also inhibit topo IV (6, 7).
Here we demonstrate that topo IV can indeed act as a replication swivel. Our demonstration depended on a use of a whole genome Escherichia coli microarray and on the observation that topo IV is a secondary target for coumarin antibiotics such as novobiocin. The evidence for the role of topo IV in DNA elongation is supported by our findings that topo IV and gyrase have overlapping roles in replication fork movement and that topo IV is sufficient to remove (+) supercoils in vivo.
Materials and Methods
Bacterial Strains and Plasmids.
The bacterial strains used in this study are listed in supplemental Table 1 (see www.pnas.org). Plasmids pBR322 and pGP509 have been described in refs. 8 and 9, respectively.
Enzymes, Reactions, and Chemicals.
Norfloxacin, novobiocin, coumermycin A1, hydroxyurea, and pancreatic DNaseI were obtained from Sigma. DNA supercoils were removed by nicking substrates with pancreatic DNase I in the presence of 300 μg/ml ethidium bromide. Assays of plasmid topology and DNA replication were done as previously described (6).
Use of Genomic Microarrays to Study DNA Replication.
Four thousand one hundred fifteen of all 4,290 annotated E. coli ORFs (10) were successfully PCR amplified by using primer pairs (Sigma–Genosys) kindly provided by Fred Blattner (University of Wisconsin, Madison, WI). These PCR products were printed onto polylysine-coated glass microscope slides as described in the MGuide at http://cmgm.stanford. edu/pbrown/mguide/index.html. W3110 ΔacrAdnaC2 cells were grown in LB + 0.2% glucose at 30°C and shifted to 42°C for 70 min. Half of the culture was then shifted to 30°C by addition of 18°C media, and the other half remained at the restrictive temperature for 20 min more before it was harvested as a nonreplicating reference. We harvested a sample of the culture 30 min after shifting it to 30°C and then added novobiocin to the cultures. The time of novobiocin addition was designated time 0. Samples taken at each successive time point were added to NaN3, and frozen in liquid nitrogen. Genomic DNA was prepared by using a Qiagen (Chatsworth, CA) column and digested with Sau3A1 (Roche Molecular Biochemicals). Genomic DNA labeling and microarray hybridization are described at http://cmgm.stanford.edu/pbrown/protocols/index.html. The 16-bit TIFF images were acquired by using the AxonScanner (genepix 1.0, Axon Instruments, Foster City, CA). After fluorescence, ratios were extracted for each element on the array by using scanalyze 2.1, all ratios were normalized on a logarithm scale such that the average ratio of 100 ORFs near terB (b#1608–1712) was equal to 1. A moving average of the fluorescence ratios for 10 adjacent ORFs was calculated, and the results were mapped onto the E. coli chromosome by using the circlemaker program (J.D., unlicensed software). Different moving averages did not affect the positioning of the replication forks. Curiously, the ratios in the replicated region averaged 1.5 to 1.6 rather than 2; this may be the result of incomplete initiation in the strain. Therefore, in Fig. 3, ORFs with a moving average ratio <1.2 were colored in gray; those with a moving average of >1.2 were colored in red.
Figure 3.
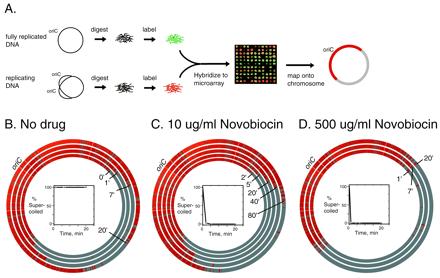
Roles of topo IV and gyrase in replication fork progression. (A) Schematic illustrating the use of genome microarrays to trace replication fork progression. Replication was synchronized in a temperature-sensitive dnaC2 initiation mutant. Fluorescently labeled genomic DNAs from a synchronized replicating culture (red) and a fully replicated reference DNA (green) were hybridized together to a glass slide containing DNA from 4,115 individual ORFs of E. coli. A representative sector of an array is shown. The fluorescence ratio in each spot indicates whether the gene was replicated. Each spot was mapped to its position on the E. coli chromosome. Replicated genes are shown in red. (B–D) Plasmid pBR322 and genomic DNAs were isolated from the cells at the indicated time points. The first time point (“0”) from the replicating DNA was taken 30 min after the shift to 30°C and immediately before the addition of novobiocin to the indicated concentrations. The positions of the clockwise replication forks at times after inhibition of gyrase or of gyrase and topo IV are shown as concentric circles superimposed on the E. coli physical map. oriC marks the origin of replication. The percent of pBR322 DNA that was supercoiled at each time point was determined by agarose gel electrophoresis and is shown in the Insets.
Gel Electrophoresis and Quantification.
To detect catenated plasmid DNA species, nicked plasmid DNA was resolved by high-resolution agarose gel electrophoresis (11). Topoisomers of closed circular DNA of pBR322 were resolved in one dimension on a 1% agarose Tris-acetate–EDTA (TAE) gel containing 0, 1, or 10 μg/ml of chloroquine. Two-dimensional gel electrophoresis was performed by resolving plasmid DNA in a 1% agarose TAE gel in the first dimension and in a 1% agarose gel in TAE + 10 μg/ml chloroquine in the second dimension. Southern blots of gels were quantified by PhosphorImager (Molecular Dynamics).
Results
Topo IV Is a Secondary Target of Novobiocin and Can Support Elongation of DNA Replication in Vivo.
If the elongation phase of bacterial DNA synthesis specifically requires gyrase to unlink the parental DNA, then inactivation of gyrase should cause a rapid arrest of DNA replication. We inactivated both subunits of the enzyme by treating the gyrAts mutant with novobiocin at 42°C. Novobiocin is a specific inhibitor of the gyrB subunit of gyrase (12). To increase sensitivity to novobiocin, we used acrA derivatives, which have impaired drug efflux (13). This double inhibition caused only a slow arrest of DNA replication (Fig. 1A). This is not a result of slow inactivation of gyrase, because inactivation of gyrase in the gyrAts mutant by temperature alone caused the supercoiling density of a reporter plasmid to drop within seconds, reaching the fullest extent of relaxation in 1–2 min (see supplemental Fig. 6 at www.pnas.org).
Figure 1.
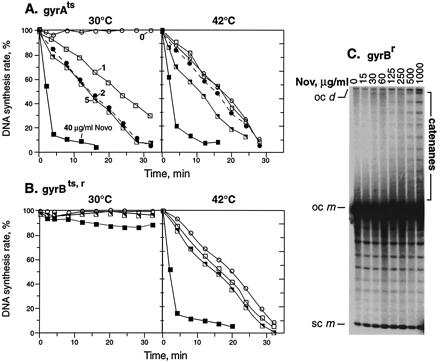
Effects on DNA replication of novobiocin addition and mutational inactivation of gyrase. Novobiocin (Nov) was added to acrAgyrA43ts cells (A) or acrA gyrBts,r novobiocin-resistant cells (B) at time 0. Cells were pulsed for 1 min with [3H]thymidine at each time point and acid precipitable counts determined. (A and B) Novobiocin concentrations were (○) 0 μg/ml; (□) 1 μg/ml; (●) 2 μg/ml; (└) 5 μg/ml; and (▪) 40 μg/ml. The rate of DNA synthesis shown is the percentage of the rate in an isogenic wild-type strain grown in parallel but without drug. (C) A acrA+gyrBr strain was treated with the indicated concentrations of novobiocin for 15 min at 30°C, and plasmid DNA from the strain was nicked and analyzed by agarose gel electrophoresis. Positions of catenanes, supercoiled monomers (sc m), open circular monomer (oc m), and open circular dimer (oc d) are indicated.
These results suggested that a second topoisomerase also acts as a swivel. The second swivel activity was sensitive to high concentrations of novobiocin. Novobiocin concentrations of 40 μg/ml rapidly shut down DNA replication at either the permissive or restrictive temperature for the gyrAts mutant (Fig. 1A). In the control with a gyrBts,r mutant that is resistant to novobiocin as well as temperature sensitive, DNA replication showed only about a 10% decrease in 40 μg/ml drug at the permissive temperature for gyrase (Fig. 1B). However, at the restrictive temperature, DNA synthesis was inhibited slowly below 10 μg/ml novobiocin but rapidly at the higher drug concentration where the secondary target is affected (Fig. 1B).
We proved in several ways that this secondary target is topo IV. The signature activity of topo IV is its essential role in decatenating the products of replication (14). We showed that catenated plasmids accumulate in novobiocin-resistant gyrase strains exposed to high novobiocin concentrations that rapidly halt DNA synthesis (Fig. 1C). The concentrations of novobiocin used are higher than in Fig. 1A, because the results shown are for an acrA+ strain.
The most direct evidence for the involvement of topo IV in elongation is provided by the use of conditional mutations in the genes encoding topo IV, parC, and parE. We combined novobiocin inhibition of gyrase with mutational inactivation of topo IV. In an acrA strain, a low concentration of novobiocin, 5 μg/ml, inhibited gyrase and caused a slow replication stop in both parE+ and parEts strains at 30°C. On shift to 42°C, DNA synthesis dropped precipitously (replication was halved in only 3.5 min) in a parEts strain but much more slowly in the parE+ control (Fig. 2A). After many unsuccessful attempts to construct strains of E. coli that would be temperature sensitive for both gyrase and topo IV, we succeeded in constructing the double mutant in Salmonella typhimurium. We compared the ability of this Salmonella strain to support replication at permissive and restrictive temperatures with single mutations in either enzyme. Inhibition of topo IV alone in this strain had only a slight effect on DNA synthesis, as expected (6, 15). Replication stopped slowly in the gyrAts single mutant at the restrictive temperature (replication was halved in 16 min). In the gyrAts parCts double mutant, replication stopped much faster at the restrictive temperature; the rate halved in only 4 min (Fig. 2B). Therefore, in E. coli and S. typhimurium it is necessary to inhibit both topo IV and gyrase to stop replication rapidly.
Figure 2.
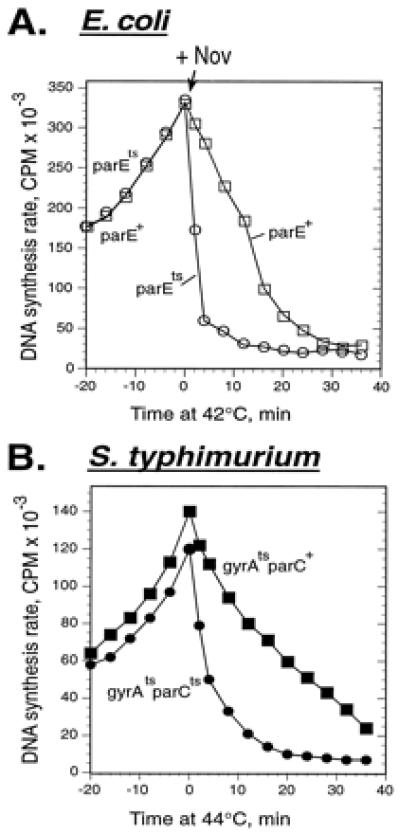
In the absence of gyrase, inhibition of topo IV causes rapid replication arrest. (A) E. coli strains acrA parE+ (□) and acrA parEts (○) were grown at 30°C. At time 0, novobiocin (Nov) was added to 5 μg/ml, and the cultures were simultaneously shifted to 42°C. The rates of DNA synthesis were measured at the indicated times. (B) S. typhimurium strains gyrA208 (gyrAts, ▪) and gyrA208parEts (●) were grown at 30°C. At time 0, they were shifted to 44°C. The rates of DNA synthesis were measured at the indicated times after temperature shift.
Use of Genomic Microarrays to Measure the Rate of Replication Fork Movement.
Direct proof of the roles of topoisomerases in DNA synthesis required measurement of replication fork progression in vivo. To do so, we developed an original method that took advantage of genomic microarray technology (16). The method is illustrated in Fig. 3A. In a synchronous population of cells, those genes that have replicated are twice as abundant as unreplicated genes. We positioned the replication forks by measuring the abundance of each gene in E. coli from the amount of DNA that hybridizes to the array. We constructed microarrays containing 96% of the E. coli ORFs amplified by PCR and printed onto glass slides. We synchronized replication in a dnaC2 mutant by shifting the temperature to 42°C for 90 min to allow the completion of a round of replication and by returning the culture to 30°C for initiation of DNA synthesis (17). We added novobiocin to the indicated concentrations to inhibit gyrase and topo IV at time “0,” 30 min after the shift to 30°C. At this time, the forks had moved about halfway around the chromosome. We took samples at times thereafter and isolated genomic DNA from the cells. We labeled this partially replicated DNA by randomly primed synthesis with Cy5 (red). Completely replicated reference DNA (from 42°C cultures) was similarly labeled with Cy3 (green), and the two differentially labeled DNAs were mixed and hybridized to the microarray. The relative copy number of each gene in the replicating chromosome could then be determined from the ratio of Cy5 to Cy3 fluorescence at the array elements corresponding to the genes. The reference DNA set a gene dosage of 1 for each ORF. Genes from the replicating culture present at the level of the reference (unreplicated genes) are colored gray on the conventional circular genetic map of E. coli; those that are more abundant (replicated genes) are in red on the map (Fig. 3 B–D). To dampen out fluctuations, a moving average was applied over 10 ORFs. The results are shown as concentric circles representing the abundance of nearly every E. coli gene at times after the zero reference (Fig. 3 B–D). There is an almost uniform red color starting from the origin of replication, uninterrupted by gray, and a near absence of red-colored genes in the unreplicated area. As a result, replication fork boundaries are sharply defined.
Fig. 3B shows the extent of replication for the control in the absence of novobiocin at 1, 7, and 20 min after time 0. Replication is bidirectional, with both forks progressing linearly at the same rate. Fig. 3B Inset shows that the supercoiling of a reporter plasmid in this strain remained maximal, as expected, because gyrase is fully functional.
In a parallel experiment, we added a high concentration of novobiocin (500 μg/ml), which we determined was sufficient to inhibit both gyrase and topo IV completely in this strain (even though this strain is acrA−, high concentrations were needed to completely inhibit the topoisomerase activity). This level of drug rapidly stopped [3H]thymidine incorporation (data not shown) and caused relaxation of an internal reference plasmid DNA within 1 min (Fig. 3D Inset). At this novobiocin concentration, the fork boundary continued to map to the same ORF over the entire time course of the experiment (Fig. 3D). We then tested the effect of intermediate levels of novobiocin: 2, 5, 10, and 50 μg/ml. The results for a time series after addition of 10 μg/ml novobiocin are shown in Fig. 3C. At this concentration of drug, gyrase is completely inhibited (Fig. 3C Inset), and thus the replication fork movement is supported by topo IV alone.
The derived positions of the replication forks as a function of time for all drug concentrations are plotted in Fig. 4. Because the two forks moved at the same rates (within error) under all conditions, the results shown in Fig. 4 are average values. Without drug, the rate of fork progression was constant, with time at 45 kb⋅min−1, which extrapolates to 51 min to replicate the entire chromosome. In this strain, 2 μg/ml of novobiocin slowly inhibits gyrase, and the rate of fork movement is reduced only slightly. Five micrograms per milliliter or 10 μg/ml of novobiocin effectively inhibit gyrase, as determined by plasmid DNA relaxation (Fig. 3C Inset) and drop the rate of fork progression to roughly 14 kb/min. We conclude that this is the rate of replication supported by topo IV alone. At later times, there is a progressive further decrease in the rate of replication, which we believe is the result of secondary effects caused by the relaxation of the chromosome.
Figure 4.
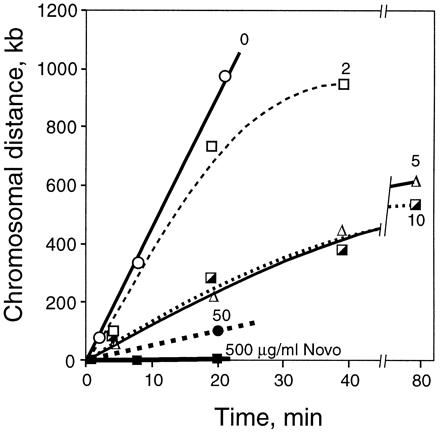
Rates of chromosomal replication supported by gyrase and topo IV. acrA dnaC2 cells were synchronized, and the position of the replication forks was determined by using hybridization to a genomic microarray as shown in Fig. 3. The plots show the average distance of the two forks relative to their respective positions at the time of novobiocin addition (time 0). Thirty minutes after the shift to 30°C to restart replication, novobiocin was added to 0 (○), 2 (□), 5 (Δ), 10 (┌), 50 (●), or 500 (▪) μg/ml. The times plotted are after addition of drug. The initial slopes (between 0 and 20 min) of the curves are: 0 drug, 45 kb/min; 2 μg/ml novobiocin, 35 kb/min; 5 and 10 μg/ml, 13.5 kb/min; 50 μg/ml, 5.1 kb/min; and 500 μg/ml, 0 kb/min.
Accumulation of Positively Supercoiled DNA Requires Inhibition of Both Gyrase and Topo IV.
Topo IV could support replication fork movement directly by removing (+) supercoils in front of the replication fork, as gyrase does. It is also possible that topo IV acts indirectly by removing (+) precatenanes behind the replication fork, as these should be in equilibrium with the (+) supercoils ahead of the fork (18, 19). Although activity on either (+) supercoils in front of the fork or (+) precatenanes behind the replication fork could constitute a swivel function, for simplicity we focused on the ability of topo IV to remove (+) supercoils in vivo.
Positive supercoils can be generated in vivo through the twin domain effect associated with transcription (20). RNA polymerase causes transitory (+) supercoils ahead of the transcription complex and (−) supercoiling behind (21). It has been thought that coumarins and quinolones generate net (+) supercoiling by inhibiting gyrase and thereby removing the activity that converts (+) supercoils to (−) supercoils (20, 22). However, these drugs inhibit topo IV as well. In all four of our E. coli conditional gyrase mutants, the mean σ of plasmids changed rapidly on temperature upshift from −0.065 to −0.01. In no case, though, was (+) supercoiled plasmid DNA produced, even after 60 min at the restrictive temperature. Therefore, inhibition of gyrase alone is not sufficient to allow (+) supercoiling of DNA.
We directly implicated topo IV in (+) supercoil removal. We shifted acrAparEts cells to the nonpermissive temperature for topo IV and treated them with a low concentration of novobiocin that selectively inhibits gyrase, conditions that cause a fast stop in DNA synthesis (Fig. 1A). Up to 30% of resident plasmid pBR322 was highly (+) supercoiled after 5 min at the restrictive temperature (Fig. 5A, lane 8). However, treating the cells with the same amount of novobiocin at the permissive temperature for topo IV, which results in inhibition of gyrase alone, only relaxed the DNA (Fig. 5A, lane 4). The gyrase and topo IV double mutants of S. typhimurium accumulated positively supercoiled plasmid DNA at the nonpermissive temperature (data not shown).
Figure 5.
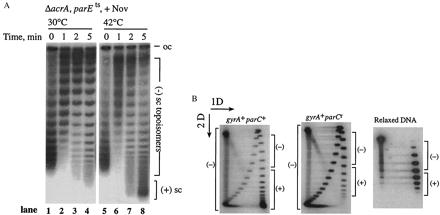
Topo IV efficiently relaxes positively supercoiled DNA in vivo. (A) pBR322 DNA was isolated from an E. coli ΔacrAparE10 strain after addition of 5 μg/ml novobiocin (Nov). Novobiocin was added at time 0, and cultures were either kept at 30°C (lanes 1–4) or shifted to 42°C (lanes 5–8). DNA was resolved on a 1% TAE gel with 10 μg/ml chloroquine. The positions of (−) supercoiled, [(−) sc], and (+) supercoiled, [(+) sc] topoisomers are shown. (B) pBR322 DNA was isolated from parC+ (Left) and parCr norfloxacin-resistant (Right) E. coli strains that had been treated with 100 μg/ml norfloxacin. The DNA was analyzed by two-dimensional agarose gel electrophoresis. The first dimension (1D) was a 1% TAE agarose gel without chloroquine, and the second dimension (2D) contained 10 μg/ml chloroquine. Reference plasmid DNA relaxed in vitro by calf thymus topoisomerase I is shown (Lower). The positions of (+) and (−) topoisomers are shown.
We independently demonstrated that topo IV efficiently relaxed (+) supercoiled DNA by using norfloxacin, a quinolone antibiotic, to inhibit gyrase. The availability of parC mutants resistant to quinolones (6) permitted a stringent control for the experiments. We used two-dimensional gel analysis to examine the entire topological distribution of plasmid DNA. When a gyrA+parC+ strain was treated with enough norfloxacin to inhibit both gyrase and topo IV, a substantial fraction of the plasmid DNA became (+) supercoiled (Fig. 5B Left, compare with relaxed DNA marker at right). The (+) supercoiled DNA was absent when the gyrA+parCr control was treated with drug (Fig. 5B Center). Similar experiments with a gyrArparC+ strain showed no significant changes in plasmid supercoiling (data not shown).
We conclude topo IV removes (+) supercoils efficiently in vivo, and that this activity may contribute to its role in the advancement of the replication fork.
Discussion
Topoisomerases play multiple roles in replication. Negative supercoiling by gyrase is needed for all three phases of replication: it aids unwinding at the origin, reduces the linking number of DNA to facilitate replication fork progression during elongation, and compacts DNA to promote decatenation and disentanglement by topo IV to complete replication (1, 4, 23). We have shown here that topo IV also has multiple roles. In addition to its established role in decatenation, topo IV promotes fork progression, at least in part, by relaxing the (+) supercoils generated by replication.
Previously, gyrase had been thought to be the sole swivel for replication. This conclusion was based in part on the rapid inhibition of DNA synthesis by quinolones and coumarins, originally identified as inhibitors of gyrase alone. The present work with coumarins and previous results with quinolones (6) show that topo IV is a secondary target of both families of drugs. A second reason that gyrase was thought to be solely responsible for elongation is that mutational inactivation of gyrase but not topo IV arrests DNA synthesis (5, 15). However, nearly all gyrase mutations cause only a slow stop in DNA synthesis under nonpermissive conditions (24–27). In our experiments with five different gyrase mutations, the rate of DNA synthesis was halved in about 20 min, or one-half of the generation time (Figs. 1 and 2 and data not shown). If gyrase were solely responsible for fork progression, the effect on DNA synthesis should be much faster. Indeed, when both gyrase and topo IV were inactivated, replication was halved within minutes (Figs. 1 and 2). These experiments imply that both topo IV and gyrase act as replication swivels. Exceptional gyrase mutants have been found (5, 28) that can cause a fast stop in DNA synthesis. We suggest that these mutations, like quinolone antibiotics, can convert gyrase into an SOS-inducing poison of replication (29, 30).
Topo IV supported fork movement at about 1/3 of the rate when both gyrase and topo IV were active (Fig. 4). We believe that the gradual cessation of fork progression apparent in the microarray analysis (Fig. 4) is a secondary consequence of the lack of supercoiling, perhaps because of impaired transcription or an inability to repair stalled forks.
We conclude that topo IV can act as a swivel for replication by relaxing (+) supercoils ahead of the fork. First, we showed this activity of topo IV is great enough to completely prevent the appearance of (+) supercoiled DNA in vivo, although it does not remove the (−) supercoils in the plasmid (Fig. 5). Second, the instant stop of DNA replication when norfloxacin targets gyrase is characteristic of a protein being converted to a physical barrier in front of the replication fork. In gyrase-deficient strains that are partially rescued by a topo I disruption, topo IV targeting by norfloxacin causes an instant replication stop (31), suggestive of topo IV action in front of the fork. Third, we discovered (N. Crisona, T. Strick, D. Bensimon, V. Croquette, and N.R.C., unpublished results) that the rate of unlinking of (+) supercoiled DNA by topo IV in vitro is 20 times higher than unlinking of (−) supercoiled DNA. This differential activity is essential for a prokaryotic swivel, as relaxation of (−) supercoils increases linking number and would negate any role in fork progression if equal to (+) supercoil relaxation. It is possible that in vivo topo IV also removes (+) precatenanes behind the replication fork (19, 32), but this has not yet been demonstrated.
In this work, we present another application of DNA microarray technology. Previously, microarrays were used to study steady-state levels of RNA or DNA (33, 34). We demonstrated that the technique is sensitive enough to study a dynamic process like DNA replication at high resolution. We traced the progression of the replication fork along the bacterial chromosome by using the relative abundance of replicated DNA over unreplicated DNA. Although this was at most 2-fold, we were able to position the fork boundary accurately. The high resolution was evidenced in two ways. Only 0.1–0.5% of the ORFs in the region of the chromosome that we defined as unreplicated had a “red” to “green” ratio higher than 1 (Fig. 3). Second, at high novobiocin concentration, the fork boundary mapped to the same ORF in all samples in the time course (Fig. 3D).
The abrupt termination of replication at high novobiocin concentrations is interesting. In the absence of topoisomerases, replication with purified enzymes of oriC- and ColEI-driven plasmids proceeds until substantial (+) supercoiling (σ = +0.048) is induced (35). In mutants of Saccharomyces cerevisiae, DNA replication goes on for several thousand base pairs in the absence of topoisomerase activity and then halts, unable to overcome the (+) torsional stress generated by replication (36). In both cases, about 10% of the parental DNA was replicated in the absence of topoisomerase. If the novobiocin-induced replication arrest likewise occurs when the supercoil density in the topological domain containing the replication fork reaches +0.048, then the rapidity of the arrest puts an upper bound on the size of this domain. Bacterial chromosome replication should cease without topoisomerase activity after 5 or 150 seconds, depending, respectively, on whether replication occurs in a 100-kb isolated supercoiled domain (37) or in a single chromosome-wide domain. We observed complete replication arrest in the first time points at less than 1 min after drug addition. This suggests that torsional stress from replication is distributed not throughout the chromosome but in a smaller topologically closed domain.
Supplementary Material
Acknowledgments
We thank C. Richmond, Y. Wei, and F .R. Blattner for their help in preparing a library of E. coli ORFs. We also thank W. Suh, C. Gross, D. Zimmer, and S. Kustu for help in preparing PCR products and microarrays. A.K. is a Program for Mathematics and Molecular Biology (PMMB) National Science Foundation Postdoctoral Fellow (National Science Foundation Grant, DMS-9406348), and his graduate work in N.R.C.'s laboratory included in the current publication was also supported by PMMB. B.P. was supported by National Institutes of Health (NIH) Molecular Biophysics Training Grant, GM08295–11. This work was supported by NIH grants to P.O.B., N.R.C., and D.B. and by the Howard Hughes Medical Institute (to P.O.B.). P.O.B. is an associate investigator at the Howard Hughes Medical Institute.
Abbreviation
- topo IV
topoisomerase IV
References
- 1.Wang J C. Annu Rev Biochem. 1996;65:635–692. doi: 10.1146/annurev.bi.65.070196.003223. [DOI] [PubMed] [Google Scholar]
- 2.Cairns J. J Mol Biol. 1963;6:208–213. doi: 10.1016/s0022-2836(63)80070-4. [DOI] [PubMed] [Google Scholar]
- 3.Drlica K, Zhao Z. Microbiol Mol Biol Rev. 1997;61:377–392. doi: 10.1128/mmbr.61.3.377-392.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kornberg A, Baker T A. DNA Replication. New York: Freeman; 1992. pp. 387–392. [Google Scholar]
- 5.Filutowicz M, Jonczyk P. Mol Gen Genet. 1983;191:282–287. doi: 10.1007/BF00334827. [DOI] [PubMed] [Google Scholar]
- 6.Khodursky A B, Zechiedrich E L, Cozzarelli N R. Proc Natl Acad Sci USA. 1995;92:11801–11805. doi: 10.1073/pnas.92.25.11801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Belland R J, Morrison S G, Ison C, Huang W M. Mol Microbiol. 1994;14:371–380. doi: 10.1111/j.1365-2958.1994.tb01297.x. [DOI] [PubMed] [Google Scholar]
- 8.Maniatis T, Fritsch E F, Sambrook J. Molecular Cloning: A Laboratory Manual. Vol. 1. Plainview, NY: Cold Spring Harbor Lab. Press; 1982. p. 1.12. [Google Scholar]
- 9.Crisona N J, Kanaar R, Gonzalez T N, Zechiedrich E L, Klippel A, Cozzarelli N R. J Mol Biol. 1994;243:437–457. doi: 10.1006/jmbi.1994.1671. [DOI] [PubMed] [Google Scholar]
- 10.Blattner F R, Plunkett G, Bloch C A, Perna N T, Burland V, Riley M, Collado-Vides J, Glasner J D, Rode C K, Mayhew G F, et al. Science. 1997;277:1453–1462. doi: 10.1126/science.277.5331.1453. [DOI] [PubMed] [Google Scholar]
- 11.Sundin O, Varshavsky A. Cell. 1980;21:103–114. doi: 10.1016/0092-8674(80)90118-x. [DOI] [PubMed] [Google Scholar]
- 12.Kampranis S, Gormley N, Tranter R, Orphanides G, Maxwell A. Biochemistry. 1999;38:1967–1976. doi: 10.1021/bi982320p. [DOI] [PubMed] [Google Scholar]
- 13.Nikaido N, Thanassi D G. Antimicrob Agents Chemother. 1993;37:1393–1399. doi: 10.1128/aac.37.7.1393. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Kato J-i, Nishimura Y, Imamura R, Niki H, Hiraga S, Susuki H. Cell. 1990;63:393–404. doi: 10.1016/0092-8674(90)90172-b. [DOI] [PubMed] [Google Scholar]
- 15.Schmid M B. J Bacteriol. 1990;172:5416–5424. doi: 10.1128/jb.172.9.5416-5424.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Schena M, Shalon D, Davis R W, Brown P O. Science. 1995;270:467–470. doi: 10.1126/science.270.5235.467. [DOI] [PubMed] [Google Scholar]
- 17.Carl P L. Mol Gen Genet. 1970;109:107–122. doi: 10.1007/BF00269647. [DOI] [PubMed] [Google Scholar]
- 18.Ullsperger C J, Vologodskii A A, Cozzarelli N R. In: Nucleic Acids and Molecular Biology. Lilley D M J, Eckstein F, editors. Vol. 9. Berlin: Springer; 1995. pp. 115–142. [Google Scholar]
- 19.Postow L, Peter B J, Cozzarelli N R. BioEssays. 1999;21:805–808. doi: 10.1002/(SICI)1521-1878(199910)21:10<805::AID-BIES1>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 20.Wu H-Y, Shyy S, Wang J C, Liu L F. Cell. 1988;53:433–440. doi: 10.1016/0092-8674(88)90163-8. [DOI] [PubMed] [Google Scholar]
- 21.Liu L F, Wang J C. Proc Natl Acad Sci USA. 1987;84:7024–7027. doi: 10.1073/pnas.84.20.7024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lockshon D, Morris D R. Nucleic Acids Res. 1983;11:2999–3017. doi: 10.1093/nar/11.10.2999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rybenkov V V, Vologodskii A V, Cozzarelli N R. J Mol Biol. 1997;267:312–323. doi: 10.1006/jmbi.1996.0877. [DOI] [PubMed] [Google Scholar]
- 24.Drlica K. Microbiol Rev. 1984;48:273–289. doi: 10.1128/mr.48.4.273-289.1984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Orr E, Fairweather N F, Holland I B, Pritchard R H. Mol Gen Genet. 1979;177:103–112. doi: 10.1007/BF00267259. [DOI] [PubMed] [Google Scholar]
- 26.Filutowicz M. Mol Gen Genet. 1980;177:301–309. doi: 10.1007/BF00267443. [DOI] [PubMed] [Google Scholar]
- 27.Mirkin S, Shmerling Z. Mol Gen Genet. 1982;188:91–95. doi: 10.1007/BF00332999. [DOI] [PubMed] [Google Scholar]
- 28.Staczek P, Higgins N P. Mol Microbiol. 1998;29:1435–1448. doi: 10.1046/j.1365-2958.1998.01025.x. [DOI] [PubMed] [Google Scholar]
- 29.Gari E, Figueroa-Bossi N, Blanc-Potard A B, Spirito F, Schmid M B, Bossi L. Mol Microbiol. 1996;21:111–122. doi: 10.1046/j.1365-2958.1996.6221338.x. [DOI] [PubMed] [Google Scholar]
- 30.Kreuzer K N, Cozzarelli N R. J Bacteriol. 1979;140:424–435. doi: 10.1128/jb.140.2.424-435.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Khodursky A B, Cozzarelli N R. J Biol Chem. 1998;273:27668–27677. doi: 10.1074/jbc.273.42.27668. [DOI] [PubMed] [Google Scholar]
- 32.Peter B J, Ullsperger C, Hiasa H, Marians K J, Cozzarelli N R. Cell. 1998;94:819–827. doi: 10.1016/s0092-8674(00)81740-7. [DOI] [PubMed] [Google Scholar]
- 33.DeRisi J, Iyer V, Brown P. Science. 1997;278:680–686. doi: 10.1126/science.278.5338.680. [DOI] [PubMed] [Google Scholar]
- 34.Pollack J, Perou C, Alizadeh A, Eisen M, Pergamenschikov A, Williams C, Jeffrey S, Botstein D, Brown P. Nat Genet. 1999;23:41–46. doi: 10.1038/12640. [DOI] [PubMed] [Google Scholar]
- 35.Hiasa H, Marians K J. J Biol Chem. 1994;269:16371–16375. [PubMed] [Google Scholar]
- 36.Kim R A, Wang J C. J Mol Biol. 1989;208:257–267. doi: 10.1016/0022-2836(89)90387-2. [DOI] [PubMed] [Google Scholar]
- 37.Pettijohn D E. Cell. 1982;30:667–669. doi: 10.1016/0092-8674(82)90269-0. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.