

Abstract
In patients with sickle cell disease, clinical complications including osteonecrosis can vary in frequency and severity, presumably due to the effects of genes that modify the pathophysiology initiated by the sickle mutation. Here, we examined the association of single nucleotide polymorphisms (SNPs) in candidate genes (cytokines, inflammation, oxidant stress, bone metabolism) with osteonecrosis in patients with sickle cell disease. Genotype distributions were compared between cases and controls using multiple logistic regression techniques. An initial screen and follow-up studies showed that individual SNPs and haplotypes composed of several SNPs in bone morphogenic protein 6, annexin A2, and klotho were associated with sickle cell osteonecrosis. These genes are important in bone morphology, metabolism, and vascular disease. Our results may provide insight into the pathogenesis of osteonecrosis in sickle cell disease, help identify individuals who are at high risk for osteonecrosis, and thus allow earlier and more effective therapeutic intervention.
Introduction
Sickle cell anemia results from homozygosity for the Glu6Val mutation in the hemoglobin beta chain gene (HBB). Osteonecrosis is a common sequela of sickle cell disease; studies suggest that by the age of 35 years, one half of all patients with sickle cell anemia have osteonecrosis.1,2 We hypothesize that the presence or absence of osteonecrosis in sickle cell patients is influenced by genetic variability in genes other than HBB that are expressed in either bone or the vasculature. To test this hypothesis, we examined the potential association of osteonecrosis with single nucleotide polymorphisms (SNPs) in candidate genes of different functional classes, including those involved in vascular function, inflammation, oxidant stress, and endothelial cell biology.
Study design
Patients provided informed consent at the time of recruitment in 1987. For this study, samples were obtained from a public repository and were de-identified. This study was approved by the Boston University Institutional Review Board.
DNA samples, clinical information, and demographic information were obtained from the Cooperative Study of Sickle Cell Disease (CSSCD) and have been described elsewhere.3,4 These studies were approved by the Boston University institutional review board. Cases are patients with sickle cell anemia, with or without concurrent α thalassemia with radiologically documented osteonecrosis of the hip and/or shoulder. Controls were patients older than 20 years who received a radiologic exam and did not have a diagnosis of osteonecrosis. We reasoned that any osteonecrosis “genotype” was less likely to be present in older, osteonecrosis-free patients.
Validated candidate gene SNPs with population frequency information and heterozygosity values of more than 0.1 were selected from public databases (http://www.ncbi.nlm.nih.gov/),5 with follow-up studies also using the Celera SNP Reference Database.6 When possible, SNPs in exons, promoters, and sequences important for RNA processing were chosen, however, none of the SNPs is known to affect protein function or gene expression levels. Genotyping was performed using the Sequenom Mass Array/Mass Spectrometer System (Sequenom, San Diego, CA) or the Applied Biosystem TaqMan probes (Applied Biosystems, Foster City, CA). Assay designs can be found in the supplemental data available at the Blood website; see the Supplemental Materials link at the top of the online article.
Approximately 3% of the DNA samples were genotyped twice for quality-control purposes. Hardy-Weinberg equilibrium (HWE) was assessed for each SNP among controls using an χ2 test. SNPs that had more than 25% missing genotypes were not considered in the analysis. This resulted in 233 SNPs being tested for association.
Genotypic counts were compared between sickle cell anemia patients with osteonecrosis (cases) and without osteonecrosis (controls) using multiple logistic regression. In our initial screen, we considered an SNP to have an association with the phenotype when the P value was equal to or less than .01, or if this and other SNPs in the same gene were significant at the .05 level. If an SNP met these criteria, additional SNPs were typed to ascertain haplotypes that could be used to elucidate and better define the pattern of association in a particular gene. Because the large number of tests conducted could inflate the rate of falsely significant associations, the final selection of significant SNPs was carried out by controlling the false discovery rate (FDR) as described previously.7 The selection of individual genotypes was based on 512 tests so that the largest P value to accept a significant association with 20% FDR was .001. The selection of pooled genotypes was based on 233 tests, so that the largest P value to accept a significant association with 20% FDR was .0309.
Pairwise linkage disequilibrium (LD) between SNPs was evaluated using the software Haploview (version 2.05, http://www.broad.mit.edu/mpg/haploview/download.php),8 which implements a maximum likelihood method to infer phase for dual heterozygotes and expresses the magnitude of LD as D′. Haplotypes were inferred using Bayesian methods as implemented in the PHASE computer program (version 2.02; http://www.stat.washington.edu/stephens/software.html).9,10 Haplotype association between cases and controls was assessed using PHASE.
Results and discussion
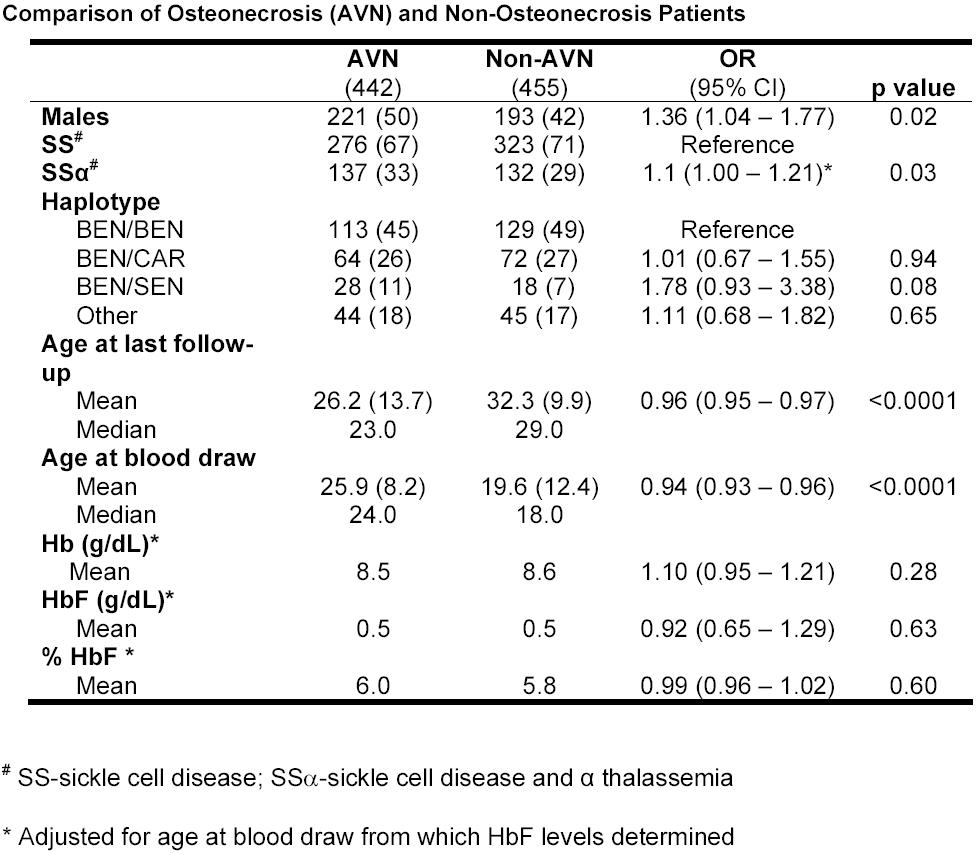
We studied 442 subjects with osteonecrosis and 455 controls. Males had a slightly higher proportion of avascular necrosis (AVN) when compared with females (P = .02); because of the study design, those with osteonecrosis, on average, were 6 years younger than the controls. Individuals with osteonecrosis had a higher prevalence of coincident α thalassemia (P = .03), and there was no difference in total hemoglobin or fetal hemoglobin (HbF) levels (P > .28). See the Supplemental Materials for detailed clinical information.
In the initial screen, 3 to 5 SNPs in 66 candidate genes involved in vascular function, inflammation, oxidant stress, and endothelial cell biology were genotyped, and significant associations were observed with 7 SNPs in 7 genes (BMP6, TGFBR2, TGFBR3, EDN1, ERG, KL, ECE1). Additional SNPs, equally distributed within the gene, were typed in all 7 genes, and a significant association with many SNPs in KL and BMP6 (Table 1) was found. SNPs in ANXA2 were also typed because of a previous finding of association between this gene and stroke among patients with sickle cell disease. In this study, we did not confirm these results in an independent population of sickle cell patients with AVN. Power calculations indicate that, for associated SNPs with an odds ratio of more than 2, examination of 100 additional cases and 200 additional controls would be sufficient to confirm our findings.
Table 1.
SNPs associated with osteonecrosis
| SNP no. | Gene | Genotype | AVN | Control | Reference genotype | AVN | Control | OR | 95% CI | P |
|---|---|---|---|---|---|---|---|---|---|---|
| rs1019856* | TGFBR2 | AA | 55 | 36 | AG | 143 | 156 | 1.75 | 1.08-2.86 | .023 |
| rs934328† | TGFBR2 | CC | 75 | 60 | T_ | 137 | 242 | 2.36 | 1.56-3.51 | < .001 |
| rs284157*† | TGFBR3 | A_ | 209 | 344 | GG | 174 | 65 | 4.95 | 3.53-6.94 | < .001 |
| rs270393*† | BMP6 | GG | 63 | 39 | C_ | 211 | 226 | 1.80 | 1.15-2.81 | .009 |
| rs267196† | BMP6 | TT | 253 | 226 | AT | 78 | 116 | 1.85 | 1.30-2.63 | .001 |
| rs267201 | BMP6 | CT | 161 | 161 | TT | 92 | 142 | 1.60 | 1.13-2.28 | .008 |
| rs449853† | BMP6 | TT | 71 | 47 | C_ | 308 | 338 | 1.68 | 1.12-2.52 | .012 |
| rs1225934† | BMP6 | CC | 308 | 265 | AC | 70 | 123 | 1.96 | 1.41-2.78 | .001 |
| rs212527† | ECE1 | GG | 75 | 22 | A_ | 123 | 196 | 5.68 | 3.33-9.62 | < .001 |
| rs5369*† | EDN1 | AG | 48 | 41 | GG | 57 | 128 | 3.04 | 1.76-5.24 | .001 |
| hCV7464888† | EDN1 | AG | 49 | 61 | AA | 209 | 141 | 2.22 | 1.41-3.45 | .001 |
| rs979091* | ERG | CC | 39 | 13 | AA | 9 | 12 | 3.85 | 1.32-11.10 | .014 |
| rs2836430 | ERG | AA | 373 | 353 | AC | 33 | 59 | 1.96 | 1.22-3.13 | .005 |
| rs7163836 | ANXA2 | GG | 149 | 119 | AA | 44 | 70 | 1.82 | 1.15-2.86 | .010 |
| hCV11770326† | ANXA2 | CC | 227 | 204 | G_ | 55 | 138 | 3.38 | 2.28-4.97 | < .001 |
| rs7170178† | ANXA2 | AA | 164 | 140 | G | 148 | 210 | 1.98 | 1.44-2.72 | < .001 |
| rs1033028† | ANXA2 | T_ | 368 | 368 | GG | 14 | 36 | 2.43 | 1.28-4.57 | .007 |
| hCV26910500† | ANXA2 | G_ | 156 | 122 | AA | 102 | 166 | 2.56 | 1.79-3.68 | < .001 |
| hCV1571628 | ANXA2 | TT | 124 | 138 | AT | 111 | 179 | 1.45 | 1.03-2.08 | .034 |
| rs538874† | STARD13 | G_ | 286 | 331 | AA | 37 | 51 | 1.79 | 1.12-2.85 | .015 |
| rs475303 | STARD13 | CT | 132 | 93 | CC | 278 | 279 | 1.42 | 1.04-1.96 | .029 |
| rs648464† | STARD13 | AG | 193 | 126 | GG | 155 | 185 | 1.91 | 1.39-2.61 | .001 |
| rs480780*† | KL | AC | 69 | 44 | CC | 6 | 5 | 2.97 | 1.83-4.84 | .001 |
| rs211235† | KL | C_ | 150 | 194 | AA | 9 | 46 | 3.97 | 1.81-8.69 | .001 |
| rs2149860† | KL | G_ | 290 | 231 | AA | 134 | 148 | 1.42 | 1.05-1.90 | .021 |
| rs685417† | KL | AG | 199 | 133 | GG | 155 | 164 | 1.66 | 1.21-2.28 | .002 |
| rs516306† | KL | T_ | 411 | 365 | CC | 8 | 19 | 2.86 | 1.19-6.90 | .019 |
| rs565587† | KL | AG | 142 | 113 | AA | 130 | 172 | 1.80 | 1.28-2.53 | .001 |
| rs211239† | KL | A_ | 388 | 314 | GG | 27 | 53 | 2.58 | 1.56-4.28 | .001 |
| rs211234† | KL | G_ | 402 | 333 | AA | 8 | 24 | 4.08 | 1.73-9.62 | .001 |
| rs2238166 | KL | C_ | 408 | 360 | TT | 8 | 16 | 2.50 | 1.02-6.16 | .046 |
| rs499091† | KL | A_ | 368 | 335 | GG | 8 | 19 | 2.85 | 1.18-6.89 | .020 |
| rs576404† | KL | C_ | 411 | 372 | AA | 5 | 16 | 4.23 | 1.40-12.82 | .010 |
| hCV3118898† | APRIN | C_ | 396 | 333 | AA | 26 | 53 | 2.51 | 1.51-4.14 | .001 |
| hCV11710292 | APRIN | AG | 174 | 125 | AA | 225 | 232 | 1.45 | 1.08-1.95 | .014 |
The selection of individual genotypes was based on 512 tests, so that the largest P value to accept a significant association with 20% FDR is .0016. The selection of pooled genotypes was based on 233 tests so that the largest P value to accept a significant association with 20% FDR was .0309. The STARD13 and APRIN genes flank the KL gene.
SNPs that were typed as part of the first, low-density SNP screen for the candidate genes
SNPs that are significant, with 20% FDR
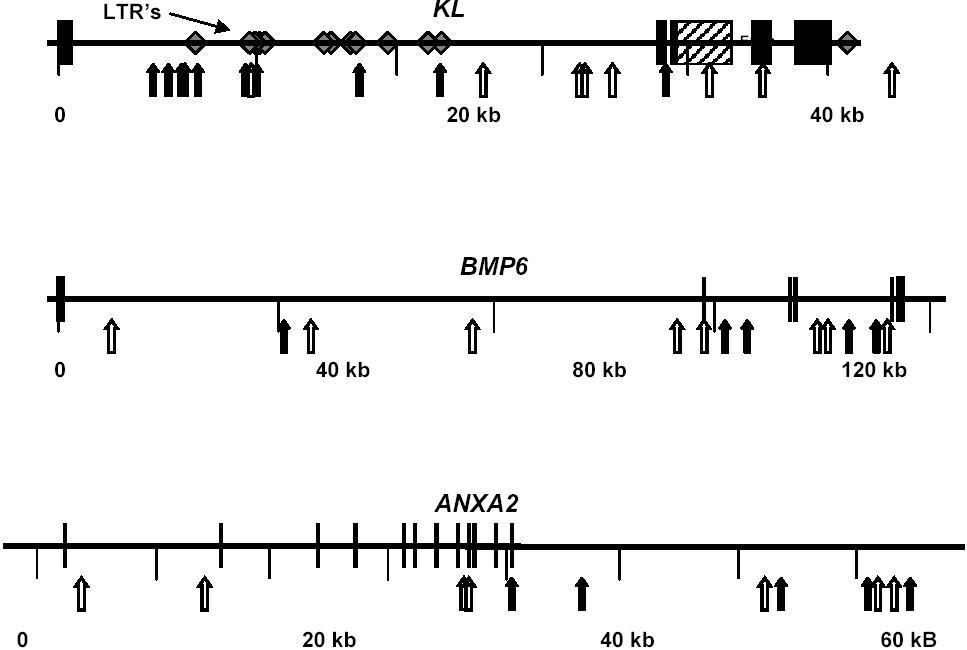
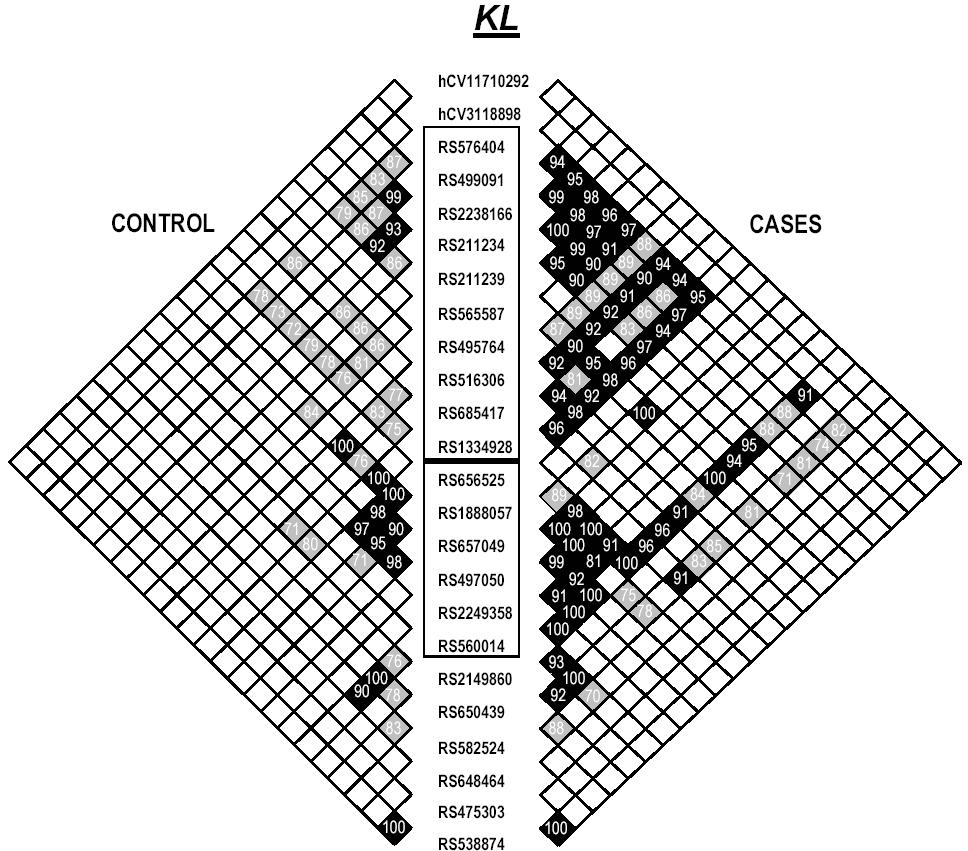
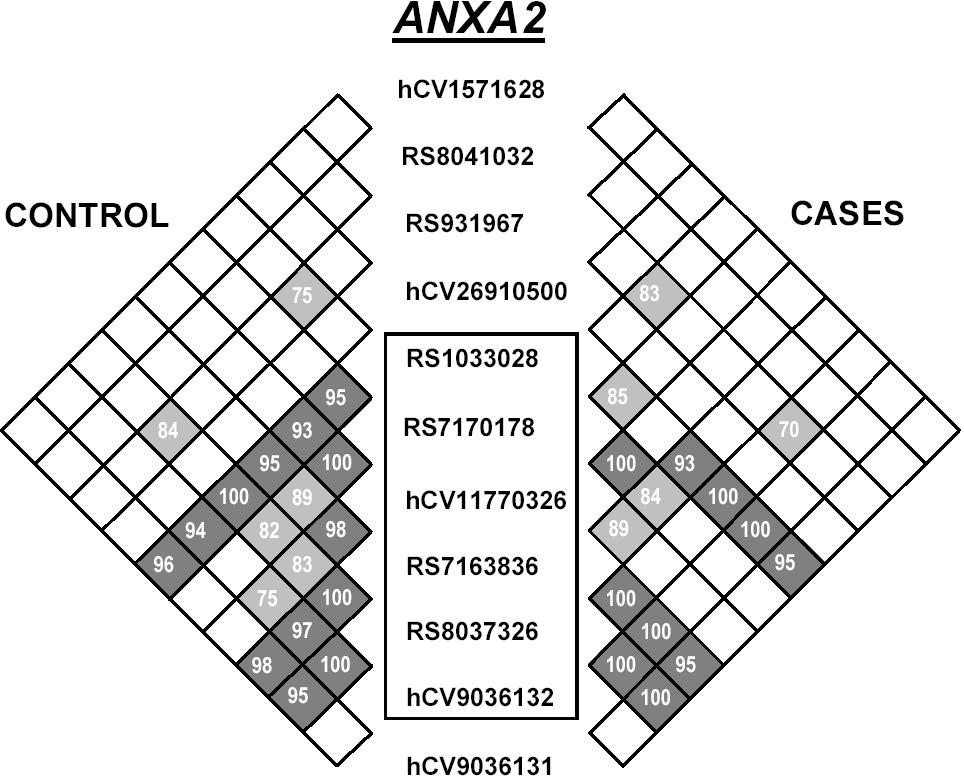
Of the 18 SNPs typed in KL, 10 were significantly associated with osteonecrosis (Table 1). Most of these SNPs were located in the 20-kilobase (kb) region representing the first half of the first KL intron and were in LD with each other. SNPs in BMP6 (5/14) and ANXA2 (6/13) were also associated with osteonecrosis (Table 1); however, these SNPs were distributed throughout the intronic and 3′ untranslated regions of the gene. Similar to the finding in KL, there was a tendency for the disease-associated SNPs to be in LD with each other.
On the basis of visual inspection of the LD pattern in these 3 genes (see Supplemental Materials), 2 haplotype blocks were defined in KL, 1 block in ANXA2, and 1 block in BMP6 (Table 2). For each of these LD blocks, haplotypes were estimated (PHASE) and for all 3 genes the distribution of haplotypes was significantly different among those with and without osteonecrosis (P = .01 for KL and BMP6; P = .03 for ANXA2).
Table 2.
Haplotype analysis of SNP markers in KL, BMP6, and ANXA2
| Control, % | Case, % | |
|---|---|---|
| KL block 1 haplotype* | ||
| CACGAATTGT | 34 | 41 |
| CACGAGATAT | 14 | 21 |
| CACGGAATGT | 12 | 8 |
| AATAGGACAG | 7 | 11 |
| CACGAAATGT | 7 | 5 |
| CACGAATTAT | 3 | 2 |
| CACGAAATAT | 2 | 1 |
| CACGAGTTAT | 2 | 1 |
| CATAGAATGT | 2 | 1 |
| CACGGATTGT | 2 | 1 |
| Rare (< 1%) | 16 | 7 |
| KL block 2 haplotype* | ||
| CCATTC | 41 | 45 |
| ACGCCC | 27 | 32 |
| ATGCCC | 7 | 7 |
| CCATCC | 9 | 5 |
| CCATTT | 4 | 6 |
| ACGCTC | 7 | 3 |
| BMP6 haplotype* | ||
| CTTTCAC | 16 | 12 |
| CTTCCCC | 13 | 13 |
| CTTCCCT | 10 | 10 |
| CTTCTCT | 8 | 9 |
| TACCTCT | 7 | 7 |
| CTTTCCC | 6 | 7 |
| CTTCTCC | 5 | 8 |
| TACCTCC | 6 | 6 |
| CTCCCCC | 4 | 4 |
| TTCCTCT | 3 | 4 |
| CTTTCCT | 3 | 3 |
| TACTCCT | 3 | 2 |
| TACCCCC | 2 | 2 |
| TTCCTCC | 1 | 2 |
| TACCCCT | 1 | 2 |
| Rare (< 1%) | 9 | 9 |
| ANXA2 haplotype† | ||
| GACACG | 28 | 23 |
| TACGCA | 14 | 18 |
| TACACG | 14 | 14 |
| TGCGCG | 12 | 12 |
| TGGGCG | 11 | 9 |
| TGGGGG | 10 | 8 |
| TACGCG | 4 | 8 |
| TGGGCA | 2 | 2 |
| TACGGG | 1 | 3 |
| TGCGCA | 2 | 1 |
| Rare (< 1%) | 3 | 3 |
Haplotypes for KL block 1 are composed of SNPs rs576404, rs499091, rs2238166, rs211234, rs211239, rs565587, rs495764, rs516306, rs685417, and rs1334928. Haplotypes in KL block 2 are composed of SNPs rs656525, rs1888057, rs657049, rs497050, rs2249358, and rs560014. Haplotypes for BMP6 are composed of SNPs rs267192, rs267196, rs267201, rs408505, rs449853, rs1225934, and rs267170. Haplotypes for ANXA2 comprise SNPs rs1033028, rs7170178, hCV11770326, rs7163836, rs8037326, and hCV9036132.
P = .01, global
P = .03, global
The 3 genes we identified are important in bone metabolism. KL encodes a glycosyl hydrolase that participates in a negative regulatory network of the vitamin D endocrine system and may be important for a wide variety of other cellular processes, including regulation of antioxidative defense, angiotensin converting enzyme activity, arteriosclerosis, and aging.11-13 Bone morphogenic proteins (BMPs), including BMP6, are pleiotropic secreted proteins structurally related to transforming growth factor β (TGF-β) and activins. BMP6 is involved in inflammatory processes14 and is important for bone formation15 and, in association with parathyroid hormone (PTH) and vitamin D, appears to be involved in inducing bone development by human bone marrow-derived mesenchymal stem cells.16 ANXA2 is a member of the calcium-dependent phospholipid-binding protein family and regulates cell growth and is involved in signal transduction pathways. It is involved in osteoblast mineralization; lipid rafts containing annexin 2 appear to be important for alkaline phosphatase activity in bone17 and the neuronal response to hypoxia.18
Although we identified genes that may play a significant role in the pathogenesis of sickle cell osteonecrosis by altering protein function or gene expression, the mechanisms by which variants in these genes predispose sickle cell patients to vascular complications are unknown. We also have observed an association between KL and priapism19 as well as between BMP6/ANXA2 and stroke20 in patients with sickle cell disease. This suggests that the vascular complications in sickle disease may have some common underlying molecular basis.
Understanding the genetic risk factors for the development of sickle cell osteonecrosis may provide new insight into the pathogenesis of this disease and eventually provide opportunities for its treatment, which now is limited.21 For example, regulating the activity of the TGF-β pathway to modulate its effects on bone may be possible.22-24 Ultimately, our results should help identify at an early age patients at high risk for osteonecrosis, and thus allow earlier and more effective therapeutic intervention.
Supplementary Material
Acknowledgments
We thank the investigators of the CSSCD who obtained blood samples for DNA-based studies and analyzed data from these studies for the original study publications cited in this paper.
Prepublished online as Blood First Edition Paper, March 22, 2005; DOI 10.1182/blood-2005-02-0548.
Supported by National Heart, Lung, and Blood Institute (NHLBI) grant HL R01 68970 (M.H.S.).
The online version of the article contains a data supplement.
The publication costs of this article were defrayed in part by page charge payment. Therefore, and solely to indicate this fact, this article is hereby marked “advertisement” in accordance with 18 U.S.C. section 1734.
References
- 1.Milner PF, Kraus AP, Sebes JI, et al. Sickle cell disease as a cause of osteonecrosis of the femoral head. N Engl J Med. 1991;325: 1476-1481. [DOI] [PubMed] [Google Scholar]
- 2.Milner PF, Kraus AP, Sebes JI, et al. Osteonecrosis of the humeral head in sickle cell disease. Clin Orthop. 1993;289: 136-143. [PubMed] [Google Scholar]
- 3.Gaston M, Rosse WF. The cooperative study of sickle cell disease: review of study design and objectives. Am J Pediatr Hematol Oncol. 1982;4: 197-201. [PubMed] [Google Scholar]
- 4.Gaston M, Smith J, Gallagher D, et al. Recruitment in the Cooperative Study of Sickle Cell Disease (CSSCD): Controlled Clinical Trials. 1987;8: 131S-140S. [DOI] [PubMed] [Google Scholar]
- 5.National Center for Biotechnology Information. http://www.ncbi.nlm.nih.gov/.
- 6.Celera SNP Reference Database. http://www.appliedbiosystems.com.
- 7.Benjamini Y, Hochberg Y. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Royal Stat Soc Series B-Methodological. 1995;57: 289-300. [Google Scholar]
- 8.Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics. 2005;21: 263-265. [DOI] [PubMed] [Google Scholar]
- 9.Stephens M, Smith NJ, Donnelly P. A new statistical method for haplotype reconstruction from population data. Am J Hum Genet. 2001;68: 978-989. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Stephens M, Donnelly P. A comparison of bayesian methods for haplotype reconstruction from population genotype data. Am J Hum Genet. 2003;73: 1162-1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Tsujikawa H, Kurotaki Y, Fujimori T, Fukuda K, Nabeshima Y. Klotho, a gene related to a syndrome resembling human premature aging, functions in a negative regulatory circuit of vitamin D endocrine system. Mol Endocrinol. 2003;17: 2393-2403. [DOI] [PubMed] [Google Scholar]
- 12.Yahata K, Mori K, Mukoyama M, et al. Regulation of stanniocalcin 1 and 2 expression in the kidney by klotho gene. Biochem Biophys Res Commun. 2003;310: 128-134. [DOI] [PubMed] [Google Scholar]
- 13.Nagai R, Saito Y, Ohyama Y, et al. Endothelial dysfunction in the klotho mouse and downregulation of klotho gene expression in various animal models of vascular and metabolic diseases. Cell Mol Life Sci. 2000;57: 738-746. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rosendahl A, Pardali E, Speletas M, Ten Dijke P, Heldin CH, Sideras P. Activation of bone morphogenetic protein/Smad signaling in bronchial epithelial cells during airway inflammation. Am J Respir Cell Mol Biol. 2002;27: 160-169. [DOI] [PubMed] [Google Scholar]
- 15.Jane JA Jr, Dunford BA, Kron A, et al. Ectopic osteogenesis using adenoviral bone morphogenetic protein (BMP)-4 and BMP-6 gene transfer. Mol Ther. 2002;6: 464-470. [DOI] [PubMed] [Google Scholar]
- 16.Sammons J, Ahmed N, El-Sheemy M, Hassan HT. The role of BMP-6, IL-6, and BMP-4 in mesenchymal stem cell-dependent bone development: effects on osteoblastic differentiation induced by parathyroid hormone and vitamin D(3). Stem Cells Dev. 2004;13: 273-280. [DOI] [PubMed] [Google Scholar]
- 17.Gillette JM, Nielsen-Preiss SM. The role of annexin 2 in osteoblastic mineralization. J Cell Sci. 2004;117: 441-449. [DOI] [PubMed] [Google Scholar]
- 18.Jin K, Mao XO, Eshoo MW, et al. cDNA microarray analysis of changes in gene expression induced by neuronal hypoxia in vitro. Neurochem Res. 2002;27: 1105-1112. [DOI] [PubMed] [Google Scholar]
- 19.Nolan V, Baldwin C, Ma Q, et al. Association of single nucleotide polymorphisms in klotho with priapism in sickle cell anaemia. Br J Haematol. 2004;128: 266-272. [DOI] [PubMed] [Google Scholar]
- 20.Sebastiani P, Ramoni M, Nolan V, Baldwin C, Steinberg M. Genetic dissection and prognostic modeling of a complex trait: overt stroke in sickle cell anemia. Nat Genet. 2005;37: 435-440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lieberman JR, Berry DJ, Mont MA, et al. Osteonecrosis of the hip: management in the 21st century. Instr Course Lect. 2003;52: 337-355. [PubMed] [Google Scholar]
- 22.Inman GJ, Nicolas FJ, Callahan JF, et al. SB-431542 is a potent and specific inhibitor of transforming growth factor-beta superfamily type I activin receptor-like kinase (ALK) receptors ALK4, ALK5, and ALK7. Mol Pharmacol. 2002;62: 65-74. [DOI] [PubMed] [Google Scholar]
- 23.Laping NJ, Grygielko E, Mathur A, et al. Inhibition of transforming growth factor (TGF)-beta1-induced extracellular matrix with a novel inhibitor of the TGF-beta type I receptor kinase activity: SB-431542. Mol Pharmacol. 2002;62: 58-64. [DOI] [PubMed] [Google Scholar]
- 24.Callahan JF, Burgess JL, Fornwald JA, et al. Identification of novel inhibitors of the transforming growth factor beta1 (TGF-beta1) type 1 receptor (ALK5). J Med Chem. 2002;45: 999-1001. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}