

Abstract
AvrXa7 is a member of the avrBs3 avirulence gene family, which encodes proteins targeted to plant cells by a type III secretion apparatus. AvrXa7, the product of avrXa7, is also a virulence factor in strain PXO86 of Xanthomonas oryzae pv. oryzae. Avirulence and virulence specificities are associated with the central repeat domain, which, in avrXa7, consists of 25.5 direct repeat units. Mutations in three C-terminal nuclear localization signal motifs eliminated avirulence and virulence activities in rice and severely reduced nuclear localization in a yeast assay system. Both pathogenicity functions and nuclear localization were restored on the addition of the sequence for the nuclear localization signal motif from SV40 T-antigen. The loss of avirulence activity because of mutations in the acidic transcriptional activation domain was restored by addition of the activation domain from the herpes simplex viral protein VP16. The activation domain was also required for virulence activity. However, the VP16 domain could not substitute for the endogenous domain in virulence assays. In gel shift assays, AvrXa7 bound double-stranded DNA with a preference for dA/dT rich sequences. The results indicate that products of the avrBs3-related genes are virulence factors targeted to host cell nuclei and have the potential to interact with the host DNA and transcriptional machinery as part of their mode of action. The results also suggest that the host defensive recognition mechanisms are targeted to the virulence factor site of action.
Keywords: avirulence, rice, disease
Two avirulence genes have been cloned from Xanthomonas oryzae pv. oryzae, the causal agent of bacterial blight of rice. Bacteria containing avrXa7 or avrXa10 elicit resistance on rice cultivars containing the resistance genes Xa7 and Xa10, respectively (1). Both genes are members of the avrBs3 gene family, which are found in a variety of species of Xanthomonas. The members of the family are nearly identical but differ in the number and apparent nature of a series of near-identical 102-bp direct repeats in the central portion of the coding sequence (reviewed ref. 2). The activities of avrBs3-related genes depend on a type III secretory pathway known as the hypersensitive reaction and pathogenicity or Hrp pathway (3, 4). Like a variety of type III-dependent proteins, the products of the avrBs3 family are secreted from the bacteria and translocated into the host cells (reviewed in ref. 5). Avirulence genes were named for the elicitation of disease resistance by the bacteria harboring the genes on inoculation to the appropriate cultivar of an otherwise susceptible plant host. However, the function of many avirulence genes from the perspective of the bacterium is their role in the virulence. In fact, many bacterial avirulence genes are dual-acting proteins (reviewed in ref. 6). These proteins elicit resistance in one context and are involved in the virulence of the bacterium on susceptible host plants. A variety of genes in the avrBs3 gene family are dual acting, including pthA from Xanthomonas citri (7), avrb6 and pthN from X. campestris pv. malvacearum (8–10), and avrXa7 (11, 12).
The biochemical functions of protein products of the avrBs3 family are unknown. Critical structural features involved in specificity for avirulence and virulence lie within the repeat domains (13–16). For example, replacement of the repeat coding region in avrXa10 with the corresponding region of avrXa7 converts avrXa10 to a gene with avirulence specificity for Xa7 and loses activity toward Xa10 (14). Exchange of the repeat domains of avrb6 and pthA demonstrated that specificity for virulence on the respective hosts was also controlled by the repeat domain (16). All members of the avrBs3 family also encode functional nuclear localization signal (NLS) motifs and acidic transcriptional activation domains (ADs) in the C-terminal coding regions (14, 17). Experimental evidence for avrBs3 and avrXa10 has indicated that the NLS and AD motifs are required for avirulence (14, 18, 19). The C-terminal features suggest that the proteins, in their capacity as virulence factors, may be targeted to the cell nucleus. However, no evidence has been reported indicating whether the NLS or AD was critical for the role of these proteins as virulence factors. We therefore examined the role of the various structural features as represented in avrXa7 on the virulence of X. oryzae pv. oryzae. Furthermore, the similarities of gene products to eukaryotic transcription factors also led us to examine the potential of AvrXa7 protein to bind DNA.
Materials and Methods
Strains, Plasmids, and DNA Manipulations.
Standard cultural methods and recombinant DNA techniques were used for Escherichia coli (20). X. oryzae pv. oryzae strain PXO99A, which lacks an endogenous copy of avrXa7 or avrXa10, was used for avirulence activity testing (1). Strain PXO86mx53, which was obtained by marker exchange of transposon mutant p29–29∷Tn5B20–53 (1), was used for virulence testing. The plasmid pZWavrXa10 contains the avrXa10 coding sequence fused to the lacZ promoter of pBluescriptKS+ (Stratagene) (14). The 3.2-kb SphI fragment of avrXa7 was derived from cosmid p29–29, which is a genomic clone from strain PXO86 (1). AD mutations in avrXa7 were prepared by replacing the SphI fragment of previously constructed mutants of avrXa10 with the SphI fragment of avrXa7 (14, 19). Codon numbering refers to corresponding codons in avrXa10. Methods for yeast manipulations were as described (21). Sequencing was performed at the Iowa State University and Kansas State University Sequencing Facilities.
Pathogenicity Tests.
Rice line IR24 is susceptible to PXO99 and PXO86 and was used for virulence assays. Rice lines IRBB10 and IRBB7 contain resistance genes Xa10 and Xa7 and were used for assaying avirulence activities of avrXa10 and avrXa7, respectively. Virulence assays were performed on 2-week-old rice plants by needleless syringe infiltration (14) and leaf-clip inoculation in growth chambers (22). Symptoms were scored by measuring lesion lengths after 11 days for the leaf-clip inoculations and noting the appearance of water soaking after 3 days for the syringe infiltrations. Lesion length measurements are averages of 25 leaves.
Nuclear Localization Assays.
The primers 5′-ACCTCTGCAGCAGAGTGGGCGTCACCGAATTCGAGC-3′ and 5′-TCAGAAGCTTCACTGAGGTTTATCATCGTCATCCTTGTAATCCAATAGCTCCATCAACCATGC-3′ were used to amplify the C-terminal coding region from pZW22 (avrXa10nls123−) (14). The second primer contains a FLAG epitope encoded immediately upstream of the stop codon. The PCR DNA was partially digested with EcoRI, completely with HindIII, and then ligated into pZWavrXa7 and pKSXa7(PH), resulting in pZWavrXa7M123 and pKSavrXa7M123, respectively. The plasmid pKSXa7(PH) was generated by ligating a partially digested PstI-HindIII fragment from pZWavrXa7 into pBluescriptKS+. Both pKSavrXa7 and pKSavrXa7M123 were cut with XhoI, partially digested with BamHI, and then ligated into vector pNIA (23) to generate pNIAXa7 and pNIAXa7 M123, respectively. Primers NLS1SV408 5′-CAGTAGATCTCCGACCAAAGAAGAAGCGCAAGGTCTCAGCTCAAACGCCGGATCAGGCGTC-3′ and NLS 5′-CAGTAGATCTCTCATGGCGCGTTAAGCGCCCACGCACTTCAGCTCAAACGCCGGATCAGG-3′, which contain NLS sequences of SV40 (RPKKKRKV) and native NLS3 (RVKRPR), respectively, were used with 3′ end primer wt2rev 5′-TCCTAAGCTTGACTGAGCCTCACTGAGGCAATAGCTCC-3′, which contains a stop codon and a HindIII site. Template DNA was pZWavrXa7M123. The BglII-HindIII fragment of the PCR product was ligated into pZWavrXa7M123 to generate plasmids pZWavrXa7SV40 and pZWavrXa7MN3, respectively. pZWavrXa7SV40 was used to generate pNIAXa7SV40. The plasmids pZWavrXa7, pZWavrXa7M123, pZWavrXa7SV40 and pZWavrXa7MN3 were cut with HindIII and ligated into pHM1, then conjugated into PXO99A and PXO86mX53, respectively, by mating with the helper strain HB101 containing pRK2013. Respective pNIA fusion plasmids (pNIA, pNIAXa7, pNIAXa7 M123, and pNIAXa7SV40) were transformed into yeast strain L40, which contained reporter genes His3 and LacZ. The transformed yeast cells were used to measure β-galactosidase activity for lacZ activation as described previously (19). Activation of His3 was assessed by growth on histidine-deficient media.
DNA-Binding Assays.
The avrXa7 coding region from pZWavrXa7 was cut by HindIII, partially with PstI, and then ligated into expression vector pPROEX (GIBCO/BRL). Primer 5′-GGGGTACCAGGAGATCTGCCATGGACCCCATTCGTTCG-3′ was used to add a BglII site upstream of the start codon of avrXa7, and the DNA was then cut with BglII and HindIII and ligated into pGEX vector (Amersham Pharmacia Biotech). The respective fusion proteins of AvrXa7 with 6His- or GST-tag were expressed and purified by affinity chromatography according to the supplier (Amersham Pharmacia Biotech). The DNA was labeled with 32P by end labeling for PolydA, T, C, G, A/T, and G/C and by PCR for random oligonucleotides, which are 5′-CAGGGCATGTGGATCCCN25GGGCCTGCAGGAATTCGA-3′, where N25 indicates 25 randomly synthesized bases. Purified protein (approximately 30 ng) was incubated in 12 mM Tris (pH 7.9)/12% glycerol/35 mM KCl/0.07 mM EDTA/8.5 mM MgCl2 with 1–2 ng of labeled DNA at room temperature for 20 min. The reaction products were separated on a 6% native PAGE gel with a running buffer of 25 mM Tris (pH 8.3)/190 mM glycine/1 mM EDTA.
Results
AvrXa7 Contains a Unique and Unusual Repeat Region That Determines Avirulence Specificity for Xa7 and Virulence on Rice.
The features of avrXa7 are shown in Fig. 1. The sequence of the 3.2-kb SphI revealed that avrXa7 had 25.5 repeats, and the order of the repeats, as defined by the twelfth and thirteenth codons of each repeat, were unique in comparison to previously characterized members of the family (Fig. 2A). The repeat coding region of avrXa7 is unusual compared with most other members of the avrBs3 family because of the presence of four repeat units 99 bp in length rather than 102 bp (Fig. 2A), having an 18-bp duplication within repeat 13 (Fig. 2B). The shorter repeat units appear to be missing the thirteenth codon of the prototypical repeat unit. Replacement of the repeat region of pZWavrXa10 with the repeat region of avrXa7 by using the central SphI fragment changes the specificity from Xa10 to Xa7 when the avirulence gene was present in strain PXO99 (Table 1). The composite gene in pZWavrXa7 was used here because further experiments required the introduction of mutations that were originally constructed in avrXa10. Because the C-terminal coding regions are interchangeable, C-terminal mutations could be readily transferred to avrXa7 simply by replacing the C-terminal coding region of pZWavrXa7 with the C-terminal coding region of avrXa10. To determine whether the composite gene had the virulence properties of the original avrXa7, pZWavrXa7 was introduced into PXO86mx53, and the resulting strain was tested for virulence (Table 1). Strain PXO86mx53 has a mutation in the endogenous copy of avrXa7 and retains an active copy of avrXa10. PXO86mx53 has no avirulence activity on IRBB7 rice containing Xa7 and impaired virulence on the susceptible rice line IR24 (Table 1). The decrease in virulence of PXO86mx53 is evidenced by reduced water soaking, a phenomenon caused by release of water into the intercellular spaces from cells in the infected area (Fig. 3A), reduced leaf lesion lengths (Fig. 3B), and lower bacterial populations in the leaves (data not shown). The gene in pZWavrXa7 was capable of restoring full virulence as well as avirulence specificity for Xa7 (Table 1; Fig. 3). Therefore, the repeat region of avrXa7 was sufficient to convert avrXa10 into a gene with the virulence and avirulence properties of avrXa7. We refer to the composite gene hereafter simply as avrXa7.
Figure 1.
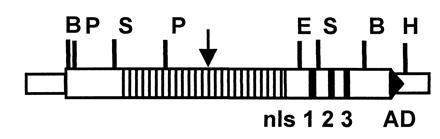
Map of avrXa7. The repeat domain is shown as a series of boxes in the middle of the gene. nls 1 2 3, NLS sequences 1, 2, and 3. AD, acidic transcriptional activation domain. E, EcoRI; P, PstI; S, SphI; H, HindIII. Arrow indicates position of Tn5-B20∷53 insertion used to generate PXO86mx53 (see Results).
Figure 2.
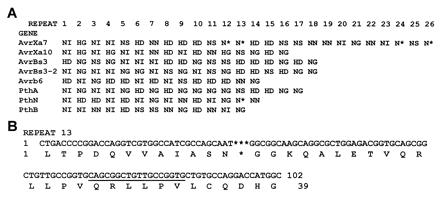
Sequence analysis of avrXa7. (A) Schematic representation of the repeat domain of the avrXa7 gene product by using the single amino acid code for the twelfth and thirteenth codons of the repeat units. GenBank accession nos. for proteins are as follows: AvrXa7, AF275317; AvrXa10, AAA92974; AvrBs3, CAA34257; AvrBs3–2, S34809; Avrb6, AAB00675; PthA, AAC43587; PthN, AAB69865; PthB, AAD01494. (B) Unusual repeat thirteen of avrXa7. Duplicated region is underlined. Asterisks indicate positions of missing nucleotides or amino acid residues of the prototypic repeat unit.
Table 1.
Dual activity of avrXa7
| Strain | BB7 (Xa7) | BB10 (Xa10) | IR24 |
|---|---|---|---|
| PXO99 (pHM1)* | S | S | S |
| PXO99 (p29-29-2A)† | R | S | S |
| PXO99 (pZWavrXa10)‡ | S | R | S |
| PXO99 (pZWavrXa7)‡ | R | S | S |
| PXO86 (pHM1) | R | R§ | S |
| PXO86mx53 (pHM1) | I | R¶ | I |
| PXO86mx53 (p29-29-2A) | R | S | S |
| PXO86mx53 (pZWavrXa7) | R | S | S |
S, susceptible reaction; R, resistant; I, impaired.
Vector alone.
Cosmid clone with avrXa7 (1).
SphI fragment of avrXa7 exchanged with SphI fragment of avrXa10 (14).
PXO86 contains a chromosomal copy of avrXa10 (1).
Although growth is poor because of the impaired growth of the bacteria, BB10 plants display a resistant phenotype.
Figure 3.
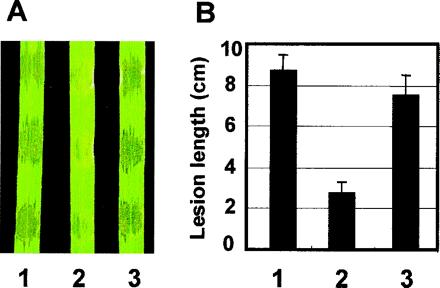
Repeat domain of avrXa7 controls virulence specificity. (A) Leaves were inoculated with wild-type PXO86 (leaf 1); PXO86mx53 (pHM1) with the mutated copy of avrXa7 and the cloning vector alone (leaf 2); and PXO86mx53 (pZWavrXa7) (leaf 3). Leaves were photographed 3 days after inoculation. Dark areas represent water-soaked tissue. (B) Lesion length measurements (cm) 11 days after leaf-clip inoculation.
The AD Is Required for Virulence and Avirulence Activities.
Deletion of the C-terminal coding region by the introduction of a stop codon (TGA1064) or replacement of the C-terminal codons 1080, 1085, and 1088 (FLL) in the AD with alanine codons was previously shown to result in the loss of transcriptional activation and avirulence activity because of avrXa10 (19). The mutations did not result in changes in the apparent stability of the protein in Xanthomonas, and the effects of the mutations could not be abrogated by expression of the genes within the host cells (14, 19). The 3.2-kb SphI fragment from avrXa7 was used to replace the corresponding fragment from avrXa10TGA1064 and avrXa10FLL to create avrXa7 versions of the same mutations (avrXa7TGA and avrXa7FLL). Both mutations caused the loss of avirulence and virulence because of avrXa7 (Table 2; Fig. 4). Similarly to AvrXa10, no differences were detected in the amount of protein synthesized in the bacterium (data not shown). Transient expression of avrXa7 in the host cells was not tested. Loss of avirulence activity could be restored to avrXa7TGA by the introduction of the AD of VP16 (Table 2; Fig. 4). However, the AD of VP16 could not restore virulence activity to avrXa7TGA (Table 2, Fig. 4).
Table 2.
Requirement of the AD for avrXa7 activity
| Strain | IRBB7 (Xa7) | IR24 |
|---|---|---|
| PXO99 (pHM1) | S | S |
| PXO99 (pZWavrXa7) | R | S |
| PXO99 (pZWavrXa7)TGA* | S | S |
| PXO99 (pZWavrXa7)FLL† | S | S |
| PXO86 (pHM1) | R | S |
| PXO86mx53 (pHM1) | I | I |
| PXO86mx53 (pZWavrXa7) | R‡ | S |
| PXO86mx53 (pZWavrXa7TGA) | I | I |
| PXO86mx53 (pZWavrXa7FLL) | I | I |
| PXO99 (pZWavrXa7VP16) | R | S |
| PXO86mx53 (pZWavrXa7VP16) | R‡ | I |
S, susceptible reaction; R, resistant; I, impaired.
TGA indicates TGA stop codon at position 1064 of AvrXa10.
FLL indicates replacement of phenylalanine, leucine, and leucine with alanines at positions 1080, 1085, and 1088, respectively, in reference to AvrXa10.
Resistance response is weak but evident.
Figure 4.

Virulence activity of AvrXa7 requires C-terminal AD. Rice leaves were syringe inoculated with PXO86mx53 (leaves 1–3) or PXO99 (leaves 4–8) containing the following genes: 1, avrXa7; 2, avrXa7TGA; 3, avrXa7VP16; 4, vector control; 5, avrXa7; 6, avrXa7TGA; 7, avrXa7VP16; 8, avrXa7VP16. Leaves 1–3 and 8 are from IR24. Leaves 4–7 are from IRBB7 (Xa7).
NLS Motifs Are Required for Avirulence and Virulence Activities of AvrXa7.
The three consensus NLS motifs in the C-terminal coding portion of avrXa7 were altered to nonconsensus sequences, creating avrXa7 M123 (pZWavrXa7M123; Fig. 5A). To directly measure the effect of the NLS mutations on nuclear localization, the ability of AvrXa7 to localize to nuclei was assayed in a yeast nuclear localization reporter system (Fig. 5B). In this system, the coding sequences of the gene are fused to the coding sequence for the LexA DNA-binding protein that lacks a NLS motif. The yeast strain contains reporter genes (lacZ and His3) that are regulated by a minimal yeast core promoter and tandem copies of the LexA consensus binding site. Expression of the lacZ and His3 genes depends on the presence of a NLS in the LexA fusion protein. When full-length wild-type avrXa7 was fused in frame to the coding region for the DNA-binding domain of LexA, the yeast strain was capable of growth without the addition of exogenous histidine and had high levels of β-galactosidase activity (Table 3). Fusion of AvrXa7 M123 to LexA resulted in weak growth on histidine-deficient media and a 76% reduction in the level of β-galactosidase (Table 3). Addition of the NLS from SV40 to avrXa7 M123 (pZWavrXa7SV40) resulted in the restoration of histidine-free growth and 77% of the β-galactosidase activity because of the wild-type protein (Table 3). AvrXa7 M123 and avrXa7SV40 were then introduced into X. oryzae pv. oryzae and tested for avirulence and virulence activities. When introduced into PXO99A and tested on IRBB7, avrXa7 M123 had no avirulence activity (Fig. 6). In PXO86mx53, avrXa7 M123 also had no virulence activity (Fig. 6). The addition of the SV40 T-antigen NLS (AvrXa7SV40, Fig. 5A) or NLS 3 of AvrXa7 (data not shown) restored both activities (Fig. 6). Protein levels in yeast or bacteria were unchanged by the NLS mutations (data not shown).
Figure 5.
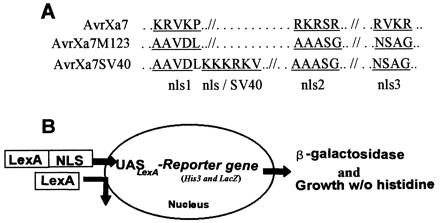
NLS mutations and yeast nuclear localization assay. (A) Amino acid replacements at NLS 1, 2, and 3 are underlined. Only regions of NLS motifs are shown. KKKRK was introduced into AvrXa7 M123 to create AvrXa7SV40. (B) Each gene was fused to the LexA coding DNA-binding domain in pNIA vector and expressed in yeast. Strain L40 contains LexA-binding sites (UASLexA) immediately upstream of lacZ and His 3 genes. Induction of lacZ and His3 depends on the nuclear localization of the LexA fusion product. Induction leads to increased β-galactosidase activity and the ability of the yeast strain to grow in histidine-deficient media.
Table 3.
Effect of nls123 mutations on nuclear localization in yeast assay
| Gene | Histidine deficiency* | % β-galactosidase activity† |
|---|---|---|
| pNIA | − | 3.1 ± 1.1 |
| pNIAXa7 | + | 100 |
| pNIAXa7M123 | ± | 20.0 ± 3.8 |
| pNIAXa7SV40 | + | 76.3 ± 2.4 |
Histidine deficiency was measured as the ability to grow on minimal media with 20 mM 3-AT
Activity is expressed as percent of activity of o-nitrophenyl β-d-galactopyranoside (ONPG) produced per hour by pNIAXa7.
Figure 6.
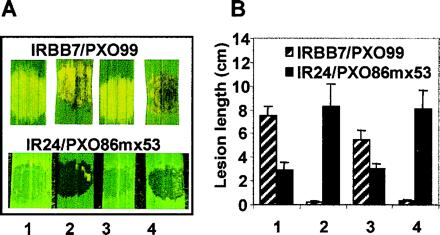
Effects of NLS mutation on the avirulence and virulence activities of avrXa7. (A) Leaves of IRBB7 rice containing the resistance gene Xa7 (Upper) and the susceptible variety IR24 (Lower) were syringe inoculated with PXO99 or PXO86mx53, respectively, containing the following genes: (1) pHM1, vector alone; (2) pZWavrXa7; (3) pZWavrXa7M123; (4) pZWavrXa7SV40. (B) Lesion length measurements 11 days after leaf-clip inoculations of IRBB7 with PXO99 (hatched bars) or IR24 with PXO86mx53 (solid bars). Numbering is as for A.
AvrXa7 Binds DNA.
The requirement for nuclear localization and transcriptional activation prompted us to consider whether the function of AvrXa7 might involve interaction with host DNA. DNA binding was assayed by gel shift assays by using AvrXa7 protein and 32P-labeled DNA oligonucleotides. In this assay, evidence for DNA binding is indicated by the slower migration of the labeled oligonucleotides that are bound to protein. AvrXa7 was synthesized in E. coli as a GST fusion. GST alone had no DNA-binding activity (Fig. 7A). None of the four single-stranded homomeric oligonucleotides [single-stranded (ss)DNA] were retarded by GST/AvrXa7 (Fig. 7A). The migration of both PolydA/dT and PolydC/dG was retarded by the GST/AvrXa7 fusion. The amount of label in the retarded bands was greater for dA/dT than dC/dG. A second assay was performed by using polyhistidine-tagged AvrXa7 and 32P-labeled random double-stranded DNA oligonucleotides. In this assay, ssDNA oligonucleotides of each base, PolydA/dT or PolydC/dG were added in 100-fold excess over the labeled random oligonucleotides. As with the GST fusion, AvrXa7 caused the gel retardation of labeled random oligonucleotides, and binding to the oligonucleotides was completely inhibited by the addition of dA/dT (Fig. 7B). Slight inhibition was observed with both PolydG and PolydC/dG. Thus, AvrXa7 will bind PolydA/dT and, to a lesser extent, PolydC/dG. Similarly, PolydC/dG is not a good competitor against a pool of random oligonucleotides (Fig. 7B).
Figure 7.
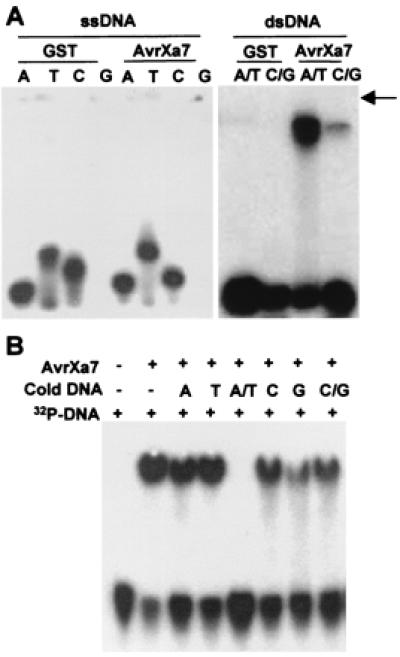
AvrXa7 is a DNA-binding protein. (A) Single-stranded and double-stranded oligonucleotides were end labeled with 32P, mixed with AvrXa7 or GST (glutathione S-transferase), and subjected to polyacrylamide electrophoresis. Arrow indicates position of well. (B) DNA-binding competition to AvrXa7. Random oligonucleotides were labeled with 32P and mixed with AvrXa7 and the indicated unlabeled DNA in 100-fold excess. A, T, C, G, indicate unlabeled single-stranded homopolymers; A/T and C/G indicate unlabeled double-stranded polymers. +/− indicate presence or absence of component.
Discussion
We have demonstrated that the specificity for avirulence activity and virulence effects of avrXa7, a dual-acting avirulence gene of the avrBs3 family, depended on the unique structural features of the products of this gene family. Specificity for enhancing virulence and Xa7-dependent avirulence activity on rice depended on the repeat domain, and both activities required functional NLS and AD motifs. The virulence properties of the avrBs3 family were first demonstrated by the identification of pthA and avrb6 (7, 16). The latter two genes have repeat domains of 13.5 and 18.5 units, respectively, and a unique repeat domain as defined by the twelfth and thirteenth codons of each repeat unit. Exchange of the domains of avrb6 and pthA, which have effects on cotton and citrus species, respectively, also resulted in a switch in the virulence specificities of the genes (16). Replacement of the repeat region of avrXa10, which has no apparent contribution to virulence, with the repeat region of avrXa7 also created a gene with all of the properties of avrXa7. The dual-acting genes are presumably under selective pressure to maintain virulence function while avoiding recognition by the host defense system. The adaptive process is presumably reflected in the variety of repeat domains in the genes from different strains and within strains. The length of a given pathogenicity gene may reflect the particular configuration of variable regions necessary for virulence that was arrived at by recombination and mutation. Strains of X. oryzae pv. oryzae contain an estimated 12 to 14 copies of the genes, which may provide ample substrate for the generation of new repeat combinations (1). How the particular repeat domains mediate specificity for virulence or resistance remains a mystery.
The requirement for NLS motifs had previously been demonstrated only for the avirulence activity of avrBs3 and avrXa10 (14, 18). These findings demonstrated that removal of the NLS motifs also resulted in the loss of virulence activity. As with AvrBs3 and AvrXa10 or any other avirulence protein, AvrXa7 has not been directly observed in the host cytoplasm or nuclei after transfer from the bacterium. Corroborative evidence, however, is abundant. Studies with truncated AvrBs3 and PthA demonstrated that the NLS motifs in the C-terminal portion of the protein could direct a β-glucuronidase fusion protein to onion cell nuclei (17, 18). Full-length fusions of AvrXa10 with the Gal4 DNA-binding domain were directed to Arabidopsis cell nuclei on the basis of transcriptional activation activity (14). The NLS signals of AvrXa7 fall into the class that function in a broad range of eukaryotic cells (24). Therefore, we used a yeast one-hybrid system in this work in an attempt to assess the behavior of full-length AvrXa7. The results indicated that full-length AvrXa7 was efficiently localized to yeast nuclei provided the endogenous NLS or heterologous SV40 motifs were intact. The formal possibility remains that the NLS sequences of AvrXa7 and related proteins are required, in general, for secretion through the type III apparatus and/or stability in the plant cells. However, a role for the NLS motifs in secretion is unlikely because of several considerations. Some secreted proteins from Xanthomonas do not have the NLS (or AD) motifs, as noted below for AvrBsP, yet remain functional (25, 26), and secretion signals for the type III systems appear, in part, to be encoded in the N-terminal mRNA structure (27). In regards to stability in the plant cells, mutations of the NLS motifs had no effect on the stability of the C terminus when fused to β-glucuronidase (18). Therefore, these proteins can localize to the nuclei of their respective host cells, and, indeed, we conclude that the proteins must localize to the nuclei to perform their function.
A domain with the features of an acidic-type AD is found at the C terminus of proteins encoded by the avrBs3 gene family (19). The AD is required for the virulence and avirulence activities of AvrXa7. Avirulence activity could be restored by introduction of the heterologous domain. However, the VP16 domain could not substitute for the endogenous domain in virulence assays. Therefore, regardless of the actual function of the domain, virulence and avirulence activities have different requirements. If the activation domain is indeed functional, the interaction of AvrXa7 with the transcriptional machinery and recognition as mediated by Xa7 (or AvrXa10 by Xa10 or AvrBs3 by Bs3) may be a relatively general phenomenon, whereas virulence may require a specific interaction with a host factor, possibly a transcription cofactor. Whether the domain acts as an activator in vivo has yet to be demonstrated. Activation activity has been demonstrated only after fusion of the protein to a DNA-binding domain from a well-characterized transcription factor. The ability of AvrXa7 by itself to promote transcription of a gene either in yeast or a plant has not been demonstrated. However, we were able to demonstrate that the protein is a general double-stranded DNA-binding protein. Further work will be required to determine which features of AvrXa7 are responsible for DNA binding and whether high-affinity sites can be identified.
The requirement of the AD and NLS for both virulence and avirulence activities of AvrXa7 is curious. The finding suggests that either the host defense factors are targeted to the site of action of the virulence factor or that R-gene products may be components of a complex that is targeted by the virulence factor. In tomato, the AvrPto protein, which also has virulence properties (28, 29), requires a myristylation motif for activity, suggesting that the protein is targeted to an inner membrane surface (personal communication). In Arabidopsis, RPM1 has been localized to the inner cytoplasmic membrane (30). Interestingly, the corresponding avirulence protein AvrRpm1, which functions in virulence for strain PsmM2 of Pseudomonas syringae pv. maculicola, also has a myristylation motif (31) and recently has been found associated with the cytoplasmic membrane on expression within the host cell (32). Loss of the myristylation site in AvrRpm1 results in loss of avirulence and virulence activities (32). Therefore, localization of the resistance gene product to the site of effector protein function may be a more general phenomenon and not specific to AvrXa7 and related proteins. Localization to the site of action may ensure that the virulence protein is effectively intercepted and the defense response initiated rather than usurped during pathogen challenge. Specific R-gene products may be targeted to and function as “guardians” of specific defense signaling complexes (33). At the same time, not all members of the avrBs3 family appear to depend on the NLS sequences for avirulence activity and therefore are not necessarily intercepted in the nucleus. AvrBs3–2, which is also referred to as avrBsP and represents another member of family with avirulence activity on tomato, was initially cloned with a truncated C-terminal coding region and therefore lacks the NLS coding regions (25, 34). One presumes that the AvrBs3–2 protein is localized to the nucleus yet intercepted in the cytoplasm.
Little is known about the function of the type III dependent virulence factors of plant pathogenic bacteria. The similarity of AvrXa7 and related proteins, regardless of the precise mechanism, suggests that all members of the family are likely to have a requirement to localize to the nuclei of their respective host plants and have the potential to interact with the host transcriptional machinery. AvrXa7 is the only avrBs3-related gene of X. oryzae pv. oryzae to be demonstrated to have virulence properties. The fact that PXO99 and other strains do not have avrXa7 activity yet remain fully virulent suggests either not all strains of X. oryzae pv. oryzae require an avrBs3 member for full virulence or that genes without Xa7 recognition can also function in virulence. Further characterization of avrXa7 and related genes in X. oryzae pv. oryzae will hopefully allow identification of the critical repeat configurations for avirulence and virulence activities.
Acknowledgments
We thank Drs. Yoon Rhee and Vitaly Citovsky for providing vectors before publication and Diana Pavlisko for assistance in the preparation of the manuscript. This work was supported by funds provided by the United States Department of Agriculture agreement 98–353303-6446 and the Kansas Agriculture Experiment Station (KAES). W.Z. was supported in part by a grant from The Rockefeller Foundation. This paper is contribution no. 00–419-J from KAES.
Abbreviations
- NLS
nuclear localization signal
- AD
transcriptional activation domain
Footnotes
Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. AF275317).
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.170286897.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.170286897
References
- 1.Hopkins C M, White F F, Choi S-H, Guo A, Leach J E. Mol Plant–Microbe Interact. 1992;5:451–459. doi: 10.1094/mpmi-5-451. [DOI] [PubMed] [Google Scholar]
- 2.Vivian A, Gibbon M J. Microbiology. 1997;143:693–704. doi: 10.1099/00221287-143-3-693. [DOI] [PubMed] [Google Scholar]
- 3.Bonas U, Schulte R, Fenselau S, Minsavage G V, Staskawicz B J, Stall R E. Mol Plant–Microbe Interact. 1991;4:81–88. [Google Scholar]
- 4.Jackson R W, Athanassopoulos E, Tsiamis G, Mansfield J W, Sesma A, Arnold D L, Gibbon M J, Murillo J, Taylor J D, Vivian A. Proc Natl Acad Sci USA. 1999;96:10875–10880. doi: 10.1073/pnas.96.19.10875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Bonas U, van den Ackerveken G. Plant J. 1997;12:1–7. doi: 10.1046/j.1365-313x.1997.12010001.x. [DOI] [PubMed] [Google Scholar]
- 6.White F F, Yang B, Johnson L B. Curr Opin Plant Biol. 2000;3:291–298. doi: 10.1016/s1369-5266(00)00082-0. [DOI] [PubMed] [Google Scholar]
- 7.Swarup S, Yang Y, Kingsley M T, Gabriel D W. Mol Plant–Microbe Interact. 1992;5:204–213. doi: 10.1094/mpmi-5-204. [DOI] [PubMed] [Google Scholar]
- 8.De Feyter R, Yang Y, Gabriel D W. Mol Plant–Microbe Interact. 1993;6:225–237. doi: 10.1094/mpmi-6-225. [DOI] [PubMed] [Google Scholar]
- 9.De Feyter R, Gabriel D W. Mol Plant–Microbe Interact. 1991;4:423–432. [Google Scholar]
- 10.Chakrabarty P K, Duan Y P, Gabriel D W. Phytopathology. 1997;87:1160–1167. doi: 10.1094/PHYTO.1997.87.11.1160. [DOI] [PubMed] [Google Scholar]
- 11.Choi S. Ph.D. thesis. Manhattan, KS: Kansas State University; 1993. [Google Scholar]
- 12.Leach J E, Zhu W, Chittoor J M, Ponciano G, Young S A, White F F. In: Biology of Plant–Microbe Interactions. Stacey G, Mullin B, Gresshoff P M, editors. St. Paul, MN: International Society for Molecular Plant–Microbe Interactions; 1996. pp. 191–196. [Google Scholar]
- 13.Herbers K, Conrads-Strauch J, Bonas U. Nature (London) 1992;356:172–174. [Google Scholar]
- 14.Zhu W, Yang B, Chittoor J M, Johnson L B, White F F. Mol Plant–Microbe Interact. 1998;11:824–832. doi: 10.1094/MPMI.1998.11.8.824. [DOI] [PubMed] [Google Scholar]
- 15.Yang Y, Gabriel D W. J Bacteriol. 1995;177:4963–4968. doi: 10.1128/jb.177.17.4963-4968.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Yang Y, De Feyter R, Gabriel D W. Mol Plant–Microbe Interact. 1994;7:345–355. doi: 10.1094/mpmi-6-225. [DOI] [PubMed] [Google Scholar]
- 17.Yang Y, Gabriel D W. Mol Plant–Microbe Interact. 1995;8:627–631. doi: 10.1094/mpmi-8-0627. [DOI] [PubMed] [Google Scholar]
- 18.Van den Ackerveken G, Marois E, Bonas U. Cell. 1996;87:1307–1316. doi: 10.1016/s0092-8674(00)81825-5. [DOI] [PubMed] [Google Scholar]
- 19.Zhu W, Yang B, Wills N, Johnson L B, White F F. Plant Cell. 1999;11:1665–1674. doi: 10.1105/tpc.11.9.1665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A. Current Protocols in Molecular Biology. New York: Wiley; 1988. [Google Scholar]
- 21.Guthrie C, Fink G R. Guide to Yeast Genetics and Molecular Biology. San Diego: Academic; 1991. [Google Scholar]
- 22.Kauffman H E, Reddy A P K, Hsiek S P V, Marca S D. Plant Dis Rep. 1973;57:537–541. [Google Scholar]
- 23.Rhee Y, Gurel F, Gafni Y, Dingwall C, Citovsky V. Nat Biotechnol. 2000;18:433–437. doi: 10.1038/74500. [DOI] [PubMed] [Google Scholar]
- 24.Smith H M S, Raikhel N V. Plant Physiol. 1999;119:1157–1163. doi: 10.1104/pp.119.4.1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Canteros B, Minsavage G, Bonas U, Pring D, Stall R. Mol Plant–Microbe Interact. 1991;4:628–632. doi: 10.1094/mpmi-4-628. [DOI] [PubMed] [Google Scholar]
- 26.Rossier O, Wengelnik K, Hahn K, Bonas U. Proc Natl Acad Sci USA. 1999;96:9368–9373. doi: 10.1073/pnas.96.16.9368. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Anderson D M, Fouts D E, Collmer A, Schneewind O. Proc Natl Acad Sci USA. 1999;96:12839–12843. doi: 10.1073/pnas.96.22.12839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Chang J H, Rathjen J P, Bernal A J, Staskawicz B J, Michelmore R W. Mol Plant–Microbe Interact. 2000;13:568–571. doi: 10.1094/MPMI.2000.13.5.568. [DOI] [PubMed] [Google Scholar]
- 29.Shan L, He P, Zhou J, Tang X. Mol Plant–Microbe Interact. 2000;13:592–598. doi: 10.1094/MPMI.2000.13.6.592. [DOI] [PubMed] [Google Scholar]
- 30.Boyes D C, Nam J, Dangl J L. Proc Natl Acad Sci USA. 1998;95:15849–15854. doi: 10.1073/pnas.95.26.15849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dangl J L, Ritter C, Gibbon M J, Mur L A J, Wood J R, Goss S, Mansfield J, Taylor J D, Vivian A. Plant Cell. 1992;4:1359–1369. doi: 10.1105/tpc.4.11.1359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Nimchuk Z, Marois E, Kjemtrup S, Leister R T, Katagiri F, Dangl J L. Cell. 2000;101:353–363. doi: 10.1016/s0092-8674(00)80846-6. [DOI] [PubMed] [Google Scholar]
- 33.Van Der Biezen E A, Jones J D. Trends Biochem Sci. 1998;23:454–456. doi: 10.1016/s0968-0004(98)01311-5. [DOI] [PubMed] [Google Scholar]
- 34.Bonas U, Conrads-Strauch J, Balbo I. Mol Gen Genet. 1993;238:261–269. doi: 10.1007/BF00279555. [DOI] [PubMed] [Google Scholar]