

Abstract
Vacuole fusion requires a coordinated cascade of priming, docking, and fusion. SNARE proteins have been implicated in the fusion itself, although their precise role in the cascade remains unclear. We now report that the vacuolar SNAP-23 homologue Vam7p is a mobile element of the SNARE complex, which moves from an initial association with the cis-SNARE complex via a soluble intermediate to the docking site. Soluble Vam7p is specifically recruited to vacuoles and can rescue a fusion reaction poisoned with antibodies to Vam7p. Both the recombinant Vam7p PX domain and a FYVE domain construct of human Hrs block the recruitment of Vam7p and vacuole fusion, demonstrating that phosphatidylinositol 3-phosphate is a primary receptor of Vam7p on vacuoles. We propose that the Vam7p cycle is linked to the availability of a lipid domain on yeast vacuoles, which is essential for coordinating the fusion reaction prior to and beyond docking.
Keywords: vacuole fusion; Vam7p; SNAP-23; SNARE complex; PX domain
Introduction
Membrane fusion along the secretory and endocytic pathway is highly coordinated and is controlled by a variety of proteins (Mellman and Warren, 2000). SNAREs are key proteins in this process: they are found on vesicles and organelles, pair in a cognate fashion, and appear to act at the center of the fusion process itself (for review see Rothman, 1994; Jahn and Südhof, 1999; Chen and Scheller, 2001). SNAREs form cis complexes on membranes that are disassembled through the action of NSF and α-SNAP (Söllner et al., 1993; Walch-Solimena et al., 1995; Otto et al., 1997; Holthuis et al., 1998; Ungermann et al., 1998a). During the fusion reaction, SNAREs from opposite membranes meet in trans to drive the membranes into close apposition (Ungermann et al., 1998b), presumably by forming a tight four-helix bundle as proposed from the crystal structure (Sutton et al., 1998). This complex is sufficient to drive liposome fusion in vitro (Weber et al., 1998). In vivo the situation is more complex. Assembly of trans-SNARE complexes requires the action of specific Rab GTPases and their effectors (Chen and Scheller, 2001; Zerial and McBride, 2001). Rab proteins mediate the initial and reversible contact of membranes by interacting with tethering factors specifically in the GTP-bound form (Cao et al., 1998; Ungermann et al., 1998b; Christoforidis et al., 1999a; Zerial and McBride, 2001). The tethering factors are required before SNARE action and may chaperone SNAREs to the fusion site (McBride et al., 1999). Most tethering proteins belong to multisubunit complexes and have no obvious homology (Zerial and McBride, 2001), though they are structurally related (Whyte and Munro, 2001). Phosphoinositides are closely linked to some of the tethering proteins (Sato et al., 2001; Simonsen et al., 2001). Vac1p and its mammalian relatives EEA1 and rabenosyn-5 localize to the endosome, in part by binding to phosphatidylinositol (PtdIns)* 3-phosphate (PtdIns 3-P) (Stenmark et al., 1996; Burd and Emr, 1998; Kutateladze et al., 1999; Peterson et al., 1999; Nielsen et al., 2000). Furthermore, the PtdIns 3-kinase Vps34p is present in two different complexes, on the trans-Golgi and on autophagosomes (Stack et al., 1995; Kihara et al., 2001). In both cases, it appears that synthesis of PtdIns 3-P is a prerequisite that facilitates a fusion process by permitting the recruitment of essential fusion factors.
In vitro fusion of yeast vacuoles serves as a model system to analyze the characteristics of a membrane fusion reaction. Vacuole fusion occurs in a coordinated cascade of priming, docking, and fusion (Wickner and Haas, 2000) and depends on a SNAP-23 homologue, Vam7p (Ungermann and Wickner, 1998). On the isolated yeast vacuole, Vam7p is part of the SNARE complex, which contains the SNAREs Vam3p, Vti1p, Nyv1p, and Ykt6p (Sato et al., 1998; Ungermann and Wickner, 1998; Ungermann et al., 1999). The HOPS tethering complex and Vac8p are also part of this complex (Price et al., 2000; Veit et al., 2001). During priming, Sec17p and Sec18p (yeast NSF and α-SNAP) disassemble the SNARE complex into its subunits, and HOPS is released and associates with Ypt7p to initiate tethering (Wickner and Haas, 2000). Docking also relies on the action of two Rho GTPases, Cdc42p and Rho1p, that act after Ypt7p (Eitzen et al., 2001; Müller et al., 2001). Later, SNAREs from opposing vacuoles meet in trans to drive the membranes in close apposition (Ungermann et al., 1998b). Fusion then depends on Ca2+, calmodulin, protein phosphatase 1, and the proteolipid of the vacuolar ATPase (Peters and Mayer, 1998; Peters et al., 1999, 2001). Two lipid requirements have been reported. PtdIns 4,5-bisphosphate and ergosterol, the yeast analogue of cholesterol, are required for priming and docking (Mayer et al., 2000; Kato and Wickner, 2001), though their mode of action remains to be discovered.
The role of Vam7p during vacuole fusion is the focus of the present study. We showed previously that extraction of Ypt7p by Gdi1p caused a release of Vam7p to the reaction supernatant, indicating that Vam7p is not firmly anchored to the vacuole (Ungermann et al., 2000). Indeed, Vam7p exists in pools in the cytosol and on the vacuole membrane (Sato et al., 1998), quite similar to the distribution of its homologue SNAP-23 between secretory granules and the cytosol (Guo et al., 1998; Faigle et al., 2000). The NH2-terminal PX domain of Vam7p has been implicated in membrane binding via PtdIns 3-P (Cheever et al., 2001). In the present study, we investigated the mobility of Vam7p and the function of this mobility for vacuole fusion. We demonstrate that Vam7p cycles between the vacuole and the cytosol: Vam7p is released rapidly from the vacuole after priming and is recovered as a soluble factor in the cytosolic fraction, the soluble protein is functional and can rescue vacuole fusion, and Vam7p is recruited to the vacuole membrane before fusion via the PX domain. The specific block of vacuole fusion by exogenously added PX or FYVE domains suggests that a PtdIns 3-P acceptor site on vacuoles is essential for vacuole fusion.
Results
A soluble intermediate of Vam7p
After priming, Vam7p retention on the vacuole depends on Ypt7p (Ungermann et al., 2000). This suggests that Vam7p must be transferred from the cis complex to the tethering machinery. We asked if we could uncouple the transfer of Vam7p by retarding the fusion reaction to characterize the mobility of Vam7p in detail. We incubated isolated vacuoles in the presence of ATP either on ice or at 26°C (Fig. 1 A). Surprisingly, a substantial fraction of Vam7p appeared in the reaction supernatant during the ice incubation (Fig. 1 A, lane 2). Since Sec17p was also observed in the supernatant, priming must have occurred. Other SNARE-associated proteins, such as Vac8p, were not released from the vacuole (Fig. 1 B). Much less Vam7p was found in the supernatant at 26°C (Fig. 1 A, lane 3). Our observations indicate that the released Vam7p becomes unstable at 26°C, whereas it is stable when released on ice, though we do not yet know whether the degradation to a shorter fragment (unpublished data) is a specific effect.
Figure 1.

Priming-dependent release of Vam7p on ice. (A) Vam7p is released from the vacuoles in an ATP-dependent reaction. BJ3505 vacuoles (12 μg) were incubated in a 60-μl reaction for 60 min on ice or at 26°C in reaction buffer containing His6-Sec18p, CoA (10 μM), and ATP as indicated. Then vacuoles were separated from the supernatant (5 min, 20,000 g, 4°C), and the reaction supernatant was precipitated by addition of TCA (13% [vol/vol]) (Ungermann et al., 1998a). Vacuole pellets and precipitated proteins from the supernatant were solubilized in sample buffer and analyzed by SDS-PAGE and immunoblotting. Immunoblots were decorated with indicated antibodies. (B) Release of Vam7p and Sec17p on ice is immediate. BJ3505 vacuoles (12 μg) were incubated on ice, separated into vacuole and supernatant fraction, and processed as in A. Immunoblots of the supernatant fractions were quantified and plotted versus time. The inset shows a quantification of Vam7p in the reaction supernatant after a 60-min incubation on ice or at 26°C. (C) Priming inhibitors block Vam7p release. BJ3505 VAM7-HA vacuoles (12 μg) were incubated in the presence of ATP and IgGs to Sec17p and Sec18p where indicated. Pellet and supernatant fractions were analyzed with HA antibodies to detect Vam7p. HA-tagged Vam7p was used to avoid interference with the added IgGs during immunoblot decoration. (D) Incubation with ATP on ice allows bypass of priming. BJ3505 and DKY6281 vacuoles (6 μg) were incubated in the presence of ATP at the indicated temperature. Inhibitors (α-Sec17p, α-Sec18p [200 μg/ml], α-Vam3p [50 μg/ml], and Gdi1p [300 μg/ml]) were added to the reactions either right away (white bars), after a 30-min ice incubation (gray bars), or after a 30-min incubation at 26°C. Each reaction was then incubated for an additional 60 min at 26°C and then assayed for alkaline phosphatase activity. (E) Vam7p is released from the cis-SNARE complex. BJ3505 vacuoles (36 μg) were incubated in a 180-μl reaction on ice for 10 min. Vacuoles were reisolated (5 min, 20,000 g, 4°C) and solubilized in lysis buffer (150 mM KCl, 0.5% Triton X-100, 20 mM Hepes/KOH, pH 7.4, 1xPIC, and 1 mM PMSF) for 10 min at 4°C and centrifuged (20,000 g, 10 min, 4°C). The soluble detergent extract was added to protein A–Sepharose–coupled antibodies to Vam3p (Ungermann et al., 1998a) and incubated for 2 h at 4°C on a nutator. Reisolated beads were washed once with lysis buffer and once with lysis buffer containing 500 mM KCl. Bound proteins were eluted from the beads with 1 ml of 0.1 M glycine, pH 2.6. The eluate was TCA precipitated and analyzed by SDS-PAGE, Western transfer and immunoblotting with antibodies against the indicated proteins. (F) Released Vam7p is a monomer. BJ 3505 vacuoles (60 μg) were incubated in the presence of ATP on ice or at 26°C as described in the legend to Fig. 1 A. The supernatant was loaded onto a continuous glycerol gradient (10–30% in PS buffer supplemented with 150 mM KCl). The samples were centrifuged at 40,000 rpm in a SW40 rotor for 18 h at 4°C. Fifteen 500-μl fractions were collected, TCA precipitated, and analyzed by SDS-PAGE and immunoblot as before.
We then analyzed the kinetics of the release reaction. About 50% of Vam7p and Sec17p were released from the vacuole within 10 min after ATP addition, significantly more Vam7p than was released if samples were incubated at 26°C (Fig. 1 B). The release of Vam7p on ice was specific. Antibodies to Sec17p and Sec18p, both inhibitors of priming, blocked the release of Sec17p and Vam7p (Fig. 1 C; for an overview of the inhibitors used see Fig. 7). Docking and fusion inhibitors did not interfere with this reaction (unpublished data). In addition, we added inhibitors to a fusion reaction after the ice incubation. Fusion is measured by biochemical complementation using two different types of vacuoles (Conradt et al., 1992; Haas et al., 1994). Vacuoles from one strain contain immature proalkaline phosphatase because of the lack of proteases. The other set of vacuoles contains proteases but lacks alkaline phosphatase. Fusion and content mixing leads to processing and activation of alkaline phosphatase, which can be measured spectrophotometrically (Haas et al., 1994). The fusion reaction had become resistant to Sec17p and Sec18p antibodies during the ice incubation (Fig. 1 D, gray bars). In contrast, the reaction was still inhibited by antibodies to the t-SNARE Vam3p that block docking, demonstrating that we uncoupled priming from docking (Fig. 1 D). Resistance to all inhibitors was achieved after 30 min at 26°C (Fig. 1 D, black bars). The released Vam7p was indeed part of the SNARE complex before release (Fig. 1 E). Vam7p, and Nyv1p and Ykt6p were isolated with Vam3p by coimmunoprecipitation and were released efficiently from Vam3p, regardless of whether the vacuoles were primed on ice or at 26°C (Fig. 1 E) (Ungermann and Wickner, 1998). In contrast, Vam3p remained associated with Vti1p during the ice incubation and was released only if vacuoles were primed at 26°C (unpublished data; Ungermann et al., 1999). Apparently, we are able to resolve multiple steps in the priming reaction.
Figure 7.

Schematic overview of Vam7p rebinding to vacuoles. The three basic stages of the vacuole fusion reaction and the respective inhibitors are shown. V7, Vam7p; PX, PX domain of Vam7p; FYVE, GST2xFYVE of Hrs.
Since the released Vam7p was completely soluble in the reaction supernatant, we analyzed it by glycerol gradient centrifugation to determine its size. Vam7p, a 37-kD protein, migrated in the gradient at 40 kD (Fig. 1 F), implying that Vam7p was not bound to other proteins while released.
Thus, priming of vacuoles on ice uncouples priming from docking and causes a significant release of monomeric Vam7p from the vacuole.
The Vam7p cycle
If after priming Vam7p existed in equilibrium between the soluble and the membrane-bound form, we would expect that repeated removal of the supernatant during the ice/ATP incubation should result in a complete removal of Vam7p. This was indeed the case (Fig. 2 A). More than 90% of Vam7p was released to the reaction supernatant when the ice incubation was done repeatedly. We then asked if the same treatment and the removal of Vam7p would also abolish fusion (Fig. 2 B). Fusion activity was lost when a fresh reaction mixture replaced the Vam7-containing supernatant after the ice incubations (Fig. 1 D, white bars). However, fusion was completely restored if the supernatant of the first ice incubation was added back to the vacuole pellet (Fig. 1 D, white compared with black bars). Repeated centrifugation did not impair fusion activity (Fig. 1 D, gray bars). This suggested that Vam7p could be a mobile fusion factor that is recruited back to the vacuoles before fusion.
Figure 2.

Released Vam7p is functional. (A) Complete Vam7p release during consecutive ice incubations. BJ3505 vacuoles (12 μg) were incubated in the presence of ATP on ice. After 30 min, the reaction was centrifuged (5 min, 20,000 g, 4°C), supernatant was removed, and a fresh reaction mix was added to the vacuoles. This procedure was repeated four times. The supernatant and the pellet were analyzed by immunoblotting with antibodies to Vam7p. Loss of Vam7p in the supernatant in the second reaction is due to the instability of released Vam7p (see text). (B) Removal of the supernatant coincides with loss of fusion activity. A fusion reaction containing the two tester strains was incubated as in A. After each incubation, vacuoles were reisolated (5 min, 20,000 g, 4°C) and resuspended either in new reaction mix containing Sec18p and CoA (white bars), the same supernatant (gray bars) to monitor loss of fusion due to centrifugation, or in the Vam7-containing supernatant removed in the first incubation (black bars). All samples were then incubated for 60 min at 26°C before assaying for fusion activity. (C) Fusion of vacuoles poisoned with anti-Vam7p. Fusion reactions containing both tester strains were incubated in a first incubation on ice or at 26°C for 30 min. Vacuoles were then reisolated (20,000 g, 5 min, 4°C), incubated for 5 min with or without anti-Vam7p on ice, and reisolated. Supernatant of a parallel reaction (30 min on ice in the presence of ATP) containing wild-type vacuoles or vam7Δ vacuoles was added to the pretreated vacuoles, incubated for 60 min at 26°C, and assayed for alkaline phosphatase activity. (D) Depletion of Vam7p from the supernatant blocks fusion. Priming on ice was done two times as described in A. The supernatant containing released Vam7p was incubated for 30 min at 4°C with protein A–Sepharose containing nonimmune IgG, anti-Vam7 (1–122) (α-PX), or anti-Vam7 (165–316) (α-cc). Then, the cleared supernatant was added to primed vacuoles as in A. (E) Soluble Vam7p can rescue fusion at a stage that is insensitive to docking inhibitors. Inhibitors were added to a fusion reaction either from the beginning (white bars) or after pretreatment with anti-Vam7p (black bars) as described in C. Fusion reactions were then incubated for 60 min at 26°C, and fusion activity was determined.
Therefore, we asked whether the released Vam7p was functional in the vacuole fusion reaction (Fig. 2 C). Vacuoles from both tester strains were mixed and incubated on ice to release Vam7p. Then IgGs to Vam7p were added to the acceptor vacuoles to block the portion of Vam7p that was still bound to the membrane (Fig. 2 C, lanes 3–7). Thus, fusion should not occur, unless we provided the reaction with Vam7p. As expected, fusion was blocked if antibodies to Vam7p were added after the ice incubation (Fig. 2 C, lane 4). For this analysis, we reisolated the vacuoles, washed them briefly, and supplied them with the supernatant from a parallel ice incubation of either wild-type (Fig. 2 C, lane 5) or vam7Δ vacuoles (Fig. 2 C, lane 6), or fresh reaction mixture (Fig. 2 C, lane 7). Then the reactions were shifted to 26°C and incubated for 60 min before being assayed for fusion. When new reaction mixture and reaction supernatant from vam7Δ vacuoles primed on ice in parallel were added to the pretreated vacuoles, no fusion activity was recovered (Fig. 2 C, lanes 6 and 7). However, the reaction supernatant from wild-type vacuoles rescued the activity (Fig. 2 C, lane 5). We confirmed this finding by a different approach. When released Vam7p was depleted from the supernatant by two specific antibodies, the supernatant did not rescue the fusion of ice-primed vacuoles (Fig. 2 D), demonstrating that soluble Vam7p is a functional intermediate in vacuole fusion.
We then asked at what stage soluble Vam7p could rescue fusion. Vacuoles were pretreated on ice with ATP to release Vam7p into the supernatant and then reisolated and incubated with anti-Vam7p as before. The indicated inhibitors and the Vam7p-containing supernatant were added to the reisolated and washed vacuoles and were incubated to allow for fusion (Fig. 2 E). Vacuoles became resistant to docking inhibitors during the repeated reisolation procedure. However, they remained sensitive to fusion inhibitors like BAPTA and MCLR (Fig. 2 E) and dependent on the soluble Vam7p provided with the reaction supernatant. Thus, Vam7p can rescue the reaction at a stage when fusion is insensitive to docking inhibitors.
To establish a recruitment assay for Vam7p, we COOH-terminally tagged Vam7p with a protein A affinity tag (Vam7-A). The modified protein replaced the endogenous protein and was completely functional in vacuole fusion (unpublished data). We then performed our release assay with vacuoles carrying either the wild-type or the tagged version of Vam7p. After the ice incubation, vacuoles were sedimented and washed briefly. Then wild-type vacuoles were resuspended in supernatant from vacuoles with the tagged Vam7p, and the reaction was shifted to 26°C. After 60 min, the reaction was separated into vacuoles and reaction supernatant, and both fractions were analyzed by Western blotting. As seen in Fig. 3 A, Vam7-A is mobilized during the ice incubation and a substantial fraction appears in the reaction supernatant (lane 1). Upon incubation of the Vam7-A–containing supernatant with the wild-type acceptor membrane, Vam7-A is efficiently recruited to the vacuole membrane (Fig. 3 A, lane 2). Similar results were obtained with an HA-tagged version of Vam7p (Fig. 3 C). The rebinding of Vam7-A to the acceptor vacuole is finished within 30 min (Fig. 3 B). In contrast, Sec17p that was released in parallel with Vam7p in the first incubation remained in the supernatant (unpublished data), suggesting that the recruitment of Vam7p does not reflect the mere reformation of a cis-SNARE complex.
Figure 3.

Recruitment of soluble Vam7p to vacuoles. (A) Wild-type vacuoles (BJ3505) and Vam7-A vacuoles were incubated in the presence of ATP for 30 min on ice. Vacuoles were reisolated (20,000 g, 5 min, 4°C), and wild-type vacuoles were resuspended in supernatant removed from Vam7-A vacuoles and incubated for 60 min at 26°C. Then the reaction was centrifuged as before and analyzed for Vam7p in pellet (P) and supernatant (S). The second band in lane 1 is a degradation product of Vam7-A. (B) Time course of recruitment. Recruitment of Vam7-A to wild-type vacuoles was done as in A but for different time points. Immunoblots showing Vam7-A were quantified by densitometry and plotted. The total amount of Vam7p in the supernatant was set to 100%. (C) Gdi1p, ATP depletion by apyrase, and neomycin block Vam7p rebinding. Wild-type and Vam7-A vacuoles were preincubated as in A. The indicated inhibitors were added to the recruitment reaction at the following concentrations: BA, 2 mM BAPTA; Gdi, 300 μg/ml Gdi1p; MC, 10 μM microcystin LR; γS, 3 mM Mg-GTPγS; Neo, 500 μM neomycin; and Apy, 20 U/ml apyrase. The pellet was analyzed as before. (D) Priming is essential to create a Vam7 binding site. Wild-type vacuoles were primed on ice for 30 min in the absence or presence of antibodies to Sec17p (200 μg/ml), then reisolated and incubated with Vam7-HA containing supernatant generated in a parallel incubation, and processed as in the legend to Fig. 1 C. Immunoblots were decorated with antibodies to Pho8p to show equal amount of vacuole membranes in each lane. (E) Lipid and protein requirements for Vam7p recruitment. Acceptor vacuoles (12 μg) were primed on ice and resuspended in PS buffer with 150 mM KCl. Trypsin (Try; 10 μg/ml) was added either alone or in the presence of soybean trypsin inhibitor (STI; 100 μg/ml) and incubated with vacuoles for 30 min on ice. STI was then added to stop the digestion. One aliquot was incubated with 2.5 U/ml of the phosphoinositide-specific PLC (Sigma-Aldrich) to remove inositol head groups. Vacuoles were reisolated and used as acceptor membranes as before. Immunoblots were decorated with anti-Pho8p to show equal loading. Note that trypsin digestion caused a truncation of the cytoplasmic tail of Pho8p.
We then analyzed whether inhibitors of the fusion reaction interfere with the recruitment of Vam7p (Fig. 3 C). Because of the frequent instability of Vam7p in the supernatant, only the recruited fraction of Vam7p was used as a read-out of the following experiments. After the incubation with ATP on ice, the indicated inhibitors were added to the reisolated vacuoles before resuspending them in the Vam7-A–containing supernatant. Rebinding of Vam7-A was efficient in the presence of fusion inhibitors (Fig. 3 C, lanes 2, 4, and 5). In contrast, addition of Gdi1p (Fig. 3 C, lane 3), neomycin (Fig. 3 C, lane 6), or apyrase (Fig. 3 C, lane 7), all of which block the docking step of vacuole fusion (Mayer et al., 1996, 2000), prevented the retrieval of Vam7-A from the supernatant completely. The characterized inhibitors may therefore relate to a specific receptor of Vam7p. Interestingly, when anti-Sec17p was added to the acceptor vacuoles before any pretreatment Vam7p binding was completely abolished (Fig. 3 D). Thus, priming is essential to create a binding site for Vam7p.
We then analyzed whether any of the known fusion proteins serves as an acceptor site for Vam7p on the vacuole. Neither IgGs to Vam3p, Nyv1p, Vam7p, Ypt7p, or Vam6p added to acceptor vacuoles, nor the use of vacuoles from deletion strains (vam3Δ, vam7Δ, ypt7Δ, vam6Δ, vac8Δ, or nyv1Δ) as acceptor membranes uncovered a clear binding site (unpublished data). These data do not exclude that the respective proteins are part of a possible binding site. However, protease pretreatment of the acceptor membrane after the ice/ATP incubation prevented rebinding of Vam7-A (Fig. 3 E, lane 2), indicating the role of protein in the recruitment reaction. Strikingly, pretreatment of primed acceptor vacuoles with phosphoinositol-specific PLC from Bacillus cereus, which removed the inositol head groups from all PIPs (Mayer et al., 2000), abolished the recruitment of Vam7p (Fig. 3 E, lane 4).
An essential role of PtdIns 3-P for the recruitment of Vam7p
The requirement for PtdIns suggested that Vam7p might interact with lipids during the recruitment and be required as a prerequisite for fusion. As mentioned before, Vam7p contains a PX domain that binds specifically to PtdIns 3-P (Cheever et al., 2001), whereas the coiled-coil domain is at the COOH terminus. We speculated that PtdIns 3-P might function as the acceptor site for Vam7p and would hence be essential for vacuole fusion.
Therefore, we first analyzed whether the addition of the purified Vam7p PX domain would influence Vam7p recruitment. We purified recombinant N- and COOH-terminal fragments of Vam7p and added them to the fusion reaction (Fig. 4 A). Out of all of the fragments, only the native NH2-terminal fragment inhibited fusion (Fig. 4 A), whereas the NH2-terminal domain containing a point mutation in the PX domain (Cheever et al., 2001) did not have any effect. We then asked at which stage the NH2 terminus would inhibit the fusion reaction. Acquisition of resistance to inhibitors over a time course permits discrimination between stages of the fusion reaction (Mayer et al., 1996). However, this assay indicates only the last step where the respective inhibitor acts. For the experiment, aliquots were removed from an ongoing fusion reaction, mixed with the indicated inhibitor, and then incubated for the remaining time (Fig. 4 B). In addition, an aliquot was placed on ice to monitor the extent of fusion at the time of inhibitor addition. The reaction becomes resistant to inhibitors of priming like anti-Sec17p shortly after the start of the reaction and then to docking inhibitors like anti-Vam7p (Fig. 4 B). Inhibitors of the fusion step inhibit with kinetics close to the ice curve. Interestingly, the reaction was sensitive to the NH2 terminus beyond the docking stage. Note that the antibody was raised to the COOH-terminal 140 amino acids and therefore may not monitor the role of the NH2 terminus precisely. This experiment thus suggests that the NH2 terminus of Vam7p is required until a late stage in the fusion reaction. We then used established assays to analyze the effects of the NH2 terminus in detail. Priming, as monitored by the ATP-dependent release of Sec17p (Mayer et al., 1996), was not inhibited by the addition of the NH2 terminus (unpublished data). Furthermore, trans-SNARE complex formation, an assay that indicates the completion of docking (Ungermann et al., 1998b), was significantly reduced when the NH2 terminus was added (Fig. 4 C). This indicates that the anchoring of Vam7p via the NH2 terminus is required for establishing stable trans-SNARE complexes. When added to the recruitment reaction, only the NH2-terminal peptide blocked Vam7p rebinding from the supernatant (Fig. 4 D). Titrating the fragments into the recruitment assay revealed that inhibition of the Vam7 recruitment completely correlated with the block in vacuole fusion (Fig. 4, A compared with E). This indicates that the NH2 terminus specifically inhibits the Vam7p binding site. In agreement with this, only the native NH2 terminus was able to bind vacuoles (Fig. 4 F). However, the NH2 terminus did not inhibit the fusion step itself. When we incubated vacuoles from the two tester strains in the presence of the fusion inhibitor BAPTA and then added inhibitors before releasing the block by addition of Ca2+ (Mayer et al., 1996; Eitzen et al., 2001), the NH2 terminus did not inhibit any more (Fig. 4 G), in agreement with the kinetic analysis (Fig. 6 B).
Figure 4.

Rebinding of Vam7p depends on the interaction with phosphoinositides. (A) A block of the fusion reaction by addition of the Vam7 PX domain. Purified Vam7p fragments were added at the indicated concentrations to fusion reactions. Abbreviations are as follows: PX, Vam7p (1–122); PX*, Vam7p (1–135) Y42A (Cheever et al., 2001); and cc, Vam7p (165–316). (B) Time course of PX domain inhibition. A 35× scale fusion reaction was started in the presence of ATP at 26°C. Aliquots (30 μl) were removed at the indicated time, antibodies (200 ng/μl) to Sec17p, Vam3p, or Vam7p or the PX domain (15 μM) were added, and samples were incubated at 26°C for a total of 90 min or set on ice before being analyzed for fusion activity. (C) Trans-SNARE formation in the presence of the PX domain. Vacuoles from BJ3505nyv1Δ and DKY6281vam3Δ were incubated in a 600-μl reaction for 40 min at 26°C in the presence of ATP, Sec18p, and CoA. Then, vacuoles were processed and analyzed for trans-SNARE complex formation by coimmunoprecipitation with antibodies to Vam3p as described (Ungermann et al., 1998b). (D) The PX domain blocks the recruitment of Vam7p. The Vam7 fragments were added to primed vacuoles before the addition of Vam7-HA–containing supernatant. Reactions were incubated for 60 min at 26°C before being analyzed. (E) The block in fusion and Vam7p recruitment of Vam7p coincide. The PX domain was added to primed acceptor vacuoles at the indicated concentrations in the presence of ATP for 10 min. Then, recruitment of Vam7-HA was performed as described above. The immunoblot of the pellet fraction was quantified by laser densitometry. (F) Specific binding of the isolated PX domain. The indicated fragments (15 nM each) were added to ice-primed vacuoles. Incubations and processing was as in the legend to Fig. 3 A. Blots were decorated with antibodies to N- and COOH-terminal Vam7p. (G) The PX domain does not block the fusion step itself. Vacuoles from both tester strains were incubated in reaction buffer containing Sec18p and CoA in the absence or presence of 3 mM BAPTA (40 min at 26°C). Inhibitors (anti-Vam3p [α-V3; 200 μg/ml], PX domain [PX; 15 μM], and GTPγS [2 mM]) were added before reversal of the BAPTA block by the addition of 3.5 mM CaCl2. The left two bars are reactions without CaCl2 addition. (H) Irreversible inhibition of fusion by the PX domain. Fusion reactions containing both tester vacuoles were incubated for 30 min on ice in the presence of ATP. Vacuoles were separated from the supernatant by centrifugation (20,000 g, 5 min, 4°C). The PX domain or the mutant form (10 μM) was incubated for 10 min at 26°C with the vacuoles. Then vacuoles were reisolated (20,000 g, 5 min, 4°C) and resuspended in reaction buffer containing ATP, CoA (10 μM), and Sec18p with 20 μM of the coiled-coil domain of Vam7p (cc) without this addition (new) or with the supernatant that had been removed after the first incubation. Fusion was continued for 60 min at 26°C, and alkaline phosphatase activity was determined. (I) The coiled-coil domain of Vam7p cannot rescue the vacuole fusion assay poisoned with antibodies to Vam7p. Pretreatment of the fusion reaction was done as in the legend to Fig. 7 A. Either a new reaction mixture containing CoA and Sec18p or the removed supernatant of the initial incubation was added to the vacuoles. cc indicates that 10 or 20 μM of the coiled-coil domain of Vam7p were added to a new reaction mixture. Reactions were performed for 60 min at 26°C before being assayed for fusion.
Figure 6.

Masking of PtdIns 3-P reveals a specific function of the PX domain. (A) Inhibition of fusion by the recombinant FYVE domain. GST-2xFYVE (as described in Materials and methods) was added at the indicated concentration to the fusion reaction. Fusion was assayed after 90 min at 26°C. (B) Inhibition by the FYVE domain is specific. The indicated concentration of wild-type FYVE domain (1 μM) or an inactive mutant (C215S; 5 μM) was added to a fusion reaction. Alkaline phosphatase activity was determined after 90 min at 26°C. (C) Both PX and FYVE domain block the recruitment of Vam7p. Acceptor vacuoles were preincubated on ice with ATP for 30 min, then reisolated (5 min, 20,000 g, 4°C), and incubated with recombinant FYVE (1 μM) or PX (15 μM) protein. The recruitment of Vam7-HA and the analysis was done as described in the legend to Fig. 3. (D) Kinetic analysis of the PX and FYVE inhibition. The experiment was performed as described in the legend to Fig. 4 B. At the indicated time points, FYVE protein (1 μM) or the PX domain (15 μM) were added. (E) Binding and release of Vam7p from Gdi1p-treated vacuoles. Wild-type acceptor vacuoles were primed on ice, then pretreated with Gdi1p (see Fig. 5 C), resuspended with Vam7-HA containing supernatant, and incubated at 26°C. At the indicated times, aliquots were removed and separated into pellet and supernatant fraction. Analysis of the pellet was done as described above.
We questioned whether we could reverse the block by the NH2 terminus (Fig. 4, H and I). Vacuoles were primed on ice and poisoned either with the NH2 terminus or the mutated version and then reisolated and supplemented with various supernatants. Whereas the NH2 terminus firmly inhibited, the mutant NH2 terminus did not interfere with the rescue of fusion by a supernatant containing released Vam7p (Fig. 4 H). We also asked whether the COOH-terminal coiled-coil domain is sufficient to rescue the fusion reaction poisoned by anti-Vam7p antibodies (Fig. 4 I). Again, rescue was possible with a supernatant containing the endogenous Vam7p but not the COOH terminus alone. Thus, even though the overexpressed COOH terminus is able to rescue vacuole morphology in vivo (Cheever et al., 2001) the anchoring of Vam7p via the NH2-terminal PX domain to vacuoles is necessary to guide the protein to the fusion site in vitro.
Our data suggested that Vam7p was binding to vacuoles via the PX domain at a PtdIns 3-P–specific site. Mayer et al. (2000) reported previously that the PtdIns 3-kinase inhibitor wortmannin inhibits PtdIns 3-P synthesis on isolated vacuoles but does not inhibit in vitro fusion; and in agreement with the results of Mayer et al. (2000), we see little or no inhibition of fusion under similar conditions (Fig. 5 A, lane 1). However, this type of experiment does not eliminate PtdIns 3-P already residing on the vacuole at the time of isolation. Therefore, we sought to deplete PtdIns 3-P by pretreating spheroplasts with wortmannin for 30 min before vacuole purification. When only one vacuole population (Pep4p or Pho8p deficient) was obtained from pretreated spheroplasts, variable degrees of fusion inhibition were noted (unpublished data). However, when both vacuole populations were obtained from wortmannin-pretreated spheroplasts the fusion signal was consistently inhibited by 60–80% and could be further suppressed by including the inhibitor in the reaction buffer (Fig. 5 A). These results indicate that a wortmannin target, probably the Vps34p PtdIns 3-kinase (but possibly also the PtdIns-4-kinase Stt4p [Cutler et al., 1997]), is required to make vacuoles fusion competent. To test this possibility, we used vacuoles deleted for VPS34 as acceptor vacuoles in the recruitment assay. To follow the kinetics of the rebinding of Vam7p in the second incubation, we removed aliquots at the indicated time points. Depletion of PtdIns 3-P caused a severe reduction in the recruitment of Vam7p (Fig. 5 B), and fusion of the vacuoles was strongly reduced (unpublished data). Both results show that Vps34p generates PtdIns 3-P as a prerequisite of efficient fusion and rebinding by Vam7p from the cytosol.
Figure 5.
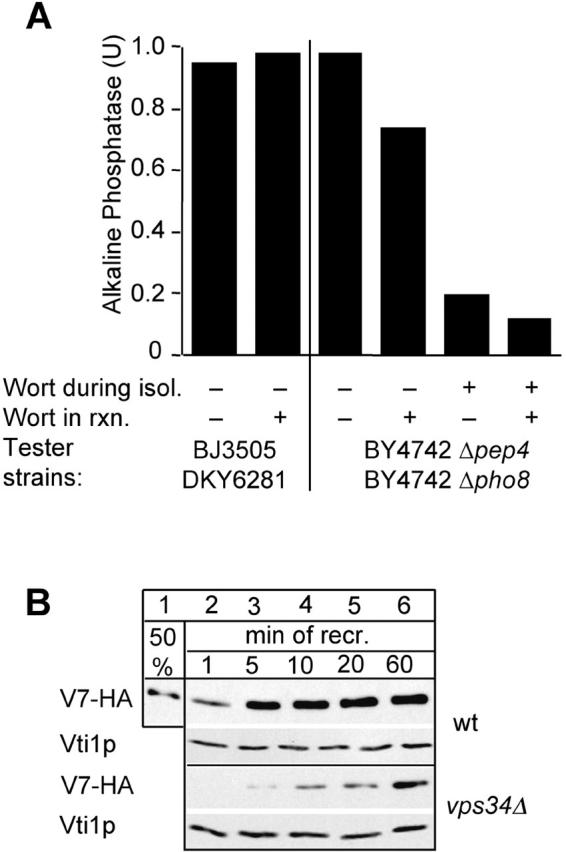
A requirement for PtdIns 3-P for vacuole fusion and Vam7 rebinding. (A) Wortmannin inhibits vacuole fusion. Vacuoles were purified from the indicated wild-type strains and incubated in reaction buffer containing ATP at 26°C for 90 min in the absence or presence of wortmannin (300 μM) before being assayed for fusion. For each reaction, a control incubation was done without ATP, which was subtracted. Where indicated, wortmannin was added during the spheroplasting reaction before vacuole isolation (as described in Materials and methods). (B) Vacuoles lacking Vps34p do not recruit Vam7p efficiently. BJ3505 vacuoles and vps34Δ vacuoles were preincubated on ice with ATP and used as acceptor membranes for soluble Vam7-HA as described in the legend to Fig. 3 A. At the indicated time points, aliquots were removed and processed for recruitment. Analysis of bound proteins was done as before. Decoration of immunoblots with anti-Vti1p served as a loading control.
To more directly target PtdIns 3-P, recombinant GST-2xFYVE domain of the mammalian receptor tyrosine kinase substrate Hrs (Gillooly et al., 2000) was used to selectively block PtdIns 3-P on the vacuole membrane. FYVE domains bind to PtdIns 3-P only (Kutateladze et al., 1999). GST-2xFYVE inhibited in vitro fusion by ∼50% at 200 nM and almost completely at 500 nM (Fig. 6 A). A mutant form of recombinant GST-2xFYVE (C215S), which cannot bind PtdIns 3-P, was noninhibitory even when added at 5 μM (Fig. 6 B). Both the FYVE domain and the PX domain block the recruitment of Vam7p from the supernatant to the vacuole membrane (Fig. 6 C), showing that PtdIns 3-P binding is essential to maintain Vam7p on the vacuole. In addition, kinetic staging experiments indicated that GST-FYVE inhibits at the docking stage of fusion, whereas the PX domain inhibits slightly later (Fig. 6 D). Although the FYVE domain just binds PtdIns 3-P, the PX domain also may bind proteins at the fusion site specifically, which could explain this difference.
However, binding of Vam7p is not limited to an interaction via PtdIns 3-P but must also be regulated. Therefore, we asked which role Ypt7p has as a possible receptor site of Vam7p. In a previous end-point analysis, Ypt7p-extraction by Gdi1p on the acceptor membrane prevented Vam7p recruitment (Fig. 3 C). However, it was possible that Vam7p had been recruited but was released once fusion had occurred. Note that fusion was partially insensitive to Gdi1p after the initial incubation (Fig. 1 E and Fig. 2 D). Therefore, we asked whether the block in recruitment in the presence of Gdi1p was instantaneous after removal of Ypt7p. Vacuoles were primed on ice and then reisolated and incubated in the presence of Gdi1p and soluble Vam7p. We then monitored the rebinding of Vam7p over time. Interestingly, Vam7p was able to rebind but was lost from the membrane at 60 min, the minimal time it takes to complete fusion in our reaction (Fig. 6 E, lanes 2–6). In contrast, Vam7p remained associated with the vacuoles in the absence of Gdi1p (Fig. 6 E, lane 1). Thus, Vam7p can reassociate with vacuoles in the absence of Ypt7p but is not maintained there. One possibility would be that Ypt7p is necessary to interact with Vam7p when a PtdIns 3-P site is not accessible or not present on the vacuole. However, after one complete fusion cycle PtdIns 3-P might be consumed due to the action of PtdIns phosphatase or Fab1p, a PtdIns 3-P 5-kinase (Yamamoto et al., 1995), and synthesis of PtdIns 3-P via the PtdIns 3-kinase Vps34p might then be necessary to maintain a docking site for Vam7p. Future experiments will be necessary to clarify this issue.
In sum, Vam7p rebinds to vacuoles via the PX domain. Our data show that PtdIns 3-P plays an important role in the assembly of Vam7p with the SNARE complex prior to fusion.
Discussion
The SNAP-23–like protein Vam7p is a mobile SNARE Fig. 7). Vam7p is initially part of the cis-SNARE complex and is released from the vacuoles as a monomer during priming on ice (Fig. 1, C and D). The recruitment requires a specific PtdIns 3-P acceptor site and occurs via the NH2-terminal PX domain (Figs. 4, 5, and 6). Finally, the released Vam7p is functional and can rescue a fusion reaction. Our data show that Vam7p associates with the vacuoles via a lipid-defined acceptor site and can act as a limiting factor for vacuole fusion controlled by the status of lipid phosphorylation on the vacuole.
Several aspects of the fusion reaction are highlighted by these observations. Priming on ice reveals that Vam7p, Ykt6p, Nyv1p, and the α-SNAP homologue Sec17p disassemble efficiently from Vam3p and Vti1p. This disassembly does not include Vti1p and Vam3p. The dissociation of Vam3p and Vti1p appears to require elevated temperature but not Sec18p (unpublished data; Ungermann et al., 1999). Therefore, we believe that priming occurs in two steps, the ATP-dependent release of Vam7p, Sec17p, Nyv1p, and Ykt6p, and the dissociation of the Vam3p-Vti1p complex. The reactions may occur fast under normal conditions, but consecutive dissociation might be needed to orchestrate defined protein–protein interactions required for tethering.
The priming-dependent release of Vam7p on ice was unexpected but is not without precedent. SNAP-23, the eukaryotic homologue, is recruited to secretory granules before their exocytic fusion at the mast cell plasma membrane (Guo et al., 1998). Interestingly, the authors take advantage of a similar behavior, namely that SNAP-23 can be recruited from distant sites to the vesicles before fusion, by incubating the cells on ice with ATP. Later, it was demonstrated that the cytoskeletal protein vimentin binds SNAP-23 and serves as an essential pool of this SNARE (Faigle et al., 2000). When cells are treated with the NSF inhibitor NEM, SNAP-23 is not found in association with vimentin but is found in association with the SNARE complex. In yeast, Sato et al. (1998) showed that Vam7p also exists in pools in the cytosol and on the vacuole. If the NSF homologue Sec18p is inactivated in a temperature-sensitive mutant, Vam7p is mostly found in association with the vacuole membrane (Sato et al., 1998). Therefore, it was assumed that SNARE complex disassembly and Vam7p partitioning were connected. We were able to show the complete cycle in a defined in vitro system. Vam7p is released from the cis-SNARE complex during the priming reaction and is recruited via PtdIns 3-P to the vacuoles as a prerequisite for efficient fusion (Figs. 1 and 2).
The released Vam7p is a functional intermediate. First, vacuoles depleted of Vam7p in a preincubation can fuse again if supplemented with a soluble pool of Vam7p (Fig. 2 B). Second, if vacuoles are poisoned with antibodies after the initial priming and Vam7p release, fusion is only possible if Vam7p is provided as a soluble pool (Fig. 2 C). Vam7p can rescue the reaction at several points during the fusion reaction. Initial data suggested that the association of Vam7p with the Rab GTPase Ypt7p was essential for Vam7p retrieval on vacuoles (Ungermann et al., 2000). This was supported by the interaction of Vam7p and Ypt7p in a two-hybrid screen (Uetz et al., 2000). A few issues are still puzzling. First, recruitment of Vam7p is possible if Ypt7p has been removed (Fig. 6 E) or blocked by antibodies (unpublished data), though the recruited Vam7p dissociates from the vacuole when Ypt7p is removed from the membrane (Fig. 6 E). Second, vacuoles lacking Ypt7p still serve as acceptor vacuoles, albeit with reduced efficiency (unpublished data). However, these vacuoles may still contain late endosome markers and therefore could have other binding sites for Vam7p. Thus, the role of Ypt7p depends on whether Vam7p is transferred directly from the SNARE complex or is recruited from the cytosol. We do not know the basis for this difference, but we can conclude that tethering does not depend on Vam7p, whereas fusion does depend on Vam7p (Fig. 2, C and D). Thus, Vam7p can enter the reaction at several stages. In this respect, our observation is comparable to the findings of Scheller and colleagues who were able to block exocytosis in PC12 cells by adding botulinum toxin and rescued this block by readdition of the removed peptide (Chen et al., 1999). However, our rescue requires an intact protein that cycles between vacuoles and cytosol in vivo and in vitro.
Our data clearly show that PtdIns 3-P is the essential phosphoinositide required for Vam7p binding. Most PtdIns 3-P is present already on the isolated vacuole, unless Vps34p was inactivated before the vacuole purification (Fig. 5 A). It appears though that synthesis of PtdIns 3-P is also occurring on the vacuole, since some wild-type vacuoles are more sensitive to wortmannin (Fig. 5 A). It is known that Rab proteins bind a variety of effectors (Zerial and McBride, 2001). Zerial and coworkers demonstrated that several proteins are complexed with the Rab5 GTPase in its active GTP-bound stage (Christoforidis et al., 1999a). The lipid kinase hVps34, which was found in the isolate, was shown to be essential for endosome fusion (Christoforidis et al., 1999b). Our data show that the PtdIns 3-P is required for efficient fusion and depends on Vps34p (Fig. 5).
We used two defined domains to map the PtdIns 3-P site on the vacuole, the PX domain of Vam7p and the FYVE domain of Hrs (Gillooly et al., 2000). Both domains interact with PtdIns-3 P and inhibit the recruitment of Vam7p similarly. Interestingly, the PX domain of Vam7p inhibited the fusion assay later than the FYVE domain does and later than the antibody to Vam7p, which was raised to the COOH terminus, not to the PX domain. How do we explain these discrepancies? An antibody binds the respective antigen and inhibits its function. The PX domain binds PtdIns 3-P tightly and therefore blocks the binding site for Vam7p. It may also interact with proteins at the docking site. Although the FYVE domain (from mammalian Hrs) can do the former, it will most likely not bind yeast vacuole proteins in a specific manner. Therefore, we conclude that the Vam7p PX domain targets Vam7p precisely to the fusion site to execute its function.
We propose that a defined lipid domain is present during docking to allow the association of Vam7p and possibly other factors with the vacuole. This is supported by our observations that the PX domain is essential for targeting Vam7p to the fusion site (Fig. 4) and by the binding and release of Vam7p in the presence of Gdi1p (Fig. 6 E). Furthermore, the COOH-terminal domain of Vam7p is not sufficient to rescue the reaction, suggesting that a lipid association via the PX domain is a prerequisite for the migration into the fusion site and supports our idea of a lipid domain present at this site. It appears that this requirement can be bypassed in vivo by the overexpression of the Vam7p coiled-coil domain (Cheever et al., 2001). However, the strong effects observed in our defined in vitro reaction suggest that cells have selected a more complex pathway to regulate the fusion reaction by setting up a defined lipid domain to target a SNARE to its place of function.
Materials and methods
Reagents and strains
Chemicals were purchased from Sigma-Aldrich unless noted otherwise. Antibodies to Vam7p were raised in White New Zealand rabbits against an NH2-terminal (amino acid 1–122) and a GST-tagged COOH-terminal fragment (amino acids 165–316) of Vam7p purified from Escherichia coli.
The yeast strains used in this study were as described (Haas et al., 1994). The COOH-terminal tagging of Vam7p with protein A or HA was done by transformation of PCR fragments containing the inserts and a TRP1 marker (Knop et al., 1999). Colonies that grew on SC-trp plates were restreaked and analyzed for a shift in the molecular weight with antibodies to Vam7p.
Biochemical techniques
GST-tagged N- (Cheever et al., 2001) and COOH-terminal fragments (amino acid 165–316) of Vam7p were expressed in BL21 E. coli cells and induced with 0.1 mM IPTG for 1 h in the presence of 1 mM PMSF (N-terminal fragments) or with 1 mM IPTG for 3 h. Cells were harvested, sonicated in the presence of 50 mM Tris, pH 7.4, and 150 mM NaCl and immediately purified via glutathione beads (Amersham Pharmacia Biotech). Fragments were cleaved from the GST by thrombin, and the protease was removed by addition of benzamidine agarose (Sigma-Aldrich). Antibodies to Vam7p were purified on CNBr-coupled GST-Vam7p (165–316) (Mayer et al., 1996). Recombinant mammalian Hrs GST-2xFYVE and the C215S mutant were expressed in E. coli BL21 cells (Stratagene), from a plasmid provided by H. Stenmark (Institute of Cancer Research, Oslo, Norway), and enriched on glutathione-sepharose to >90% purity as described (Gillooly et al., 2000).
Vacuole purification and wortmannin pretreatment
Vacuoles were purified by lyticase treatment and DEAE lysis as described (Haas et al., 1994). Wortmannin pretreatment of spheroplasts was performed as follows. Wortmannin (Sigma-Aldrich) was dissolved in DMSO immediately before use. Wortmannin (50 μM final) was added to cells suspended in spheroplasting buffer, and the suspension was incubated for 5 min at 30°C, then oxalyticase enzyme was added, and the suspension was incubated at 30°C for additional 25 min before fractionation. Vacuoles were obtained after this treatment at normal yield. Vam7p levels were similar or slightly lower (<50% reduction).
Vacuole fusion assay
Vacuole fusion is measured by a biochemical complementation assay (Wickner and Haas, 2000). Standard fusion reactions were done in reaction buffer (10 mM PIPES/KOH, pH 6.8, 120 mM KCl, 5 mM MgCl2, 200 mM sorbitol) containing an ATP-regenerating system (0.5 mM ATP, 40 mM creatine phosphate, 0.1 mg/ml creatine kinase), a protease inhibitor cocktail, 10 μM CoA, and 1 μg/ml His6-Sec18p.
Acknowledgments
We are grateful to Scott Emr, Trey Sato, Harald Stenmark, and Bill Wickner for plasmids and advice, and members of the Ungermann laboratory for comments.
This work was supported by the Boehringer Ingelheim Fonds (to C. Boeddinghaus), the Cancer Research Fund of the Damon Runyon-Walter Winchell Foundation (to A.J. Merz), the Deutsche Forschungsgemeinschaft (UN111/2-2), and the National Institutes of Health (GM23377).
Rico Laage's present address is Axaron Bioscience AG, Im Neuenheimer Feld 515, 69120 Heidelberg, Germany.
Footnotes
Abbreviations used in this paper: PtdIns, phosphatidylinositol; PtdIns 3-P, PtdIns 3-phosphate.
References
- Burd, C.G., and S.D. Emr. 1998. Phosphatidylinositol(3)-phosphate signaling mediated by specific binding to RING FYVE domains. Mol. Cell. 2:157–162. [DOI] [PubMed] [Google Scholar]
- Cao, X., N. Ballew, and C. Barlowe. 1998. Initial docking of ER-derived vesicles requires Uso1p and Ypt1p but is independent of SNARE proteins. EMBO J. 17:2156–2165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheever, M.L., T.K. Sato, T. de Beer, T.G. Kutateladze, S.D. Emr, and M. Overduin. 2001. Phox domain interaction with PtdIns(3)P targets the Vam7 t-SNARE to vacuole membranes. Nat. Cell Biol. 3:613–618. [DOI] [PubMed] [Google Scholar]
- Chen, Y.A., and R.H. Scheller. 2001. SNARE-mediated membrane fusion. Nat. Rev. Mol. Cell Biol. 2:98–106. [DOI] [PubMed] [Google Scholar]
- Chen, Y.A., S.J. Scales, S.M. Patel, Y.C. Doung, and R.H. Scheller. 1999. SNARE complex formation is triggered by Ca2+ and drives membrane fusion. Cell. 97:165–174. [DOI] [PubMed] [Google Scholar]
- Christoforidis, S., H.M. McBride, R.D. Burgoyne, and M. Zerial. 1999. a. The Rab5 effector EEA1 is a core component of endosome docking. Nature. 397:621–625. [DOI] [PubMed] [Google Scholar]
- Christoforidis, S., M. Miaczynska, K. Ashman, M. Wilm, L. Zhao, S.C. Yip, M.D. Waterfield, J.M. Backer, and M. Zerial. 1999. b. Phosphatidylinositol-3-OH kinases are Rab5 effectors. Nat. Cell Biol. 1:249–252. [DOI] [PubMed] [Google Scholar]
- Conradt, B., J. Shaw, T. Vida, S. Emr, and W. Wickner. 1992. In vitro reactions of vacuole inheritance in Saccharomyces cerevisiae. J. Cell Biol. 119:1469–1479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cutler, N.S., J. Heitman, and M.E. Cardenas. 1997. STT4 is an essential phosphatidylinositol 4-kinase that is a target of wortmannin in Saccharomyces cerevisiae. J. Biol. Chem. 272:27671–27677. [DOI] [PubMed] [Google Scholar]
- Eitzen, G., N. Thorngren, and W. Wickner. 2001. Rho1p and Cdc42p act after Ypt7p to regulate vacuole docking. EMBO J. 20:5650–5656. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Faigle, W., E. Colucci-Guyon, D. Louvard, S. Amigorena, and T. Galli. 2000. Vimentin filaments in fibroblasts are a reservoir for SNAP23, a component of the membrane fusion machinery. Mol. Biol. Cell. 11:3485–3494. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gillooly, D.J., I.C. Morrow, M. Lindsay, R. Gould, N.J. Bryant, J.M. Gaullier, R.G. Parton, and H. Stenmark. 2000. Localization of phosphatidylinositol 3-phosphate in yeast and mammalian cells. EMBO J. 19:4577–4588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guo, Z., C. Turner, and D. Castle. 1998. Relocation of the t-SNARE SNAP-23 from lamellipodia-like cell surface projections regulates compound exocytosis in mast cells. Cell. 94:537–548. [DOI] [PubMed] [Google Scholar]
- Haas, A., B. Conradt, and W. Wickner. 1994. G-protein ligands inhibit in vitro reactions of vacuole inheritance. J. Cell Biol. 126:87–97. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holthuis, J.C., B.J. Nichols, S. Dhruvakumar, and H.R. Pelham. 1998. Two syntaxin homologues in the TGN/endosomal system of yeast. EMBO J. 17:113–126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jahn, R., and T.C. Südhof. 1999. Membrane fusion and exocytosis. Annu. Rev. Biochem. 68:863–911. [DOI] [PubMed] [Google Scholar]
- Kato, M., and W. Wickner. 2001. Ergosterol is required for the Sec18/ATP-dependent priming step of homotypic vacuole fusion. EMBO J. 20:4035–4040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kihara, A., T. Noda, N. Ishihara, and Y. Ohsumi. 2001. Two distinct Vps34 phosphatidylinositol 3-kinase complexes function in autophagy and carboxypeptidase Y sorting in Saccharomyces cerevisiae. J. Cell Biol. 152:519–530. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knop, M., K. Siegers, G. Pereira, W. Zachariae, B. Winsor, K. Nasmyth, and E. Schiebel. 1999. Epitope tagging of yeast genes using a PCR-based strategy: more tags and improved practical routines. Yeast. 15:963–972. [DOI] [PubMed] [Google Scholar]
- Kutateladze, T.G., K.D. Ogburn, W.T. Watson, T. de Beer, S.D. Emr, C.G. Burd, and M. Overduin. 1999. Phosphatidylinositol 3-phosphate recognition by the FYVE domain. Mol. Cell. 3:805–811. [DOI] [PubMed] [Google Scholar]
- Mayer, A., W. Wickner, and A. Haas. 1996. Sec18p (NSF)-driven release of Sec17p (alpha-SNAP) can precede docking and fusion of yeast vacuoles. Cell. 85:83–94. [DOI] [PubMed] [Google Scholar]
- Mayer, A., D. Scheglmann, S. Dove, A. Glatz, W. Wickner, and A. Haas. 2000. Phosphatidylinositol 4,5-bisphosphate regulates two steps of homotypic vacuole fusion. Mol. Biol. Cell. 11:807–817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McBride, H.M., V. Rybin, C. Murphy, A. Giner, R. Teasdale, and M. Zerial. 1999. Oligomeric complexes link Rab5 effectors with NSF and drive membrane fusion via interactions between EEA1 and syntaxin 13. Cell. 98:377–386. [DOI] [PubMed] [Google Scholar]
- Mellman, I., and G. Warren. 2000. The road taken: past and future foundations of membrane traffic. Cell. 100:99–112. [DOI] [PubMed] [Google Scholar]
- Müller, O., D.I. Johnson, and A. Mayer. 2001. Cdc42p functions at the docking stage of yeast vacuole membrane fusion. EMBO J. 20:5657–5665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nielsen, E., S. Christoforidis, S. Uttenweiler-Joseph, M. Miaczynska, F. Dewitte, M. Wilm, B. Hoflack, and M. Zerial. 2000. Rabenosyn-5, a novel Rab5 effector, is complexed with hVPS45 and recruited to endosomes through a FYVE finger domain. J. Cell Biol. 151:601–612. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Otto, H., P.I. Hanson, and R. Jahn. 1997. Assembly and disassembly of a ternary complex of synaptobrevin, syntaxin, and SNAP-25 in the membrane of synaptic vesicles. Proc. Natl. Acad. Sci. USA. 94:6197–6201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peters, C., and A. Mayer. 1998. Ca2+/calmodulin signals the completion of docking and triggers a late step of vacuole fusion. Nature. 396:575–580. [DOI] [PubMed] [Google Scholar]
- Peters, C., P.D. Andrews, M.J. Stark, S. Cesaro-Tadic, A. Glatz, A. Podtelejnikov, M. Mann, and A. Mayer. 1999. Control of the terminal step of intracellular membrane fusion by protein phosphatase 1. Science. 285:1084–1087. [DOI] [PubMed] [Google Scholar]
- Peters, C., M.J. Bayer, S. Buhler, J.S. Andersen, M. Mann, and A. Mayer. 2001. Trans-complex formation by proteolipid channels in the terminal phase of membrane fusion. Nature. 409:581–588. [DOI] [PubMed] [Google Scholar]
- Peterson, M.R., C.G. Burd, and S.D. Emr. 1999. Vac1p coordinates Rab and phosphatidylinositol 3-kinase signaling in Vps45p-dependent vesicle docking/fusion at the endosome. Curr. Biol. 9:159–162. [DOI] [PubMed] [Google Scholar]
- Price, A., D. Seals, W. Wickner, and C. Ungermann. 2000. The docking stage of yeast vacuole fusion requires the transfer of proteins from a cis-SNARE complex to a Rab/Ypt protein. J. Cell Biol. 148:1231–1238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rothman, J.E. 1994. Mechanisms of intracellular protein transport. Nature. 372:55–63. [DOI] [PubMed] [Google Scholar]
- Sato, T.K., T. Darsow, and S.D. Emr. 1998. Vam7p, a SNAP-25-like molecule, and Vam3p, a syntaxin homolog, function together in yeast vacuolar protein trafficking. Mol. Cell. Biol. 18:5308–5319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato, T.K., M. Overduin, and S.D. Emr. 2001. Location, location, location: membrane targeting directed by PX domains. Science. 294:1881–1885. [DOI] [PubMed] [Google Scholar]
- Simonsen, A., A.E. Wurmser, S.D. Emr, and H. Stenmark. 2001. The role of phosphoinositides in membrane transport. Curr. Opin. Cell Biol. 13:485–492. [DOI] [PubMed] [Google Scholar]
- Söllner, T., S.W. Whiteheart, M. Brunner, H. Erdjument-Bromage, S. Geromanos, P. Tempst, and J.E. Rothman. 1993. SNAP receptors implicated in vesicle targeting and fusion. Nature. 362:318–324. [DOI] [PubMed] [Google Scholar]
- Stack, J.H., D.B. DeWald, K. Takegawa, and S.D. Emr. 1995. Vesicle-mediated protein transport: regulatory interactions between the Vps15 protein kinase and the Vps34 PtdIns 3-kinase essential for protein sorting to the vacuole in yeast. J. Cell Biol. 129:321–334. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stenmark, H., R. Aasland, B.H. Toh, and A. D'Arrigo. 1996. Endosomal localization of the autoantigen EEA1 is mediated by a zinc-binding FYVE finger. J. Biol. Chem. 271:24048–24054. [DOI] [PubMed] [Google Scholar]
- Sutton, R.B., D. Fasshauer, R. Jahn, and A.T. Brunger. 1998. Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4 A resolution. Nature. 395:347–353. [DOI] [PubMed] [Google Scholar]
- Uetz, P., L. Giot, G. Cagney, T.A. Mansfield, R.S. Judson, J.R. Knight, D. Lockshon, V. Narayan, M. Srinivasan, P. Pochart, et al. 2000. A comprehensive analysis of protein-protein interactions in Saccharomyces cerevisiae. Nature. 403:623–627. [DOI] [PubMed] [Google Scholar]
- Ungermann, C., and W. Wickner. 1998. Vam7p, a vacuolar SNAP-25 homolog, is required for SNARE complex integrity and vacuole docking and fusion. EMBO J. 17:3269–3276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungermann, C., B.J. Nichols, H.R. Pelham, and W. Wickner. 1998. a. A vacuolar vt-SNARE complex, the predominant form in vivo and on isolated vacuoles, is disassembled and activated for docking and fusion. J. Cell Biol. 140:61–69. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungermann, C., K. Sato, and W. Wickner. 1998. b. Defining the functions of trans-SNARE pairs. Nature. 396:543–548. [DOI] [PubMed] [Google Scholar]
- Ungermann, C., G.F. von Mollard, O.N. Jensen, N. Margolis, T.H. Stevens, and W. Wickner. 1999. Three v-SNAREs and two t-SNAREs, present in a pentameric cis-SNARE complex on isolated vacuoles, are essential for homotypic fusion. J. Cell Biol. 145:1435–1442. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ungermann, C., A. Price, and W. Wickner. 2000. A new role for a SNARE protein as a regulator of the Ypt7/Rab-dependent stage of docking. Proc. Natl. Acad. Sci. USA. 97:8889–8891. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Veit, M., R. Laage, L. Dietrich, L. Wang, and C. Ungermann. 2001. Vac8p release from the SNARE complex and its palmitoylation are coupled and essential for vacuole fusion. EMBO J. 20:3145–3155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walch-Solimena, C., J. Blasi, L. Edelmann, E.R. Chapman, G.F. von Mollard, and R. Jahn. 1995. The t-SNAREs syntaxin 1 and SNAP-25 are present on organelles that participate in synaptic vesicle recycling. J. Cell Biol. 128:637–645. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Weber, T., B.V. Zemelman, J.A. McNew, B. Westermann, M. Gmachl, F. Parlati, T.H. Sollner, and J.E. Rothman. 1998. SNAREpins: minimal machinery for membrane fusion. Cell. 92:759–772. [DOI] [PubMed] [Google Scholar]
- Whyte, J.R., and S. Munro. 2001. The Sec34/35 Golgi transport complex is related to the exocyst, defining a family of complexes involved in multiple steps of membrane traffic. Dev. Cell. 1:527–537. [DOI] [PubMed] [Google Scholar]
- Wickner, W., and A. Haas. 2000. Yeast homotypic vacuole fusion: a window on organelle trafficking mechanisms. Annu. Rev. Biochem. 69:247–275. [DOI] [PubMed] [Google Scholar]
- Yamamoto, A., D.B., DeWald, I.V. Boronenkov, R.A. Anderson, S.D. Emr, and D. Koshland. 1995. Novel PI(4)P 5-kinase homologue, Fab1p, essential for normal vacuole function and morphology in yeast. Mol. Biol. Cell. 6:525–539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zerial, M., and H. McBride. 2001. Rab proteins as membrane organizers. Nat. Rev. Mol. Cell Biol. 2:107–117. [DOI] [PubMed] [Google Scholar]