

Summary
Gene transcription is a central timekeeping process in animal clocks. In Drosophila, the basic helix-loop helix (bHLH)-PAS transcription factor heterodimer, CLOCK (CLK)/CYCLE(CYC) transcriptionally activates the clock components period (per), timeless (tim), Par domain protein 1 (Pdp1), and vrille (vri) that feedback and regulate distinct features of CLK/CYC function [1]. Microarray studies have identified numerous rhythmically expressed transcripts [2-7], some of which are potential direct CLK targets [7]. Here we demonstrate a circadian function for one such target, a bHLH-Orange repressor CG17100/CLOCKWORK ORANGE (CWO). cwo is rhythmically expressed and levels are reduced in Clk mutants, suggesting that cwo is CLK-activated in vivo. cwo mutants display reduced amplitude molecular and behavioral rhythms with lengthened periods. Molecular analysis suggests CWO acts, in part, by repressing CLK target genes. We propose that CWO acts as a transcriptional and behavioral rhythm amplifier.
Results and Discussion
Only two (out of five) microarray studies had initially identified CG17100 as a rhythmically expressed gene [4, 5]. To test whether CG17100 exhibits robust rhythms, we used real-time quantitative RT-PCR and found significant rhythms in both LD and constant dark (DD) conditions (Figure 1A, B). We also assayed CG17100 in ClkJrk mutants and found that cwo levels are at trough levels, suggesting that CG17100 is a CLK-activated gene (Figure 1C). We identified a remarkable 20 CLK target CACGTG E-box sequences in the 5’ region and in the large first intron, suggesting direct CLK activation (Figure 1D). Given its potential clock function and the presence of an Orange domain, commonly found in bHLH repressors [8], we dubbed it clockwork orange [9].
Figure 1. CG17100/clockwork orange Transcript Rhythms in Drosophila Heads Using Real-Time PCR.
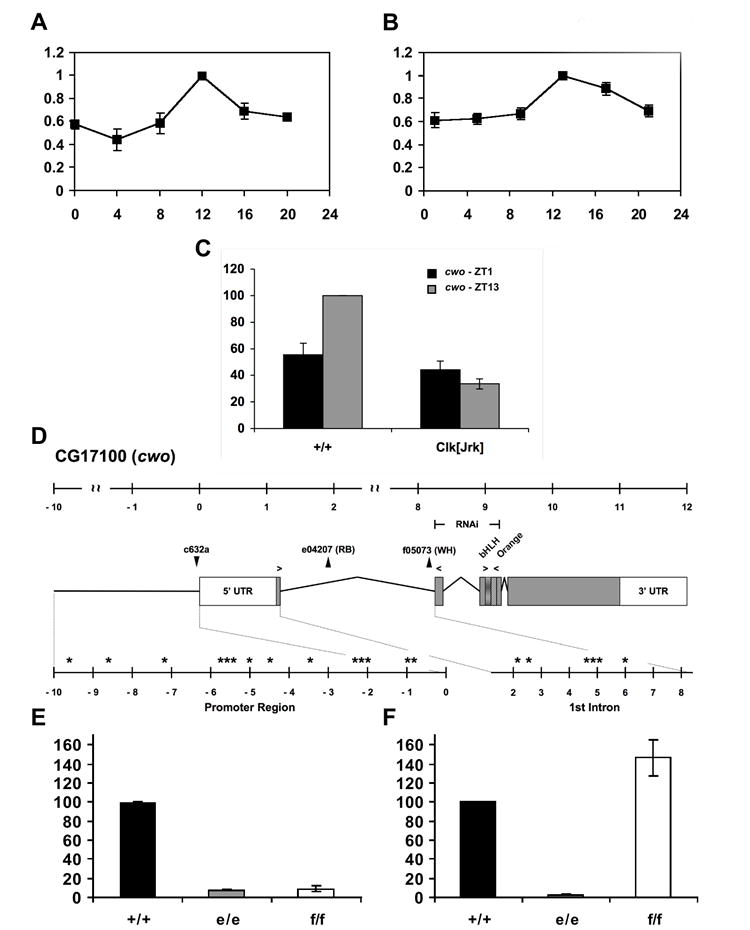
Quantitative real-time RT-PCR (A, B) experiments performed in light-dark (LD; A) and dark-dark conditions (DD; B). X-axis indicates either Zeitgeber Time (ZT; A), where ZT0 is lights-on, or circadian time (CT; B). For real-time experiments (A, B), relative transcript levels have been normalized with the peak level (ZT or CT13) set to 100. (C) Quantitative real-time RT-PCR analysis of cwo transcript levels in wild-type (+/+) and ClkJrk (Clk[Jrk]) mutants at ZT1 and ZT13. Relative transcript levels have been normalized with the WT peak (ZT13) set to 100. (D) Full-length transcript profile and domain organization of wild-type cwo. White and gray boxes indicate untranslated and protein coding regions, respectively. Positions of the various cwo transposon insertions used are shown as black triangles above the diagram. The genomic regions used for the UAS-CWORNAi construct is shown above the transcript profile (labelled as RNAi). Arrows over the diagram denote the location of the primer sets used in real-time PCR experiments. Asterisks indicate the physical location of canonical E-box elements (CACGTG) within the promoter and first intron, as identified by Fly Enhancer (genomeenhancer.org/fly). The figure has been drawn to scale and all units are provided in kilobases. (E,F) cwo transcript levels in wild-type (+/+; =100) and homozygous cwo mutants cwoe (e/e) and cwof (f/f). Primer sets spanning either exons 1 and 2 (E) or exon 3 (F), as shown in (D), were used to measure relative transcript levels. N experiments ≥ 3. Error bars indicate SEM.
We then examined mutants containing transposon insertions in the first cwo intron, cwoe04207 (cwoe) and cwof05073 (cwof; Figure 1D). To determine if these insertions disrupt cwo, we performed qRT-PCR using primers spanning this 7 kb intron. Amplification in homozygous mutants was reduced to ~10% of wild-type (p<0.001; Figure 1E). The only amplicon detected in the mutants was of wild-type size, although we likely failed to detect intron-containing transcripts due to the large size of the potential amplified product (>7 kb). Assaying levels of downstream exon 3 suggests that cwoe-containing transcript destabilizes the un/mis-spliced transcript resulting in reduced levels (Figure 1F). In cwof, reduced apparent transcript amplification across intron 1 (Figure 1E) likely reflects inefficient amplification of the large un/mis-spliced product. Nonetheless, as exon 1 contains the putative initiating ATG (Figure 1D), disruption of splicing between exon 1 and 2 in both cwo mutants would have a dramatic consequence on protein function.
We then tested transheterozygous mutants (cwoe/cwof) as well as homozygous cwo mutants (cwoe/cwoe, cwof/cwof) and observed dramatic reductions in the strength of behavioral circadian rhythms in DD (Figure S1A-D, Table S1 in the Supplementary Data available with this article online; p<0.001). Those flies that demonstrated detectable rhythms often had lengthened circadian periods with reduced strength (Table S1). While rhythmicity is evident immediately upon transfer of flies to DD, rhythms damp in DD (Figure 2C, Figure S1). This phenotype is recessive and excision of either insertion can substantially improve mutant rhythmicity and period (~24 h; Table S1, Figure 2A, Figure S1E; p<0.05). We also performed complementation testing with flies heterozygous for a deletion that removes cwo, Df(3R)ED5495. Flies trans-heterozygous for cwoe and cwof with Df(3R)ED5495 have very poor rhythms, indicating a failure to complement (Table S1, Figure 2A-C, Figure S1F; p<0.001). These data reveal a critical role for cwo in rhythm amplitude and additionally on period length.
Figure 2. Circadian and Diurnal Behavior of cwo Mutants Under Constant Darkness and Light-Dark Conditions.
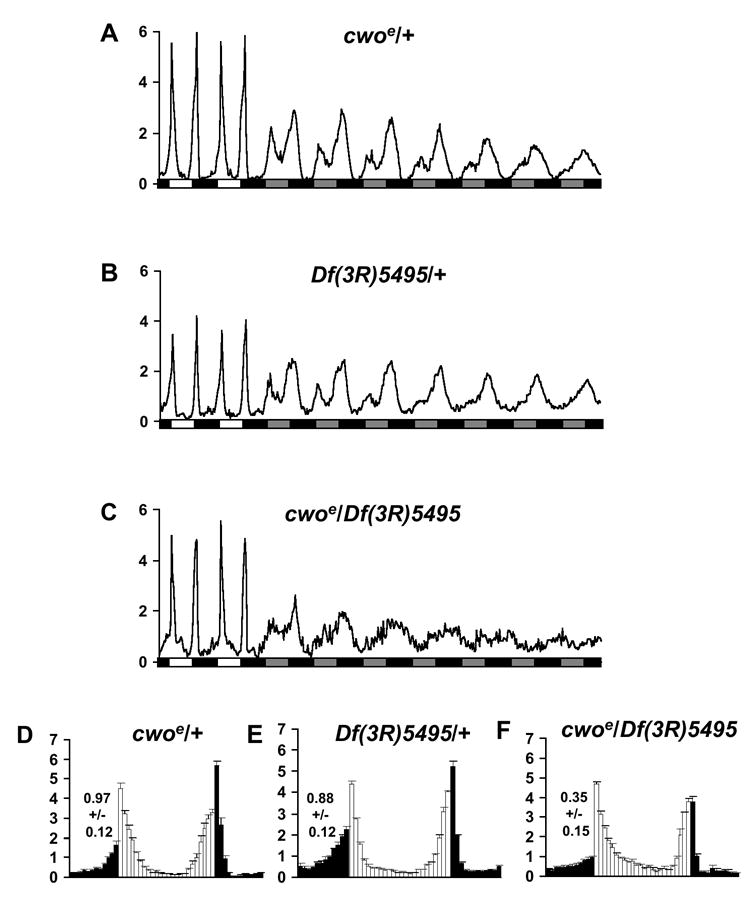
(A-C) Behavior under constant darkness conditions of cwoe/+ (A) (n=80), Df(3R)5495/+ (B) (n=89), and cwoe/Df(3R)5495 (C) (n=43). White and black boxes indicate light and dark periods. Gray boxes indicates subjective day in constant darkness. (D-F) Normalized activity profiles during diurnal conditions of cwoe/+ (D) (n=77), Df(3R)5495/+ (E) (n=79), and cwoe/Df(3R)5495 (F) (n=35). Light and dark bars indicate activity during the light and dark phase, respectively. N experiments= 2-4. Numerical values indicate measures of morning anticipation. Error bars indicate SEM.
Under light-dark conditions, wild-type flies display a morning peak around the time of lights-on and an evening peak around the time of lights-off. Flies increase their activity in anticipation of these transitions, reflecting circadian clock function. Quantitative analysis of morning anticipation indicates a reduction in the degree of anticipation, i.e., the magnitude of the activity increase preceding lights-on, in cwoe/Df(3R)ED5495 relative to heterozygous cwoe/+ or Df(3R)ED5495/+ control flies (p<0.05), consistent with a defect in LD clock function (Figure 2D-F).
To independently confirm the cwo phenotype, we also expressed a dsRNA targeting cwo in transgenic flies (Figure 1D). Expression using the circadian timGAL4 driver, that is expressed in all ~100 pacemaker neurons controlling circadian behavior [10], also resulted in period lengthening phenotypes (Table S2; p<0.001). In contrast, cwo overexpression by timGAL4 resulted in only a modest reduction of rhythmic power when compared to the timGAL4/+ control (Table S2). The RNAi phenotypes are due to specific knockdown of cwo as RNAi directed against GFP (GFPRNAi) does not result in detectable phenotypes (data not shown), the cwoRNAi period phenotype can be partially rescued by wild-type cwo (p<0.001), and cwoRNAi specifically reduces cwo, but not cyc transcript levels, a bHLH family member (Figure S2). Period effects appear to be mediated by a core set of PIGMENT DISPERSING FACTOR (PDF) expressing ventral lateral pacemaker neurons (LNv) as the pdfGAL4 driver leads to lengthened periods in combination with UAScwoRNAi #44 (p<0.001) [11]. pdf promoter driven GAL80 [12], a GAL4 inhibitor, can block the period lengthening effects seen with timGAL4 (p<0.001). These data are consistent with reports that cwo may be expressed specifically in pacemaker neurons [13]. We also analyzed the expression of a GAL4 enhancer trap, c632a [14], inserted just upstream of the cwo transcription start site (Figure 1D) and find expression in the PDF+ LNv (Figure S3).
We next assayed whole head transcriptional oscillations of three CLK target genes, vrille (vri), Pdp1ε, and period (per; see Figure S4), over the first day of constant darkness in cwo mutants (Figure 3A). These whole-head molecular rhythms largely reflect clock function in the eye as opposed to the ~100 brain neurons that drive behavior. In different cwo mutants, we observed elevated vri transcript levels at trough times, CT1 (cwoe/cwof, cwof/cwof only) and CT5 (all cwo mutants tested,p<0.05). In all cwo mutants, we also found reduced transcript levels at the peak CT13 (p<0.05). We observed similar results examining another CLK target, Pdp1ε. Pdp1ε levels exhibited increased trough levels at CT1 and CT5 in cwo mutants (Figure 3B), consistent with a CWO role as a repressor of CLK activated transcripts.
Figure 3. Altered Rhythmic Expression of vri, pdp1ε, and per in cwo Mutants.
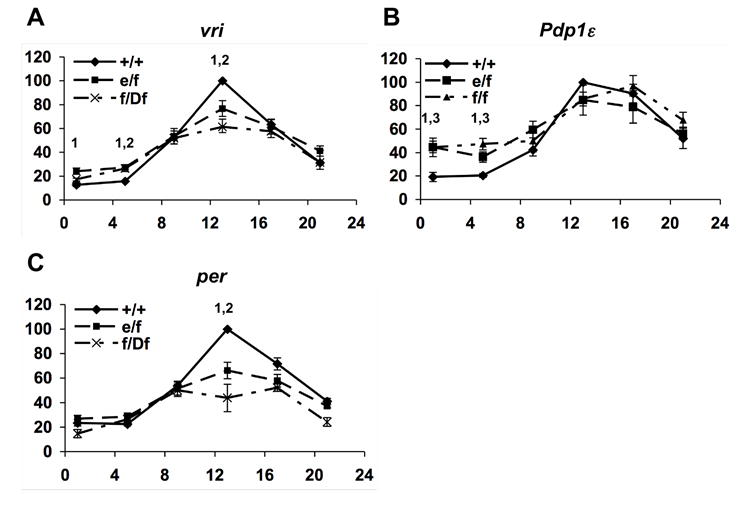
Quantitative real-time RT-PCR analysis of vri (A), pdp1ε (B), and per (C) expression during the first day of DD. X-axis indicates circadian time. Wild-type (+/+) levels at CT 13 (peak) are set to 100 and indicated as a closed line. cwo mutants cwoe/cwof (e/f), cwof/Df(3R)ED5495 (f/Df), and cwof/cwof (f/f) are indicated as dashed lines. Data for f/f not shown for vri but is similar to e/f. Statistical significance (p<.05) is indicated with a “1” for comparing +/+ with e/f, “2” for comparing +/+ with f/Df, and “3” for comparing +/+ with f/f. N experiments ≥ 3 except f/Df CT5,9,17,21, where N=2. Error bars indicate SEM.
per mRNA (Figure 3C) and pre-mRNA levels (Figure S5a) were also altered but with reduced peak levels at CT13 in cwo mutants (p<0.05). Clk expression in cwo mutants is comparable to wild-type indicating that reduced peak per levels cannot be explained by reduced Clk expression (Figure S5b). cwo mutant effects were not evident by DD day 4, although oscillations were also not detectable due to damping of eye clocks (data not shown). The finding of transcript phenotypes on DD day 1 when behavioral phenotypes are subtle suggest that eye clocks may be more sensitive to cwo loss than behaviorally relevant pacemaker neurons. Importantly, two CLK target genes, vri and Pdp1ε, show increased transcript levels at trough times in cwo mutants.
cwo encodes for a bHLH-Orange (bHLH-O) protein that are often DNA binding transcriptional repressors [8]. Interestingly, two proteins implicated in control of mammalian circadian rhythms, Dec1 and Dec2, are also bHLH-O proteins, that can repress mouse Clock action [15]. However, genetic inactivation of Dec1 (also known as Stra13) does not have a core clock phenotype [16]. Definitive tests of the in vivo function of Dec1 and Dec2 will require the analysis of double knockout animals.
To determine if CWO can repress CLK function, we co-transfected cwo and Clk into Drosophila S2 cells. cwo represses CLK activation of several clock gene promoters (Figure 4A; p<0.005). CWO does not directly interact with CLK nor does it significantly affect CLK levels (data not shown). Interestingly, cwo transfection alone also reduced baseline activity of CLK target promoters (Figure 4B; p<0.05). Importantly, CWO does not repress (or only weakly represses) promoters that are not CLK-activated: Clk itself and the heterologous thymidine kinase (TK) promoter (Figure 4B). CWO also selectively represses a CACGTG E-box containing promoter but not other artificial promoters (Figure S6; p<0.005). CWO CWO repression depends on intact CLK target CACGTG sequence (Figure 4C; p<0.005). Fusion of the VP16 activation domain to CWO converts CWO into an E-box dependent transcriptional activator (p<0.001). Gel shift analyses with recombinant CWO indicated specific binding to a CACGTG E box but not mutant E box probes. This binding is partially competed by an unlabeled E-box but not a mutated E-box fragment, and is super-shifted by anti-GST antibodies (Figure 4D). We found similar results using extracts from FLAG-tagged CWO bHLH domain transfected 293T cells (Figure S7). These results suggest that CWO specifically binds to E boxes and represses CLK-activated promoters. Importantly, these in vitro results are consistent with our in vivo data indicating elevated vri and Pdp1ε transcript levels in cwo mutants at times of maximum repression (Figure 3A, B).
Figure 4. CWO Specifically Represses and Binds Clock Gene Promoters.
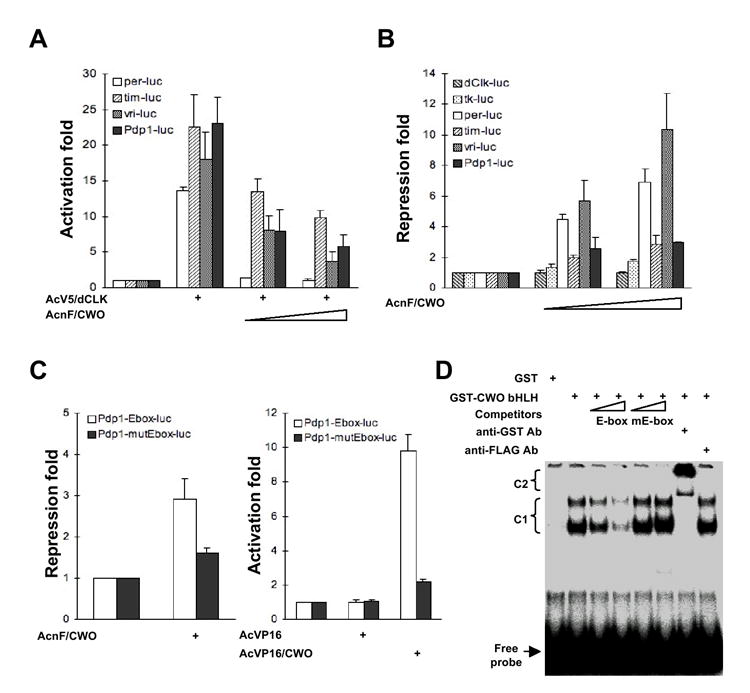
(A, B) S2 cells were cotransfected with reporter plasmids (1 μg except 0.1 μg for vri-mluc) and the increasing amounts (0.25 μg and 1 μg) of expression vector for FLAG-tagged CWO (AcF/CWO) in the presence (A) or absence (B) of expression vector for V5-tagged CLK (AcV5/CLK; 10 ng for per-luc, 0.25 ng for tim-luc and pdp1-mluc, 1 ng for vri-mluc). (C) S2 cells were cotransfected with reporter plasmids (1 μg) and the expression vector for FLAG-tagged CWO or VP16 activation domain-fused CWO (1 μg). Activation fold was calculated by normalizing values to luciferase activity in the presence of reporter plasmid, which was set to 1, while repression fold by inversely normalizing them. For (A-C), N experiments = 3, and standard deviations are depicted by error bars. (D) Electrophoretic mobility shift assay for CWO binding to E boxes (CACGTG). Increasing amount (10- and 50- fold molar excess) of unlabeled competitors containing wild-type (E-box) or mutant E-box (mE-box) or 2 μg of anti-GST or anti-FLAG was preincubated with GST-fusion proteins prior to the addition of labeled probe. C1, shift by GST-CWO bHLH:DNA complex; C2, supershift by GST-bHLH:DNA:anti-GST antibody complex.
Here we have demonstrated an in vivo role for CWO in the Drosophila circadian clock. Our data demonstrate reduced morning anticipation, lengthened periods and damping rhythms in DD. Given that these alleles may not be nulls (Figure 1E,F), we cannot determine definitively if CWO is essential for clock function. Nonetheless, our data argue strongly for a CWO role in driving high amplitude transcriptional oscillations. Indeed, the strength of the observed phenotypes is comparable to or greater than loss-of-function alleles in the PDP1/VRI feedback loop [17-19]. Mechanistic analysis suggests this may be accomplished, in part, by binding to CLK target E-boxes and repressing E-box driven transcription. Two other groups have also found similar in vivo results for CWO (M. Rosbash and H. Ueda, personal communications).
It is interesting to compare the CWO repressor with the well-studied transcriptional repressor PER. Both are rhythmically expressed [20], are CLK-activated in vivo [21] and in turn, repress CLK activation in S2 cells [22], and genetic disruption leads to circadian molecular and behavioral phenotypes. Interestingly, both display differential effects on CLK target genes. In per01, vri and Pdp1ε transcripts are at wild-type peak levels consistent with its proposed repressor function[18], while per transcript or transcription is intermediate between peak and trough [20, 23]. Reduced per transcription has been explained by low Clk levels in per01 but then why do vri and Pdp1ε levels remain at peak levels? In cwo mutants, vri and Pdp1ε transcripts are elevated at trough times while per transcript is reduced only at peak times (Figure 3). One possibility to explain the complexity of per regulation is that full repression by PER and/or CWO may be required to get subsequent full per activation. Alternatively, CWO and/or PER may activate per transcription under some conditions.
The identification of clockwork orange further emphasizes the pivotal role of the Clk gene in the circadian clock. CLK appears to directly activate five clock components, all of which feedback and control Clk gene activity at distinct steps (Figure S3). PER/TIM regulate CLK/CYC DNA-binding [24, 25], PDP1/VRI control Clk transcription [18, 26], while CWO is activated by CLK, and feeds back by binding and repressing through CLK/CYC target sites. Taken together, the multiplicity of feedback controls highlights the central role of Clk, one consistent with a master regulator function [27].
We propose that CWO and CLK are principally involved in regulating pacemaker amplitude [28] while the PER/TIM loop plays a pre-eminent role in dictating period or phase of the rhythms. Interestingly, our CWO results are similar to those of Clk mutants in both flies and mice in which reduced amplitude circadian rhythms are observed [21, 29, 30]. Mutants in per and tim and their phosphorylation regulators [31] can lead to large (>2 hour) period changes while largely sparing rhythmicity. Given their evolutionary conservation, we predict that genetic inactivation of both Dec1 and Dec2 will reveal similar roles in mammals.
Experimental Procedures
Plasmids
Total RNA from adult fly heads was isolated using the TRIzol reagent (Invitrogen) and reverse-transcribed using the M-MuLV reverse transcriptase according to the manufacturer’s instructions (Roche). The cwo cDNA was amplified by PCR with the appropriate primer set, inserted into pAcnF for N-terminally FLAG-tagged expression in S2 cells, and confirmed by sequencing. It was also inserted into pAcVP16 for VP16 activation domain-fusion protein expression in S2 cells. The cDNA corresponding to the CWO bHLH domain (aa 1-126) was subcloned into pFLAG-CMV2 (Sigma) for N-terminally FLAG-tagged expression in mammalian cells and pGEX4T-1 (Amersham Biosciences) for glutathione-S-transferase (GST)-fusion protein expression in bacteria. The dClk cDNA [24] were similarly inserted into pAc5/V5-His (Invitrogen) for V5- and His-tagged expression in S2 cells. The per-luc, tim-luc, dClk-luc, and tk-luc constructs were described previously[22]. Promoter regions of vri gene (from -2.8 kb to 48 bp relative to the transcriptional start site)[17] and Pdp1 gene (from -3.5 kb to +196 bp relative to the transcriptional start site) were amplified from fly genomic DNA, inserted into pGL3-basic (Promega) which was modified to remove a putative binding site of E4BP-4/VRI/PDP1[18, 32], and designated as vri-luc and pdp1-luc, respectively. Promoter region from -1.0 kb to +196 bp of Pdp1 gene was similarly subcloned and designated as Pdp1-Ebox-luc. This region contains one canonical E-box element (CACGTG) at -758 bp upstream from the transcriptional start site, which was mutated (CCCGGG) in Pdp1-mutEbox-luc construct.
Drosophila stocks
All flies were reared with standard cornmeal-yeast-agar medium at 25°C under LD (12-h light/12-h dark) cycles. pdf-GAL4, tim-GAL4-62, and pdf-GAL80 were described previously[10, 12, 33]. GMR-GAL4, UAS-GFPRNAi [34], UAS-mRFP (myristylated red fluorescence protein) were obtained from Bloomington Drosophila stock center. For CWO overexpression in a transgenic fly via the GAL4/UAS system[35], cwo cDNA was inserted into pUAST which was modified to express N-terminally FLAG-tagged CWO. The RNAi construct for cwo gene was designed according to the genomic cDNA hybrid method[36]. cDNA corresponding to the second and third exons of cwo gene (nt 60-460) and genomic DNA including the cDNA with the internal and adjacent 3’ introns were ligated together into the pUAST. Transgenic constructs were injected with pUCHspπΔ2-3 into w1118 embryos, from which several germ-line transformants were established. UAS-CWO lines contain transgenes on the second chromosome, while two independent UAS-CWORNAi lines (#44 and #58) contain them on the third chromosome.
Behavioral analysis
Locomotor activity of individual male flies was measured using Drosophila Activity Monitors (Trikinetics). Monitoring conditions included LD cycles for 2-5 days, followed by DD cycles for a week. Data were analyzed using ClockLab analysis software (Actimetrics) with the significance level of the χ2 periodogram set to α = 0.05. Flies with a χ2 statistic ≥10 over the significance line were scored as rhythmic. Results from at least two independent experiments were averaged. Normalized activity plots for LD and DD were generated by normalizing the average activity of each individual fly to 1. Flies with little or no activity over the final day of the analysis, or throughout the entire analysis, were considered potentially sick and removed. To calculate Morning Anticipation Index values, we determined the largest two-hour increase in normalized average activity of each genotype over the last 6 hr of the dark phase. For all genotypes, the largest two-hour increase in activity occurred between ZT22-ZT24. Anticipation index values were compared between genotypes and their appropriate controls using one-way ANOVA and Tukey post-hoc tests at a significance level of p=0.05.
Cell culture and transient transfection
Drosophila Schneider 2 (S2) cells were maintained in Shields and Sang M3 insect medium (Sigma) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin (Invitrogen). 293T cells were maintained in Dulbecco’s modified Eagle’s medium (Invitrogen) supplemented with 10% fetal bovine serum. Cells were transiently transfected using the standard calcium precipitation method. The quantity of total DNA used in transfection was kept constant by including an appropriate blank vector. For reporter assays, cells were harvested at 36 h after transfection and luciferase assays were performed according to manufacturer’s instructions (Promega). In parallel, cell extracts used in the luciferase assays were immunoblotted with anti-V5 antibody to routinely monitor the constant expression of V5-tagged dCLK protein.
Quantitative RT-PCR
Fly heads were isolated at the indicated time points and total RNA was isolated using the TRIzol reagent (Invitrogen). After the removal of contaminating genomic DNA by DNase I digestion, it was reverse-transcribed by the M-MuLV reverse transcriptase and oligo(dT) primer according to the manufacturer’s instructions (Promega). Alternatively, total RNA from heads was directly amplified using the QuantiTect SYBR green RT-PCR kit (QIAGEN). Semi-quantitative PCR was performed under non-saturating conditions and PCR products were resolved by 1% agarose gel electrophoresis. The relative amount of clock gene transcripts was quantified as described previously[37]. The amplified regions of clock genes for S2 real-time experiments are the full-length cDNAs of vri, Pdp1, and cyc genes, the C-terminal 2.2 kb cDNA of tim gene, and the N-terminal 1 kb cDNA of dClk gene. For quantitative RT-PCR experiments, the following primer sets were used: for rp49, RP49-F: 5’-CGACGCTTCAAGGGACAGTATC-3’, RP49-R: 5’-TTACGACACCAAACGATCGA-3’; for vri (exon 3) - Vri-F: 5′-TGTTTTTTGCCGCTTCGGTCA-3′, Vri-R: 5′-TTACGACACCAAACGATCGA-3′; for Pdp1 (exon 1-2) – Pdp1RD-F: 5’-GAACCCAAGTGTAAAGACAATGCG-3’, Pdp1RD-R: 5’-CTGGAAATACTGCGACAATGTGG-3; for per (exon 1-2) - Per-F: 5′-CAGCAGCAGCCTAATCG-3′, Per-R: 5′-GAGTCGGACACCTTGG-3′; per pre-mRNA (intron 1) – Per[915-936]-F: 5′- AACCCCTACGATTTGGATAGCC -3′, Per[1046-1067]-R: 5’- TGGATAACAGTCGCATAACCCG-3’; Clk (exon 1) – Clk-F: 5’-TACTGCGTGAGGATATCG-3’, Clk-R: 5’-GTTGTTGTTCTGGTTGC-3’; cwo (exon 1)-2 - Cwo-F: 5′-CCCTATTGGAACGAGACGAA-3′, Cwo-R: 5′-GGCATATTCAGCATCGTCCT-3′; cwo (exon 3): Cwo(3)-F: 5’-CCGTATCGAGAAGACGGAGA-3’, Cwo(3)-R: 5’-GCATGTGAACGTCGTAGAGG-3’.
Circadian Cell Expression of cwo
The c632a-GAL4 enhancer trap line (Fly-Trap, J. Douglas Armstrong) was identified as having circadian expression by eGFP as part of an unrelated behavioral screen. The insertion site of the GAL4 was then mapped to the promoter region of cwo using inverse PCR (Dr. Eric Spana, Model Systems Genomics Group, Duke University). 5-7 day old male flies expressing c632a-GAL4 driven UAS-eGFP were dissected, fixed using 3.7% paraformaldehyde, and then immunostained using rabbit-anti-pigment dispersing factor primary antibody[38] at a 1:10,000 concentration, and donkey-anti-rabbit Alexa Fluor 594 secondary antibody (Invitrogen, Molecular Probes) at a 1:500 concentration. Whole mount brains were mounted in 80% ultra-pure glycerol (Invitrogen) and were imaged using laser scanning confocal microscopy (Nikon).
Electrophoretic Mobility Shift Assay
293T cells in 6-well plates were transfected with blank vector or expression vector for FLAG-tagged CWO bHLH domain (aa 1-126). Cells were harvested 24 h after transfection and lysed in hypotonic lysis buffer (10 mM HEPES pH 7.9 and 1.5 mM MgCl2) at 4°C for 15 min. After centrifugation, nuclear proteins were prepared by extracting the pellet with high salt lysis buffer (20 mM HEPES pH 7.9, 500 mM KCl, 10% glycerol, 1.5 mM MgCl2, 0.3 mM EDTA, 0.2 mM dithiothreitol, 0.1% Nonidet P-40, and 1 mM phenylmethylsulfonyl fluoride) at 4°C for 30 min and stored at -70°C before use. Synthetic oligonucleotides containing E-box (5’-AAA GCC GCC GCT CAC GTG GCG AAC TGC GTG-3’) or mutated E-box (mE) (5’-AAA GCC GCC GCT CAG CTG GCG AAC TGC GTG-3’) were labeled with gamma-32P ATP using T4 polynucleotide kinase and annealed to complementary strands. Labeled probe (approximately 200 fmol per reaction) was incubated with 5 μg of nuclear extract or 50 ng of bacterially purified GST-fusion proteins in binding buffer (4 mM HEPES pH 7.9, 100 mM KCl, 2% glycerol, 0.3 mM MgCl2, 0.06 mM EDTA, 0.04 mM dithiothreitol, 0.02% Nonidet P-40, 0.2 mM phenylmethylsulfonyl fluoride, and 1 μg poly[dI-dC]) at room temperature for 30 min. For a competition assay, a 10 or 50 fold molar excess of unlabeled probes containing wild-type or mutated E-box was preincubated with the proteins prior to the addition of labeled probe. For a supershift assay, 2 μg of anti-FLAG antibody (Sigma) or anti-GST antibody (Upstate) was preincubated with the proteins prior to the addition of labeled probe. The reactions were terminated by electrophoresis on a 6% native polyacrylamide gel in Tris-borate-EDTA running buffer. The gel was dried and subjected to autoradiography.
Statistical Analysis
Time point and genotype experiments were performed using one-way ANOVA with Tukey post-hoc tests at significance level p=0.05.
Supplementary Material
Supplemental Data include seven figures and two tables and are available with this article online.
Acknowledgments
We thank Michael Rosbash and Hiroki Ueda for communication of results prior to publication, Monica Villar for expert technical support, Eric Spana (Duke University) for inverse PCR analysis, Terrance Lee for behavioral analysis software improvements, the following for fly strains: Bloomington Drosophila stock center, Harvard Exelixis Drosophila stock collection, J. Douglas Armstrong (c632a) and Michael Rosbash, and the following for clones: Justin Blau (Clk-mluc, tk-mluc), Choogon Lee (Clk cDNA), Charles Weitz (54bp-luc) and Steve Kay (per-luc, tim-luc). This work is supported by the following grants: the Brain Research Center (grant # M103KV010003-06K2201-00310) of the 21st Century Frontier Research Program funded by the Ministry of Science and Technology, the Republic of Korea (J.C.), the Basic Research Promotion Fund (KRF-2005-201-C00035) of the Korea Research Foundation funded by the Ministry of Education and Human Resources Development, the Republic of Korea (J.C.), the Protein Network Research Center at Yonsei University funded by the Ministry of Science and Technology, the Republic of Korea (J.C.), National Institutes of Health (R.A.) and the March of Dimes (R.A.). The authors have no competing interests to report.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Hardin PE. The circadian timekeeping system of Drosophila. Curr Biol. 2005;15:R714–722. doi: 10.1016/j.cub.2005.08.019. [DOI] [PubMed] [Google Scholar]
- 2.Claridge-Chang A, Wijnen H, Naef F, Boothroyd C, Rajewsky N, Young MW. Circadian regulation of gene expression systems in the Drosophila head. Neuron. 2001;32:657–671. doi: 10.1016/s0896-6273(01)00515-3. [DOI] [PubMed] [Google Scholar]
- 3.Wijnen H, Naef F, Boothroyd C, Claridge-Chang A, Young MW. Control of Daily Transcript Oscillations in Drosophila by Light and the Circadian Clock. PLoS Genet. 2006;2:e39. doi: 10.1371/journal.pgen.0020039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Ueda HR, Matsumoto A, Kawamura M, Iino M, Tanimura T, Hashimoto S. Genome-wide transcriptional orchestration of circadian rhythms in Drosophila. J Biol Chem. 2002;277:14048–14052. doi: 10.1074/jbc.C100765200. [DOI] [PubMed] [Google Scholar]
- 5.Lin Y, Han M, Shimada B, Wang L, Gibler TM, Amarakone A, Awad TA, Stormo GD, Van Gelder RN, Taghert PH. Influence of the period-dependent circadian clock on diurnal, circadian, and aperiodic gene expression in Drosophila melanogaster. Proc Natl Acad Sci U S A. 2002;99:9562–9567. doi: 10.1073/pnas.132269699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Ceriani MF, Hogenesch JB, Yanovsky M, Panda S, Straume M, Kay SA. Genome-wide expression analysis in Drosophila reveals genes controlling circadian behavior. J Neurosci. 2002;22:9305–9319. doi: 10.1523/JNEUROSCI.22-21-09305.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.McDonald MJ, Rosbash M. Microarray analysis and organization of circadian gene expression in Drosophila. Cell. 2001;107:567–578. doi: 10.1016/s0092-8674(01)00545-1. [DOI] [PubMed] [Google Scholar]
- 8.Dawson SR, Turner DL, Weintraub H, Parkhurst SM. Specificity for the hairy/enhancer of split basic helix-loop-helix (bHLH) proteins maps outside the bHLH domain and suggests two separable modes of transcriptional repression. Mol Cell Biol. 1995;15:6923–6931. doi: 10.1128/mcb.15.12.6923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Burgess A. A clockwork orange. New York: W.N. Norton and Company; 1962. [Google Scholar]
- 10.Kaneko M, Hall JC. Neuroanatomy of cells expressing clock genes in Drosophila: transgenic manipulation of the period and timeless genes to mark the perikarya of circadian pacemaker neurons and their projections. J Comp Neurol. 2000;422:66–94. doi: 10.1002/(sici)1096-9861(20000619)422:1<66::aid-cne5>3.0.co;2-2. [DOI] [PubMed] [Google Scholar]
- 11.Park JH, Helfrich-Forster C, Lee G, Liu L, Rosbash M, Hall JC. Differential regulation of circadian pacemaker output by separate clock genes in Drosophila. Proc Natl Acad Sci U S A. 2000;97:3608–3613. doi: 10.1073/pnas.070036197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Stoleru D, Peng Y, Agosto J, Rosbash M. Coupled oscillators control morning and evening locomotor behaviour of Drosophila. Nature. 2004;431:862–868. doi: 10.1038/nature02926. [DOI] [PubMed] [Google Scholar]
- 13.Shafer OT, Helfrich-Forster C, Renn SC, Taghert PH. Reevaluation of Drosophila melanogaster’s neuronal circadian pacemakers reveals new neuronal classes. J Comp Neurol. 2006;498:180–193. doi: 10.1002/cne.21021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Armstrong JD. Fly-trap. 2005 www.fly-trap.org.
- 15.Honma S, Kawamoto T, Takagi Y, Fujimoto K, Sato F, Noshiro M, Kato Y, Honma K. Dec1 and Dec2 are regulators of the mammalian molecular clock. Nature. 2002;419:841–844. doi: 10.1038/nature01123. [DOI] [PubMed] [Google Scholar]
- 16.Grechez-Cassiau A, Panda S, Lacoche S, Teboul M, Azmi S, Laudet V, Hogenesch JB, Taneja R, Delaunay F. The transcriptional repressor STRA13 regulates a subset of peripheral circadian outputs. J Biol Chem. 2004;279:1141–1150. doi: 10.1074/jbc.M305369200. [DOI] [PubMed] [Google Scholar]
- 17.Blau J, Young MW. Cycling vrille expression is required for a functional Drosophila clock. Cell. 1999;99:661–671. doi: 10.1016/s0092-8674(00)81554-8. [DOI] [PubMed] [Google Scholar]
- 18.Cyran SA, Buchsbaum AM, Reddy KL, Lin MC, Glossop NR, Hardin PE, Young MW, Storti RV, Blau J. vrille, Pdp1, and dClock form a second feedback loop in the Drosophila circadian clock. Cell. 2003;112:329–341. doi: 10.1016/s0092-8674(03)00074-6. [DOI] [PubMed] [Google Scholar]
- 19.Benito J, Zheng H, Hardin PE. PDP1epsilon functions downstream of the circadian oscillator to mediate behavioral rhythms. J Neurosci. 2007;27:2539–2547. doi: 10.1523/JNEUROSCI.4870-06.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hardin PE, Hall JC, Rosbash M. Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nature. 1990;343:536–540. doi: 10.1038/343536a0. [DOI] [PubMed] [Google Scholar]
- 21.Allada R, White NE, So WV, Hall JC, Rosbash M. A mutant Drosophila homolog of mammalian Clock disrupts circadian rhythms and transcription of period and timeless. Cell. 1998;93:791–804. doi: 10.1016/s0092-8674(00)81440-3. [DOI] [PubMed] [Google Scholar]
- 22.Darlington TK, Wager-Smith K, Ceriani MF, Staknis D, Gekakis N, Steeves TDL, Weitz CJ, Takahashi JS, Kay SA. Closing the circadian loop: CLOCK-induced transcription of its own inhibitors per and tim. Science. 1998;280:1599–1603. doi: 10.1126/science.280.5369.1599. [DOI] [PubMed] [Google Scholar]
- 23.So WV, Rosbash M. Post-transcriptional regulation contributes to Drosophila clock gene mRNA cycling. Embo J. 1997;16:7146–7155. doi: 10.1093/emboj/16.23.7146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lee C, Bae K, Edery I. PER and TIM inhibit the DNA binding activity of a Drosophila CLOCK-CYC/dBMAL1 heterodimer without disrupting formation of the heterodimer: a basis for circadian transcription. Mol Cell Biol. 1999;19:5316–5325. doi: 10.1128/mcb.19.8.5316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Yu W, Zheng H, Houl JH, Dauwalder B, Hardin PE. PER-dependent rhythms in CLK phosphorylation and E-box binding regulate circadian transcription. Genes Dev. 2006;20:723–733. doi: 10.1101/gad.1404406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Glossop NR, Houl JH, Zheng H, Ng FS, Dudek SM, Hardin PE. VRILLE Feeds Back to Control Circadian Transcription of Clock in the Drosophila Circadian Oscillator. Neuron. 2003;37:249–261. doi: 10.1016/s0896-6273(03)00002-3. [DOI] [PubMed] [Google Scholar]
- 27.Zhao J, Kilman VL, Keegan KP, Peng Y, Emery P, Rosbash M, Allada R. Drosophila clock can generate ectopic circadian clocks. Cell. 2003;113:755–766. doi: 10.1016/s0092-8674(03)00400-8. [DOI] [PubMed] [Google Scholar]
- 28.Kim EY, Bae K, Ng FS, Glossop NR, Hardin PE, Edery I. Drosophila CLOCK protein is under posttranscriptional control and influences light-induced activity. Neuron. 2002;34:69–81. doi: 10.1016/s0896-6273(02)00639-6. [DOI] [PubMed] [Google Scholar]
- 29.Allada R, Kadener S, Nandakumar N, Rosbash M. A recessive mutant of Drosophila Clock reveals a role in circadian rhythm amplitude. Embo J. 2003;22:3367–3375. doi: 10.1093/emboj/cdg318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Vitaterna MH, Ko CH, Chang AM, Buhr ED, Fruechte EM, Schook A, Antoch MP, Turek FW, Takahashi JS. The mouse Clock mutation reduces circadian pacemaker amplitude and enhances efficacy of resetting stimuli and phase-response curve amplitude. Proc Natl Acad Sci U S A. 2006;103:9327–9332. doi: 10.1073/pnas.0603601103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Harms E, Kivimae S, Young MW, Saez L. Posttranscriptional and posttranslational regulation of clock genes. J Biol Rhythms. 2004;19:361–373. doi: 10.1177/0748730404268111. [DOI] [PubMed] [Google Scholar]
- 32.Yamaguchi S, Mitsui S, Yan L, Yagita K, Miyake S, Okamura H. Role of DBP in the circadian oscillatory mechanism. Mol Cell Biol. 2000;20:4773–4781. doi: 10.1128/mcb.20.13.4773-4781.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Renn SC, Park JH, Rosbash M, Hall JC, Taghert PH. A pdf neuropeptide gene mutation and ablation of PDF neurons each cause severe abnormalities of behavioral circadian rhythms in Drosophila. Cell. 1999;99:791–802. doi: 10.1016/s0092-8674(00)81676-1. [DOI] [PubMed] [Google Scholar]
- 34.Roignant JY, Carre C, Mugat B, Szymczak D, Lepesant JA, Antoniewski C. Absence of transitive and systemic pathways allows cell-specific and isoform-specific RNAi in Drosophila. Rna. 2003;9:299–308. doi: 10.1261/rna.2154103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. doi: 10.1242/dev.118.2.401. [DOI] [PubMed] [Google Scholar]
- 36.Kalidas S, Smith DP. Novel genomic cDNA hybrids produce effective RNA interference in adult Drosophila. Neuron. 2002;33:177–184. doi: 10.1016/s0896-6273(02)00560-3. [DOI] [PubMed] [Google Scholar]
- 37.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 38.Nitabach MN, Wu Y, Sheeba V, Lemon WC, Strumbos J, Zelensky PK, White BH, Holmes TC, Nitabach MN, Sheeba V, Vera DA, Blau J, Holmes TC. Electrical hyperexcitation of lateral ventral pacemaker neurons desynchronizes downstream circadian oscillators in the fly circadian circuit and induces multiple behavioral periods. J Neurosci. 2006;26:479–489. doi: 10.1523/JNEUROSCI.3915-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Data include seven figures and two tables and are available with this article online.