

Abstract
Adult neural progenitor cells (NPCs) are an attractive source for functional replacement in neurodegenerative diseases and traumatic central nervous systems (CNS) injury. It has been shown that transplantation of neural stem cells or NPCs into the lesioned region partially restores CNS function. However, the capacity of endogenous NPCs in neuronal replacement and functional recovery of spinal cord injury (SCI) is apparently poor. More significantly, the early response of endogenous adult NPCs to SCI remains largely undefined. A comprehensive understanding of the temporal and spatial responses of NPCs to SCI is essential for the development of effective NPC therapy to restore neurological function. To this end, we have analyzed the early organization, distribution, and potential function of NPCs in response to SCI using nestin promoter controlled LacZ reporter transgenic mice. We showed that there was an increase of NPC proliferation, migration, and neurogenesis in adult spinal cord after traumatic compression SCI. The proliferation of NPCs detected by BrdU incorporation and LacZ staining was restricted to the ependymal zone (EZ) of the central canal. During acute SCI, NPCs in the EZ of the central canal migrated vigorously toward the dorsal direction, where the compression lesion is generated. The optimal NPC migration occurred in the adjacent region close to the epicenter. More significantly, there was an increased de novo neurogenesis from NPCs 24 hours after SCI. The enhanced proliferation, migration, and neurogenesis of (from) endogenous NPCs in the adult spinal cord in response to SCI suggest a potential role for NPCs in attempting to restore SCI-mediated neuronal dysfunction.
Keywords: Neural progenitor cells, Neurogenesis, Astrogenesis, Spinal cord Injury, Nestin, Radial glia
Introduction
Traumatic spinal cord injury (SCI) causes neuronal and glial cell damage and tissue disruption, leading to neurological dysfunction. Two major pathological stages occur in SCI: The primary injury involves mechanical force-mediated cell necrosis and tissue damage, and the secondary injury results in a cascade of biochemical events that produce progressive destruction on the spinal cord tissues (1–4). Together, the death of neurons, astroglia, and oligodendroglia in and around the lesion site disrupts neural circuitry and leads to neurological dysfunction (5). Although the biochemical events leading to the temporal and spatial patterns of cell death and neurological dysfunction have been well-characterized, therapy to repair or prevent cell and tissue damage, and to restore neurological function remain largely unresolved (6). Recent studies have shown that transplantation of neuroepithelial cells, or neural stem/progenitor cells into the injured spinal cord can promote functional recovery in adult animal models. For example, McDonald et al.(7) have demonstrated that transplanted embryonic stem cells can not only survive, proliferate, differentiate, but also promote functional recovery in injured rat spinal cord. Akiyama et al. (8) have shown that transplanted neural precursor cells derived from the adult human brain can facilitate functional remyelination in the demyelinated spinal cord. More recently, Vroemen et al. (9) have demonstrated that adult neural progenitor cell grafts can integrate along axonal pathways, and connect with the existing neural networks after transplanted into acutely lesioned spinal cord. Furthermore, intravenously injected neural progenitor cells can migrate to the injured spinal cord and differentiate into neurons, astrocytes and oligodendrocytes (10). Nevertheless, it has been shown that majority of the survived neural stem cells, glial progenitor cells or NPCs after transplantation to the lesioned spinal cord differentiate into astroglial phenotypes (11,12). The molecular nature of differentiation of transplanted neural stem cells toward glial cell direction remains largely unknown. To some extent, the selective differentiation may be due to the anti-neurogenic function of bone morphogenetic proteins (BMPs) released after SCI (13,14). Transplantation of fetal neural precursor cells overexpressing BMP inhibitor promoted differentiation of neural precursor cells into neurons and oligodendrocytes, and more significantly, increased functional recovery in recipient mice after SCI (14). In addition, transplantation of other cell types, such as olfactory ensheathing cells and Schwann cells into the injured spinal cord has been demonstrated to increase tissue regenerative capacity, even though the structural and functional recovery was relatively moderate (15,16). Together, the transplantation of a variety of cells, particularly the neural stem cells or NPCs in multiple injury paradigms has provided to a certain degree encouraging results for functional recovery.
Neural progenitor cells have been shown to exist in adult mammalian CNS (17). Neurogenesis from NPCs occurs in developmental, growth and aging processes (18–21). For example, neurogenesis generates functional neurons in vitro from adult human and primate brain (22–24). In addition, pathological processes promote neurogenesis as reported in human patients with Alzheimer’s disease (25), and Huntington’s disease (26). Brain and spinal cord injury facilitates neurogenesis in traumatic animal models (27–30). These findings suggest that promotion of neurogenesis from adult NPCs may be potentially able to functionally replace degenerated (damaged) cells during pathogenesis or injury. More recently, in an amyotrophic lateral sclerosis (ALS)-like mouse model, we showed that motor neuron degeneration facilitate NPC proliferation, migration and differentiation in mouse spinal cord, particularly at the disease onset and progression stages. These results together suggest that adult endogenous NPCs may be a potential source for de novo neurogenesis. In fact, substantial evidence has demonstrated that adult mammalian spinal cords contain regenerative NPCs. More importantly, these NPCs are activated following injury and contribute to some functional repair, although the self-repair ability is limited (29–30).
A comprehensive understanding of NPCs responsible for neurogenesis is essential to the development of therapies aimed at promoting neural circuitry for functional recovery after SCI. The early response of NPCs to SCI remains largely unknown and the role of NPCs in functional recovery in SCI remains to be elucidated. Expression of nestin in the CNS is generally considered as a reliable NPC marker and has been extensively used for the characterization of NPCs in vitro and in vivo (31–36). To this end, we chose the nestin promoter controlled LacZ transgenic mice to analyze the temporal response of NPCs in the spinal cords of the compression lesioned mouse model of SCI. We demonstrated that there was an increase of NPC proliferation in the EZ of the central canal. In addition, we showed that there was an increase of NPC migration from EZ of the central canal to the lesion regions. More importantly, we demonstrated that there was increase of neurogenesis from NPCs following SCI. The enhanced NPC proliferation, migration and differentiation suggest that the endogenous NPCs may play an important role in attempting to repair SCI damaged neural circuitry.
Materials and Methods
Transgenic mouse lines
Adult (70–80 days of age) nestin promoter controlled reporter gene LacZ transgenic mice (pNes-Tg) (Jackson Laboratory, Bar Harbor, ME) were used for compression spinal cord injury (37,38). Transgenic progeny were identified by regular PCR amplification of tail DNA using specific primers. The experimental protocols for SCI studies were approved by the Institutional Animal Use and Care Committee and are in close agreement with the National Institutes of Health guideline for the care and use of laboratory animals.
Compression Spinal cord injury
The experimental model for mouse severe compression spinal cord injury was essentially similar to that previously described by Farooque (39) and with minor modifications (40). Briefly, animals were deeply anesthetized with pentobarbital in a dose 20 mg/kg body weight by intraperiotineal (i.p.) approach. After skin decontamination, a 15–20 mm midline incision was made, and a laminectomy of T12 to L3 vertebra was performed under a dissection microscope. Animals were then placed in a modified stereotaxic apparatus, and a 15 to 30 g of weight was applied to the spinal cord for 5 min with a 1×2-mm rectangular plastic plate. After injury, skin was sutured and mice were kept under a heating lamp for recovery.
In vivo 5-bromodeoxyuridine (BrdU) labeling
BrdU at 50 mg/kg/day was administrated by I.P. for 5 days to adult pNes-Tg mice. On day 5 of BrdU administration, mice were processed for spinal cord injury. Mice were continuously labeled with BrdU for 1 or 2 days before the spinal cords were processed for analyzing the early response of NPCs to acute traumatic injury. BrdU immunostaining was described in the following section.
LacZ staining, immunostaining, image analysis and quantification
The lumbar region of the spinal cord was used to analyze the organization and distribution of NPCs in response to SCI. For LacZ staining, sections (12 μm) were incubated in 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal) solution for 16 h at room temperature as previously described. For immunohistochemical staining, sections were incubated in blocking buffer (10% goat serum/0.2% triton X-100 in 1×PBS, pH 7.5) for 1 h at room temperature. Primary antibody was then added to the blocking buffer (1:250) and the section was incubated at 4 °C overnight. The next day, sections were washed 5 times (5 min each) in 1×PBS (pH 7.5) containing 0.5% triton X-100, followed by incubation with specific fluorescein-conjugated secondary antibody for 2 h at room temperature. After extensive washes, sections were covered with anti-fade medium and sealed for fluorescent analysis. For negative control staining, sections were incubated without primary antibody.
Image collection and analysis
All images were collected and analyzed with a Nikon fluorescent microscope E800 equipped with the Spot digital camera and Photoshop software. Quantifications of NPC distribution were performed by counting and analyzing the number of LacZ positive nuclei in the dorsal and ventral horn regions (5 sections/mouse, and 3 mice/group). Quantifications of LacZ staining intensity and BrdU staining intensity at the ependymal zone surrounding central canal of mouse spinal cords were performed with the NIH software Image J. At least 5 sections/mouse, and 3 mice were analyzed. The arbitrary units were used to express the LacZ and BrdU staining intensity of EZ.
Statistical analysis
Statistical analysis of SCI vs. normal control was performed using the paired Student t test. P < 0.05 was considered significant.
Results
Tissue damage and neurological dysfunction in acute compression SCI mice
The acute compression injury mouse model was selected to analyze the early response of NPCs to SCI according to a similar procedure described by Farooque (39), with minor modifications (40). Mild (15g of weight for 5 min), moderate (20g of weight for 5 min) and severe (30g of weight for 5 min) lesion conditions were applied to generate different degrees of SCI. The mouse hind limbs remained partial paralysis under mild lesion conditions, and complete paralysis under moderate and severe lesion conditions within 24 to 48 hours, while the control mice had minimum alteration in walking behaviors. The morphological and pathological changes adjacent to the epicenter of the lesioned spinal cords are shown in Figure 1A. There was extensive swelling, hemorrhage and tissue degeneration in the epicenter as the degree of injury was increased, shown by the haematoxylin and eosin (HE) staining. With the different lesion conditions, we carried out studies to analyze the early responses of adult endogenous NPCs to SCI.
Figure 1. Spinal cord injury promoted early responses of NPCs to tissue damage.
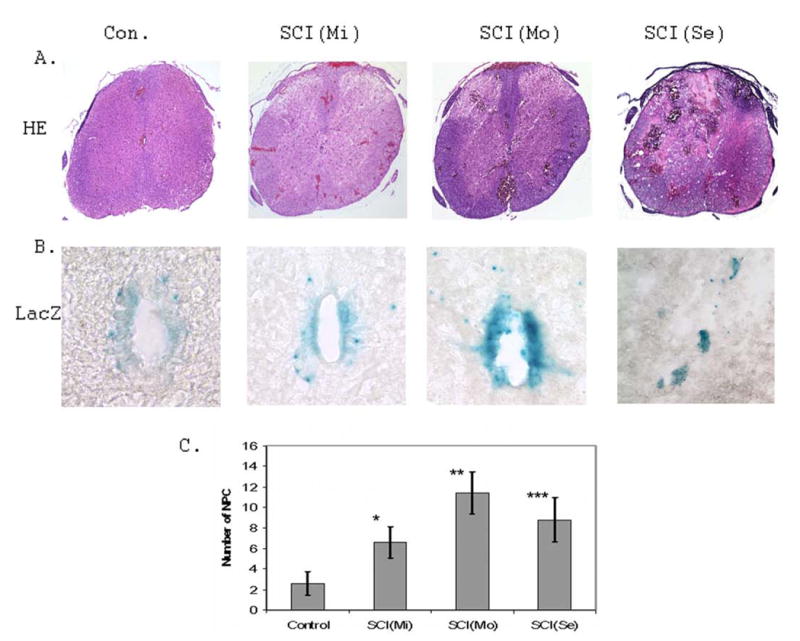
A. Spinal cord injury generates different degrees of tissue damage analyzed by HE staining. Different injury conditions [SCI(Mi):mild; SCI(Mo): moderate; SCI(Se): severe] generate different degree of spinal cord lesions. B. Identification of NPCs in the ependymal zone of the central canal region of adult mouse spinal cord was performed by LacZ staining. C. Spinal cord injury significantly increased the number of cells distributed in the central canal region compared to that of surgical control (n=3 mice; 5 sections/mouse; *p < 0.02; **p< 0.01; ***p<0.02).
Early responses of NPCs to compression SCI
Nestin promoter controlled reporter gene activity assay revealed that there was increased LacZ staining in the ependymal zone (EZ) of the central canal region upon SCI (Fig. 1B). More significantly, the number of NPCs migrating out toward the dorsal direction was dramatically increased in mild, moderate and severe SCI compared to that of surgical control mice (Fig. 1C). Interestingly, under severe SCI conditions, large LacZ positively stained aggregates apparently dissociated from the EZ of the central canal region migrated out toward the lesioned region in the dorsal areas (Fig. 1B).
Proliferation of NPCs in the ependymal zone (EZ) of the central canal
A combination of LacZ staining and BrdU labeling was used to study proliferation of NPCs in the pNes-LacZ mouse model after SCI (41,42). We focused on the moderate lesion conditions to analyze the organization and distribution of NPCs to SCI. We demonstrated that the EZ of the central canal contains NPCs that are positively stained with LacZ and BrdU antibody. More significantly, there was an increase of LacZ (Fig. 1B; Fig. 2) and BrdU (Fig. 2) staining intensity in the EZ of the central canal of the SCI mice compared to the surgical control mice. Most LacZ positive NPCs in the EZ of the central canal region were colocalized with BrdU staining, suggesting that there is an increase of NPC proliferation in the EZ of the central canal region upon SCI.
Figure 2. Spinal cord injury increased NPC proliferation in the ependymal zone of the central canal.
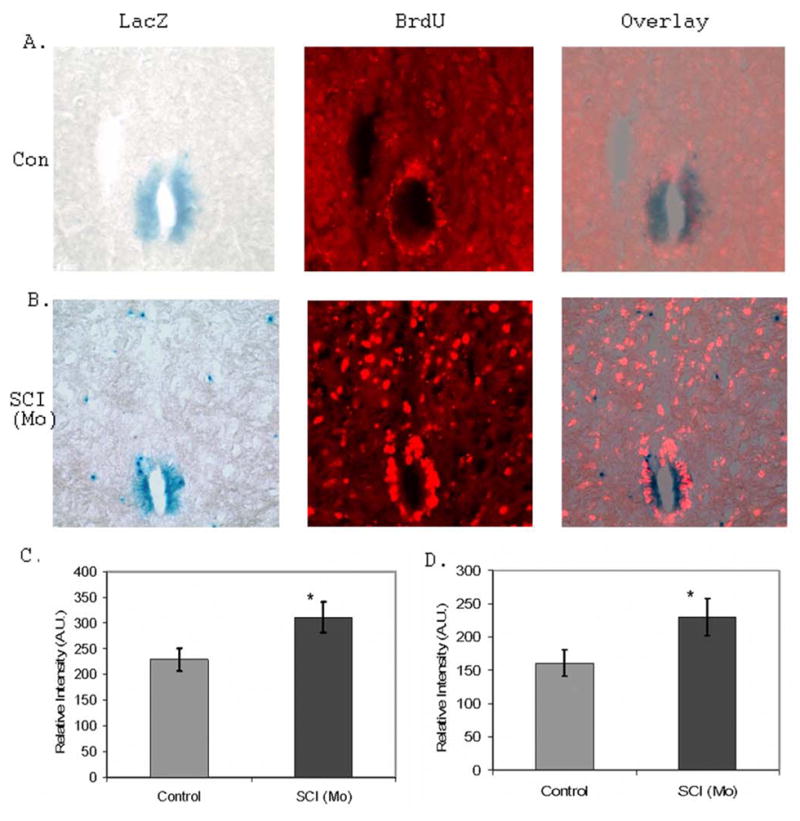
A. The representative LacZ staining and BrdU labeling of NPCs in the ependymal zone (EZ) of the central canal region in the control mouse. B. The representative LacZ staining and BrdU labeling of NPCs in the EZ of the central canal region in the moderate SCI mice [SCI(Mo)]. C. SCI significantly increased LacZ staining intensity in the EZ of the central canal compared to surgical controls (n=3 mice; 5 sections/mouse; *p<0.05). D. SCI significantly increased BrdU staining intensity in the EZ of the central canal compared to surgical controls (n=3 mice; 5 sections/mouse; *p<0.05).
On the other hand, most of the LacZ positive cells outside of the central canal region did not colocalize with BrdU, suggesting that these cells were not proliferative (Fig.2). However, these cells were highly migratory and could mobilize an immediate response to SCI. The increased number of NPCs in the dorsal horn regions of the SCI mouse spinal cord was largely attributed to the migration of NPCs from the EZ of the central canal region (Fig. 2B).
Migration of NPCs from the ependymal zone (EZ) of the central canal to the dorsal direction in the lesioned mice spinal cords
Two lesion conditions, moderate injury and severe injury were used to analyze NPC migration in response to SCI. We showed that the number of NPCs migrated out from the EZ of the central canal toward the dorsal direction was significant increased in SCI mice compared to that of the surgical control (Fig. 3). More significantly, in addition to the individual NPCs, the large cell aggregates apparently dissociated from the EZ of the central canal also migrated toward the lesion direction under the severe lesion conditions (Fig. 1B and Fig. 3C).
Figure 3. Migration of NPCs from the EZ of the central canal to the lesion region of dorsal horn area after SCI.
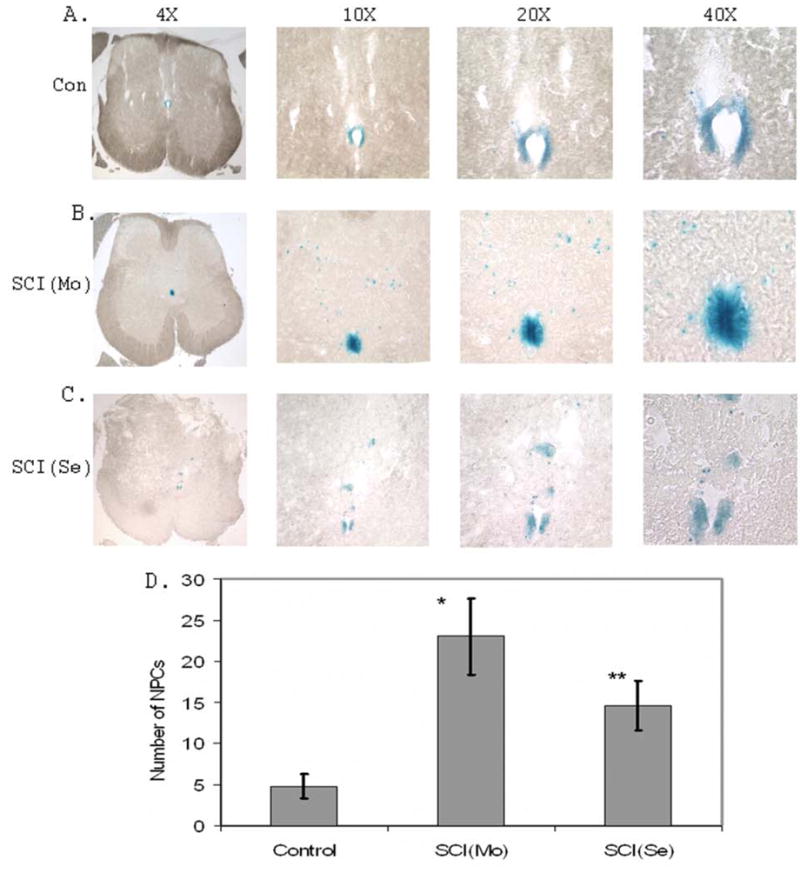
A. Representative spinal cord images of LacZ staining at different magnifications demonstrating the distribution of NPCs in the EZ of the central canal in the surgical control mice. Only a few of NPCs migrate out from EZ of the central canal to the dorsal direction. B. Representative spinal cord images of LacZ staining at different magnifications demonstrating the migration of NPCs from the EZ of the central canal to the dorsal direction in the moderate SCI mice. C. Representative spinal cord image of LacZ staining at different magnifications demonstrating the migration of NPCs from the EZ of the central canal to the dorsal direction in the severe SCI mice. D. Quantification of NPCs migrating out from the EZ of the central canal to the lesion area (n=3 mice; 5 sections/mouse; * p<0.01; **p<0.02 compared to control respectively).
Distribution and organization of NPCs in dorsal and ventral horn regions upon SCI
To study the early response of NPCs to SCI, we also analyzed the distribution and organization of NPCs in the dorsal and ventral horn regions. We demonstrated that the LacZ stained NPCs distributed in all the regions of the adult mouse spinal cord in addition to the EZ of the central canal. There was a polarity distribution of NPCs in the spinal cord. The number of NPCs distributed in the dorsal horn region was far more than in the ventral horn region of the spinal cord (Fig. 4 and Fig. 5). After SCI, the number of NPCs in the dorsal and ventral regions of the spinal cord was significantly increased compared to the specific regions of the surgical control mice (Fig. 4). The quantification of NPCs in the dorsal, ventral and central canal regions of the control and SCI mice were shown in Fig. 4 C. The detailed distribution of NPCs in the dorsal horn regions was shown in Fig. 5.
Figure 4. Distribution of NPCs in the dorsal horn and ventral horn regions of SCI mice compared to surgical control mice.

A. Representative images of LacZ staining demonstrating the distribution of NPCs in the dorsal horn regions of the control and moderate SCI mouse spinal cords. B. Representative images of LacZ staining demonstrating the distribution of NPCs in the ventral horn regions of the control and moderate SCI mouse spinal cords. C. Quantification of NPC distribution in the central canal (C.C), upper central canal (up-C.C.), left dorsal horn (LDH) and right dorsal horn (RDH) of SCI mice compared to surgical control mice (n=3 mice; 5 sections/mouse; *p<0.05; **p<0.02 compared to control respectively). D. Quantification of NPC distribution in the left ventral horn (LVH) and right ventral horn (RVH) of SCI mice compared to surgical control mice (n=3 mice; 5 sections/mouse; *p<0.01; **p<0.05 compared to control respectively).
Figure 5. Analysis of the distribution of NPCs in the dorsal horn regions of SCI mice compared to surgical control mice.
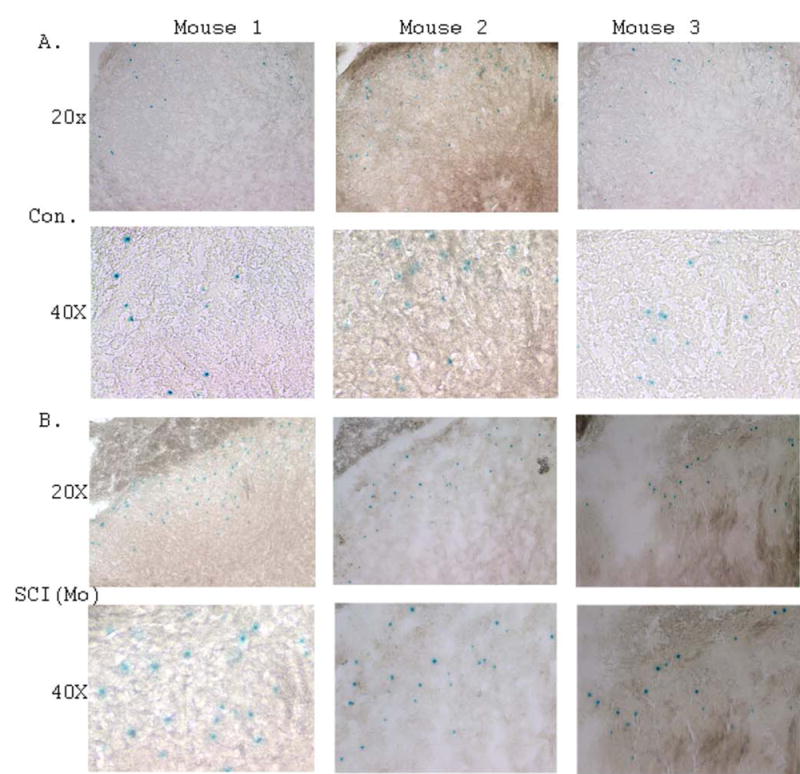
A. Representative images of LacZ staining demonstrating the distribution of NPCs in the dorsal horn regions of surgical control mice at 20x and 40x magnifications. B. Representative images of LacZ staining demonstrating the distribution of NPCs in the dorsal horn regions of moderate SCI mice at 20x and 40x magnifications.
Enhanced neurogenesis from NPCs in response to SCI
The increased proliferation of NPCs in the EZ of the central canal, and enhanced migration of NPCs to the lesioned regions suggest that NPCs may attempt to repair SCI-mediated damage. To further study the potential functionality of the NPCs in response to SCI, we analyzed the possibility of neurogenesis, astrogenesis and oligogenesis from the NPCs adjacent to the lesioned epicenter. We showed that there was an increased of neurogenesis from NPCs determined with neuronal markers, NeuN (Fig. 6) and TuJ1 (data not shown) respectively. In the same way, we also analyzed astrogenesis and oligogenesis from NPCs using specific astrocyte and oligodendrocyte markers. To a large extent, there was no astrogenesis (Fig. 7) and oligogenesis (data not shown) from the NPCs following SCI.
Figure 6. Analysis of neurogenesis of NPCs in the spinal cords of SCI mice compared to surgical control mice.

A. Representative images of LacZ and NeuN staining in surgical control mouse spinal cord. B. Representative images of LacZ and NeuN staining in moderate SCI mouse spinal cord. C. Representative images of LacZ and NeuN staining demonstrating the colocalization of NPCs and neurons in the dorsal horn region of the surgical control mouse spinal cord. D. Representative images of LacZ and NeuN staining demonstrating the colocalization of NPCs and neurons in the dorsal horn region of the moderate SCI mouse spinal cord. E. Quantification of neurogenesis of NPCs in response to SCI compared to surgical control (n=5; *p<0.02).
Figure 7. Analyses of astrogenesis of NPCs in the spinal cord of SCI mouse spinal cords compared with that of surgical control.
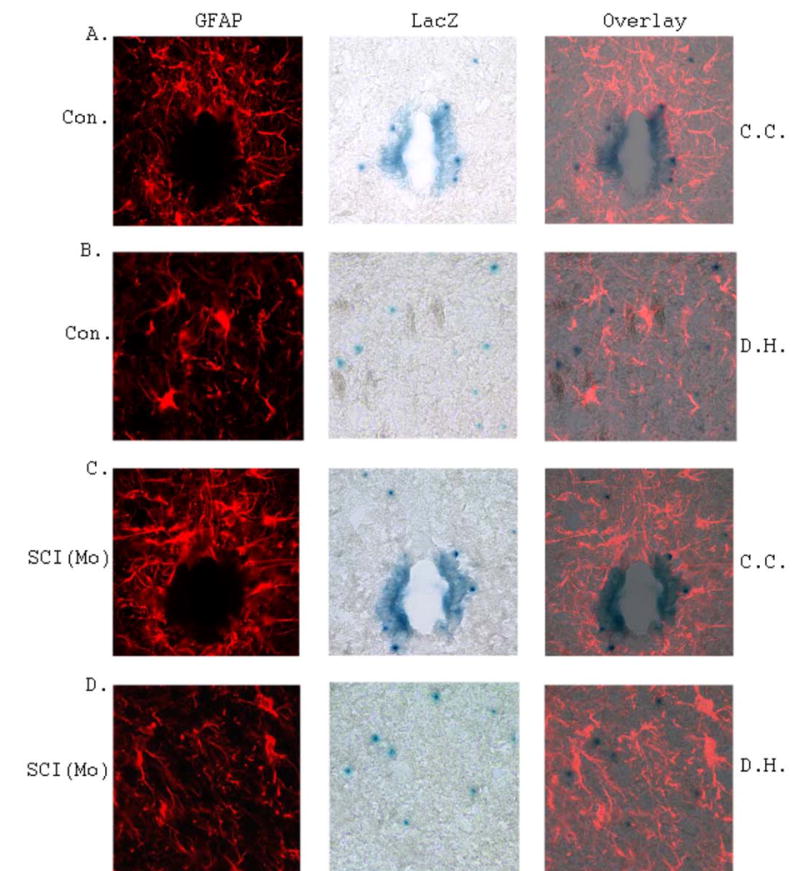
A. Representative images of GFAP and LacZ staining in the central canal (C.C.) region in the surgical control mouse spinal cord. B. Representative images of GFAP and LacZ staining in the dorsal horn (D.H.) region in the surgical control mouse spinal cord. C. Representative images of GFAP and LacZ staining in the central canal (C.C.) region in the moderate SCI mouse spinal cord. B. Representative images of GFAP and LacZ staining in the dorsal horn (D.H.) region in the moderate SCI mouse spinal cord. No astrogenesis was detected from NPCs in response to SCI.
Discussion
Many lines of evidence have demonstrated that NPCs are present in the adult CNS(43,44). More significantly, these NPCs participate actively in normal and neurodegenerative disease-mediated neurogenesis (23–26). Spinal cord injury causes destruction of CNS tissue and death of neurons, astrocytes and oligodendrocytes, leading to neurological dysfunction. The temporal and spatial mechanisms of neuronal cell degeneration by SCI have been well defined (6). However, effective therapy to prevent neuronal cell death and repair neural circuitry remains a major problem. Toward the goal of effective therapy of SCI damage, transplantation of exogenous stem cells/NPCs, to some extent, has achieved a moderate level of neurological function recovery (6–10). Nevertheless, the early responses of endogenous NPCs to SCI remain largely unexplored, although certain levels of neurogenesis have been observed. For this reason, we have focused our effort to analyze the organization, distribution and de novo neurogenesis of NPCs in the pNestin-LacZ transgenic mouse model upon SCI. Nestin has been generally considered a reliable marker for NPCs and has been applied for identification and characterization of NPCs in embryonic and adult CNS. Therefore, we selected pNestin-LacZ reporter transgenic mice to characterize the early response of adult spinal NPCs to SCI. The present study with different lesion conditions demonstrates three major findings: 1). SCI induces NPC proliferation in the EZ of the central canal of the adult mouse spinal cord; 2). SCI promotes NPC migration from the EZof the central canal toward the dorsal horn, where the lesion occurs; 3). SCI increases de novo neurogenesis from NPCs in and adjacent to the lesioned regions. The enhanced proliferation, migration and neurogenesis of the endogenous adult spinal cord NPCs in response to acute SCI during the early phase suggest that adult endogenous NPCs can be potentially utilized for functional recovery.
The early response of adult NPCs to traumatic SCI can be divided into three closely related stages as we have defined in the mouse model, i.e., NPC proliferation, migration and differentiation. In light of the above-mentioned findings, we have first identified and characterized one proliferative population of NPCs which were labeled with BrdU and colocalized with LacZ staining in the adult mouse spinal cord after SCI. Notably, the LacZ staining intensity and BrdU staining intensity were increased in the EZ of the central canal region as the degree of injury was increased, compared with that of surgical controls. The LacZ staining and BrdU labeling together suggested that there was an increase of NPC proliferation in response to SCI (Figs. 1 and 2). The proliferative NPCs identified with 6 to 7 days of BrdU labeling were primarily restricted in the EZ of the central canal region (Fig. 2A and 2B). In addition, we have demonstrated that SCI promoted migration of NPCs from the EZ of the central canal region to the lesioned dorsal horn area. Interestingly, most of the NPCs distributed outside of the EZ of the central canal were not labeled with BrdU, suggesting they were not proliferative. The NPCs cells located outside of the EZ of the central canal tend to migrate toward the lesioned area first. Apparently, the NPCs that migrated out of the central canal lost proliferative ability, because they are not labeled with BrdU (Fig. 2).
Compared with the surgical control mice, migration of NPCs from the EZ of the central canal towards the lesioned dorsal regions was greatly enhanced. Several findings related to adult NPC proliferation and migration in the early response to SCI are worth of mentioning. First, increased proliferation and migration of NPCs occurred as early as 6 hours after SCI (data not shown). By 24 hours after SCI, there was a dramatic increase in the number of NPCs in the dorsal horn region (Figs. 4 and 5). Second, the migratory path of NPCs in response to SCI was toward the dorsal horn direction, where the lesion occurred. In surgical control mice, there were only a few NPCs migrated out from the EZ of the central canal. In contrast, there was a dramatic increase in the number of NPCs that migrated out toward the lesion direction. Third, there is a polarity of NPC distribution in the spinal cord. In addition to the NPCs in the EZ of the central canal, majority of the NPCs located in the dorsal horn region (Lamina I, II and III regions), only a few of NPCs distributed in the ventral region (Fig. 4). The mechanism of the polar organization and distribution of NPCs remain largely unknown. In response to spinal cord injury, there was an increase in the number of NPCs to both dorsal and ventral horn regions (Figs. 4 and 5). Based on the analysis of the organization and distribution of NPCs, the increased number of NPCs in the dorsal and ventral regions was from the EZ of the central canal. By BrdU labeling and LacZ staining, the proliferative NPCs are primarily located in the central canal, while the non-proliferative NPCs were distributed unevenly across spinal cord. Understanding the organization and distribution of NPCs in response to SCI is important to the functional recovery of SCI damage by stimulating endogenous NPCs for regeneration.
The current study also provides important insights into the regenerative potential of adult NPCs toward neuronal direction in response to SCI. We have demonstrated that there was an increase of neurogenesis, but not astrogenesis and oligogenesis from endogenous NPCs in the mouse model of compression SCI. We demonstrated that about 26% of the endogenous adult NPCs in the dorsal horn region adjacent to the lesioned area differentiate toward neurons by immunohistochemical staining with neuronal markers, NeuN (Fig. 6) and Tuj1(data not shown) respectively. The observation of increased neurogenesis from NPCs in the lesioned spinal cord suggests that the nestin-positive adult NPCs may contribute to functional replacement. The early response of enhanced neuronal differentiation from NPCs in SCI supported previous findings that traumatic and ischemic brain injury, and neuronal cell death in neurodegenerative diseases promoted cortical, hippocampus and striatum neurogenesis in animal models (27–30). On the other hand, the current report was different from previous findings that demonstrated gliogenesis, not neurogenesis from endogenous adult neural stem cells (or NPCs), or transplanted NPCs was the major event following SCI (12,14,20). Several factors may be attributed to the differences of the observations in the SCI paradigms. In the current studies, the NPC population responding to SCI was defined by nestin promoter controlled LacZ staining. As demonstrated previously, these NPCs were likely derived from radical glia (45–47). These NPCs apparently have default differentiation toward neuronal direction. On the other hand, the neural stem cells that differentiate into astrocytes or other glial cells upon SCI are likely derived from glial progenitor cells (GPCs) (12–14). Although both types of cells express nestin, they do have different differentiation defaults, neurons vs. glial cells. In the transplantation models, cultured GPCs, neuroepithelial cells, or neural stem cells transplanted into the lesioned spinal cords become glia predominately in response to SCI (11,12). This may reflect that the molecular cues from SCI selectively induce transplanted cells toward glial direction (14). On the other hand, the culture conditions may predetermine the differentiation pattern toward glial lineage. In contrast, the unique population defined by nestin promoter controlled LacZ staining in the adult spinal cord can give rise to neurons predominately (45–47). It becomes clear that the temporal and spatial responses of endogenous adult NPCs were different from that of transplanted cells after SCI. Thus, analyzing the early response of endogenous NPCs to the SCI may offer a therapeutic potential to differentiate specific population of NPCs toward neuronal direction for functional replacement.
In a separate study, we demonstrated that motor neuron degeneration promoted differentiation of adult NPCs toward neuronal cells in the ALS-like mouse spinal cord (data in preparation). Similarly, in the current study, we showed that spinal cord injury enhanced neurogenesis from adult endogenous NPCs in the lesioned spinal cord. The early responses (proliferation, migration and neurogenesis) of adult NPCs to SCI suggest that therapeutic approaches that promote de novo neurogenesis may potentially be useful for repairing neurological circuitry damaged by SCI.
Acknowledgments
This study was supported in part by U.S. Public Health Service Grants AG23923, NS45829, HL75034 and Muscular Dystrophy Association grant 3334 and KSCHIRT grant 1-9A.
Reference List
- 1.Blight AR. Spinal cord injury models: neurophysiology. J Neurotrauma. 1992;9:147–149. doi: 10.1089/neu.1992.9.147. [DOI] [PubMed] [Google Scholar]
- 2.Blight A. Mechanical factors in experimental spinal cord injury. J Am Paraplegia Soc. 1988;11:26–34. doi: 10.1080/01952307.1988.11735792. [DOI] [PubMed] [Google Scholar]
- 3.Juurlink BH, Paterson PG. Review of oxidative stress in brain and spinal cord injury: suggestions for pharmacological and nutritional management strategies. J Spinal Cord Med. 1998;21:309–334. doi: 10.1080/10790268.1998.11719540. [DOI] [PubMed] [Google Scholar]
- 4.Anderson DK, Hall ED. Pathophysiology of spinal cord trauma. Ann Emerg Med. 1993;22:987–992. doi: 10.1016/s0196-0644(05)82739-8. [DOI] [PubMed] [Google Scholar]
- 5.Beattie MS, Farooqui AA, Bresnahan JC. Review of current evidence for apoptosis after spinal cord injury. J Neurotrauma. 2000;17:915–925. doi: 10.1089/neu.2000.17.915. [DOI] [PubMed] [Google Scholar]
- 6.Grossman SD, Rosenberg LJ, Wrathall JR. Temporal-spatial pattern of acute neuronal and glial loss after spinal cord contusion. Exp Neurol. 2001;168:273–282. doi: 10.1006/exnr.2001.7628. [DOI] [PubMed] [Google Scholar]
- 7.Mcdonald JW, Liu XZ, Qu Y, et al. Transplanted embryonic stem cells survive, differentiate and promote recovery in injured rat spinal cord. Nat Med. 1999;5:1410–1412. doi: 10.1038/70986. [DOI] [PubMed] [Google Scholar]
- 8.Akiyama Y, Honmou O, Kato T, Uede T, Hashi K, Kocsis JD. Transplantation of clonal neural precursor cells derived from adult human brain establishes functional peripheral myelin in the rat spinal cord. Exp Neurol. 2001;167:27–39. doi: 10.1006/exnr.2000.7539. [DOI] [PubMed] [Google Scholar]
- 9.Vroemen M, Aigner L, Winkler J, Weidner N. Adult neural progenitor cell grafts survive after acute spinal cord injury and integrate along axonal pathways. Eur J Neurosci. 2003;18:743–751. doi: 10.1046/j.1460-9568.2003.02804.x. [DOI] [PubMed] [Google Scholar]
- 10.Fujiwara Y, Tanaka N, Ishida O, et al. Intravenously injected neural progenitor cells of transgenic rats can migrate to the injured spinal cord and differentiate into neurons, astrocytes and oligodendrocytes. Neurosci Lett. 2004;366:287–291. doi: 10.1016/j.neulet.2004.05.080. [DOI] [PubMed] [Google Scholar]
- 11.Hill CE, Proschel C, Noble M, et al. Acute transplantation of glial-restricted precursor cells into spinal cord contusion injuries: survival, differentiation, and effects on lesion environment and axonal regeneration. Exp Neurol. 2004;190:289–310. doi: 10.1016/j.expneurol.2004.05.043. [DOI] [PubMed] [Google Scholar]
- 12.Cao QL, Zhang YP, Howard RM, Walters WM, Tsoulfas P, Whittemore SR. Pluripotent stem cells engrafted into the normal or lesioned adult rat spinal cord are restricted to a glial lineage. Exp Neurol. 2001;167:48–58. doi: 10.1006/exnr.2000.7536. [DOI] [PubMed] [Google Scholar]
- 13.Setoguchi T, Nakashima K, Takizawa T, et al. Treatment of spinal cord injury by transplantation of fetal neural precursor cells engineered to express BMP inhibitor. Exp Neurol. 2004;189:33–44. doi: 10.1016/j.expneurol.2003.12.007. [DOI] [PubMed] [Google Scholar]
- 14.Setoguchi T, Yone K, Matsuoka E, et al. Traumatic injury-induced BMP7 expression in the adult rat spinal cord. Brain Res. 2001;921:219–225. doi: 10.1016/s0006-8993(01)03123-7. [DOI] [PubMed] [Google Scholar]
- 15.Li Y, Field PM, Raisman G. Repair of adult rat corticospinal tract by transplants of olfactory ensheathing cells. Science. 1997;277:2000–2002. doi: 10.1126/science.277.5334.2000. [DOI] [PubMed] [Google Scholar]
- 16.Martin D, Robe P, Franzen R, et al. Effects of Schwann cell transplantation in a contusion model of rat spinal cord injury. J Neurosci Res. 1996;45:588–597. doi: 10.1002/(SICI)1097-4547(19960901)45:5<588::AID-JNR8>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- 17.Weiss S, Dunne C, Hewson J, et al. Multipotent CNS stem cells are present in the adult mammalian spinal cord and ventricular neuroaxis. J Neurosci. 1996;16:7599–7609. doi: 10.1523/JNEUROSCI.16-23-07599.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Clarke D, Frisen J. Differentiation potential of adult stem cells. Curr Opin Genet Dev. 2001;11:575–580. doi: 10.1016/s0959-437x(00)00235-5. [DOI] [PubMed] [Google Scholar]
- 19.Clarke DL, Johansson CB, Wilbertz J, et al. Generalized potential of adult neural stem cells. Science. 2000;288:1660–1663. doi: 10.1126/science.288.5471.1660. [DOI] [PubMed] [Google Scholar]
- 20.Johansson CB, Momma S, Clarke DL, Risling M, Lendahl U, Frisen J. Identification of a neural stem cell in the adult mammalian central nervous system. Cell. 1999;96:25–34. doi: 10.1016/s0092-8674(00)80956-3. [DOI] [PubMed] [Google Scholar]
- 21.Lie DC, Song H, Colamarino SA, Ming GL, Gage FH. Neurogenesis in the adult brain: new strategies for central nervous system diseases. Annu Rev Pharmacol Toxicol. 2004;44:399–421. doi: 10.1146/annurev.pharmtox.44.101802.121631. [DOI] [PubMed] [Google Scholar]
- 22.Roy NS, Wang S, Jiang L, et al. In vitro neurogenesis by progenitor cells isolated from the adult human hippocampus. Nat Med. 2000;6:271–277. doi: 10.1038/73119. [DOI] [PubMed] [Google Scholar]
- 23.Bedard A, Parent A. Evidence of newly generated neurons in the human olfactory bulb. Brain Res Dev Brain Res. 2004;151:159–168. doi: 10.1016/j.devbrainres.2004.03.021. [DOI] [PubMed] [Google Scholar]
- 24.Bernier PJ, Bedard A, Vinet J, Levesque M, Parent A. Newly generated neurons in the amygdala and adjoining cortex of adult primates. Proc Natl Acad Sci U S A. 2002;99:11464–11469. doi: 10.1073/pnas.172403999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Jin K, Peel AL, Mao XO, et al. Increased hippocampal neurogenesis in Alzheimer's disease. Proc Natl Acad Sci U S A. 2004;101:343–347. doi: 10.1073/pnas.2634794100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Curtis MA, Penney EB, Pearson AG, et al. Increased cell proliferation and neurogenesis in the adult human Huntington's disease brain. Proc Natl Acad Sci U S A. 2003;100:9023–9027. doi: 10.1073/pnas.1532244100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Tattersfield AS, Croon RJ, Liu YW, Kells AP, Faull RL, Connor B. Neurogenesis in the striatum of the quinolinic acid lesion model of Huntington's disease. Neuroscience. 2004;127:319–332. doi: 10.1016/j.neuroscience.2004.04.061. [DOI] [PubMed] [Google Scholar]
- 28.Rice AC, Khaldi A, Harvey HB, et al. Proliferation and neuronal differentiation of mitotically active cells following traumatic brain injury. Exp Neurol. 2003;183:406–417. doi: 10.1016/s0014-4886(03)00241-3. [DOI] [PubMed] [Google Scholar]
- 29.Jin K, Sun Y, Xie L, et al. Directed migration of neuronal precursors into the ischemic cerebral cortex and striatum. Mol Cell Neurosci. 2003;24:171–189. doi: 10.1016/s1044-7431(03)00159-3. [DOI] [PubMed] [Google Scholar]
- 30.Yagita Y, Kitagawa K, Ohtsuki T, et al. Neurogenesis by progenitor cells in the ischemic adult rat hippocampus. Stroke. 2001;32:1890–1896. doi: 10.1161/01.str.32.8.1890. [DOI] [PubMed] [Google Scholar]
- 31.Schwartz PH, Bryant PJ, Fuja TJ, Su H, O'Dowd DK, Klassen H. Isolation and characterization of neural progenitor cells from post-mortem human cortex. J Neurosci Res. 2003;74:838–851. doi: 10.1002/jnr.10854. [DOI] [PubMed] [Google Scholar]
- 32.Yaworsky PJ, Kappen C. Heterogeneity of neural progenitor cells revealed by enhancers in the nestin gene. Dev Biol. 1999;205:309–321. doi: 10.1006/dbio.1998.9035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Fujiwara Y, Tanaka N, Ishida O, et al. Intravenously injected neural progenitor cells of transgenic rats can migrate to the injured spinal cord and differentiate into neurons, astrocytes and oligodendrocytes. Neurosci Lett. 2004;366:287–291. doi: 10.1016/j.neulet.2004.05.080. [DOI] [PubMed] [Google Scholar]
- 34.Takahashi M, Arai Y, Kurosawa H, Sueyoshi N, Shirai S. Ependymal cell reactions in spinal cord segments after compression injury in adult rat. J Neuropathol Exp Neurol. 2003;62:185–194. doi: 10.1093/jnen/62.2.185. [DOI] [PubMed] [Google Scholar]
- 35.Englund U, Bjorklund A, Wictorin K. Migration patterns and phenotypic differentiation of long-term expanded human neural progenitor cells after transplantation into the adult rat brain. Brain Res Dev Brain Res. 2002;134:123–141. doi: 10.1016/s0165-3806(01)00330-3. [DOI] [PubMed] [Google Scholar]
- 36.Roy NS, Wang S, Jiang L, et al. In vitro neurogenesis by progenitor cells isolated from the adult human hippocampus. Nat Med. 2000;6:271–277. doi: 10.1038/73119. [DOI] [PubMed] [Google Scholar]
- 37.Mitsuhashi T, Aoki Y, Eksioglu YZ, et al. Overexpression of p27Kip1 lengthens the G1 phase in a mouse model that targets inducible gene expression to central nervous system progenitor cells. Proc Natl Acad Sci U S A. 2001;98:6435–6440. doi: 10.1073/pnas.111051398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Aoki Y, Huang Z, Thomas SS, et al. Increased susceptibility to ischemia-induced brain damage in transgenic mice overexpressing a dominant negative form of SHP2. FASEB J. 2000;14:1965–1973. doi: 10.1096/fj.00-0105com. [DOI] [PubMed] [Google Scholar]
- 39.Farooque M. Spinal cord compression injury in the mouse: presentation of a model including assessment of motor dysfunction. Acta Neuropathol (Berl) 2000;100:13–22. doi: 10.1007/s004010051187. [DOI] [PubMed] [Google Scholar]
- 40.Xu W, Chi L, Xu R, et al. Increased production of reactive oxygen species contributes to motor neuron death in a compression mouse model of spinal cord injury. Spinal Cord. 2005;43:204–213. doi: 10.1038/sj.sc.3101674. [DOI] [PubMed] [Google Scholar]
- 41.Maslov AY, Barone TA, Plunkett RJ, Pruitt SC. Neural stem cell detection, characterization, and age-related changes in the subventricular zone of mice. J Neurosci. 2004;24:1726–1733. doi: 10.1523/JNEUROSCI.4608-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lie DC, Dziewczapolski G, Willhoite AR, Kaspar BK, Shults CW, Gage FH. The adult substantia nigra contains progenitor cells with neurogenic potential. J Neurosci. 2002;22:6639–6649. doi: 10.1523/JNEUROSCI.22-15-06639.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Emsley JG, Mitchell BD, Kempermann G, Macklis JD. Adult neurogenesis and repair of the adult CNS with neural progenitors, precursors, and stem cells. Prog Neurobiol. 2005 doi: 10.1016/j.pneurobio.2005.04.002. [DOI] [PubMed] [Google Scholar]
- 44.Romanko MJ, Rola R, Fike JR, et al. Roles of the mammalian subventricular zone in cell replacement after brain injury. Prog Neurobiol. 2004;74:77–99. doi: 10.1016/j.pneurobio.2004.07.001. [DOI] [PubMed] [Google Scholar]
- 45.Anthony TE, Klein C, Fishell G, Heintz N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 2004;41:881–890. doi: 10.1016/s0896-6273(04)00140-0. [DOI] [PubMed] [Google Scholar]
- 46.Merkle FT, Tramontin AD, Garcia-Verdugo JM, varez-Buylla A. Radial glia give rise to adult neural stem cells in the subventricular zone. Proc Natl Acad Sci U S A. 2004;101:17528–17532. doi: 10.1073/pnas.0407893101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Hasegawa K, Chang YW, Li H, et al. Embryonic radial glia bridge spinal cord lesions and promote functional recovery following spinal cord injury. Exp Neurol. 2005;193:394–410. doi: 10.1016/j.expneurol.2004.12.024. [DOI] [PubMed] [Google Scholar]