

Abstract
Teratomas are benign tumors that form after ectopic injection of embryonic stem (ES) cells into mice and contain derivatives of all primitive germ layers. To study the role of β1 integrin during teratoma formation, we compared teratomas induced by normal and β1-null ES cells. Injection of normal ES cells gave rise to large teratomas. In contrast, β1-null ES cells either did not grow or formed small teratomas with an average weight of <5% of that of normal teratomas. Histological analysis of β1-null teratomas revealed the presence of various differentiated cells, however, a much lower number of host-derived stromal cells than in normal teratomas. Fibronectin, collagen I, and nidogen were expressed but, in contrast to normal teratomas, diffusely deposited in β1-null teratomas. Basement membranes were present but with irregular shape and detached from the cell surface.
Normal teratomas had large blood vessels with a smooth inner surface, containing both host- and ES cell–derived endothelial cells. In contrast, β1-null teratomas had small vessels that were loosely embedded into the connective tissue. Furthermore, endothelial cells were always of host-derived origin and formed blood vessels with an irregular inner surface. Although β1- deficient endothelial cells were absent in teratomas, β1-null ES cells could differentiate in vitro into endothelial cells. The formation of a complex vasculature, however, was significantly delayed and of poor quality in β1-null embryoid bodies. Moreover, while vascular endothelial growth factor induced proliferation of endothelial cells as well as an extensive branching of blood vessels in normal embryoid bodies, it had no effect in β1-null embryoid bodies.
A hallmark of tumor cells is their ability to grow anchorage independent. Proliferation and survival of tumor cells, determining progression of solid tumors, are independent of signals elicited by interactions with the surrounding extracellular matrix (ECM1; Folkman and Moscona, 1978). In contrast, normal diploid cells require anchorage to the ECM for proliferation as well as survival (Dike and Farmer, 1988). Several lines of direct evidence show that integrins transduce these signals (Varner and Cheresh, 1996).
Integrins are the most important family of cell surface receptors that mediate cell–matrix interactions (Hynes, 1992). They are heterodimers of noncovalently linked α and β subunits. So far 15 different α subunits and 8 different β subunits are known. The β1 subunit can associate with at least 10 different α subunits forming the largest subfamily of integrins. Members of the β1 integrin subfamily primarily bind to components of the ECM such as fibronectin, collagens, and laminins, but some of them also participate in direct cell–cell adhesion (Hynes, 1992; Haas and Plow, 1994). The cytoplasmic domain of β1 integrin can directly interact with cytoskeletal proteins such as talin and α-actinin and with signal transducing proteins such as focal adhesion kinase (FAK; Schaller et al., 1995) and integrin-linked kinase (Hannigan et al., 1996).
Integrin engagement and clustering regulate shape, motility, survival, and proliferation of cells. These events are executed by integrin-mediated cascades of intracellular signals that include tyrosine phosphorylation of FAK (Guan and Shalloway, 1992), increases in intracellular Ca2+ levels (Schwartz, 1993), intracellular pH (Schwartz et al., 1989, 1990), inositol lipid synthesis (McNamee et al., 1993), and expression of cyclins (Guadagno et al., 1993). Furthermore, it has been demonstrated that integrins can also mediate the activation of protein kinase C (Vuori and Ruoslahti, 1993), mitogen-activated protein kinase (Morino et al., 1995) and NF-κB (Yebra et al., 1995). In addition to these adhesion-mediated signaling pathways, many cells depend on growth factor–mediated signals for appropriate cell cycle progression and proliferation.
In the present study we have used β1 integrin–deficient embryonic stem (ES) cells (Fässler al., 1995) to induce teratomas in syngeneic mice. ES cells as well as pre- or early postimplantation embryos of most mouse strains develop into tumors when transplanted into an ectopic location of syngeneic animals (Damjanov and Solter, 1974; Damjanov, 1978). These tumors are composed of various differentiated somatic tissues and are called teratomas. We show that β1-null ES cells give rise to either very small or no teratomas. The most prominent changes that are associated with the impaired growth in β1-null teratomas are abnormal depositon of ECM proteins and various defects in basement membranes. Furthermore, β1-null teratomas showed an inefficient angiogenesis. A number of studies have demonstrated convincingly that tumor growth is dependent on angiogenesis (Folkman, 1996). Tumor angiogenesis is regulated by factors produced by tumor cells as well as by cell adhesion molecules expressed on endothelial cells. Systemic or local administration of antibodies or cyclic RGD peptides blocking αvβ3 integrin function inhibits tumor angiogenesis and as a consequence promotes tumor regression (Brooks et al., 1994a ). This anti-angiogenic effect of the αvβ3 antagonists results from the activation of apoptosis of newly sprouting blood vessels (Brooks et al., 1994b ). We now show that β1 integrin plays an essential role during angiogenesis in teratomas. In normal teratomas both host- and ES cell–derived endothelial cells contribute to angiogenesis. In contrast, in β1-null teratomas, all vascular cells are exclusively derived from the host. Furthermore, we report that vascular endothelial growth factor (VEGF) treatment–induced proliferation of endothelial cells and extensive branching of blood vessels in normal but not in β1-null embryoid bodies.
MATERIALS AND METHODS
Cells and Cell Culture
The following ES cells were used to induce teratomas: wild-type ES cell line D3 (+/+; Doetschman et al., 1985); G119 (+/−), which is heterozygous for the β1 integrin gene mutation; G101 (+/+), which is wild-type for β1 integrin gene but mock transfected; and G201 (−/−) which is β1 integrin deficient (Fässler et al., 1995). In the cell clones G119 and G201, a fusion DNA of β-galactosidase and neomycin is inserted in frame with the ATG of the β1 integrin gene (Fässler et al., 1995). The cell clone G101 contains a randomly integrated β-galactosidase gene that is ubiquitously expressed.
ES cells were cultured in the absence of a fibroblast feeder layer in DME, supplemented with 20% heat-inactivated FCS (GIBCO BRL, Gaithersburg, MD), 0.1 mM β-mercaptoethanol (Sigma Chemical Co., St. Louis, MO), 1× non-essential amino acids (GIBCO BRL), and 1,000 U/ml recombinant leukemia inhibiting factor (GIBCO BRL).
For differentiation, ES cells were cultured in hanging drops as described previously (Fässler et al., 1996). Briefly, 600 cells were cultured in 20 μl of DME, supplemented with 20% FCS hanging from the lid of the culture dish for 5 d, which allows the formation of cell aggregates (embryoid bodies). Subsequently, the aggregates were plated on Tissue Tek chambers (Nunc, Wiesbaden, Germany) and incubated for 7, 15, or 20 d, respectively, fixed in 4% paraformaldehyde and immunostained for von Willebrand Factor (Behringwerke Ag, Marburg Lahn, Germany) and platelet endothelial cell adhesion molecule (PECAM) (Vecchi et al., 1994).
Alternatively, after outgrowth of cell aggregates for 20 d, cells were trypsinized and cultured on gelatin-coated glass cover slips for 24 h, fixed, and stained for von Willebrand Factor (vWF) or PECAM. Positive cells were counted using an Axiophot fluorescence microscope (Carl Zeiss, Oberkochen, Germany).
For VEGF treatment, ES cells were cultured in hanging drops for 2 d, in bacteriological dishes for 3 d, and then plated on gelatinized glass cover slips for another 12 d. The culture medium was DME, containing 20% FCS with either 10 or 20 ng/ml VEGF (R&D Sytems, Wiesbaden, Germany).
Antibodies
The following primary antibodies were used: rabbit anti–rat β1 integrin; hamster anti–mouse β3 integrin; hamster anti–mouse αv integrin (both from PharMingen, San Diego, CA); hen anti–human FN (Johansson and Höök, 1984); rabbit anti–mouse FN (GIBCO BRL); goat anti-collagen type I; goat anti-collagen type III (both from Southern Biotechnology Associates Inc., Birmingham, AL); rat anti–mouse nidogen (Dziadek et al., 1988); rat anti–mouse PECAM-1 (Vecchi et al., 1994); and rabbit anti– mouse vWF (Behringwerke).
The following secondary antibodies were used: goat anti–rabbit FITC; goat anti–rabbit Cy3; rabbit anti–hamster FITC; goat anti–chicken FITC; donkey anti–goat Cy3; goat anti–rat Cy3 (Jackson Immunoresearch Lab. Inc., West Grove, PA), rabbit anti-digoxygenin FITC (Boehringer Mannheim, Mannheim, Germany), biotinylated goat anti–rabbit Ig (Vector Laboratories, Inc., Burlingame, CA), biotinylated goat anti–rat Ig (Amersham Intl., Little Chalfont, UK), and streptavidin-horseradish peroxidase conjugate (Amersham Intl.).
Teratoma Induction
107 ES cells were trypsinized, washed twice, suspended in 100 μl PBS, and then injected subcutaneously on the back of syngeneic 129/SV male mice. After 21 or 28 d, tumors were surgically removed and frozen in ice-cold isopentan. To analyze cell proliferation, 25 mg per 100 g body weight of the thymidine analogue bromodeoxyuridine (BrdU) was injected intraperitoneally 2.5 h before the excision of the tumors.
Microscopical Analysis of Embryoid Bodies and Tumor Tissue
Light microscopy.
For light microscopical examination, small pieces of teratomas and 20-μm-thick immunostained cryosections were dehydrated conventionally in a graded ethanol series, and finally infiltrated with and embedded in araldite (Serva, Heidelberg, Germany). Semithin sections of 1–2 μm were analyzed using a Zeiss Axiophot microscope (Carl Zeiss), with or without methylene blue staining.
Histochemistry.
Teratomas were surgically removed, frozen in ice-cold isopentan, and stored at −80°C. Tissue specimens were cut into 10-μm-thick sections and collected on glass slides (Shandon, Frankfurt, Germany). Hematoxylin/Eosin and lacZ staining followed published protocols (Fässler et al., 1995). Stained sections were analyzed using a Zeiss Axiophot microscope.
Cell proliferation.
Incorporation of intraperitoneally injected BrdU into the DNA of replicating teratoma cells was analyzed using anti-BrdU monoclonal antibodies, following the protocol supplied by the manufacturer (Boehringer Mannheim).
Similarly, proliferation of cells in embryoid bodies was assayed after addition of BrdU to the culture medium and incubation for another 2 h. Embryoid bodies were fixed as described above, and then processed following the manufacturer's protocol. For development of the immunostaining 3,3-diaminobenzidine-tetrahydrochloride (DAB) with nickel enhancer (12 μl 0.1 PBS, 300 μl DAB, 300 μm NH4Cl, 300 μl NiSO4, 10% glucose, and 50 μl glucose oxidase) was used. This gives rise to a black reaction product that is localized in the nuclei of proliferating cells. Subsequently, sections were incubated with vWF antibody or PECAM-1 antibody (1:800) and processed as described below. Counterstaining was done with methyl green for 35 s. Afterwards, embryoid bodies were dehydrated in a graded ethanol series, coverslipped, and analyzed using a Zeiss Axiophot microscope.
Apoptosis.
Cell death was analyzed following the protocol supplied by the manufacturer (Oncor, Heidelberg, Germany). Briefly, tissue sections were fixed in 4% paraformaldehyde in PBS, postfixed in ethanol/acetic acid (2:1), and incubated with terminal deoxynucleotidyl transferase, which catalyzes the addition of digoxygenin-conjugated nucleotides to free 3′-hydroxyl groups in apoptotic cells. Incorporation of labeled nucleotides was detected by FITC-conjugated, anti-digoxygenin antibodies. The number of apoptotic cells was determined from random sections of three normal and four β1-null teratomas.
Immunofluorescence.
Tumor sections, which were 6-μm thick, were incubated with primary antibody in PBS, supplemented with 1% BSA at dilutions recommended by the suppliers for 1 h, at room temperature. Subsequently, the sections were washed three times in PBS and incubated with fluorescence-labeled secondary antibodies at dilutions recommended by the manufacturer for 1 h, at room temperature. After washing three times in PBS, the fluorescent specimens were mounted using elvanol (Fässler et al., 1995).
For analyzing the expression of αv and β3 integrin on endothelial cells, embryoid bodies were fixed in 4% paraformaldehyde and indirectly double labeled with hamster antibodies against αv or β3 integrin (PharMingen), respectively, and rat antibodies against PECAM-1 (Vecchi et al., 1994). Subsequently, embryoid bodies were washed three times in PBS and incubated with fluorescence-labeled secondary antibodies at dilutions recommended by the manufacturer for 1 h at room temperature. After washing three times in PBS, the fluorescent specimens were mounted using elvanol (Fässler et al., 1995). Analysis was performed using a Zeiss Axiophot fluorescence microscope.
Immunohistochemistry.
For immunohistochemistry, 20-μm-thick cryosections of sucrose-embedded teratomas prefixed with 4% paraformaldehyde in 0.1× PBS, and paraformaldehyde-fixed embryoid bodies were stained. Endogenous peroxidase was blocked by 3% H2O2 in 60% methanol. Subsequently, cells were permeabilized with 0.2% Triton X-100 (Sigma Chemical Co.) in 0.1× PBS. The sections were incubated with von Willebrand antibody (1:1,000) or PECAM antibody (1:800) in PBS. Embryoid bodies were incubated in PBS, supplemented with 0.8% BSA overnight at 4°C. Subsequent sections and embryoid bodies were washed four times in PBS and incubated with biotinylated secondary antibodies for 1 h at room temperature. A streptavidin–horseradish peroxidase complex was then used as a detection system (1:200; incubation for 1 h at room temperature). The staining was developed for 15 min with DAB in 5 mM Tris-HCl buffer, pH 7.4, supplemented with 0.1% H2O2. Analysis was performed using a Zeiss Axiophot microscope. Morphometrical analyses of vessels identified by PECAM antibody were performed with Optimas 5.2 (Optimas Corporation, Bothwell, WA).
Electron Microscopy
Tumor tissue was obtained from 6-wk-old chimeric animals, which were killed by cervical dislocation and subsequently transcardially perfused with a 0.1 mol/liter cacodylate buffer containing 2% paraformaldehyde and 2% glutaraldehyde at a perfusion pressure of 60 cm H2O. Tumors were removed, fixed for an additional 4 h in the same fixative, and then cut into small pieces and fixed in 0.1 mol/liter cacodylate buffer, pH 7.3, containing 2% osmium tetroxide for 2 h at 4°C. Tissue pieces were rinsed three times in cacodylate buffer, block stained for 8 h in 70% ethanol containing 1% uranyl acetate, dehydrated in a series of graded ethanol, and embedded in araldite. Ultrathin sections (30–60 nm) obtained with a diamond knife on a Reichert ultramicrotome (Reichert, Bensheim, Germany) were placed on copper grids and examined with a Zeiss EM 902A electron microscope.
Embryoid bodies were fixed in a 100 mM Hepes/Pipes buffer, pH 7.35, containing 1.75% paraformaldehyde, 2% glutaraldehyde, and 15% picric acid for 1 h at room temperature. Afterwards embryoid bodies were treated with 100 mM Hepes/Pipes buffer containing 1% tannic acid for 30 min at room temperature and finally osmificated with 0.5% OsO4. Before embedding in Epon resin (Agar Scientific, Stansted, UK) embryoid bodies were dehydrated in a graded series of ethanol. For light microscopy, semithin sections were stained with methylene blue. For electron microscopy, ultrathin sections (30–60 nm) were mounted on formvar-coated copper grids, stained with 0.2% uranyl acetate and lead citrate, and then examined with a Zeiss EM 902A electron microscope.
RESULTS
ES Cells Lacking β1 Integrin Form No or Only Small Teratomas
To test whether the lack of β1 integrin has an influence on the development of teratomas normal and β1-null ES cells were injected subcutaneously into syngeneic male mice. After 21 d of incubation, wild-type tumors were clearly visible on the back of the mice. In contrast, β1-null tumors either did not form or were significantly reduced in size (Fig. 1).
Figure 1.

Male 129/Sv mice inoculated with 107 D3 cells (β1+/+; right side of the back) and 107 G 201 cells (β1−/−, left side of the back). The right side of the back was inoculated with normal ES cells. After an incubation period of 3 wk, both mice developed a big teratoma (arrows). The left side of the back was inoculated with β1-null ES cells. Note that a small teratoma formed and is visible in the left mouse (arrowheads) but not in the right mouse.
To investigate the growth of β1-null tumors in more detail, mice were killed by cervical dislocation and examined under the stereomicroscope. In 9 out of 18 animals injected with β1-null ES cells, no tumor could be detected either at the site of injection, or in distant organs. In the remaining nine animals, a small tumor had formed, which was significantly reduced in weight (mean value 47 mg) when compared with tumors derived from normal (0.93 g) or β1 integrin–heterozygous (1.18 g) ES cells (Fig. 2). No difference in tumor growth was observed in mice carrying either a β1-null tumor alone or together with a normal or heterozygous tumor.
Figure 2.
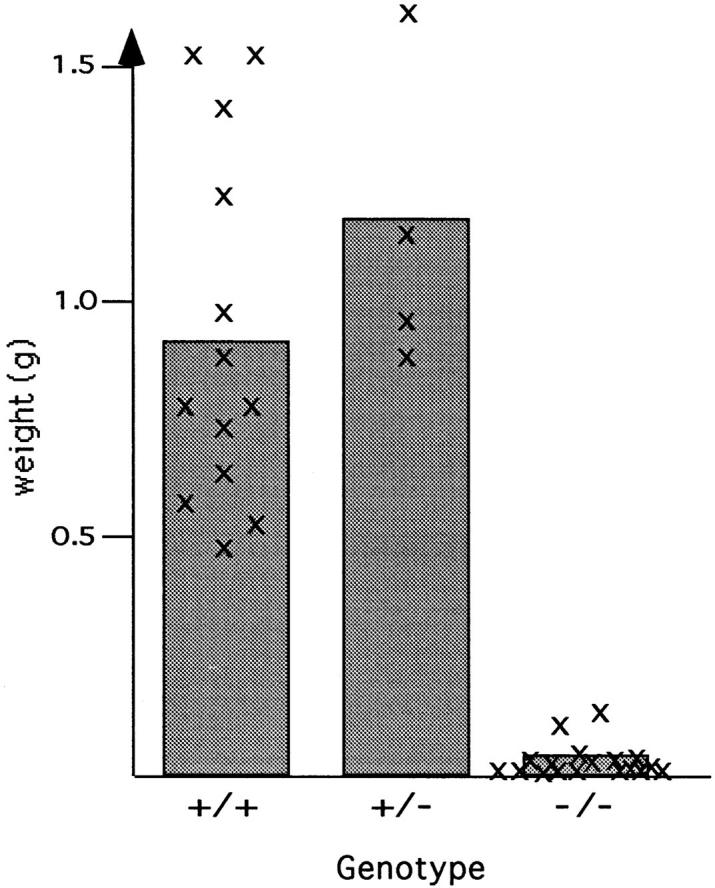
Weights of teratomas derived from normal (+/+), β1-heterozygous (+/−), and β1-null (−/−) ES cells. The mean values were calculated from 13 +/+, 4 +/−, and 9 −/− teratomas, respectively. Individual values are designated by an x.
Cell Proliferation and Apoptosis Is Not Significantly Altered in β1-null Teratomas
To test whether cell proliferation is reduced in β1-null tumors BrdU was injected into tumor-bearing mice. Detection of BrdU in tissue sections with specific antibodies showed that the distribution of replicating cells is similar in normal and β1-null teratomas. Some areas in both tumor types contained nests of many proliferating cells, whereas other areas show a homogenous distribution of a few proliferating cells (Fig. 3). This heterogeneity of the tumor architecture results from the wide range of different tissues that form in teratomas. As a consequence, the number of proliferating as well as apoptotic cells varied tremendously from one area to another; normal tumors grown for 10 d had between 84 and 176 BrdU-postive cells per mm2 tissue section and tumors grown for 21 d had between 220 and 279 BrdU-positive cells per mm2 tissue section. In corresponding sections of β1-null teratomas, the number of proliferating cells ranged between 42 and 116 cells in 10-d-old teratomas, and between 156 and 265 cells in 21-d-old teratomas. This large range in proliferating cells in both tumor types did not provide a statistically significant difference but a tendency towards a reduced proliferation rate in β1-null teratomas.
Figure 3.

Analysis of cell proliferation and apoptosis in a normal (+/+) teratoma and a β1-null (−/−) teratoma. ES cells were injected under the skin of the back and incubated for 21 d. 2 h before tumor isolation BrdU was injected intraperitoneally. Incorporated BrdU was detected using specific antibodies labeled with FITC (A and B). Apoptotic cells were identified by staining for the presence of free 3′-hydroxyl groups in tissue sections of +/+ and −/− teratomas (C and D). Bar, 100 μm.
Similarly, the number of apoptotic cells was very inhomogenously distributed in the tissue of normal and β1-null teratomas: β1+/+, β1+/−, and β1−/− teratomas showed higher apoptosis rates in areas that were lacZ-positive, and thus ES cell–derived and lower rates in areas that were lacZ-negative, and thus host cell–derived (data not shown). Both tumor types showed areas with small cell clusters with a high rate of apoptosis, whereas in other areas the apoptotic cells were evenly distributed (Fig. 3). The number of apoptotic cells ranged between 250 and 712 cells per mm2 tissue in normal teratomas, and between 249 and 805 cells per mm2 tissue section in β1-null teratomas. Also here, the large variation did not provide a statistically significant difference but again a tendency towards a higher apoptosis rate in β1-null teratomas was apparent.
β1-Null Teratomas Contain Differentiated Cells but Have an Altered Tissue Architecture with Less Host-derived Cells
Hematoxylin/Eosin staining revealed that β1 +/+ teratomas (G101) (Fig. 4 A) and β1 +/− teratomas (not shown) are composed of a variety of differentiated cells and tissues, e.g., glandular epithelium, cartilage, connective tissue, and neuronal cells. β1-Null tumors appeared cell dense and contained fewer cords of fibrotic material (Fig. 4 B). Histochemical analysis of several β1-null tumors revealed that many differentiated cells are present in β1-null teratomas. ES cell–derived cells could be differentiated by lacZ activity, since the lacZ gene was introduced in the β1 integrin gene and in the genome of the mock-transfected ES cells (Fässler et al., 1995; and data not shown). Mock-transfected, as well as β1 integrin–heterozygous tumors contained large areas devoid of lacZ activity proving their host-derived origin (Fig. 4 C). In contrast, the number of lacZ-negative cells was low in all β1-null tumors analyzed (Fig. 4 D) indicating that the migration of normal host cells into mutant tumor tissue is reduced.
Figure 4.
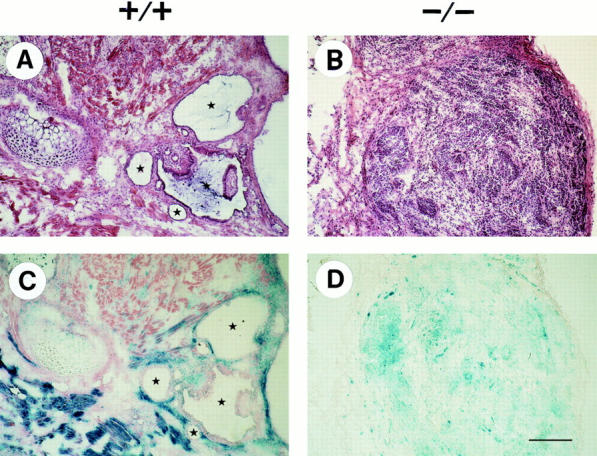
Histology of normal (+/+) and β1-null (−/−) teratomas. Tumors derived from normal ES cells (β1 +/+) and from β1-null ES cells (β1 −/−) were stained with Hematoxylin/Eosin (A and B) and lacZ/eosin (C and D), respectively. Blue, lacZ-positive cells are ES cell-derived whereas lacZ-negative cells are host-derived stromal cells that have migrated into the tumor tissue. Stars indicate lumina of glandular structures. Bar, 100 μm.
Deposition of Extracellular Proteins Is Altered in β1-null Teratomas
To test whether the deposition of matrix proteins is affected by the lack of β1 integrin, the distribution of fibronectin, collagen I, and nidogen were compared between normal and β1-null tumors. In normal tumors most of the fibronectin (Fig. 5 C) was associated with cells expressing large amounts of β1 integrin (Fig. 5 A). Only in a few areas fibronectin staining did not colocalize with β1 integrin staining (data not shown). In β1-null tumors, on the other hand, fibronectin was deposited throughout the tumor tissue (Fig. 5 D) and did not correlate with host cell–derived β1 integrin expression (Fig. 5 B). Similarly, in sections derived from β1-null tumors, collagen I (Fig. 5 F) and nidogen (not shown) were diffusely distributed. This was not the case in normal tumors in which collagen I (Fig. 5 E) and nidogen (not shown) were mainly restricted to areas with strong β1 integrin staining.
Figure 5.
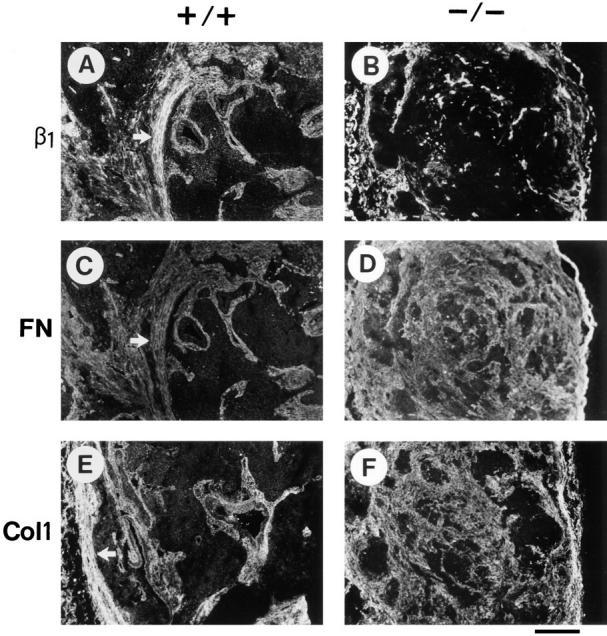
Immunostaining for ECM components in normal (+/+) and in β1-null (−/−) teratomas. The antibodies used were directed against β1 integrin (A and B), fibronectin (C and D), and collagen I (E and F). The staining shown in A and C, and B and D, respectively, are double stainings of the same tissue specimens. Arrows indicate dense cords of connective tissue. Bar, 100 μm.
To evaluate whether the ultrastructural morphology of basement membranes is altered by the lack of β1 integrin expression, electronmicroscopy was performed with tumor tissues and embryoid bodies derived from normal and β1-null ES cells. In normal tumors and normal embryoid bodies the basement membrane was continuously present along the basal surface of epithelial cells (Fig. 6, A and D) or around muscle cells (not shown) with a constant width that never exceeded 30 nm. In contrast, most cells in β1-null teratomas and embryoid bodies were covered by partially detached basement membranes, which resulted in the formation of irregular clefts between plasma membrane and lamina densa (Fig. 6, B and F). In other areas, cells were covered by several layers of basement membranes (Fig. 6 C) or by only partially developed basement membranes (Fig. 6, E and F).
Figure 6.
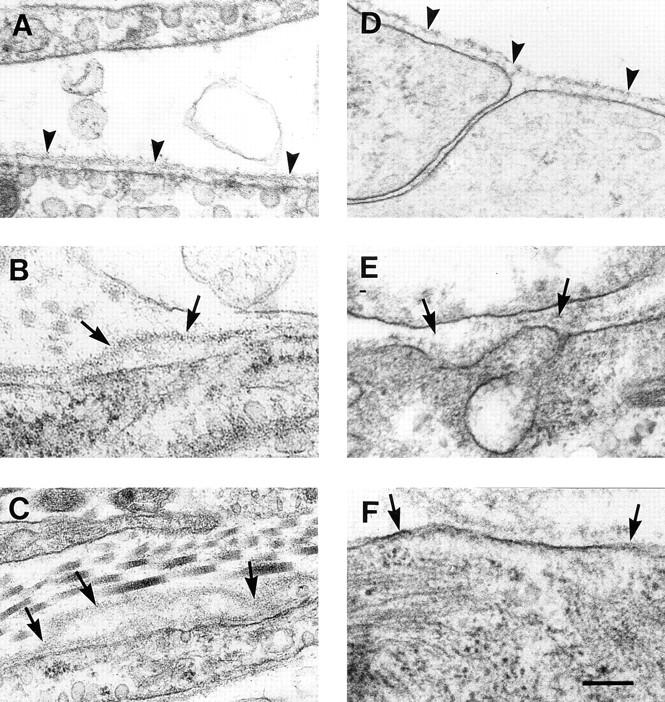
Electronmicroscopy of normal (A and D) and β1-null teratomas and embryoid bodies (B, C, E, and F). Normal cells in teratomas (A) and embryoid bodies (D) are covered by a smooth basement membrane that is strictly located in close vicinity of the cell surface (arrowhead). In β1-null teratomas as well as β1-null embryoid bodies, the basement membranes are partially detached (B and F, arrows), multilayered (C), or show an increased thickness and a loss of typical structure (E). Bar, 250 nm.
Blood Vessels of β1-null Teratomas Are Small and Irregular in Shape and Lack ES Cell-derived Endothelial Cells
Hematoxylin/Eosin staining, as well as immunostaining for the endothelial markers vWF and PECAM revealed a large number of circular vessels of various sizes in β1 +/+ teratomas (Fig. 7 A). In contrast, in β1-null tumors, vessels were smaller in size and of irregular shape (Fig. 7 B).
Figure 7.

Immunostaining of a normal (+/+) and a β1-null (−/−) teratoma for the expression of vWF and for β1 integrin expression. Teratomas were removed 21 d after inoculation and sectioned. Neighboring sections were stained for vWF and β1 integrin expression, respectively. Whereas normal teratomas have large vessels, β1-null teratomas have small vessels. Note that like in normal tumors vessels of β1-null tumors coexpress vWF and β1 integrin indicating that endothelial cells are host derived. Bar, 100 μm.
To test whether β1-null ES cells can contribute to the formation of blood vessels, endothelial cells were characterized in β1 +/+ and −/− teratomas. In β1 +/+ and +/− teratomas, all cells that expressed vWF or PECAM showed strong staining for β1 integrin (Fig. 7 C). Furthermore, many cells that expressed vWF were also lacZ-positive (Fig. 8, A and C) clearly indicating that ES cells can contribute to angiogenesis in developing teratomas. In β1-null tumors, however, cells that expressed vWF (Fig. 7 B) always expressed high levels of β1 integrin, indicating their host-derived origin (Fig. 7 D). These results were confirmed in serial sections of six β1-null teratomas, which were analyzed by lacZ and vWF stainings. In all β1-null teratomas analyzed so far, cells that were positive for vWF staining (Fig. 8 B) were negative for lacZ expression (Fig. 8 D).
Figure 8.


Localization of vWF and lacZ expression in normal (+/+) and β1-null (−/−) teratomas. Teratomas were removed 21 d after inoculation of ES cells and sectioned. Sections were stained overnight for lacZ expression and subsequently immunostained for vWF expression. In normal teratomas, vWF-positive areas (A, arrows) are found to be positive and negative for lacZ expression (C, corresponding arrows) indicating that endothelial cells are both host- and ES cell–derived. In contrast, in β1-null teratomas, all vWF-positive areas (B, arrows) are lacZ-negative (D, corresponding arrows) indicating that endothelial cells are solely host derived. Many other areas, however, that are vWF-negative (B, arrowhead) are clearly lacZ-positive (D, corresponding arrowhead) indicating that many other regions in β1-null teratomas are ES cell–derived. Bar, 100 μm.
During the analysis of tissue sections derived from β1-null teratomas it became evident that also the host cell–derived vasculature has abnormalities. Whereas blood vessels in normal teratomas have a smooth inner surface and are tightly anchored to the surrounding connective tissue (Fig. 9 A), blood vessels in β1-null tumors have an irregular inner surface and display large gaps between endothelial cells and the surrounding tissue (Fig. 9 B).
Figure 9.

Semithin sections of a normal and a β1-null teratoma stained with methylene blue and immunostained for vWF. Vessels (V) in normal teratomas (A) have a smooth inner surface and are tightly embedded within the surrounding tissue (arrows). Vessels of β1-null teratomas (B) have an irregular surface and have lost contacts to the surrounding tissue (arrows). Bar, 20 μm.
β1-Null ES Cells Can Differentiate into Endothelial Cells In Vitro but Formation of Vessels Is Delayed
ES cells were cultured for 5 d in suspension to form embryoid bodies, and then were plated on tissue culture dishes. Initially, PECAM-positive cells were aggregated in dense clusters (Fig. 10 A). These clusters then started to form thin tubes and branched. After 7 d on the culture dish about half of the PECAM-positive cells were part of blood vessels in embryoid bodies derived from normal ES cells. After 15 d of incubation, PECAM-positive cell clumps were no longer visible, the blood vessels were significantly grown in diameter, extensively branched (Fig. 10 B), and often contained many blood cells (Fig. 10 C).
Figure 10.
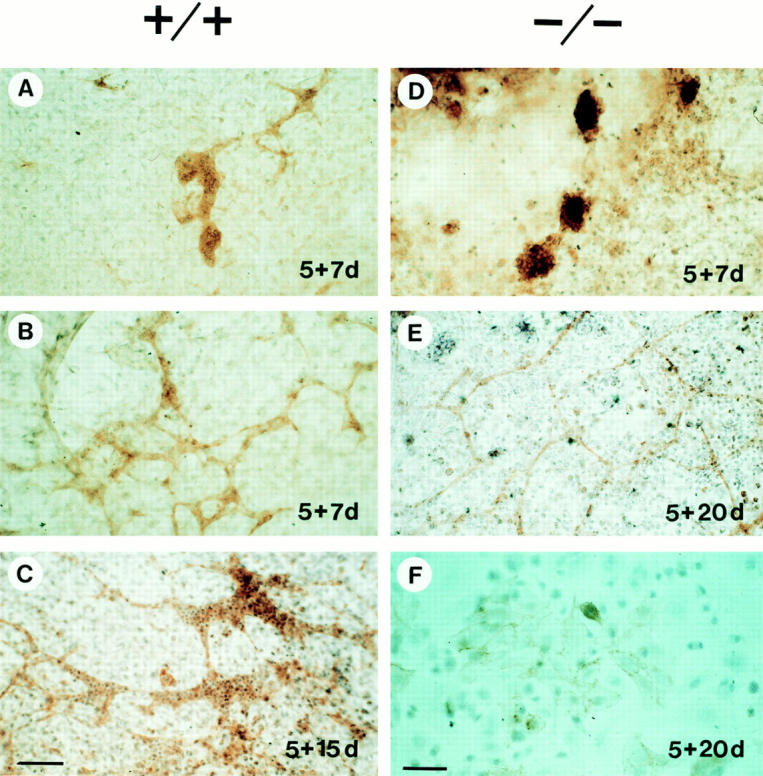
Normal (+/+) and β1-null (−/−) embryoid bodies immunostained for PECAM. ES cells were differentiated in hanging drops for 2 d, cultured in bacteriological dishes for another 3 d (which gives a total culture period of 5 d in suspension), and plated on gelatin-coated cover slips. After various periods on the cover slips (7, 15, 20 d) outgrowth were fixed and stained for PECAM. The culture periods are designated in each picture. Note the presence of blood cells in vessels of normal embryoid body outgrowths (C). Bars: (A–D) 80 μm; (F) 40 μm.
When β1-null embryoid bodies were analyzed 7 or 15 d after plating, only PECAM-positive cell clusters were found without any signs of vessel formation (Fig. 10 D). After 20 d in culture, a fine network of small vessels began to form in some areas of the embryoid bodies (Fig. 10 E). These vessels were small in diameter and never contained blood cells. In other areas, PECAM-positive cells were still organized in small cell patches, often with loose cell– cell contacts (Fig. 10 F). The mean value of the diameter of normal and β1-null vessels in seven embryoid bodies after a 3-wk culture period was 18.1 ± 3.0 μm and 6.8 ± 2.1 μm, respectively (Fig. 11).
Figure 11.

Diameter of vessel lumen in normal (+/+) and β1-null (−/−) embryoid bodies. •, −/− PECAM1; □, +/+ PECAM1. Morphometrical analysis of PECAM-positive vessels after 5 + 20 d in culture. Normal embryoid bodies have variously sized vessels with diam up to 70 μm. In β1-null embryoid bodies most vessels have a diam of 10 μm and only a few are larger.
To determine whether the absence of β1 integrin expression interferes with the differentiation of endothelial cells, we counted the number of PECAM-positive cells in normal as well as β1-null embryoid bodies. After a plating period of 28 d, 100 embryoid bodies (derived from normal and β1-null ES cells, respectively) were trypsinized; single cells were plated and cultured overnight, and then stained for PECAM expression. Out of 9,600 cells counted, 96 PECAM- positive cells were found in normal embryoid bodies and 9 cells in β1-null embryoid bodies. This 10-fold decrease of PECAM-positive cells in the β1-null embryoid bodies indicates a delayed or less efficient differentiation of β1-null endothelial cells.
Expression of β3 and αv Integrin Is not Significantly Altered on β1-null Endothelial Cells
Proliferating endothelial cells express large amounts of αvβ3 and αvβ5 integrins. To determine whether the expression of β3 or αv integrin is altered on β1-null endothelial cells, normal and β1-null embryoid bodies were double-immunostained for either PECAM and β3 integrin (Fig. 12, A–D), or PECAM and αv integrin (Fig. 12, E–H), respectively. Vessels in embryoid bodies derived from normal and β1-null ES cells express similar amounts of β3 (Fig. 12, B and D) and αv integrin (Fig. 12, F and H). These data indicate that the absence of β1 integrin is not associated with an altered expression of other endothelial cell integrins.
Figure 12.
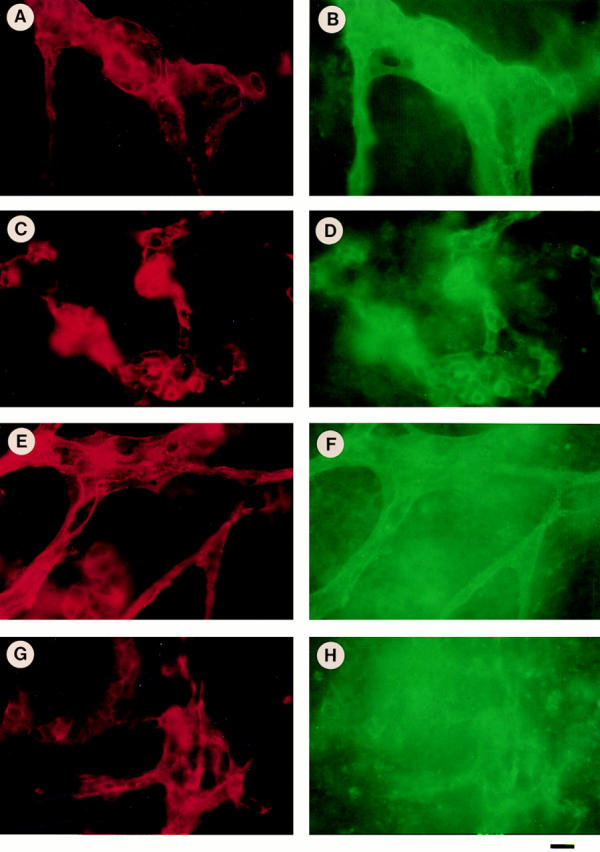
Double immunostaining of normal (+/+) and β1-null (−/−) embryoid bodies for β3 integrin and PECAM (A–D), and αv integrin and PECAM (E–H). Normal and mutant ES cells were differentiated for 20 d, fixed and stained for PECAM (A and C, red) and β3 (B and D, green), or PECAM (E and G, red) and αv (F and H, green), respectively. In normal embryoid bodies large vessels stain for PECAM (A and C) and β3 (B), or αv integrin (D). In β1-null embryoid bodies the vessel diameters are smaller but the staining for PECAM (E and G), β3 (D), and αv (H) is similar, like in normal bodies. Note the high background for αv staining, which is expressed on many PECAM-negative cells in the embryoid bodies. Bar, 15 μm.
VEGF Does Not Promote Angiogenesis in β1-null Embryoid Bodies
VEGF induces proliferation of endothelial cells and sprouting of vessels and therefore is a potent promoter of angiogenesis. To test whether VEGF can overcome abnormal angiogenesis in β1-null embryoid bodies, normal and β1 integrin-deficient ES cells were differentiated in the presence of 10 or 20 ng/ml VEGF, respectively. After 5 d in suspension culture and 12 d on gelatinized cover slips, embryoid bodies were treated with BrdU for 2 h and subsequently stained for PECAM expression and BrdU incorporation. Treatment of normal embryoid bodies with either 10 or 20 ng/ml VEGF significantly increased the number and the extent of blood vessel branching (Fig. 13, A and C; and Table I). In contrast, VEGF at both concentrations had no significant effect on blood vessel branching in β1-null embryoid bodies (Fig. 13, B and D; and Table I) indicating that VEGF cannot overcome the lack of β1 integrin function. VEGF treatment also increased the number of BrdU-positive cells in normal (Fig. 13, A and C, arrows) but not in β1-null embryoid bodies (Fig. 13, B and D, arrows).
Figure 13.
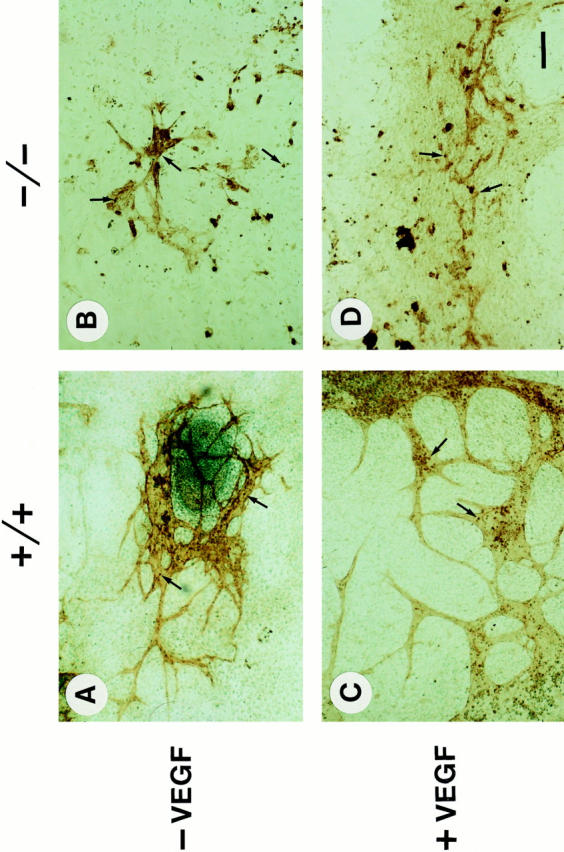
VEGF treatment of normal (+/+) and β1-null (−/−) embryoid bodies. Normal or β1-null embryoid bodies were treated either without (A and B) or with (C and D) 10 ng/ml VEGF during a period of 5 d in suspension and 12 d on a gelatinized glass cover slip. 2 h before fixation, the cultures were treated with BrdU. Sections were first stained for BrdU incorporation (arrows), then for PECAM-1, and finally counterstained with methyl green. Note that VEGF treatment increases the branching of blood vessels (C) and the proliferation rate of PECAM-positive cells in normal (C, arrows) but not in β1-null embryoid bodies (D, arrows). Bar, 100 μm.
Table I.
Quantification of Branches of Blood Vessels in Normal (+/+) and β1-null (−/−) Embryoid Bodies Treated Without or With VEGF
| Without VEGF | 10 ng/ml VEGF | 20 ng/ml VEGF | ||||
|---|---|---|---|---|---|---|
| (1/1) | 273 ± 103 (n = 9) | 363 ± 103 (n = 8) | 372 ± 86 (n = 7) | |||
| (2/2) | 41 ± 30 (n = 10) | 32 ± 20 (n = 8) | 76 ± 85 (n = 7) |
Embryoid bodies were cultured 5 d in suspension and 12 d on gelatinized glass cover slips in the absence or presence of 10 or 20 ng/ml VEGF, respectively. After the end of the culture period, embryoid bodies were fixed and stained for PECAM expression. Data show all vessel branches per embryoid body and are expressed in mean + SD. n is number of embryoid bodies analyzed.
DISCUSSION
In the present study we show that the growth of ES cell– derived teratomas is dependent on the expression of β1 integrin. Whereas subcutaneous injection of normal ES cells under the skin of the back of syngeneic mice gave rise to large teratomas, β1-null ES cells either produced no, or only very small tumors. This reduced growth of β1-null teratomas suggests a growth-supporting role of β1 integrin.
These results were unexpected since many tumors lack α5β1 integrin function, a phenomenon that has been linked with anchorage-independent growth and high tumorigenicity (Hirst et al., 1986; Plantefaber and Hynes, 1990; Varner et al., 1995). Conversely, overexpression of α5β1 integrin in Chinese hamster ovary cells is accompanied by a loss of tumorigenicity and reduced proliferation in vitro (Giancotti and Ruoslahti, 1990). A similar growth-suppressing role was recently demonstrated for α2β1 integrin, which is frequently absent in breast cancer cells. Reexpression of α2β1 integrin in such cells also leads to a diminution of the malignant phenotype (Zutter et al., 1995).
Several explanations could account for this discrepancy. First, ES cells are normal embryonal cells that differentiate into many somatic tissues and thus give rise to a tumor called teratoma (Damjanov and Solter, 1974; Damjanov, 1978). Since these cells are not transformed, they may still be dependent on anchorage to the extracellular matrix for growth (Dike and Farmer, 1988) and survival (Meredith et al., 1993; Boudreau et al., 1995). Integrins execute cell– matrix interactions and lack of anchorage in β1-null ES cells could therefore be an explanation for the reduced tumor growth. To test this hypothesis we analyzed cell proliferation by injecting BrdU into teratoma-bearing mice and stained for apoptosis. Extensive analysis of normal and β1-null teratomas revealed that cell proliferation and apoptosis are not significantly affected in β1-null tumors. It has to be noted, however, that the extents of proliferation and apoptosis were very different between different sections of the same tumor, as well as between individual tumors. This heterogeneity is due to the ability of ES cells to differentiate into a wide range of somatic tissues with different apoptosis and proliferation rates. For this reason we obtained, in normal as well as β1-null teratomas, enormous variations in the numbers of proliferating and apoptotic cells. Despite this variation, there were more areas with higher numbers of proliferating and lower numbers of apoptotic cells in normal teratomas when compared with β1-null teratomas. Such alterations would be in agreement with reports that have linked cell survival with integrin function (for review see Ruoslahti and Reed, 1994). The binding of α5β1 integrin to fibronectin is inducing bcl-2 expression, which is one of the best studied cell survival factors (Zhang et al., 1995). Other studies showed that disrupting the binding to basement membranes can induce apoptosis (Boudreau et al., 1995; Coucouvanis and Martin, 1995). Biochemical studies of β1-null teratomas (Sasaki, T., E. Forsberg, W. Bloch, K. Addicks, R. Fässler, and R. Timpl, submitted for publication), together with our electronmicroscopical analysis indicate that the integrity of basement membranes is severely affected by the absence of β1 integrin. Whereas epithelial cells of normal teratomas are tightly attached to a basement membrane, β1-null cells have either partly or completely lost this tight attachment leading to the dislocation of basement membranes into the interstitium. This absence of normal interaction between cells and basement membranes could not only explain the increased cell death in teratomas, but also the altered differentiation of keratinocytes (Bagutti et al., 1996) and cardiac muscle cells (Fässler et al., 1996). Similar defects in basement membrane morphology have been reported in α3-null mice, which have disorganized and fragmented glomerular basement membranes, and podocytes that completely lack foot processes (Kreidberg et al., 1996). In addition, these mice develop skin blisters that are due to an inefficient maintenance of the epidermal basement membrane (DiPersio et al., 1997).
A second explanation for the reduced tumor growth could be the altered composition or distribution of ECM proteins in β1-null teratomas. The ECM has an important role as reservoir for growth factors, proteases, and protease inhibitors (Alexander and Werb, 1991; Nathan and Sporn, 1991). Basement membranes, for example, can store and release many growth factors. The various abnormalities of basement membranes as seen in β1-null teratomas and β1-null embryoid bodies may contribute to the diminished growth and to defects in differentiation (Fässler et al., 1996). In addition, the disturbed matrix deposition can alter mechanochemical properties, which may reduce the growth rate of β1-null teratomas (Ingber, 1991). Indeed, immunostainings for fibronectin and collagens revealed an altered distribution in β1-null teratomas. In normal teratomas both matrix proteins concentrate in streaklike structures and often colocalize with high β1 integrin staining. In contrast, both proteins are diffusely deposited in β1-null teratomas and never concentrate along β1 integrin-positive, host-derived cells. This striking finding was also supported by routine histological analysis. Tissue specimens of β1-null teratomas contained little ECM, with many mutant and only a few normal, host-derived (lacZ negative) cells.
A third explanation for the formation of small teratomas could be abnormal angiogenesis in the absence of β1 integrin. In contrast to tumors derived from somatic tissue, the vasculature of teratomas is derived from the host and from the ES cells (Risau et al., 1988; Vittet et al., 1996). Immunostaining demonstrated host-derived endothelial cells expressing β1 integrin in normal as well as β1-null tumors. Whereas lacZ staining indicated ES cell–derived (lacZ positive) endothelial cells in β1 integrin–heterozygous as well as mock-transfected tumors, no ES cell–derived endothelial cells were detected in β1-null teratomas. This complete absence of β1-null endothelial cells is also seen in the highly vascularized fetal and adult liver of β1-null chimeric mice (Fässler and Meyer, 1995; Hirsch et al., 1996). These data suggest, that β1-deficient ES cells either do not differentiate in vivo into endothelial cells or that they differentiate into endothelial cells but cannot organize into blood vessels. A detailed inspection of the vasculature revealed furthermore that blood vessels were significantly smaller in β1-null teratomas as compared to normal teratomas, which may result from the small tumor size. In addition, however, we found that the shape of smaller vessels is also different in β1-null tumors: the inner surface is irregular and the vessels are loosely embedded into the surrounding tissue. Apparently, even endothelial cells expressing β1 integrin are not able to make normal blood vessels in β1-null teratoma, indicating lack of an essential interaction between endothelial cells and the surrounding cells or the extracellular matrix.
To test whether the β1-null ES cells have a cell autonomous impairment of differentiation into endothelial cells, we differentiated ES cells into embryoid bodies in vitro and assessed for the presence of endothelial cells (Fässler et al., 1996). To our surprise, we found that differentiation of endothelial cells occurs in normal and β1-null embryoid bodies. After 12 d (5 d in suspension culture, and 7 d on plastic surface) normal endothelial cells start to sprout and to form a large network of vessels of various diameters that are also often filled with blood cells. In contrast, β1-null endothelial cells initially form compact cell nests with tight cell–cell contacts. Only after a culture period of 3 wk, β1-null endothelial cells start to migrate and to form small vessels. The ability of β1-null endothelial cells to differentiate may also occur in vivo. This hypothesis is supported by the presence of β1-null hematopoietic stem cells in chimeric mice (Hirsch et al., 1996) that, together with endothelial cells, develop from a common precursor cell called hemangioblast. Currently, we have no experimental explanation for the complete lack of β1-null endothelial cells both in teratomas and in livers of chimeric mice (Fässler and Meyer, 1995), despite the fact that they are readily formed in vitro. It is possible that the in vitro environment favors the survival of these cells, which is not the case in vivo.
An important role for angiogenesis has been reported for αvβ3 and αvβ5 integrins (Varner and Cheresh, 1996). Immunostaining of β1-null embryoid bodies with antibodies detecting αv or β3 integrin clearly revealed that both integrin subunits are not downregulated. Therefore, we can exclude that the absence of β1 integrin expression influences endothelial cell function by altering the expression of other integrins. Growth factors and angiogenic signals can upregulate αvβ3 expression on endothelial cells and induce blood vessel branching (Brooks et al., 1994a ; Friedlander et al., 1995). Moreover, several growth factors were shown to exert their angiogenic signals in concert with endothelial integrins. For example, tumor necrosis factor-α and basic FGF depend on αvβ3 integrin, and VEGF and TGF-α depend on αvβ5 integrin (Friedlander et al., 1995). VEGF is the most potent stimulator of neovascularization by regulating proliferation of endothelial cells and branching of blood vessels. Testing whether VEGF can override the defective branching of β1-null endothelial cells, we found that VEGF had no effect on β1-null endothelial cells: neither the proliferation rate nor the branching could be enhanced in β1-null embryoid bodies. Endothelial cells in normal embryoid bodies, however, responded to VEGF by an increased proliferation rate and by an extensive formation of new vessel branches. These data demonstrate for the first time that β1 integrins play an important role during angiogenesis and moreover, that VEGF signaling depends on the presence of β1 integrin function. This finding is unexpected since earlier reports showed that β1 integrin function is not essential for tumor-induced angiogenesis (Brooks et al., 1994b ). Brooks et al. (1994b) treated chicken embryos that were exposed to various tumors with anti-β1 integrin antibodies and were unable to influence tumor growth or angiogenesis. One explanation for this discrepancy may be an incomplete inhibition of β1 integrin function with a single injection of antibodies. An alternative explanation could be the ubiquitous expression of β1 integrin. Antibodies against β1 integrin find their antigen on almost all cells and may be eliminated before endothelial cells proliferate and form new blood vessels.
Acknowledgments
We thank M. Grüne and S. Benkert for technical assistance and E. Dejana for antibody gifts.
This work was supported by the Deutsche Forschungsgemeinschaft (Fa 296/1-2) and the Hermann and Lilly Schilling Stiftung.
Abbreviations used in this paper
- BrdU
bromodeoxyuridine
- ECM
extracellular matrix
- ES
embryonic stem
- PECAM
platelet/endothelial cell adhesion molecule
- VEGF
vascular endothelial growth factor
- vWF
von Willebrand Factor
Footnotes
W. Bloch and E. Forsberg have contributed equally to this work.
Address all correspondence to Reinhard Fässler, Max Planck Institut für Biochemie, am Klopferspitz 18 A, 82 152 Martinsried, Germany. Tel.: (49) 89-8578-2215. FAX: (49) 89-8578-2422. e-mail: faessler@biochem.mpg.de
REFERENCES
- Alexander, C.M., and Z. Werb. 1991. Degradation of extracellular matrix. In Cell Biology of Extracellular Matrix. E. Hay, editor. Plenum Press, New York. pp. 255–294.
- Bagutti C, Wobus AM, Fässler R, Watt FM. Differentiation of embryonal stem cells into keratinocytes: comparison of wild-type and β1 integrin-deficient cells. Dev Biol. 1996;179:184–196. doi: 10.1006/dbio.1996.0250. [DOI] [PubMed] [Google Scholar]
- Boudreau N, Sympson CJ, Werb Z, Bissel MJ. Suppression of ICE and apoptosis in mammary epithelial cells by extracellular matrix. Science (Wash DC) 1995;267:891–893. doi: 10.1126/science.7531366. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooks PC, Clark RAF, Cheresch DA. Requirement of vascular integrin αvβ3 for angiogenesis. Science (Wash DC) 1994a;264:569–571. doi: 10.1126/science.7512751. [DOI] [PubMed] [Google Scholar]
- Brooks PC, Montgomery AMP, Rosenfeld M, Reisfeld RA, Hu T, Klier G, Cheresh DA. Integrin αvβ3antagonists promote tumor regression by inducing apoptosis of angiogenic blood vessels. Cell. 1994b;79:1157–1164. doi: 10.1016/0092-8674(94)90007-8. [DOI] [PubMed] [Google Scholar]
- Coucouvanis E, Martin GR. Signals for death and survival: a two-step mechanism for cavitation in the vertebrate embryo. Cell. 1995;83:279–287. doi: 10.1016/0092-8674(95)90169-8. [DOI] [PubMed] [Google Scholar]
- Damjanov I. Teratoma and teratocarcinoma in experimental animals. Natl Cancer Inst Monogr. 1978;49:305–306. [PubMed] [Google Scholar]
- Damjanov I, Solter D. Experimental teratoma. Curr Top Pathol. 1974;59:69–130. doi: 10.1007/978-3-642-65857-0_2. [DOI] [PubMed] [Google Scholar]
- Dike LE, Farmer SR. Cell adhesion induces expression of growth-associated genes in suspension-arrested fibroblasts. Proc Natl Acad Sci USA. 1988;85:6792–6796. doi: 10.1073/pnas.85.18.6792. [DOI] [PMC free article] [PubMed] [Google Scholar]
- DiPersio CM, Hodivila-Dilke KM, Jaenisch R, Kreidberg JA, Hynes RO. α3β1 integrin is required for normal development of the epidermal basement membrane. J Cell Biol. 1997;137:729–742. doi: 10.1083/jcb.137.3.729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Doetschman TC, Eistetter H, Katz M, Schmidt W, Kemler R. The in vitro development of blastocyst derived embryonic stem cell lines: formation of visceral yolk sac, blood islands and myocardium. J Embryol Exp Morph. 1985;87:27–45. [PubMed] [Google Scholar]
- Dziadek M, Clements R, Mitrangas K, Reiter H, Fowler K. Analysis of degradation of the basement membrane protein nidogen, using a specific monoclonal antibody. Eur J Biochem. 1988;172:219–225. doi: 10.1111/j.1432-1033.1988.tb13876.x. [DOI] [PubMed] [Google Scholar]
- Fässler R, Meyer M. Consequences of lack of β1 integrin gene expression in mice. Genes Dev. 1995;9:1896–1908. doi: 10.1101/gad.9.15.1896. [DOI] [PubMed] [Google Scholar]
- Fässler R, Pfaff M, Murphy J, Noegel A, Johansson S, Timpl R, Albrecht R. Lack of β1 integrin gene in embryonic stem cells affects morphology, adhesion, and migration but not integration into the inner cell mass of blastocysts. J Cell Biol. 1995;128:979–988. doi: 10.1083/jcb.128.5.979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fässler R, Rohwedel J, Maltsev V, Bloch W, Lentini S, Guan K, Gullberg D, Hescheler J, Addicks K, Wobus AM. Differentiation and integrity of cardiac muscle cells are impaired in the absence of β1 integrin. J Cell Sci. 1996;109:2989–2999. doi: 10.1242/jcs.109.13.2989. [DOI] [PubMed] [Google Scholar]
- Folkman J. Fighting cancer by attacking its blood supply. Sci Am. 1996;275:150–154. doi: 10.1038/scientificamerican0996-150. [DOI] [PubMed] [Google Scholar]
- Folkman J, Moscona A. Role of cell shape in growth control. Nature (Lond) 1978;273:345–349. doi: 10.1038/273345a0. [DOI] [PubMed] [Google Scholar]
- Friedlander M, Brooks PC, Shaffer RW, Kincaid CM, Varner JA, Cheresh DA. Definition of two angiogenetic pathways by distinct αv integrins. Science (Wash DC) 1995;270:1500–1502. doi: 10.1126/science.270.5241.1500. [DOI] [PubMed] [Google Scholar]
- Giancotti FG, Ruoslahti E. Elevated levels of the α5β1 fibronectin receptor suppress the transformed phenotype of chinese hamster ovary cell variants. Cell. 1990;60:849–859. doi: 10.1016/0092-8674(90)90098-y. [DOI] [PubMed] [Google Scholar]
- Guadagno TM, Ohtsubo M, Roberts JM, Assoian RK. A link between cyclin A expression and adhesion-dependent cell cycle progression. Science (Wash DC) 1993;262:1572–1575. doi: 10.1126/science.8248807. [DOI] [PubMed] [Google Scholar]
- Guan JL, Shalloway D. Regulation of focal adhesion associated protein tyrosine kinase by both cellular adhesion and oncogenic transformation. Nature (Lond) 1992;358:690–692. doi: 10.1038/358690a0. [DOI] [PubMed] [Google Scholar]
- Haas TA, Plow EF. Integrin-ligand interactions: a year in review. Curr Opinion Cell Biol. 1994;6:656–662. doi: 10.1016/0955-0674(94)90091-4. [DOI] [PubMed] [Google Scholar]
- Hannigan GE, Leung-Hagesteijn C, Fitz-Gibbon L, Coppolino MG, Radeva G, Filmus J, Bell JC, Dedhar S. Regulation of cell adhesion and anchorage-dependent growth by a new β1 integrin-linked protein kinase. Nature (Lond) 1996;379:91–96. doi: 10.1038/379091a0. [DOI] [PubMed] [Google Scholar]
- Hirsch E, Iglesias A, Potocnik AJ, Hartmann U, Fässler R. Impaired migration but not differentiation of haematopoietic stem cells in the absence of β1 integrins. Nature (Lond) 1996;380:171–175. doi: 10.1038/380171a0. [DOI] [PubMed] [Google Scholar]
- Hirst R, Horwitz A, Buck C, Rohrschneider L. Phosphorylation of the fibronectin receptor y oncogenes that encode tyrosine kinases. Proc Natl Acad Sci USA. 1986;83:6470–6474. doi: 10.1073/pnas.83.17.6470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hynes RO. Integrins: versatility, modulation, and signaling in cell adhesion. Cell. 1992;69:11–25. doi: 10.1016/0092-8674(92)90115-s. [DOI] [PubMed] [Google Scholar]
- Ingber D. Integrins as mechanochemical transducers. Curr Opin Cell Biol. 1991;3:841–848. doi: 10.1016/0955-0674(91)90058-7. [DOI] [PubMed] [Google Scholar]
- Johansson S, Höök M. Substrate adhesion of rat hepatocytes: on the mechanism of attachment to fibronectin. J Cell Biol. 1984;98:810–817. doi: 10.1083/jcb.98.3.810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kreidberg JA, Donovan MJ, Goldstein SL, Renke H, Shepherd K, Jones RC, Jaenisch R. α3β1 integrin has a crucial role in kidney and lung organogenesis. Development (Camb) 1996;122:3537–3547. doi: 10.1242/dev.122.11.3537. [DOI] [PubMed] [Google Scholar]
- McNamee HP, Ingber DE, Schwartz MA. Adhesion to fibronectin stimulates inositol lipid synthesis and enhances PDGF-induced inositol lipid breakdown. J Cell Biol. 1993;121:673–678. doi: 10.1083/jcb.121.3.673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meredith JE, Fazeli B, Schwartz MA. The extracellular matrix as a cell survival factor. Mol Biol Cell. 1993;4:953–961. doi: 10.1091/mbc.4.9.953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morino N, Mimura T, Hamasaki K, Tobe K, Ueki K, Kikuchi K, Takehara K, Kadowaki T, Yazaki Y, Nojima Y. Matrix/integrin interaction activates the mitogen-activated protein kinase, p44erk-1 and p42erk-2. J Biol Chem. 1995;270:269–273. doi: 10.1074/jbc.270.1.269. [DOI] [PubMed] [Google Scholar]
- Nathan C, Sporn M. Cytokines in context. J Cell Biol. 1991;113:981–986. doi: 10.1083/jcb.113.5.981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plantefaber LC, Hynes RO. Changes in integrin receptors on oncogenically transformed cells. Cell. 1990;56:281–290. doi: 10.1016/0092-8674(89)90902-1. [DOI] [PubMed] [Google Scholar]
- Risau W, Sariola H, Zerwes HG, Sasse J, Ekblom P, Kemler R, Doetschman T. Vasculogenesis and angiogenesis in embryonic stem cell- derived embryoid bodies. Development (Camb) 1988;102:471–478. doi: 10.1242/dev.102.3.471. [DOI] [PubMed] [Google Scholar]
- Ruoslanti E, Reed JC. Anchorage dependence, integrins and apoptosis. Cell. 1994;77:477–478. doi: 10.1016/0092-8674(94)90209-7. [DOI] [PubMed] [Google Scholar]
- Schaller MD, Otey CA, Hildebrand JD, Parsons JT. Focal adhesion kinase and paxillin bind to peptides mimicking β integrin cytoplasmic domains. J Cell Biol. 1995;130:1181–1187. doi: 10.1083/jcb.130.5.1181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz MA. Signaling by integrins: implications for tumorigenesis. Cancer Res. 1993;53:1503–1506. [PubMed] [Google Scholar]
- Schwartz MA, Both G, Lechene C. Effect of cell spreading on cytoplasmic pH in normal and transformed fibroblasts. Proc Natl Acad Sci USA. 1989;86:4525–4529. doi: 10.1073/pnas.86.12.4525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schwartz MA, Cragoe EJ, Jr, Lechene CP. pH regulation in spread cells and round cells. J Biol Chem. 1990;265:1327–1332. [PubMed] [Google Scholar]
- Varner JA, Emerson DA, Juliano RL. Integrin α5β1 expression negatively regulates cell growth: reversal by attachment to fibronectin. Mol Biol Cell. 1995;6:725–740. doi: 10.1091/mbc.6.6.725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Varner JA, Cheresh DA. Integrins and cancer. Curr Opin Cell Biol. 1996;8:724–730. doi: 10.1016/s0955-0674(96)80115-3. [DOI] [PubMed] [Google Scholar]
- Vecchi A, Garlanda C, Lampugnani MG, Resnati M, Matteucci C, Stoppacciaro A, Schnurch H, Risau W, Ruco L, Mantovani A, et al. Monoclonal antibodies specific for endothelial cells of mouse blood vessels. Their application in the identification of adult and embryonic endothelium. Eur J Cell Biol. 1994;63:247–254. [PubMed] [Google Scholar]
- Vittet D, Prandini MH, Berthier R, Schweitzer A, Martin-Sisteron H, Uzan G, Dejana E. Embryonic stem cells differentiate in vitro to endothelial cells through successive maturation steps. Blood. 1996;88:3424–3431. [PubMed] [Google Scholar]
- Vuori K, Ruoslahti E. Activation of protein kinase C precedes α5β1 integrin-mediated cell spreading on fibronectin. J Biol Chem. 1993;268:21459–21462. [PubMed] [Google Scholar]
- Yebra M, Filardo EJ, Bayna EM, Kawahara E, Becker JC, Cheresh DA. Induction of carcinoma cell migration on vitronectin by NF-κ B-dependent gene expression. Mol Biol Cell. 1995;6:841–850. doi: 10.1091/mbc.6.7.841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang Z, Vuori K, Reed JC, Ruoslahti E. The α5β1 integrin supports survival of cells on fibronectin and up-regulates Bcl-2 expression. Proc Natl Acad Sci USA. 1995;92:6161–6165. doi: 10.1073/pnas.92.13.6161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zutter MM, Santoro SA, Staatz WD, Tsung YL. Re-expression of the α2β1 integrin abrogates the malignant phenotype of breast carcinoma cells. Proc Natl Acad Sci USA. 1995;92:7411–7415. doi: 10.1073/pnas.92.16.7411. [DOI] [PMC free article] [PubMed] [Google Scholar]