

Abstract
This objective of this study was to investigate the toxicogenomics and the spatial regulation of global gene expression profiles elicited by Endoplasmic Reticulum (ER) stress inducer Tunicamycin (TM) in mouse small intestine and liver as well as to identify TM-modulated Nuclear Factor-E2-related factor 2 (Nrf2)–dependent genes. Gene expression profiles were analyzed using 45,000 Affymetrix mouse genome 430 2.0 array and GeneSpring 7.2 software. Microarray results were validated by quantitative real-time reverse transcription-PCR analyses. Clusters of genes that were either induced or suppressed more than two fold by TM treatment compared with vehicle in C57BL/6J/Nrf2(−/−; knockout)and C57BL/6J Nrf2 (+/+; wildtype) mice genotypes were identified. Amongst these, in small intestine and liver, 1291 and 750 genes respectively were identified as Nrf2-dependent and upregulated, and 1370 and 943 genes respectively as Nrf2-dependent and downregulated. Based on their biological functions, these genes can be categorized into molecular chaperones and heat shock proteins, ubiquitination/proteolysis, apoptosis/cell cycle, electron transport, detoxification, cell growth/differentiation, signaling molecules/interacting partners, kinases and phosphatases, transport, biosynthesis/metabolism, nuclear assembly and processing, and genes related to calcium and glucose homeostasis. Phase II detoxification/antioxidant genes as well as putative interacting partners of Nrf2 such as nuclear corepressors and coactivators, were also identified as Nrf2-dependent genes. The identification of TM-regulated and Nrf2-dependent genes in the unfolded protein response to ER stress not only provides potential novel insights into the gestalt biological effects of TM on the toxicogenomics and spatial regulation of global gene expression profiles in cancer pharmacology and toxicology, but also points to the pivotal role of Nrf2 in these biological processes.
Keywords: Tunicamycin, endoplasmic reticulum stress, Nuclear Factor-E2-related factor 2, microarray, global gene expression profiles
1. Introduction
The endoplasmic reticulum (ER) is an important organelle in which newly synthesized secretory and membrane-associated proteins destined to the extracellular space, plasma membrane, and the exo/endocytic compartments are correctly folded and assembled [1, 2]. An imbalance between the cellular demand for protein synthesis and the capacity of the ER in promoting protein maturation and transport can lead to an accumulation of unfolded or malfolded proteins in the ER lumen. This condition has been designated “ER stress” [2, 3]. Interestingly, the accumulation of misfolded protein in the ER triggers an adaptive stress response – termed the unfolded protein response (UPR) – mediated by the ER transmembrane protein kinase and endoribonuclease inositol-requiring enzyme-1α (IRE1α) [4]. The glucosamine-containing nucleoside antibiotic, Tunicamycin (TM, Fig.1), produced by genus Streptomyces, is an inhibitor of N-linked glycosylation and the formation of N-glycosidic protein-carbohydrate linkages [5]. It specifically inhibits dolichol pyrophosphate-mediated glycosylation of asparaginyl residues of glycoproteins [6] and induces “ER stress”.
Fig. 1.

Chemical Structure of Tunicamycin (TM).
Pivotal to the antioxidant response [7–10] typical in mammalian homeostasis and oxidative stress is the important transcription factor Nrf2 or Nuclear Factor-E2-related factor 2 that has been extensively studied by many research groups cited above as well as this laboratory [11–14]. Under homeostatic conditions, Nrf2 is mainly sequestered in the cytoplasm by a cytoskeleton-binding protein called Kelch-like erythroid CNC homologue (ECH)-associated protein 1 (Keap1) [11, 15, 16]. When challenged with oxidative stress, Nrf2 is quickly released from Keap1 retention and translocates to the nucleus [11, 17]. We have recently identified [11] a canonical redox-insensitive nuclear export signal (NES) (537LKKQLSTLYL546) located in the leucine zipper (ZIP) domain of the Nrf2 protein as well as a redox-sensitive NES (173LLSI-PELQCLNI186) in the transactivation (TA) domain of Nrf2 [18]. Once in the nucleus, Nrf2 not only binds to the specific consensus cis-element called antioxidant response element (ARE) present in the promoter region of many cytoprotective genes [12, 16, 19], but also to other trans-acting factors such as small Maf (MafG and MafK) [20] that can coordinately regulate gene transcription with Nrf2. We have previously reported [12] that different segments of Nrf2 transactivation domain have different transactivation potential; and that different MAPKs have differential effects on Nrf2 transcriptional activity, with ERK and JNK pathways playing an unequivocal role in positive regulation of Nrf2 transactivation domain activity. To better understand the biological basis of signaling through Nrf2, it has also become imperative to identify possible interacting partners of Nrf2 such as coactivators or corepressors apart from trans-acting factors such as small Maf.
Recently, it was reported [21] that Nrf1, another member of the Cap’ n’ Collar (CNC) family of basic leucine zipper proteins that is structurally similar to Nrf2, is normally targeted to the ER membrane, and that ER stress induced by TM in vitro may play a role in modulating Nrf1 function as a transcriptional activator. We sought to investigate the potential role of ER stress in modulating Nrf2 function as a transcriptional activator in vivo. Nrf2 knockout mice are greatly predisposed to chemical-induced DNA damage and exhibit higher susceptibility towards cancer development in several models of chemical carcinogenesis [19]. In the present study, we have investigated, by microarray expression profiling, the global gene expression profiles elicited by oral administration of TM in small intestine and liver of Nrf2 knockout (C57BL/6J/Nrf2−/−) and wild type (C57BL/6J) mice to enhance our understanding of TM-regulated toxicological effects mediated through Nrf2. We have identified clusters of TM-modulated genes that are Nrf2-dependent in small intestine and liver and categorized them based on their biological functions. The identification of TM-regulated Nrf2-dependent genes will yield valuable insights into the role of Nrf2 in TM-modulated gene regulation with respect to cancer pharmacology and toxicology. This study also enables the identification of novel molecular targets that are regulated by TM via Nrf2. The current study is also the first to investigate the global gene expression profiles elicited by TM in an in vivo murine model where the role of Nrf2 is also examined.
2.0. Materials and Methods
2.1. Animals and Dosing
The protocol for animal studies was approved by the Rutgers University Institutional Animal Care and Use Committee (IACUC). Nrf2 knockout mice Nrf2 (−/−) (C57BL/SV129) have been described previously.[22]. Nrf2 (−/−) mice were backcrossed with C57BL/6J mice (The Jackson Laboratory, ME USA). DNA was extracted from the tail of each mouse and genotype of the mouse was confirmed by polymerase chain reaction (PCR) by using primers (3′-primer, 5′-GGA ATG GAA AAT AGC TCC TGC C-3′; 5′-primer, 5′-GCC TGA GAG CTG TAG GCC C-3′; and lacZ primer, 5′-GGG TTT TCC CAG TCA CGA C-3′). Nrf2(−/−) mice-derived PCR products showed only one band of ~200bp, Nrf2 (+/+) mice-derived PCR products showed a band of ~300bp while both bands appeared in Nrf2(+/−) mice PCR products. Female C57BL/6J/Nrf2(−/−) mice from third generation of backcrossing were used in this study. Age-matched female C57BL/6J mice were purchased from The Jackson Laboratory (Bar Harbor, ME). Mice in the age-group of 9–12 weeks were housed at Rutgers Animal Facility with free access to water and food under 12 h light/dark cycles. After one week of acclimatization, the mice were put on AIN-76A diet (Research Diets Inc. NJ USA) for another week. The mice were then administered TM (Sigma-Aldrich, St.Louis, MO) at a dose of 2 mg/kg (dissolved in 50% PEG 400 aqueous solution) by oral gavage. The control group animals were administered only vehicle (50% PEG 400 aqueous solution). Each treatment was administrated to a group of four animals for both C57BL/6J and C57BL/6J/Nrf2(−/−) mice. Mice were sacrificed 3h after TM treatment or 3 h after vehicle treatment (control group). Livers and small intestines were retrieved and stored in RNA Later (Ambion, Austin,TX) solution.
2.2. Sample Preparation for Microarray Analyses
Total RNA from liver and small intestine tissues were isolated by using a method of TRIzol (Invitrogen, Carlsbad, CA) extraction coupled with the RNeasy kit from Qiagen (Valencia, CA). Briefly, tissues were homogenized in trizol and then extracted with chloroform by vortexing. A small volume (1.2 ml) of aqueous phase after chloroform extraction and centrifugation was adjusted to 35% ethanol and loaded onto an RNeasy column. The column was washed, and RNA was eluted following the manufacturer’s recommendations. RNA integrity was examined by electrophoresis, and concentrations were determined by UV spectrophotometry.
2.3. Microarray Hybridization and Data Analysis
Affymetrix (Affymetrix, Santa Clara, CA) mouse genome 430 2.0 array was used to probe the global gene expression profiles in mice following TM treatment. The mouse genome 430 2.0 Array is a high-density oligonucleotide array comprised of over 45,101 probe sets representing over 34,000 well-substantiated mouse genes. The library file for the above-mentioned oligonucleotide array is readily available at http://www.affymetrix.com/support/technical/libraryfilesmain.affx. After RNA isolation, all the subsequent technical procedures including quality control and concentration measurement of RNA, cDNA synthesis and biotin-labeling of cRNA, hybridization and scanning of the arrays, were performed at CINJ Core Expression Array Facility of Robert Wood Johnson Medical School (New Brunswick, NJ). Each chip was hybridized with cRNA derived from a pooled total RNA sample from four mice per treatment group, per organ, and per genotype (a total of eight chips were used in this study) (Fig.2). Briefly, double-stranded cDNA was synthesized from 5 μg of total RNA and labeled using the ENZO BioArray RNA transcript labeling kit (Enzo Life Sciences, Inc.,Farmingdale, NY, USA) to generate biotinylated cRNA. Biotin-labeled cRNA was purified and fragmented randomly according to Affymetrix’s protocol. Two hundred microliters of sample cocktail containing 15 μg of fragmented and biotin-labeled cRNA was loaded onto each chip. Chips were hybridized at 45°C for 16 h and washed with fluidics protocol EukGE-WS2v5 according to Affymetrix’s recommendation. At the completion of the fluidics protocol, the chips were placed into the Affymetrix GeneChip Scanner where the intensity of the fluorescence for each feature was measured. The expression value (average difference) for each gene was determined by calculating the average of differences in intensity (perfect match intensity minus mismatch intensity) between its probe pairs. The expression analysis file created from each sample (chip) was imported into GeneSpring 7.2 (Agilent Technologies, Inc., Palo Alto, CA) for further data characterization. Briefly, a new experiment was generated after importing data from the same organ in which data was normalized by array to the 50th percentile of all measurements on that array. Data filtration based on flags present in at least one of the samples was first performed, and a corresponding gene list based on those flags was generated. Lists of genes that were either induced or suppressed more than two fold between treated versus vehicle group of same genotype were created by filtration-on-fold function within the presented flag list. By use of color-by-Venn-Diagram function, lists of genes that were regulated more than two fold only in C57BL/6J mice in both liver and small intestine were created. Similarly, lists of gene that were regulated over two fold regardless of genotype were also generated.
Fig. 2.
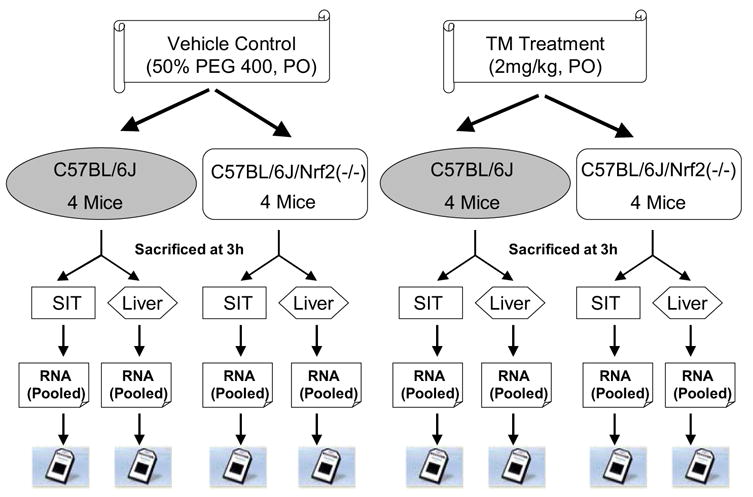
Schematic representation of experimental design; SIT, Small Intestine.
2.4. Quantitative Real-time PCR for Microarray Data Validation
To validate the microarray data, several genes of interest were selected from various categories for quantitative real-time PCR analyses. Glyceraldehyde-3-phosphate-dehydrogenase (GAPDH) served as the “housekeeping” gene. The specific primers for these genes listed in Table I were designed by using Primer Express 2.0 software (Applied Biosystems, Foster City, CA) and were obtained from Integrated DNA Technologies, Coralville, I A. The specificity of the primers was examined by a National Center for Biotechnology Information Blast search of the mouse genome. Instead of using pooled RNA from each group, RNA samples isolated from individual mice as described earlier were used in real-time PCR analyses. For the real-time PCR assays, briefly, first-strand cDNA was synthesized using 4μg of total RNA following the protocol of SuperScript III First-Strand cDNA Synthesis System (Invitrogen) in a 40 μl reaction volume. The PCR reactions based on SYBR Green chemistry were carried out using 100 times diluted cDNA product, 60 nM of each primer, and SYBR Green master mix (Applied Biosystems, Foster City, CA) in 10 μl reactions. The PCR parameters were set using SDS 2.1 software (Applied Biosystems, Foster City, CA) and involved the following stages : 50°C for 2min, 1 cycle; 95°C for 10 mins, 1 cycle; 95°C for 15 secs → 55 °C for 30 secs → 72°C for 30 secs, 40 cycles; and 72°C for 10 mins, 1 cycle. Incorporation of the SYBR Green dye into the PCR products was monitored in real time with an ABI Prism 7900HT sequence detection system, resulting in the calculation of a threshold cycle (CT) that defines the PCR cycle at which exponential growth of PCR products begins. The carboxy-X-rhodamine (ROX) passive reference dye was used to account for well and pipetting variability. A control cDNA dilution series was created for each gene to establish a standard curve. After conclusion of the reaction, amplicon specificity was verified by first-derivative melting curve analysis using the ABI software; and the integrity of the PCR reaction product and absence of primer dimers was ascertained. The gene expression was determined by normalization with control gene GAPDH. In order to validate the results, the correlation between corresponding microarray data and real-time PCR data was evaluated by the statistical ‘coefficient of determination’, r2=0.97.
Table I.
Representative oligonucleotide primers used in quantitative real-time PCR
| Gene Name | GenBank Accession No | Forward Primer | Reverse Primer |
|---|---|---|---|
| ATP-binding cassette, sub-family B (MDR/TAP),1A (Abcb1b) | NM_011075 | 5′-GAATGTCCAGTGGCTCCGA-3′ | 5′-CGGCTGTTGTCTCCATAGGC-3′ |
| ATP-binding cassette, sub-family C (CFTR/MRP), 1(Abcc1) | NM_008576 | 5′-CTCACGATTGCTCATCGGCT-3′ | 5′-AATCACCCGCGTGTAGTCCA-3′ |
| CASP8 and FADD-like apoptosis regulator (Cflar) | NM_207653 | 5′-CCAGCTTTTCTTGTTTCCCAAG-3′ | 5′-CGGCGAACAATCTGGGTTAT-3′ |
| Glutamate cysteine ligase, modifier subunit (Gclm) | NM_008129 | 5′-CGAGGAGCTTCGGGACTGTA-3′ | 5′-TGGTGCATTCCAAAACATCTG-3′ |
| Glutathione S-transferase, alpha 4 | NM_010357 | 5′-AGGAGTCATGGCAGCCAAAC-3′ | 5′-CCTCAAACTCCACTCCAGCC-3′ |
| Glutathione S-transferase, mu3 | NM_010359 | 5′-ATCCGCTTGCTCCTGGAATA-3′ | 5′-TTCTCACTCAGCCACTGGCTT-3′ |
| Inhibitor of kappaB kinase gamma (Ikbkg) | NM_010547 | 5′-CTGAAAGTTGGCTGCCATGAG-3′ | 5′-GAGTGGTGAGCTGGAGCAGG-3′ |
| Nuclear receptor coactivator 3 (Ncoa5) | NM-144892 | 5′-GAGGTGTCAGAGACGCCCAG-3′ | 5′-TTTCTTGTGGCCTTTGCTTTC-3′ |
| Nuclear receptor interacting protein 1 (Nrip1) | NM_173440 | 5′-AACAGTGAGCTGCCAACCCT-3′ | 5′-CTTCGGGACCATGCAGATGT-3′ |
| P300/CBP-associated factor (Pcaf) | NM_020005 | 5′-AGAGAGGCAGACAACGATCGA-3′ | 5′-TTGATGCGGTTCAGAAACATCT-3′ |
| Protein kinase C, epsilon (Prkce) | NM_011104 | 5′-ACGCTCCTATCGGCTACGAC-3′ | 5′-CGAACTGGATGGTGCAGTTG-3′ |
| Src family associated phosphoprotein 2 (Scap2) | NM_018773 | 5′- GCTGGCTACCTGGAAAAACG -3′ | 5′-TTCAAACCCCAGAAAGCTGTG-3′ |
| Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | NM_008084 | 5′-CACCAACTGCTTAGCCCCC-3′ | 5′-TCTTCTGGGTGGCAGTGATG-3′ |
3.0. Results
3.0.1. TM-Modulated Gene Expression Patterns in Mouse Small Intestine and Liver
Subsequent to data normalization, 48.76% (21,991) of the probes passed the filtration based on flags present in at least one of four small intestine sample arrays depicted in Figure 2. Expression levels of 1291 probes were elevated or of 1370 probes were suppressed over two fold by TM only in the wild-type mice, while 3471 probes were induced or 2024 probes were inhibited over two fold by TM only in the Nrf2(−/−) mice small intestine (Fig.3a). Similarly, changes in gene expression profiles were also observed in mice liver. Overall, the expression levels of 51.495% (23,225) probes were detected in least in one of four liver sample arrays depicted in Figure 2. In comparison with the results from small intestine sample arrays, a smaller proportion of well-defined genes were either elevated (750) or suppressed (943) over two fold by TM in wild-type mice liver alone; whereas 39 well-defined genes were induced or 3170 genes were inhibited in Nrf2(−/−) mice liver. (Fig.3b).
Fig. 3. Regulation of Nrf2-dependent gene expression by TM in mouse small intestine and liver.

Gene expression patterns were analyzed at 3h after administration of a 2mg/kg single oral dose of TM; Nrf2-dependent genes that were either induced or suppressed over two fold were listed. The positive numbers on the y-axis refer to the number of genes being induced; the negative numbers on the y-axis refer to the number of genes being suppressed.
3.0.2. Quantitative Real–Time PCR Validation of Microarray Data
To validate the data generated from the microarray studies, several genes from different categories (Table I) were selected to confirm the TM-regulative effects by the use of quantitative real-time PCR analyses as described in detail under Materials and Methods. After ascertaining the amplicon specificity by first-derivative melting curve analysis, the values obtained for each gene were normalized by the values of corresponding GAPDH expression levels. The fold changes in expression levels of treated samples over control samples were computed by assigning unit value to the control (vehicle) samples. Computation of the correlation statistic showed that the data generated from the microarray analyses are well-correlated with the results obtained from quantitative real-time PCR (coefficient of determination, r2 = 0.97, Fig.4).
Fig. 4. Correlation of microarray data with quantitative real-time PCR data.

Fold changes in gene expression measured by quantitative real-time PCR for each sample in triplicate (n=3) were plotted against corresponding fold changes from microarray data (coefficient of determination, r2 = 0.97).
3.0.3. TM-Induced Nrf2-Dependent Genes in Small Intestine and Liver
Genes that were induced only in wild-type mice, but not in Nrf2(−/−) mice, by TM were designated as TM-induced Nrf2-dependent genes. Based on their biological functions, these genes were classified into categories, including ubiquitination and proteolysis, electron transport, chaperones and unfolded protein response genes, detoxification enzymes, transport, apoptosis and cell cycle control, cell adhesion, kinases and phosphatases, transcription factors and interacting partners, glucose-related genes, ER and Golgi-related genes, translation factors, RNA/Protein processing and nuclear assembly, biosynthesis and metabolism, cell growth and differentiation, and G protein-coupled receptors (Table II lists genes relevant to our interest).
Table II .
TM-induced Nrf2-dependent genes in mouse small intestine and liver.
| GenBank Accession | Gene Symbol | Gene Title | SIT* | Liver** |
|---|---|---|---|---|
| Cell Adhesion | ||||
| NM_009864 | Cdh1 | Cadherin 1 | 6.77 | |
| XM_283264 | Cdh10 | cadherin 10 | 7.01 | |
| NM_007664 | Cdh2 | cadherin 2 | 9.72 | |
| XM_488510 | Cspg2 | chondroitin sulfate proteoglycan 2 | 2.72 | 2.82 |
| NM_009903 | Cldn4 | claudin 4 | 4.86 | |
| NM_018777 | Cldn6 | claudin 6 | 2.32 | |
| NM_031174 | Dscam | Down syndrome cell adhesion molecule (Dscam) | 2.25 | |
| NM_010103 | Edil3 | EGF-like repeats and discoidin I-like domains 3 | 9.2 | |
| NM_008401 | Itgam | integrin alpha M | 2.42 | |
| NM_008405 | Itgb2l | integrin beta 2-like | 9.64 | |
| Jam3 | Junction adhesion molecule 3 | 2.2 | ||
| NM_007736 | Col4a5 | procollagen, type IV, alpha 5 | 2.54 | |
| XM_139187 | Pcdh9 | protocadherin 9 | 2.33 | |
| Apoptosis and Cell cycle control | ||||
| XM_194020 | Acvr1c | activin A receptor, type IC | 26.49 | |
| NM_178655 | Ank2 | ankyrin 2, brain | 17.4 | |
| NM_153287 | Axud1 | AXIN1 up-regulated 1 | 2.71 | |
| Bcl2 | B-cell leukemia/lymphoma 2 (Bcl2), transcript variant 1 | 2.68 | ||
| NM_009744 | Bcl6 | B-cell leukemia/lymphoma 6 | 2.02 | |
| NM_207653 | Cflar | CASP8 and FADD-like apoptosis regulator | 2.12 | |
| NM_026373 | Cdk2ap2 | CDK2-associated protein 2 | 2.35 | |
| XM_484088 | Cdc27 | cell division cycle 27 homolog (S. cerevisiae) | 2.36 | |
| NM_009862 | Cdc45l | cell division cycle 45 homolog (S. cerevisiae)-like | 3.38 | |
| NM_026201 | Ccar1 | cell division cycle and apoptosis regulator 1 | 9.84 | |
| NM_013538 | Cdca3 | cell division cycle associated 3 | 2.18 | |
| NM_011806 | Dmtf1 | cyclin D binding myb-like transcription factor 1 | 2.88 | |
| NM_028399 | Ccnt2 | cyclin T2 | 9.26 | |
| NM_009874 | Cdk7 | cyclin-dependent kinase 7 (homolog of Xenopus MO15 cdk-activating kinase) | 15.94 | |
| NM_007837 | Ddit3 | DNA-damage inducible transcript 3 | 13.72 | 9 |
| NM_007950 | Ereg | epiregulin | 5.85 | |
| NM_008087 | Gas2 | Growth arrest specific 2 | 2.01 | |
| XM_137276 | Gas2l3 | growth arrest-specific 2 like 3 | 4.26 | |
| NM_146071 | Muc20 | mucin 20 | 2.96 | |
| NM_009044 | Rel | reticuloendotheliosis oncogene | 2.35 | |
| NM_133810 | Stk17b | serine/threonine kinase 17b (apoptosis-inducing) | 2.27 | |
| NM_028769 | Syvn1 | synovial apoptosis inhibitor 1, synoviolin | 4.77 | |
| NM_021897 | Trp53inp1 | transformation related protein 53 inducible nuclear protein 1 | 2.49 | |
| Biosynthesis and Metabolism | ||||
| --- | Acyl-CoA synthetase long-chain family member 5 | 17.14 | ||
| NM_029901 | Akr1c21 | aldo-keto reductase family 1, member C21 | 2.14 | |
| NM_023179 | Atp6v1g2 | ATPase, H+ transporting, V1 subunit G isoform 2 | 2.23 | |
| 1451144_at | Bxdc2 | brix domain containing 2 | 2.19 | |
| NM_023525 | Cad | carbamoyl-phosphate synthetase 2, aspartate transcarbamylase, and dihydroorotase | 2.4 | |
| NM_198415 | Ckmt2 | creatine kinase, mitochondrial 2 | 29.85 | |
| NM_007710 | Ckm | creatine kinase, muscle | 21.08 | |
| NM_030225 | Dlst | dihydrolipoamide S-succinyltransferase (E2 component of 2-oxo-glutarate complex) | 3.28 | |
| NM_021896 | Gucy1a3 | guanylate cyclase 1, soluble, alpha 3 | 5.6 | |
| NM_011846 | Mmp17 | matrix metallopeptidase 17 | 24.22 | |
| NM_138656 | Mvd | mevalonate (diphospho) decarboxylase | 3.46 | |
| NM_009127 | Scd1 | stearoyl-Coenzyme A desaturase 1 | 2.26 | |
| Calcium homeostasis | ||||
| NM_013471 | Anxa4 | Annexin A4 | 5.2 | |
| NM_009722 | Atp2a2 | ATPase, Ca++ transporting, cardiac muscle, slow twitch 2 | 2.65 | |
| NM_023116 | Cacnb2 | Calcium channel, voltage-dependent, beta 2 subunit | 14.13 | |
| NM_009781 | Cacna1c | Calcium channel, voltage-dependent, L type, alpha 1C subunit | 7.94 | |
| NM_028231 | Kcnmb2 | potassium large conductance calcium-activated channel, subfamily M, beta member 2 | 4.62 | |
| Cell Growth and Differentiation | ||||
| NM_010111 | Efnb2 | ephrin B2 | 2.67 | |
| NM_177390 | Myo1d | Myosin ID | 2.68 | |
| NM_145610 | Ppan | peter pan homolog (Drosophila) | 2.04 | |
| NM_021883 | Tmod1 | tropomodulin 1 | 2.7 | |
| NM_009394 | Tnnc2 | troponin C2, fast | 16.76 | |
| ER/Golgi transport and ER/Golgi biosynthesis/metabolism | ||||
| NM_025445 | Arfgap3 | ADP-ribosylation factor GTPase activating protein 3 | 2.58 | |
| NM_025505 | Blzf1 | basic leucine zipper nuclear factor 1 | 7.72 | |
| NM_009938 | Copa | coatomer protein complex subunit alpha | 2.4 | |
| NM_025673 | Golph3 | Golgi phosphoprotein 3 | 2.57 | |
| NM_146133 | Golph3l | golgi phosphoprotein 3-like | 2.41 | |
| NM_008408 | Itm1 | intergral membrane protein 1 | 6.62 | 2.56 |
| NM_027400 | Lman1 | Lectin, mannose-binding | 2.79 | |
| NM_025408 | Phca | phytoceramidase, alkaline | 3.04 | |
| NM_009178 | Siat4c | ST3 beta-galactoside alpha-2,3-sialyltransferase 4 | 2.4 | |
| NM_020283 | B3galt1 | UDP-Gal:betaGlcNAc beta 1,3-galactosyltransferase, polypeptide 1 | 2.54 | |
| NM_011716 | Wfs1 | Wolfram syndrome 1 homolog (human) | 2.02 | |
| Electron Transport | ||||
| NM_015751 | Abce1 | ATP-binding cassette, sub-family E (OABP), member 1 | 2.48 | |
| NM_010001 | Cyp2c37 | cytochrome P450, family 2. subfamily c, polypeptide 37 | 5.58 | |
| NM_023913 | Ern1 | Endoplasmic reticulum (ER) to nucleus signalling 1 | 2.12 | |
| XM_129326 | Gucy2g | guanylate cyclase 2g | 2.39 | |
| NM_007952 | Pdia3 | protein disulfide isomerase associated 3 | 3.11 | |
| NM_009787 | Pdia4 | protein disulfide isomerase associated 4 | 3.19 | |
| XM_907880 | Pdia6 | protein disulfide isomerase associated 6 | 2.9 | |
| XM_284053 | Steap2 | six transmembrane epithelial antigen of prostate 2 | 3.23 | |
| NM_198295 | 730024F05Ri | Thioredoxin domain containing 10 | 2.91 | |
| NM_029572 | Txndc4 | thioredoxin domain containing 4 (endoplasmic reticulum) | 2.47 | |
| NM_023140 | Txnl2 | Thioredoxin-like 2 | 8.25 | |
| G-protein coupled receptors | ||||
| NM_008158 | Gpr27 | G protein-coupled receptor 27 | 2.72 | |
| NM_145066 | Gpr85 | G protein-coupled receptor 85 | 3.78 | |
| AK015353 | Grm2 | G protein-coupled receptor, family C, group 1, member B | 2.18 | |
| NM_008177 | Grpr | gastrin releasing peptide receptor | 2.18 | |
| NM_010314 | Gngt1 | guanine nucleotide binding protein (G protein), gamma transducing activity polypeptide 1 | 2.02 | |
| NM_139270 | Pthr2 | parathyroid hormone receptor 2 | 2.12 | |
| NM_011056 | Pde4d | phosphodiesterase 4D, cAMP specific | 2.78 | |
| NM_022881 | Rgs18 | regulator of G-protein signaling 18 | 2.36 | |
| Kinases and Phosphatases | ||||
| NM_153066 | Ak5 | adenylate kinase 5 | 2.33 | |
| NM_144817 | Camk1g | calcium/calmodulin-dependent protein kinase I gamma | 2.32 | |
| NM_139059 | Csnk1d | Casein kinase 1, delta (Csnk1d), transcript variant 2 | 2.08 | |
| NM_177914 | MGI:3580254 | diacylglycerol kinase kappa | 13.2 | |
| NM_130447 | Dusp16 | dual specificity phosphatase 16 | 3.17 | 2.13 |
| NM_019987 | Ick | intestinal cell kinase | 2.06 | |
| XM_283179 | Mast4 | microtubule associated serine/threonine kinase family member 4 | 3.62 | |
| NM_016700 | Mapk8 | mitogen activated protein kinase 8 | 12.43 | |
| Mapk8 | mitogen activated protein kinase 8 | 7.41 | ||
| NM_172688 | Map3k7 | mitogen activated protein kinase kinase kinase 7 | 3.29 | |
| NM_011101 | Prkca | Protein kinase C, alpha | 2.04 | |
| NM_011104 | Prkce | protein kinase C, epsilon | 3.3 | |
| NM_021880 | Prkar1a | protein kinase, cAMP dependent regulatory, type I, alpha | 15.23 | |
| NM_175638 | Prkwnk4 | Protein kinase, lysine deficient 4 | 8.16 | |
| NM_016979 | Prkx | protein kinase, X-linked | 2.27 | |
| NM_133485 | Ppp1r14c | Protein phosphatase 1, regulatory (inhibitor) subunit 14c | 5.03 | |
| NM_012024 | Ppp2r5e | protein phosphatase 2, regulatory subunit B (B56), epsilon isoform | 2.49 | |
| NM_008913 | Ppp3ca | Protein phosphatase 3, catalytic subunit, alpha isoform | 14.5 | |
| AK134422 | Ptp | Protein tyrosine phosphatase | 3.27 | |
| NM_028259 | Rps6kb1 | ribosomal protein S6 kinase, polypeptide 1 | 2.05 | |
| NM_031880 | Tnk1 | tyrosine kinase, non-receptor, 1 | 2.54 | |
| Nuclear Assembly and Processing | ||||
| NM_010613 | Khsrp | KH-type splicing regulatory protein | 2.44 | |
| NM_008671 | Nap1l2 | nucleosome assembly protein 1-like 2 | 4.7 | |
| NM_026175 | Sf3a1 | splicing factor 3a, subunit 1 | 2.77 | |
| NM_009408 | Top1 | Topoisomerase (DNA) I | 5.94 | |
| NM_008717 | Zfml | Zinc finger, matrin-like | 2.19 | |
| Glucose biosynthesis/metabolism | ||||
| NM_009605 | Adipoq | adiponectin, C1Q and collagen domain containing | 2.23 | |
| NM_018763 | Chst2 | Carbohydrate sulfotransferase 2 | 2.02 | |
| NM_008079 | Galc | galactosylceramidase | 2.95 | |
| NM_029626 | Glt8d1 | glycosyltransferase 8 domain containing 1 | 2.17 | |
| NM_013820 | Hk2 | hexokinase 2 | 2.46 | |
| NM_010705 | Lgals3 | Lectin, galactose binding, soluble 3 | 3.2 | |
| NM_199446 | Phkb | phosphorylase kinase beta | 2.06 | |
| NM_016752 | Slc35b1 | solute carrier family 35, member B1 | 2.13 | |
| Signaling molecules and interacting partners | ||||
| NM_029291 | Ascc2 | Activating signal cointegrator 1 complex subunit 2 | 3.24 | |
| NM_007498 | Atf3 | activating transcription factor 3 | 8.73 | |
| NM_016707 | Bcl11a | B-cell CLL/lymphoma 11A (zinc finger protein) | 4.2 | |
| NM_033601 | Bcl3 | B-cell leukemia/lymphoma 3 | 2.08 | |
| NM_007553 | Bmp2 | bone morphogenetic protein 2 | 2.55 | |
| NM_007558 | Bmp8a | bone morphogenetic protein 8a | 2.39 | |
| NM_178661 | Creb3l2 | cAMP responsive element binding protein 3-like 2 | 2.01 | |
| NM_010016 | Daf1 | decay accelerating factor 1 | 2.29 | |
| NM_007897 | Ebf1 | early B-cell factor 1 | 8.98 | |
| NM_023580 | Epha1 | Eph receptor A1 | 2.037 | |
| NM_133753 | Errfi1 | ERBB receptor feedback inhibitor 1 | 2.53 | |
| NM_0010058 | Erbb2ip | Erbb2 interacting protein | 2.11 | |
| NM_0010058 | Erbb2ip | Erbb2 interacting protein | 2.04 | |
| NM_007906 | Eef1a2 | eukaryotic translation elongation factor 1 alpha 2 | 3.67 | |
| NM_007917 | Eif4e | eukaryotic translation initiation factor 4E | 3.05 | |
| NM_173363 | Eif5 | eukaryotic translation initiation factor 5 | 2.13 | |
| NM_010515 | Igf2r | Insulin-like growth factor 2 receptor | 2.22 | |
| NM_010591 | Jun | Jun oncogene | 2.29 | |
| NM_010592 | Jund1 | Jun proto-oncogene related gene d1 | 2.43 | |
| NM_008416 | Junb | Jun-B oncogene | 2.36 | |
| NM_013602 | Mt1 | metallothionein 1 | 2.15 | |
| NM_008630 | Mt2 | metallothionein 2 | 2.79 | |
| NM_170671 | Mycbpap | Mycbp associated protein | 3.97 | |
| NM_177619 | Myst2 | MYST histone acetyltransferase 2 | 2 | |
| NM_009123 | Nkx1-2 | NK1 transcription factor related, locus 2 (Drosophila) | 2.45 | |
| NM_030612 | Nfkbiz | nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, zeta | 2.67 | |
| NM_017373 | Nfil3 | nuclear factor, interleukin 3, regulated | 12.68 | 3.34 |
| NM_144892 | Ncoa5 | nuclear receptor coactivator 5 | 14.65 | |
| NM_173440 | Nrip1 | nuclear receptor interacting protein 1 | 2.87 | |
| BC032981 | Nfxl1 | nuclear transcription factor, X-box binding-like 1 | 3.13 | |
| NM_020005 | Pcaf | P300/CBP-associated factor | 2.33 | |
| NM_027924 | Pdgfd | platelet-derived growth factor, D polypeptide | 6.17 | |
| NM_017463 | Pbx2 | pre B-cell leukemia transcription factor 2 | 2.84 | |
| NM_026383 | Pnrc2 | proline-rich nuclear receptor coactivator 2 | 2.08 | |
| NM_145495 | Rin1 | Ras and Rab interactor 1 | 7.01 | 2.87 |
| NM_011651 | Stk22s1 | serine/threonine kinase 22 substrate 1 | 2.36 | |
| NM_175246 | Snip1 | Smad nuclear interacting protein 1 | 2.07 | |
| NM_007707 | Socs3 | suppressor of cytokine signaling 3 | 2.45 | |
| NM_080843 | Socs4 | suppressor of cytokine signaling 4 | 2 | |
| NM_009365 | Tgfb1i1 | transforming growth factor beta 1 induced transcript 1 | 2.22 | |
| NM_0010130 | Tgfbrap1 | transforming growth factor, beta receptor associated protein 1 | 2.7 | |
| NM_013869 | Tnfrsf19 | tumor necrosis factor receptor superfamily, member 19 | 2.4 | |
| NM_010755 | Maff | v-maf musculoaponeurotic fibrosarcoma oncogene family, protein F (avian) | 2.92 | |
| NM_009524 | Wnt5a | wingless-related MMTV integration site 5A | 8.25 | |
| Transport | ||||
| NM_007511 | Atp7b | ATPase, Cu++ transporting, beta polypeptide | 2.34 | |
| NM_011075 | Abcb1b | ATP-binding cassette, sub-family B (MDR/TAP), member 1B | 4.65 | |
| NM_008576 | Abcc1 | ATP-binding cassette, sub-family C (CFTR/MRP), member 1 | 2.37 | |
| NM_172621 | Clic5 | Chloride intracellular channel 5, mRNA | 2.39 | |
| NM_024406 | Fabp4 | Fatty acid binding protein 4, adipocyte | 3.7 | |
| NM_146188 | Kctd15 | potassium channel tetramerisation domain containing 15 | 2.82 | |
| Kctd7 | potassium channel tetramerisation domain containing 7 | 2.65 | ||
| NM_148938 | Slc1a3 | solute carrier family 1 (glial high affinity glutamate transporter), member 3 | 4.03 | |
| NM_019481 | Slc13a1 | solute carrier family 13 (sodium/sulphate symporters), member 1 | 2.09 | |
| NM_0010041 | Slc13a5 | solute carrier family 13 (sodium-dependent citrate transporter), member 5 | 6.88 | |
| NM_011395 | Slc22a3 | solute carrier family 22 (organic cation transporter), member 3 | 2.07 | |
| NM_172980 | Slc28a2 | solute carrier family 28 (sodium-coupled nucleoside transporter), member 2 | 2 | |
| NM_078484 | Slc35a2 | solute carrier family 35 (UDP-galactose transporter), member 2 | 6.5 | |
| NM_011990 | Slc7a11 | solute carrier family 7 (cationic amino acid transporter, y+ system), member 11 | 5.93 | |
| NM_080852 | Slc7a12 | solute carrier family 7 (cationic amino acid transporter, y+ system), member 12 | 2.95 | |
| NM_011406 | Slc8a1 | Solute carrier family 8 (sodium/calcium exchanger), member 1 | 2.86 | |
| NM_178892 | Tiparp | TCDD-inducible poly(ADP-ribose) polymerase | 2.27 | |
| Ubiquitination and Proteolysis | ||||
| NM_027926 | Cpa4 | carboxypeptidase A4 | 2.29 | |
| NM_011931 | Cop1 | Constitutive photomorphogenic protein | 2.52 | |
| NM_013868 | Hspb7 | heat shock protein family, member 7 (cardiovascular) | 2.25 | |
| NM_146042 | Ibrdc2 | IBR domain containing 2 | 6.77 | |
| NM_009174 | Siah2 | seven in absentia 2 | 2.46 | |
| Siah2 | seven in absentia 2 | 2.12 | ||
| NM_025692 | Ube1dc1 | ubiquitin-activating enzyme E1-domain containing 1 | 2.79 | |
| NM_173443 | Vcpip1 | valosin containing protein (p97)/p47 complex interacting protein 1 | 2.12 | |
| Molecular chaperones and Heat Shock Proteins | ||||
| NM_0010124 | Hspb6 | heat shock protein, alpha-crystallin-related, B6 | 2.13 | |
| NM_010918 | Nktr | natural killer tumor recognition sequence | 2.02 | |
| NM_030201 | Stch | stress 70 protein chaperone, microsome-associated, human homolog | 2.47 | |
| Stch | stress 70 protein chaperone, microsome-associated, human homolog | 2.37 | ||
| Miscellaneous | ||||
| NM_008161 | Gpx3 | glutathione peroxidase 3 | 5.79 | |
| NM_028733 | Pacsin3 | Protein kinase C and casein kinase II substrate 3 (Pacsin3) | 2.12 | |
| NM_009409 | Top2b | Topoisomerase (DNA) II beta (Top2b), mRNA | 3.39 | |
| NM_020283 | B3galt1 | UDP-Gal:betaGlcNAc beta 1,3-galactosyltransferase, polypeptide 1 | 2.45 | |
Genes that were induced >2-fold by TM only in small intestine of Nrf2 wild-type mice but not in small intestine of Nrf2 knockout mice compared with vehicle treatment at 3h. The relative mRNA expression levels of each gene in treatment group over vehicle group (fold changes) are listed.
Genes that were induced >2-fold by TM only in liver of Nrf2 wild-type mice but not in liver of Nrf2 knockout mice compared with vehicle treatment at 3h. The relative mRNA expression levels of each gene in treatment group over vehicle group (fold changes) are listed.
In response to TM-induced ER stress, several unfolded protein response genes were identified as Nrf2-regulated including, amongst others, heat shock protein, alpha-crystallin-related, B6 (Hspb6) in liver, heat shock protein family, member 7,cardiovascular (Hspb7) in small intestine, and stress 70 protein chaperone, microsome-associated, human homolog (Stch) in both liver and small intestine. A large number of apoptosis and cell-cycle related genes were also upregulated in response to TM treatment. Representative members included B-cell leukemia/lymphoma 2 (Bcl2), CASP8 and FADD-like apoptosis regulator (Cflar), Epiregulin (Ereg), Growth arrest specific 2 (Gas2) and synovial apoptosis inhibitor 1, synoviolin (Syvn1). Interestingly, several important transcription/translation factors and interacting partners were identified as Nrf2-dependent and TM-regulated. These included P300/CBP-associated factor (Pcaf), Smad nuclear interacting protein 1 (Snip1), nuclear receptor coactivator 5 (Ncoa5), nuclear receptor interacting protein 1 (Nrip1), nuclear transcription factor, X-box binding-like 1 (Nfxl1), eukaryotic translation initiation factors 1α 2, 4e and 5 (Eif 1a2, 4e and 5), Erbb2 interacting protein (Erbb2ip), cAMP responsive element binding protein 3-like 2 (Creb3l2) and Jun oncogene (Jun).
Other categories of genes induced by TM in an Nrf2-dependent manner included cell adhesion (cadherins 1, 2, and 10), glucose-related genes (hexokinase 2), transport (solute carrier family members Slc13a1, Slc22a3, Slc8a1 and others), and ubiquitination and proteolysis (Constitutive photomorphogenic protein and carboxypeptidase A4). The glutathione peroxidase 3 (Gpx3) gene was also upregulated in liver in an Nrf2-dependent manner in response to TM treatment.
3.0.4. TM-Suppressed Nrf2-Dependent Genes in Small Intestine and Liver
As shown in Table III which lists genes relevant to our interest, TM treatment also inhibited the expression of many genes falling into similar functional categories in an Nrf2-dependent manner. Major Phase II detoxifying genes identified as Nrf2-regulated and TM-modulated included several isoforms of Glutathione-S-transferase (Gst), and glutamate cysteine ligase, modifier subunit (Gclm). Additionally, Phase I genes such as cytochrome P450 family members Cyp3a44, Cyp39a1 and Cyp8b1 were also downregulated in response to TM-treatment in an Nrf2-dependent manner. Moreover, many transport genes, which may be regarded as Phase III genes, including members of solute carrier family (Slc23a2, Slc23a1, Slc37a4, Slc4a4, Slc40a1, Slc9a3) and multidrug-resistance associated proteins (Abcc3) were also downregulated via Nrf2 and regulated through TM. Thus, a co-ordinated response involving Phase I, II and III genes was observed on TM treatment in an Nrf2-dependent manner.
Table III.
TM-suppressed Nrf2-dependent genes in mouse small intestine and liver.
| GenBank Accession | Gene Symbo | Gene Title | SIT* | Liver** |
|---|---|---|---|---|
| Cell Adhesion | ||||
| NM _174988 | Cdh22 | cadherin 22 | 0.47 | |
| NM_053096 | Cml2 | camello-like 2 | 0.45 | |
| NM_009818 | Catna1 | Catenin (cadherin associated protein), alpha 1 | 0.13 | |
| NM_008729 | Catnd2 | Catenin (cadherin associated protein), delta 2 | 0.5 | |
| XM_488510 | Cspg2 | chondroitin sulfate proteoglycan 2 | 0.43 | |
| NM_018764 | Pcdh7 | protocadherin 7 | 0.21 | |
| NM_053134 | Pcdhb9 | protocadherin beta 9 | 0.28 | |
| NM_033595 | Pcdhga12 | Protocadherin gamma subfamily A, 10, mRNA | 0.4 | |
| Apoptosis and Cell cycle control | ||||
| NM_007566 | Birc6 | baculoviral IAP repeat-containing 6 | 0.49 | |
| NM_009741 | Bcl2 | B-cell leukemia/lymphoma 2 | 0.25 | |
| NM_009950 | Cradd | CASP2 and RIPK1 domain containing adaptor with death domain | 0.43 | |
| NM_007609 | Casp11 | caspase 11, apoptosis-related cysteine peptidase | 0.45 | |
| NM_009811 | Casp6 | caspase 6 | 0.36 | |
| NM_025680 | Ctnnbl1 | catenin, beta like 1 | 0.45 | |
| NM_025866 | Cdca7 | cell division cycle associated 7 | 0.36 | |
| NM_026560 | Cdca8 | cell division cycle associated 8 | 0.37 | |
| XM_181420 | Cgref1 | cell growth regulator with EF hand domain 1 | 0.47 | |
| NM_009131 | Clec11a | C-type lectin domain family 11, member a | 0.2 | |
| NM_146207 | Cul4a | cullin 4A | 0.49 | |
| NM_009873 | Cdk6 | cyclin-dependent kinase 6 | 0.47 | |
| NM_009876 | Cdkn1c | cyclin-dependent kinase inhibitor 1C (P57) | 0.48 | |
| NM_007892 | E2f5 | E2F transcription factor 5 | 0.25 | |
| NM_008655 | Gadd45b | growth arrest and DNA-damage-inducible 45 beta | 0.18 | |
| NM_183358 | Gadd45gip1 | growth arrest and DNA-damage-inducible, gamma interacting protein 1 | 0.45 | |
| NM_010578 | Itgb1 | integrin beta 1 (fibronectin receptor beta) | 0.12 | |
| NM_019745 | Pdcd10 | programmed cell death 10 | 0.46 | |
| NM_009383 | Tial1 | Tial1 cytotoxic granule-associated RNA binding protein-like 1 | 0.07 | 0.36 |
| NM_009425 | Tnfsf10 | tumor necrosis factor (ligand) superfamily, member 10 | 0.33 | |
| NM_009517 | Wig1 | wild-type p53-induced gene 1 | 0.5 | |
| Biosynthesis and Metabolism | ||||
| NM_177470 | Acaa2 | acetyl-Coenzyme A acyltransferase 2 (mitochondrial 3-oxoacyl-Coenzyme A thiolase) | 0.49 | |
| NM_133904 | Acacb | Acetyl-Coenzyme A carboxylase beta | 0.14 | |
| NM_009695 | Apoc2 | apolipoprotein C-II | 0.41 | |
| NM_010174 | Fabp3 | Fatty acid binding protein 3, muscle and heart | 0.23 | |
| NM_008609 | Mmp15 | matrix metallopeptidase 15 | 0.46 | |
| NM_023792 | Pank1 | pantothenate kinase 1 | 0.29 | 0.3 |
| NM_144844 | Pcca | propionyl-Coenzyme A carboxylase, alpha polypeptide | 0.49 | |
| NM_013743 | Pdk4 | pyruvate dehydrogenase kinase, isoenzyme 4 | 0.4 | |
| NM_019437 | Rfk | riboflavin kinase | 0.48 | |
| NM_138758 | Tmlhe | trimethyllysine hydroxylase, epsilon | 0.37 | |
| NM_133995 | Upb1 | ureidopropionase, beta | 0.42 | |
| NM_009471 | Umps | uridine monophosphate synthetase | 0.36 | |
| Calcium homeostasis | ||||
| NM_013472 | Anxa6 | annexin A6 | 0.43 | |
| NM_007590 | Calm3 | calmodulin 3 | 0.43 | |
| NM_023051 | Clstn1 | calsyntenin 1 | 0.5 | |
| Electron Transport | ||||
| XM_485295 | Cyb561d1 | cytochrome b-561 domain containing 1 | 0.44 | |
| NM_013809 | Cyp2g1 | cytochrome P450, family 2, subfamily g, polypeptide 1 | 0.43 | |
| NM_177380 | Cyp3a44 | cytochrome P450, family 3, subfamily a, polypeptide 44 | 0.39 | |
| NM_018887 | Cyp39a1 | cytochrome P450, family 39, subfamily a, polypeptide 1 | 0.41 | |
| NM_010012 | Cyp8b1 | cytochrome P450, family 8, subfamily b, polypeptide 1 | 0.5 | |
| NM_170778 | Dpyd | dihydropyrimidine dehydrogenase | 0.49 | |
| NM_010231 | Fmo1 | flavin containing monooxygenase 1 | 0.42 | |
| NM_008631 | Mt4 | metallothionein 4 | 0.13 | |
| NM_026614 | Ndufa5 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 5 | 0.4 | |
| NM_026610 | Ndufb4 | NADH dehydrogenase (ubiquinone) 1 beta subcomplex 4 | 0.47 | |
| NM_010887 | Ndufs4 | NADH dehydrogenase (ubiquinone) Fe-S protein 4 | 0.47 | |
| NM_178239 | Ndor1 | NADPH dependent diflavin oxidoreductase 1 | 0.44 | |
| XM_128552 | Pdia2 | protein disulfide isomerase associated 2 | 0.43 | |
| NM_025848 | Sdhd | succinate dehydrogenase complex, subunit D, integral membrane protein | 0.45 | |
| NM_013711 | Txnrd2 | thioredoxin reductase 2 | 0.4 | |
| NM_011743 | Zfp106 | zinc finger protein 106 | 0.1 | |
| Golgi assembly and glycosylation | ||||
| NM_007454 | Ap1b1 | adaptor protein complex AP-1, beta 1 subunit | 0.49 | |
| NM_028758 | Gga2 | Golgi associated, gamma adaptin ear containing, ARF binding protein 2 | 0.44 | |
| NM_008315 | St3gal2 | ST3 beta-galactoside alpha-2,3-sialyltransferase 2 | 0.5 | |
| G-protein coupled receptors | ||||
| NM_008315 | Htr7 | 5-hydroxytryptamine (serotonin) receptor 7 (Htr7), mRNA | 0.47 | |
| NM_177231 | Arrb1 | arrestin, beta 1 | 0.38 | |
| NM_030258 | Gpr146 | G protein-coupled receptor 146 | 0.49 | |
| NM_010309 | Gnas | GNAS (guanine nucleotide binding protein, alpha stimulating) complex locus | 0.38 | |
| NM_023121 | Gngt2 | guanine nucleotide binding protein (G protein), gamma transducing activity polypeptide 2 | 0.47 | |
| NM_008142 | Gnb1 | guanine nucleotide binding protein, beta 1 | 0.5 | |
| NM_053235 | V1rc5 | vomeronasal 1 receptor, C5 | 0.43 | |
| Kinases and Phosphatases | ||||
| NM_177343 | Camk1d | Calcium/calmodulin-dependent protein kinase 1D | 0.18 | |
| NM_009793 | Camk4 | Calcium/calmodulin-dependent protein kinase IV (Camk4) | 0.11 | |
| NM_013642 | Dusp1 | dual specificity phosphatase 1 | 0.44 | |
| NM_010765 | Mapkapk5 | MAP kinase-activated protein kinase 5 | 0.47 | |
| NM_011951 | Mapk14 | mitogen activated protein kinase 14 | 0.46 | |
| NM_011944 | Map2k7 | mitogen activated protein kinase kinase 7 | 0.16 | |
| NM_023538 | Mulk | multiple substrate lipid kinase | 0.45 | |
| NM_145962 | Pank3 | pantothenate kinase 3 | 0.4 | |
| NM_00102495 | Pik3r1 | phosphatidylinositol 3-kinase, regulatory subunit, polypeptide 1 (p85 alpha) | 0.46 | |
| NM_145401 | Prkag2 | protein kinase, AMP-activated, gamma 2 non-catalytic subunit | 0.49 | 0.12 |
| NM_017374 | Ppp2cb | Protein phosphatase 2a, catalytic subunit, beta isoform | 0.44 | |
| NM_008914 | Ppp3cb | protein phosphatase 3, catalytic subunit, beta isoform | 0.44 | |
| NM_019651 | Ptpn9 | Protein tyrosine phosphatase, non-receptor type 9 | 0.23 | |
| NM_011213 | Ptprf | protein tyrosine phosphatase, receptor type, F | 0.4 | |
| NM_009184 | Ptk6 | PTK6 protein tyrosine kinase 6 | 0.37 | |
| NM_013845 | Ror1 | Receptor tyrosine kinase-like orphan receptor 1 | 0.43 | |
| NM_019924 | Rps6ka4 | ribosomal protein S6 kinase, polypeptide 4 | 0.43 | |
| Nuclear assembly and processing | ||||
| NM_148948 | Dicer1 | Dicer1, Dcr-1 homolog (Drosophila) | 0.43 | |
| XM_131040 | Hist2h2bb | Histone 2, H2bb | 0.37 | |
| NM_019786 | Tbk1 | TANK-binding kinase 1 | 0.4 | |
| Glucose biosynthesis/metabolism | ||||
| NM_019395 | Fbp1 | fructose bisphosphatase 1 | 0.47 | |
| NM_025799 | Fuca2 | fucosidase, alpha-L- 2, plasma | 0.44 | |
| NM_008155 | Gpi1 | glucose phosphate isomerase 1 | 0.49 | |
| NM_008061 | G6pc | glucose-6-phosphatase, catalytic | 0.26 | |
| NM_00101337 | Lman2l | lectin, mannose-binding 2-like | 0.49 | |
| NM_008548 | Man1a | mannosidase 1, alpha | 0.45 | |
| NM_010956 | Ogdh | Oxoglutarate dehydrogenase (lipoamide) | 0.25 | |
| NM_00101336 | Prkaa1 | protein kinase, AMP-activated, alpha 1 catalytic subunit | 0.49 | |
| Signaling molecules and interacting partners | ||||
| NM_009755 | Bmp1 | bone morphogenetic protein 1 | 0.42 | |
| NM_00101336 | E2f8 | E2F transcription factor 8 | 0.41 | |
| NM_010141 | Epha7 | Eph receptor A7 | 0.49 | |
| NM_020273 | Gmeb1 | glucocorticoid modulatory element binding protein 1 | 0.48 | |
| NM_010323 | Gnrhr | Gonadotropin releasing hormone receptor | 0.28 | |
| NM_176958 | Hif1an | hypoxia-inducible factor 1, alpha subunit inhibitor | 0.49 | |
| NM_010547 | Ikbkg | inhibitor of kappaB kinase gamma | 0.45 | |
| NM_010515 | Igf2r | insulin-like growth factor 2 receptor | 0.44 | |
| NM_010513 | Igf1r | insulin-like growth factor I receptor | 0.35 | 0.5 |
| NM_009697 | Nr2f2 | Nuclear receptor subfamily 2, group F, member 2 | 0.45 | |
| Pdap1 | PDGFA associated protein 1 | 0.47 | ||
| NM_013634 | Pparbp | peroxisome proliferator activated receptor binding protein | 0.44 | |
| NM_027230 | Prkcbp1 | protein kinase C binding protein 1 | 0.48 | |
| NM_026880 | Pink1 | PTEN induced putative kinase 1 | 0.48 | |
| NM_018773 | Scap2 | src family associated phosphoprotein 2 | 0.49 | |
| NM_007706 | Socs2 | Suppressor of cytokine signaling 2 | 0.44 | |
| NM_178111 | Trp53inp2 | tumor protein p53 inducible nuclear protein 2 | 0.47 | |
| NM_011703 | Vipr1 | vasoactive intestinal peptide receptor 1 | 0.4 | |
| NM_010153 | Erbb3 | v-erb-b2 erythroblastic leukemia viral oncogene homolog 3 (avian) | 0.46 | |
| Transport | ||||
| NM_009727 | Atp8a1 | ATPase, aminophospholipid transporter (APLT), class I, type 8A, member 1 | 0.44 | |
| NM_024173 | Atp6v1g1 | ATPase, H+ transporting, V1 subunit G isoform 1 | 0.46 | |
| NM_029600 | Abcc3 | Multidrug resistance-associated protein 3 (Abcc3) | 0.46 | |
| NM_010604 | Kcnj16 | potassium inwardly-rectifying channel, subfamily J, member 16 | 0.15 | |
| NM_018824 | Slc23a2 | Sodium-dependent vitamin C transporter type 2 (Slc23a1) | 0.45 | |
| NM_011397 | Slc23a1 | solute carrier family 23 (nucleobase transporters), member 1 | 0.3 | |
| NM_008063 | Slc37a4 | solute carrier family 37 (glycerol-6-phosphate transporter), member 4 | 0.46 | |
| NM_018760 | Slc4a4 | solute carrier family 4 (anion exchanger), member 4 | 0.5 | |
| NM_016917 | Slc40a1 | Solute carrier family 40 (iron-regulated transporter), member 1 | 0.45 | |
| XM_127434 | Slc9a3 | solute carrier family 9 (sodium/hydrogen exchanger), member 3 | 0.41 | |
| Detoxifying enzymes | ||||
| NM_008129 | Gclm | glutamate-cysteine ligase , modifier subunit | 0.45 | |
| NM_029555 | Gstk1 | glutathione S-transferase kappa 1 | 0.45 | |
| NM_010357 | Gsta4 | glutathione S-transferase, alpha 4 | 0.49 | |
| NM_010359 | Gstm3 | glutathione S-transferase, mu 3 | 0.22 | |
| XM_359308 | Gstm7 | glutathione S-transferase, mu 7 | 0.46 | |
| NM_008185 | Gstt1 | glutathione S-transferase, theta 1 | 0.43 | |
| NM_025304 | Lcmt1 | leucine carboxyl methyltransferase 1 | 0.48 | |
| NM_019946 | Mgst1 | microsomal glutathione S-transferase 1 | 0.18 | |
| NM_025569 | Mgst3 | microsomal glutathione S-transferase 3 | 0.47 | |
| NM_019878 | Sult1b1 | sulfotransferase family 1B, member 1 | 0.41 | |
| Ubiquitination and Proteolysis | ||||
| NM_011780 | Adam23 | A disintegrin and metallopeptidase domain 23 | 0.34 | |
| NM_007754 | Cpd | carboxypeptidase D | 0.45 | |
| NM_134015 | Fbxw11 | F-box and WD-40 domain protein 11 | 0.27 | |
| NM_177703 | Fbxw19 | F-box and WD-40 domain protein 19 | 0.11 | |
| NM_028705 | Herc3 | hect domain and RLD 3 | 0.35 | |
| NM_145486 | Mar 2 | membrane-associated ring finger (C3HC4) 2 | 0.5 | 0.28 |
| NM_020487 | Prss21 | protease, serine, 21 | 0.12 | |
| NM_008944 | Psma2 | proteasome (prosome, macropain) subunit, alpha type 2 | 0.49 | |
| NM_013640 | Psmb10 | proteasome (prosome, macropain) subunit, beta type 10 | 0.5 | |
| XM_483996 | Usp34 | ubiquitin specific peptidase 34 | 0.49 | |
| NM_013918 | Usp25 | Ubiquitin-specific processing protease | 0.47 | |
| Molecular Chaperones and Heat Shock Proteins | ||||
| NM_146036 | Ahsa1 | AHA1, activator of heat shock 90kDa protein ATPase homolog 1 (yeast) | 0.4 | |
| NM_025384 | Dnajc15 | DnaJ (Hsp40) homolog, subfamily C, member 15 | 0.5 | |
| NM_139139 | Dnajc17 | DnaJ (Hsp40) homolog, subfamily C, member 17 | 0.43 | |
| NM_024219 | Hsbp1 | heat shock factor binding protein 1 | 0.46 | |
| Hspa1b | heat shock protein 1B | 0.41 | ||
| NM_019960 | Hspb3 | heat shock protein 3 | 0.3 | |
| Miscellaneous | ||||
| NM_008708 | Nmt2 | N-myristoyltransferase 2 | 0.14 | |
| NM_007453 | Prdx6 | peroxiredoxin 6 | 0.46 | |
| NM_011434 | Sod1 | Superoxide dismutase 1, soluble | 0.25 | |
Genes that were suppressed >2-fold by TM only in small intestine of Nrf2 wild-type mice but not in small intestine of Nrf2 knockout mice compared with vehicle treatment at 3h. The relative mRNA expression levels of each gene in treatment group over vehicle group (fold changes) are listed.
Genes that were suppressed >2-fold by TM only in liver of Nrf2 wild-type mice but not in liver of Nrf2 knockout mice compared with vehicle treatment at 3h. The relative mRNA expression levels of each gene in treatment group over vehicle group (fold changes) are listed.
Other categories of genes affected included apoptosis and cell cycle-related genes (Caspases 6 and 11, growth arrest and DNA-damage-inducible 45 β), electron transport (Cyp450 members and NADH dehydrogenase isoforms), kinases and phosphatases (mitogen activated protein kinase family members, ribosomal protein S6 kinase), transcription factors and interacting partners (inhibitor of kappa B kinase gamma and src family associated phosphoprotein 2), and glucose-related genes (glucose-6-phosphatase, catalytic, fructose bisphosphatase 1, and glucose phosphate isomerase 1). Superoxide dismutase (Sod1) was also identified as an Nrf2-regulated and TM-modulated gene that was suppressed. Furthermore, cell adhesion genes (cadherin 22), ubiquitination and proteolysis genes (Usp25 and Usp34), and some unfolded protein response genes (heat shock proteins 1B and 3) were also observed to be downregulated in response to TM treatment via Nrf2.
4.0. Discussion
The major goal of this study was to identify toxicant Tunicamycin-regulated Nrf2-dependent genes in mice liver and small intestine by using C57BL/6J Nrf2 (+/+; wildtype) and C57BL/6J/Nrf2(−/−; knockout) mice and genome-scale microarray analyses. We sought to investigate by transcriptome expression profiling the potential role of ER stress stimulus in modulating Nrf2 function as a transcriptional activator in vivo. As a protein-folding compartment, the ER is exquisitely sensitive to alterations in homeostasis, and provides stringent quality control systems to ensure that only correctly folded proteins transit to the Golgi and unfolded or misfolded proteins are retained and ultimately degraded. A number of biochemical and physiological stimuli, such as perturbation in calcium homeostasis or redox status, elevated secretory protein synthesis, expression of misfolded proteins, sugar/glucose deprivation, altered glycosylation, and overloading of cholesterol can disrupt ER homeostasis, impose stress to the ER, and subsequently lead to accumulation of unfolded or misfolded proteins in the ER lumen [23]. The ER has evolved highly specific signaling pathways called the unfolded protein response (UPR) to cope with the accumulation of unfolded or misfolded proteins [4, 23]. ER stress stimulus by Thapsigargin has also been shown [24] to activate the c-Jun N-terminal kinase (JNK) or stress-activated protein kinase (SAPK) that is a member of the mitogen-activated protein kinase (MAPK) cascade [25]. Moreover, it has been reported that the coupling of ER stress to JNK activation involves transmembrane protein kinase IRE1 by binding to an adaptor protein TRAF2, and that IRE1α−/− fibroblasts were impaired in JNK activation by ER stress [26]. We have previously reported that phenethyl isothiocyanate (PEITC) from cruciferous vegetables activates JNK1 [27] and that the activation of the antioxidant response element (ARE) by PEITC involves both Nrf2 and JNK1 [13] in HeLa cells. We have also reported [12] that extracellular signal-regulated kinase (ERK) and JNK pathways play an unequivocal role in positive regulation of Nrf2 transactivation domain activity in vitro in HepG2 cells. Recently, it was shown [21] that Nrf1, another member of the Cap’ n’ Collar (CNC) family of basic leucine zipper proteins that is structurally similar to Nrf2, is normally targeted to the ER membrane, and that ER stress induced by TM in vitro may play a role in modulating Nrf1 function as a transcriptional activator. Here, we investigated the role of Nrf2 in modulating transcriptional response to ER stress stimulus by TM in vivo in an Nrf2 (−/−; deficient) murine model, thus providing new biological insights into the diverse cellular and physiological processes that may be regulated by the UPR in cancer pharmacology and toxicology.
Interestingly, a co-ordinated response involving Phase I, II and III genes that has not been demonstrated earlier was observed in vivo on ER stress induction with TM in an Nrf2-dependent manner. Phase I drug-metabolizing enzymes (DMEs) such as cytochrome P450 family members Cyp3a44, Cyp39a1 and Cyp8b1 were downregulated in response to TM-treatment in an Nrf2-dependent manner. Additionally, major Phase II detoxifying genes identified as Nrf2-regulated and TM-modulated included several isoforms of Glutathione-S-transferase (Gst), and glutamate cysteine ligase, modifier subunit (Gclm). Moreover, many transport genes, which may be regarded as Phase III genes, including members of solute carrier family (Slc23a2, Slc23a1, Slc37a4, Slc4a4, Slc40a1, Slc9a3) and multidrug-resistance associated proteins (Abcc1, Abcc3 and Mdr1b or Abcb1b) were also downregulated via Nrf2 and regulated through TM. The co-ordinated regulation of these genes could have significant effects in toxicology by enhancing the cellular defense system, preventing the activation of procarcinogens/reactive intermediates, and increasing the excretion/efflux of reactive carcinogens or metabolites.
There could be two possible outcomes of prolonged ER stress: (1) an adaptive response promoting cell survival; or (2) the induction of apoptotic cell death [3]. Indeed, several genes related to apoptosis and cell cycle control were modulated in response to TM stimulus in vivo in an Nrf2-dependent manner. The major genes upregulated in this category included the anti-apoptotic B-cell leukemia/lymphoma 2 (Bcl2) family gene, CASP8 and FADD-like apoptosis regulator (Cflar), Epiregulin (Ereg), Growth arrest specific 2 (Gas2), cyclin T2 (Ccnt2) and cyclin-dependent kinase 7 (Cdk7) all in small intestine apart from mucin 20 (Muc20) and synovial apoptosis inhibitor 1, synoviolin (Syvn1) in liver ; whereas genes downregulated in this category included cyclin-dependent kinase 6 (Cdk6) and Bcl2 in liver, baculoviral inhibitor of apoptosis (IAP)-repeat containing 6 (Birc6) and Caspases 6 and 11 in small intestine, and growth arrest and DNA-damage-inducible 45 – β (Gadd45b), and gamma interacting protein 1 (Gadd45gip1) - in liver and small intestine respectively amongst others. To our knowledge, this is the first report in vivo of apoptosis and cell cycle-related genes that are both modulated by the ER stress inducer TM and regulated via Nrf2. Moreover, it has been noted [28] that although the basic machinery to carry out apoptosis appears to be present in essentially all mammalian cells at all times, the activation of the suicide program is regulated by many different signals that originate from both the intracellular and the extracellular milieu. Notably, transcription factor NF-κB is critical for determining cellular sensitivity to apoptotic stimuli by regulating both mitochondrial and death receptor apoptotic pathways. Recently, it was reported [29] that autocrine tumor necrosis factor alpha links ER stress to the membrane death receptor pathway through IRE1alpha-mediated NF-κB activation and down-regulation of TRAF2 expression. In our study, we saw a downregulation of inhibitor of kappaB kinase gamma (Iκbkg or IKKγ) in liver in an Nrf2-dependent manner in response to TM-induced ER stress. Since the catalytic subunits, IKK and IKKβ, require association with the regulatory IKKγ (NEMO) component to gain full basal and inducible kinase activity and since tetrameric oligomerization of IκB Kinase γ (IKKγ) is obligatory for IKK Complex activity and NF-κB activation [30], our results appear to be validated from a functional standpoint and underscore the complexity of factors involved in making the decision between cell survival and cell death in response to TM-mediated ER stress in vivo, not excluding the possibility of potential cross-talk between Nrf2/ARE pathway and other signaling pathways that may converge at multiple levels in the cell.
Interestingly, impaired proteasome function through pharmacological inhibition, or by accumulation of malfolded protein in the cytoplasm, can ultimately block ER-associated degradation (ERAD) [31] which is important for eviction of malfolded proteins from the ER to the cytoplasm where they are subsequently ubiquitinated and degraded via the proteasome. In our study, several genes associated with the ubiquitin/proteasome pathway were regulated in response to TM in an Nrf2-dependent manner. These included, amongst others, constitutive photomorphogenic protein (Cop1), carboxypeptidase A4 (Cpa4), ubiquitin-specific peptidase 34 (Usp34), and ubiquitin-specific processing protease (Usp25). Furthermore, UPR genes such as various heat shock proteins (Hspb3, Hspb6, Hspb7, Hspa1B) and molecular chaperones and folding enzymes, e.g., stress 70 protein chaperone (Stch) were also seen to be regulated by TM-induced ER stress and modulated by Nrf2. Since the UPR directs gene expression important for remediating accumulation of malfolded protein in the ER, the identification of UPR-responsive genes in our study validates our results from a biological perspective. Moreover, important genes related to glycosylation modifications (e.g., galactosyltransferase, B3galt1), ER to Golgi transport (ADP-ribosylation factor GTPase activating protein 3, Arfgap3; coatomer protein complex subunit alpha, Copa; Lectin, mannose-binding 1,Lman1), and intra-Golgi transport (Golgi associated, gamma adaptin ear containing, ARF binding protein 2, Gga2) were also seen to be regulated by TM in an Nrf2-dependent manner. Genes related to biogenesis of ribosomes on rough ER where proteins are synthesized from mRNA, e.g., brix domain containing 2 (Bxdc2) and ribosomal protein S6 kinase, polypeptides 1 (Rps6ka1) and 4 (Rps6ka4), were also regulated via Nrf2 and modulated by TM treatment. To our knowledge, this is the first in vivo investigation examining the potential role of Nrf2 and TM-induced ER stress in the simultaneous modulation of UPR-responsive genes, clearance by the ubiquitin/proteasome pathway members, and cellular biosynthetic-secretory pathway involving ribosomal biogenesis genes and ER to Golgi transport genes.
Additionally, many genes related to glucose biosynthesis and metabolism including glucose phosphate isomerase 1 (gluconeogenesis/glycolysis), fructose bisphosphatase 1(gluconeogenesis), glucose-6-phosphatase (glycogen biosynthesis), hexokinase 2 (glycolysis), adiponectin (glucose metabolism), lectins (galactose- and mannose-binding) and the solute carrier family member Slc 35b1 (sugar porter) were all seen to be regulated through Nrf2 and modulated by TM-induced ER stress. The simultaneous modulation of genes encoding for insulin like growth factor receptors 1 and 2 point to a potential role for glucose- and ER stress-mediated insulin resistance [32] wherein the potential role of Nrf2 has never been examined earlier.
In recent times, there is a renewed interest in dissecting the interacting partners of Nrf2 such as coactivators and corepressors which are co-regulated with Nrf2 to better understand the biochemistry of Nrf2. In a recent microarray study [33], we have reported that CREB-binding protein (CBP) was upregulated in mice liver on treatment with (-)epigallocatechin-3-gallate (EGCG) in an Nrf2-dependent manner. We have also demonstrated [12] previously, using a Gal4-Luc reporter co-transfection assay system in HepG2 cells, that the nuclear transcriptional coactivator CBP, which can bind to Nrf2 transactivation domain and can be activated by extracellular signal-regulated protein kinase (ERK) cascade, showed synergistic stimulation with Raf on the transactivation activities of both the chimera Gal4-Nrf2 (1–370) and the full-length Nrf2. In the current study, we observed the upregulation of the P300/CBP-associated factor (P/CAF), transacting factor v-maf musculoaponeurotic fibrosarcoma oncogene family, protein F (Maf F), nuclear receptor co-activator 5 (Ncoa5), nuclear receptor co-repressor interacting protein (Nrip1) and Smad nuclear interacting protein 1 (Snip1) ; as well as downregulation of the src family associated phosphoprotein 2 (Scap2) in an Nrf2-dependent manner. Although microarray expression profiling cannot provide evidence of binding between partners, this is the first investigation to potentially suggest that co-repressor Nrip1 and co-activators P/CAF and Ncoa5, similar to CBP in our previous studies, may serve as putative TM-regulated nuclear interacting partners of Nrf2 in eliciting the UPR-responsive events in vivo. We have also shown recently [34] that coactivator P/CAF could transcriptionally activate a chimeric Gal4-Nrf2-Luciferase system containing the Nrf2 transactivation domain in HepG2 cells. In addition, P/CAF which is known [35] to be a histoneacetyl transferase protein has recently been shown [36] to mediate DNA damage-dependent acetylation on most promoters of genes involved in the DNA-damage and ER-stress response, which validates our observation of P/CAF induction via Nrf2 in response to TM-induced ER stress. Taken together, it is tempting to speculate that the TM-regulated pharmacological and toxicological effects may be regulated by a multimolecular complex, which involves Nrf2 along with the transcriptional co-repressor Nrip1 and the transcriptional co-activators P/CAF and Ncoa5, in addition to the currently known trans-acting factors such as small Maf [20], with multiple interactions between the members of the putative complex as we have shown recently with the p160 family of proteins [34]. Indeed, further studies of a biochemical nature would be needed to substantiate this hypothesis and extend our understanding of Nrf2 regulation in TM-mediated ER stress.
Many important transcription factors affecting diverse signaling pathways were identified as regulated through Nrf2 and modulated by TM treatment. For example, Jun oncogene, platelet-derived growth factor, metallothionein 1 and 2, transforming growth factor beta 1 and ErbB2 interacting protein were upregulated ; whereas hypoxia-inducible factor 1, alpha subunit inhibitor, peroxisome proliferator activated receptor binding protein, v-erb-b2 erythroblastic leukemia viral oncogene homolog 3 (avian) and protein kinase C binding protein 1 were downregulated via Nrf2 in response to TM. Since these transcription factors can modulate the expression of many different gene transcripts encoding various proteins, their identification as Nrf2-regulated and ER-stress- or TM-modulated would be important in enhancing our current understanding of UPR responsive genes and in providing new biological insights into the diverse cellular and physiological processes that may be regulated by the UPR in Nrf2-regulated cancer pharmacology and toxicology.
In the category of kinases and phosphatases, several members of the MAPK cascade such as Map2k7, Mapk14, Mapk8, Map3k7 as well as MAPK-activated protein kinase 5 (Mapkapk5) were identified as regulated by TM via Nrf2. Moreover, members of the calcium/calmodulin signaling pathway such as calcium/calmodulin-dependent - protein kinase I gamma (Camkg), -protein kinase 1D (Camk1d) and -protein kinase IV (Camk4) were shown to be regulated by TM in an Nrf2-dependent manner. Interestingly, glutathione peroxidase 3 (Gpx3) was upregulated and superoxide dismutase 1 (Sod1) was downregulated by TM via Nrf2 which can have important implications in oxidative stress-mediated [37] pathophysiology or ER stress caused by perturbations in redox circuitry [23, 37, 38]
Indeed, there is a growing interest amongst researchers in targeting the UPR in cancerous tumor growth [39]. Recently, it was shown [40] that the proteasomal inhibitor bortezomib induces a unique type of ER stress characterized by an absence of eif2alpha phosphorylation, ubiquitylated protein accumulation, and proteotoxicity in human pancreatic cancer cells. It was also reported [41] that malignant B cells may be highly dependent on ER-Golgi protein transport and that targeting and inhibiting this process by brefeldin A may be a promising therapeutic strategy for B-cell malignancies, especially for those that respond poorly to conventional treatments, e.g., fludarabine resistance in chronic lymphocytic leukemia (CLL). However, the role of Nrf2 in modulating the UPR in vivo has never been examined before.
The current study, thus, addresses the spatial regulation in mouse small intestine and liver of global gene expression profiles elicited by TM-mediated ER stress via Nrf2. Several common clusters of genes such as that for ubiquitin/proteasome, cell adhesion, transcription factors were observed in this study that were also observed in previous studies with Nrf2 activators[9, 33, 42–44] which validates our studies from a functional standpoint. In addition, three clusters of genes – calcium homeostasis, ER/Golgi transport & ER/Golgi biosynthesis/metabolism genes, and glucose homeostasis genes – were uniquely observed as modulated via Nrf2 in response to TM-mediated ER stress that were not discernible in previous studies with Nrf2 activators. Indeed, the involvement of the three clusters mentioned above is a rational response to alteration in the homeostatic environment brought about by the toxicant TM-induced ER stress, and is reflective of their potential role in the UPR to ER stress that is naturally not observed in previous studies on cancer chemoprevention with Nrf2 activators that do not induce ER stress. The presence of the three unique clusters as mentioned above that relate to the putative role of these genes in the UPR is an effect that appears to be elicited in a toxicant-specific manner. In addition, classical Phase II genes such as Gst isoforms and Gclm were downregulated in a Nrf2-dependent fashion in response to the toxicant TM at 3 hours in this study. We were able to see the downregulation of classical Phase II genes in qRT-PCR experiments performed at a 12 hour time-point (data not shown) with the extent of downregulation being more pronounced at 12 hours than at 3 hours in response to the toxicant TM. Interestingly, this contrasts with the delayed response reported for the classical Phase II gene NQO1 in response to Nrf2 activator BHA (Butylated hydroxyanisole) wherein the induction of the gene peaked at 12 hours[43] with no gene induction at 3 hours. Taken together, the downregulation of classical Phase II genes in response to TM-induced ER stress should be viewed in the light of a complex of physiological factors including partitioning across the gastrointestinal tract, intestinal transit time, uptake into the hepatobiliary circulation, exposure parameters such as Cmax, Tmax and AUC, and pharmacokinetics of disposition after oral administration of TM. Further studies will be necessary to address the effect(s) of temporal dependence on pharmacokinetic parameters and gene expression profiles to further enhance our current understanding of TM-mediated ER stress response, the complexity of kinetics of Phase II gene expression response to a toxicant and the role of Nrf2.
In conclusion, our microarray expression profiling study provides some novel insights into the pharmacogenomics and spatial regulation of global gene expression profiles elicited in the mouse small intestine and liver by TM in an Nrf2-dependent manner from a biological perspective. Amongst these TM-regulated genes, clusters of Nrf2-dependent genes were identified by comparing gene expression profiles between C57BL/6J Nrf2(+/+) and C57BL/6J/Nrf2(−/−) mice. The identification of novel molecular targets that are regulated by TM via Nrf2 in vivo raises possibilities for targeting the UPR proteins in future to augment or suppress the ER stress response and modulate disease progression. This study clearly extends the current latitude of thought on the molecular mechanisms underlying TM-mediated UPR effects as well as the role(s) of Nrf2 in its biological functions. Future in vivo and in vitro mechanistic studies exploring the germane molecular targets or signaling pathways as well as Nrf2-dependent genes related to the significant functional categories uncovered in the current study would greatly extend our understanding of the diverse cellular and physiological processes that may be regulated by the UPR in cancer pharmacology and toxicology, and the potential role of ER stress in modulating Nrf2 function as a transcriptional activator. .
Acknowledgments
The authors are deeply grateful to Mr. Curtis Krier at the Cancer Institute of New Jersey (CINJ) Core Expression Array Facility for his expert assistance with the microarray analyses. The authors are also deeply indebted to Ms. Donna Wilson of the Keck Center for Collaborative Neuroscience, Rutgers University as well as the staff of the Human Genetics Institute of New Jersey at Rutgers University for their great expertise and help with the quantitative real-time PCR analyses. This work was supported in part by NIH grant R01- 094828.
ABBREVIATIONS
- TM
Tunicamycin
- ER
Nuclear Factor-E2 -related factor 2, Nrf2, Endoplasmic Reticulum
- UPR
Unfolded Protein Response
- Mapk
Mitogen-activated protein kinase
- ARE
Antioxidant response element
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.van Huizen R, Martindale JL, Gorospe M, Holbrook NJ. P58IPK, a novel endoplasmic reticulum stress-inducible protein and potential negative regulator of eIF2alpha signaling. J Biol Chem. 2003;278:15558–64. doi: 10.1074/jbc.M212074200. [DOI] [PubMed] [Google Scholar]
- 2.Kaufman RJ. Stress signaling from the lumen of the endoplasmic reticulum: coordination of gene transcriptional and translational controls. Genes Dev. 1999;13:1211–33. doi: 10.1101/gad.13.10.1211. [DOI] [PubMed] [Google Scholar]
- 3.Reimertz C, Kogel D, Rami A, Chittenden T, Prehn JH. Gene expression during ER stress-induced apoptosis in neurons: induction of the BH3-only protein Bbc3/PUMA and activation of the mitochondrial apoptosis pathway. J Cell Biol. 2003;162:587–97. doi: 10.1083/jcb.200305149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Hetz C, Bernasconi P, Fisher J, Lee AH, Bassik MC, Antonsson B, Brandt GS, Iwakoshi NN, Schinzel A, Glimcher LH, Korsmeyer SJ. Proapoptotic BAX and BAK modulate the unfolded protein response by a direct interaction with IRE1alpha. Science. 2006;312:572–6. doi: 10.1126/science.1123480. [DOI] [PubMed] [Google Scholar]
- 5.Mahoney WC, Duksin D. Biological activities of the two major components of tunicamycin. J Biol Chem. 1979;254:6572–6. [PubMed] [Google Scholar]
- 6.Olden K, Pratt RM, Jaworski C, Yamada KM. Evidence for role of glycoprotein carbohydrates in membrane transport: specific inhibition by tunicamycin. Proc Natl Acad Sci U S A. 1979;76:791–5. doi: 10.1073/pnas.76.2.791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Alam J, Stewart D, Touchard C, Boinapally S, Choi AM, Cook JL. Nrf2, a Cap'n'Collar transcription factor, regulates induction of the heme oxygenase-1 gene. J Biol Chem. 1999;274:26071–8. doi: 10.1074/jbc.274.37.26071. [DOI] [PubMed] [Google Scholar]
- 8.McMahon M, Itoh K, Yamamoto M, Chanas SA, Henderson CJ, McLellan LI, Wolf CR, Cavin C, Hayes JD. The Cap'n'Collar basic leucine zipper transcription factor Nrf2 (NF-E2 p45-related factor 2) controls both constitutive and inducible expression of intestinal detoxification and glutathione biosynthetic enzymes. Cancer Res. 2001;61:3299–307. [PubMed] [Google Scholar]
- 9.Thimmulappa RK, Mai KH, Srisuma S, Kensler TW, Yamamoto M, Biswal S. Identification of Nrf2-regulated genes induced by the chemopreventive agent sulforaphane by oligonucleotide microarray. Cancer Res. 2002;62:5196–203. [PubMed] [Google Scholar]
- 10.Prochaska HJ, De Long MJ, Talalay P. On the mechanisms of induction of cancer-protective enzymes: a unifying proposal. Proc Natl Acad Sci U S A. 1985;82:8232–6. doi: 10.1073/pnas.82.23.8232. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Li W, Jain MR, Chen C, Yue X, Hebbar V, Zhou R, Kong AN. Nrf2 Possesses a redox-insensitive nuclear export signal overlapping with the leucine zipper motif. J Biol Chem. 2005;280:28430–8. doi: 10.1074/jbc.M410601200. [DOI] [PubMed] [Google Scholar]
- 12.Shen G, Hebbar V, Nair S, Xu C, Li W, Lin W, Keum YS, Han J, Gallo MA, Kong AN. Regulation of Nrf2 transactivation domain activity. The differential effects of mitogen-activated protein kinase cascades and synergistic stimulatory effect of Raf and CREB-binding protein. J Biol Chem. 2004;279:23052–60. doi: 10.1074/jbc.M401368200. [DOI] [PubMed] [Google Scholar]
- 13.Keum YS, Owuor ED, Kim BR, Hu R, Kong AN. Involvement of Nrf2 and JNK1 in the activation of antioxidant responsive element (ARE) by chemopreventive agent phenethyl isothiocyanate (PEITC) Pharm Res. 2003;20:1351–6. doi: 10.1023/a:1025737622815. [DOI] [PubMed] [Google Scholar]
- 14.Chen C, Kong AN. Dietary chemopreventive compounds and ARE/EpRE signaling. Free Radic Biol Med. 2004;36:1505–16. doi: 10.1016/j.freeradbiomed.2004.03.015. [DOI] [PubMed] [Google Scholar]
- 15.Itoh K, Wakabayashi N, Katoh Y, Ishii T, Igarashi K, Engel JD, Yamamoto M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Genes Dev. 1999;13:76–86. doi: 10.1101/gad.13.1.76. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Dhakshinamoorthy S, Jaiswal AK. Functional characterization and role of INrf2 in antioxidant response element-mediated expression and antioxidant induction of NAD(P)H:quinone oxidoreductase1 gene. Oncogene. 2001;20:3906–17. doi: 10.1038/sj.onc.1204506. [DOI] [PubMed] [Google Scholar]
- 17.Wakabayashi N, Dinkova-Kostova AT, Holtzclaw WD, Kang MI, Kobayashi A, Yamamoto M, Kensler TW, Talalay P. Protection against electrophile and oxidant stress by induction of the phase 2 response: fate of cysteines of the Keap1 sensor modified by inducers. Proc Natl Acad Sci U S A. 2004;101:2040–5. doi: 10.1073/pnas.0307301101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Li W, Yu SW, Kong AN. Nrf2 Possesses a Redox-Sensitive NES in the Neh5 transactivation Domain. J Biol Chem. 2006 doi: 10.1074/jbc.M602746200. In Press. [DOI] [PubMed] [Google Scholar]
- 19.Yu X, Kensler T. Nrf2 as a target for cancer chemoprevention. Mutat Res. 2005;591:93–102. doi: 10.1016/j.mrfmmm.2005.04.017. [DOI] [PubMed] [Google Scholar]
- 20.Dhakshinamoorthy S, Jaiswal AK. Small maf (MafG and MafK) proteins negatively regulate antioxidant response element-mediated expression and antioxidant induction of the NAD(P)H:Quinone oxidoreductase1 gene. J Biol Chem. 2000;275:40134–41. doi: 10.1074/jbc.M003531200. [DOI] [PubMed] [Google Scholar]
- 21.Wang W, Chan JY. Nrf1 is targeted to the ER membrane by a N-terminal transmembrane domain: inhibition of nuclear translocation and transacting function. J Biol Chem. 2006 doi: 10.1074/jbc.M602802200. [DOI] [PubMed] [Google Scholar]
- 22.Chan K, Lu R, Chang JC, Kan YW. NRF2, a member of the NFE2 family of transcription factors, is not essential for murine erythropoiesis, growth, and development. Proc Natl Acad Sci U S A. 1996;93:13943–8. doi: 10.1073/pnas.93.24.13943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Zhang K, Kaufman RJ. Protein folding in the endoplasmic reticulum and the unfolded protein response. Handb Exp Pharmacol. 2006:69–91. doi: 10.1007/3-540-29717-0_3. [DOI] [PubMed] [Google Scholar]
- 24.Srivastava RK, Sollott SJ, Khan L, Hansford R, Lakatta EG, Longo DL. Bcl-2 and Bcl-X(L) block thapsigargin-induced nitric oxide generation, c-Jun NH(2)-terminal kinase activity, and apoptosis. Mol Cell Biol. 1999;19:5659–74. doi: 10.1128/mcb.19.8.5659. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kyriakis JM, Banerjee P, Nikolakaki E, Dai T, Rubie EA, Ahmad MF, Avruch J, Woodgett JR. The stress-activated protein kinase subfamily of c-Jun kinases. Nature. 1994;369:156–60. doi: 10.1038/369156a0. [DOI] [PubMed] [Google Scholar]
- 26.Urano F, Wang X, Bertolotti A, Zhang Y, Chung P, Harding HP, Ron D. Coupling of stress in the ER to activation of JNK protein kinases by transmembrane protein kinase IRE1. Science. 2000;287:664–6. doi: 10.1126/science.287.5453.664. [DOI] [PubMed] [Google Scholar]
- 27.Yu R, Jiao JJ, Duh JL, Tan TH, Kong AN. Phenethyl isothiocyanate, a natural chemopreventive agent, activates c-Jun N-terminal kinase 1. Cancer Res. 1996;56:2954–9. [PubMed] [Google Scholar]
- 28.Steller H. Mechanisms and genes of cellular suicide. Science. 1995;267:1445–9. doi: 10.1126/science.7878463. [DOI] [PubMed] [Google Scholar]
- 29.Hu P, Han Z, Couvillon AD, Kaufman RJ, Exton JH. Autocrine tumor necrosis factor alpha links endoplasmic reticulum stress to the membrane death receptor pathway through IRE1alpha-mediated NF-kappaB activation and down-regulation of TRAF2 expression. Mol Cell Biol. 2006;26:3071–84. doi: 10.1128/MCB.26.8.3071-3084.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tegethoff S, Behlke J, Scheidereit C. Tetrameric oligomerization of IkappaB kinase gamma (IKKgamma) is obligatory for IKK complex activity and NF-kappaB activation. Mol Cell Biol. 2003;23:2029–41. doi: 10.1128/MCB.23.6.2029-2041.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Jiang HY, Wek RC. Phosphorylation of the alpha-subunit of the eukaryotic initiation factor-2 (eIF2alpha) reduces protein synthesis and enhances apoptosis in response to proteasome inhibition. J Biol Chem. 2005;280:14189–202. doi: 10.1074/jbc.M413660200. [DOI] [PubMed] [Google Scholar]
- 32.Wang H, Kouri G, Wollheim CB. ER stress and SREBP-1 activation are implicated in beta-cell glucolipotoxicity. J Cell Sci. 2005;118:3905–15. doi: 10.1242/jcs.02513. [DOI] [PubMed] [Google Scholar]
- 33.Shen G, Xu C, Hu R, Jain MR, Nair S, Lin W, Yang CS, Chan JY, Kong AN. Comparison of (-)-epigallocatechin-3-gallate elicited liver and small intestine gene expression profiles between C57BL/6J mice and C57BL/6J/Nrf2 (−/−) mice. Pharm Res. 2005;22:1805–20. doi: 10.1007/s11095-005-7546-8. [DOI] [PubMed] [Google Scholar]
- 34.Lin W, Shen G, Yuan X, Jain MR, Yu S, Zhang A, Chen JD, Kong AN. Regulation of Nrf2 Transactivation Domain Activity by p160 RAC3/SRC3 and Other Nuclear Co-Regulators. J Biochem Mol Biol. 2006;39:304–10. doi: 10.5483/bmbrep.2006.39.3.304. [DOI] [PubMed] [Google Scholar]
- 35.Chen H, Tini M, Evans RM. HATs on and beyond chromatin. Curr Opin Cell Biol. 2001;13:218–24. doi: 10.1016/s0955-0674(00)00200-3. [DOI] [PubMed] [Google Scholar]
- 36.Ceribelli M, Alcalay M, Vigano MA, Mantovani R. Repression of New p53 Targets Revealed by ChIP on Chip Experiments. Cell Cycle. 2006;5 doi: 10.4161/cc.5.10.2777. [DOI] [PubMed] [Google Scholar]
- 37.Kim BR, Hu R, Keum YS, Hebbar V, Shen G, Nair SS, Kong AN. Effects of glutathione on antioxidant response element-mediated gene expression and apoptosis elicited by sulforaphane. Cancer Res. 2003;63:7520–5. [PubMed] [Google Scholar]
- 38.Jones DP. Extracellular redox state: refining the definition of oxidative stress in aging. Rejuvenation Res. 2006;9:169–81. doi: 10.1089/rej.2006.9.169. [DOI] [PubMed] [Google Scholar]
- 39.Garber K. Researchers target unfolded protein response in cancerous tumor growth. J Natl Cancer Inst. 2006;98:512–4. doi: 10.1093/jnci/djj174. [DOI] [PubMed] [Google Scholar]
- 40.Nawrocki ST, Carew JS, Dunner K, Jr, Boise LH, Chiao PJ, Huang P, Abbruzzese JL, McConkey DJ. Bortezomib inhibits PKR-like endoplasmic reticulum (ER) kinase and induces apoptosis via ER stress in human pancreatic cancer cells. Cancer Res. 2005;65:11510–9. doi: 10.1158/0008-5472.CAN-05-2394. [DOI] [PubMed] [Google Scholar]
- 41.Carew JS, Nawrocki ST, Krupnik YV, Dunner K, Jr, McConkey DJ, Keating MJ, Huang P. Targeting endoplasmic reticulum protein transport: a novel strategy to kill malignant B cells and overcome fludarabine resistance in CLL. Blood. 2006;107:222–31. doi: 10.1182/blood-2005-05-1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kwak MK, Wakabayashi N, Itoh K, Motohashi H, Yamamoto M, Kensler TW. Modulation of gene expression by cancer chemopreventive dithiolethiones through the Keap1-Nrf2 pathway. Identification of novel gene clusters for cell survival. J Biol Chem. 2003;278:8135–45. doi: 10.1074/jbc.M211898200. [DOI] [PubMed] [Google Scholar]
- 43.Nair S, Xu C, Shen G, Hebbar V, Gopalakrishnan A, Hu R, Jain MR, Lin W, Keum YS, Liew C, Chan JY, Kong AN. Pharmacogenomics of Phenolic Antioxidant Butylated Hydroxyanisole (BHA) in the Small Intestine and Liver of Nrf2 Knockout and C57BL/6J Mice. Pharm Res. 2006 doi: 10.1007/s11095-006-9099-x. Epub. [DOI] [PubMed] [Google Scholar]
- 44.Shen G, Xu C, Hu R, Jain MR, Gopalkrishnan A, Nair S, Huang MT, Chan JY, Kong AN. Modulation of nuclear factor E2-related factor 2-mediated gene expression in mice liver and small intestine by cancer chemopreventive agent curcumin. Mol Cancer Ther. 2006;5:39–51. doi: 10.1158/1535-7163.MCT-05-0293. [DOI] [PubMed] [Google Scholar]