

Abstract
Drosophila melanogaster cuticular pheromones consist of unsaturated hydrocarbons with at least one double bond in position 7: 7 tricosene (T) in males and 7,11 heptacosadiene (HD) in females. However, in many African populations like the Tai strain, females possess low levels of 7,11 HD and high levels of its positional isomer 5,9 HD. We have previously isolated a desaturase gene, desat1, from the Canton-S strain (CS), a 7,11 HD-2-rich morph of D. melanogaster. This desaturase is located in 87C, a locus that has been involved in the difference between 7,11 HD and 5,9 HD morphs. Therefore, we have searched for different desaturase isoforms in both strains. We first cloned desat1 in the Tai strain and report here functional expression of desat1 in CS and Tai. In both strains, the Desat1 enzymes have the same Δ9 specificity and preferentially use palmitate as a substrate, leading to the synthesis of ω7 fatty acids. Also found was a desaturase sequence, named desat2, with a homologous catalytic domain and a markedly different N-terminal domain compared with desat1. In CS genome, it lies 3.8 kb upstream of desat1 and is not transcribed in either sex. In the Tai strain, it is expressed only in females and acts preferentially on myristate, leading to the synthesis of ω5 fatty acids. We suggest, therefore, that desat2 might play a control role in the biosynthesis of 5,9 HD hydrocarbons in Tai females and could explain the dienic hydrocarbon polymorphism in D. melanogaster.
Keywords: acyl-Coa desaturase, pheromone biosynthesis
Unsaturated fatty acids are structural components of membrane lipids. In animals, fatty acids are desaturated by a membrane-bound enzyme complex involving cytochrome b5, cytochrome b5 reductase, and a desaturase (1–3). Desaturases catalyze the introduction of a double bond into the hydrocarbon chain of a fatty acyl-CoA, at a position determined by the enzyme specificity. A low number of animal desaturases have been both molecularly and functionally characterized by heterologous expression in yeast or in Arabidopsis: Δ9 and Δ6 desaturases from rat (4, 5), Δ6 desaturase from mouse (6), ω3, Δ6, and Δ5 desaturases from Caenorhabditis elegans (7–9), and Δ11 and Δ9 desaturases from the moth Trichoplusia ni (10, 11).
In the Drosophila melanogaster subgroup, flies use double bonds in cuticular hydrocarbons among other structural parameters to signal sex or species (12, 13). Males have high levels of monoenes, whereas females are rich in dienes, for example 7-tricosene (7T) and 7,11 heptacosadiene (7,11 HD) in the Canton-S strain (CS) (14).
Studies in D. melanogaster strongly suggest that the biosynthesis of male monoenes and female dienes follows the same elongation-decarboxylation mechanism characterized in Musca domestica (15, 16) and shares early steps up to vaccenic acid, a common precursor (refs. 17 and 18; Fig. 1). The involvement of a desaturase with a Δ9 specificity to introduce the common double bond has also been hypothesized (18). We have previously isolated a CS D. melanogaster desaturase gene, desat1, expressed in both males and females, and localized in 87C (19). We report here the functional characterization of the CS desat1 gene by heterologous expression in yeast and show that it does encode a Δ9 desaturase acting preferentially on palmitate, thus leading to ω7 fatty acids. (The location of a double bond in a fatty acid is described by Δ and ω positions: the position corresponds to the double bond position relative to the carboxyl end of fatty acids and the ω position relative to the methyl end.)
Figure 1.
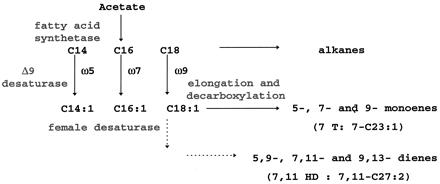
Summary of biosynthetic pathways in D. melanogaster for producing hydrocarbons. The specificity of the female-specific desaturase is not known (arrows in dotted lines).
In D. melanogaster, there is a marked geographical polymorphism with female dienes. For example, females of populations from sub-Saharan Africa, which includes the Tai strain, possess about 1/10 the amount of 7,11 HD relative to females from the rest of the world but exhibit much larger amounts of its positional isomer, 5,9 HD (20, 21). This double bond position polymorphism is sex specific in the African strains, and the hydrocarbon difference has been mapped to a single locus at 87C,D, which is close to that of the desat gene (22). In the biosynthesis of polyunsaturated fatty acids of many animals, the double bond closer to the methyl end is often introduced first (3). Thus it seemed reasonable to search for isoforms of the enzyme introducing the first double bond; for example 5,9 HD-rich Tai flies might have a Δ9 desaturase isoform able to produce ω5 hydrocarbons rather than ω7 as in 7,11 HD-rich CS flies. We have first cloned the desat1 ORF from the Tai strain and found that it is expressed in both sexes and encodes a Δ9 desaturase with the same specificity and preference for palmitate as in the CS strain. We then screened a CS genomic library with desat1 and found a second putative desaturase gene, named desat2, located 3.7 kb upstream of the desat1 gene. In contrast to desat1, which is expressed in both males and females of CS and Tai strains, desat2 is expressed in Tai females and not in males. When expressed in yeast, desat2 encodes also a Δ9 desaturase, but with a different substrate specificity. This desaturase preferentially acts on myristate, therefore leading to the synthesis of ω5 hydrocarbons.
Materials and Methods
Drosophila Stocks and Strains.
Two strains of D. melanogaster were used: CS, an old standard laboratory strain from Ohio, characterized by a high 7,11 HD phenotype, and Tai, a stock composed of a mixture of isofemale lines collected in Tai, Ivory Coast, in 1980, and characterized by a high 5,9 HD phenotype (20).
Flies were reared at 25°C on standard yeast/cornmeal/agar medium with a 12:12-h light/dark cycle. For RNA extraction, flies were sexed within 1 hr after emergence and were kept separately in rearing bottles between 0 and 5 days.
CS Genomic–DNA Library Screen.
A partial desat1 cDNA containing the beginning of the cDNA and the complete desat1 ORF was labeled by random hexamer priming by using [−32 P]dATP (Stratagene). The probe was used to screen randomly sheared Drosophila CS DNA cloned into Charon-4 A (23). Plaque lifts on nitrocellulose (Schleicher & Schuell) were hybridized overnight in hybridization buffer containing 5 × SSC, 5 × Denhardt's solution (0.02% polyvinylpyrrolidone/0.02% Ficoll/0.02% BSA), 50% formamide, and 0.5% SDS, at 42°C. The filters were washed twice at room temperature for 10 min in 2 × SSC/0.1% SDS buffer. Positive clones were isolated. EcoRI fragments were subcloned into Bluescript SK and partially sequenced by the dideoxy chain termination method (24).
RNA Extraction and Reverse Transcription–PCR (RT-PCR) Analysis.
Total RNA was isolated from CS or Tai Drosophila by the lithium chloride/urea method. Poly(A+) RNA and cDNA were prepared as previously described (19). A putative desat2 gene ORF was determined by the webgene program (http://www.itba.mi.cnr.it/webgene/) from one EcoRI fragment, which resulted from the screen on CS genomic DNA.
Two pairs of specific oligodeoxynucleotides complementary to the 21-nt terminal sequences of desat1 or desat2 ORF were synthesized. The direct primer corresponded to the initiator ATG codon and the six following codons of desat1 or desat2 ORF, and the reverse primer to the seven last codons of desat1 or desat2 ORF including the stop codon. BamHI and SacI restriction sites were added at the 5′ end of the direct and reverse primers, respectively. These primers were designed as follows: (ds1 was an abbreviation of desat1 and ds2 of desat2) ds1-dir 5′-GGGGGATCCATGCCGCCCAACGCCCAAGCC-3′, ds1-rev 5′-GGGGAGCTCCTATTCGCTTTTTTTGTG(T/A)GT-3′, ds2-dir 5′-GGGGGATCCATGGCGCCCTACTCAAGGATA-3′ and ds2-rev 5′-GGGGAGCTCTCATCTACTTTTATCCACAAG-3′. The locations of these primers are described in Fig. 4.
Figure 4.
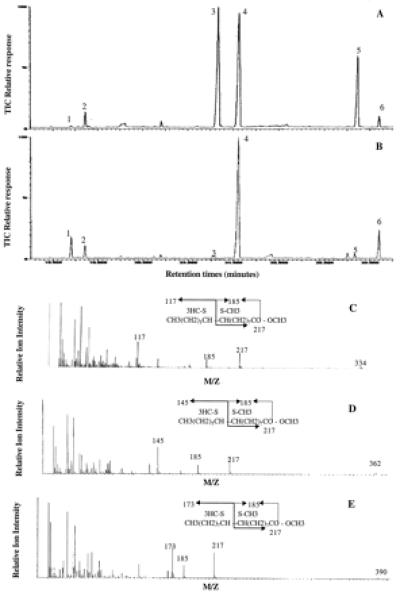
Identification of monounsaturated fatty acid resulting from the Tai desat1 and desat2 gene expression in yeast fatty acid methyl esters of yeast transformed with Yeptaidesat1 (A) and Yeptaidesat2 (B) were analyzed by GC/MS. The common peaks were identified as Δ9 C14:1 (peak 1), C14:0 (peak 2), Δ9 C16:1 (peak 3), C16:0 (peak 4), Δ9 C18:1 (peak 5), and C18:0 (peak 6). Mass spectrum of the DMDS adducts of 0.9 monounsaturated fatty acid methyl esters were represented: Δ9–14 Me:DMDS (C), Δ9–16 Me:DMDS (D), and Δ9–18 Me:DMDS (E). These mass spectra exhibit the characteristic Δ9 unsaturated fragments ions at m/z 217 and 185.
cDNA (500 ng) was amplified in a Taq polymerase (Appligene, Strasbourg, France) buffer supplemented to a final concentration of 1.5 mM MgCl2/0.2 mM dNTPs/0.2 μM of each primer in a final 50-μl volume. The mixtures were amplified in a thermal cycler (MJ Research, Cambridge, MA) for 40 cycles (92°C, 30 s; 55°C, 1 min; 72°C, 1 min) preceded by a 2-min denaturation step at 92°C and followed by a 5-min elongation step at 72°C.
Molecular Cloning.
The resulting CS or Tai desat1 and desat2 PCR fragments were digested by the BamHI and SacI restriction enzymes and were subcloned in a Bluescript SK−, which was also cut by BamHI and SacI. Several clones were sequenced to determine the putative mutations introduced by Taq polymerase during RT-PCR. A clone without mutations resulting from each RT-PCR amplification was then restricted by BamHI and SacI and ligated to the digested BamHI-SacI derivative vector of YEP352/OLE4.8, which was used for the functional assay of the rat Δ9 desaturase (4) and the T.ni Δ9 and Δ11 desaturases (10, 11). The plasmid constructions contain chimeric ORFs that start with the 27 first codons of the yeast Δ9 desaturase ORF followed by an in-frame four-codon linker and a complete Drosophila desaturase ORF. Each chimeric ORF is flanked by the promoter and terminator sequences of the OLE1 gene of yeast Δ9 desaturase. The resulting constructions were named YEPcsdesat1, YEPtaidesat1, and YEPtaidesat2.
Yeast Transformation and Growth.
The YEPcsdesat1, YEPtaidesat1, and YEPtaidesat2 constructs were transformed in the Δ9 desaturase deficient Saccharomyces cerevisiae strain L8-14C [Mat α, ole1Δ∷LEU2, leu2–2, leu2–112, trp1–1, ura3–52, his4] (4), by using a standard method (alkali-cation yeast transformation kit, Bio 101, Vista, CA). Transformed yeasts were plated onto complete synthetic dextrose medium containing 1% tergitol, 0.5 mM oleic acid, and 0.5 mM palmitoleic acid and lacking uracil. To test genetic complementation of the mutant yeast strain, transformed yeasts were plated onto complete yeast extract/peptone/dextrose medium lacking unsaturated fatty acids.
Fatty Acid Analysis.
Transformants were grown to ≈1.107 cells/ml at 30°C in yeast extract/peptone/dextrose medium (1% bacto-yeast extract/2% bacto-peptone/2% glucose), washed with water, and extracted with chloroform/methanol (2:1). Yeast total fatty acids were analyzed by GC/MS of methyl esters by using a nonpolar BP1 column (Scientific Glass Engineering, Ringwood, Victoria, Australia), and the double bond positions of the monounsaturated methyl esters were determined by MS of their dimethyldisulfide adducts (25, 26).
Results
Cloning of desat1 in Tai Strain and Heterologous Expression of CS and Tai desat1 in Yeast.
desat1 primers were designed from the CS sequence to amplify the complete Tai desaturase ORF by RT-PCR. Amplifications were performed on either male or female Tai DNA, and all produced a single band of 1.1 kb as in CS strain (Fig. 2). Tai desat1 amplification products were subcloned in Bluescript SK, and several clones corresponding to each amplification were sequenced. Only one ORF was obtained, which encodes a polypeptide of 383 amino acids (Fig. 3). At the nucleic acid level, Tai desat1 ORF sequence shows only four substitutions leading to three amino acid replacements in comparison with the CS desat1 ORF sequence (19). These amino acid changes occurred as follows: C to S in position 44, A to V in position 77, and L to F in position 81.
Figure 2.
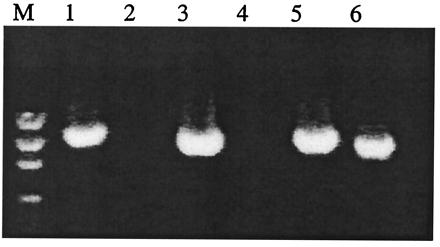
Products of RT-PCR reactions from mixed male-female CS RNA (lanes 1 and 2) and male or female Tai RNA (males, lanes 3 and 4; females, lanes 5, and 6). desat1 primers (lanes 1, 3 and 5) or desat2 primers (lanes 2, 4, and 6) were used. Molecular weight standards (M) were included.
Figure 3.

The two amino acid sequences of D. melanogaster desaturases (Tai strain). Sequences show 65.0% identity. The three histidine domains are boxed, and the putative transmembrane domains are indicated by underlines. * shows the amino acid modifications between CS and Tai Desat sequences. Identical and homologous amino acids are indicated with : and ., respectively. desat1 and desat2 cDNA sequences from Tai have been introduced in European Molecular Biology Laboratory (accession nos.: AJ245747 and AJ271415).
To study the function of the CS and Tai desat1 genes, both ORFs were first subcloned in the expression derivative vector of YEP352/OLE4.8. Resulting plasmids were used to transform a Δ9 desaturase-deficient yeast strain. Yeasts transformed with either plasmid were able to grow on a medium lacking unsaturated fatty acids, indicating that both desaturases were functional in yeast, produced monounsaturated fatty acids, and could complement the Δ9 desaturase mutation of the mutant yeast.
The fatty acid compositions of the transformants as determined by the GC/MS analysis of fatty acid methyl esters are shown in Fig. 3. The CS and Tai desat1 desaturases have the same specificity, with predominantly C16:1 and C18:1 being produced by both transformants in a ratio higher than 2–3:1. Data are shown only for Tai desat1, but similar ones were obtained for CS (Fig. 4A). The Δ9 position of the double bond in all monounsaturated fatty acids was determined by the dimethyl disulfide derivatization and the analysis of the resulting products (Fig. 4 D and E). Δ9 C14:1 fatty acids were also observed but in very low amounts (less than 1% of the total fatty acid content).
Genomic Organization of the desat Locus in CS.
To characterize further the desaturase locus, a CS genomic library was hybridized at high stringency with a CS desat1 cDNA. One positive phage was obtained, and its insert liberated two fragments with similar sizes around 9 kb when cut with EcoRI. Both fragments were subcloned in a Bluescript SK−. Both gave a hybridization signal with the desat1 probe but differed in their HindIII restriction profile because one fragment had no HindIII site. The second EcoRI fragment produced three fragments when cut with HindIII: two with a positive hybridization signal, one 4.5-kb HindIII subfragment, and one 1.3-kb EcoRI-HindIII subfragment (Fig. 5). These subfragments were sequenced.
Figure 5.
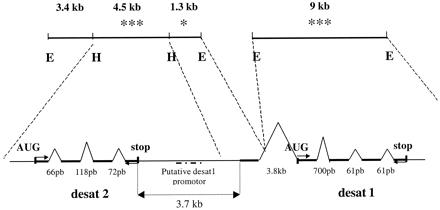
Genomic organization of the desat locus in CS strain. Two hybridization-positive EcoRI fragments were obtained after hybridization of a CS genomic library with desat1 probe. The number of * show the intensity of the hybridization signal with the desat1 probe. EcoRI and HindIII restriction sites are noted E and H, respectively. Putative desat2 ORF and desat1 complete cDNA are in bold. The intron sizes of desat1 and putative desat2 genes are noted under their representations. Primers used for RT-PCR are indicated by small arrows. Dotted line indicates the position of the putative desat1 promoter.
The 4.5-kb HindIII subfragment sequence revealed a region showing homology with desat1 and contained another putative desaturase sequence that was named desat2. The putative ORF of desat2 was identified with the webgene program, which suggested that this gene would be functional, if expressed, and would have three introns of 66 pb, 118 pb, and 72 pb. However, no promoter could be found.
The 1.3-kb hybridization-positive EcoRI-HindIII sequence contained the 140 first nucleotides of the CS desat1 cDNA followed by the beginning of an intron (155 nucleotides) and then an EcoRI site (Fig. 5).
The other 9-kb EcoRI fragment began with the end of an intron (3.65 kb) and was followed by the complete CS desat1 sequence (Fig. 5). Therefore, desat1 has a 3.8-kb-long intron just before the first coding AUG. Three other introns have been described already in the CS desat1 gene, with 700 pb, 61 pb, and 61 pb, respectively (19).
The desat2 stop codon was separated from the first nucleotide of the CS desat1 cDNA by 3.7 kb. A putative promoter of desat1 gene was localized 2.5 kb upstream of the beginning of desat1 cDNA in the 4.5-kb HindIII subfragment.
The genomic organization of the desat locus was confirmed by the genomic sequences from the isogenic strain y2, cn bw sp resulting from the Drosophila sequencing project (accession no. AC 17132). The desat1 and putative desat2 genes were present in these sequences but have not yet been molecularly characterized. According to the databases, the desat locus is localized in region 87B-C on polytene chromosomes.
The desat2 Gene Is Expressed Only in Adult Tai Females.
To determine whether this new potential gene desat2 could be transcribed in the two D. melanogaster strains, RT-PCR was performed with desat2 primers with either mixed male–female CS cDNA and Tai cDNA from either males and females (Fig. 2).
An amplification was obtained only with Tai female cDNA and yielded a single amplification fragment. The female Tai desat2 ORF is shorter than the Tai desat1 ORF: 1.08 kb instead of 1.15 kb. It encodes a 361-aa protein (instead of 383). The alignment of Desat1 and Desat2 reveals 65% identity and 81% similarity between the two proteins. The Tai desat2 protein shows a markedly modified N-terminal end including a deletion of 21 amino acids in its N-terminal region in comparison to the desat1 protein (Fig. 3). But the Tai desat2 ORF shows only one modification compared with CS desat2 ORF: a serine instead of a glycine in position 30.
The functional expression of Tai desat2 was performed as described for desat1. The desat2 ORF was subcloned in the expression derivative vector of YEP352/OLE4.8, and the resulting plasmid was used to transform an ole Δ9 desaturase-deficient yeast strain. The yeast desaturase deficiency was again complemented. The fatty acid produced by the cultures transformed with desat2 was analyzed by GC/MS (Fig. 4B). A Δ9 specificity was characterized, but the major unsaturated acid produced had 14 carbons, and only trace amounts of Δ9 C16:1 and Δ9 C18:1 fatty acids were found. Thus desat2 encodes a Δ9 desaturase preferring myristic acid as a substrate.
Discussion
Research has been conducted to determine why Tai female flies have large quantities of the unusual 5,9-heptacosadiene on their cuticle, rather than the more common 7,11-heptacosadiene found in CS female flies. Characterization of two desaturase genes, desat1 and desat2, has provided an answer because of the specific expression and unusual unsaturated product of the latter gene.
The desaturase 1 ORFs are similar in both Tai and CS strains of the D. melanogaster species, with only four nucleotide substitutions resulting in three amino acid modifications in the N-terminal region. These amino acid replacements in the Tai protein do not result in any marked functional change. In both strains, the encoded enzymes have a Δ9 specificity and the same preference for palmitic acid as a substrate. Thus Desat1 can produce the unsaturated acid precursors for ω7 hydrocarbons in flies of the CS and Tai strains.
The other desaturase gene, desat2, is expressed only in adult females of the Tai strain. The encoded peptide sequences of desat1 and desat2 show 65% identity. Desat2 shows the same putative transmembrane regions as Desat1 but is smaller (361 amino acids instead of 383) because of a shorter N-terminal region. The specificity of Desat2 is different from that of Desat1, as it produces myristoleic acid (Δ9 C14:1) when expressed in the same desaturase-deficient yeast and very little palmitoleic and oleic acids. In comparison, Desat1 preferentially produces palmitoleic acid (Δ9 C16:1) and the cabbage looper moth, Trichoplusia ni, Δ9 desaturase preferentially produces oleic acid (Δ9 C18:1) (10). These three desaturases share the same Δ9 regio-specificity but differ in their substrate specificities. If one compares Desat1 with Desat2 in the cytosolic region localized between the two transmembrane domains, all amino acids but one are identical or have conservative substitutions. This region contains two of three conserved histidine motifs shown to be essential for catalysis (27). It is interesting to note that the region between the second transmembrane domain and the third histidine motif (at 298–302 in Desat1) differs significantly between the two sequences, and we suggest that the specificity for substrate chain length could be determined by this region.
In the Tai, strain desat2 is expressed but in a sex-specific way. This suggests that this gene might be regulated by the female sex determination cascade (28, 29). Finally, the correlation between the expression of desat2 and the abundance of 5,9 dienes, which is null in both sexes of the CS strain but high in females of the Tai strain, strongly suggests an involvement of desat2 in the biosynthesis of these dienes. The desat2 gene seems able to explain much of the 7,11 HD/5,9 HD polymorphism. Moreover, it is located in the same chromosomal region where female polymorphism has actually been mapped (22).
The presence of desat2 in the genomic sequence of CS strain, where it is not expressed, is intriguing. The CS desat2 sequence is very similar to the Tai one. However, the presence of a defective promoter in CS strain and a functional one only in Tai females might explain why desat2 is transcribed only in Tai females.
Acknowledgments
We are grateful to Patricia Marsella-Herrick, Dr. Claire-Lise Rosenfield, Professor Michel Jacquet, and Dr. Georges Renault for technical assistance with the functional assay and to Maialene Letbatz for help in sequencing. Financial support from Centre National de la Recherche Scientifique (CNRS) and Ministère de l'Education Nationale et de la Recherche (M.E.N.R.) is acknowledged. This work was also supported by grants to D.C.K. from the National Science Foundation/United States Environmental Protection Agency (BES-9728367) and the United States Department of Agriculture (97–35302-4345), and to W.L.R. from the National Science Foundation (IBN-9870669). Financial support from CNRS and M.E.N.R. is also acknowledged.
Abbreviations
- T
tricosene
- HD
heptacosadiene
- CS
Canton-S
- RT-PCR
reverse transcription–PCR
Footnotes
Data deposition: The sequences reported in this paper have been deposited in the GenBank database [accession nos. desat1 from Canton-S (U73160), desat1 from Tai (AJ245747), and desat2 from Tai (AJ271415)].
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.150243997.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.150243997
References
- 1.Holloway P W. Biochemistry. 1971;10:1556–1560. doi: 10.1021/bi00785a008. [DOI] [PubMed] [Google Scholar]
- 2.Spatz L, Strittmatter P. J Biol Chem. 1971;248:793–799. [PubMed] [Google Scholar]
- 3.Jeffcoat R. Eur J Biochem. 1979;101:447–453. doi: 10.1111/j.1432-1033.1979.tb19738.x. [DOI] [PubMed] [Google Scholar]
- 4.Stukey J E, McDonough V M, Martin C E. J Biol Chem. 1990;265:20144–20149. [PubMed] [Google Scholar]
- 5.Aki T, Shimada Y, Inagaki K, Higashimoto H, Kawamoto S, Shigeta S, Ono K, Suzuki O. Biochem Biophys Res Commun. 1999;255:575–579. doi: 10.1006/bbrc.1999.0235. [DOI] [PubMed] [Google Scholar]
- 6.Cho H, Nakamura M T, Clarke S D. J Biol Chem. 1999;274:471–477. doi: 10.1074/jbc.274.1.471. [DOI] [PubMed] [Google Scholar]
- 7.Spychalla J P, Kinney A J, Browse J. Proc Natl Acad Sci USA. 1997;94:1142–1147. doi: 10.1073/pnas.94.4.1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Napier J A, Hey S J, Lacey D J, Shewry P R. Biochem J. 1998;330:611–614. doi: 10.1042/bj3300611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Watts J L, Browse J. Arch Biochem Biophys. 1999;362:175–182. doi: 10.1006/abbi.1998.1024. [DOI] [PubMed] [Google Scholar]
- 10.Knipple D C, Rosenfield C L, Miller S J, Liu W, Tang J, Ma P W K, Roelofs W L. Proc Natl Acad Sci USA. 1998;95:15287–15292. doi: 10.1073/pnas.95.26.15287. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Liu W, Ma P W K, Marsella-Herrick P, Rosenfield C L, Knipple D C, Roelofs W. Insect Biochem Mol Biol. 1999;29:435–443. doi: 10.1016/s0965-1748(99)00020-x. [DOI] [PubMed] [Google Scholar]
- 12.Jallon J M. Behav Genet. 1984;14:441–478. doi: 10.1007/BF01065444. [DOI] [PubMed] [Google Scholar]
- 13.Coyne J A, Oyama R. Proc Natl Acad Sci USA. 1995;92:9505–9509. doi: 10.1073/pnas.92.21.9505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jallon J M, David J R. Evolution (Lawrence, KS) 1987;4:294–302. [Google Scholar]
- 15.Reed J R, Vanderwel D, Choi S, Pomonis J G, Reitz R C, Blomquist G J. Proc Natl Acad Sci USA. 1994;91:10000–10004. doi: 10.1073/pnas.91.21.10000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Reed J R, Quilici D R, Blomquist G J, Reitz R C. Biochemistry. 1995;34:16221–16227. doi: 10.1021/bi00049a038. [DOI] [PubMed] [Google Scholar]
- 17.Chan Yong T P, Jallon J M. C R Hebd Séances Acad Sci. 1986;303:197–202. [Google Scholar]
- 18.Pennanec'h M, Bricard L, Kunesh G, Jallon J M. J Insect Physiol. 1997;43:1111–1116. doi: 10.1016/s0022-1910(97)00082-6. [DOI] [PubMed] [Google Scholar]
- 19.Wicker-Thomas C, Henriet C, Dallerac R. Insect Biochem Mol Biol. 1997;27:963–972. doi: 10.1016/s0965-1748(97)00077-5. [DOI] [PubMed] [Google Scholar]
- 20.Jallon J M, Péchiné J M. C R Acad Sci. 1989;309:1551–1556. [Google Scholar]
- 21.Ferveur J F, Cobb M, Boukella H, Jallon J M. Genetica. 1996;97:73–80. doi: 10.1007/BF00132583. [DOI] [PubMed] [Google Scholar]
- 22.Coyne J A, Wicker-Thomas C, Jallon J M. Genet Res. 1999;73:189–203. doi: 10.1017/s0016672398003723. [DOI] [PubMed] [Google Scholar]
- 23.Maniatis T, Hardison R C, Lacy E, Lauer J, O'Connell C, Quon D, Sim G K, Efstratiadis A. Cell. 1978;15:687–701. doi: 10.1016/0092-8674(78)90036-3. [DOI] [PubMed] [Google Scholar]
- 24.Sanger F, Nickler S, Coulson A R. Proc Natl Acad Sci USA. 1977;74:5463–5467. doi: 10.1073/pnas.74.12.5463. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Buser H R, Arn H, Guerin P, Rauscher S. Anal Chem. 1983;55:818–822. [Google Scholar]
- 26.Dunkelblum E, Tan S H, Silk P J. J Chem Ecol. 1985;11:265–277. doi: 10.1007/BF01411414. [DOI] [PubMed] [Google Scholar]
- 27.Shanklin J, Whittle E, Fox B G. Biochemistry. 1994;33:12787–12794. doi: 10.1021/bi00209a009. [DOI] [PubMed] [Google Scholar]
- 28.Jallon J M, Laugé G, Orssaud L, Antony C. Genet Res Camb. 1988;51:17–22. [Google Scholar]
- 29.Arthur B I, Jr, Jallon J M, Caflisch B, Choffat Y, Nothiger R. Curr Biol. 1998;8:1189–1190. doi: 10.1016/s0960-9822(07)00491-5. [DOI] [PubMed] [Google Scholar]