

Abstract
Given their accessibility, multipotent skin-derived cells might be useful for future cell replacement therapies. We describe the isolation of multipotent stem cell–like cells from the adult trunk skin of mice and humans that express the neural crest stem cell markers p75 and Sox10 and display extensive self-renewal capacity in sphere cultures. To determine the origin of these cells, we genetically mapped the fate of neural crest cells in face and trunk skin of mouse. In whisker follicles of the face, many mesenchymal structures are neural crest derived and appear to contain cells with sphere-forming potential. In the trunk skin, however, sphere-forming neural crest–derived cells are restricted to the glial and melanocyte lineages. Thus, self-renewing cells in the adult skin can be obtained from several neural crest derivatives, and these are of distinct nature in face and trunk skin. These findings are relevant for the design of therapeutic strategies because the potential of stem and progenitor cells in vivo likely depends on their nature and origin.
Introduction
Embryonic, fetal, and adult tissues are used as sources to investigate the developmental and therapeutic potential of stem cells. Because of their accessibility and the possibility that the patient could act as a stem cell donor, adult stem cells from the skin have received particular attention (Slack, 2001). Apart from multipotent epithelial stem cells that form hair follicles, sebaceous glands, and epidermis (Taylor et al., 2000; Oshima et al., 2001; Blanpain et al., 2004; Claudinot et al., 2005) and so-called melanocyte stem cells that generate pigmented cells (Nishimura et al., 2002), a multipotent cell dubbed skin-derived precursor cell (SKP) has been isolated from both the murine and human skin (Toma et al., 2001). SKPs have the potential to produce in vitro cell types normally not found in the skin, such as neuronal cells. Subsequently, several laboratories reported the existence of self-renewing cells present in the skin of mice, pigs, and humans and able to differentiate in vitro into cells expressing neuronal, glial, osteoblast, chondrocyte, smooth muscle, melanocyte, and adipocyte lineage markers (Belicchi et al., 2004; Dyce et al., 2004; Joannides et al., 2004; Sieber-Blum et al., 2004; Amoh et al., 2005; Shih et al., 2005; Toma et al., 2005).
The formation of cells normally not present in skin might be due to transdifferentiation, which describes the conversion of a cell type of a specific tissue lineage into a cell type of another lineage (Wagers and Weissman, 2004). Alternatively, cells from a given lineage might dedifferentiate into a more naive state that allows the cell to redifferentiate along new lineages. Finally, multipotent cells with stem cell features might persist until adulthood, able to generate a broad variety of cells, depending on their environment. To distinguish among these possibilities, the origin and nature of the cell in question has to be determined and its developmental potential has to be analyzed at the single cell level (Wagers and Weissman, 2004).
The developmental origin and exact localization of skin cells giving rise to neural and nonneural progeny is unclear in many of the reported cases. Multipotent skin-derived cells have been enriched by means of markers found on hematopoietic stem cells (Belicchi et al., 2004) or have been isolated from transgenic animals expressing GFP from promoter elements of Nestin (Amoh et al., 2005), a gene also expressed in neural progenitor cells. One source that has been associated with sphere-forming SKPs is the dermal papilla from whisker follicles (Fernandes et al., 2004). Whisker follicles are large hair follicles of the face that serve as sensory organs for a wide range of mammals, excluding humans. Genetic in vivo cell fate mapping revealed that the dermal papilla of these follicles is of neural crest origin (Fernandes et al., 2004). Similarly, culturing explants of bulge and dermal sheath of whisker follicles allowed the identification of neural crest–derived multipotent cells in the upper part of the whisker follicle (Sieber-Blum and Grim, 2004; Sieber-Blum et al., 2004).
A neural crest origin might explain the multipotency of at least some stem and progenitor cells in the skin. Indeed, the neural crest contributes during vertebrate development to a variety of tissues, including the peripheral nervous system and nonneural cell types such as melanocytes in the skin (Le Douarin and Dupin, 2003). Clonal analysis revealed that multipotent, self-renewing neural crest stem cells (NCSCs) cannot only be isolated from migratory neural crest but also from different tissues at later stages and even from the adult organism (Stemple and Anderson, 1992; Bixby et al., 2002; Kruger et al., 2002). Thus, it is conceivable that apart from the whisker follicle, other neural crest–derived compartments in the skin might contain multipotent neural crest–derived cells.
Results
p75/Sox10-positive neural crest–derived cells with stem cell properties can be isolated from the adult murine and human skin
Floating sphere cultures have previously been used to identify self-renewing cells in both murine and human skin (Toma et al., 2001, 2005; Belicchi et al., 2004; Fernandes et al., 2004; Joannides et al., 2004). To further characterize sphere-forming cells derived from the trunk skin of adult mice, dorsal and ventral skin biopsies comprising both dermis and epidermis were dissociated and cultured, and formation of spheres was observed within 4–7 d of culture. These spheres could be passaged for several months without overt morphological changes (Fig. 1 A), pointing to the self-renewing capacity of cells present in the spheres. Intriguingly, unlike SKPs enriched by marker selection (Belicchi et al., 2004) or cultured in slightly different conditions than used here (Fernandes et al., 2004), 100% of all primary, secondary, and later passage spheres generated from mouse trunk skin (n > 50 spheres) contained cells expressing the low-affinity neurotrophin receptor p75 and the transcription factor Sox10, both markers for NCSCs (Fig. 1, C and E; Stemple and Anderson, 1992; Paratore et al., 2001). In spheres passaged >20 times, 67.0 ± 10.5% of all cells expressed p75, 76.6 ± 4.5% of all cells expressed Sox10, and 58.6 ± 10.5% of all cells were double positive for p75 and Sox10. 15.0 ± 6.2% of all cells were negative for these markers, pointing to a cellular heterogeneity within skin-derived spheres, as also observed in sphere cultures from other tissues (Reynolds and Rietze, 2005). Thus, skin-derived cells expressing NCSC markers can be propagated in culture for prolonged time periods.
Figure 1.

Skin-derived spheres contain cells that express the NCSC markers p75 and Sox10. Spheres were generated from murine adult trunk skin (A) and human adult thigh skin (B). Spheres passaged >22 times were allowed to spread on FN-coated plates (C–F) and fixed after 8 h. Immunocytochemical analysis revealed that both murine and human skin–derived spheres contain numerous cells expressing p75 (visualized by Alexa 488 fluorescence, green) and Sox10 (revealed by Cy3 fluorescence, red; E and F). The arrow shows a double-positive cell, and the arrowhead shows a negative cell. (C and D) Corresponding DAPI staining.
Similarly, spheres readily formed from dissociated surgical samples of adult human thigh and face skin (Fig. 1 B). These spheres could be expanded by passaging, such that after 3 mo >109 cells had been generated from a 16-cm2 skin sample used as starting material. Similar to mouse cultures, all spheres contained p75/Sox10-positive cells, which accounted for >60% of all cells (Fig. 1, D and F). However, other markers for premigratory or migratory NCSCs, such as Sox9 and HNK-1, were not expressed.
As p75 and Sox10 are markers for NCSCs (Stemple and Anderson, 1992; Paratore et al., 2001), we next examined whether the mouse trunk skin–derived spheres originate from the neural crest. The fate of neural crest cells was mapped in vivo by mating ROSA26 Cre reporter (R26R) mice, which express β-galactosidase upon Cre-mediated recombination, with mice expressing Cre recombinase under the control of the Wnt1 promoter (Jiang et al., 2000; Lee et al., 2004). In Wnt1-Cre/R26R double-transgenic mice, virtually all NCSCs express β-galactosidase (Brault et al., 2001; Lee et al., 2004). Importantly, despite the transient expression of Cre recombinase, the progeny of neural crest cells continue to express β-galactosidase because of the genomic recombination event. Anti–β-galactosidase antibody staining revealed that all primary and late passage spheres generated from the back skin of adult Wnt1-Cre/R26R double-transgenic mice were composed of neural crest–derived cells (Fig. 2 and not depicted). In particular, 100% of all p75-positive cells coexpressed β-galactosidase, as revealed by a typical punctuated staining pattern (Lutolf et al., 2002). Because 87.3 ± 6.0% of all p75-positive cells also expressed Sox10 (three independent experiments with spheres obtained after 20–35 passages), the data demonstrate that sphere-forming, p75/Sox10-expressing cells from the adult mouse skin are neural crest derivatives.
Figure 2.

Skin-derived spheres are of neural crest origin. Neural crest–derived cells were identified by lineage tracing in spheres generated from Wnt1-Cre/R26R adult mouse skin samples. Primary spheres (not depicted) and spheres at passage 20 were analyzed immunocytochemically for p75 (A, green) and β-galactosidase (B, red) expression. Overlay of A and B reveals that spheres contain cells coexpressing p75 and β-galactosidase (D). (C) Corresponding DAPI staining. Bar, 20 μm.
To test the developmental potential of sphere cells derived from murine and human skin, spheres containing p75/Sox10-positive neural crest cells were allowed to differentiate at high cellular density. The formation of glia expressing glial fibrillary acidic protein (GFAP), βIII tubulin (TuJ1)–positive neuronal cells, and smooth muscle actin (SMA)–expressing nonneural cells was readily detectable in both mouse and human cell cultures (Fig. 3, A–C and G–I), although the number of neuronal cells generated was highly variable and low in comparison to that of glia and smooth muscle cells. Upon addition of ascorbic acid and bone morphogenic protein (BMP) 2, the generation of chondrocytes was observed (Fig. 3, D and J), whereas treatment with stem cell factor and endothelin-3 resulted in formation of a few melanocytes (Fig. 3, E and K). Finally, occasional adipocytes were detected (Fig. 3, F and L). However, we never observed the generation of keratinocytes as assessed by staining with a pan-keratin antibody (unpublished data), demonstrating that neural crest–derived sphere-forming cells are distinct from epithelial stem cells of the skin.
Figure 3.

Cells from skin-derived spheres differentiate into neural crest cell lineages and adipocytes. Late-passage spheres obtained from murine and human skin were allowed to spread at high densities on different substrates and were incubated in media permissive for cell differentiation. Marker expression indicated the generation of GFAP-positive glia (A and G), TuJ1-positive neuronal cells (B and H), and SMA-positive smooth muscle cells (C and I). Some plates (A, B, G, and H) were counterstained with DAPI. Cells with features of chondrocytes, melanocytes, and adipocytes were identified by Alcian blue staining (D and J), DOPA reaction (E and K), or with oil red O (F and L), respectively. Bar, 20 μm.
The aforementioned data are consistent with the idea that skin-derived spheres contain multipotent cells capable of generating neural and nonneural cell types. In analogy to NCSCs isolated from other stages and locations, it is likely that this broad potential is inherent to the p75/Sox10-expressing neural crest–derived cells found in the spheres. To address this hypothesis, we plated cells from mouse trunk skin–derived spheres at clonal density and prospectively identified and mapped single undifferentiated, unpigmented p75-positive clone founder cells (Stemple and Anderson, 1992; Hagedorn et al., 1999; Lee et al., 2004; Kleber et al., 2005). The clone founder cells were then incubated in culture conditions permissive for neurogenesis, gliogenesis, and nonneural cell formation (Stemple and Anderson, 1992). 57.9% of all p75-positive founder cells were at least tripotent, giving rise to clones consisting of neural and nonneural cell types (Fig. 4 A). Virtually no p75-positive cell was restricted to a single cell lineage. Thus, p75/Sox10-positive neural crest–derived cells prepared from the adult trunk skin are multipotent and can be expanded in culture. Upon isolation, these cells therefore exhibit properties of NCSCs.
Figure 4.

Clonal analysis of multipotent neural crest–derived cells from adult skin. Spheres derived from mouse skin were dissociated after passage 17, and single cells were plated at clonal density with or without the addition of instructive growth factors. Cells were prospectively identified with p75 staining and mapped on the plate. A clone was counted positive for a given cell type when it contained at least one cell expressing the appropriate marker (TuJ1, neuronal cells; NG2, glia; SMA, smooth muscle). N, neuronal cells; G, glia; S, smooth muscle; O, other; death, lost clones. (A) Culture without the addition of instructive growth factors was permissive for the generation of heterogeneous clones. Multipotent cells generated three or four different fates. Clones containing exclusively smooth muscle, neuronal, or glia cells were not identified. (B) Cultures containing BMP2 generated S-only clones, S/O, and N/S/O clones. Clone types containing glial cells were significantly decreased. (C) Cultures containing NRG1 also generated S/O and S-only clones. Clone types containing neuronal cells were decreased. (D) TGFβ addition increased the numbers of S/O clones and S-only clones. Each bar represents the mean ± SD of three independent experiments, counting at least 50 clones per experiment.
Several instructive growth factors, including Wnt, BMP, neuregulin (NRG), and TGFβ, have been shown to promote specific fate decisions in NCSCs at the expense of other possible fates. Although Wnt responsiveness is lost at later developmental stages (Kleber et al., 2005), postmigratory NCSCs isolated from various structures maintain their responsiveness to BMP2, NRG1, and TGFβ, although the biological activity of these factors changes with time and location (Bixby et al., 2002; Kruger et al., 2002). Similarly, single prospectively identified p75-positive neural crest cells isolated from the adult back skin were sensitive to BMP2, NRG1, and TGFβ (Fig. 4, B–D). All three instructive growth factors suppressed multipotency without affecting survival of founder cells and promoted the generation of clones containing nonneural cells that were mostly SMA positive. However, we were unable to identify growth factors inducing exclusively neuro- or gliogenesis in skin-derived neural crest cells, whereas NCSCs isolated from other sources give rise to neurons and glia, respectively, in response to BMP2 and NRG1 (Sommer, 2001; Le Douarin and Dupin, 2003). Hence, adult skin–derived neural crest cells, although displaying NCSC features, are intrinsically different from other types of NCSCs and show altered factor responsiveness.
Multiple sources of sphere-forming neural crest–derived cells in the whisker follicle
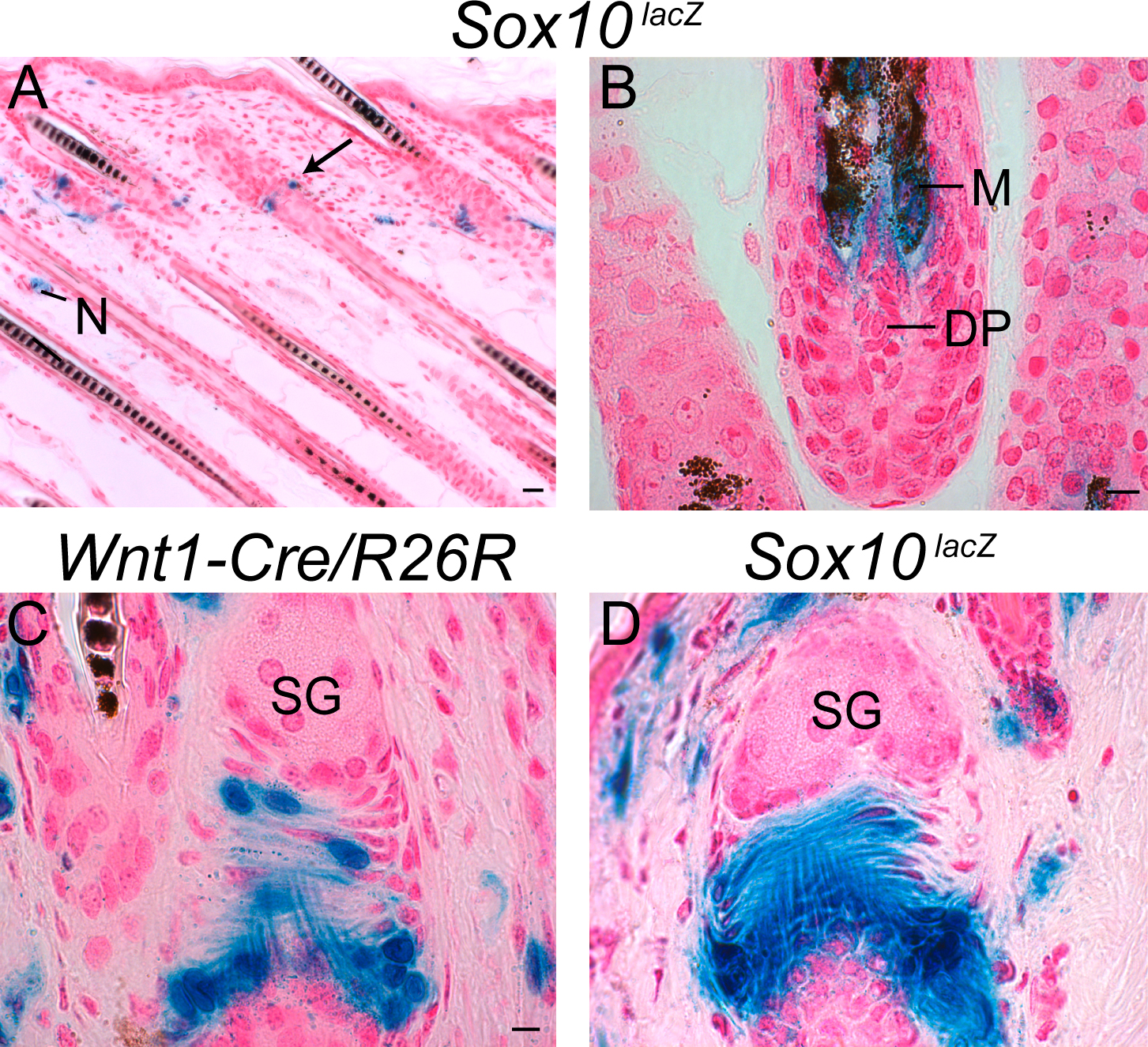
Apart from back skin–derived p75/Sox10-positive multipotent cells (Figs. 1 and 2), the neural crest origin of sphere-forming cells in the adult skin has been demonstrated for whisker follicle–derived SKPs, which, however, are negative for the NCSC markers p75 and Sox10 (Fernandes et al., 2004). This could either reflect differential regulation of NCSC markers in the same cell type because of varying culture conditions or indicate sphere-forming capacity of skin cells from different neural crest derivatives. To address this issue, we first mapped neural crest derivatives in the adult skin and investigated which of these neural crest derivatives express the NCSC marker Sox10 in vivo. We initially focused on the whisker follicle because this structure has been identified before as a source of multipotent neural crest–derived cells (Fernandes et al., 2004; Sieber-Blum et al., 2004). In the head, the neural crest contributes to many mesenchymal structures (Santagati and Rijli, 2003). Thus, many mesenchymal structures in whisker follicles isolated from Wnt1-Cre/R26R double-transgenic mice expressed β-galactosidase (Fig. 5 A). In particular, the capsula, the ringwulst, the dermal sheath, and, as previously published (Fernandes et al., 2004; Sieber-Blum et al., 2004), the dermal papilla turned out to be neural crest derived. The neural crest origin of all these structures was confirmed by fate mapping experiments performed in human tissue plasminogen activator (Ht-PA)-Cre/R26R mice, in which Cre recombinase is expressed in neural crest cells independently from Wnt1 promoter activity (Pietri et al., 2003; Fig. 5 B). As revealed by X-gal staining of whisker follicles isolated from Sox10lacZ mice (that express β-galactosidase from the Sox10 locus; Britsch et al., 2001), capsula, ringwulst, and dermal papilla did not express Sox10 in vivo, whereas the dermal sheath, glial cells in nerve endings, and melanocytes were Sox10 positive (Fig. 5 C). Thus, the whisker follicle comprises various Sox10-positive and -negative tissues of neural crest origin.
Figure 5.

Many mesenchymal structures of whisker hair follicles derive from the neural crest and harbor cells capable of generating spheres. (A) Whisker follicles from Wnt1-Cre/R26R mice were incubated in X-gal solution to reveal neural crest–derived cells in the dermal sheath (DS), capsula (C), dermal papilla (DP), ringwulst (RW), nerve (N), and regions above the sebaceous gland (SG). The sebaceous gland itself was negative. Melanocytes (M) were not or only very weakly X-gal positive, as reported previously (Fernandes et al., 2004). (B) Whiskers dissected from Ht-PA-Cre/R26R mice and treated with X-gal solution. The same structures as in the Wnt1-Cre/R26R follicles were shown to be derived from the neural crest. (C) Whiskers dissected from Sox10lacZ mice and incubated in X-gal solution. Cells in which the Sox10 promoter was active were identified in dermal sheath, nerve, and melanocytes. Capsula, dermal papilla, ringwulst, and sebaceous gland were negative. (D) Different neural crest–derived structures were microdissected from Wnt1-Cre/R26R whisker follicles (A) and cultured in medium permissive for sphere formation. All structures appear to contain cells with the potential to form X-gal–positive spheres. The ringwulst was isolated from rat whisker follicles and therefore not stained with X-gal. Six independent experiments were performed for each structure. Bar, 50 μm.
To investigate which of these neural crest derivatives contain cells with sphere-forming potential, dermal papilla, capsula, the upper part of the dermal sheath (without the bulge), and the lower part of the dermal sheath were isolated from whiskers of adult Wnt1-Cre/R26R double-transgenic mice by microdissection, dissociated, and cultured in the same conditions as used before for trunk skin–derived multipotent neural crest cells. In addition, rat whiskers were used to dissect the ringwulst, which in mice was too small to be isolated without contamination from other tissues. Strikingly, all these whisker follicle structures appear to harbor cells with the capacity to generate spheres (Fig. 5 D). X-gal staining of Wnt1-Cre/R26R mouse cell cultures confirmed that the spheres were neural crest derived. Therefore, neural crest cells with sphere-forming potential are not confined to a particular niche in the whisker follicle.
Glial cells as well as the melanocyte lineage are associated with sphere-forming p75/Sox10-positive cells in the adult back skin
Unlike in the head, the mesenchyme in the trunk is not derived from the neural crest (Santagati and Rijli, 2003), and β-galactosidase expression in back skin of Wnt1-Cre/R26R mice was thus restricted to a few locations (Fig. 6, A and C). The same structures were also labeled in the back skin of Ht-PA-Cre/R26R mice (Fig. 6, B and D). In particular, both in the anagen and telogen stage, X-gal staining was found in the permanent part of the pelage follicle, including the bulge region below the sebaceous gland (Fig. 6, A and B; and Fig. S1, available at http://www.jcb.org/cgi/content/full/jcb.200606062/DC1). This area comprises the location of melanocyte stem cells (Nishimura et al., 2002) and glial cells in nerve endings (Botchkarev et al., 1997). In addition, pigmented melanocytes in the bulb region (the lower part of the hair follicles; Fig. 6, C and D) and nerves expressed β-galactosidase. In contrast, other hair follicle structures such as the dermal papilla, dermal sheath, and the outer and inner root sheaths were X-gal negative and, in the trunk skin, do not originate from the neural crest (Fig. 6, A–D).
Figure 6.

Localization of neural crest–derived and p75/Sox10-positive cells in murine back skin. Back skin from Wnt1-Cre/R26R (A, C, E, G, I, and K) and Ht-PA-Cre/R26R (B, D, F, H, J, and L) mice with pelage follicles in anagen phase was stained with X-gal solution. In both transgenic mouse models, positive cells were localized in nerves (N), in the hair follicle bulge region below the sebaceous gland (arrows; A and B), and in the bulb containing melanocytes (M; C and D). E, F, I, and J show pictures of back skin sections stained with X-gal and either antibodies against p75 or against Sox10 visualized by immunofluorescence. Enlarged areas show X-gal–positive cells in the bulge area coexpressing p75 (G and H) and Sox10 (K and L). The panels to the right represent overlays of X-gal and marker stainings. Note that although p75/X-gal– and Sox10/X-gal–positive cells in the bulge area point to the existence of NCSC-like cells in this location, p75 and Sox10 are not specific for neural crest derivatives in the skin and also label X-gal–negative structures (white arrows). Moreover, not all neural crest–derived cells coexpress p75 and Sox10 (arrowheads). The dermal papilla (DP) was X-gal negative in both mouse lines. Bars: (A, C, E, and I) 20 μm; (G) 10 μm.
To determine the potential origin of Sox10-positive sphere-forming cells in the skin (Fig. 1), we assessed Sox10 expression by virtue of β-galactosidase activity in the back skin of Sox10lacZ mice in vivo. Interestingly, Sox10-expressing cells were confined to exactly the same areas as were X-gal–positive cells in Wnt1-Cre/R26R and Ht-PA-Cre/R26R mice, including nerves, melanocytes, and a domain consistently found below the sebaceous gland in anagen and telogen stage that encompasses the hair follicle bulge with the niche for melanocyte stem cells and nerve endings (Fig. S1). Importantly, in both Wnt1-Cre/R26R and Ht-PA-Cre/R26R mice, X-gal–positive cells in the region below the sebaceous gland coexpressed Sox10 and p75 protein (Fig. 6, E–L). Thus, p75/Sox10-positive multipotent neural crest–derived cells from the trunk skin (Figs. 1–4 ) are connected to the glial or the melanocyte lineage or to both of these lineages.
To elucidate whether p75/Sox10 expression and the capacity to form spheres are associated with glial cells from skin, we made use of desert hedgehog (Dhh)-Cre mice that express Cre recombinase in the peripheral glial lineage from early stages onward, but not in migrating neural crest cells or in neural crest–derived cells of other than glial lineages (Jaegle et al., 2003). β-Galactosidase activity was detectable in nerves and nerve endings in the back skin of adult Dhh-Cre/R26R mice (Fig. 7 A). As predicted from the proposed location of glial cells associated with nerve endings in the hair follicle (Botchkarev et al., 1997), X-gal staining in pelage follicles of Dhh-Cre/R26R mice was confined to a region around the bulge (Fig. 7 A), corresponding to the area that also contains β-galactosidase–expressing cells in Wnt1-Cre/R26R, Ht-PA-Cre/R26R, and Sox10lacZ mice (Fig. 6, A and B; and Fig. S1). In Dhh-Cre/R26R mice, X-gal–labeled cells of the bulge region were also labeled with anti-Sox10 antibody (Fig. 7 C) and anti-p75 antibody (Fig. 7 E). Pigmented melanocytes in the hair follicle bulb were X-gal negative, however, indicating that cells labeled in Dhh-Cre/R26R mice do not give rise to melanocyte s and thus are not related to the melanocyte lineage (Fig. 7 G).
Figure 7.

Lineage tracing of cells from the glial or the melanocyte lineage in murine back skin. Back skin from Dhh-Cre/R26R (A and G) and Dct-Cre/R26R (B and H) mice was stained with X-gal solution to identify progenitors and differentiated cells from the glial and melanocyte lineage, respectively, by lineage tracing. (A) X-gal–expressing cells on sections from Dhh-Cre/R26R mice were localized inside hair follicles in the bulge region below the sebaceous gland (arrow) and in glial cells (N) outside hair follicles. (G) Melanocytes (M) in the hair follicle bulb were not stained. (B) X-gal–expressing cells on Dct-Cre/R26R sections were localized inside hair follicles in the bulge region (arrow). (H) Melanocytes also stained for X-gal in Dct-Cre/R26R skin. Enlarged areas show X-gal–positive cells in the bulge area costained for Sox10 (C and D) and p75 (E and F). Bars: (A and G) 20 μm; (C) 10 μm.
To directly demonstrate that cells from the glial lineage tracked by Dhh-Cre promoter activity possess sphere-forming potential, these cells have to be prospectively identified and freshly isolated. One possibility to achieve this would be by using specific surface antigen markers. However, such markers for the early glial lineage are currently unavailable. Furthermore, nerves present in the skin cannot be isolated by microdissection. Therefore, we used a genetic strategy to prospectively identify and directly isolate cells associated with the glial lineage. Dhh-Cre mice were mated with R26R-EYFP mice that express EYFP upon Cre-mediated recombination (Srinivas et al., 2001). Cells expressing EYFP in the trunk skin of Dhh-Cre/R26R-EYFP double-transgenic mice were isolated by FACS and transferred into medium permissive for sphere formation (Fig. 8, A and B). Although from unselected skin samples >106 cells were used to generate ∼50 spheres (Fig. 1; see Materials and methods), <10,000 cells from both the EYFP-positive and -negative cell fraction were seeded in these experiments, to assess a possible enrichment in the spherogenic potential of FACS-selected cells. In two independent experiments, the EYFP-positive (Fig. 8 A, green frame), but not the EYFP-negative (Fig. 8 A, blue frame), cell population gave rise to spheres. Moreover, acutely fixed primary spheres of EYFP-positive cells were composed of cells expressing both p75 and Sox10 (Fig. 8 C). Thus, p75/Sox10-positive cells related to the glial lineage can be isolated from the skin and form spheres.
Figure 8.

Prospective identification and direct isolation of sphere-forming cells from glial and melanocyte lineages. Sorting of cells from the skin of Dhh-Cre/R26R-EYFP (A) and Dct-Cre/R26R-EYFP mice (E) by FACS. Green and blue regions indicate EYFP-positive and -negative cells, respectively. Mice not carrying the corresponding Cre were used as negative controls for EYFP fluorescence (B and F). Primary spheres derived from EYFP- positive cells sorted from Dhh-Cre/R26R-EYFP (C) or Dct-Cre/R26R-EYFP (G) trunk skin contain cells positive for both Sox10 (red) and p75 (green). (D and H) Quantification (two independent experiments) of the number of spheres formed from EYFP-positive and -negative cells sorted from Dhh-Cre/R26R-EYFP and Dct-Cre/R26R-EYFP skin. Bars, 20 μm.
We next asked whether sphere-forming potential is a common feature of peripheral glia. Therefore, we investigated whether sphere cultures can also be established from adult peripheral nerves. In agreement with others (Toma et al., 2001), we were unable to obtain spheres from cultures of dissociated sciatic and trigeminal nerves from adult mice (unpublished data). Thus, nerves or nerve endings in skin, but not peripheral nerves in general, contain cells with sphere-forming potential.
In cell preparations from the trunk skin of Dhh-Cre/R26R mice, only a fraction of all p75/Sox10-positive cells also expressed β-galactosidase (unpublished data). This could point to inefficient Cre-mediated recombination in Dhh-Cre/R26R mice. Alternatively, sources in the skin other than the glial lineage might yield sphere-forming neural crest–related cells. To address whether spherogenic neural crest–derived cells might be connected to the melanocyte lineage, we traced the fate of trunk skin cells in Dct-Cre/R26R mice (Guyonneau et al., 2002). Dct codes for the enzyme dopachrome tautomerase (also called Trp-2), which is required for melanin synthesis and already expressed in melanocyte stem cells (Nishimura et al., 2002). As expected, β-galactosidase activity in the back skin of Dct-Cre/R26R mice was detectable in melanocytes (Fig. 7 H) and in the hair follicle bulge region corresponding to the location of melanocyte stem cells (Fig. 7 B; Nishimura et al., 2002). Moreover, some X-gal–positive cells in the bulge region also expressed Sox10 (Fig. 7 D) and p75 (Fig. 7 F).
To investigate whether, in addition to cells of the glial lineage, the early melanocyte lineage also comprises undifferentiated neural crest–derived cells with the capacity to generate spheres, we isolated EYFP-expressing cells prospectively identified in the skin of Dct-Cre/R26R-EYFP mice. Intriguingly, in two independent experiments, FACS isolation and culturing of <10,000 cells revealed that only EYFP-expressing (Fig. 8 E, green frame), but not EYFP-negative, cells (Fig. 8 E, blue frame) were able to form spheres (Fig. 8 H). Analysis of acutely fixed primary spheres revealed many cells coexpressing p75 and Sox10, whereas pigmented differentiated melanocytes were absent (Fig. 8 G). These data indicate that the early melanocyte lineage comprises p75/Sox10-positive cells that can be propagated as spheres. Thus, as in whisker follicles of the face, the trunk skin contains more than one source of sphere-forming neural crest–derived cells, namely, cells of the glial and melanocyte lineages.
Discussion
In the present study, we show that cells with NCSC features can be isolated from the adult trunk skin of both mouse and human. Like NCSCs from other embryonic and postnatal sources, these neural crest–derived cells in the skin express p75 and Sox10 and are multipotent, able to generate several neural and nonneural lineages. Moreover, multipotent neural crest–derived cells from the adult skin display a self-renewing capacity, in that mouse and human skin–derived cells can be grown and expanded for months in floating sphere cultures. Intriguingly, in whisker follicles of facial skin, several structures of neural crest origin appear to comprise cells with sphere-forming capacity. In the trunk skin, however, genetic cell fate mapping, p75/Sox10 expression analysis in vivo, and, importantly, prospective identification and direct isolation demonstrate that cells displaying NCSC properties do not reside in mesenchymal structures of hair follicles but, rather, are associated with the melanocyte and glial lineages. Thus, self-renewing neural crest–derived cells from the skin are not confined to a particular niche but can be attributed to distinct locations in face and trunk skin.
Sphere-forming neural crest–derived cells reside in distinct structures of the adult skin
Several reports have described the isolation of multipotent cells from murine and human skin (Belicchi et al., 2004; Dyce et al., 2004; Joannides et al., 2004; Sieber-Blum et al., 2004; Amoh et al., 2005; Shih et al., 2005; Toma et al., 2005), raising the question about the origin of the endogenous cells able to self-renew and to generate multiple cell lineages, including cell types normally not found in the skin. Multipotent cells expressing GFP under the control of a Nestin regulatory element have been isolated from the hair follicle bulge area of transgenic mice and reported to undergo neurogenesis in vivo upon transplantation into the murine subcutis (Amoh et al., 2005). In these mice, Nestin-GFP–expressing cells are associated with the outer root sheath (Li et al., 2003), which, however, does not originate from the neural crest (Figs. 5 and 6). The nature of Nestin-expressing cells in the skin remains to be determined, though, as Nestin-GFP in another transgenic mouse line (Kawaguchi et al., 2001) marks the inner but not outer root sheath (unpublished data). Moreover, Nestin may be widely expressed in multiple structures of human skin (Wang et al., 2006).
The dermal papilla from whisker follicles has been reported to be of neural crest origin and to harbor sphere-forming SKPs that are p75/Sox10 negative (Fernandes et al., 2004). However, many structures of the whisker follicle turned out to be neural crest derived, including the dermal sheath, the ringwulst, and the capsula, apart from the dermal papilla (Fig. 5). Intriguingly, upon microdissection, all these tissues appeared to have cells with sphere-forming potential, although we cannot formally exclude possible contamination of the microdissected material by other structures. Thus, the capacity to self-renew appears to be a widespread feature of neural crest–derived cells in the adult skin of the face. Moreover, at least some of these self-renewing cells are also multipotent, as both SKP spheres from whisker pads and explant cultures of the upper part of whisker follicles with dermal sheath and the bulge region contain multipotent neural crest–derived cells (Fernandes et al., 2004; Sieber-Blum et al., 2004).
It has been proposed that, similar to SKPs from face skin, p75-negative cells isolated from the mouse back skin or the human foreskin are also neural crest derived, although this has not been addressed yet (Fernandes et al., 2004; Toma et al., 2005). Most likely, however, these latter sphere-forming cells are not associated with the dermal papilla of hair follicles, because the human foreskin is devoid of hair follicles. Moreover, using in vivo cell fate mapping, we demonstrate that, unlike in whisker follicles of the face, only few structures of pelage follicles in the trunk skin are actually neural crest derived. Dermal papilla, dermal sheath, and other supportive structures do not appear to be neural crest derivatives (Figs. 5 and 6). This is consistent with studies in chicken and mouse, which showed that mesenchymal tissues in the trunk are not neural crest derived (Santagati and Rijli, 2003). These differences in facial versus trunk neural crest contribution might also be relevant for the study of hair follicle development, given that whisker follicles are a widely used model system to investigate mechanisms regulating follicular cell fates (Alonso and Fuchs, 2003; Gambardella and Barrandon, 2003).
Skin structures harboring multipotent neural crest–derived cells are presumably marked by p75 and Sox10 expression in vivo, given that cells displaying NCSC properties after isolation from the adult trunk skin express these markers. Indeed, in Wnt1-Cre/R26R, Ht-PA-Cre/R26R, Dhh-Cre/R26R, and Dct-Cre/R26R mice, β-galactosidase–expressing cells positive for p75 and Sox10 were found in the bulge region encompassing melanocyte stem cells and glial cells in nerve endings (Botchkarev et al., 1997; Nishimura et al., 2002; Figs. 6 and 7). However, the only way to unambiguously demonstrate that these lineages contain resident multipotent cells with the potential to self- renew is by prospectively identifying such cells in the adult skin and testing their potential upon acute isolation. Because of the lack of specific surface markers, we were not able to use antibodies to directly isolate multipotent cells from back skin. In particular, p75 expression does not distinguish between glial and melanocyte lineages (Fig. 7) and, in addition, is found in regions of the skin that are not neural crest derived, such as the outer root sheath of hair follicles, and, at early stages of hair follicle morphogenesis, the dermal papilla (Fig. 6; Botchkareva et al., 1999; Rendl et al., 2005). Similarly, the melanocyte marker c-Kit is not suitable for isolation of prospective multipotent cells from the melanocyte lineage because it is not expressed in the bulge of anagen hair follicles and because in vivo it is also found in epithelial skin cells not originating from the neural crest (Peters et al., 2002, 2003). Moreover, in preliminary experiments, we failed to obtain p75/Sox10-positive spheres from c-Kit–positive skin cells isolated by FACS (unpublished data). Nonetheless, we were able to identify spherogenic neural crest–derived cells in the skin by using a genetic approach. Thereby, the lineage-specific activity of Dhh-Cre and of Dct-Cre, respectively, combined with a Cre reporter allele (Srinivas et al., 2001), led to EYFP expression in cells from either the glial or the melanocyte lineage. Consistent with a dual origin of multipotent neural crest–derived cells in the trunk skin, EYFP-positive cells isolated from both Dhh-Cre/R26R-EYFP and Dct-Cre/R26R-EYFP mice formed spheres of p75/Sox10-positive cells (Fig. 8). Thus, we conclude that in the trunk skin of mice, spherogenic neural crest–derived cells are associated with the glial as well as the melanocyte lineage. Based on their high similarities to mouse cells in terms of marker expression and potential, and based on the fact that humans do not have whisker follicles, we assume that the p75/Sox10-positive multipotent cells obtained from human skin (Figs. 1 and 3) are also related to glial and melanocyte rather than mesenchymal lineages.
That cells of the bulge region marked in Dct-Cre/R26R mice generate melanocytes is in agreement with earlier findings (Nishimura et al., 2002), which identified DctlacZ-positive cells of the bulge region as so-called melanocyte stem cells generating differentiated melanocytes in the lower part of the hair follicle. Moreover, a considerable fraction of Dct-positive cells in the bulge also expresses Sox10 (Osawa et al., 2005), in agreement with our results (Fig. 7). Our analysis of neural crest–derived cells isolated from Dct-Cre/R26R mice supports the hypothesis that melanocyte progenitors in the bulge region are not only self-renewing (Nishimura et al., 2002) but indeed represent multipotent cells (Sommer, 2005).
The combined data indicate that the adult skin is host to sphere-forming cells with different identities. In addition to multipotent, undifferentiated neural crest–related cells (Fernandes et al., 2004; this study), the potential to self-renew has also been attributed to pigment cells and possibly other developmentally restricted neural crest–derived cell types (Dupin et al., 2000; Trentin et al., 2004). Similar to NCSCs isolated at different time points and from different peripheral nervous system regions (Kleber and Sommer, 2004), the multipotent neural crest–derived skin cells described in this study display altered responsiveness to instructive growth factors as compared with migratory NCSCs (Fig. 4). Thus, multipotent, self-renewing neural crest–derived cells change intrinsic properties with time and location. This presumably reflects their specific functional requirements, although the physiological role of multipotent neural crest–derived cells in the skin remains to be determined. In particular, it is unclear whether these cells display properties of NCSCs only upon transfer into culture or whether they are multifated and self-renewing in vivo as well. Moreover, it will be interesting to elucidate whether and how these cells functionally interact with other, unrelated stem cell types of the skin, such as epithelial stem cells also found in the bulge region of hair follicles (Taylor et al., 2000; Oshima et al., 2001; Blanpain et al., 2004; Claudinot et al., 2005). Finally, it should be addressed whether spherogenic neural crest–derived cells persisting in the adult skin might represent a target for mutational transformation leading to cancers such as melanoma (Pardal et al., 2003).
Limited developmental and therapeutic potential of multipotent neural crest–derived cells from the adult skin
Adult stem cells as an alternative to embryonic stem cells are a prime target of applied research that seeks to treat degenerative diseases by cell replacement therapies (Wagers and Weissman, 2004). The skin might represent an ideal source for adult stem cells in tissue repair because it is easily accessible. Because skin-derived multipotent cells described here are of neural crest origin, their capacity to give rise to neural and nonneural cell types at clonal density (Fig. 4) does apparently not reflect transdifferentiation but, rather, the broad potential inherent to NCSCs. The fact that these cells can be easily expanded in culture, even when isolated from the skin of adult humans, might make them a valuable source for cell replacement therapies because sufficient cell material could be obtained for such purposes. However, in our hands, spherogenic neural crest–derived cells from the adult mouse skin displayed a rather restricted potential in vivo (unpublished data). In particular, skin-derived spheres of neural crest origin, when dissociated and injected into the lateral ventricles of rat and chicken embryos or transplanted onto hippocampal brain slices, remained largely undifferentiated in cell aggregates close to the injection site and failed to integrate into the host central nervous system (CNS) tissue (unpublished data). This is in contrast to neural progenitors obtained from embryonic stem cells or neural stem cells from the CNS assessed in the same experimental paradigms (Brüstle et al., 1997; Benninger et al., 2003; Wernig et al., 2004). Moreover, we did not observe neural differentiation, tissue integration, or behavioral improvement upon transplantation of skin-derived multipotent neural crest–derived cells into the striatum of a 6-hydroxydopamine–lesioned mouse model for Parkinson's disease (Bjorklund et al., 2002; unpublished data), or upon intravenous injection into mice with experimental autoimmune encephalomyelitis, a model for multiple sclerosis previously used to test the therapeutic potential of adult neural stem cells from the CNS (Pluchino et al., 2003; unpublished data). These trials suggest that multipotent cells of neural crest origin that are present in the adult skin cannot transdifferentiate into neural cell types of the CNS. Likewise, SKPs failed to generate CNS neurons upon transplantation into hippocampal slices (Fernandes et al., 2006). This does not exclude the possibility that adult neural crest–related cells with stem cell properties might be of high value for the generation of Schwann cells, cartilage, or other neural crest–derived tissues potentially useful in clinical applications. Hence, our study underlines the importance of choosing the appropriate stem cell type for a given task.
Materials and methods
Skin sphere cultures
Human thigh skin from an adult man (∼45 yr of age) and face skin from an adult woman (∼57 yr of age; provided by G. Beer, University Hospital of Zurich, Zurich, Switzerland) were obtained in the frame of cosmetic surgery according to the guidelines of the University Hospital of Zurich. Murine skin was taken from adult C57/BL6 mice of at least 8 wk of age. Skin samples (composed of both dermis and epidermis) were dissected, cut into small pieces, and digested in 0.1% Trypsin-EDTA (Invitrogen) in HBSS without Ca2+ and Mg2+ (Animed) and digested for 50 min at 37°C. Partially digested skin pieces were dissociated mechanically and filtered through a 40-μm cell strainer (BD Biosciences). The cell suspension was centrifuged and washed with medium, and the cell pellet was resuspended in growth medium (GM) consisting of dME-F12 1:1 containing 1× B-27 supplement (Invitrogen), 20 ng/ml FGF2 (PeproTech), 10 ng/ml EGF (PeproTech), penicillin/streptomycin (P/S), and Fungizone. GM for human cells also contained 10 ng/ml leukemia inhibitory factor (Sigma-Aldrich). 2.5–4 million cells were seeded in GM into an uncoated T-25 cell culture flask (BD Biosciences). After 4–7 d in culture, sphere formation was observed. For FACS analysis, skin was taken from mice between 10 and 16 d of age. Samples were incubated in 0.5 mg/ml Dispase (Roche) in HBSS for 30 min at 4°C. Fat tissue was removed with forceps, and the rest of the skin was cut into small pieces and digested in 1 mg/ml collagenase (Worthington) in HBSS for 45 min at 37°C. After a final digestion with 0.1% Trypsin-EDTA in HBSS for 5 min at 37°C, the partially digested skin pieces were dissociated mechanically and treated as described above. FACS was performed with a FACS Aria (Becton Dickinson).
Skin sphere passaging
Once a week, the sphere suspension was transferred into a 15-ml Falcon tube. Cells adhering to the flask bottom were discarded. Spheres were centrifuged, and one third of the supernatant was transferred as conditioned medium into a new T-25 flask. Spheres were incubated with 300 μl Trypsin-EDTA solution (0.25%) for 3–5 min at RT. 400 μl of Ovomucoid solution (1 mg/ml Trypsin inhibitor [Sigma-Aldrich] and 10 mg DNase [Roche] in 25 ml medium) were added, and spheres were dissociated mechanically, centrifuged, resuspended in fresh GM, and seeded into a new flask containing one third conditioned medium. After some passages, spheres were cultured in flasks coated with Poly(2-hydroxyethylmethacrylate) (Poly-Hema; Sigma-Aldrich). Coating was performed at RT with a solution of 16 mg/ml Poly-Hema in 95% ethanol.
Microdissection and sphere cultures of whisker follicles
Anagen-phase whisker follicles from mouse and rat were dissected out of the whisker pad and microdissected as described previously (Kobayashi et al., 1993). Follicle structures were incubated in 0.05% Trypsin-EDTA in dME for 1.5–2 h at 37°C. Trypsin activity was stopped with dME containing 10% FCS. After two washing steps with GM, cells of each structure were plated in a well of a 24-well dish. Sphere formation was observed within 1–2 wk. Passaging was performed as described for skin spheres using 0.05% Trypsin-EDTA for 2 min at RT.
Cell differentiation
All differentiation assays were performed using spheres plated on dishes coated with fibronectin (FN) or poly-d-lysine/FN as described previously (Stemple and Anderson, 1992). Neurogenesis, gliogenesis, and smooth muscle formation were observed after 3–7 d in GM. Chondrocyte formation was obtained after 9 d in dME containing 10% FCS, 50 μg/ml ascorbic acid 2-phosphate (Sigma-Aldrich), 10 ng/ml FGF2, and P/S, followed by dME containing 10% FCS, 50 μg/ml ascorbic acid 2-phosphate, and 10 ng/ml BMP2 (PeproTech) for another 3 d. Adipocytes were occasionally observed when spheres were cultured in DME-F12 containing B-27 supplement and 10 ng/ml BMP2. Melanocytes were observed when cultured in MEM containing 10% FCS, 50 ng/ml murine stem cell factor (PeproTech), 100 nM endothelin-3 (Sigma-Aldrich), and P/S for at least 10 d.
Clonal analysis of neural crest–derived stem cells
Murine skin–derived spheres passaged >17 times were dissociated with Trypsin-EDTA as described and plated at clonal density on pDL/FN-coated 35-mm dishes (Corning) in standard medium prepared as reported previously (Stemple and Anderson, 1992). Single p75-positive cells were labeled and mapped as described previously (Hagedorn et al., 1999) and incubated in standard medium alone or supplemented with 100 ng/ml BMP2, 1 nM NRG1 (R&D Systems), or 0.1 ng/ml TGFβ (R&D Systems). After 10 d, the cells were fixed and analyzed immunocytochemically.
Immunofluorescence on cells and tissue sections
Anti-p75, anti-Sox10, anti-SMA, and anti-GFAP antibody stainings on cells were done as described by Kleber et al. (2005). Anti-TuJ1 antibody (1:200; Sigma-Aldrich) and anti-Keratin antibody (1:500; Abcam) were used for 2 h at RT, whereas anti–β-galactosidase (1:100; Roche) and anti-NG2 (1:200; Chemicon) antibodies were used with incubation overnight at 4°C. The following secondary antibodies were used for 1 h at RT: Cy3-conjugated goat anti-mouse (1:200; Jackson ImmunoResearch Laboratories), Cy3-conjugated goat anti-rabbit (1:200; Jackson ImmunoResearch Laboratories), Alexa 488–conjugated goat anti-mouse (1:100; Invitrogen), and Alexa 488–conjugated goat anti-rabbit (1:100; Invitrogen). Cell nuclei were stained with DAPI. Paraffin sections of X-gal–treated skin were stained for Sox10 as described previously (Dutt et al., 2006) using a controlled antigen-retrieval device (FSG 120-T/T; Milestone). Heat unmasking for the p75 staining was done in 10 mM trisodium citrate, pH 6.0, using the same antigen-retrieval device. The antibody (Chemicon) was used at a dilution of 1:5,000. Alexa 594–conjugated goat anti-mouse and Alexa 488–conjugated goat anti-rabbit (1:200; Invitrogen) were used as secondary antibodies. Immunofluorescence of cells was analyzed using a microscope (Axiovert 100; Carl Zeiss MicroImaging, Inc.) and magnifications of 32×. Pictures were made with a camera (AxioCam MRm) and Axiovision 4.3 software (Carl Zeiss MicroImaging, Inc.). Immunofluorescence of sections was analyzed using a microscope (Axioskop 2; Carl Zeiss MicroImaging, Inc.) with 20×, 63×, and 100× magnifications. Pictures were made with a camera (AxioCam HRc) and Axiovision 4.2 software.
Other stainings
For Alcian blue staining, cells fixed with 4% formaldehyde were incubated with a 3% solution of glacial acetic acid in distilled water for 3 min at RT followed by a 1% Alcian blue solution (Chroma Gesellschaft) in 3% acetic acid for 5 min at RT. For oil red O staining, fixed cells were incubated in 60% isopropanol in water for 15 min at RT followed by another 15 min in an oil red O mixture (0.35 g oil red O [Sigma-Aldrich] in 50 ml isopropanol and water in a 3:2 dilution). For DOPA reaction, cells were incubated in a 0.1% solution of 3-(3,4-Dihydroxyphenyl)-l-alanine (L-DOPA) in PBS for 5 h at 37°C.
Resin sections
Skin pieces from 12-d-old mice for anagen or 8-wk-old mice for telogen stage and single whiskers from Wnt1-Cre/R26R, Ht-PA-Cre/R26R, and Sox10lacZ mice were fixed and incubated in X-gal solution (Paratore et al., 2002). Samples were washed three times with PBS, incubated for 24 h at RT in Bouin's fixative, washed twice for 15 min in H2O, and dehydrated in 80%, 95%, and three times 100% ethanol. Samples were transferred into a 1:1 mixture of 100% ethanol and glycomethacrylate (Leica), left for infiltration at 4°C for 1–2 wk, and embedded in glycomethacrylate resin. 4–5-μm sections were cut on a microtome using a glass knife. Counterstainings were performed with Fast red (Merck) for 30 min at 60°C.
Paraffin sections
Skin from 12-d-old mice from Dhh-Cre/R26R and Dct-Cre/R26R mice was stained with X-gal, embedded in paraffin, and sectioned as described previously (Wurdak et al., 2005).
Online supplemental material
Fig. S1 shows localization of neural crest–derived cells expressing p75/Sox10 in the adult skin. Online supplemental material is available at http://www.jcb.org/cgi/content/full/jcb.200606062/DC1.
Supplementary Material
Acknowledgments
We thank Ned Mantei for critical reading of the manuscript; Andrew McMahon, Philippe Soriano, Michael Wegner, and Frank Costantini for providing transgenic animals; and Michael Wegner for the anti-Sox10 antibody. We thank Gertrude Beer for providing human skin samples, Liliane Schnell for excellent technical assistance, and Eva Niederer and the Zentrallabor für Zellsortierung ETH/UNI for support with the FACS. We are grateful to Oliver Brüstle, Gudrun Gossrau, Alberto Perez-Bouza, Patrick Bargsten, Burkhard Becher, Esther Stoeckli, and Boris Ferger for their support with in vivo transplantation experiments.
This work was supported by the Swiss National Science Foundation (grant PP00A-108344), the National Center of Competence in Research on Neural Plasticity and Repair, the Neuroscience Center Zurich, the Bonizzi-Theler Foundation, the Ecole Polytechnique Fédérale de Lausanne, and the Lausanne University Hospital.
C.E. Wong and C. Paratore contributed equally to this paper.
T. Pietri's present address is Institute of Neuroscience, University of Oregon, Eugene, OR 97403.
Abbreviations used in this paper: BMP, bone morphogenic protein; CNS, central nervous system; FN, fibronectin; GFAP, glial fibrillary acidic protein; GM, growth medium; NCSC, neural crest stem cell; NRG, neuregulin; SKP, skin-derived precursor cell; SMA, smooth muscle actin.
References
- Alonso, L., and E. Fuchs. 2003. Stem cells of the skin epithelium. Proc. Natl. Acad. Sci. USA. 100(Suppl. 1):11830–11835. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amoh, Y., L. Li, K. Katsuoka, S. Penman, and R.M. Hoffman. 2005. Multipotent nestin-positive, keratin-negative hair-follicle bulge stem cells can form neurons. Proc. Natl. Acad. Sci. USA. 102:5530–5534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belicchi, M., F. Pisati, R. Lopa, L. Porretti, F. Fortunato, M. Sironi, M. Scalamogna, E.A. Parati, N. Bresolin, and Y. Torrente. 2004. Human skin-derived stem cells migrate throughout forebrain and differentiate into astrocytes after injection into adult mouse brain. J. Neurosci. Res. 77:475–486. [DOI] [PubMed] [Google Scholar]
- Benninger, F., H. Beck, M. Wernig, K.L. Tucker, O. Brüstle, and B. Scheffler. 2003. Functional integration of embryonic stem cell-derived neurons in hippocampal slice cultures. J. Neurosci. 23:7075–7083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bixby, S., G.M. Kruger, J.T. Mosher, N.M. Joseph, and S.J. Morrison. 2002. Cell-intrinsic differences between stem cells from different regions of the peripheral nervous system regulate the generation of neural diversity. Neuron. 35:643–656. [DOI] [PubMed] [Google Scholar]
- Bjorklund, L.M., R. Sanchez-Pernaute, S. Chung, T. Andersson, I.Y. Chen, K.S. McNaught, A.L. Brownell, B.G. Jenkins, C. Wahlestedt, K.S. Kim, and O. Isacson. 2002. Embryonic stem cells develop into functional dopaminergic neurons after transplantation in a Parkinson rat model. Proc. Natl. Acad. Sci. USA. 99:2344–2349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanpain, C., W.E. Lowry, A. Geoghegan, L. Polak, and E. Fuchs. 2004. Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell. 118:635–648. [DOI] [PubMed] [Google Scholar]
- Botchkarev, V.A., S. Eichmuller, O. Johansson, and R. Paus. 1997. Hair cycle-dependent plasticity of skin and hair follicle innervation in normal murine skin. J. Comp. Neurol. 386:379–395. [DOI] [PubMed] [Google Scholar]
- Botchkareva, N.V., V.A. Botchkarev, L.H. Chen, G. Lindner, and R. Paus. 1999. A role for p75 neurotrophin receptor in the control of hair follicle morphogenesis. Dev. Biol. 216:135–153. [DOI] [PubMed] [Google Scholar]
- Brault, V., R. Moore, S. Kutsch, M. Ishibashi, D.H. Rowitch, A.P. McMahon, L. Sommer, O. Boussida, and R. Kemler. 2001. Inactivation of the β-catenin gene by Wnt1-Cre-mediated deletion results in dramatic brain malformation and failure of craniofacial development. Development. 128:1253–1264. [DOI] [PubMed] [Google Scholar]
- Britsch, S., D.E. Goerich, D. Riethmacher, R.I. Peirano, M. Rossner, K.A. Nave, C. Birchmeier, and M. Wegner. 2001. The transcription factor Sox10 is a key regulator of peripheral glial development. Genes Dev. 15:66–78. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brüstle, O., A.C. Spiro, K. Karram, K. Choudhary, S. Okabe, and R.D. McKay. 1997. In vitro-generated neural precursors participate in mammalian brain development. Proc. Natl. Acad. Sci. USA. 94:14809–14814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Claudinot, S., M. Nicolas, H. Oshima, A. Rochat, and Y. Barrandon. 2005. Long-term renewal of hair follicles from clonogenic multipotent stem cells. Proc. Natl. Acad. Sci. USA. 102:14677–14682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dupin, E., C. Glavieux, P. Vaigot, and N.M. Le Douarin. 2000. Endothelin 3 induces the reversion of melanocytes to glia through a neural crest-derived glial-melanocytic progenitor. Proc. Natl. Acad. Sci. USA. 97:7882–7887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dutt, S., M. Kleber, M. Matasci, L. Sommer, and D.R. Zimmermann. 2006. Versican V0 and V1 guide migratory neural crest cells. J. Biol. Chem. 281:12123–12131. [DOI] [PubMed] [Google Scholar]
- Dyce, P.W., H. Zhu, J. Craig, and J. Li. 2004. Stem cells with multilineage potential derived from porcine skin. Biochem. Biophys. Res. Commun. 316:651–658. [DOI] [PubMed] [Google Scholar]
- Fernandes, K.J., I.A. McKenzie, P. Mill, K.M. Smith, M. Akhavan, F. Barnabe-Heider, J. Biernaskie, A. Junek, N.R. Kobayashi, J.G. Toma, et al. 2004. A dermal niche for multipotent adult skin-derived precursor cells. Nat. Cell Biol. 6:1082–1093. [DOI] [PubMed] [Google Scholar]
- Fernandes, K.J., N.R. Kobayashi, C.J. Gallagher, F. Barnabe-Heider, A. Aumont, D.R. Kaplan, and F.D. Miller. 2006. Analysis of the neurogenic potential of multipotent skin-derived precursors. Exp. Neurol. 201:32–48. [DOI] [PubMed] [Google Scholar]
- Gambardella, L., and Y. Barrandon. 2003. The multifaceted adult epidermal stem cell. Curr. Opin. Cell Biol. 15:771–777. [DOI] [PubMed] [Google Scholar]
- Guyonneau, L., A. Rossier, C. Richard, E. Hummler, and F. Beermann. 2002. Expression of Cre recombinase in pigment cells. Pigment Cell Res. 15:305–309. [DOI] [PubMed] [Google Scholar]
- Hagedorn, L., U. Suter, and L. Sommer. 1999. P0 and PMP22 mark a multipotent neural crest-derived cell type that displays community effects in response to TGF-β family factors. Development. 126:3781–3794. [DOI] [PubMed] [Google Scholar]
- Jaegle, M., M. Ghazvini, W. Mandemakers, M. Piirsoo, S. Driegen, F. Levavasseur, S. Raghoenath, F. Grosveld, and D. Meijer. 2003. The POU proteins Brn-2 and Oct-6 share important functions in Schwann cell development. Genes Dev. 17:1380–1391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiang, X., D.H. Rowitch, P. Soriano, A.P. McMahon, and H.M. Sucov. 2000. Fate of the mammalian cardiac neural crest. Development. 127:1607–1616. [DOI] [PubMed] [Google Scholar]
- Joannides, A., P. Gaughwin, C. Schwiening, H. Majed, J. Sterling, A. Compston, and S. Chandran. 2004. Efficient generation of neural precursors from adult human skin: astrocytes promote neurogenesis from skin-derived stem cells. Lancet. 364:172–178. [DOI] [PubMed] [Google Scholar]
- Kawaguchi, A., T. Miyata, K. Sawamoto, N. Takashita, A. Murayama, W. Akamatsu, M. Ogawa, M. Okabe, Y. Tano, S.A. Goldman, and H. Okano. 2001. Nestin-EGFP transgenic mice: visualization of the self-renewal and multipotency of CNS stem cells. Mol. Cell. Neurosci. 17:259–273. [DOI] [PubMed] [Google Scholar]
- Kleber, M., and L. Sommer. 2004. Wnt signaling and the regulation of stem cell function. Curr. Opin. Cell Biol. 16:681–687. [DOI] [PubMed] [Google Scholar]
- Kleber, M., H.Y. Lee, H. Wurdak, J. Buchstaller, M.M. Riccomagno, L.M. Ittner, U. Suter, D.J. Epstein, and L. Sommer. 2005. Neural crest stem cell maintenance by combinatorial Wnt and BMP signaling. J. Cell Biol. 169:309–320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi, K., A. Rochat, and Y. Barrandon. 1993. Segregation of keratinocyte colony-forming cells in the bulge of the rat vibrissa. Proc. Natl. Acad. Sci. USA. 90:7391–7395. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruger, G.M., J.T. Mosher, S. Bixby, N. Joseph, T. Iwashita, and S.J. Morrison. 2002. Neural crest stem cells persist in the adult gut but undergo changes in self-renewal, neuronal subtype potential, and factor responsiveness. Neuron. 35:657–669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le Douarin, N.M., and E. Dupin. 2003. Multipotentiality of the neural crest. Curr. Opin. Genet. Dev. 13:529–536. [DOI] [PubMed] [Google Scholar]
- Lee, H.Y., M. Kleber, L. Hari, V. Brault, U. Suter, M.M. Taketo, R. Kemler, and L. Sommer. 2004. Instructive role of Wnt/beta-catenin in sensory fate specification in neural crest stem cells. Science. 303:1020–1023. [DOI] [PubMed] [Google Scholar]
- Li, L., J. Mignone, M. Yang, M. Matic, S. Penman, G. Enikolopov, and R.M. Hoffman. 2003. Nestin expression in hair follicle sheath progenitor cells. Proc. Natl. Acad. Sci. USA. 100:9958–9961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lutolf, S., F. Radtke, M. Aguet, U. Suter, and V. Taylor. 2002. Notch1 is required for neuronal and glial differentiation in the cerebellum. Development. 129:373–385. [DOI] [PubMed] [Google Scholar]
- Nishimura, E.K., S.A. Jordan, H. Oshima, H. Yoshida, M. Osawa, M. Moriyama, I.J. Jackson, Y. Barrandon, Y. Miyachi, and S. Nishikawa. 2002. Dominant role of the niche in melanocyte stem-cell fate determination. Nature. 416:854–860. [DOI] [PubMed] [Google Scholar]
- Osawa, M., G. Egawa, S.S. Mak, M. Moriyama, R. Freter, S. Yonetani, F. Beermann, and S. Nishikawa. 2005. Molecular characterization of melanocyte stem cells in their niche. Development. 132:5589–5599. [DOI] [PubMed] [Google Scholar]
- Oshima, H., A. Rochat, C. Kedzia, K. Kobayashi, and Y. Barrandon. 2001. Morphogenesis and renewal of hair follicles from adult multipotent stem cells. Cell. 104:233–245. [DOI] [PubMed] [Google Scholar]
- Paratore, C., D.E. Goerich, U. Suter, M. Wegner, and L. Sommer. 2001. Survival and glial fate acquisition of neural crest cells are regulated by an interplay between the transcription factor Sox10 and extrinsic combinatorial signaling. Development. 128:3949–3961. [DOI] [PubMed] [Google Scholar]
- Paratore, C., C. Eichenberger, U. Suter, and L. Sommer. 2002. Sox10 haploinsufficiency affects maintenance of progenitor cells in a mouse model of Hirschsprung disease. Hum. Mol. Genet. 11:3075–3085. [DOI] [PubMed] [Google Scholar]
- Pardal, R., M.F. Clarke, and S.J. Morrison. 2003. Applying the principles of stem-cell biology to cancer. Nat. Rev. Cancer. 3:895–902. [DOI] [PubMed] [Google Scholar]
- Peters, E.M., D.J. Tobin, N. Botchkareva, M. Maurer, and R. Paus. 2002. Migration of melanoblasts into the developing murine hair follicle is accompanied by transient c-Kit expression. J. Histochem. Cytochem. 50:751–766. [DOI] [PubMed] [Google Scholar]
- Peters, E.M., M. Maurer, V.A. Botchkarev, K. Jensen, P. Welker, G.A. Scott, and R. Paus. 2003. Kit is expressed by epithelial cells in vivo. J. Invest. Dermatol. 121:976–984. [DOI] [PubMed] [Google Scholar]
- Pietri, T., O. Eder, M. Blanche, J.P. Thiery, and S. Dufour. 2003. The human tissue plasminogen activator-Cre mouse: a new tool for targeting specifically neural crest cells and their derivatives in vivo. Dev. Biol. 259:176–187. [DOI] [PubMed] [Google Scholar]
- Pluchino, S., A. Quattrini, E. Brambilla, A. Gritti, G. Salani, G. Dina, R. Galli, U. Del Carro, S. Amadio, A. Bergami, et al. 2003. Injection of adult neurospheres induces recovery in a chronic model of multiple sclerosis. Nature. 422:688–694. [DOI] [PubMed] [Google Scholar]
- Rendl, M., L. Lewis, and E. Fuchs. 2005. Molecular dissection of mesenchymal-epithelial interactions in the hair follicle. PLoS Biol. 3:e331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds, B.A., and R.L. Rietze. 2005. Neural stem cells and neurospheres— re-evaluating the relationship. Nat. Methods. 2:333–336. [DOI] [PubMed] [Google Scholar]
- Santagati, F., and F.M. Rijli. 2003. Cranial neural crest and the building of the vertebrate head. Nat. Rev. Neurosci. 4:806–818. [DOI] [PubMed] [Google Scholar]
- Shih, D.T., D.C. Lee, S.C. Chen, R.Y. Tsai, C.T. Huang, C.C. Tsai, E.Y. Shen, and W.T. Chiu. 2005. Isolation and characterization of neurogenic mesenchymal stem cells in human scalp tissue. Stem Cells. 23:1012–1020. [DOI] [PubMed] [Google Scholar]
- Sieber-Blum, M., and M. Grim. 2004. The adult hair follicle: cradle for pluripotent neural crest stem cells. Birth Defects Res. C. Embryo Today. 72:162–172. [DOI] [PubMed] [Google Scholar]
- Sieber-Blum, M., M. Grim, Y.F. Hu, and V. Szeder. 2004. Pluripotent neural crest stem cells in the adult hair follicle. Dev. Dyn. 231:258–269. [DOI] [PubMed] [Google Scholar]
- Slack, J. 2001. Skinny dipping for stem cells. Nat. Cell Biol. 3:E205–E206. [DOI] [PubMed] [Google Scholar]
- Sommer, L. 2001. Context-dependent regulation of fate decisions in multipotent progenitor cells of the peripheral nervous system. Cell Tissue Res. 305:211–216. [DOI] [PubMed] [Google Scholar]
- Sommer, L. 2005. Checkpoints of melanocyte stem cell development. Sci. STKE. 2005:pe42. [DOI] [PubMed]
- Srinivas, S., T. Watanabe, C.S. Lin, C.M. William, Y. Tanabe, T.M. Jessell, and F. Costantini. 2001. Cre reporter strains produced by targeted insertion of EYFP and ECFP into the ROSA26 locus. BMC Dev. Biol. 1:4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stemple, D.L., and D.J. Anderson. 1992. Isolation of a stem cell for neurons and glia from the mammalian neural crest. Cell. 71:973–985. [DOI] [PubMed] [Google Scholar]
- Taylor, G., M.S. Lehrer, P.J. Jensen, T.T. Sun, and R.M. Lavker. 2000. Involvement of follicular stem cells in forming not only the follicle but also the epidermis. Cell. 102:451–461. [DOI] [PubMed] [Google Scholar]
- Toma, J.G., M. Akhavan, K.J. Fernandes, F. Barnabe-Heider, A. Sadikot, D.R. Kaplan, and F.D. Miller. 2001. Isolation of multipotent adult stem cells from the dermis of mammalian skin. Nat. Cell Biol. 3:778–784. [DOI] [PubMed] [Google Scholar]
- Toma, J.G., I.A. McKenzie, D. Bagli, and F.D. Miller. 2005. Isolation and characterization of multipotent skin-derived precursors from human skin. Stem Cells. 23:727–737. [DOI] [PubMed] [Google Scholar]
- Trentin, A., C. Glavieux-Pardanaud, N.M. Le Douarin, and E. Dupin. 2004. Self-renewal capacity is a widespread property of various types of neural crest precursor cells. Proc. Natl. Acad. Sci. USA. 101:4495–4500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wagers, A.J., and I.L. Weissman. 2004. Plasticity of adult stem cells. Cell. 116:639–648. [DOI] [PubMed] [Google Scholar]
- Wang, Y., Y. Zhang, Y. Zeng, Y. Zheng, G. Fu, Z. Cui, and T. Yang. 2006. Patterns of nestin expression in human skin. Cell Biol. Int. 30:144–148. [DOI] [PubMed] [Google Scholar]
- Wernig, M., F. Benninger, T. Schmandt, M. Rade, K.L. Tucker, H. Bussow, H. Beck, and O. Brüstle. 2004. Functional integration of embryonic stem cell-derived neurons in vivo. J. Neurosci. 24:5258–5268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wurdak, H., L.M. Ittner, K.S. Lang, P. Leveen, U. Suter, J.A. Fischer, S. Karlsson, W. Born, and L. Sommer. 2005. Inactivation of TGFbeta signaling in neural crest stem cells leads to multiple defects reminiscent of DiGeorge syndrome. Genes Dev. 19:530–535. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}