

Abstract
In vitro delivery of the diphtheria toxin catalytic (C) domain from the lumen of purified early endosomes to the external milieu requires the addition of both ATP and a cytosolic translocation factor (CTF) complex. Using the translocation of C-domain ADP-ribosyltransferase activity across the endosomal membrane as an assay, the CTF complex activity was 650–800-fold purified from human T cell and yeast extracts, respectively. The chaperonin heat shock protein (Hsp) 90 and thioredoxin reductase were identified by mass spectrometry sequencing in CTF complexes purified from both human T cell and yeast. Further analysis of the role played by these two proteins with specific inhibitors, both in the in vitro translocation assay and in intact cell toxicity assays, has demonstrated their essential role in the productive delivery of the C-domain from the lumen of early endosomes to the external milieu. These results confirm and extend earlier observations of diphtheria toxin C-domain unfolding and refolding that must occur before and after vesicle membrane translocation. In addition, results presented here demonstrate that thioredoxin reductase activity plays an essential role in the cytosolic release of the C-domain. Because analogous CTF complexes have been partially purified from mammalian and yeast cell extracts, results presented here suggest a common and fundamental mechanism for C-domain translocation across early endosomal membranes.
Keywords: endosome; Hsp 90; thioredoxin reductase; geldanamycin; radicicol
Introduction
Diphtheria toxin (DT;* 58 kD) is a typical single-chain AB toxin composed of three functional domains: the amino-terminal catalytic (C) domain corresponds to fragment A (21 kD), and the transmembrane (T) and carboxy-terminal receptor-binding domains comprise fragment B (37 kD) of the toxin (Choe et al., 1992). A disulfide bond between Cys186 and Cys201 subtends a protease-sensitive loop and connects fragment A with fragment B. Furin-mediated cleavage within this loop and retention of the disulfide bond have been shown to be prerequisites for intoxication of eukaryotic cells (Ariansen et al., 1993; Tsuneoka et al., 1993). Substitution of the native receptor domain with human interleukin-2 (IL-2) has resulted in the formation of a fusion protein toxin, DAB389IL-2, whose cytotoxic action is specifically targeted only to cells expressing the high affinity IL-2 receptors (Bacha et al., 1988; Waters et al., 1990; vanderSpek 2002>Ratts and vanderSpek, 2002).
Because DAB389IL-2 binds with greater affinity to its receptor compared with native DT, this fusion protein toxin has proven to be an effective and novel probe for studying internalization of the C-domain by target cells (Williams et al., 1990). Although much is known about the mechanisms of receptor-binding and receptor-mediated endocytosis of native DT and the DT-related fusion proteins, little is known about the precise molecular mechanisms of C-domain translocation across the endosomal membrane and its release into the cytosol.
Unfolding of the DT C-domain was first postulated as a prerequisite for translocation by Kagan et al. (1981) and Donovan et al. (1981). The necessity for complete denaturation of the DT C-domain before translocation was then indirectly demonstrated by Wiedlocha et al. (1992) and by Falnes et al. (1994). At present, there are two conflicting hypotheses for translocation of denatured DT C-domain across the early endosomal membrane. Studies using artificial lipid bilayers suggest that the DT T-domain itself exhibits chaperonin-like properties and is solely sufficient to promote C-domain delivery across the bilayer (Oh et al., 1999; Ren et al., 1999). In contrast, studies using partially purified early endosomes that were preloaded with toxin suggest that C-domain translocation across the vesicle membrane is dependent on ATP and the presence of cytosolic components which include β-COP (Lemichez et al., 1997). Because protease digestion patterns of DT inserted into planar lipid bilayers differ from those of DT inserted into the plasma membrane (Moskaug et al., 1991; Cabiaux et al., 1994), it seems likely that interaction(s) between the toxin and proteins associated with the endosomal membrane (e.g., receptor) influence the orientation and/or stoichiometry of insertion of the T-domain and translocation of the C-domain. In addition, Ren et al. (1999) and Hammond et al. (2002) have shown that although the DT T-domain has chaperonin-like properties, it has a significantly greater affinity for other molten globule-like polypeptides compared with its own C-domain.
To further define the requirements of C-domain translocation across the endosomal membrane, we have used an in vitro C-domain translocation assay essentially as described by Lemichez et al. (1997). This assay uses purified early endosomes that have been preloaded with DAB389IL-2 and monitors the translocation of ADP-ribosyltransferase activity from the endosomal lumen to the external milieu. We have used translocated ADP-ribosyltransferase activity to monitor the purification of cytosolic components that are required for this process. In the present paper, we demonstrate by mass spectrometry (MS) sequence identification and the effect of specific inhibitors that the chaperonin heat shock protein (Hsp) 90 and thioredoxin reductase (TrR-1) are components of a cytosolic translocation factor (CTF) complex that is essential for the translocation and release of C-domain from early endosomes. Furthermore, the identification of CTF complex homologues in partially purified yeast extracts suggests that DT C-domain translocation may proceed by a fundamental mechanism of entry.
Results
Partial purification of the DT C-domain CTF complex
The requirements for DT C-domain translocation across the early endosomal membrane and release into the external milieu were monitored using an in vitro translocation assay modified from Umata et al. (1990) and Lemichez et al. (1997). The early endosomal compartment of HUT102 6TG cells was preloaded with DAB389IL-2 in the presence of bafilomycin A1. Early endosomes were purified by sucrose density gradient centrifugation, and then incubated in the presence of ATP and cytosolic extracts from either HUT102 6TG cells or yeast. After incubation at 37°C, translocation of the C-domain across the endosomal membrane and release into the external medium was monitored by ADP-ribosyltransferase activity of both the pellet and supernatant fluid fractions after ultracentrifugation. The [32P]ADP ribosylation of elongation factor 2 (EF-2) was measured by autoradiography of SDS-PAGE of reaction mixtures (Chung and Collier, 1977). The limit of sensitivity of this assay is in the range of 10−14–10−16 M C-domain, a level well below that of detection by immunoblot (unpublished data).
As shown in Fig. 1 A, on dilution of bafilomycin A1 and the addition of both ATP and cytosolic extracts to the reaction mixture, the C-domain is translocated across the endosomal membrane and released into the external medium. Moreover, preboiling the cytosolic extracts before their addition to the reaction mixture abolishes C-domain translocation. These results suggest that the C-domain translocation across the membrane of early endosomes requires cytosolic protein(s). The time course of C-domain translocation was examined using the epitope-labeled fusion protein toxin DAB189(VSV-G)B389IL-2. The cytotoxic potency of the epitope-tagged fusion toxin is almost identical to that of DAB389IL-2 (IC50 = 3 × 10−11 M vs. 4 × 10−12 M). As shown in Fig. 1 B, the ADP-ribosyltransferase activity as measured by densitometry of the combined 32P-labeled EF-2 from each paired pellet and supernatant fluid fraction is plotted as percentage of ADP-ribosyltransferase activity in the supernatant fluid. As can be seen, translocation of the C-domain is linear for up to 45 min, at which time ∼80% of the total activity is found in the supernatant fluid fraction. As reported by Lemichez et al. (1997), whereas ADP-ribosyltransferase activity was translocated to the external medium, cointernalized HRP activity was found to remain in the pellet fraction throughout the incubation period (unpublished data). These results strongly suggest that C-domain translocation is specific, and not the result of spontaneous endosomal lysis during the incubation period. Finally, in the presence of added ATP and cytosolic extracts, the translocation of the C-domain is dependent on membrane fluidity and does not occur at temperatures below 15°C.
Figure 1.

Partial purification of cytosolic proteins required to mediate DT C-domain translocation from the lumen of early endosomes in vitro. (A) Early endosomes from human T cells (HUT102/6TG), preloaded with DAB389IL-2, were incubated for 30 min at 37°C with 2 mM ATP (A) and/or 4 μg HUT102/6TG crude cytosol (C). C* denotes heat inactivation of cytosol before incubation with endosomes. Endosomes were pelleted, and the supernatant fraction (S) was assayed for DT C-domain ADP-ribosyltransferase activity by measuring the incorporation of [32P]NAD+ into AD[32P]ribosyl-EF-2 after 7% SDS-PAGE and autoradiography. (B) Time course for translocation of ADP-ribosyltransferase activity from DA189(VSV-G)B389IL-2 across the early endosomal membrane. Translocations were performed as described above, except the reactions were incubated at different temperatures (0, 15, and 37°C) for 15, 30, and 45 min. Both the supernatant and the pellet fractions were assayed for ADP-ribosyltransferase activity, and the autoradiographic signals were measured by densitometry. The sum of densitometry units from each pair of supernatant fluid and pellet fractions is plotted as the percentage of activity in the supernatant fractions at that time point. (n = 3; error bar denotes SD).
To rule out the possibility that the crude T cell and yeast extracts contained an allosteric regulator(s) of vesicular ATPase activity rather than protein(s) that are required for C-domain translocation, early endosomes were charged with a 70-kD dextran conjugated with the pH-sensitive fluorescent dye, SNARF-1. As shown in Fig. 2 A, compared with pH 7.5, the fluorescence emission of 1 ng/ml SNARF-1 is decreased approximately fourfold at pH 4.5. As measured by the quenching of fluorescence emission of SNARF-1, in vitro acidification of the early endosomal lumen occurs on dilution of bafilomycin A1 and requires the addition of 2 mM ATP to the reaction mixture (Fig. 2 B). Moreover, the time course for the acidification of early endosomes in vitro is virtually identical after the addition of either 2 mM ATP or 2 mM ATP plus partially purified T cell CTF complex.
Figure 2.

The in vitro acidification of early endosomes requires ATP and does not require any cytosolic protein factors. (A) Fluorescence emission of 1 ng/ml SNARF-1 70 kD dextran conjugate standards at pH 7.5 and 4.5 was measured at an excitation wavelength of 534 nM and an emission wavelength of 645 nM. (B) Purified early endosomes preloaded with the pH-sensitive SNARF-1 70 kD dextran conjugates were incubated in translocation assay buffer for 20 min at 37°C with 2 mM ATP and/or 0.1 μg/μL of Mono Q-purified cytosol. In each instance, assays were performed in triplicate and fluorescence was monitored using a fluorescence detector (model 650S; PerkinElmer). Error bars denote SD.
Partial purification of human T cell and yeast cytosolic factors required for the in vitro translocation of ADP-ribosyltransferase activity across the membrane of early endosomes
Because C-domain translocation across the endosomal vesicle membrane requires the addition of cytosolic components to the reaction mixture, we used translocation of ADP-ribosyltransferase activity to monitor the partial purification of the active component(s) from both human T cell (HUT102/6TG) and yeast (NLY22−) extracts. After DEAE anion exchange chromatography, translocation-active fractions (150 mM–190 mM NaCl) were pooled and applied to a Sephacryl® 200 sizing column. The translocation-active fractions (250–100 kD) were pooled and further fractionated by Mono Q HPLC under conditions free of reducing agents. The translocation-active fraction was found to elute from the Mono Q column at 27.3 mS. As shown in Fig. 3 A, after fractionation on Mono Q, CTF complex activity from human T cell and yeast cell extracts was increased by 650-fold and 800-fold, respectively. Further analysis of the Mono Q-pooled fractions by SDS-PAGE and colloidal Coomassie staining revealed multiple protein bands ranging in apparent molecular mass from ∼12–100 kD (Fig. 3 B).
Figure 3.
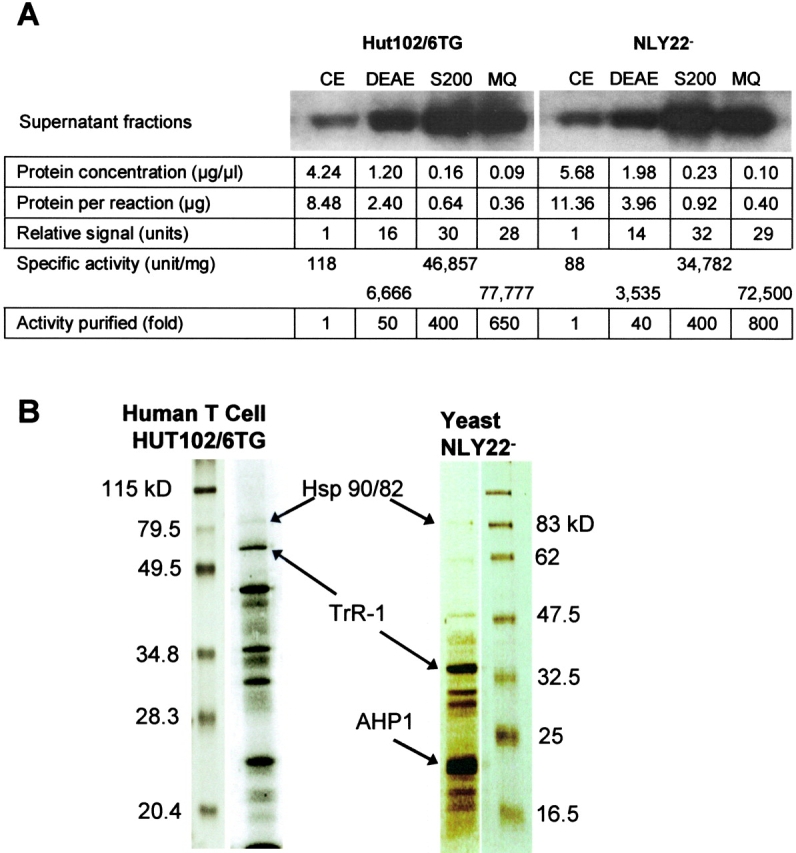
The partial purification of CTFs results in the increase of translocation in vitro specific activity. (A) Translocation in vitro specific activity of CTFs increases after each stage of purification. Reactions were performed as described in Fig. 1, and only the ADP-ribosyltransferase activity of the supernatant fluid fractions is shown. CE, crude extract; DEAE, DEAE-Sepharose anion exchange chromatography (150–190 mM NaCl fractions); S200, Sephacryl® 200 sizing chromatography (250–100-kD fractions); MQ, Mono Q anion exchange chromatography (27.3-mS fractions). (B) Colloidal Coomassie stained 10% SDS-PAGE protein band profiles after Mono Q anion exchange chromatography. Partially purified CTF complex fractions from both T cells and yeast cells were eluted at a conductance of 27.3 mS.
Identification of individual components of the CTF complex
Tryptic peptides from “in-gel” digestion of individual protein bands resolved by SDS-PAGE were subjected to analysis by mass spectroscopy using matrix-assisted laser desorption/ionization–time-of-flight (MALDI-TOF) and nano-electrospray ionization (ESI) quadrupole/orthogonal TOF spectrometers (Jensen et al., 1999). Peptide maps and tandem MS sequence data allowed for the unequivocal identification of Hsp 90 (α and β) and TrR-1 in the partially purified CTF complex mixture from human T cells (Fig. 4; Table I). Importantly, the corresponding yeast homologues, Hsp 82 and TrR-1, as well as thioredoxin peroxidase, were identified in the partially purified CTF complex from yeast cells (Table I). The cumulative peptide coverage for each protein identified through LC-MS/MS sequencing was between 65 and 85% of the total protein (Table S1, available at www.jcb.org/cgi/content/full/jcb.200210028/DC1). Ions unassigned in the LC-MS/MS spectra were indicative of truncation, sequence variation, and/or post-translational modification.
Figure 4.

Identification of putative CTF(s) using MS. (A) Representative total ion chromatogram from an online capillary liquid chromatography mass spectrometric analysis of the in-gel tryptic digest after immunoprecipitation of the 84-kD band (Fig. 2 B and Fig. 5 B) from human partially purified CTFs using rabbit polyclonal anti-Hsp 90 antibodies. (B) Mass spectrum from LC-MS elution at time 19.5 to 20.5 min, as indicated by the shaded region in (A). Peaks are labeled with the m/z value, the charge state, the corresponding amino acid segment, and specification of the Hsp 90 isoform. (C) Tandem mass spectrum for m/z 575.984+ (redundant sequence from Hsp 90 α and β; see inset on B). Complementary a and b (NH2-terminal derived) as well as y (COOH-terminal derived) ions are labeled in the spectrum with m/z value and charge state. All observed a, b, and y ions are indicated in the peptide diagram. Data were analyzed using BioAnalyst™ (Applied Biosystems) reconstruction algorithms. For initial screening and searches, acquired mass values were compared with theoretical protein digests using the Mascot search engine (Matrix Science Ltd.).
Table I. Summary of the data obtained for each putative CTF component identified in this work: MALDI, ESI-MS/MS, LC-MS/MS, Western blot (WB), in vitro translocation assay (TA), and mammalian cell cytotoxicity assay (CA).
| Putative CTFs | Data obtained |
|---|---|
| Human T cell (Hut 102/6TG) |
|
| Hsp 90 α | MALDI, ESI-MS/MS, LC-MS/MS, WB, TA, CA |
| Hsp 90 β | MALDI, ESI-MS/MS, LC-MS/MS, WB, TA, CA |
| TrR-1 | MALDI, ESI-MS/MS, WB, TA, CA |
| Yeast (NLY22−) | |
| Hsp 82 | MALDI, ESI-MS/MS, WB, TA, CA |
| TrR-1 | MALDI, LC-MS/MS, TA, CA |
| Thioredoxin peroxidase (AHP1) |
MALDI, ESI-MS/MS |
Hsp 90 is essential (but not sufficient) for C-domain translocation in vitro
To determine whether Hsp 90 was a component of the CTF complex and to establish a functional role for this chaperonin in C-domain translocation, we conducted a series of experiments to examine the effects of using both polyclonal anti-Hsp 90 antibodies and specific inhibitors. As shown in Fig. 5 A, either immunoprecipitation of Hsp 90 from the CTF complex or addition of anti-Hsp 90 to the CTF complex before initiation of the translocation reaction resulted in marked loss of ADP-ribosyltransferase activity in the supernatant fluid fraction. There was no significant loss of ADP-ribosyltransferase activity in the pellet fractions, which argues against the possibility of any translocated but nonrefolded pool of DT C-domain. Because attempts to reconstitute the immunodepleted CTF complex with human recombinant (hr) Hsp 90 failed to restore translocation activity, we conclude that additional, as yet unknown, protein(s) required for translocation was also removed by immunoprecipitation (Fig. 5 B).
Figure 5.

Hsp 90 is a component of the CTF complex. (A) Hut 102/6TG (H) and yeast NLY22− (Y) partially purified CTF complexes were coincubated with 1 μg anti-Hsp 90 and 1 μg anti-Hsp 82 antibodies, respectively, for 10 min at RT before standard analysis in the in vitro translocation assay. Analogously, CTF complexes were depleted of Hsp 90 or Hsp 82 by immunoprecipitation and assayed for translocation activity in vitro. Either partially purified CTF complex or hrHsp 90 was added back to Hsp 90- depleted CTF complexes as indicated, and translocation activity in vitro was assayed. (B) Colloidal Coomassie-stained 10% SDS-PAGE protein band profiles of immunoprecipitated human Hsp 90 (IP), marker (M). Arrow indicates Hsp 90 as identified by ESI LC-MS/MS analysis.
Because geldanamycin and radicicol are well-known inhibitors of Hsp 90, we next examined the effect of these two agents on C-domain translocation in vitro. These agents are known to bind to the ATPase site of the chaperone and block ATP hydrolysis, thereby inhibiting refolding and release of substrate (Grenert et al., 1997; Schulte et al., 1998). Fig. 6 shows that neither the addition of geldanamycin nor radicicol alone was capable of inhibiting C-domain translocation. However, when both inhibitors were used in combination, C-domain translocation was inhibited. There are several reports demonstrating the synergistic inhibitory effects of geldanamycin and radicicol on Hsp 90, and inhibition is thought to result from either the disruption of substrate binding or the interaction with cochaperonins (Rosenhagen et al., 2001). This phenomenon appears to be Hsp 90-specific because the addition of hrHsp 90 to geldanamycin/radicicol-treated human T cell CTF complexes restored C-domain translocation (Fig. 6). Interestingly, the addition of rhHsp 90 to the geldanamycin/radicicol treated CTF complex from yeast only partially restored C-domain translocation in vitro, suggesting that these agents disrupt a species-specific Hsp 82 cochaperone interaction necessary for reconstitution of translocation activity.
Figure 6.

Hsp 90 is essential for mediating DAB 389 IL-2 C-domain translocation from the lumen of early endosomes to the external milieu. The Hsp 90-specific inhibitors, geldanamycin and radicicol, were preincubated with partially purified CTF complex as indicated for 15 min at RT before assaying for translocation activity in vitro. Excess partially purified HUT 102/6TG CTF complex and hrHsp 90 were added to geldanamycin/radicicol treated CTF complexes as indicated and assayed for translocation activity in vitro.
TrR-1 is essential (but not sufficient) for DT C-domain translocation in vitro
Because TrR-1 was also identified by MS sequence analysis of CTF complexes from human T cell and yeast extracts, we have used both immunoprecipitation and specific inhibitors to demonstrate a functional role of TrR-1 in the translocation and/or release of the C-domain from early endosomes. As shown in Fig. 7 A, immunoprecipitation of TrR-1 from human CTF complexes and 2′,5′-ADP-Sepharose affinity chromatographic depletion of yeast TrR-1 from CTF complex mixtures abolished C-domain translocation in vitro. Because there was no significant loss of ADP-ribosyltransferase activity in the pellet fractions (unpublished data), we conclude that there is no pool of translocated but nonrefolded DT C-domain. Reconstitution experiments in which bovine recombinant (br) TrR-1 was added back to TrR-1-depleted CTF complexes, from both T cell and yeast, failed to restore C-domain translocation (unpublished data). Because these experiments were performed under conditions known to reduce the interchain disulfide bond between the C- and T-domains, these results suggest that TrR-1 is a component of a complex and that another factor(s) essential for translocation was codepleted with TrR-1 (Fig. 7 B).
Figure 7.

TrR-1 is a component of the CTF complex. (A) Hut 102/6TG (H) and yeast NLY22− (Y) partially purified CTF complexes were depleted of TrR-1 by immunoprecipitation with anti-human TrR-1 antibodies or affinity purification using 2′,5′ ADP-Sepharose. TrR-1–depleted CTF complexes were then assayed for translocation activity in vitro under reducing conditions. Either partially purified CTF complex or brTrR-1 was added back to TrR-1–depleted CTF complexes as indicated, and translocation activity in vitro was assayed. (B) Colloidal Coomassie-stained 10% SDS-PAGE protein band profiles of 2′,5′ ADP-Sepharose affinity-purified yeast TrR-1 (AP), marker (M). Arrow indicates yeast TrR-1 as identified by ESI LC-MS/MS analysis.
Next, we examined the effect of the TrR-1 stereo-specific inhibitor cis-13-retinoic acid on C-domain translocation (Schallreuter and Wood, 1989). The addition of cis-13-retinoic acid, but not trans-13-retinoic acid, to either human or yeast CTF complex mixtures resulted in the complete inhibition of C-domain translocation in vitro under nonreducing conditions (Fig. 8). Importantly, the addition of excess brTrR-1 to cis-13-retinoic–treated complex restored C-domain translocation activity in vitro, suggesting that cis-13-retinoic acid inhibition is TrR-1 specific. Finally, when assayed under reducing conditions (20 mM DTT), cis-13-retinoic acid had no effect on C-domain translocation (unpublished data). Together, these results indicate that TrR-1 activity plays an essential role in the translocation and/or release of the C-domain from early endosomes.
Figure 8.

TrR-1 function is essential for mediating DAB389IL-2 C-domain translocation from the lumen of early endosomes to the external milieu under nonreducing conditions. Partially purified CTF complex, both human and yeast, were assayed for translocation activity in vitro under nonreducing conditions using translocation buffer containing 10 μM NADPH without DTT. The TrR-1 stereospecific inhibitor cis-13-retinoic acid and the inactive trans-13-retinoic acid isomer were preincubated with partially purified CTF complex as indicated for 15 min at RT before assaying for translocation activity in vitro under nonreducing conditions. Excess brHsp 90 was added to c13RA-treated CTF complexes as indicated, and translocation activity in vitro was assayed under nonreducing conditions.
Geldanamycin/radicicol and cis-13-retinoic acid protect HUT102 6TG cells from the cytotoxic action of DAB389IL-2
Because geldanamycin and radicicol were found to have a synergistic effect in blocking the in vitro translocation of ADP-ribosyltransferase from purified early endosomes, we examined the ability of these agents to protect intact cells from DAB389IL-2. A series of dose-response experiments showed that neither the addition of 10 nM geldanamycin nor 10 nM radicicol alone confer protection against the fusion protein toxin. However, as seen in in vitro translocation assays, these agents in combination (10 nM each) were able to affect a two-log shift in the DAB389IL-2 dose-response curve (IC50 ≈ 10−8 M) compared with the untreated control (IC50 ≈ 5 × 10−10 M). In a similar fashion, cis-13-retinoic acid was found to affect a similar dose-dependent shift in the DAB389IL-2 dose-response curve for HUT102 6TG cells (unpublished data), confirming and extending early observations made by Sandvig and Olsnes (1981).
Discussion
In the present report, we demonstrate that the in vitro translocation of the DAB389IL-2 ADP-ribosyltransferase activity across the membrane of early endosomes and its release into the external milieu requires a CTF complex. Using translocated ADP-ribosyltransferase activity as an assay, we have partially purified the CTF complex from both human T cell and yeast extracts. MS sequencing of individual protein bands revealed by colloidal Coomassie staining of SDS-polyacrylamide gels has allowed the identification of Hsp 90 and TrR-1 from human T cells, and the homologous Hsp 82 and TrR-1 from yeast extracts. A functional role for these proteins in the translocation and/or cytosolic release of ADP-ribosyltransferase activity was established through immunoprecipitation and the use of specific inhibitors.
After depletion of either Hsp 90 or TrR-1 from partially purified human T cell and yeast CTF complexes, we were not successful in reconstituting in vitro translocation of the C-domain by the addition of hrHsp 90 or brTrR-1, either alone or in combination. These results suggest that Hsp 90 chaperonin and TrR-1 are components of a complex(es) that is (are) necessary for facilitating C-domain translocation across the early endosomal membrane. In marked contrast, we were able to reconstitute in vitro C-domain translocation activity in either geldanamycin/radicicol or cis-13-retinoic acid treated CTF complexes by the addition of recombinant proteins to the mixture. Together, these observations lead us to conclude that both the chaperonin Hsp 90 and TrR-1 are required for C-domain translocation, but are not in themselves sufficient.
Hsp 90 is ubiquitously expressed and is known to be a component of several multi-molecular chaperonin complexes that are highly conserved in eukaryotes (Chang and Lindquist, 1994). The interaction of Hsp 90 with other cochaperonins and the formation of discrete complexes is known to mediate Hsp 90 substrate recognition (Caplan, 1999). Although Hsp 90 does not usually directly bind nor refold nascent polypeptides, it is known to refold a growing list of newly synthesized proteins including membrane-associated protein kinases (Bijlmakers and Marsh, 2000). In addition to its refolding activity, Hsp 90 complexes are also known to regulate the trafficking of membrane-associated proteins through interactions with cytoskeleton motors (Pratt et al., 1999).
The CTF complex is capable of refolding thermally denatured DT fragment A in vitro, and refolding requires the ATPase activity of Hsp 90 (unpublished data). However, the inhibition of Hsp 90 ATPase activity by either geldanamycin or radicicol alone does not inhibit translocation of ADP-ribosyltransferase activity across the early endosomal membrane. As such, it would appear that refolding of denatured C-domain into an active conformation and translocation are mutually exclusive events. The synergistic effects of geldanamycin and radicicol on the inhibition of ADP-ribosyltransferase translocation are of interest, and is consistent with previous reports (Rosenhagen et al., 2001). It is possible that when used in combination, these inhibitors result in either a disruption of Hsp90 substrate recognition and/or the disruption of Hsp 90–cochaperone interactions, thereby leading to an inhibition of C-domain translocation. Although a firm conclusion cannot yet be reached, the inability to reconstitute yeast CTF complexes with mammalian factors supports the later hypothesis. We are currently investigating whether or not Hsp 90 interacts directly with the fusion protein toxin during translocation or is simply an architectural component of the complex.
After furin-mediated nicking of the α-carbon backbone of either DT or DAB389IL-2, retention of the interchain disulfide bond between the C- and T-domains of the toxin presumably is essential for insertion and threading of the denatured C-domain into and through the nascent channel formed by the T-domain (vanderSpek et al., 1994). Moreover, post-translocation reduction of this disulfide bond is also required for the release of the C-domain into the cytosol because unreduced C-domain and membrane-inserted T-domain are both targeted for proteolytic degradation (Moskaug et al., 1993; Madshus et al., 1994). Indeed, the pivotal role of this event is underscored by the observation that reduction of this interchain disulfide bond is the rate-limiting step in the diphtherial intoxication of eukaryotic cells (Papini et al., 1993). Observations reported here confirm and extend these earlier findings, and strongly suggest that TrR-1 is a component of the CTF complex required for the release of the C-domain from the early endosome. These observations also confirm and extend the earlier observations of Sandvig and Olsnes (1981), who reported that retinoic acids inhibit the action of several AB toxins, including DT, on eukaryotic cells.
Although the data reported here clearly demonstrate that TrR-1 activity is required for at least the cytosolic release of the DAB389IL-2 C-domain from purified early endosomes, we cannot conclude whether or not TrR-1 is directly involved in the reduction of the interchain disulfide bond. Because we have identified thioredoxin peroxidase in CTF complexes purified from yeast, it is possible that TrR-1 functions indirectly through a cascade of reductases (e.g., thioredoxin; Moskaug et al., 1987).
It is widely accepted that anthrax lethal toxin and edema factor, as well as the botulinum neurotoxins, must pass through an acidic early endosomal compartment in order to deliver their respective C-domain into the cytosol of targeted cells. The unfolding of the C-domains of anthrax lethal factor (Wesche et al., 1998) and botulinum toxin serotype D (Bade et al., 2002), as well as the TrR-1–mediated reduction of the botulinum neurotoxins (Kistner and Habermann, 1992; Bigalke and Shoer, 2000), have been postulated to be prerequisites for their delivery to the cytosol. Accordingly, the findings reported here may have wider implications. Importantly, several protein complexes of similar composition have been described in protein-trapping proteomic analysis of yeast. For example, Ho et al. (2002) has shown that cyclophilin-trapped complexes from yeast contain Hsp 82, TrR-1, and Sec 27. Moreover, cyclophilin is required for the cytosolic entry of HIV (Braaten et al., 1996), the vacuolar import of fructose-1,6-bisphophatase (Brown et al., 2001), and the activation of peroxiredoxins (Lee et al., 2001). It should also be noted that trafficking mechanisms mediated by cyclophilin–Hsp 90 complexes are synergistically affected by geldanamycin and radicicol (Meyer et al., 2000). In aggregate, observations reported here confirm and extend the hypothesis that multiple pathogens from diverse phylogenetic backgrounds, as well as many of their virulence determinants have convergently evolved to recruit host cell proteins (e.g., CTF complexes) in order to facilitate their membrane translocation and release into the cytosol of eukaryotic cells.
Materials and methods
Cell culture
HUT1026TG cells (TIB 1620; American Type Culture Collection), were maintained in RPMI 1640 (BioWhittaker) supplemented with 10% FBS (HyClone), 2 mM glutamine (BioWhittaker), 50 IU/ml penicillin, and 50 μg/ml streptomycin (BioWhittaker) at 37°C in 5% CO2. Yeast strain NLY22− (a gift from Dr. Kevin Jarrell, Modular Genetics Inc., Lincoln, MA) was maintained in YPD media (Difco®) and on YPD agar plates at 30°C.
Purification of EF-2
EF-2 was partially purified using a procedure by Chung and Collier (1977). After purification, fractions containing EF-2 were identified by ADP-ribosyltransferase using DAB389IL-2 (see In vitro ribosylation assay). EF-2 was further purified by DEAE-Sepharose (Reactifs IBF) anion exchange chromatography. EF-2 was eluted with a linear gradient, 0–200 mM NaCl, in 50 mM Tris-HCl, pH 8.0, 50 mM Mg(OAc)2, 0.1 M KCl, 4 mM CaCl2, 5 mM 2-ME and 1 μg PMSF (Sigma-Aldrich) per ml. Fractions containing EF-2 were identified as above. Aliquots were adjusted to a final concentration of 2 mM DTT, 5% glycerol, and stored at −70°C. Purified EF-2 was ∼80% homogeneous as resolved by 7% SDS-PAGE and stained with colloidal Coomassie (Invitrogen). Protein concentration was determined by Bradford Assay according to standard protocols using Coomassie Protein Assay Reagent (Pierce Chemical Co.).
Purification of early endosomes
Early endosomes were isolated from HUT102/6TG cells according to a protocol by Duprez and Dautry-Varsat (1986). The early endosomal compartment was loaded with 1 μM DAB389IL-2, 1 μM DA189(VSV-G)B389IL-2, 8 mg/ml 70-kD SNARF1-dextran conjugate (Molecular Probes, Inc.), and/or 5 mg/ml HRP (Sigma-Aldrich) using 1 μM bafilomycin A1–primed cells (Sigma-Aldrich).
Purification of HUT102/6TG CTF complex
Crude cytosolic extract was isolated from HUT 102/6TG cells according to the protocol modified from Bomsel et al. (1990). In brief, cells were washed three times with cold PBS containing 5 mg/ml BSA, once with cold PBS alone, and twice with cold cytosol buffer (CB; 3% sucrose in 100 mM Hepes-KOH, pH 7.9, 1.4 M KCl, 30 mM MgCl2, 2 mM EDTA, and 5 mM DTT). Cells were lysed by 20 passages through a 25 G needle in CB containing protease inhibitors as follows: 10 μg/ml aprotinin, 1 μg/ml pepstatin, 1 μg/ml antipain, and 1 μm PMSF (all obtained from Sigma-Aldrich) The lysate was centrifuged at 1,000 g for 15 min at 4°C. The post-nuclear supernatant was then centrifuged at 170,000 g for 1 h at 4°C. The supernatant fraction was dialyzed overnight at 4°C against cytosol dialysis buffer (CDB; 1% sucrose in 20 mM Tris-HCl, pH 8.0, 2 mM EDTA, and 2 mM 2-ME) containing protease inhibitors as described in CB.
Crude cytosol was fractionated according to standard chromatographic protocols. In brief, crude extract was loaded onto an in-house packed DEAE-Sepharose (Reactifs IBF) XK 26 column (Amersham Biosciences) for anion exchange chromatography. A peristaltic FPLC pump (P-1; Amersham Biosciences) and Single Path Monitor (UV-1; Amersham Biosciences) were used during chromatography. The column was preequilibrated with buffer B3 (containing 50 mM Tris-HCl, pH 8.0, 1 mM EDTA, 5 mM 2-mercaptoethanol, and 1 μg PMSF per ml), and “loaded” sample was washed using the same buffer. CTFs were eluted with a linear gradient, 0–400 mM NaCl, in buffer B3 at a flow rate of 5 ml/min. Fractions containing CTFs were identified using an in vitro translocation assay and in vitro ribosylation assay in series (see Materials and methods). Fractions containing in vitro translocation activity eluted between 150 to 190 mM NaCl, and were pooled and concentrated using Centriplus Centrifugal Filters (YM-10; Amicon) according to manufacturer's directions. Protein concentration was determined as described by Bradford assay.
Next, CTFs were fractionated by size exclusion chromatography using Sephacryl® S200 (Amersham Biosciences) XK 26 column (Amersham Biosciences) equilibrated with buffer B3. A Single Path Monitor (UV-1; Amersham Biosciences) was used to monitor chromatography. Sample loads of 5 ml were isocratically eluted in buffer B3. Flow rate was gravitationally determined at ∼2 ml per min. Resolution of the mobile phase was monitored by 7–12% SDS-PAGE and staining with colloidal Coomassie. CTFs were identified using an in vitro translocation assay and an in vitro ribosylation assay in series, and correlated with elution of 100 to 250 kD sized proteins, but contained proteins as small as 20–25 kD when visualized by 7%-12% SDS-PAGE and stained with colloidal Coomassie.
Partially purified CTFs were further purified by anion exchange chromatography using a column (Mono Q HR 5/5; Amersham Biosciences) on an HPLC (Biosys2000; Beckman Coulter). The column was preequilibrated with buffer B4 (containing 50 mM Tris-HCl, pH 8.0, and 1 mM EDTA). Sample loads of 2 ml were washed using buffer B4 and CTFs were eluted using serial hyperbolic step gradients, 0 to 1.0 M NaCl, in buffer B4 at a flow rate 2 ml/min. CTFs were identified using an in vitro translocation assay and an in vitro ribosylation assay in series and eluted at a conductance of 27.3 mS. Translocation in vitro-competent fractions were pooled, dialyzed against 50 mM Tris-HCl, pH 7.4, and 1% sucrose overnight at 4°C, and then concentrated using Microcon Centrifugal Filters (YM-10; Amicon) according to manufacturer's directions. Protein concentration was determined as by Bradford assay. Controls indicated that the purified CTF complex had no intrinsic ADP-ribosyltransferase activity.
Purification of NLY22− CTF complex
Yeast crude cytosolic extract was isolated using the same procedure described above for HUT 102/6TG cells, except NLY22− cells were lysed by vortexing cells with 212–300 micrometer glass beads (Sigma-Aldrich). Cell lysis was monitored by decrease in exclusion of Trypan Blue dye (GIBCO BRL). Controls indicated that the purified CTF complex had no intrinsic ADP-ribosyltransferase activity.
In vitro translocation assay
Translocation of the C-domain was performed using protocol modified by Lemichez et al. (1997) as follows: 25-μl reaction mixtures containing 4 μl early endosomes in translocation buffer (TB; 50 mM Tris-HCl, pH 7.4, and 25 mM EDTA). For reducing conditions, TB contained 20 mM DTT. For nonreducing conditions, TB contained 10 μM NADPH (Qbiogene). ATP and cytosol were added to 2 mM and 5.0 to 0.09 μg/μl as indicated, respectively. Translocation mixtures were incubated at 37°C for 30 min, and the supernatant fluid and pellet were separated by ultracentrifugation at 180,000 g at 4°C for 20 min. The pellet fraction was resuspended in 25 μl TB containing 0.2% Triton X-100 (Sigma-Aldrich), and both the lysed pellet and supernatant fluid were boiled for 5 min. The inhibitors geldanamycin (Alomone Labs), radicicol (Sigma-Aldrich), cis-13-retinoic acid (Sigma-Aldrich), and trans-13-retinoic acid (Sigma-Aldrich) were added as indicated. hrHsp 90 (StressGen Biotechnologies), brTrR-1 (American Diagnostica, Inc.), and hrTrx (American Diagnostica, Inc.) were added as indicated. The membrane integrity of purified early endosomes in the assay system was verified using HRP as described by Lemichez et al. (1997).
In vitro ADP-ribosylation assay
The in vitro NAD+-dependent ADP-ribosylation of EF-2 was performed according to a protocol by Chung and Collier (1977). Reaction mixtures contained 3 pM [32P]-NAD+ (800 μCi/mmol; Dupont-NEN Life Science Products), and when indicated 1 mM ATP and/or 0.5 mg/ml crude HUT102/6TG cytosol. Where indicated, autoradiographic signals on X-OMAT AR film (Kodak) were analyzed by ImageQuant™ software (Molecular Dynamics) and Kodak ID software (Kodak) according to manufacturer's directions.
Immunoprecipitation and affinity chromatography
Immunoprecipitation of human Hsp 90 (both α and β), yeast Hsp 82, and human TrR-1 were performed according to standard protocols using rabbit IgG polyclonal anti–human Hsp 90 antibodies (Santa Cruz Biotechnology, Inc.), rabbit polyclonal anti-Hsp82 antiserum (a gift from S. Lindquist, Massachusetts Institute of Technology, Cambridge, MA), and rabbit polyclonal anti–human TrR-1 antibodies (Upstate Biotechnology). Antibody was first cross-linked to Protein A Agarose (Santa Cruz Biotechnology, Inc.) before immunoprecipitation. In each instance, 2–4 μg rabbit polyclonal was incubated with 100 μl or 200 μl of resuspended volume of Protein A Agarose in 50 mM Tris-HCl and 1 mM EDTA containing 1% NP-40 and 100 mM NaCl on a rocker overnight at 4°C. Bound antibody was collected by centrifugation at 1,000 g for 5 min at 4°C, and washed 2× with 10× current volume with 0.2 M sodium borate (Sigma-Aldrich), pH 9.0, for 5 min at 25°C. Dimethyl Pimelimidate.2HCl (Sigma-Aldrich) was added to a final concentration of 20 mM, and the reaction mixture was incubated for 30 min at 25°C. Cross-linked antibody was pelleted by centrifugation at 1,000 g for 5 min at 4°C, and the pellet was washed 2× with 10× current volume 0.2 M ethanolamine (Sigma-Aldrich) for 30 min at 25°C, and 2× with PBS for 30 min at 25°C.
Immunoprecipitations using the cross-linked antibody agarose conjugates were performed according to standard protocols. In brief, 200 μl of Mono Q partially purified CTFs (∼0.1 μg/μl) in 50 mM Tris-HCl and 1% sucrose, containing 1% NP-40 and 25 mM NaCl, was incubated with 20 μl of antibody-agarose conjugate on a rocker overnight at 4°C. Immunoprecipitates were collected by centrifugation at 1,000 g for 5 min at 4°C, and supernatant fluid was evaluated in the in vitro translocation assay. Pellet was washed 3× with 100 μl cold 50 mM Tris-HCl and 1 mM EDTA containing 1% NP-40 and 50 mM NaCl, and resuspended in 50 μl 1× SDS-PAGE loading buffer and boiled for 5 min. Antibody-agarose beads were pelleted by centrifugation at 1,000 g for 5 min at 25°C and the supernatant was analyzed by 10% SDS-PAGE, stained with colloidal Coomassie, and selected bands were evaluated by LC-MS/MS.
Yeast TrR-1 was affinity purified using 2′,5′ ADP-Sepharose agarose (Amersham Biosciences) using a protocol modified from Hunt et al. (1983). In brief, 20 μg of 2′,5′ ADP-Sepharose agarose was washed 2× with 200 μl 50 mM Tris-HCl and 1 mM EDTA for 20 min. Mono Q partially purified CTFs (200 μl of ∼0.1 μg/μl) in 50 mM Tris-HCl, 1 mM EDTA, 1% sucrose, and 25 mM NaCl was incubated with 2′,5′ ADP-Sepharose on a rocker overnight at 4°C. Affinity-purified TrR-1 was collected by centrifugation at 1,000 g for 5 min at 4°C. The supernatant fluid was assayed for translocation activity in vitro. The pellet was washed 2× in 100 μl 50 mM Tris-HCl, pH 7.5, 1 mM EDTA, and 1% sucrose, and was then resuspended in 50 μl 50 mM Tris-HCl, pH 7.5, 1 mM EDTA, and 1% sucrose containing 20 μM NADPH and incubated for 2 h at 25°C. The supernatant fluid was collected after centrifugation at 1,000 g for 5 min at 4°C, and the supernatant fluid was analyzed by 10% SDS-PAGE, stained with colloidal Coomassie, and selected bands were evaluated by LC-MS/MS.
Western blots
Confirmation of CTF identification by MS was performed by Western blot analysis according to standard protocols. In addition to using antibodies (see Immunoprecipitation and affinity chromatography), horse polyclonal anti-DT antibody (Massachusetts Antitoxin and Vaccine Laboratories) was used. In brief, samples were analyzed by 7–12% SDS-PAGE, transferred to Immobilon-P (Millipore), probed with the appropriate primary and secondary antibodies, and detected using either 3,3′-DAB (Sigma-Aldrich) or ECL (Amersham Biosciences) according to the manufacturer's directions.
In-gel reduction, alkylation, and digestion of partially purified CTFs
The preparation of partially purified CTFs for identification by MS was performed using a modified procedure from Shevchenko et al. (1996). In brief, partially purified CTFs were separated by 10% SDS-PAGE, stained with colloidal Coomassie, and selected bands were excised and chopped into small pieces. Gel pieces were washed 3× in 50 mM ammonium bicarbonate (Sigma-Aldrich) in 50% acetonitrile (ACN; Acros) for 20 min at 25°C. Gel pieces were washed with 100% ACN for 10 min at 25°C. Supernatant was discarded, and the gel pieces were dried in a SpeedVac® for 15 min. Gel pieces were reduced in 20 mM DTT, 50 mM ammonium bicarbonate, and 5% ACN for 1 h at 55°C. Supernatant was discarded and the pieces were washed with 100 μl 50 mM ammonium bicarbonate for 10 min at 25°C and subsequently with 100 μl 100% ACN for 10 min at 25°C.
Gel pieces were alkylated in 100 μl 100 mM iodoacetamide (ICN Biomedicals) and 50 mM ammonium bicarbonate for 30 min in the dark at 25°C. Supernatant was discarded and the pieces were washed with 100 μl 50 mM ammonium bicarbonate for 10 min at 25°C and subsequently dried with 100 μl 100% ACN for 10 min at 25°C. The washing and drying steps were repeated before drying the pieces in a SpeedVac® for 15 min.
Gel pieces were rehydrated in digestion buffer (50 mM ammonium bicarbonate) and MS Sequencing Grade Trypsin (Roche) at an estimated 1:100 enzyme to substrate ratio on ice for 45 min. 50 mM Ammonium bicarbonate was added when necessary to keep the gel pieces wet. Digestions were incubated for 6–8 h at 37°C. Peptides were extracted from the gel pieces using 100 μl 20 mM ammonium bicarbonate for 20 min, followed by 2× 200 μl 1% TFA in 50% ACN for 20 min, and finally 1× 100 μl 100% ACN for 10 min. Supernatant fluids were pooled and dried in a SpeedVac®. The pellets were resuspended in 0.1% TFA and desalted using ZipTip® C18 pipette tips (Millipore) according to manufacturer's directions.
Capillary HPLC of tryptic peptides
HPLC was performed using a capillary LC system (LC Packings; Dionex Corp.) composed of a Famous autosampler, a Switchos microcolumn switching unit and an Ultimate pump. Sample loads of 5 μl were preconcentrated and desalted online with a “small molecule” C18 CapTrap™ (Michrom BioResources) using a solution of 5% FA and 0.1% TFA at a flow rate of 50 μl per min for 4 min. Capillary HPLC columns were prepared in house as follows: 300 μm ID × 15-cm fused silica capillaries were pressure bomb–packed (Mass Evolution, Inc.) at 2,000 PSI with Magic C18 3-μm 200 Å pore-reversed phase-packing material (Michrom BioResources) using 2-propanol as a carrier solvent. Columns were washed with 10% acetic acid, followed by methanol, then the HPLC mobile phase before use at a flow rate of 2 μl per min. Elution was by linear gradient; 95% A (5% ACN, 0.1% formic acid) to 55% B (85% ACN, 10% 2-propanol, 0.1% formic acid) over 50 min followed by 60 min of column regeneration.
MALDI and ESI MS, tandem MS, and LC-MS/MS
MALDI MS was acquired in positive polarity on a mass spectrometer (Reflex IV; Bruker) with delayed extraction in the reflectron mode using a UV nitrogen laser. A laser power of 28–45% was used, and 50–100 laser shots were summed for each spectrum. The matrix used was 2,5-dihydroxybenzoic acid (Sigma-Aldrich). Data were analyzed using BioAnalyst™ (Applied Biosystems) reconstruction algorithms. For initial screening and searches, acquired mass values were compared with theoretical protein digests using the Mascot search engine (Matrix Science, Ltd.). Reported scores, based on a probability of match, were statistically significant for each protein identified in Table I.
ESI MS and MS/MS were performed using an ESI quadrupole/orthogonal acceleration time-of-flight mass spectrometer (QSTARi® Pulsar; Applied Biosystems). MS and MS/MS were acquired in the positive polarity mode over the range of m/z 320–1800 (MS) and m/z 100–1800 (MS/MS) with resolution >1:9,000 (full width half maximum) and better than 50 ppm mass accuracy (external calibration). For nanospray, a Protana source was used using uncoated glass nanospray tips pulled in house to 1 μm ID using a capillary puller (Sutter Instrument Co.) ESI was initiated at ∼1,200 V via a Pt wire inserted into the glass tip. Tandem mass spectra were acquired using Ar as the collision gas and sufficient collision energy to obtain complete sequence information of the precursor. Pulsed ion enhancement of product ions was used for MS/MS of low S/N precursors. For LC-MS, the LC was coupled to the mass spectrometer using 50 μm ID distal coated nanospray tips pulled to 15 μm ID, 75 μm OD at the tip (New Objectives Inc.). ESI was performed at 4,500 V. Information-dependent acquisition was used to obtain MS/MS spectra of peaks during elution from the LC system. MS peaks that exceeded a threshold of 10 counts/s were subjected to MS/MS using preset collision energies proportional to the m/z value of the precursor (∼18–60 V, lab frame). Pulsed ion enhancement was used for all LC-MS/MS spectra.
Cytotoxicity assays
Cytotoxicity assays for the fusion protein toxins were performed essentially as described by vanderSpek et al. (1994). Cytotoxicity assays to evaluate the affects of geldanamycin, radicicol, and retinoic acid on DAB389IL-2 intoxication were modified from vanderSpek et al. (1994) as such: cells were seeded at 5 × 104 cells per well and preincubated with inhibitors geldanamycin, radicicol, cis-13-retinoic acid, for 30 min at 37°C, 5% CO2 and subsequently incubated with varying concentrations of DAB389IL-2 and inhibitor for 15 min at 37°C, 5% CO2. Cells were pelleted and washed free of toxin with media containing inhibitor and incubated for 8–12 h at 37°C, 5% CO2. Cells were then washed and pulsed with minimal media (leucine depleted; BioWhittaker) containing [14C]leucine (NEN Life Science Products) for 2 h at 37°C, 5% CO2, and protein synthesis was analyzed according to vanderSpek et al. (1994). Media alone and media plus inhibitor alone served as controls. Assays were performed in quadruplicate.
Online supplemental material
Table of peptide coverage from LC-MS/MS is available as online supplemental material at http://www.jcb.org/cgi/content/full/jcb.200210028/DC1.
Supplemental Material
Acknowledgments
Dedicated in memory of Dr. Harold Amos.
We would like to thank E. Simons and H. Long for help with fluorescence studies, G. Belfort for thoughtful discussions, E. Keller for thoughtful discussions and critical reading of the manuscript, and S. Lindquist for the gift of anti-Hsp 82 antibodies.
R. Ratts is supported by NIH training grant DK07201. J.R. Murphy is supported by NIH grant RO1 CA60934. C.E. Costello is supported by NIH Grants P41-RR10888 and S10 RR15942.
The online version of this article includes supplemental material.
H. Zeng's present address is Dept. of Pathology, Harvard Medical School, Boston, MA 02115.
C. Blue's present address is Division of Infection and Immunity, University of Glasgow, Glasgow G12 8QQ, UK.
Footnotes
Abbreviations used in this paper: br, bovine recombinant; C, catalytic; CTF, cytosolic translocation factor; DT, diphtheria toxin; EF-2, elongation factor 2; ESI, electrospray ionization; hr, human recombinant; Hsp, heat shock protein; IL-2, interleukin-2; MALDI, matrix-assisted laser desorption ionization; MS, mass spectrometry; T, transmembrane; TrR-1, thioredoxin reductase.
References
- Ariansen, S., B.N. Afanasiev, J.O. Moskaug, H. Stenmark, I.H. Madshus, and S. Olsnes. 1993. Membrane translocation of diphtheria toxin A-fragment: role of carboxy-terminal region. Biochemistry. 32:83–90. [DOI] [PubMed] [Google Scholar]
- Bacha P., C. Waters, J. Williams, J.R. Murphy, and T.B. Strom. 1988. Interleukin 2 receptor-targeted cytotoxicity. Interleukin 2 receptor-mediated action of a diphtheria toxin-related interleukin 2 fusion protein. J. Exp. Med. 167:612–622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bade, S., A. Rummel, J. Alves, H. Bigalke, and T. Binz. 2002. New insights into the translocation process of botulinum neurotoxins. Naunyn. Schmiedebergs Arch. Pharmacol. 365(Sup. 2):R13. [Google Scholar]
- Bigalke, H., and L.F. Shoer. 2000. Clostridial neurotoxins. Bacterial Protein Toxins. K. Aktories and I. Just, editors. Springer-Verlag, Berlin. 407–444.
- Bijlmakers, M.J., and M. Marsh. 2000. Hsp90 is essential for the synthesis and subsequent membrane association, but not the maintenance, of the Src-kinase p56(lck). Mol. Biol. Cell. 11:1585–1595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bomsel, M., R. Parton, S.A. Kuznetsov, T.A. Schroer, and J. Gruenberg. 1990. Microtubule- and motor-dependent fusion in vitro between apical and basolateral endocytic vesicles from MDCK cells. Cell. 62:719–731. [DOI] [PubMed] [Google Scholar]
- Braaten, D., E.K. Franke, and J. Luban. 1996. Cyclophilin A is required for the replication of group M human immunodeficiency virus type 1 (HIV-1) and simian immunodeficiency virus SIV(CPZ)GAB but not group O HIV-1 or other primate immunodeficiency viruses. J. Virol. 70:4220–4227. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, C.R., D.Y. Cui, G.G. Hung, and H.L. Chiang. 2001. Cyclophilin A mediates Vid22p function in the import of fructose-1,6-bisphosphatase into Vid vesicles. J. Biol. Chem. 276:48017–48026. [DOI] [PubMed] [Google Scholar]
- Cabiaux, V., P. Quertenmont, K. Conrath, R. Brasseur, C. Capiau, and J.M. Ruysschaert. 1994. Topology of diphtheria toxin B fragment inserted in lipid vesicles. Mol. Microbiol. 11:43–50. [DOI] [PubMed] [Google Scholar]
- Caplan, A.J. 1999. Hsp90's secrets unfold: new insights from structural and functional studies. Trends Cell Biol. 9:262–268. [DOI] [PubMed] [Google Scholar]
- Chang, H.C., and S. Lindquist. 1994. Conservation of Hsp90 macromolecular complexes in Saccharomyces cerevisiae. J. Biol. Chem. 269:24983–24988. [PubMed] [Google Scholar]
- Choe, S., M.J. Bennett, G. Fujii,P.M. Curmi, K.A. Kantardjieff, R.J. Collier, and D. Eisenberg. 1992. The crystal structure of diphtheria toxin. Nature. 357:216–222. [DOI] [PubMed] [Google Scholar]
- Chung, D.W., and R.J. Collier. 1977. The mechanism of ADP-ribosylation of elongation factor 2 catalyzed by fragment A from diphtheria toxin. Biochim. Biophys. Acta. 483:248–257. [DOI] [PubMed] [Google Scholar]
- Donovan, J.J., M.I. Simon, R.K. Draper, and M. Montal. 1981. Diphtheria toxin forms transmembrane channels in planar lipid bilayers. Proc. Natl. Acad. Sci. USA. 78:172–176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duprez, V., and A. Dautry-Varsat. 1986. Receptor-mediated endocytosis of interleukin 2 in a human tumor T cell line. Degradation of interleukin 2 and evidence for the absence of recycling of interleukin receptors. J. Biol. Chem. 261:15450–15454. [PubMed] [Google Scholar]
- Falnes, P.O., S. Choe, I.H. Madshus, B.A. Wilson, and S. Olsnes. 1994. Inhibition of membrane translocation of diphtheria toxin A-fragment by internal disulfide bridges. J. Biol. Chem. 269:8402–8407. [PubMed] [Google Scholar]
- Grenert, J.P. W.P. Sullivan, P. Fadden, T.A. Haystead, J. Clark, E. Mimnaugh, H. Krutzsch, H.J. Ochel, T.W. Schulte, E. Sausville, et al. 1997. The amino-terminal domain of heat shock protein 90 (hsp90) that binds geldanamycin is an ATP/ADP switch domain that regulates hsp90 conformation. J. Biol. Chem. 272:23843–23850. [DOI] [PubMed] [Google Scholar]
- Hammond, K., G.A. Caputo, and E. London. 2002. Interaction of the membrane-inserted diphtheria toxin T domain with peptides and its possible implications for chaperone-like T domain behavior. Biochemistry. 41:3243–3253. [DOI] [PubMed] [Google Scholar]
- Ho, Y., A. Gruhler, A. Heilbut, G.D. Bader, L. Moore, S.L. Adams, A. Millar, P. Taylor, K. Bennett, K. Boutilier, et al. 2002. Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry. Nature. 415:180–183. [DOI] [PubMed] [Google Scholar]
- Hunt, T., P. Herbert, E.A. Campbell, C. Delidakis, and R.J. Jackson. 1983. The use of affinity chromatography on 2′5′ ADP-sepharose reveals a requirement for NADPH, thioredoxin and thioredoxin reductase for the maintenance of high protein synthesis activity in rabbit reticulocyte lysates. Eur. J. Biochem. 131:303–311. [DOI] [PubMed] [Google Scholar]
- Jensen, O.N., M. Wilm, A. Shevchenko, and M. Mann. 1999. Peptide sequencing of 2-DE gel-isolated proteins by nanoelectrospray tandem mass spectrometry. Methods Mol. Biol. 112:571–588. [DOI] [PubMed] [Google Scholar]
- Kagan, B.L., A. Finkelstein, and M. Colombini. 1981. Diphtheria toxin fragment forms large pores in phospholipid bilayer membranes. Proc. Natl. Acad. Sci. USA. 78:4950–4954. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kistner, A., and E. Habermann. 1992. Reductive cleavage of tetanus toxin and botulinum neurotoxin A by the thioredoxin system from brain. Evidence for two redox isomers of tetanus toxin. Naunyn Schmiedebergs Arch. Pharmacol. 345:227–234. [DOI] [PubMed] [Google Scholar]
- Lee, S.P., Y.S. Hwang, Y.J. Kim, K.S. Kwon, H.J. Kim, K. Kim, and H.Z. Chae. 2001. Cyclophilin a binds to peroxiredoxins and activates its peroxidase activity. J. Biol. Chem. 276:29826–29832. [DOI] [PubMed] [Google Scholar]
- Lemichez, E., M. Bomsel, G. Devilliers, J. vanderSpek, J.R. Murphy, E.V. Lukianov, S. Olsnes, and P. Boquet. 1997. Membrane translocation of diphtheria toxin fragment A exploits early to late endosome trafficking machinery. Mol. Microbiol. 23:445–457. [DOI] [PubMed] [Google Scholar]
- Madshus, I.H., A. Wiedlocha, and K. Sandvig. 1994. Intermediates in translocation of diphtheria toxin across the plasma membrane. J. Biol. Chem. 269:4648–4652. [PubMed] [Google Scholar]
- Meyer, B.K., J.R. Petrulis, and G.H. Perdew. 2000. Aryl hydrocarbon (Ah) receptor levels are selectively modulated by hsp90-associated immunophilin homolog XAP2. Cell Stress Chaperones. 5:243–254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moskaug, J.O., K. Sandvig, and S. Olsnes. 1987. Cell-mediated reduction of the interfragment disulfide in nicked diphtheria toxin. A new system to study toxin entry at low pH. J. Biol. Chem. 262:10339–10345. [PubMed] [Google Scholar]
- Moskaug, J.O., H. Stenmark, and S. Olsnes. 1991. Insertion of diphtheria toxin B-fragment into the plasma membrane at low pH. Characterization and topology of inserted regions. J. Biol. Chem. 266:2652–2659. [PubMed] [Google Scholar]
- Moskaug, J.O., K. Prydz, and S. Olsnes. 1993. Rapid cellular removal of a membrane-inserted foreign polypeptide. Biochem. J. 291:473–477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Oh, K.J., L. Senzel, R.J. Collier, and A. Finkelstein. 1999. Translocation of the catalytic domain of diphtheria toxin across planar phospholipid bilayers by its own T domain. Proc. Natl. Acad. Sci. USA. 96:8467–8470. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papini, E., R. Rappuoli, M. Murgia, and C. Montecucco. 1993. Cell penetration of diphtheria toxin. Reduction of the interchain disulfide bridge is the rate-limiting step of translocation in the cytosol. J. Biol. Chem. 268:1567–1574. [PubMed] [Google Scholar]
- Pratt, W.B., A.M. Silverstein, and M.D. Galigniana. 1999. Model for the cytoplasmic trafficking of signalling proteins involving the hsp90-binding immunophilins and p50cdc37. Cell. Signal. 11:839–851. [DOI] [PubMed] [Google Scholar]
- Ratts, R., and J.C. vanderSpek. 2002. Diphtheria toxin: structure function and its clinical applications. Chimeric Toxins. H. Lorberboum-Galski and P. Lazarovici, editors. Taylor and Francis, London/New York. 14–36.
- Ren, J., K. Kachel, H. Kim, S. Malenbaum, R.J. Collier, and E. London. 1999. Interaction of diphtheria toxin T domain with molten globule-like proteins and its implications for translocation. Science. 284:955–957. [DOI] [PubMed] [Google Scholar]
- Rosenhagen, M.C., J.C. Young, G.M. Wochnik, A.S. Herr, U. Schmidt, F.U. Hartl, F. Holsboer, and T. Rein. 2001. Synergistic inhibition of the glucocorticoid receptor by radicicol and benzoquinone ansamycins. Biol. Chem. 382:499–504. [DOI] [PubMed] [Google Scholar]
- Sandvig, K., and S. Olsnes. 1981. Effects of retinoids and phorbol esters on the sensitivity of different cell lines to the polypeptide toxins modeccin, abrin, ricin and diphtheria toxin. Biochem. J. 194:821–827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schallreuter, K.U., and J.M. Wood. 1989. The stereospecific suicide inhibition of human melanoma thioredoxin reductase by 13-cis-retinoic acid. Biochem. Biophys. Res. Commun. 160:573–579. [DOI] [PubMed] [Google Scholar]
- Schulte, T.W., S. Akinaga, S. Soga, W. Sullivan, B. Stensgard, D. Toft, and L.M. Neckers. 1998. Antibiotic radicicol binds to the N-terminal domain of Hsp90 and shares important biologic activities with geldanamycin. Cell Stress Chaperones. 3:100–108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shevchenko, A., M. Wilm, O. Vorm, and M. Mann. 1996. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 68:850–858. [DOI] [PubMed] [Google Scholar]
- Tsuneoka, M., K. Nakayama, K. Hatsuzawa, M. Komada, N. Kitamura, and E. Mekada. 1993. Evidence for involvement of furin in cleavage and activation of diphtheria toxin. J. Biol. Chem. 268:26461–26465. [PubMed] [Google Scholar]
- Umata, T., Y. Moriyama, M. Futai, and E. Mekada. 1990. The cytotoxic action of diphtheria toxin and its degradation in intact Vero cells are inhibited by bafilomycin A1, a specific inhibitor of vacuolar-type H+-ATPase. J. Biol. Chem. 265:21940–21945. [PubMed] [Google Scholar]
- vanderSpek, J.C., K. Howland, T. Friedman, and J.R. Murphy. 1994. Maintenance of the hydrophobic face of the diphtheria toxin amphipathic transmembrane helix 1 is essential for the efficient delivery of the catalytic domain to the cytosol of target cells. Protein Eng. 7:985–989. [DOI] [PubMed] [Google Scholar]
- Waters, C.A., P.A. Schimke, C.E. Snider, K. Itoh, K.A. Smith, J.C. Nichols, T.B. Strom, and J.R. Murphy. 1990. Interleukin-2 receptor-targeted cytotoxicity. Receptor binding requirements for entry of a diphtheria toxin-related interleukin 2 fusion protein into cells. Eur. J. Immunol. 20:785–791. [DOI] [PubMed] [Google Scholar]
- Wesche, J., J.L. Elliott, P.O. Falnes, S. Olsnes, and R.J. Collier. 1998. Characterization of membrane translocation by anthrax protective antigen. Biochemistry. 37:15737–15746. [DOI] [PubMed] [Google Scholar]
- Wiedlocha, A., I.H. Madshus, H. Mach, C.R. Middaugh, and S. Olsnes. 1992. Tight folding of acidic fibroblast growth factor prevents its translocation to the cytosol with diphtheria toxin as vector. EMBO J. 11:4835–4842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williams, D.P., C.E. Snider, T.B. Strom, and J.R. Murphy. 1990. Structure/function analysis of interleukin-2-toxin (DAB486-IL-2). Fragment B sequences required for the delivery of fragment A to the cytosol of target cells. J. Biol. Chem. 265:11885–11889. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.