

Abstract
Importin β is a major mediator of import into the cell nucleus. Importin β binds cargo molecules either directly or via two types of adapter molecules, importin α, for import of proteins with a classical nuclear localization signal (NLS), or snurportin 1, for import of m3G-capped U snRNPs. Both adapters have an NH2-terminal importin β–binding domain for binding to, and import by, importin β, and both need to be returned to the cytoplasm after having delivered their cargoes to the nucleus. We have shown previously that CAS mediates export of importin α. Here we show that snurportin 1 is exported by CRM1, the receptor for leucine-rich nuclear export signals (NESs). However, the interaction of CRM1 with snurportin 1 differs from that with previously characterized NESs. First, CRM1 binds snurportin 1 50-fold stronger than the Rev protein and 5,000-fold stronger than the minimum Rev activation domain. Second, snurportin 1 interacts with CRM1 not through a short peptide but rather via a large domain that allows regulation of affinity. Strikingly, snurportin 1 has a low affinity for CRM1 when bound to its m3G-capped import substrate, and a high affinity when substrate-free. This mechanism appears crucial for productive import cycles as it can ensure that CRM1 only exports snurportin 1 that has already released its import substrate in the nucleus.
Keywords: nuclear transport, nuclear pore complex, importin, exportin, snurportin 1
Eukaryotic cells have to exchange macromolecules between their nuclear and cytoplasmic compartments. This nucleocytoplasmic transport proceeds through nuclear pore complexes (NPCs)1 and is largely mediated by specific transport receptors that shuttle between the nucleus and the cytoplasm in order to accomplish multiple rounds of transport (for recent reviews see Dahlberg and Lund, 1998; Izaurralde and Adam, 1998; Mattaj and Englmeier, 1998). Directionality in transport has been suggested to be conferred by the GTPase Ran and the asymmetric nucleocytoplasmic distribution of its regulators (Görlich et al., 1996b, 1996c). Ran switches between a GDP and a GTP-bound form by GTP hydrolysis and nucleotide exchange. Its nucleotide exchange factor RCC1 is nuclear, while the RanGTPase-activating protein (RanGAP1) is cytoplasmic. This differential compartmentalization of RCC1 and RanGAP is thought to result in a high concentration of RanGTP in the nucleus and a low level in the cytoplasm. Transport receptors are RanGTP-binding proteins that respond to this gradient by loading and unloading their cargoes in the appropriate compartment. Importins bind cargo molecules initially in the cytoplasm, release them upon binding to RanGTP in the nucleus, and return to the cytoplasm as RanGTP complexes without their cargo (Rexach and Blobel, 1995; Chi et al., 1996; Görlich et al., 1996c; Izaurralde et al., 1997; Schlenstedt et al., 1997; Siomi et al., 1997; Jäkel and Görlich, 1998). RanGTP then needs to be removed from the importins to allow binding of another import substrate. This is accomplished by the combined action of RanBP1 and RanGAP1 and results in the hydrolysis of the Ran-bound GTP (Bischoff and Görlich, 1997; Floer et al., 1997; Lounsbury and Macara, 1997). Exportins, on the other hand, bind their substrates in the nucleus forming a trimeric complex with RanGTP (Fornerod et al., 1997a; Kutay et al., 1997, 1998; Arts et al., 1998). The trimeric complex is then transferred to the cytoplasm where GTP hydrolysis results in Ran's irreversible dissociation from the complex, allowing the exportin to release its substrate, reenter the nucleus, and bind and export the next cargo molecule. Because these transport cycles should constantly deplete Ran from the nucleus, Ran needs to be efficiently recycled back to the nucleus. This is accomplished through the action of NTF2 (Ribbeck et al., 1998).
Transport receptors are at least distantly related to importin β and form a superfamily whose members are of a similar size (90–130 kD) and share an NH2-terminal RanGTP-binding domain. (Fornerod et al., 1997b; Görlich et al., 1997). In simple cases, they bind their substrates directly. Examples include nuclear import of hnRNP proteins mediated by transportin (Pollard et al., 1996; Fridell et al., 1997) and import of ribosomal proteins, which is mediated by at least four different transport receptors in higher eukaryotes (Jäkel and Görlich, 1998) and two in yeast (Rout et al., 1997; Schlenstedt et al., 1997). Paradigms for transport of proteins and RNAs out of the nucleus include nuclear export of importin α by CAS (Kutay et al., 1997), export of leucine-rich nuclear export signal (NES) containing proteins mediated by CRM1 (Fornerod et al., 1997a; Stade et al., 1997), and the recently characterized tRNA export pathway (Arts et al., 1998; Kutay et al., 1998; see also Hellmuth et al., 1998; Sarkar and Hopper, 1998).
Importin β (Chi et al., 1995; Görlich et al., 1995a; Imamoto et al., 1995; Radu et al., 1995) is special among the import receptors in that it can extend its substrate specificity by the use of adapter molecules such as importin α. The importin α/β complex mediates import of proteins carrying a classical basic nuclear localization signal (NLS; for reviews see Görlich, 1997; Nigg, 1997). Importin α recognizes the substrate in the cytoplasm, while importin β mediates the translocation of the importin α/NLS complex through the NPC. Importin α is characterized by an NH2-terminal importin β–binding (IBB) domain that confers binding to and import by importin β (Görlich et al., 1996a; Weis et al., 1996).
After nuclear entry, the importin α/β complex is dissociated by direct binding of RanGTP to importin β. The importin β/RanGTP complex is directly returned to the cytoplasm. As indicated above, importin α requires a specialized exportin, CAS, for its reexport. CAS has a clear preference for NLS-free importin α, explaining why NLS proteins stay in the nucleus.
The biogenesis of the spliceosomal snRNPs U1, U2, U4, and U5 in higher eukaryotes is a complex process that includes cytoplasmic and nuclear maturation steps. These U snRNAs are initially synthesized in the nucleus as monomethyl (m7G)-capped RNA polymerase II transcripts and are exported to the cytoplasm with the aid of the nuclear cap-binding complex (CBC) and the exportin CRM1 (Hamm and Mattaj, 1990; Izaurralde et al., 1994, 1995; Fornerod et al., 1997a). The U snRNAs then bind Sm core proteins in the cytoplasm to form a ribonucleoprotein complex denoted Sm core, which is followed by the hypermethylation of the m7G-cap to the mature 2,2,7-trimethylguanosine G cap (m3G-cap) (Mattaj, 1986; Plessel et al., 1994). The m3G-cap and the Sm core constitute two distinct import signals that redirect the mature snRNP particles to the nucleus (Fischer and Lührmann, 1990; Hamm et al., 1990; Fischer et al., 1991, 1993). Both signals apparently access the importin β–dependent import pathway through adapters that are distinct from importin α (Palacios et al., 1997). The Sm core–specific adapter is not yet known. However, the m3G-cap receptor, snurportin 1, has been identified recently and shown to mediate nuclear import of m3G-capped uridine-rich small nuclear RNPs (U snRNPs; Huber et al., 1998). Snurportin 1 and importin α share the NH2-terminal IBB domain for importin β binding, but are otherwise unrelated in sequence.
Just like importin α, snurportin 1 also needs to be returned back to the cytoplasm after each round of import. Unlike importin α, which uses CAS for recycling, we show here that reexport of snurportin 1 is mediated by CRM1. Snurportin 1 is a potent competitor of other CRM1-dependent pathways and interacts with CRM1 orders of magnitude more avidly than previously described export signals. Binding of CRM1 and of a m3G-cap to snurportin 1 appear to be mutually exclusive, suggesting that CRM1 only reexports substrate-free snurportin 1 and thereby ensures productive import cycles.
Materials and Methods
Recombinant Protein Expression and Purification
The following proteins were expressed in Escherichia coli BLR/Rep4 and purified as previously described: Xenopus importin α (Görlich et al., 1994), human importin α (Görlich et al., 1996a), NTF2 (Ribbeck et al., 1998), His-tagged RanQ69L and Ran wild type (Görlich et al., 1996c), Rna1p (Bischoff et al., 1995a), RanBP1 (Bischoff et al., 1995b), and CAS (Kutay et al., 1997). Snurportin 1 was cloned into the BamHI-XmaI sites of pQE30 (Qiagen), expressed with an NH2-terminal His tag and purified on nickel-NTA agarose followed by dialysis in 20 mM Hepes-KOH, pH 7.5, 200 mM NaCl, 2 mM magnesium acetate, 250 mM sucrose. GST-snurportin 1 was expressed from pGEX T4 and purified on glutathione Sepharose 4B, followed by chromatography on MonoQ. zz-snurportin 1 (full length and fragments) were cloned into the NcoI-BamHI sites of pQE60 and expressed with a COOH-terminal His tag and with NH2-terminal IgG binding domains from Staphylococcus aureus protein A. Human CRM1 was cloned into the NdeI-BamHI sites of pET3a and was expressed in BL21 DE3 without addition of IPTG at 30°C. This recombinant CRM1 is untagged and was purified on nickel-NTA agarose followed by chromatography on MonoQ. The cDNA coding for the HIV Rev protein was cloned into the NcoI-BamHI site of the 6z60 vector (Jäkel and Görlich, 1998). Expression was with an NH2-terminal 6z tag and a COOH-terminal his tag, purification was on nickel agarose.
Labeling of Proteins
Labeling of snurportin 1 with fluorescein 5′ maleimide (Calbiochem) was performed at a 1:1 molar ratio in 50 mM Hepes-KOH, pH 7.5, 200 mM NaCl, 2 mM magnesium acetate, 250 mM sucrose for 2 h on ice. Free label was removed on a NAP5 column equilibrated with 50 mM Hepes-KOH, 150 mM NaCl, 2 mM magnesium acetate. Labeling of Xenopus importin α has been described before (Görlich et al., 1996a).
Antibodies
Antibodies against the following antigens have been described before: human importin β (Görlich et al., 1995b), Xenopus RanBP7 (Görlich et al., 1997), and human CAS (Jäkel and Görlich, 1998). Anti-CRM1 antibodies were raised in rabbits against a CRM1 peptide (cys-EKHKRQMSV). Antibodies were affinity-purified on sulfoLink (Pierce) to which the antigens had been coupled.
Import Assays
Permeabilization of HeLa cells and nuclear import reactions in suspension was performed essentially as described before (Adams et al., 1990; Görlich et al., 1996c; Kutay et al., 1997). The import buffer contained: 2 mg/ml nucleoplasmin core (to block nonspecific binding), 20 mM Hepes-KOH, pH 7.5, 140 mM potassium acetate, 5 mM magnesium acetate, 250 mM sucrose, 0.5 mM EGTA. Reactions were supplemented with an energy-regenerating system (0.5 mM ATP, 0.5 mM GTP, 10 mM creatine phosphate, 50 μg/ml creatine kinase), 5 μM RanGDP, and 1 μM NTF2.
Binding Assays
zz-tagged RanQ69L and snurportin 1 prebound to IgG-Sepharose were used as affinity matrices. The z domain is the IgG binding domain from S. aureus protein A. 500 μl of cytoplasmic HeLa extract or 200 μl of E. coli lysate expressing the recombinant proteins was incubated with 20 μl of affinity matrix for 4 h at 4°C in binding buffer (50 mM Hepes-KOH, pH 7.5, 200 mM NaCl, 5 mM magnesium acetate, 0.005% digitonin). The beads were recovered by mild centrifugation and washed three times with 1 ml binding buffer. Elution was with 100 μl of 50 mM Tris-HCl, pH 7.5, 1 M magnesium chloride for 10 min at room temperature. Proteins were precipitated with 90% isopropanol (final concentration), dissolved in SDS sample buffer, and analyzed by SDS-PAGE.
Kinetic measurement of the RanGTPase was as described before (Bischoff et al., 1995a, 1995b; Kutay et al., 1997) with modifications described in the figure legends. The m3G-cap (m3GpppAmpUmpA-oligonucleotide) was described previously (Huber et al., 1998) and the m7GpppG-cap dinucleotide was purchased from Pharmacia.
Oocyte Injection
Oocyte injection and analysis of microinjected RNA by denaturing gel electrophoresis and autoradiography were performed as described (Jarmolowski et al., 1994). The mutant RNAs used (U1ΔSm, U5ΔSm) lack protein-binding sites required for the nuclear import of these RNAs (Hamm and Mattaj, 1990; Jarmolowski et al., 1994) and thus remain in the cytoplasm after export from the nucleus.
Results
Identification of Potential Snurportin 1 Export Receptors
Snurportin 1 mediates the m3G-cap–dependent import of U snRNAs (Huber et al., 1998), most likely in conjunction with importin β (Palacios et al., 1997; J. Huber and R. Lührmann, unpublished observations). To achieve multiple rounds of import, snurportin 1 has to return to the cytoplasm after delivering its import substrate to the nucleus. Since importin α and snurportin 1 both contain an IBB domain for binding to and import by importin β, it was of interest to know if snurportin 1 would also exit the nucleus in the same way as importin α, by using CAS for reexport (Kutay et al., 1997). Surprisingly however, we could not detect any interaction between CAS and snurportin 1 (see below), suggesting that snurportin 1 export is mediated by a distinct factor. To identify this factor, we immobilized snurportin 1 and tested which proteins from a HeLa extract it would bind (Fig. 1). Without further addition, three major bands were recovered in the bound fraction and identified by Western blotting as importin β, CRM1, and importin 7 (formerly called RanBP7). The binding of importin β to snurportin 1 was expected from previous studies. Importin 7 was apparently recovered only because it binds importin β (not shown, but see Görlich et al., 1997). Binding of the export receptor CRM1 to snurportin 1 might suggest that CRM1 mediates export of snurportin 1. Indeed, the pattern of snurportin 1–bound proteins changed when RanQ69L GTP was included in the binding reaction. This Ran mutant is GTPase-deficient, remains GTP-bound even in the presence of RanGAP1, and can therefore be used to mimic a nuclear environment in a cytoplasmic extract. Under these conditions, the binding of the two importins was greatly reduced. In contrast, binding of CRM1 was enhanced by the presence of RanGTP. Thus, CRM1 behaved as expected for a snurportin-specific export receptor.
Figure 1.
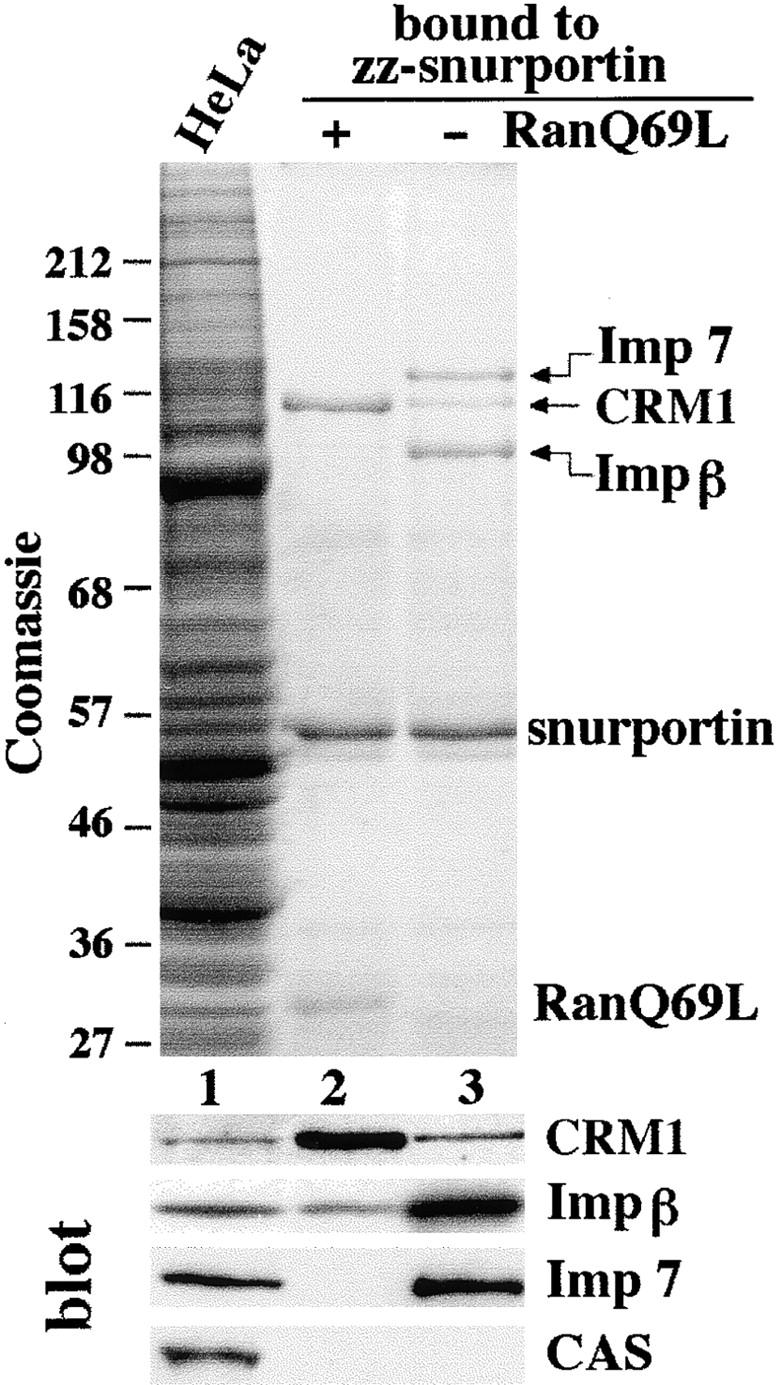
Identification of potential snurportin 1 export factors. A HeLa cell extract was subjected to binding to immobilized snurportin 1 in the presence or absence of 1.3 μM RanQ69L (GTP-bound form). Starting material and bound fractions were analyzed by SDS-PAGE followed by Coomassie staining and Western blotting. The load in the bound fractions corresponds to 20 times the starting material. Note that CRM1 preferentially bound to snurportin 1 in the presence of RanQ69L, but importin β and importin 7 in its absence.
The interaction between snurportin 1 and CRM1 could be either direct or mediated by an unidentified factor in the HeLa extract. To exclude the second possibility we repeated the binding experiments using E. coli lysates containing recombinant transport receptors. CRM1 bound to immobilized snurportin 1 (Fig. 2 A, lane 3), and this binding was enhanced by the addition of RanQ69L. Ran was also recovered in the bound fraction, indicating the formation of a trimeric snurportin/CRM1/RanGTP complex (Fig. 2 A, lane 2). Conversely, the trimeric complex could also be assembled using immobilized RanQ69L, free CRM1, and GST-snurportin 1 (Fig. 2 B). The high cooperativity of complex formation was evident from the observation that CRM1 binding to RanGTP was not detectable in the absence of snurportin 1 but was strong in its presence.
Figure 2.
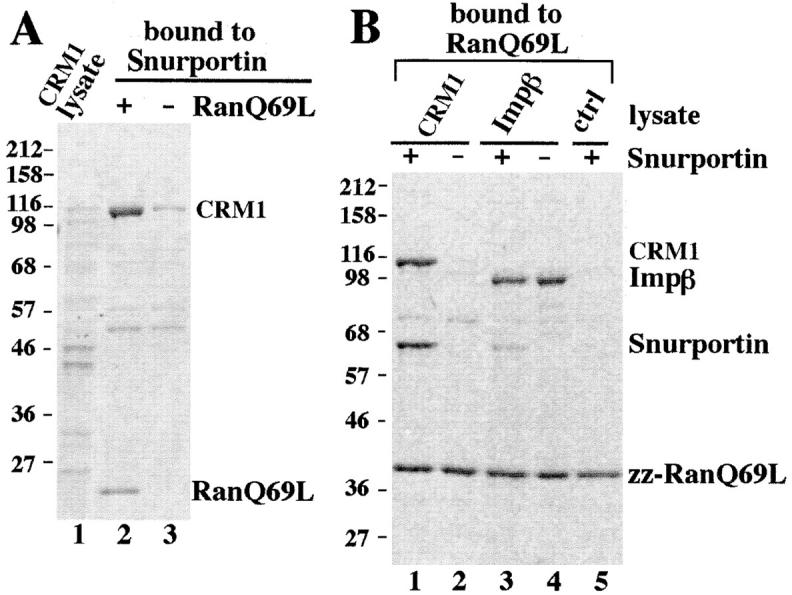
Snurportin 1 interacts directly with CRM1 and importin β in a RanGTP-regulated manner. (A) Immobilized snurportin 1 was used to bind recombinant CRM1 from a total E. coli lysate. Where indicated, 1.3 μM RanQ69L (GTP) was also added. Starting material and bound fractions were analyzed by SDS-PAGE followed by Coomassie staining. The load in the bound fractions corresponded to 20 times the starting material. (B) Immobilized RanQ69L was incubated with total E. coli lysates expressing either recombinant CRM1 (lanes 1 and 2), importin β (lanes 3 and 4), or a control lysate (lane 5). Where indicated, (+) 300 μl of total E. coli lysate expressing a GST-snurportin 1 fusion protein was also included (lanes 1, 3, and 5). The interaction between snurportin 1 and CRM1 in the presence of RanQ69L is direct and a trimeric snurportin/CRM1/RanQ69L complex is formed.
To obtain more quantitative data on the formation of the trimeric complex, we used a kinetic assay (Fig. 3, and see for example Kutay et al., 1997). Binding of an exportin to RanGTP prevents GTPase activation by RanGAP1. GTP hydrolysis can easily be quantified and used to calculate the proportion of Ran that is exportin-bound. From the dose dependence of the effects one can estimate apparent dissociation constants (K d) for the complex formation. Without further addition, CRM1 bound RanGTP only very weakly (apparent K d ≫ 1 μM, Fig. 3 A); In the presence of a saturating concentration of snurportin 1 (2 μM), however, the K d shifted to 4–5 nM, emphasizing the highly cooperative nature of the snurportin/CRM1/RanGTP complex formation. The specificity of the effect was verified by two controls. First, the presence of importin α had no effect on the RanGTP/CRM1 interaction. Second, CAS responded only to its specific export substrate importin α, but not to snurportin 1 (Fig. 3 B).
Figure 3.

Kinetic characterization of the snurportin/CRM1/ RanGTP interaction. (A) The kinetic assay to measure complex formation is based on the observation that binding of RanGTP to an importin β–like factor prevents GTPase activation by RanGAP. 50 pM Ran-[γ-32P]GTP was preincubated at 15°C with the indicated concentrations of CRM1 in the absence or presence of 2 μM snurportin 1 or 2 μM importin α (human Rch1p). After 30 min, a 30-s GTPase reaction was started by addition of 40 nM Rna1p, the S. pombe RanGAP. Hydrolysis of Ran-bound GTP was determined as released [32P]phosphate. Note that Ran and snurportin 1 bound to CRM1 in a highly cooperative manner, with snurportin 1 increasing the apparent affinity of CRM1 for RanGTP roughly 1,000-fold. The apparent constant for dissociation of RanGTP from the trimeric complex is 4–5 nM. The presence of 15 nM RanBP1 relieved the GAP resistance of the complex completely. (B) Measurements were performed exactly as in A except that CAS was added instead of CRM1. Note that snurportin 1 bound selectively to CRM1, but not to CAS, while importin α bound to CAS but not to CRM1.
Disassembly of the Snurportin/CRM1/RanGTP Complex
Export complexes form in the nucleus and need to be disassembled after export in the cytoplasm. In the case of CAS and exportin-t, this disassembly is brought about by the concerted action of RanBP1 and RanGAP1. However, there is a complication in respect to CRM1, namely that RanBP1 and RanGAP1 apparently also contain leucine-rich NESs (Richards et al., 1996; Matunis et al., 1998) and are thus also potential export substrates for CRM1 which could stabilize a CRM1/RanGTP complex. Therefore, it was quite surprising that RanBP1 rendered also the snurportin/CRM1/RanGTP complex sensitive to RanGAP and thereby caused disassembly (see corresponding curves in Fig. 3 A).
CRM1 Is a Functional Nuclear Export Receptor for Snurportin
The observation that CRM1 binds snurportin 1 with high affinity and specificity strongly suggests that CRM1 mediates nuclear export of snurportin. To test this directly, we performed the pulse-chase export experiment shown in Fig. 4. Nuclei of permeabilized HeLa cells were first loaded for 10 min with fluorescein-labeled snurportin 1 and Texas red–labeled importin α in the presence of importin β, Ran, NTF2, and an energy-regenerating system. The sample was then split into four. The first aliquot was fixed immediately and confocal fluorescence microscopy confirmed that snurportin 1 and importin α had both efficiently accumulated in the nuclei. The three remaining aliquots were supplemented with either CRM1, CAS, or buffer and incubated for another 10 min before these samples were fixed also. The addition of CRM1 caused a significant depletion of the nuclear pool of snurportin and an increased NPC signal that probably represented import and export intermediates. However, CRM1 did not change the localization of importin α. Addition of CAS had exactly the opposite effect. It led to nuclear export of importin α but had no effect on snurportin. Therefore, we can conclude that CRM1 specifically mediates export of snurportin. The experiment also confirms our previous data that CAS is the importin α–specific exportin (Kutay et al., 1997) and contradicts the suggestion by Boche and Fanning (1997) that importin α would exit the nucleus by virtue of a Rev-like NES, i.e., in a CRM1-dependent manner.
Figure 4.

CRM1 promotes the export of snurportin 1 from the nucleus. Nuclear import of 1.6 μM fluorescein-labeled snurportin 1 and 1.6 μM Texas red–labeled importin α was allowed for 10 min in the presence of importin β (0.7 μM) and nucleoplasmin (1.4 μM) as described in Materials and Methods. After 10 min, one aliquot of the sample was fixed. The remaining sample was split in three. Either 3 μM CRM1, 3 μM CAS, or buffer was added and the incubation was continued for another 10 min before fixation. Nuclei were spun onto coverslips and fluorescent proteins were detected by confocal microscopy (63× oil objective). Note that addition of CRM1 specifically leads to a depletion of the intranuclear snurportin 1 and the appearance of an NPC staining, but had no effect on importin α distribution. In contrast, CAS promoted export of importin α, but not of snurportin.
Binding of m3G-cap and of CRM1 to Snurportin 1 Are Mutually Exclusive
Productive import cycles require snurportin 1 to enter the nucleus with cargo and to exit without cargo. Although nuclear retention of the import substrate might contribute to this directionality, transport would be most efficient if the affinity of snurportin 1 for m3G-capped U snRNPs was high during nuclear entry and low during reexport. β-Family transport receptors use RanGTP to regulate interactions with their transport substrates. This mechanism is not available to snurportin 1 because it does not bind Ran (Fig. 2 B, lane 5). However, it uses importin β for nuclear entry and CRM1 for exit from the nucleus and the binding of the two transport receptors might regulate snurportin's affinity for the m3G-cap. Therefore, we tested for a cross-talk between the binding sites for the m3G-cap and CRM1. In the absence of RanGTP, CRM1 bound weakly to snurportin 1 (Fig. 5 A). A 10-fold excess of a m3G-cap oligo prevented CRM1 binding completely, suggesting that m3G-cap and CRM1 binding to snurportin 1 are mutually exclusive. However, the m3G-cap oligo could only reduce, but not prevent, CRM1 binding in the presence of Ran-GTP, i.e., when the exportin was in its high affinity form.
Figure 5.

Binding of m3G-cap and of CRM1 to snurportin 1 are mutually exclusive. (A) Immobilized snurportin 1 was used to bind recombinant CRM1 out of total E. coli lysate either in the presence (lanes 3 and 5) or absence (lanes 2 and 4) of a 10-fold molar excess of a m3G-cap oligonucleotide (5 μM). Where indicated, 11 μM RanQ69L was also added (lanes 4 and 5). In the absence of Ran-GTP, m3G-cap prevented binding of CRM1 to snurportin, while in the presence of Ran-GTP binding of CRM1 to snurportin 1 is reduced but not abolished by m3G-cap. (B) Formation of the trimeric snurportin/CRM1/ RanGTP complex was measured as in Fig. 2 A, with the modification that the concentration of CRM1 was kept constant at 300 nM and the concentration of snurportin 1 was varied. Where indicated, snurportin 1 had been preincubated with either 5 μM m3G-capped oligonucleotide or m7GpppG dinucleotide. Note that the m3G-cap RNA oligonucleotide specifically inhibited trimeric complex formation, while the m7G-cap analogue had no effect. (C) Measurements were performed exactly as in B except that a Rev-NES-BSA conjugate was used instead of snurportin. Note that the m3G-cap RNA oligonucleotide had no effect on the Rev-NES/CRM1/RanGTP interaction. (D) Measurements were performed exactly as in B, varying the concentrations of the following export substrates: snurportin 1; HIV Rev protein; Rev-NES peptide 1, which is a synthetic peptide (cys-LPPLERLTL) corresponding to the minimum Rev activation domain, Rev-NES peptide 2, which is a slightly larger peptide (cys-PVPLQLPPLERLTLD) that also includes NH2- and COOH-terminally flanking residues, Mut. NES peptide (cys-LPPDLRLTL), which corresponds to a loss-of-function mutant of the activation domain, was used as a negative control.
The antagonism of CRM1 and m3G-cap binding was also clearly evident in the kinetic assay that measures snurportin/CRM1/RanGTP complex formation (Fig. 5 B). The presence of 5 μM m3G-cap shifted the equilibrium and reduced the apparent affinity of snurportin 1 for the CRM1/RanGTP complex ∼40-fold. The effect was highly specific as verified by two controls. First, a m7G-cap analogue, which does not bind snurportin 1, had no effect on the snurportin/CRM1/RanGTP interaction. Second, the m3G-cap did not interfere with the interaction between CRM1 and BSA conjugated to HIV Rev NES peptides and is thus snurportin 1–specific (Fig. 5 C).
CRM1 Binds Snurportin 1 Far Stronger Than an NES from HIV Rev
A mutual displacement of CRM1 and m3G-cap from snurportin 1 should occur twice per import cycle. When the trimeric export complex has been exported and RanGTP removed, a still quite stable CRM1/snurportin 1 dimer ends up in the cytoplasm. Binding of the import substrate to snurportin 1 displaces the low affinity form of the exportin (Fig. 5 A). Conversely, following nuclear entry, the U snRNPs need to be displaced from snurportin 1 by the high affinity form of CRM1 in the presence of RanGTP. Because the m3G-cap binds snurportin 1 very tightly, displacement would only be efficient if the CRM1/RanGTP complex would bind with at least comparable affinity. In fact, Fig. 5 D shows that CRM1 binds snurportin 1 50 times more tightly (K d ≈ 10 nM) than the HIV Rev protein (K d ≈ 0.5 μM). The difference is even more dramatic when compared with the isolated NES from HIV Rev. A peptide comprising the Rev activation domain plus a few flanking residues bound CRM1 700 times weaker (K d ≈ 7 μM), and the minimum Rev activation domain itself even 5,000-fold weaker (K d ≈ 50 μM) than snurportin 1.
Snurportin 1 Interacts with CRM1 Not through a Small NES, but Rather via a Large Domain
The 5,000-fold higher affinity of CRM1 for snurportin 1 compared with the affinity for the minimum Rev activation domain (Rev-NES) raised the question as to the nature of snurportin's export signal. Snurportin 1 contains an IBB domain for importin β binding (residues 26–65), while m3G-cap binding has been attributed to the part of the protein that is COOH-terminal to the IBB (Huber et al., 1998). Analysis of the amino acid sequence of snurportin 1 did not reveal the existence of a single amino acid sequence matching the consensus leucine-rich NES; rather, several regions were found which distantly resemble it. To delineate the binding site for CRM1 on snurportin 1 we expressed NH2- and COOH-terminal snurportin 1 deletion mutants in E. coli, immobilized them, and tested their capacity to bind CRM1 in the presence or absence of RanQ69L (Fig. 6 A). As a control, we tested the same mutants for binding to importin β and found, as expected, that all deletion mutants containing the IBB domain (residues 26–65) also bound importin β (Fig. 6 B).
Figure 6.
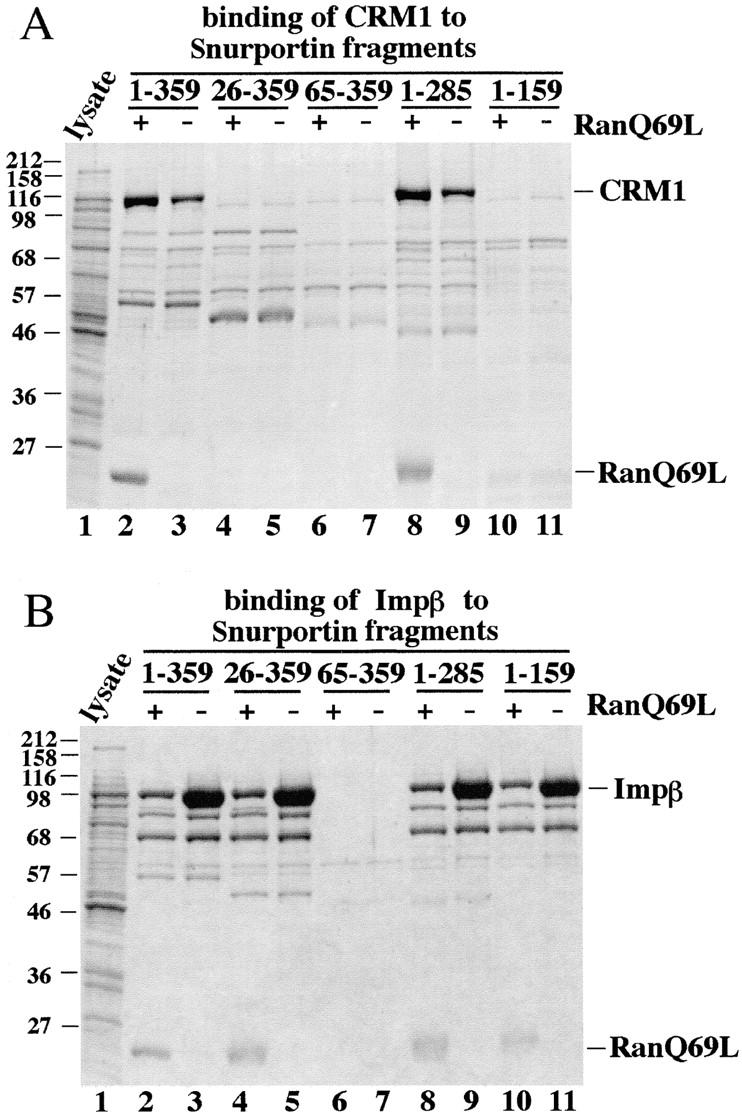
Binding properties of snurportin 1 deletion mutants. Immobilized full-length snurportin 1 (lanes 2 and 3) or deletion mutants (lanes 4–11) were tested for binding of (A) CRM1 and (B) importin β from total E. coli lysates. Where indicated, 1.5 μM RanQ69L GTP had also been added. Analysis was as in Fig. 2. Note that deletion of the 26 NH2-terminal residues of snurportin 1 abolished binding to CRM1. Deletion of >74 residues from the COOH terminus of snurportin 1 also resulted in the loss of the CRM1 interaction. All fragments that contained the IBB domain (residues 25–65) bound importin β.
Deletion of as few as 26 amino acids from the NH2 terminus prevented CRM1 binding and also nuclear export in the permeabilized HeLa cell assay (Fig. 6 A and data not shown). A snurportin 1 fragment lacking 74 amino acids from the COOH terminus (residues 1–285) retained the ability to bind CRM1 in the solution binding assay (Fig. 6 A); however, the kinetic assay indicated that the deletion lowered the affinity for CRM1 by 60% (not shown). A smaller fragment comprising residues 1–159 lost CRM1 binding entirely. Therefore, it appears that binding to CRM1 is sensitive to deletions from both the NH2 and the COOH termini. Thus, in contrast to the short export signals characterized previously, snurportin 1 uses a large domain of at least 159 residues, but probably >285 residues, to bind CRM1. Such a domain could make more contacts to CRM1 and thereby bind more tightly than a simple NES. However, the main advantage might be that a conformational change in this domain can allow regulation of affinity.
Snurportin 1 Is a Potent Competitor of CRM1-dependent Pathways In Vivo
We next wanted to know if snurportin 1 would also use CRM1 for reexport to the cytoplasm in Xenopus oocytes. As an initial experiment, we found that snurportin 1 indeed gets rapidly exported when injected into nuclei of Xenopus oocytes (not shown). If this rapid export is mediated by CRM1, then one would expect that snurportin 1 also competes other CRM1-mediated export pathways. To test this, we injected a mixture of 32P-labeled RNAs consisting of DHFR mRNA, histone H4 mRNA, U1ΔSm RNA, U5ΔSm RNA, U6Δss RNA, and tRNAi Met into nuclei of Xenopus oocytes. After 180 min, the mRNAs, the U1 and U5 RNAs, and the tRNA had been efficiently exported to the cytoplasm (Fig. 7). U snRNA export is known to require CRM1 (Fornerod et al., 1997a) and when GST-snurportin 1 was coinjected, export of U1 and U5 was severely inhibited, but export of tRNA and the two mRNAs remained unaffected. Thus, one can conclude that snurportin 1 accesses the CRM1 export pathway also in this cell type. Furthermore, the very high affinity for CRM1 should make snurportin 1 a quite useful tool to characterize export of other substrates, and sensitivity towards competition by snurportin 1 should be a stringent test for a CRM1 requirement.
Figure 7.

Effects of nuclear injection of snurportin 1 on RNA export. Xenopus laevis oocyte nuclei were coinjected with a mixture of 32P-labeled RNAs and, where indicated, with purified recombinant GST-snurportin 1 fusion protein at the concentrations indicated above the lanes. The mixture of RNAs consisted of DHFR mRNA, histone H4 mRNA, U1ΔSm snRNA, U5ΔSm snRNA, U6Δss snRNA, and human initiator methionyl tRNA. The ΔSm U snRNAs lack the Sm binding site required for reimport into the nucleus. U6Δss does not leave the nucleus and is an internal control for nuclear integrity and proper nuclear injection. Synthesis of DHFR and histone H4 mRNA, and U1ΔSm and U5ΔSm, was primed with the m7GpppG-cap dinucleotide and synthesis of U6Δss was primed with γ-mGTP. RNA samples from total oocytes (T) or cytoplasmic (C) and nuclear (N) fractions were collected immediately after injection in lanes 1–3 or 180 min after injection in lanes 4–15. RNAs were resolved on 8% acrylamide/7 M urea denaturing gels. Note that coinjection of snurportin 1 specifically competed export of U1 and U5 snRNA but had only a negligible effect on mRNA export.
Discussion
Transport between nucleus and cytoplasm appears to be largely mediated by adapter molecules and Ran-binding transport receptors that shuttle between the two compartments. Multiple rounds of import require a cyclic process that results not only in the transport of the cargo to the other side of the nuclear membrane, but also in the subsequent restoration of the original state of the transport system (recycling).
We have investigated here a recycling reaction for U snRNP import, namely the reexport of snurportin 1 back to the cytoplasm (Fig. 8). Snurportin 1 binds m3G-capped U snRNP in the cytoplasm (Huber et al., 1998) and is imported by importin β into the nucleus (Fig. 4; Palacios et al., 1997; Huber et al., 1998). There, importin β binds RanGTP, which destabilizes the interaction with snurportin 1 (Fig. 6). Interestingly, the snurportin/importin β/RanGTP complex is considerably more stable than the corresponding complex with importin α and can even be isolated in small amounts (Figs. 2 B and 6). This could indicate that the actual dissociation of snurportin 1 from importin β does not occur immediately after NPC passage, but further inside the nucleus. Snurportin 1, then, has to release the m3G-capped import substrate before it can form a trimeric export complex with CRM1 and RanGTP. The formation of this complex is highly cooperative and we estimate that snurportin 1 increases the affinity of CRM1 for RanGTP ∼1,000 fold (Fig. 3 A). The export complex would then be transferred to the cytoplasm, where it must be disassembled.
Figure 8.

The snurportin 1 nucleocytoplasmic transport cycle. (1) In the cytoplasm, snurportin 1 binds m3G-capped U snRNAs and (2) is translocated by importin β into the nucleus. Translocation is terminated by direct binding of nuclear RanGTP to importin β (3), which results in the dissociation of importin β from snurportin. (4) Release of the m3G-capped U snRNA from snurportin 1 is prerequisite for (5) incorporation of snurportin 1 into a trimeric export complex with CRM1 and RanGTP. The trimeric complex is exported to the cytoplasm where (6) RanGTP is removed from CRM1 through the action of RanBP1 and RanGAP. (7) CRM1 is displaced from snurportin 1 by the next m3G-capped import substrate and a new transport cycle can proceed. β stands for importin β, Snp for snurportin 1; Sm for the Sm core domain; and m3G for the m3G-cap import signal of U snRNPs.
This disassembly apparently occurs in several steps, involves GTP hydrolysis, and requires RanBP1 and RanGAP1 (Fig. 3 A). CRM1-bound RanGTP resists GTPase activation by RanGAP. Therefore, RanBP1 probably causes an initial release of Ran in the form of a RanBP1/ RanGTP complex in which the RanGTPase can be activated by RanGAP1 (Bischoff and Görlich, 1997). GTP hydrolysis is the irreversible step of the disassembly because RanGDP cannot rebind to CRM1. Removal of RanGTP could alternatively be accomplished by the RanBP2/ SUMO-RanGAP1 complex that is located at the cytoplasmic filaments of the NPC (Mahajan et al., 1997; Saitoh et al., 1997).
Although Ran-free CRM1 is the low affinity form, the remaining CRM1/snurportin 1 heterodimer is still quite stable. However, binding of an m3G-capped substrate displaces CRM1 (Fig. 5). The snurportin/importin β complex would then form and mediate import of the next U snRNP.
For productive transport cycles, carrier molecules need to have different affinities for their substrates during nuclear entry and exit. Ran-binding transport receptors can regulate their affinity for cargo with the aid of the RanGTPase. Adapter molecules do not directly bind RanGTP and, instead, they apparently use differential binding of transport receptors to coordinate the interaction with their cargoes. The first example for that has been import by importin α. Binding of importin β on the way into the nucleus increases the affinity importin α for the NLS (Rexach and Blobel, 1995; Görlich et al., 1996c). Conversely, CAS preferentially binds and exports NLS-free importin α (Kutay et al., 1997). A second precedence is the nuclear CBC which binds 7 monomethyl-cap structures and promotes export of m7G-capped RNAs (Izaurralde et al., 1995). After export, CBC needs to release the RNA before it is reimported by the importin α/β heterodimer. CBC can simultaneously bind a m7G-cap and importin α. However, binding of importin β to the importin α/CBC/ RNA complex causes the displacement of the RNA, allowing CBC to reenter the nucleus without its export substrate (Görlich et al., 1996b).
Here we show that the affinity of snurportin 1 for its import substrate is regulated by CRM1. The molecular basis for the mutual exclusivity of CRM1 and m3G-cap binding to snurportin 1 is still unknown. The two binding sites in snurportin 1 are not identical because the 65 NH2-terminal residues are dispensable for m3G-cap binding (Huber et al., 1998) but are required for the interaction with CRM1 (Fig. 6). However, the binding sites could overlap and thereby exclude simultaneous binding. Alternatively, snurportin 1 might adopt two distinct conformations, one with a high affinity for a m3G-cap, but weak CRM1 binding, and the other with the opposite preference. U snRNP release from snurportin 1 could occur spontaneously and rebinding could be prevented by CRM1. Alternatively, the release would be more efficient if CRM1 could actively displace m3G from snurportin, i.e., not only shift the equilibrium, but also increase the off-rate. In addition, it should be noted that formation of spliceosomes would also retain the mature U snRNP in the nucleus.
Since its discovery as the receptor for leucine-rich NESs (Fornerod et al., 1997b; Stade et al., 1997), CRM1 has been implicated in nuclear export of a number of substrates including MAPKs (Ferrigno et al., 1998), cyclin B1 (Hagting et al., 1998; Yang et al., 1998), NFAT (Kehlenbach et al., 1998), actin (Wada et al., 1998), Dsk1p (Fukuda et al., 1997), and Pap1p (Kuge et al., 1998; Toone et al., 1998). The involvement of CRM1 in the export of the majority of these substrates was suggested by the identification of a leucine-rich NES and/or their sensitivity to the drug leptomycin B, a CRM1 inhibitor (Nishi et al., 1994). A direct physical interaction between CRM1 and its potential export substrates has not always been demonstrated and it is still possible that additional bridging factors are involved. Apart from mediating the export of RRE-containing viral RNAs through Rev, CRM1 is also believed to participate in the export of U snRNPs (Fischer et al., 1995; Fornerod et al., 1997a) and possibly in mRNA export in yeast (Segref et al., 1997; Stade et al., 1997; Watkins et al., 1998). Therefore, it appears that from the three exportins identified so far CRM1 has the broadest range of substrates. In contrast, CAS (Kutay et al., 1997) and exportin-t (Arts et al., 1998; Hellmuth et al., 1998; Kutay et al., 1998; Sarkar and Hopper, 1998) appear to have each specialized in the export of a single extremely abundant class of export substrates, the members of the importin α family and tRNA, respectively.
The question of whether CRM1 is involved in mRNA export in higher eukaryotes has been a controversial issue recently (reviewed by Stutz and Rosbash, 1998). In contrast to the experimental evidence against a role of Crm1p in mRNA export provided by Fischer et al. (1995) and Fornerod et al. (1997a), a later study arrived at the opposite conclusion (Pasquinelli et al., 1997). The fact that mRNA export from Xenopus oocytes nuclei resists competition by snurportin 1 (Fig. 7) confirms the conclusions by Fornerod et al. (1997a) and Fischer et al. (1995) and makes it very unlikely that mRNA export in higher eukaryotes is mediated by CRM1.
CRM1 substrates other than snurportin 1 have been suggested to interact with CRM1 by virtue of a leucine-rich stretch of 8–10 amino acids, a prototype being the Rev activation domain (Fischer et al., 1995; Wen et al., 1995; Bogerd et al., 1996). Surprisingly, however, the isolated activation domain binds CRM1 100 times weaker than the full-length Rev protein (Fig. 5 D). This could have two explanations. First, a leucine-rich export signal might need an appropriate protein context to adopt the conformation required for high-affinity CRM1 binding. In addition, our data indicate that residues flanking a given NES can also significantly contribute to its affinity for CRM1 (Fig. 5 D).
CRM1 binds snurportin 1 still 50-fold stronger than the full-length Rev protein. The probable reason for this difference is that CRM1 has to displace the avidly binding m3G-capped import substrate. The structural basis for the high affinity of the snurportin/CRM1 interaction is still unclear, but nearly the entire snurportin 1 molecule appears to be required for it. The amino acid sequence of snurportin 1 does not contain a single perfect match to the consensus leucine-rich NES, but several regions that appear to resemble it. The affinity might be that high because several of these imperfect NESs interact synergistically with the same CRM1 molecule. However, the interaction of CRM1 with snurportin 1 might also be completely different from that with the prototype NES. This would not be the first case for a transport receptor recognizing very different signals. Transportin, for example, can bind the M9 import signal, which does not have a single essential basic residue, and it also binds the extremely basic import signal from the ribosomal protein L23a (Pollard et al., 1996; Jäkel and Görlich, 1998).
Acknowledgments
We would like to thank Petra Schwarzmaier for excellent technical help, Dr. Sekine for the kind gift of the m3G-cap oligonucleotide, Drs. Maarten Fornerod and Iain W. Mattaj for a Crm1 clone, and Dr. Howard Fried for critical reading of the manuscript.
This work was supported by grants from the Deutsche Forschungsgemeinschaft (SFBs 352 and 286) and the Human Frontier Science Programme Organization (grant RG0198/1998M).
Abbreviations used in this paper
- CBC
cap-binding complex
- GAP
GTPase-activating protein
- IBB domain
importin β–binding domain
- NES
nuclear export signal
- NLS
nuclear localization signal
- NPC
nuclear pore complex
- U snRNP
uridine-rich small nuclear RNP
References
- Adam SA, Sterne R, Marr, Gerace L. Nuclear protein import in permeabilized mammalian cells requires soluble cytoplasmic factors. J Cell Biol. 1990;111:807–816. doi: 10.1083/jcb.111.3.807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arts GJ, Fornerod M, Mattaj IW. Identification of a nuclear export receptor for tRNA. Curr Biol. 1998;8:305–314. doi: 10.1016/s0960-9822(98)70130-7. [DOI] [PubMed] [Google Scholar]
- Bischoff FR, Görlich D. RanBP1 is crucial for the release of RanGTP from importin β-related nuclear transport factors. FEBS Lett. 1997;419:249–254. doi: 10.1016/s0014-5793(97)01467-1. [DOI] [PubMed] [Google Scholar]
- Bischoff FR, Krebber H, Kempf T, Hermes I, Ponstingl H. Human RanGTPase activating protein RanGAP1 is a homolog of yeast RNA1p involved in messenger RNA processing and transport. Proc Natl Acad Sci USA. 1995a;92:1749–1753. doi: 10.1073/pnas.92.5.1749. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bischoff FR, Krebber H, Smirnova E, Dong WH, Ponstingl H. Coactivation of RanGTPase and inhibition of GTP dissociation by Ran GTP binding protein RanBP1. EMBO (Eur Mol Biol Organ) J. 1995b;14:705–715. doi: 10.1002/j.1460-2075.1995.tb07049.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boche I, Fanning E. Nucleocytoplasmic recycling of the nuclear localization signal receptor alpha subunit in vivo is dependent on a nuclear export signal, energy, and RCC1. J Cell Biol. 1997;139:313–325. doi: 10.1083/jcb.139.2.313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bogerd HP, Fridell RA, Benson RE, Hua J, Cullen BR. Protein sequence requirements for function of the human T-cell leukemia virus type 1 Rex nuclear export signal delineated by a novel in vivo randomization-selection assay. Mol Cell Biol. 1996;16:4207–4214. doi: 10.1128/mcb.16.8.4207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chi NC, Adam EJH, Adam SA. Sequence and characterization of cytoplasmic nuclear import factor p97. J Cell Biol. 1995;130:265–274. doi: 10.1083/jcb.130.2.265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chi NC, Adam EJH, Visser GD, Adam SA. RanBP1 stabilizes the interaction of Ran with p97 in nuclear protein import. J Cell Biol. 1996;135:559–569. doi: 10.1083/jcb.135.3.559. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dahlberg JE, Lund E. Functions of the GTPase Ran in RNA export from the nucleus. Curr Opin Cell Biol. 1998;10:400–408. doi: 10.1016/s0955-0674(98)80017-3. [DOI] [PubMed] [Google Scholar]
- Ferrigno P, Posas F, Koepp D, Saito H, Silver PA. Regulated nucleo/cytoplasmic exchange of HOG1 MAPK requires the importin beta homologs NMD5 and XPO1. EMBO (Eur Mol Biol Organ) J. 1998;17:5606–5614. doi: 10.1093/emboj/17.19.5606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer U, Lührmann R. An essential signaling role for the m3G cap in the transport of U1 snRNP to the nucleus. Science. 1990;249:786–790. doi: 10.1126/science.2143847. [DOI] [PubMed] [Google Scholar]
- Fischer U, Darzynkiewicz E, Tahara SM, Dathan NA, Lührmann R, Mattaj IW. Diversity in the signals required for nuclear accumulation of U snRNPs and variety in the pathways of nuclear transport. J Cell Biol. 1991;113:705–714. doi: 10.1083/jcb.113.4.705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer U, Sumpter V, Sekine M, Satoh T, Lührmann R. Nucleo-cytoplasmic transport of U snRNPs: definition of a nuclear location signal in the Sm core domain that binds a transport receptor independently of the m3G cap. EMBO (Eur Mol Biol Organ) J. 1993;12:573–583. doi: 10.1002/j.1460-2075.1993.tb05689.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fischer U, Huber J, Boelens WC, Mattaj IW, Lührmann R. The HIV-1 Rev activation domain is a nuclear export signal that accesses an export pathway used by specific cellular RNAs. Cell. 1995;82:475–483. doi: 10.1016/0092-8674(95)90436-0. [DOI] [PubMed] [Google Scholar]
- Floer M, Blobel G, Rexach M. Disassembly of RanGTP-karyopherin beta complex, an intermediate in nuclear protein import. J Biol Chem. 1997;272:19538–19546. doi: 10.1074/jbc.272.31.19538. [DOI] [PubMed] [Google Scholar]
- Fornerod M, Ohno M, Yoshida M, Mattaj IW. Crm1 is an export receptor for leucine rich nuclear export signals. Cell. 1997a;90:1051–1060. doi: 10.1016/s0092-8674(00)80371-2. [DOI] [PubMed] [Google Scholar]
- Fornerod M, Vandeursen J, Vanbaal S, Reynolds A, Davis D, Murti KG, Fransen J, Grosveld G. The human homologue of yeast Crm1 is in a dynamic subcomplex with Can/Nup214 and a novel nuclear pore component Nup88. EMBO (Eur Mol Biol Organ) J. 1997b;16:807–816. doi: 10.1093/emboj/16.4.807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fridell RA, Truant R, Thorne L, Benson RE, Cullen BR. Nuclear import of hnRNP A1 is mediated by a novel cellular cofactor related to karyopherin-β. J Cell Sci. 1997;110:1325–1331. doi: 10.1242/jcs.110.11.1325. [DOI] [PubMed] [Google Scholar]
- Fukuda M, Asano S, Nakamura T, Adachi M, Yoshida M, Yanagida M, Nishida E. CRM1 is responsible for intracellular transport mediated by the nuclear export signal. Nature. 1997;390:308–311. doi: 10.1038/36894. [DOI] [PubMed] [Google Scholar]
- Görlich D. Nuclear protein import. Curr Opin Cell Biol. 1997;9:412–419. doi: 10.1016/s0955-0674(97)80015-4. [DOI] [PubMed] [Google Scholar]
- Görlich D, Prehn S, Laskey RA, Hartmann E. Isolation of a protein that is essential for the first step of nuclear protein import. Cell. 1994;79:767–778. doi: 10.1016/0092-8674(94)90067-1. [DOI] [PubMed] [Google Scholar]
- Görlich D, Kostka S, Kraft R, Dingwall C, Laskey RA, Hartmann E, Prehn S. Two different subunits of importin cooperate to recognize nuclear localization signals and bind them to the nuclear envelope. Curr Biol. 1995a;5:383–392. doi: 10.1016/s0960-9822(95)00079-0. [DOI] [PubMed] [Google Scholar]
- Görlich D, Vogel F, Mills AD, Hartmann E, Laskey RA. Distinct functions for the two importin subunits in nuclear protein import. Nature. 1995b;377:246–248. doi: 10.1038/377246a0. [DOI] [PubMed] [Google Scholar]
- Görlich D, Henklein P, Laskey RA, Hartmann E. A 41 amino acid motif in importin alpha confers binding to importin beta and hence transit into the nucleus. EMBO (Eur Mol Biol Organ) J. 1996a;15:1810–1817. [PMC free article] [PubMed] [Google Scholar]
- Görlich D, Kraft R, Kostka S, Vogel F, Hartmann E, Laskey RA, Mattaj IW, Izaurralde E. Importin provides a link between nuclear protein import and U snRNA export. Cell. 1996b;87:21–32. doi: 10.1016/s0092-8674(00)81319-7. [DOI] [PubMed] [Google Scholar]
- Görlich D, Panté N, Kutay U, Aebi U, Bischoff FR. Identification of different roles for RanGDP and RanGTP in nuclear protein import. EMBO (Eur Mol Biol Organ) J. 1996c;15:5584–5594. [PMC free article] [PubMed] [Google Scholar]
- Görlich D, Dabrowski M, Bischoff FR, Kutay U, Bork P, Hartmann E, Prehn S, Izaurralde E. A novel class of RanGTP binding proteins. J Cell Biol. 1997;138:65–80. doi: 10.1083/jcb.138.1.65. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hagting A, Karlsson C, Clute P, Jackman M, Pines J. MPF localization is controlled by nuclear export. EMBO (Eur Mol Biol Organ) J. 1998;17:4127–4138. doi: 10.1093/emboj/17.14.4127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hamm J, Mattaj IW. Monomethylated cap structures facilitate RNA export from the nucleus. Cell. 1990;63:109–118. doi: 10.1016/0092-8674(90)90292-m. [DOI] [PubMed] [Google Scholar]
- Hamm J, Darzynkiewicz E, Tahara SM, Mattaj IW. The trimethylguanosine cap structure of U1 snRNA is a component of a bipartite nuclear targeting signal. Cell. 1990;62:569–577. doi: 10.1016/0092-8674(90)90021-6. [DOI] [PubMed] [Google Scholar]
- Hellmuth K, Lau DM, Bischoff FR, Kunzler M, Hurt E, Simos G. Yeast los1p has properties of an exportin-like nucleocytoplasmic transport factor for tRNA. Mol Cell Biol. 1998;18:6374–6386. doi: 10.1128/mcb.18.11.6374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huber J, Cronshagen U, Kadokura M, Marshallsay C, Wada T, Sekine M, Lührmann R. Snurportin 1, an m3G-cap-specific nuclear import receptor with a novel domain structure. EMBO (Eur Mol Biol Organ) J. 1998;17:4114–4126. doi: 10.1093/emboj/17.14.4114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Imamoto N, Shimamoto T, Kose S, Takao T, Tachibana T, Matsubae M, Sekimoto T, Shimonishi Y, Yoneda Y. The nuclear pore targeting complex binds to nuclear pores after association with a karyophile. FEBS Lett. 1995;368:415–419. doi: 10.1016/0014-5793(95)00699-a. [DOI] [PubMed] [Google Scholar]
- Izaurralde E, Adam SA. Transport of macromolecules between the nucleus and the cytoplasm. RNA. 1998;4:351–364. [PMC free article] [PubMed] [Google Scholar]
- Izaurralde E, Lewis J, McGuigan C, Jankowska M, Darzynkiewicz E, Mattaj IW. A nuclear cap binding protein complex involved in pre-mRNA splicing. Cell. 1994;78:657–668. doi: 10.1016/0092-8674(94)90530-4. [DOI] [PubMed] [Google Scholar]
- Izaurralde E, Lewis J, Gamberi C, Jarmolowski A, McGuigan C, Mattaj IW. A cap-binding protein complex mediating U snRNA export. Nature. 1995;376:709–712. doi: 10.1038/376709a0. [DOI] [PubMed] [Google Scholar]
- Izaurralde E, Kutay U, von Kobbe C, Mattaj IW, Görlich D. The asymmetric distribution of the constituents of the Ran system is essential for transport into and out of the nucleus. EMBO (Eur Mol Biol Organ) J. 1997;16:6535–6547. doi: 10.1093/emboj/16.21.6535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jäkel S, Görlich D. Importin beta, transportin, RanBP5, and RanBP7 mediate nuclear import of ribosomal proteins in mammalian cells. EMBO (Eur Mol Biol Organ) J. 1998;17:4491–4502. doi: 10.1093/emboj/17.15.4491. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jarmolowski A, Boelens WC, Izaurralde E, Mattaj IW. Nuclear export of different classes of RNA is mediated by specific factors. J Cell Biol. 1994;124:627–635. doi: 10.1083/jcb.124.5.627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kehlenbach RH, Dickmanns A, Gerace L. Nucleocytoplasmic shuttling factors including Ran and CRM1 mediate nuclear export of NFAT in vitro. J Cell Biol. 1998;141:863–874. doi: 10.1083/jcb.141.4.863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuge S, Toda T, Iizuka N, Nomoto A. Crm1 (XpoI) dependent nuclear export of the budding yeast transcription factor yAP-1 is sensitive to oxidative stress. Genes Cells. 1998;3:521–532. doi: 10.1046/j.1365-2443.1998.00209.x. [DOI] [PubMed] [Google Scholar]
- Kutay U, Bischoff FR, Kostka S, Kraft R, Görlich D. Export of importin alpha from the nucleus is mediated by a specific nuclear transport factor. Cell. 1997;90:1061–1071. doi: 10.1016/s0092-8674(00)80372-4. [DOI] [PubMed] [Google Scholar]
- Kutay U, Lipowsky G, Izaurralde E, Bischoff FR, Schwarzmaier P, Hartmann E, Görlich D. Identification of a tRNA-specific nuclear export receptor. Mol Cell. 1998;1:359–369. doi: 10.1016/s1097-2765(00)80036-2. [DOI] [PubMed] [Google Scholar]
- Lounsbury KM, Macara IG. Ran-binding protein 1 (RanBP1) forms a ternary complex with Ran and karyopherin beta and reduces GTPase-activating protein (RanGAP) inhibition by karyopherin beta. J Biol Chem. 1997;272:551–555. doi: 10.1074/jbc.272.1.551. [DOI] [PubMed] [Google Scholar]
- Mahajan R, Delphin C, Guan T, Gerace L, Melchior F. A small ubiquitin-related polypeptide involved in targeting RanGAP1 to nuclear pore complex protein RanBP2. Cell. 1997;88:97–107. doi: 10.1016/s0092-8674(00)81862-0. [DOI] [PubMed] [Google Scholar]
- Mattaj IW. Cap trimethylation of U snRNA is cytoplasmic and dependent on U snRNP protein binding. Cell. 1986;46:905–911. doi: 10.1016/0092-8674(86)90072-3. [DOI] [PubMed] [Google Scholar]
- Mattaj IW, Englmeier L. Nucleocytoplasmic transport: the soluble phase. Ann Rev Biochem. 1998;67:265–306. doi: 10.1146/annurev.biochem.67.1.265. [DOI] [PubMed] [Google Scholar]
- Matunis MJ, Wu J, Blobel G. SUMO-1 modification and its role in targeting the Ran GTPase-activating protein, RanGAP1, to the nuclear pore complex. J Cell Biol. 1998;140:499–509. doi: 10.1083/jcb.140.3.499. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nigg EA. Nucleocytoplasmic transport: signals, mechanisms and regulation. Nature. 1997;386:779–787. doi: 10.1038/386779a0. [DOI] [PubMed] [Google Scholar]
- Nishi K, Yoshida M, Fujiwara D, Nishikawa M, Horinouchi S, Beppu T. Leptomycin B targets a regulatory cascade of crm1, a fission yeast nuclear protein, involved in control of higher order chromosome structure and gene expression. J Biol Chem. 1994;269:6320–6324. [PubMed] [Google Scholar]
- Palacios I, Hetzer M, Adam SA, Mattaj IW. Nuclear import of U snRNPs requires importin beta. EMBO (Eur Mol Biol Organ) J. 1997;16:6783–6792. doi: 10.1093/emboj/16.22.6783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pasquinelli AE, Powers MA, Lund E, Forbes D, Dahlberg JE. Inhibition of mRNA export in vertebrate cells by nuclear export signal conjugates. Proc Natl Acad Sci USA. 1997;94:14394–14399. doi: 10.1073/pnas.94.26.14394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plessel G, Fischer U, Lührmann R. m3G cap hypermethylation of U1 small nuclear ribonucleoprotein (snRNP) in vitro: evidence that the U1 small nuclear RNA-(guanosine-N2)-methyltransferase is a non-snRNP cytoplasmic protein that requires a binding site on the Sm core domain. Mol Cell Biol. 1994;14:4160–4172. doi: 10.1128/mcb.14.6.4160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pollard VW, Michael WM, Nakielny S, Siomi MC, Wang F, Dreyfuss G. A novel receptor mediated nuclear protein import pathway. Cell. 1996;88:985–994. doi: 10.1016/s0092-8674(00)80173-7. [DOI] [PubMed] [Google Scholar]
- Radu A, Blobel G, Moore MS. Identification of a protein complex that is required for nuclear protein import and mediates docking of import substrate to distinct nucleoporins. Proc Natl Acad Sci USA. 1995;92:1769–1773. doi: 10.1073/pnas.92.5.1769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rexach M, Blobel G. Protein import into nuclei: association and dissociation reactions involving transport substrate, transport factors, and nucleoporins. Cell. 1995;83:683–692. doi: 10.1016/0092-8674(95)90181-7. [DOI] [PubMed] [Google Scholar]
- Ribbeck K, Lipowsky G, Kent HM, Stewart M, Görlich D. NTF2 mediates nuclear import of Ran. EMBO (Eur Mol Biol Organ) J. 1998;17:6587–6598. doi: 10.1093/emboj/17.22.6587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richards SA, Lounsbury KM, Carey KL, Macara IG. A nuclear export signal is essential for the cytosolic localization of the Ran binding protein, RanBP1. J Cell Biol. 1996;134:1157–1168. doi: 10.1083/jcb.134.5.1157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rout MP, Blobel G, Aitchison JD. A distinct nuclear import pathway used by ribosomal proteins. Cell. 1997;89:715–725. doi: 10.1016/s0092-8674(00)80254-8. [DOI] [PubMed] [Google Scholar]
- Saitoh H, Pu R, Cavenagh M, Dasso M. RanBP2 associates with Ubc9p and a modified form of RanGAP1. Proc Natl Acad Sci USA. 1997;94:3736–3741. doi: 10.1073/pnas.94.8.3736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarkar S, Hopper AK. tRNA nuclear export in Saccharomyces cerevisiae: in situ hybridization analysis. Mol Biol Cell. 1998;9:3041–3055. doi: 10.1091/mbc.9.11.3041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlenstedt G, Smirnova E, Deane R, Solsbacher J, Kutay U, Görlich D, Ponstingl H, Bischoff FR. Yrb4p, a yeast Ran GTP binding protein involved in import of ribosomal protein L25 into the nucleus. EMBO (Eur Mol Biol Organ) J. 1997;16:6237–6249. doi: 10.1093/emboj/16.20.6237. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Segref A, Sharma K, Doye V, Hellwig A, Huber J, Lührmann R, Hurt E. Mex67p, a novel factor for nuclear mRNA export, binds to both Poly(a)(+) RNA and nuclear pores. EMBO (Eur Mol Biol Organ) J. 1997;16:3256–3271. doi: 10.1093/emboj/16.11.3256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Siomi MC, Eder PS, Kataoka N, Wan L, Liu Q, Dreyfuss G. Transportin-mediated nuclear import of heterogenous RNP proteins. J Cell Biol. 1997;138:1181–1192. doi: 10.1083/jcb.138.6.1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stade K, Ford CS, Guthrie C, Weis K. Exportin 1 (CRM1) is an essential nuclear export factor. Cell. 1997;90:1041–1050. doi: 10.1016/s0092-8674(00)80370-0. [DOI] [PubMed] [Google Scholar]
- Stutz F, Rosbash M. Nuclear RNA export. Genes Dev. 1998;12:3303–3319. doi: 10.1101/gad.12.21.3303. [DOI] [PubMed] [Google Scholar]
- Toone WM, Kuge S, Samuels M, Morgan BA, Toda T, Jones N. Regulation of the fission yeast transcription factor Pap1 by oxidative stress: requirement for the nuclear export factor Crm1 (exportin) and the stress-activated MAP kinase Sty1/Spc1. Genes Dev. 1998;12:1453–1463. doi: 10.1101/gad.12.10.1453. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wada A, Fukuda M, Mishima M, Nishida E. Nuclear export of actin: a novel mechanism regulating the subcellular localization of a major cytoskeletal protein. EMBO (Eur Mol Biol Organ) J. 1998;17:1635–1641. doi: 10.1093/emboj/17.6.1635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watkins JL, Murphy R, Emtage JL, Wente SR. The human homologue of Saccharomyces cerevisiaeGle1p is required for poly(A)+ RNA export. Proc Natl Acad Sci USA. 1998;95:6779–6784. doi: 10.1073/pnas.95.12.6779. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weis K, Ryder U, Lamond AI. The conserved amino terminal domain of hSRP1α is essential for nuclear protein import. EMBO (Eur Mol Biol Organ) J. 1996;15:1818–1825. [PMC free article] [PubMed] [Google Scholar]
- Wen W, Meinkoth JL, Tsien RY, Taylor SS. Identification of a signal for rapid export of proteins from the nucleus. Cell. 1995;82:463–473. doi: 10.1016/0092-8674(95)90435-2. [DOI] [PubMed] [Google Scholar]
- Yang J, Bardes ES, Moore JD, Brennan J, Powers MA, Kornbluth S. Control of cyclin B1 localization through regulated binding of the nuclear export factor CRM1. Genes Dev. 1998;12:2131–2143. doi: 10.1101/gad.12.14.2131. [DOI] [PMC free article] [PubMed] [Google Scholar]