

Abstract
Background
Mycobacteria have developed a number of pathways that provide partial protection against both reactive oxygen species (ROS) and reactive nitrogen species (RNS). We recently identified a locus in Mycobacterium marinum, mel2, that plays a role during infection of macrophages. The molecular mechanism of mel2 action is not well understood.
Results
To better understand the role of the M. marinum mel2 locus, we examined these genes for conserved motifs in silico. Striking similarities were observed between the mel2 locus and loci that encode bioluminescence in other bacterial species. Since bioluminescence systems can play a role in resistance to oxidative stress, we postulated that the mel2 locus might be important for mycobacterial resistance to ROS and RNS. We found that an M. marinum mutant in the first gene in this putative operon, melF, confers increased susceptibility to both ROS and RNS. This mutant is more susceptible to ROS and RNS together than either reactive species alone.
Conclusion
These observations support a role for the M. marinum mel2 locus in resistance to oxidative stress and provide additional evidence that bioluminescence systems may have evolved from oxidative defense mechanisms.
Background
Mycobacteria appear to have numerous molecular pathways responsible for their inherent resistance to reactive oxygen species (ROS) [1-3]. In most bacteria, oxidative stress induces a global regulator, OxyR, that induces detoxifying enzymes such as alkyl hydroperoxide reductase (AhpC) and catalase/hydroperoxidase I (KatG) [4,5]. During normal aerobic metabolism bacteria produce superoxide (O2 •-) that is converted to hydrogen peroxide (H2O2) and oxygen (O2) by superoxide dismutase and H2O2 is converted to water (H2O) and O2 by KatG [6] or AhpC [7]. The two superoxide dismutase (SOD) genes present in mycobacteria, sodA and sodC, have been suggested to play a role in resistance to ROS. A sodC mutant is more susceptible to ROS, including hydrogen peroxide (H2O2), and displays a defect in growth within activated macrophages [8,9]. The sodA gene has been down-regulated by antisense methods, resulting in increased sensitivity to H2O2 [10]. Mycobacteria also express a catalase, KatG, that affects resistance to ROS produced by NADPH oxidase activity in activated macrophages [2]. Other pathways must play an important role in resistance of M. tuberculosis to oxidative stress because oxyR is inactive [11], katG is absent or mutated in numerous human clinical isolates [12-16] and ahpC is expressed at very low levels [17,18].
Similar to ROS, there are several pathways involved in mycobacterial resistance to reactive nitrogen species (RNS), including noxR1, noxR3 [19,20], dlaT [21], msrA [22,23], cysH [3], DNA repair, protein degradation in the proteasome and flavin cofactor synthesis [24]. In addition to its role in resistance to ROS, the mycobacterial ahpC is also involved in resistance to the RNS peroxynitrite, but not nitric oxide [25]. Peroxynitrite is produced by SOD in the presence of H2O2 and nitric oxide, linking these two important mechanisms of oxidative stress-mediated cell death [26]. This observation may help to explain the inherent resistance of M. tuberculosis to peroxynitrite as compared to less pathogenic mycobacteria [27].
Bioluminescence systems can protect cells against ROS [28-32] through a catalase-like reaction between the electron donating ROS and oxidized luciferase-bound flavin mononucleotide, producing water and light [33]. The similarity of luciferases to oxidases [34] suggests that bioluminescence systems could have evolved from oxygen defense mechanisms [35]. During genetic analysis of factors that affect macrophage infection, we identified the M. marinum mel2 locus, which displays similarity to lux genes involved in bioluminescence [36]. In the current study, more detailed analysis of the genes in the mel2 locus suggests functional similarity between mel2 and bioluminescence systems. Based on this similarity, we asked whether the M. marinum mel2 locus is involved in resistance of mycobacteria to oxidative stress. We constructed an M. marinum mutant that carries a transposon insertion in the first gene in the mel2 locus, melF, by allelic exchange and demonstrated that this mutant displays increased susceptibility to both ROS and RNS. Since this mutation may have polar effects on downstream genes, we complemented this mutant with two constructs, one that carries the melF gene alone and another with the entire mel2 locus. The melF mutant defect is partially complemented by melF alone, but fully complemented by the entire mel2 locus. We recently found that the mel2 mutant displays a defect for growth in activated macrophages that is alleviated by the presence of either ROS scavengers or nitric oxide synthase inhibitors [37], suggesting that the mel2 mutant is more susceptible to ROS and RNS than wild type bacteria. The data obtained in the current study support and extend these observations through demonstration that the mel2 locus plays a role in susceptibility to several different compounds that produce ROS and RNS in laboratory media. Our results indicate that the M. marinum mel2 locus is the first of a newly identified class of genes with similarity to bioluminescence genes involved in resistance to both ROS and RNS.
Results
Similarity of the genes in the mel2 locus to bioluminescence genes
The initial analysis of the genes present in the mel2 locus indicated that the melF, melG and melH genes display similarity to luxA [38], luxG [39] and luxH [39] genes involved in bioluminescence [36]. In order to obtain a better understanding of these findings and explore the possibility of additional functional similarities, we conducted detailed analysis of the conserved motifs present within the melF-melK genes. We first conducted an NCBI Conserved Domain (CD) Search with MelF. We obtained a 100% alignment (E = 1 × 10-28) for the 323 amino acid (a.a.) bacterial luciferase-like monooxygenase motif (pfam00296.11; Figure 1A). This motif is conserved in all bacterial luciferase genes, including the luxA and luxB genes from Vibrio harveyi, for which crystal structures have been previously determined [40,41]. Many of the residues responsible for catalytic activity and FMNH2 binding for LuxA and LuxB are also present in MelF [42,43], suggesting that these proteins have related activities. Analysis of the relatedness of MelF to LuxA and LuxB places MelF on an independent branch (Figure 1B), indicating it is nearly equally related to both, with a slightly closer relationship to LuxA than LuxB.
Figure 1.
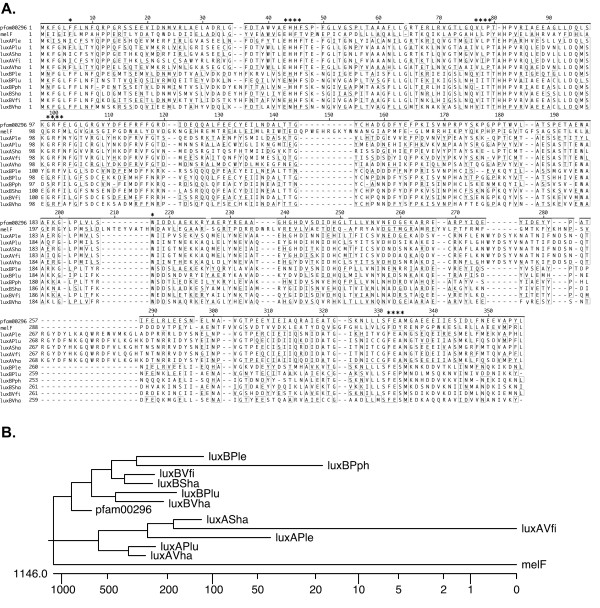
Similarity of MelF to the bioluminescence protein luciferase. Alignment of MelF with LuxA and LuxB proteins from bioluminescent bacteria and the concensus (pfam00296) (A) and a dendogram (B) showing the relationship between them. Numbers to the left of the alignment indicate the position within each protein and above indicate the position within pfam00296. Residues that match the concensus are shown outlined by boxes. Asterixes above alignment indicate conserved regions thought to be involved in enzyme activity. Abbreviations for aligned proteins and their accession numbers are as follows: melF, M. marinum MelF (AAV32084); pfam00296, luciferase-like monooxygenases conserved domain; luxAPle, Photobacterium leiognathi LuxA (P29238); luxAPlu, Photorhabdus luminescens LuxA (AAK98554); luxASha, Shewanella hanedai LuxA (BAB40796); luxAVfi, Vibrio fishcheri LuxA (AAD48477); luxAVha, Vibrio harveyi LuxA (CAA41597); luxBPle, Photobacterium leiognathi LuxB (P09141); luxBPlu, Photorhabdus luminescens LuxB (AAK98555); luxBPph, Photobacterium phosphoreium LuxB (P12744); luxBSha, Shewanella hanedai LuxB (BAB40797); luxBVfi, Vibrio fishcheri LuxB (AAD48478); luxBVha, Vibrio harveyi LuxB (AAA88686). The dendogram was rendered from the alignment using MegAlign (DNASTAR). Tree length is shown to the left of the log scale ruler beneath the dendogram.
Analysis of conserved domains within melG-melK also demonstrated striking similarity to genes involved in bioluminescence (Figure 2A). Functional domains that display similarity to LuxC (MelK), LuxD (MelH), LuxG (MelG) and LuxH (MelJ) were identified. Although there is no clear homologue of LuxE within the mel2 locus, MelH carries domains with similarity to aminopeptidases and lysophospholipases, suggesting that this protein could serve in the role of both the transferase and synthetase activities found in the lux pathway. Additional putative functional domains were present within MelG, MelH and MelI that were not present within the lux genes. Some of these differences may be due to differences in substrate specificity between these pathways and may help to explain why mycobacteria are not luminescent. With the differences and similarities between the mel2 and lux proteins in mind, we constructed a working model for the putative biochemical roles of the proteins encoded by the mel2 locus (Figure 2B). Since mycobacteria face significant ROS during infections, we reasoned that a role in protection against ROS could help to explain the presence of conserved domains between the mel2 locus and bioluminescent systems.
Figure 2.
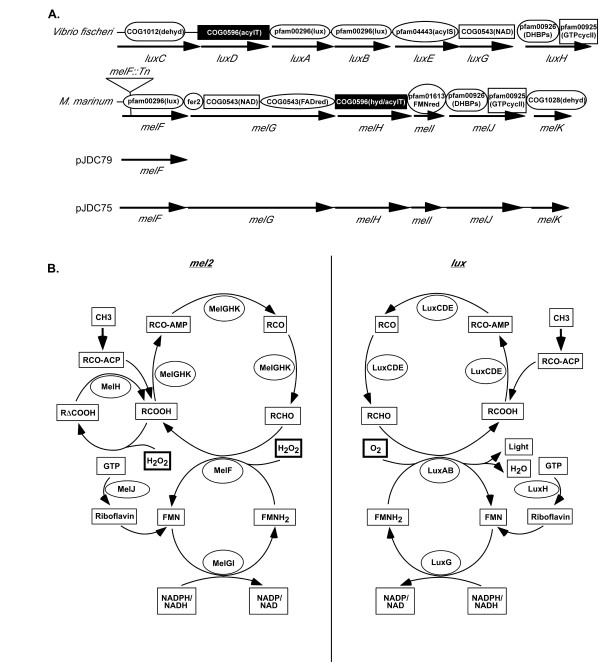
Conserved motifs within the mel2 locus and our working model. Conserved domains found within the Vibrio fisheri lux loci and Mycobacterium marinum mel2 locus (A) and proposed biochemical roles (B). Position of the M. marinum mel2 mutant transposon insertion is shown as a triangle above the mel2 locus (melF::Tn) and the structures of the complementing constructs, pJDC79 and pJDC75, are below the mel2 locus. The luxC-luxG region is located from the V. fisheri chromosome II and the luxH gene is from chromosome I (A). Conserved domains were identified using the NCBI conserved domain search. The designation for each conserved domain(s) is shown above each gene with an abbreviation for its proposed biochemical function in parenthesis. Abbreviations for proposed biochemical functions are as follows with additional conserved domains not shown in the figure in parenthesis: lux, luciferase; fer2, 2Fe-2S iron-sulfur cluster binding domain (cd00207); NAD, flavodoxin oxidoreductases and oxidoreductase NAD-binding domain (pfam00175); FADred, FAD-dependent oxidoreductases; hyd, abhydrolase alpha/beta hydrolase fold (pfam00561); acylT, predicted acyltransferases; FMNred, flavin reductase like domain; DHBPs, 3,4-dihydroxy-2-butanone 4-phosphate synthase; GTPcycII, GTP cyclohydrolase II; dehyd, dehydrogenase; acylS, acyl-protein synthetase. We constructed a hypothetical model for how these proteins might interact to reduce reactive oxygen species (B). The resulting pathways for mel2 are similar to the lux pathways, but are better adapted to serve as a potential defense against oxidative stress through the presence of an epoxide hydrolase (melH), in addition to the reduction of fatty acid aldehydes observed with lux. R represents a number of potential fatty acid molecules that could be used as substrates for these reactions. The LuxCDE proteins function as a complex to produce the aldehydes used to reduce oxygen by LuxAB and we have depicted a similar situation for MelGHK.
M. marinum mel2 mutant and complementing strains
An M. marinum mel2 mutant was constructed by in vitro mutagenesis of the mel2 locus with mini-Mu and replacement of the wild type gene by allelic exchange [36]. We confirmed the presence of the appropriate insertion in the melF gene (Figure 2A) by Southern analysis and PCR. Since insertion mutations can have polar effects on downstream genes and the genes in the mel2 locus are very closely juxtaposed to each other, we asked whether the insertion in melF affects transcript levels for the melG-melK genes. RT-PCR with primer pairs upstream of the melF insertion mutation produces relatively similar levels of product for the M. marinum melF mutant and wild type strains, but RT-PCR with primer pairs within the downstream genes produce less product in the melF mutant than the wild type strain (Figure 3). These observations suggest that the Mu insertion in melF has polar effects on downstream genes and full complementation of this mutation will most likely require the entire mel2 locus. Since the luxA gene plays a pivotal role in bioluminescence and similar genes, including melF, are thought to be oxidoreductases, it is possible that only the melF gene will be required for the role of mel2 in resistance to ROS. In order to differentiate between these possibilities we complemented the M. marinum melF mutant with both melF alone (pJDC79) and the entire mel2 locus (pJDC75) (Figure 2).
Figure 3.
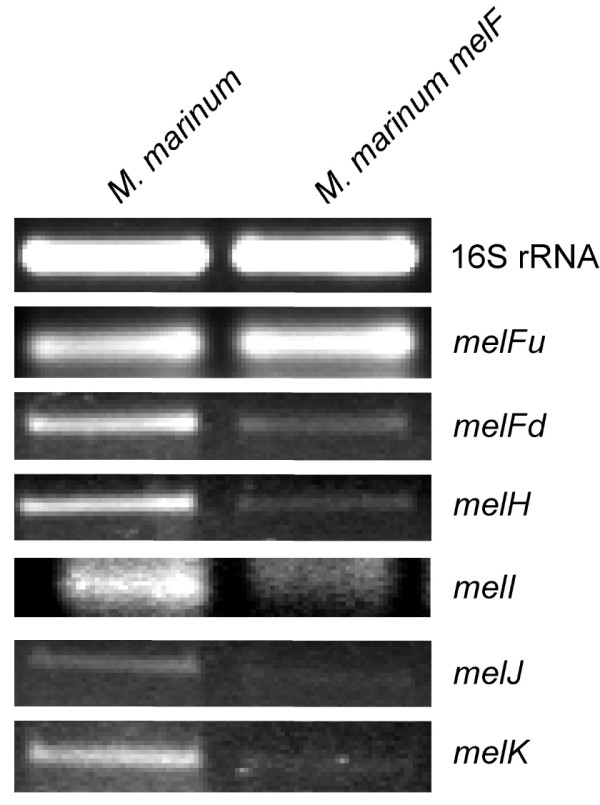
Examination of the polar effects due to the melF insertion mutation. RT-PCR studies to evaluate the effects of the Mu insertion on mel2 transcription upstream (melFu) and downstream of the insertion site (melFd, melH, melI, melJ, melK). Equivalent amounts of RNA from M. marinum wild type and the M. marinum melF mutant were reverse transcribed and subjected to PCR with specific oligonucleotides within each gene. Equal amounts of each PCR product were loaded on 0.8% agarose gels and compared to the 16S rRNA control RT-PCR reaction for each strain (16S rRNA) carried out on the same samples. Data shown are representative of two independent experiments.
The mycobacterial mel2 locus affects susceptibility to ROS
We first compared the ROS susceptibility of wild type M. marinum with that of M. tuberculosis and the non-pathogenic mycobacterial species M. smegmatis (Figure 4A). We found that M. marinum displays similar levels of resistance to H2O2 as M. tuberculosis at various concentrations and times of treatment. In contrast, M. smegmatis is readily killed, even at 1 mM H2O2 where the pathogenic strains are nearly completely resistant (P < 0.001). Interestingly, we found that a mel2 mutant that carries an insertion in the melF gene is much more susceptible than wild type M. marinum to H2O2 (Figure 4B–D; P < 0.01). This difference is more pronounced at 5 mM (between 76–84% for wild type vs. 4–10% survival for the mutant after 2 h) than at 1 mM (between 85–97% for wild type vs. 52–70% survival for the mutant after 2 h). Resistance to H2O2 cannot be restored to the mel2 mutant with the melF gene alone, even expressed from a plasmid (pJDC79), but can be restored by a single copy integrated plasmid carrying the entire mel2 locus (pJDC75). In contrast, no difference in the growth rate or survival of these mycobacterial strains in standard laboratory medium without H2O2 is observed (data not shown). These observations suggests that the melF gene alone is not sufficient to confer resistance to H2O2, and that the melF insertion mutation has polar effects on downstream genes involved in H2O2 resistance.
Figure 4.
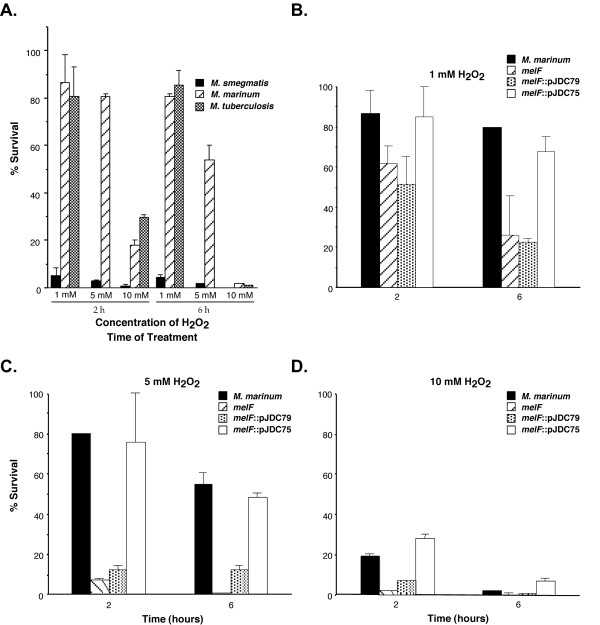
Susceptibility of mycobacterial species and the mel2 mutant to H2O2. Susceptibility of M. smegmatis, M. marinum and M. tuberculosis to different concentrations of H2O2 for 2 or 6 h (A). The data points are absent for M. tuberculosis at 5 mM H2O2 because these bacteria were only tested at 1 and 10 mM concentrations. Susceptibility of M. marinum wild type as compared to the mel2 mutant that carries an insertion in melF (melF), the mutant carrying a plasmid expressing melF alone (melF::pJDC79) and an integrated single copy plasmid expressing the entire mel2 locus (melF::pJDC75) to 1 mM (B), 5 mM (C) and 10 mM (D) H2O2. The % survival = (CFU Tx/CFU initial) × 100. Data are the means and standard deviations of assays done in triplicate, representative of three experiments.
We further probed the role of the mel2 locus in resistance to ROS through the use of two additional ROS generating compounds, cumene hydroperoxide and t-BOOH. Both of these compounds are organic peroxides that produce ROS inside the bacterial cell, but are more stable in aqueous solutions than H2O2. Organic peroxides decompose to alkoxyl and peroxyl radicals in addition to H2O2 [44,45]. The mel2 mutant was more susceptible to both cumene hydroperoxide and t-BOOH than wild type M. marinum (Figure 5; P < 0.01). Interestingly, at the two-hour time point partial complementation of the resistance defect was observed, but once again, the entire mel2 locus confers wild type resistance levels. These observations indicate that the mel2 locus plays a role in resistance to ROS, including the diverse radicals produced by organic peroxides.
Figure 5.
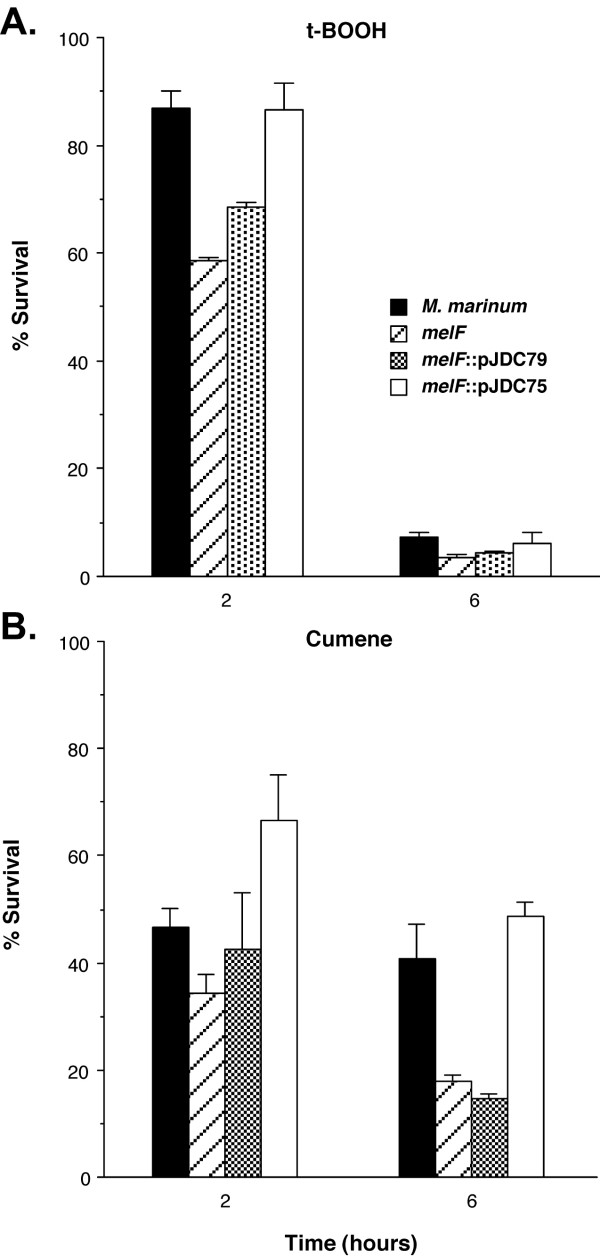
Susceptibility of the mel2 mutant to reactive oxygen species. Susceptibility of M. marinum wild type as compared to the mel2 mutant that carries an insertion in melF (melF), the mutant carrying a plasmid expressing melF alone (melF::pJDC79) and an integrated single copy plasmid expressing the entire mel2 locus (melF::pJDC75) to 10 mM tert-butyl hydroperoxide (t-BOOH, A) or 100 mM cumene hydroperoxide (Cumene, B). The % survival = (CFU Tx/CFU initial) × 100. Data are the means and standard deviations of assays done in triplicate, representative of three experiments.
The mel2 locus affects susceptibility to RNS
Since the mel2 locus plays a role in resistance to ROS, it is also possible that it will affect resistance to RNS. The ROS and RNS pathways are linked in the reaction of nitric oxide with superoxide to produce peroxynitrite [1,46,47]. Because of the importance of RNS in protection against mycobacterial infections [48-51], pathways that affect susceptibility are likely to be important for pathogenesis. We examined the susceptibility of the mel2 mutant to acidified NaNO2, which is a source of nitric oxide [24,48], and SNAP, which releases nitric oxide under neutral pH in the presence of trace metals [52,53]. Similar to ROS, the mel2 mutant displays greater susceptibility than wild type M. marinum to RNS (P < 0.01) and this phenotype can be complemented partially by the melF gene alone and completely by the entire mel2 locus (Figure 6).
Figure 6.
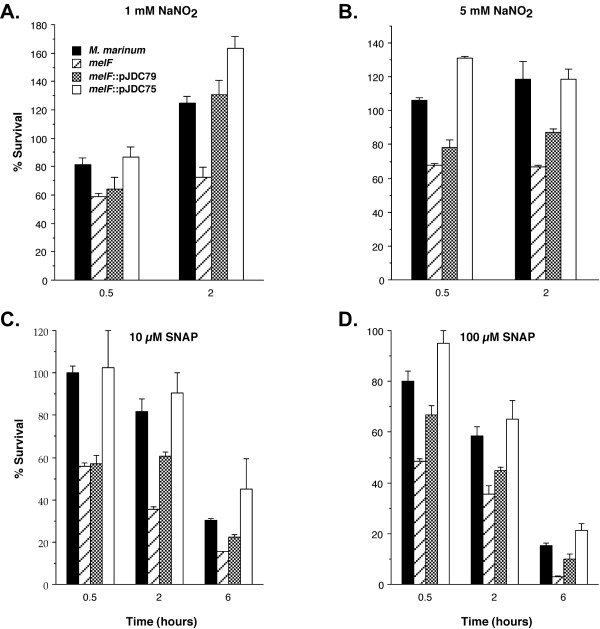
Susceptibility of the mel2 mutant to reactive nitrogen species. Susceptibility of M. marinum wild type as compared to the mel2 mutant that carries an insertion in melF (melF), the mutant carrying a plasmid expressing melF alone (melF::pJDC79) and an integrated single copy plasmid expressing the entire mel2 locus (melF::pJDC75) to 1 mM (A) or 5 mM (B) sodium nitrite (NaNO2) and 10 μM (C) or 100 μM (D) S-nitroso-N-acetyl penicillamine (SNAP). The % survival = (CFU Tx/CFU initial) × 100. Data are the means and standard deviations of assays done in triplicate, representative of three experiments.
The mel2 locus affects susceptibility to the combination of ROS and RNS
Since these observations suggest that the mel2 locus is involved in resistance of mycobacteria to both ROS and RNS, we examined whether the presence of both ROS and RNS simultaneously would have a more dramatic effect upon this mutant. Interestingly, the mel2 mutant is much more susceptible to treatment with both H2O2 and SNAP together than either compound alone (Figure 7; P < 0.01). These observations suggest that the mel2 mutant plays a role in susceptibility to both ROS and RNS, whether treated with them together, as most likely occurs in vivo, or separately.
Figure 7.
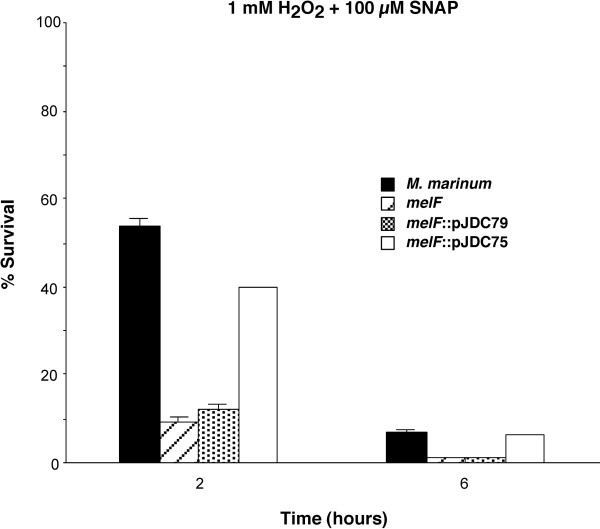
Susceptibility to reactive nitrogen and oxygen species together. Susceptibility of M. marinum wild type as compared to the mel2 mutant that carries an insertion in melF (melF), the mutant carrying a plasmid expressing melF alone (melF::pJDC79) and an integrated single copy plasmid expressing the entire mel2 locus (melF::pJDC75) to H2O2 plus S-nitroso-N-acetyl penicillamine (SNAP). The % survival = (CFU Tx/CFU initial) × 100. Data are the means and standard deviations of assays done in triplicate, representative of three experiments.
Discussion
The molecular mechanisms of mycobacterial resistance to ROS and RNS have been an area of intense investigation and suggest that there are multiple pathways involved in resistance [1]. In the current study, we identified a novel set of genes in the mel2 locus that play a role in resistance to both ROS and RNS. As shown in our previous studies, this locus is also important for survival in activated macrophages and virulence in the mouse footpad model of infection [37]. To the best of our knowledge, this is the first description of a mycobacterial pathway that impacts susceptibility to both of these reactive species. Since RNS and ROS are linked through the production of peroxynitrite from nitric oxide and superoxide [1,46,47], the mel2 system may be specifically involved in resistance to this reactive species. The presence of the mel2 locus in the tuberculosis complex and M. marinum [36] and absence in avirulent mycobacteria that are more susceptible to peroxynitrite [27] supports this concept.
The similarity of the mel2 locus to bioluminescence systems at the amino acid level and the presence of conserved domains between them are intriguing observations. These data are particularly interesting in light of the recent observations that bioluminescent systems can protect cells against oxidative stress [28-32]. In search of a biological role for bioluminescence in bacteria that would explain how such an energy-consuming system could have developed evolutionarily, it has been proposed that these pathways protect against ROS generated in an aerobic atmosphere [35,54]. Interestingly, it has been observed that ROS play a pivotal role in host-symbiont interactions with bioluminescent bacteria [55]. At present, our model for the biochemical function of mel2 (Figure 2B) is purely hypothetical and is in need of more experimental support, but the large number of conserved functional domain similarities between the lux and mel2 loci suggests that they may have related functions. However, it seems unlikely that this function is bioluminescence, since mycobacteria are not normally bioluminescent and we did not observe any bioluminescence associated with our mutant or complemented strains (data not shown). Our observation that the mel2 locus plays a role in resistance to ROS helps to explain the presence of loci similar to bioluminescence genes in non-luminescent bacterial pathogens.
The inherent resistance of M. marinum to ROS is impacted by a mutation in the mel2 locus. This observation suggests that mel2 has an important role in either directly scavenging oxygen radicals or repairing damage caused by them. Since the mel2 mutant affects susceptibility to H2O2 and the organic peroxides cumene hydroperoxide and t-BOOH, which generate alkoxyl radicals, peroxyl radicals and H2O2 [44,45], it is unclear whether mel2 is specific to a particular type of ROS. The apparent absence of specificity could be the result of this pathway utilizing an unknown oxidizable substrate that is recycled, similar to luciferin in bioluminescent systems [35], direct scavenging of H2O2, which all three compounds produce, or repair of damaged DNA, proteins or lipids [6]. Interestingly, luciferase can produce light using H2O2 alone, in the absence of luciferin, suggesting that luciferase can scavenge H2O2, superoxide and hydroxyl radicals [33]. Overall, these data suggest that MelF functions as a FMN-dependent non-heme catalase. The presence of the mel2 locus in pathogenic mycobacteria may at least partially explain why the catalase (katG) gene can be mutated during acquisition of isoniazid resistance [56], yet katG negative M. tuberculosis are responsible for numerous clinical infections in humans [12-16]. Since oxidative stress increases susceptibility of mycobacteria to isoniazid [57], it is possible that in some cases there is a relationship between isoniazid susceptibility and the mel2 locus. This possibility can be tested by comparing the effects of a double and single katG and mel2 mutants on virulence and isoniazid resistance.
The role of bioluminescence systems from other bacteria in resistance to RNS has not been examined, but our observations with mel2 suggest that this possibility is worth investigating. Since susceptibility to both SNAP and acidified NaNO2 are impacted by the mel2 mutation, this phenotype is not the result of greater susceptibility to the acidic pH used with NaNO2. The fact that the mel2 mutant displays an obvious defect when exposed to a combination of both ROS and RNS would imply that this locus is important for growth in environments where both of these reactive species are present, such as during infection of mammals. We found that the M. marinum luxA homologue, melF, may play an important role in resistance to both RNS and ROS, since this gene alone can partially complement what may be a polar mutation. Alternatively, this observation could be the result of low levels of expression of the remainder of genes within mel2, as a result of the polar mutation. This polar mutation would allow only low levels of the putative Mel2 protein complex to be formed and provide partial complementation once a functional melF gene is expressed. A better understanding of the biochemical roles of each of the mel2 genes and their importance in susceptibility to ROS and RNS will require analysis of each gene individually as well as in the presence or absence of each of the different Mel2 components.
Conclusion
In this study, we confirmed that the mel2 locus plays a role in the susceptibility of M. marinum to ROS and RNS. Although this locus displays similarity to bioluminescent systems in other bacterial species, further biochemical studies are necessary to demonstrate the functional significance of the conserved domains that are present. These observations suggest that mel2 represents a previously unrecognized pathway for resistance of bacterial pathogens to ROS and RNS and support the concept that bioluminescence systems may have evolved from oxidative stress defense mechanisms.
Methods
Strains and growth conditions
M. marinum strain M, a clinical isolate obtained from the skin of a patient [58], was used in these studies. M. marinum strains were grown at 33°C in 7H9 broth (Difco, Detroit, Mich.) supplemented with 0.5% glycerol, 10% albumin-dextrose complex (ADC) and 0.25% Tween 80 (M-ADC-TW) for 5 days. M. smegmatis strain mc2155 [59] cultures were grown in M-ADC-TW for 3 days at 37°C and M. tuberculosis strain Erdman (ATCC35801) cultures were grown in M-ADC-TW for 10 days at 37°C. The number of viable bacteria was determined for each assay using the LIVE/DEAD assay (Molecular Probes, Eugene, OR.) and by plating dilutions for colony forming units (cfu) on 7H9 (M-ADC) agar (Difco, Detroit, Mich.). All inocula used were > 99% viable. E. coli strains were grown in Luria-Bertani (LB, Difco) media at 37°C. Where appropriate, kanamycin was added at a concentration of 25 μg/ml (E. coli) or 10 μg/ml (M. marinum).
Construction of M. marinum mel2 mutant and complementing strains
The M. marinum mel2 mutant carries a mini-Mu transposon insertion near the amino terminus of the melF gene as described previously [36]. Our previous studies have found no functional differences between the M. tuberculosis and M. marinum mel2 loci, both confer wild type host cell infection and growth in macrophages to the M. marinum melF insertion mutant [36,37], so either can be used for complementation studies. The M. marinum melF::pJDC79 strain is the melF mutant that carries the plasmid pMV262 [60] expressing the melF gene from M. tuberculosis that has been previously shown to complement the macrophage infection defect of the M. marinum mel2 mutant [36]. The M. marinum melF::pJDC75 strain is the melF mutant that carries the single-copy integrating plasmid pYUB178 [61] with the entire M. tuberculosis mel2 locus cloned into its single NheI site. Construction of all strains was confirmed by Southern analyses and PCR as described previously [36].
RT-PCR analyses
RT-PCR for the mel2 transcripts was performed using the ThermoScript RT-PCR System (Invitrogen) according to the manufacturer's instructions. Basically, 300–500 ng of DNase treated, total bacterial RNA was mixed with gene specific reverse primers, dNTP mix and 40U of RNaseOUT and incubated at 65°C for 5 min and then placed on ice prior to use. The annealed primers were extended with 15 U of Thermoscript RT at 55°C for 60 min followed by heat inactivation of the enzyme at 85°C for 5 min. The residual, non-transcribed RNA were removed with 2U of E. coli RNaseH at 37°C for 20 min. 2 μl of the cDNA was used in PCR amplification with 1 mM appropriate forward and reverse primers and 5U of Thermopol enzyme (NEB) in a total volume of 50 ul. All primers used for RT-PCR reactions are shown in Table 1. The concentrations of RNA in wild type and mutant strains were normalized against the respective 16s rRNA. 150 ng of M. marinum total genomic DNA of was used as positive control for the PCR reaction and RT-PCR reactions without reverse transcriptase was included in all experiments as negative control. The amplified products were analyzed by 0.8% agarose gel electrophoresis and the products measured by densitometry semiquantitatively using an Alpha Imager (Alpha Innotech) and Alpha Ease FC software.
Table 1.
Oligonucleotides
| Namea | Targetb | Sequence (5' -> 3') |
| MelFuF | melFu | CAGAAGACGCGATCACGGCG |
| MelFuR | GGGTCGGCGAACACTTCACC | |
| MelFdF | melFd | CCTGCTGCCCTATCATCACC |
| MelFdR | CCTCCAAAGCCCGAAGCCGC | |
| MelFtnF | melFtn | CAGAAGACGCGATCACGGCG |
| MelFtnR | CCTGCTGCCCTATCATCACC | |
| MelGF | melG | GCGAGAAGGGCACCGCCATG |
| MelGR | CTCAGATCACCCACGGTCAC | |
| MelHF | melH | GTGACCGTGGGTGATCTGAG |
| MelHR | GAAGGCGCAACTCACTGCCG | |
| MelIF | melI | CGGCAGTGAGTTGCGCCTTC |
| MelIR | CATCCAGGCTCCGTTGCGGG | |
| MelJF | melJ | CCCGCAACGGAGCCTGGATG |
| MelJR | GCCGCGAGTGGCGTGTCTGC | |
| MelKF | melK | GCAGACACGCCACTCGCGGC |
| MelKR | GACGCTCACCACAGTGCGGC | |
| rRNAF | rRNA | AGAGTTTGATCCTGGCTCAG |
| rRNAR | CACGCTCACAGTTAAGCTGT |
aDesignation for each oligonucleotide used in this study for RT-PCR analyses. F indicates a forward primer and R indicates a reverse primer.
bTarget gene mRNA transcript for RT-PCR analyses. Oligonucleotides are listed as primer pairs. The reverse primers were used for cDNA synthesis and the forward and reverse used together for RT-PCR. The melFu target is the region of the mel2 locus transcript upstream of the transposon insertion in the melF mutant but down stream of the start codon, melFd target is the region downstream of the transposon insertion but still within melF and melFtn is across the transposon insertion in melF.
In silico analysis of the melF-melK genes
Detailed analysis of the amino acid sequence of MelF-MelK was carried out initially using protein-protein National Center for Biotechnology Information (NCBI) BLAST [62] and Conserved Domain Search [63] as described previously [36]. Once motifs of interest were identified, they were compared to the appropriate bioluminescence genes and the mel2 gene and homologues were aligned and dendograms constructed using MegAlign (DNASTAR). Domain scores were considered significant if greater than 150 and the expectation values were less than 1 × 10-10.
Susceptibility to reactive oxygen species
Mycobacterial strains were exposed to ROS generated by H2O2, cumene hydroperoxide and tert-butyl hydroperoxide (t-BOOH). The susceptibility of mycobacteria to these compounds was determined by treatment for various periods of time at the appropriate growth temperature for the mycobacterial strain used and plating dilutions on M-ADC agar to determine CFU at each time point as compared to the original inoculum (To), i.e. percent survival = (CFU Tx/CFU To) × 100. Dimethyl sulfoxide (DMSO) was used as a solvent for t-BOOH and was tested for effects on viability of all mycobacterial strains and no solvent affected mycobacterial viability during the time periods examined or at the final concentrations used.
Susceptibility to reactive nitrogen species
Mycobacterial strains were exposed to RNS generated by S-nitroso-N-acetyl penicillamine (SNAP) and acidification of sodium nitrite (NaNO2) to pH 5.2 for various periods of time. Susceptibility was determined in the same manner as that described for ROS. DMSO was used as a solvent for SNAP and had no effects on viability of mycobacteria at the concentrations and time periods used.
Statistical analyses
All experiments were carried out in triplicate and repeated at least three times. The significance of the results was determined using the Student t-test. P values of < 0.05 were considered significant.
Authors' contributions
S.S. carried out the majority of these studies, participated in data analysis and participated in preparation of the manuscript. P.K.M. carried out some of the assays and participated in data analysis. S.L.G.C. carried out some of the assays and participated in data analysis. J.D.C. conceived the study, designed the experiments, completed the data analysis and prepared the final draft of the manuscript. All authors read the manuscript, participated in editing the manuscript and approved the final version.
Acknowledgments
Acknowledgements
This work was supported by grant AI47866 from the National Institutes of Health. We thank Drs. David McMurray and James Samuel for critical review of this manuscript.
Contributor Information
Selvakumar Subbian, Email: SSubbian@medicine.tamhsc.edu.
Parmod K Mehta, Email: pkmehta3@hotmail.com.
Suat LG Cirillo, Email: slcirillo@medicine.tamhsc.edu.
Jeffrey D Cirillo, Email: jdcirillo@medicine.tamhsc.edu.
References
- Nathan C, Shiloh MU. Reactive oxygen and nitrogen intermediates in the relationship between mammalian hosts and microbial pathogens. Proc Natl Acad Sci U S A. 2000;97:8841–8848. doi: 10.1073/pnas.97.16.8841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ng VH, Cox JS, Sousa AO, MacMicking JD, McKinney JD. Role of KatG catalase-peroxidase in mycobacterial pathogenesis: countering the phagocyte oxidative burst. Mol Microbiol. 2004;52:1291–1302. doi: 10.1111/j.1365-2958.2004.04078.x. [DOI] [PubMed] [Google Scholar]
- Senaratne RH, De Silva AD, Williams SJ, Mougous JD, Reader JR, Zhang T, Chan S, Sidders B, Lee DH, Chan J, Bertozzi CR, Riley LW. 5'-Adenosinephosphosulphate reductase (CysH) protects Mycobacterium tuberculosis against free radicals during chronic infection phase in mice. Mol Microbiol. 2006;59:1744–1753. doi: 10.1111/j.1365-2958.2006.05075.x. [DOI] [PubMed] [Google Scholar]
- Storz G, Toledano MB. Regulation of bacterial gene expression in response to oxidative stress. Methods Enzymol. 1994;236:196–207. doi: 10.1016/0076-6879(94)36017-0. [DOI] [PubMed] [Google Scholar]
- Christman MF, Storz G, Ames BN. OxyR, a positive regulator of hydrogen peroxide-inducible genes in Escherichia coli and Salmonella typhimurium, is homologous to a family of bacterial regulatory proteins. Proc Natl Acad Sci U S A. 1989;86:3484–3488. doi: 10.1073/pnas.86.10.3484. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imlay JA. Pathways of oxidative damage. Annu Rev Microbiol. 2003;57:395–418. doi: 10.1146/annurev.micro.57.030502.090938. [DOI] [PubMed] [Google Scholar]
- Seaver LC, Imlay JA. Alkyl hydroperoxide reductase is the primary scavenger of endogenous hydrogen peroxide in Escherichia coli. J Bacteriol. 2001;183:7173–7181. doi: 10.1128/JB.183.24.7173-7181.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dussurget O, Stewart G, Neyrolles O, Pescher P, Young D, Marchal G. Role of Mycobacterium tuberculosis copper-zinc superoxide dismutase. Infect Immun. 2001;69:529–533. doi: 10.1128/IAI.69.1.529-533.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piddington DL, Fang FC, Laessig T, Cooper AM, Orme IM, Buchmeier NA. Cu,Zn superoxide dismutase of Mycobacterium tuberculosis contributes to survival in activated macrophages that are generating an oxidative burst. Infect Immun. 2001;69:4980–4987. doi: 10.1128/IAI.69.8.4980-4987.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edwards KM, Cynamon MH, Voladri RKR, Hager C, DeStefano MS, Tham KT, Lakey DL, Bochan MR, Kernodle DS. Iron-cofactored superoxide dismutase inhibits host responses to Mycobacterium tuberculosis. Am J Respir Crit Care Med. 2001;164:2213–2219. doi: 10.1164/ajrccm.164.12.2106093. [DOI] [PubMed] [Google Scholar]
- Sherman DR, Sabo PJ, Hickey MJ, Arain TM, Mahairas GG, Yuan Y, Barry CE, 3rd, Stover CK. Disparate responses to oxidative stress in saprophytic and pathogenic mycobacteria. Proc Natl Acad Sci U S A. 1995;92:6625–6629. doi: 10.1073/pnas.92.14.6625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marttila HJ, Soini H, Huovinen P, Viljanen MK. katG mutations in isoniazid-resistant Mycobacterium tuberculosis isolates recovered from Finnish patients. Antimicrob Agents Chemother. 1996;40:2187–2189. doi: 10.1128/aac.40.9.2187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marttila HJ, Soini H, Eerola E, Vyshnevskaya E, Vyshnevskiy BI, Otten TF, Vasilyef AV, Viljanen MK. A Ser315Thr substitution in KatG is predominant in genetically heterogeneous multidrug-resistant Mycobacterium tuberculosis isolates originating from the St. Petersburg area in Russia. Antimicrob Agents Chemother. 1998;42:2443–2445. doi: 10.1128/aac.42.9.2443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Musser JM, Kapur V, Williams DL, Krieswirth BN, van Soolingen D, van Embden JD. Characterization of the catalase-peroxidase gene (katG) and inhA locus in isoniazid-resistant and -susceptible strains of Mycobacterium tuberculosis by automated DNA sequencing: restricted array of mutations associated with drug resistance. J Infect Dis. 1996;173:196–202. doi: 10.1093/infdis/173.1.196. [DOI] [PubMed] [Google Scholar]
- Nikolayevsky V, Brown T, Balabanova Y, Ruddy M, Fedorin I, Drobniewski F. Detection of mutations associated with isoniazid and rifampin resistance in Mycobacterium tuberculosis isolate from Samara Region, Russian Federation. J Clin Microbiol. 2004;42:4498–4502. doi: 10.1128/JCM.42.10.4498-4502.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramaswamy S, Musser JM. Molecular genetic basis of antimicrobial agent resistance in Mycobacterium tuberculosis: 1998 update. Tuber Lung Dis. 1998;79:3–29. doi: 10.1054/tuld.1998.0002. [DOI] [PubMed] [Google Scholar]
- Dhandayuthapani S, Zhang Y, Mudd MH, Deretic V. Oxidative stress response and its role in sensitivity to isoniazid in mycobacteria: characterization and inducibility of ahpC by peroxides in Mycobacterium smegmatis and lack of expression in M. aurum and M. tuberculosis. J Bacteriol. 1996;178:3641–3649. doi: 10.1128/jb.178.12.3641-3649.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Springer B, Master S, Sander P, Zahrt T, McFalone M, Song J, Papavinasasundaram KG, Colston MJ, Boettger E, Deretic V. Silencing of oxidative stress response in Mycobacterium tuberculosis: expression patterns of ahpC in virulent and avirulent strains and effect of ahpC inactivation. Infect Immun. 2001;69:5967–5973. doi: 10.1128/IAI.69.10.5967-5973.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrt S, Shiloh MU, Ruan J, Choi M, Gunzburg S, Nathan C, Xie Q, Riley L. A novel antioxidant gene from Mycobacterium tuberculosis. J Exp Med. 1997;186:1885–1896. doi: 10.1084/jem.186.11.1885. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ruan J, St. Joh G, Ehrt S, Riley L, Nathan C. noxR3, a novel gene from Mycobacterium tuberculosis, protects Salmonella typhimurium from nitrosative and oxidative stress. Infect Immun. 1999;67:3276–3283. doi: 10.1128/iai.67.7.3276-3283.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi S, Ehrt S. Dihydrolipoamide acyltransferase is critical for Mycobacterium tuberculosis pathogenesis. Infect Immun. 2006;74:56–63. doi: 10.1128/IAI.74.1.56-63.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- St John G, Brot N, Ruan J, Erdjument-Bromage H, Tempst P, Weissbach H, Nathan C. Peptide methionine sulfoxide reductase from Escherichia coli and Mycobacterium tuberculosis protects bacteria against oxidative damage from reactive nitrogen intermediates. Proc Natl Acad Sci U S A. 2001;98:9901–9906. doi: 10.1073/pnas.161295398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Douglas T, Daniel DS, Parida BK, Jagannath C, Dhandayuthapani S. Methionine sulfoxide reductase A (MsrA) deficiency affects the survival of Mycobacterium smegmatis within macrophages. J Bacteriol. 2004;186:3590–3598. doi: 10.1128/JB.186.11.3590-3598.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Darwin KH, Ehrt S, Gutierrez-Ramos JC, Weich N, Nathan CF. The proteasome of Mycobacterium tuberculosis is required for resistance to nitric oxide. Science. 2003;302:1963–1966. doi: 10.1126/science.1091176. [DOI] [PubMed] [Google Scholar]
- Master SS, Springer B, Sander P, Boettger EC, Deretic V, Timmins GS. Oxidative stress response genes in Mycobacterium tuberculosis: role of ahpC in resistance to peroxynitrite and stage-specific survival in macrophages. Microbiology. 2002;148:3139–3144. doi: 10.1099/00221287-148-10-3139. [DOI] [PubMed] [Google Scholar]
- McBride AG, Borutaite V, Brown GC. Superoxide dismutase and hydrogen peroxide cause rapid nitric oxide breakdown, peroxynitrite production and subsequent cell death. Biochim Biophys Acta. 1999;1454:275–288. doi: 10.1016/s0925-4439(99)00046-0. [DOI] [PubMed] [Google Scholar]
- Yu K, Mitchell C, Xing Y, Magliozzo RS, Bloom BR, Chan J. Toxicity of nitrogen oxides and related oxidants on mycobacteria: M. tuberculosis is resistant to peroxynitrite anion. Tuber Lung Dis. 1999;79:191–198. doi: 10.1054/tuld.1998.0203. [DOI] [PubMed] [Google Scholar]
- Szpilewska H, Czyz A, Wegrzyn G. Experimental evidence for the physiological role of bacterial luciferase in the protection of cells against oxidative stress. Curr Microbiol. 2003;47:379–382. doi: 10.1007/s00284-002-4024-y. [DOI] [PubMed] [Google Scholar]
- Barros MP, Bechara EJ. Bioluminescence as a possible auxiliary oxygen detoxifying mechanism in elaterid larvae. Free Radic Biol Med. 1998;24:767–777. doi: 10.1016/S0891-5849(97)00335-3. [DOI] [PubMed] [Google Scholar]
- Barros MP, Bechara EJ. Luciferase and urate may act as antioxidant defenses in larval Pyrearinus termitilluminans (Elateridae: Coleoptera) during natural development and upon 20-hydroxyecdysone treatment. Photochem Photobiol. 2000;71:648–654. doi: 10.1562/0031-8655(2000)071<0648:LAUMAA>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Barros MP, Bechara EJ. Daily variations of antioxidant enzyme and luciferase activities in the luminescent click-beetle Pyrearinus termitilluminans: cooperation against oxygen toxicity. Insect Biochem Mol Biol. 2001;31:393–400. doi: 10.1016/S0965-1748(00)00132-6. [DOI] [PubMed] [Google Scholar]
- Katsev AM, Wegrzyn G, Szpilewska H. Effects of hydrogen peroxide on light emission by various strains of marine luminescent bacteria. J Basic Microbiol. 2004;44:178–184. doi: 10.1002/jobm.200310330. [DOI] [PubMed] [Google Scholar]
- Watanabe H, Nagoshi T, Inaba H. Luminescence of a bacterial luciferase intermediate by reaction with H2O2: the evolutionary origin of luciferase and source of endogenous light emission. Biochim Biophys Acta. 1993;1141:297–302. doi: 10.1016/0005-2728(93)90056-L. [DOI] [Google Scholar]
- Lei X, Cho KW, Herndon ME, Tu SC. Elicitation of an oxidase activity in bacterial luciferase by site-directed mutation of a noncatalytic residue. J Biol Chem. 1990;265:4200–4203. [PubMed] [Google Scholar]
- Rees JF, De Wergifosse B, Noiset O, Dubuisson M, Janssens B, Thompson EM. The origins of marine bioluminescence: turning oxygen defence mechanisms into deep-sea communication tools. J Exp Biol. 1998;201:1211–1221. doi: 10.1242/jeb.201.8.1211. [DOI] [PubMed] [Google Scholar]
- El-Etr SH, Subbian S, Cirillo SL, Cirillo JD. Identification of two Mycobacterium marinum loci that affect interactions with macrophages. Infect Immun. 2004;72:6902–6913. doi: 10.1128/IAI.72.12.6902-6913.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subbian S, Mehta PK, Cirillo SL, Bermudez LE, Cirillo JD. A Mycobacterium marinum mel2 Mutant is Defective for Growth in Macrophages Producing Reactive Oxygen and Nitrogen Species. Infect Immun. 2007 doi: 10.1128/IAI.01000-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Escher A, O'Kane DJ, Szalay AA. The beta subunit polypeptide of Vibrio harveyi luciferase determines light emission at 42 degrees C. Mol Gen Genet. 1991;230:385–393. doi: 10.1007/BF00280295. [DOI] [PubMed] [Google Scholar]
- Swartzman E, Miyamoto C, Graham A, Meighen E. Delineation of the transcriptional boundaries of the lux operon of Vibrio harveyi demonstrates the presence of two new lux genes. J Biol Chem. 1990;265:3513–3517. [PubMed] [Google Scholar]
- Fisher AJ, Thompson TB, Thoden JB, Baldwin TO, Rayment I. The 1.5-A resolution crystal structure of bacterial luciferase in low salt conditions. J Biol Chem. 1996;271:21956–21968. doi: 10.1074/jbc.271.36.21956. [DOI] [PubMed] [Google Scholar]
- Thoden JB, Holden HM, Fisher AJ, Sinclair JF, Wesenberg G, Baldwin TO, Rayment I. Structure of the beta 2 homodimer of bacterial luciferase from Vibrio harveyi: X-ray analysis of a kinetic protein folding trap. Protein Sci. 1997;6:13–23. doi: 10.1002/pro.5560060103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li CH, Tu SC. Probing the functionalities of alphaGlu328 and alphaAla74 of Vibrio harveyi luciferase by site-directed mutagenesis and chemical rescue. Biochemistry. 2005;44:13866–13873. doi: 10.1021/bi051182i. [DOI] [PubMed] [Google Scholar]
- Li CH, Tu SC. Active site hydrophobicity is critical to the bioluminescence activity of Vibrio harveyi luciferase. Biochemistry. 2005;44:12970–12977. doi: 10.1021/bi050935y. [DOI] [PubMed] [Google Scholar]
- Alia M, Ramos S, Mateos R, Bravo L, Goya L. Response of the antioxidant defense system to tert-butyl hydroperoxide and hydrogen peroxide in a human hepatoma cell line (HepG2) J Biochem Mol Toxicol. 2005;19:119–128. doi: 10.1002/jbt.20061. [DOI] [PubMed] [Google Scholar]
- Simic MG, Bergtold DS, Karam LR. Generation of oxy radicals in biosystems. Mutat Res. 1989;214:3–12. doi: 10.1016/0027-5107(89)90192-9. [DOI] [PubMed] [Google Scholar]
- Koppenol WH, Moreno JJ, Pryor WA, Ischiropoulos H, Beckman JS. Peroxynitrite, a cloaked oxidant formed by nitric oxide and superoxide. Chem Res Toxicol. 1992;5:834–842. doi: 10.1021/tx00030a017. [DOI] [PubMed] [Google Scholar]
- Ischiropoulos H, Zhu L, Beckman JS. Peroxynitrite formation from macrophage-derived nitric oxide. Arch Biochem Biophys. 1992;298:446–451. doi: 10.1016/0003-9861(92)90433-W. [DOI] [PubMed] [Google Scholar]
- Chan J, Xing Y, Magliozzo RS, Bloom BR. Killing of virulent Mycobacterium tuberculosis by reactive nitrogen intermediates produced by activated murine macrophages. J Exp Med. 1992;175:1111–1122. doi: 10.1084/jem.175.4.1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan J, Tanaka K, Carroll D, Flynn J, Bloom BR. Effects of nitric oxide synthase inhibitors on murine infection with Mycobacterium tuberculosis. Infect Immun. 1995;63:736–740. doi: 10.1128/iai.63.2.736-740.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ehrt S, Schnappinger D, Bekiranov S, Drenkow J, Shi S, Gingeras TR, Gaasterland T, Schoolnik G, Nathan C. Reprogramming of the macrophage transcriptome in response to interferon-gamma and Mycobacterium tuberculosis: signaling roles of nitric oxide synthase-2 and phagocyte oxidase. J Exp Med. 2001;194:1123–1140. doi: 10.1084/jem.194.8.1123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- MacMicking JD, North RJ, LaCourse R, Mudgett JS, Shah SK, Nathan CF. Identification of nitric oxide synthase as a protective locus against tuberculosis. Proc Natl Acad Sci U S A. 1997;94:5243–5248. doi: 10.1073/pnas.94.10.5243. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fass U, Panickar K, Williams K, Nevels K, Personett D, McKinney M. The role of glutathione in nitric oxide donor toxicity to SN56 cholinergic neuron-like cells. Brain Res. 2004;1005:90–100. doi: 10.1016/j.brainres.2004.01.046. [DOI] [PubMed] [Google Scholar]
- Holm P, Kankaanranta H, Metsa-Ketela T, Moilanen E. Radical releasing properties of nitric oxide donors GEA 3162, SIN-1 and S-nitroso-N-acetylpenicillamine. Eur J Pharmacol. 1998;346:97–102. doi: 10.1016/S0014-2999(98)00009-0. [DOI] [PubMed] [Google Scholar]
- McElroy WD, Seliger HH. Origin and evolution of bioluminescence. In: Kasha M and Pullman B, editor. Horizons in biochemistry. New York, Academic Press; 1962. pp. 91–101. [Google Scholar]
- Ruby EG, McFall-Ngai MJ. Oxygen-utilizing reactions and symbiotic colonization of the squid light organ by Vibrio fischeri. Trends Microbiol. 1999;7:414–420. doi: 10.1016/S0966-842X(99)01588-7. [DOI] [PubMed] [Google Scholar]
- Cohn ML, Kovitz C, Oda U, Middlebrook G. Studies on isoniazid and tubercle bacilli. II. The growth requirements, catalase activities, and pathogenic properties of isoniazid-resistant mutants. Am Rev Tuberc. 1954;70:641–664. doi: 10.1164/art.1954.70.4.641. [DOI] [PubMed] [Google Scholar]
- Bulatovic VM, Wengenack NL, Uhl JR, Hall L, Roberts GD, Cockerill FR, 3rd, Rusnak F. Oxidative stress increases susceptibility of Mycobacterium tuberculosis to isoniazid. Antimicrob Agents Chemother. 2002;46:2765–2771. doi: 10.1128/AAC.46.9.2765-2771.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ramakrishnan L, Falkow S. Mycobacterium marinum persists in cultured mammalian cells in a temperature-restricted fashion. Infect Immun. 1994;62:3222–3229. doi: 10.1128/iai.62.8.3222-3229.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snapper SB, Melton RE, Mustafa S, Kieser T, Jacobs WR., Jr. Isolation and characterization of efficient plasmid transformation mutants of Mycobacterium smegmatis. Mol Microbiol. 1990;4:1911–1919. doi: 10.1111/j.1365-2958.1990.tb02040.x. [DOI] [PubMed] [Google Scholar]
- Stover CK, de la Cruz VF, Fuerst TR, Burlein JE, Benson LA, Bennet LT, Bansal GP, Young JF, Lee MH, Hatfull GF, Snapper SB, Barletta RG, Jacobs WR, Jr., Bloom BR. New use of BCG for recombinant vaccines. Nature. 1991;351:456–460. doi: 10.1038/351456a0. [DOI] [PubMed] [Google Scholar]
- Pascopella L, Collins FM, Martin JM, Lee MH, Hatfull GF, Stover CK, Bloom BR, Jacobs WR., Jr. Use of in vivo complementation in Mycobacterium tuberculosis to identify a genomic fragment associated with virulence. Infect Immun. 1994;62:1313–1319. doi: 10.1128/iai.62.4.1313-1319.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nuc Acids Res. 1997;25:3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marchler-Bauer A, Anderson JB, DeWeese-Scott C, Fedorova ND, Geer LY, He S, Hurwitz DI, Jackson JD, Jacobs AR, Lanczycki CJ, Liebert CA, Liu C, Madej T, Marchler GH, Mazumder R, Nikolskaya AN, Panchenko AR, Rao BS, Shoemaker BA, Simonyan V, Song JS, Thiessen PA, Vasudevan S, Wang Y, Yamashita RA, Yin JJ, Bryant SH. CDD: a curated Entrez database of conserved domain alignments. Nucleic Acids Res. 2003;31:383–387. doi: 10.1093/nar/gkg087. [DOI] [PMC free article] [PubMed] [Google Scholar]