

Abstract
Many cellular components are transported using a combination of the actin- and microtubule-based transport systems. However, how these two systems work together to allow well-regulated transport is not clearly understood. We investigate this question in the Xenopus melanophore model system, where three motors, kinesin II, cytoplasmic dynein, and myosin V, drive aggregation or dispersion of pigment organelles called melanosomes. During dispersion, myosin V functions as a “molecular ratchet” to increase outward transport by selectively terminating dynein-driven minus end runs. We show that there is a continual tug-of-war between the actin and microtubule transport systems, but the microtubule motors kinesin II and dynein are likely coordinated. Finally, we find that the transition from dispersion to aggregation increases dynein-mediated motion, decreases myosin V–mediated motion, and does not change kinesin II–dependent motion. Down-regulation of myosin V contributes to aggregation by impairing its ability to effectively compete with movement along microtubules.
Keywords: dynein; kinesin II; myosin V; melanophore; organelle transport
Introduction
Many vesicles and other cellular cargos require transport to different cellular destinations at specific times. To accomplish this task, cells employ molecular motors and two sets of directional polymer tracks: microtubules and actin filaments. These tracks are potentially complimentary; although microtubules in animal cells are typically arranged radially, actin filaments are found mostly in a randomly oriented meshwork. Indeed, utilization of both systems has been observed for vesicles in squid axoplasm (Kuznetsov et al., 1992), mitochondria (Morris and Hollenbeck, 1995; Krendel et al., 1998); ER-derived vesicles (Tabb et al., 1998), cytoplasmic particles in Drosophila embryos (Mermall et al., 1994), postendocytic (Maples et al., 1997) or secretory vesicles (Hirschberg et al., 1998), and pigment organelles (Rodionov et al., 1998; Rogers and Gelfand, 1998; Wu et al., 1998; for review see Brown, 1999).
The reason for a single cargo to employ both types of transport is not clear. One attractive hypothesis is that microtubules are used for rapid long distance travel, whereas actin tracks serve for local transport to locations not accessible to the microtubules (Langford, 1995). A second model is that the microtubule-based transport system provides the majority of the transport, while the actin cytoskeleton anchors the cargos in the appropriate location, once they are delivered there through microtubule transport (Wu et al., 1998). Both models seem compatible with a coordinated transfer from the microtubules to actin. Biochemical studies have established direct interactions between myosin and microtubule motors that could in principle help to mediate such a transfer (Huang et al., 1999).
Contrary to such sequential activity models, other work suggests that both systems may be active simultaneously, engaged in a tug-of-war where each class of motor tries to move along its respective polymer. The result of this competition is to modulate each other's function. In support of this theory, biochemical and immunolocalization studies indicate that in many cases both actin and microtubule motors are present on the same cargos at the same time (Tabb et al., 1998). Further, impairment of myosin V activity increases microtubule-based organelle transport in mouse melanocytes and neurons, as judged by comparing motion in cells derived from wild-type versus dilute (lacking myosin Va) mice (Wu et al., 1998; Bridgman, 1999).
How a cell controls transport will be determined by the type and extent of interactions between different transport systems: if there is a coordinated transfer of an organelle from actin to microtubules (or vice versa), the transfer itself can be regulated to determine which cytoskeletal track is used. In contrast, if there exists a continual tug-of-war, regulation would alter how likely one set of motors was to win over the other.
We have chosen to investigate the question of motor interactions and transport regulation in Xenopus melanophores. In this study, we use quantitative analysis of organelle movement in different genetic backgrounds to determine if microtubule- and actin-based transport functions independently or in concert. We also investigated how the two transport systems are modulated by physiological signals in order to achieve regulated intracellular transport.
In melanophores, pigment granules have been shown to move using a combination of kinesin II, dynein, and myosin V (Rogers and Gelfand, 1998; Tuma et al., 1998; unpublished data), and there is unsurpassed ability to manipulate organelle transport. Xenopus melanophores disperse pigment organelles (melanosomes) throughout the cytoplasm in response to melanocyte-stimulating hormone (MSH)* and aggregate them in response to melatonin. The action of both hormones is mediated by cAMP. MSH increases cytoplasmic cAMP, whereas melatonin decreases it (Daniolos et al., 1990). Furthermore, dominant negative constructs can be used to selectively block either kinesin II or myosin V activity (Tuma et al., 1998; Rogers et al., 1999), allowing the investigation of the role of these motors in organelle transport and their functional interactions. These constructs can be used for disrupting motor activity for two reasons. First, they lack a motor domain and cannot bind to the actin filaments or microtubules and thus interfere with activity of other motors. Second, because they have the stalk and tail domains they can presumably be incorporated into any complexes in a wild-type manner. Our previous work (Tuma et al., 1998; Rogers et al., 1999) showed that inhibition of either kinesin II or myosin V impairs the ability of the cells to achieve pigment dispersion; however, the individual contribution of each motor to the common task of pigment dispersion remained unknown.
Here we have combined dominant negative manipulations with a biophysical study of transport. Individual melanosome tracking and movement analysis were used to quantify how the motion of pigment granules changed due to either regulation by hormones or motor inactivation. This approach allowed us to answer two questions: (1) what properties of motor activity are regulated to result in net dispersion or net aggregation? and (2) what types of functional interactions are there between the microtubule and actin motors, and are these interactions regulated?
Results
Phenylthiourea treatment allows quantitative characterization of individual melanosome motion
To understand the role of individual motors in organelle transport and how transport is regulated during aggregation versus dispersion, we combined tracking of individual melanosomes with quantitative analysis of movement. We compared the motion in wild-type, kinesin II dominant negative and myosin V dominant negative backgrounds.
It is impossible to follow individual melanosomes in regular melanophores because of the high spatial density of organelles (Fig. 1 A). To solve this problem, we reduced the number of pigment granules by treating cells with the tyrosinase inhibitor phenylthiourea (PTU). Because tyrosinase is a key enzyme in melanin biosynthesis, PTU inhibited formation of new melanosomes: after several passages in the presence of PTU, cells become completely devoid of pigment (Fig. 1 B). PTU is not toxic, and melanophores can proliferate indefinitely (≥100 passages) in its presence.
Figure 1.
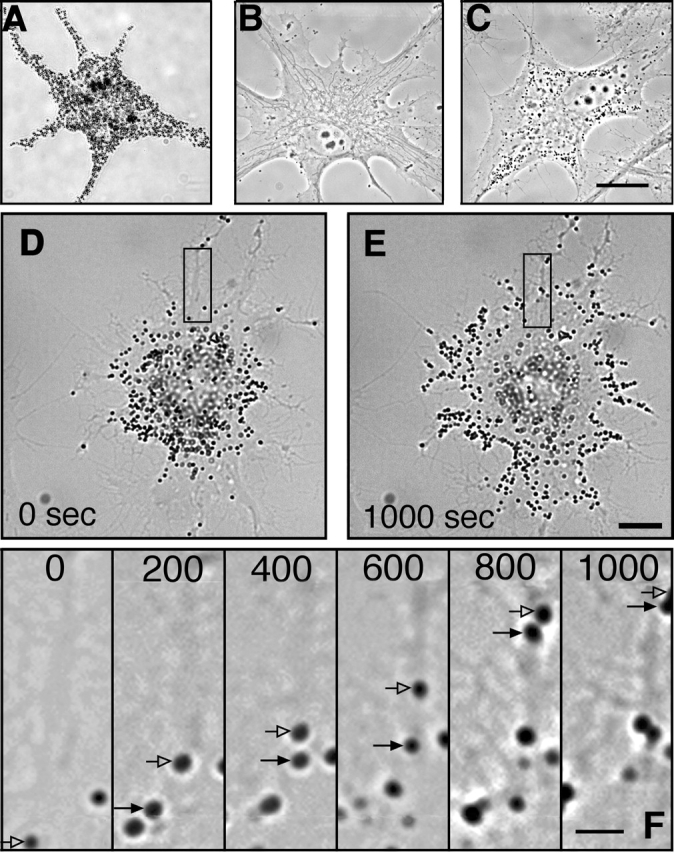
Tracking organelles in cells with a limited number of melanosomes. (A) Cultured Xenopus melanophore. The large number of melanosomes makes tracking individual organelles impossible. (B) Cell after 4 wk of culture in 1 mM PTU. No melanosomes are seen in the cytoplasm. (C) Melanophore after 24 h recovery from PTU has a small number of melanosomes. (D and E) Melanophore after 24 h recovery from PTU before (D) and 1000 s after (E) addition of MSH, showing that the melanosomes in PTU-treated cells disperse in response to MSH. (F) Sequence of images taken during pigment dispersion of the cell shown in D and E. Arrows show positions of two melanosomes in subsequent frames. Time after addition of MSH (in seconds) is shown on each frame. Bright-field microscopy (A and D–F); phase–contrast microscopy (B and C). Bars: (A–E), 10 μm; (F) 2.5 μm.
The effect of PTU is completely reversible, since removal of the inhibitor from the medium allows recovery of melanin synthesis and melanosome biogenesis. After 24–48 h without PTU, typical cells contain only ∼100 melanosomes (Fig. 1 C). These melanosomes undergo normal aggregation and dispersion (Fig. 1, D and E) and can be easily tracked by eye (Fig. 1 F) and automatically using image processing software. All the quantitative studies reported here examined motion in such cells.
Myosin V and kinesin II are required for pigment dispersion
Knowing that pigment dispersion requires both kinesin II and myosin V, we sought first to determine the role of these motors in the process. Since microtubules are relatively straight and radially oriented (Fig. 2 A) and actin filaments tend to be more randomly organized (Fig. 2 B), one would intuitively expect that in vivo motion driven by microtubule motors would be linear and radial, whereas motion driven by myosin V would appear less linear and more random. This appears to be the case; in general, trajectories of individual melanosomes are more linear in latrunculin-treated cells or cells expressing the myosin V dominant negative construct than in nocodazole-treated cells (Fig. 3, A and B) . We confirmed that long linear tracks correspond to microtubule-based motion by tracking melanosome movement in cells microinjected with fluorescent tubulin and stimulated to disperse pigment 60 min after injection. Overlay of melanosomes trajectories over one of the microtubule images (Fig. 3 C) shows that long linear movements of organelles correspond to microtubule profiles, confirming that these movements are indeed microtubule based. As expected, outward linear trajectories are absent predominantly in the kinesin II dominant negative cells. Inward linear trajectories are observed in both the kinesin II and the myosin V dominant negative cells, suggesting that the use of the dominant negative constructs to inhibit kinesin II or myosin V activity does not impair cytoplasmic dynein.
Figure 2.

Distribution of microtubules and actin filaments in Xenopus melanophores. (A) Microtubules stained by a tubulin antibody. Note radial pattern of distribution. (B) Actin filament distribution as revealed by rhodamine-phalloidin staining. In addition to thin bundles, most of the staining is distributed diffusely. Since rhodamine-phalloidin does not bind nonpolymerized actin, diffuse staining suggests random arrangement of actin filaments. Bar, 20 μm.
Figure 3.
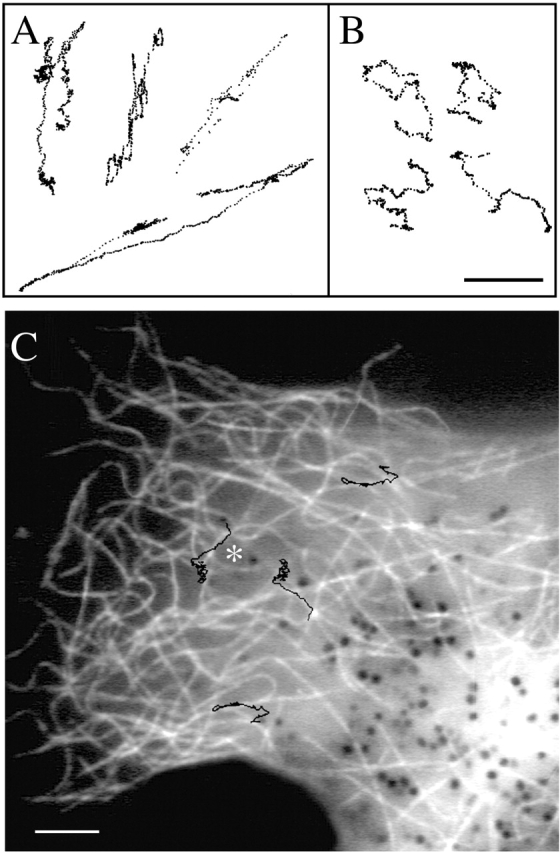
Linear melanosome tracks correlate with the position of microtubules. Movement of melanosomes in cells transfected with dominant negative myosin V (A) or treated with nocodazole (B). Note that in A melanosomes move along linear tracks, whereas movement in B is more random. Position of melanosomes was registered at 30 Hz, but for clarity only every sixth frame is displayed. Bar, 1 μm. (C) Overlay of melanosome tracks and fluorescently labeled microtubules. Microtubules were labeled by injecting cells with Alexa 488 tubulin. Movement of melanosomes was tracked using a series of bright-field images taken every 3 s. The resulting tracks were overlayed onto a fluorescent image showing the microtubule distribution. Note that linear movements correlate with the position of microtubules. The track labeled with an asterisk does not exactly overlap with the image of the underlying microtubule because this microtubule moved substantially in the course of the tracking. Bar, 5 μm.
We first determined how well the dominant negative constructs block the activity of their targets. In kinesin II/myosin V double dominant negative cells, most melanosomes are aggregated in the cell center. The few individual granules not in the aggregated “clump” also show dramatically reduced motion (unpublished data). Thus, the majority of outward motion is blocked by the combined dominant negative constructs. This is consistent with our previous characterization (Tuma et al., 1998; Rogers et al., 1999).
Because we can use nocodazole to depolymerize all the microtubules, it is possible to directly observe motion due entirely to myosin V activity (Fig. 4) . Thus, we could directly quantify the magnitude of the myosin V dominant negative's effect: we tracked the motion of individual melanosomes and calculated the average distance from the point we started, tracking the melanosome as a function of time, that is, r2(t) (see Materials and methods for details). In nocodazole-treated cells, the r2(t) analysis indicates robust myosin V activity, but this activity completely disappears in myosin V dominant negative cells treated with nocodazole (Fig. 4). Thus, the dominant negative construct fully blocks myosin V function. In addition, the negligible residual r2(t) indicates both that nocodazole does indeed eliminate microtubule-based motion and that passive diffusion of melanosomes is insignificant. Identical results were obtained with a different dominant negative myosin V construct that encodes only the globular portion of myosin V tail and is thus incapable of dimerization (unpublished data). This result suggests that inhibition of myosin V motility by dominant negative constructs is due to competition of the truncated protein with the full-length myosin V for binding to organelles.
Figure 4.

Quantification of displacement generated by myosin V. The r2(t) plots show the square of the average displacement of the pigment granules as a function of time. Each curve represents an average of ∼100 plots for individual moving granules (∼25 melanosomes chosen at random in four different cells). The indicated error bars are the standard error of the mean. Nocodazole was used to depolymerize microtubules in order to quantify myosin V (M-V)–driven motion. The decrease in motion in aggregating versus dispersing cells indicates that myosin V–driven motion is down-regulated as part of the dispersion to aggregation transition. The lowest curve shows that in the absence of microtubules expression of the myosin V dominant negative construct completely eliminates motion, demonstrating the efficacy of the construct. The magnitude of dispersion in a wild-type background driven by kinesin II, dynein, and myosin V (top curve) is similar to dispersion driven only by myosin V (second curve).
Regulation occurs by changes in dynein- and myosin-based transport
The ability to independently quantify the actin- or microtubule-based components of motion allowed us to investigate how the cell regulates motor activity resulting in net aggregation and dispersion. To quantify the microtubule component of motion, we used analysis software (Gross et al., 2000) to resolve the long linear paths into individual “runs,” that is, periods of uninterrupted plus or minus end motion. This data was analyzed using two complementary approaches: the mean travel distance was determined using a modified averaging procedure (Figs. 5 and 6) , whereas the properties of long runs were examined by fitting the distribution histogram (Fig. 7 and Table I; see Materials and methods). Initially, we used the dominant negative myosin V construct (Rogers et al., 1999) to block myosin V activity and looked at the lengths and velocities of plus and minus end runs in cells treated with MSH or melatonin. Comparing these measurements, we found that the mean outward (plus end) run length is constant, regardless of the state of the cells (i.e., MSH versus melatonin stimulation) (Fig. 5 A and Table I), whereas the mean inward (minus end) run length changes significantly between MSH- and melatonin-treated cells (Fig. 5 A). Thus, one of the mechanisms that cells use to shift transport from net aggregation to net dispersion is to alter the length of minus end (dynein-powered) runs, while keeping plus end runs constant.
Figure 5.

Regulation of microtubule-based motion. To investigate regulation of microtubule motors, their activity was studied in the absence of functional myosin V. Shown are (modified) averages of travel distances (as described in Materials and methods) and velocities of plus and minus end runs. A run was defined as a period of uninterrupted motion along the microtubule (as described in Materials and methods). (A) The change from dispersion to aggregation coincides with a change in the length of (long-type) minus end runs, whereas the length of plus end runs remains constant. (B) The average velocity of (short-type) plus and minus end runs. There is no significant change in run velocity in either direction due to the change from dispersion to aggregation. These data show an average of ∼400 individual runs from four or more cells. M-V, myosin V. The error bars are the standard error of the mean.
Figure 6.
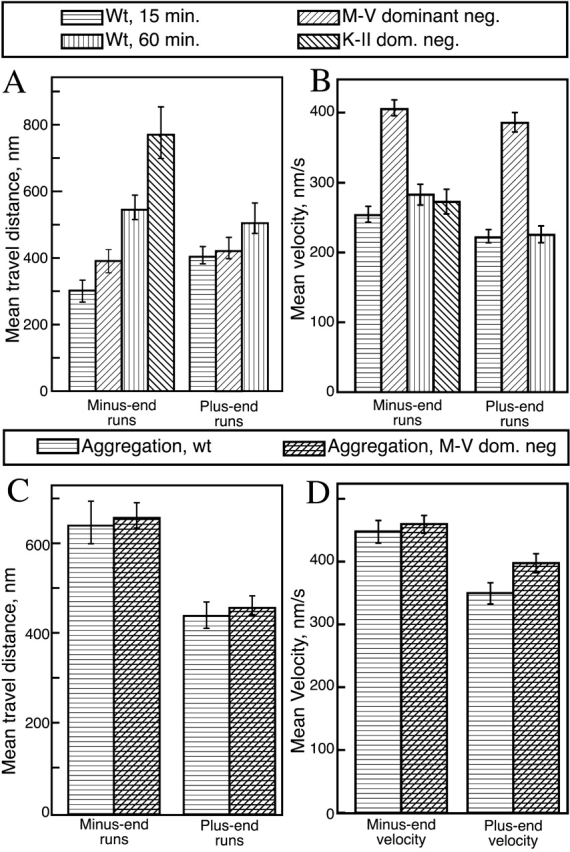
Microtubule motion as a function of time and as altered by the expression of the dominant negative constructs. Average travel distances (modified as described in Materials and methods) and velocities were measured during dispersion (A and B) and aggregation (C and D). Run lengths, dispersion (A); velocities, dispersion (B); run lengths, aggregation (C); velocities, aggregation (D). The myosin V and kinesin II constructs both alter the length of minus end runs (A), but myosin V does not alter the length of plus end runs. Blocking myosin V function results in increased velocities in both plus and minus end travel (B), but decreasing kinesin II function does not alter minus end velocities (B). During aggregation, blocking myosin V activity has no effect on either run lengths (C) or velocities (D). M-V, myosin V. Error bars are the standard error of the mean.
Figure 7.
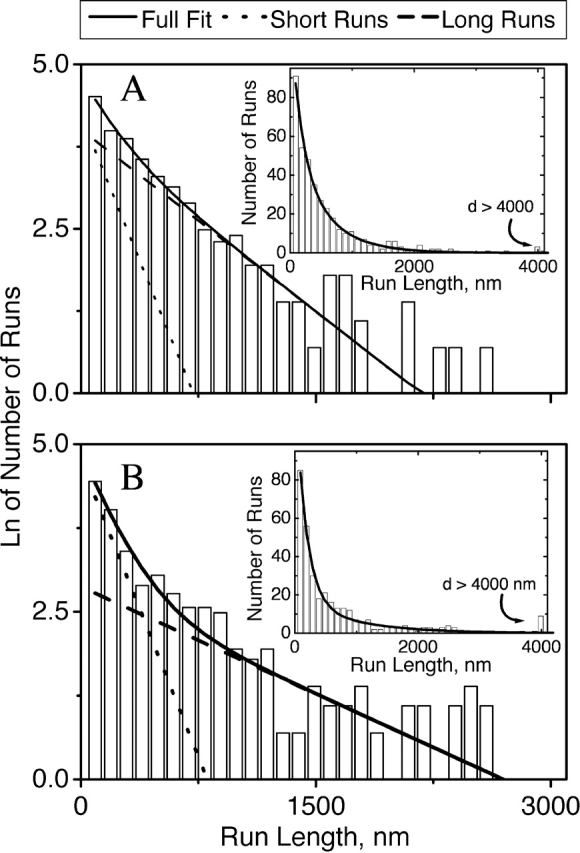
Distribution of run lengths for minus end motion in wild-type (A) and myosin V (B) cells stimulated with MSH. Individual run lengths were determined as described in Materials and methods. In all cases examined, the distribution was well described by the sum of two decaying exponentials (Table I). The original histogram (bars) together with the double-decaying exponential fit (solid line) is shown in the inserts. Note that there are relatively more long runs in the myosin V cells (B) than in wild-type cells (A). The full graphs show the natural logarithm of the number of runs versus run length. The solid line is the complete fit, whereas the dotted and dashed lines indicate the contributions of the short and long runs, respectively. The long runs (tail of the distribution) are significantly altered due to myosin V activity, but the short runs are not (compare the slope of the long-runs line in A and B).
Table I. Physical parameters of melanophore motion.
| Long distance constant D
L, minus end |
Number ratio N
SL, minus end |
X
V
2, P(X
V
2), minus end motion |
Long distance constant D
L, plus end |
Number ratioN SL, plus end |
X
V
2, P(X
V
2), plus end motion |
|
|---|---|---|---|---|---|---|
| nm | nm | |||||
| Wild-type dispersion |
519 ± 67 | 0.29 ± 0.22 | 0.42, 0.96 | 665 ± 46 | 0.25 ± 0.20 | 0.41, 0.95 |
| Wild-type aggregation |
1,206 ± 200 | 0.16 ± 0.12 | 0.95, 0.42 | 845 ± 118 | 0.11 ± 0.08 | 0.71, 0.69 |
| Dominant negative myosin V dispersion |
974 ± 117 | 1.25 ± 1.06 | 0.67, 0.80 | 831 ± 43 | 0.65 ± 0.43 | 0.32, 0.99 |
| Latrunculin dispersion |
1,318 ± 206 | 1.12 ± 0.48 | 0.74, 0.85 | 1,180 ± 150 | 0.55 ± 0.30 | 0.86, 0.72 |
| Dominant negative myosin V aggregation |
1,284 ± 150 | 0.16 ± 0.14 | 1.12, 0.30 | 876 ± 145 | 0.15 ± 0.09 | 1.31, 0.15 |
Melanosome motion was characterized by tracking analysis. The distance constant D L was found by fitting histograms of travel distance, D, to the sum of two exponential functions: y(D) = exp(−D/D s) 1 A L exp(−D/D L). D S did not vary appreciably, with an approximate value in all cases of ∼130 ± 40 nm. The long distance constant D L is an estimate of mean run length of long runs because for a decaying exponential the decay constant is identical to the average travel distance. The number ratio N SL is the number of short runs divided by the number of long runs and thus measures the relative frequency of the two travel states. The X V 2 values (with corresponding probabilities) indicate the goodness of the fit.
To quantify the regulation of myosin V–based motion, we used the r2(t) statistic, which measures the average travel distance of a melanosome from the point at which we started tracking it. We depolymerized microtubules with nocodazole, stimulated cells with melatonin or MSH, and computed r2(t) from tracking many melanosomes chosen at random in each class of cells (Fig. 4). Myosin V–driven motion is decreased ∼50% in melatonin- versus MSH-treated cells as judged by comparing the slope of r2(t) under these two conditions (Fig. 4). Thus, a second component of the switch from dispersion to aggregation includes a down-regulation of myosin V–driven motion.
Other work (Reese and Haimo, 2000) suggested that the change from net aggregation to net dispersion resulted from changes in the ability of both kinesin II and dynein to interact with microtubules. Their biochemical analysis found that during dispersion, binding of dynein to microtubules was somewhat decreased after 15 min of treatment with MSH and completely lost after 60 min with the hormone. We compared the microtubule component of motion in cells 15 and 60 min after addition of MSH and found minus end movements are present even after 60 min in MSH. Furthermore, the length of both plus end and minus end runs increased at 60 min so the minus end motion clearly was not impaired (Fig. 6 A), whereas the mean travel velocity did not change. Because melanosomes show bidirectional motion even 60 min after the cells receive the dispersion or aggregation signal, the data of (Reese and Haimo, 2000) are inconsistent with our observations.
There is a formal possibility that although the motion of individual melanosomes appears similar, the number of plus or minus moving organelles is different. To investigate this possibility, we identified all melanosomes that appeared moving on microtubule tracks in a given cell in a 30-s period, 15 and 60 min after addition of MSH. At 15 min, we observed that 53 ± 10% (four cells; mean ± SEM) of the moving melanosomes included a minus end run, whereas at 60 min 75 ± 4% (four cells; mean ± SEM) of the moving melanosomes included a minus end run. Thus, there is no evidence of a decrease in dynein activity as a function of time in dispersing cells.
Finally, in biochemical experiments directly repeating the protocol published by Reese and Haimo (2000) we were unable to reproduce the change in microtubule binding. In our experiments, the amount of kinesin II and cytoplasmic dynein that binds to microtubules in the presence of AMP-PNP did not change after 60 min treatment with melatonin or MSH (Fig. 8 B).
Figure 8.

The ability of motors to interact with melanosomes and microtubules does not change between aggregation and dispersion. (A) Melanosomes were purified from cells treated with the aggregating stimulus, melatonin, or the dispersing stimulus, MSH, and Western blots were performed with antibodies against myosin V (M-V), kinesin II (K-II), and cytoplasmic dynein (Dy). Note that the amount of microtubule motors on melanosomes does not change due to aggregation (A) or dispersion (D), whereas the amount of myosin V is higher on melanosomes purified from cells dispersing the pigment than from cells aggregating the pigment. (B) Binding of kinesin II (K-II) or cytoplasmic dynein (Dy) to microtubules in the presence of AMP-PNP. Bovine brain microtubules were added to extracts from cells aggregating (A) or dispersing (D) pigment, the mixture was spun through a glycerol cushion, and the pellets were analyzed for the presence of motors by Western blotting (experimental protocol from Reese and Haimo (2000).
Regulation of myosin V involves changes in the number of cargo-bound motors
The tracking experiments demonstrate that regulation alters both the dynein-driven component of motion and the myosin V component of motion. To determine if these changes are due to an alteration of the number of motors bound to the cargo, we measured the amount of melanosome-bound motors on purified melanosomes using quantitative Western blot analysis. We found no change in the overall amount of microtubule motors bound to melanosomes purified from dispersing versus aggregating cells (Fig. 8 A). Thus, for the microtubule motors alteration of motor cargo binding does not appear to play a significant role in regulation of net transport. This is consistent with previously published work (Reese and Haimo, 2000).
However, the situation is clearly different in the case of myosin V. Melanosomes from aggregating cells have substantially less myosin V than melanosomes from dispersing cells (Fig. 8 A, M-V). We quantified the number of molecules of myosin V on the surface of melanosomes from aggregating and dispersing cells, using Western blotting with a recombinant fragment of myosin V as a standard. This procedure yields an estimate of 65 ± 10 and 88 ± 7 molecules of myosin V per melanosome in aggregating and dispersing cells, respectively. This 35% change is comparable to the decrease in myosin V–based movement (Fig. 4); therefore, an attractive model is that the decrease in myosin V–based motion is the result of a change in melanosome-bound myosin V (see Discussion).
Interactions between the microtubule and actin transport systems
How does myosin V alter microtubule-based transport? To understand interactions between the two systems during melanosome transport, we studied how the function of one system is altered by the inactivation of the other. Inactivation of myosin V alters microtubule-based motion: the travel velocity along microtubules in each direction is increased (Fig. 6 B) as is the mean length of minus end runs (Fig. 6 A).
Because a distribution's mean is an incomplete description of the underlying process, we determined how the loss of myosin V activity altered the distribution of microtubule-based run lengths. The length of runs was well described in all cases by the sum of two decaying exponentials (Table I, reduced chi-squared values, and Fig. 7), suggesting that there are two classes of motion, that is, short runs and long runs. From the fit, we determine distance constants characterizing both types of runs (D S and D L) and the relative proportion of short versus long runs (N SL). D L provides an estimate for the average length of long runs (Table I). During dispersion, impairment of myosin V activity due to latrunculin or the myosin V dominant negative construct resulted in an increase in D L (Table I and Fig. 7) and an increase in N SL. The effect was much greater on minus end motion than plus end motion (Table I). D S remained approximately constant (unpublished data). Thus, during dispersion myosin V has two opposite effects: it improves microtubule-based transport by suppressing short runs but also impairs microtubule transport by shortening long runs.
Further comparing motion in the wild-type and myosin V dominant negative cells, we observed that in wild-type cells there appear to be fewer melanosomes that remained stationary. To quantify this effect, we tracked ≥80 randomly chosen melanosomes in three different backgrounds: wild-type, myosin V dominant negative, and latrunculin-treated cells. We operationally defined as stationary any melanosome that did not move at least 500 nm away from the starting point at least once over the course of 33 s of observation (this corresponds to an average velocity of <15 nm/s). In the wild-type cells, only 6.8% of the melanosomes fall into the stationary category compared with 17.1% in a latrunculin-treated cells and 21.6% in myosin V dominant negative cells.
However, inactivation of myosin V activity either by latrunculin or the myosin V construct resulted in a subset of the microtubule-driven motion that was particularly effective. In the wild-type (88 granules tracked), there were no displacements of more than 4,000 nm over the course of 33 s (0%), whereas there were 5 such events (out of 97) in the myosin V dominant negative background (5.1%) and 11 such events (out of 82) in the latrunculin background (13.4%).
To find if the interaction between the two systems is regulated, we compared the microtubule-based motion in control cells (aggregating and dispersing) and myosin V dominant negative cells (aggregating and dispersing). In aggregating cells, the microtubule-based motion is not altered by the myosin V inactivation (Fig. 6, C and D, and Table I) in contrast to the observed effect during dispersion (Fig. 6, A and B, Table I). Thus, cells can regulate the extent to which myosin V interferes with the microtubule-based motion.
Interactions between the microtubule motors
How does kinesin II alter dynein-based motion? To investigate, we compared minus end motion in wild-type and kinesin II dominant negative cells. We find no difference in travel velocity in wild-type versus the kinesin II dominant negative background (Fig. 6 B); however, loss of kinesin II activity does result in an increase in the mean minus end travel distance (Fig. 6 A). Further work remains to more fully understand the effects of disabling kinesin II on the minus end motion.
Discussion
The use of dominant negative mutants for analysis of organelle movement
In this work, we used dominant negative versions of myosin V and kinesin II to investigate the role of individual motors in pigment organelle movement in melanophores, and regulation of these motors. To interpret the results of particle tracking analysis, we addressed whether expression of truncated proteins results in simple loss of function of their wild-type counterparts or has a more complex effect. Our results with myosin V dominant negative constructs show that both the fragment that includes only the globular portion of the tail and the fragment that includes both the globular part and a part of coiled-coil completely suppress movement of melanosomes along actin filaments and have the same effect upon microtubule-based motion. We demonstrated that the fragment that includes a portion of coiled-coil can bind to the wild-type heavy chain, forming heterodimers with the full-length heavy chain and can also bind to melanosomes, competing out the wild-type protein (unpublished data). However, the construct encoding just the globular portion of the tail that is unable to form dimers has the same effect on movement. This result suggests that either the one-headed heterodimer cannot effectively interact with actin or more likely the level of expression of the truncated protein is much higher than that of endogenous myosin V. In the latter case, the number of one-headed heterodimers would be much less that of truncated homodimers, and homodimers would effectively displace both heterodimers and wild-type myosin V from the surface of organelles. In either case, experimental results demonstrate that overexpression of the tail fragment in melanophores results in the complete loss of myosin V function and therefore is a reliable tool for the functional studies.
In the case of kinesin II, we have demonstrated previously that truncated 85-kD subunit (Xklp-3) without the motor domain is still capable of binding to the two other subunits of the protein (Tuma et al., 1998), and therefore it is possible that the motor domain on the 95-kD subunit interacts with microtubules. However, this interaction is not sufficient to power organelle movement along microtubules during dispersion (Tuma et al., 1998). Further, in vitro studies that examined the activity of one-headed kinesin heterodimers found that four to six single-headed kinesins could move microtubules in a gliding assay, suggesting that the single-headed kinesins were not in a rigor state that opposed motion (Hancock and Howard, 1998). Almost complete inhibition of movement in kinesin II/myosin V double mutant shows that kinesin II activity is mostly inhibited also. Combined, we believe that overexpression of the kinesin II dominant negative constructs predominantly blocks kinesin II–mediated motion by causing a loss of function but is unlikely to result in rigor binding of the cargo to the microtubules blocking movement irreversibly.
Contribution of kinesin II and myosin V to pigment dispersion
The observed r2(t) due to myosin V activity alone (Fig. 4) is almost as high as that observed in wild-type cells, suggesting that myosin V is able to contribute significantly to the transport.
Equally important, myosin V alters the efficiency of microtubule-based dispersion: without myosin V, during dispersion the plus and minus runs are approximately the same length so that one would not expect significant dispersion (Fig. 5 A). However, active myosin V changes this balance because it impairs minus end motion more significantly than plus end motion. This effect is reflected in the alteration of mean minus end travel distance (Fig. 6 A) but is particularly evident when one looks at its effect on the length of long runs. In the absence of myosin V, long minus end runs are 88% longer (974 versus 519 nm) (Table I and Fig. 7), whereas long plus end runs are only 25% longer (831 versus 665 nm). Thus, myosin V activity particularly shortens long minus end runs and helps kinesin II–based motion to dominate dynein-based motion. At a global level, the results of this effect can be seen by observing that in cells without myosin V activity (i.e., in latrunculin-treated or myosin V dominant negative cells) giving the cells the dispersion stimulus does not result in overall dispersion; however, pigment granules are found predominantly in the cell center (unpublished data; Tuma et al., 1998).
Finally, myosin V activity acts to regularize the motion of individual particles: eliminating movement along actin filaments leads to an increase in stalled particles, short runs, and particles which travel extremely far distances.
These findings result in a modification of the capture model proposed for melanosome movement in mouse melanocytes, which suggests that myosin V acts merely to anchor the cargos after that are moved by the microtubule transport system (Wu et al., 1998).
Mechanisms regulating melanosome transport
One goal of this work was to investigate regulation of organelle transport, specifically the extent and significance of interactions between three types of motor proteins (kinesin II, cytoplasmic dynein, and myosin V) present on melanosomes. We show that the change from net aggregation to net dispersion of melanosomes results from the regulation of two aspects of motion: the length of dynein-driven runs is decreased, and the activity of myosin V is increased.
How are these changes in motion accomplished? Our previous work (Rogers et al., 1999; Karcher et al., 2001) demonstrated that alteration of cargo binding was used to alter organelle transport by myosin V during the cell cycle. We believe that a similar mechanism operates during pigment aggregation. The 35% decrease in the amount of myosin V bound to melanosomes is roughly consistent with the 50% decrease in myosin V driven r2(t), suggesting that changes in the amount of motor bound to the organelle may explain changes in the myosin V component of melanosome motion during aggregation. Therefore, we favor the hypothesis that both the magnitude of r2(t) and the ability of myosin V to compete with the microtubule motors are determined by controlling the binding of myosin V to melanosome.
In contrast, we and others (Reese and Haimo, 2000) found that the quantity of microtubule motors bound to the melanosomes does not change, and therefore regulation of motor number is probably not used to control the length of microtubule runs. However, it remains unclear how direct regulation of dynein or kinesin II could result in changes in run length. One possibility is that the coordination between the microtubule motors of opposite polarity discussed below allows a switch to be used by the cell to regulate the transition between opposite directions of travel. Such a switch was hypothesized to control bidirectional transport of lipid droplets in early Drosophila embryos (Gross et al., 2000). In both cases, lipid droplets in Drosophila and melanosomes in Xenopus melanophores, regulation of net transport occurs by altering motion in only one direction of travel. How such a switch works is still not clear; however, the work on Drosophila lipid droplets suggests that proteins such as klarsicht are required both for coordination of opposite polarity motors and regulation of the switching probability (Welte et al., 1998).
Coordination of microtubule motors and tug-of-war with myosin V
The observation that, unlike impairment of myosin V, the dominant negative impairment of kinesin II function does not increase the velocity of minus end transport suggests that there is not a tug-of-war between the microtubule motors, but instead their activity is coordinated to prevent simultaneous activity. Certainly, the molecular mechanism of such coordination is not yet known.
We and others (Wu et al., 1998; Bridgman, 1999) observed significant stimulation of microtubule-based motion due to a loss of myosin V function. This result shows that microtubule- and actin-based transport are not coordinated but rather in a tug-of-war.
We believe that myosin V alters microtubule-based transport in two important ways. First, myosin V selectively ends minus end runs, thus helping to generate net plus end–directed motion (see the model below). In this respect, by making plus end runs longer than minus end runs, myosin V makes the transport along microtubules more directional. Second, in the presence of myosin V activity melanosomes are less likely to pause or become stuck. Thus, by making travel along actin filaments accessible myosin V may help melanosomes detour around blockages that could not be avoided if they were constrained to microtubules.
Finally, a remarkable property of the regulated transport in melanophores is that the tug-of-war between myosin V and the microtubule motors is only observed during pigment dispersion. Down-regulation of myosin V transport during aggregation impairs its ability to compete with microtubule-based transport, and therefore this tug-of-war itself is the subject of physiological regulation (see the model below).
Model
The observations discussed above can be integrated into a model presented in Fig. 9 . This model describes the behavior of each of the three motors, dynein, kinesin II, and myosin V, in cells dispersing and aggregating melanosomes. We hypothesize that during dispersion, melanosomes are simultaneously linked to both microtubules and actin. As a result, there is a continual competition between the two transport systems. One effect of this tug-of-war is that microtubule motor-based transport is slowed down by myosin V interference (Wu et al., 1998; Bridgman, 1999).
Figure 9.

Model for transport of melanosomes by actin- and microtubule-based motors. A pigment organelle can be moved in three different ways: along the microtubule by kinesin II (A and D) or dynein (B and E), or along the actin filaments by myosin V (C and F). Direction of movement is shown by double arrows. Although at any moment motion is dominated by one of the two transport systems (i.e., microtubules or actin), the motors of the other transport system are transiently active and interacting with their substrate (i.e., actin or microtubules). During dispersion, these transient interactions (indicated by T) are significant and allow myosin V to reduce the velocity of microtubule-based transport (A and B). The interactions also play a role in the switch between the two transport systems, that is, between B and C. Because myosin V activity predominantly decreases the length of minus end microtubule-based motion, we hypothesize that the switch from microtubule- to actin-based transport occurs only between state B and C (from dynein movement to myosin V movement) and that transitions from kinesin II movement (A) to myosin V movement (C) are rare. Because kinesin II but not dynein appears to win in tug-of-wars with myosin V, we suggest that the C to A transition is possible but not the C to B transition; however, we have no direct evidence on this point. During aggregation, myosin V activity is decreased, which results in reduced weak transient interactions (indicated by weak T) in D and E, and myosin V is no longer able to interfere with microtubule-based motion. Due to the weakness of the interactions, any time the microtubule motors are in contact with the microtubules they win the tug-of-war with myosin V and there is a transfer (F to D or F to E). Similarly, the reverse transfer from microtubules to actin-based transport (D to F or E to F) does not occur. M-V, myosin V; K-II, kinesin II; Dy, dynein; Ms,melanosome.
A key feature of the model is that during dispersion the transition from microtubule-dominant transport to actin-dominant transport occurs predominantly during dynein-driven motion (transition 9 B to 9 C). The result of such a switch is to end a minus end run by switching onto actin. This decreases minus end motion, favoring dispersion. Since the length of kinesin II runs are much less affected by the activity of myosin V, we hypothesize that any kinesin II/myosin V tug-of-war tends to end in kinesin II's favor. Thus, the model shows that the 9 A to 9 C transition is not allowed. Furthermore, the fact that kinesin II tends to win such tug-of-wars more than dynein makes the 9 C to 9 A transition more likely than the 9 C to 9 B transition. In turn, this also tends to bias the transport toward the plus end direction.
Another likely reason that cells need to use both systems for pigment dispersion is that unlike actin filaments microtubules do not uniformly cover the cytoplasm. During dispersion, the goal is to uniformly spread melanosomes out so transfer from microtubules to actin is necessary for the cell to achieve homogeneous pigment dispersion (Rodionov et al., 1998).
During aggregation, we observe no tug-of-war between actin and microtubule transport because the myosin V component is down-regulated. Nevertheless, its residual activity is still sufficient to move melanosomes along actin filaments. This is essential because diffusional movement of organelles is negligible (Fig. 4) and melanosomes that are not in contact with microtubules would never reach them and therefore would not be delivered to the cell center. At the same time, down-regulated myosin V transport is unable to compete effectively with the microtubule motors. As a result, melanosomes that are in contact with the actin filaments only (Fig. 9 F) are moved along them until they encounter a microtubule at which point the microtubule motors pull the melanosome off the actin and onto the microtubules (transition from 9 F to 9 D and 9 F to 9 E). These melanosomes then stay on microtubules, traveling back and forth with an inward bias (Fig. 9, D and E) until they reach the cell center.
Although our model proposes a tug-of-war between myosin V and the microtubule motors, we believe there is no such tug-of-war between the microtubule motors of the opposite polarity. Instead, kinesin II and dynein activity is coordinated, with a switch controlling whether the plus or the minus end component of movement is active at any particular moment (Fig. 9, A and B).
In summary, the model proposes that both cytoskeletal transport systems contribute significantly to melanosome motion and that regulation of net transport involves changing both the motility properties of dynein and myosin V and also altering the interactions of myosin V with the microtubule motors.
Relation of this model to existing work
The data presented here show that myosin V alters microtubule-driven transport. Thus, our model alters the capture model proposed by Wu et al. (1998): we show that the activity of myosin V does indeed help to block dynein-based minus end travel, not by holding the melanosome in place but rather by terminating a minus end run by moving it away from the microtubule. In contrast to a delivery-plus-capture picture, in our model these processes are quite dynamic: melanosomes are constantly moving on and off the microtubules, and myosin V contributes a significant amount of motility.
In this paper, we have demonstrated that regulation of net transport occurs both through alteration of dynein run length and myosin V motility. Further, our observations suggest that myosin V plays two roles in the melanosome transport system. The use of the randomly oriented and positioned actin filament network helps the system to achieve a uniform distribution of melanosomes for highly effective color change of the cells as originally proposed by Rodionov et al. (1998). In addition, by selectively interrupting minus end motion myosin V blocks dynein-based transport and increases net dispersion. From these studies, two major questions emerge. First, what is the nature of the process governing the reversals of microtubule transport? Second, is regulation of myosin V explained purely by decrease in the amount of bound motors, or does it reflect direct regulation of myosin V motor activity as well? These questions have to be answered by direct studies of organelle motility in a purified system.
Materials and methods
Cell culture
An immortalized Xenopus melanophore cell line (a gift from Dr. Michael Lerner, University of Texas Southwestern Medical Center, Dallas, TX) was cultured as described (Rogers et al., 1997). For elimination of melanosomes, the medium was supplemented with PTU, an inhibitor of tyrosinase, at a final concentration of 1 mM and maintained in this medium for at least 3 wk. Cells with a small number of melanosomes used for organelle-tracking experiments were generated by transferring of melanosome-free melanophores into medium without PTU for 24–48 h.
Aggregation and dispersion of pigment were induced by 10 nM melatonin or 100 nM MSH, respectively. Microtubules were depolymerized by treatment of melanophores with 10 μM nocodazole for 60 min. Depolymerization was verified by immunofluorescent staining with an antitubulin antibody DM1α (Sigma-Aldrich). Plasmids encoding GFP- and myc-tagged dominant negative versions of myosin V and kinesin II (Xklp-3) were described previously (Tuma et al., 1998; Rogers et al., 1999). Transfection was performed using FuGene-6 (Roche Diagnostics).
Melanosome tracking and movement analysis
Movement of melanosomes in vivo was imaged with bright-field microscopy using Microphot SA upright microscope (Nikon) with a 60× 1.4 NA plan apochromat lens. A filter cutting light with a wavelength <715 nm was installed in the illumination path. Melanosome movement was captured using a Newvicon camera (Hamamatsu Photonics) with a 4× projection lens (∼66 nm/pixel), processed using Argus-10 processor, and recorded using a S-VHS VCR AG-6730 (Panasonic) at a rate of 30 Hz. Cells transfected with dominant negative constructs were identified using fluorescent microscopy. For cotransfections with dominant negative mutants of kinesin II and myosin V, we used GFP-tagged version of headless kinesin II and myc-tagged version of headless myosin V. In these experiments, GFP-positive cells were used for live recording, and then cultures were fixed and stained with anti-myc antibody (Evan et al., 1985) to confirm that they expressed both proteins.
Video sequences of ∼1 min were transferred from videotape onto the hard disk of a Silicon Graphics O2 workstation using the SGI frame-grabbing software and hardware. Melanosomes were tracked using the nanotrack routine of Isee image processing software (Inovision Corp.).
The trajectories resulting from this particle tracking were analyzed in two different ways. If the motion in question involved myosin V activity and thus contained periods of apparent random nondirectional travel, we used the r2(t) statistic, which is simply a measure of how far the particle moved in any direction from the point at which we started tracking it, as a function of time. Since the motion was reminiscent of diffusion, we used the square of the displacement, since for a true two-dimensional diffusion the square of the displacement is proportional to the time, that is, (r2) = 4Dt. In most cases (Fig. 4), the plot of r2(t) does not appear to be well fit by a single line, suggesting that the motion was inconsistent with simple diffusion.
If the microtubule component of the motion was being analyzed, we wanted to look at the component of motion along the microtubule axis. In this case, a line fit to the trajectory was used to estimate the microtubule axis, and then motion along the microtubule was determined by projecting the trajectory onto this best fit line (Welte et al., 1998). This record of displacement along the microtubule versus time was then analyzed by breaking it into periods of uninterrupted plus and minus end motion that is, runs, as described (Gross et al., 2000) with the exception that due to slightly more noise in the acquisition system, for a displacement to be scored as a run it needed to be ≥40 nm (instead of 30 nm) and last ≥0.23 s (instead of 0.16 s). Similarly, pauses were operationally defined as periods when there was <40 nm displacement for a duration of ≥0.23 s.
Two different definitions of runs were used: long-type and short-type runs. For all conclusions regarding displacement, we used an intuitive definition: a run was ended only by a reversal in travel or cessation of motion along the microtubule (i.e., an apparent switch to actin-based motion). Short pauses do not end long-type runs. Pauses are ignored because they don't result in any net transport. In addition to overall transport that was quantified using long-type runs, we wanted to quantify the velocity of travel because a tug-of-war can cause a decrease in travel velocity. However, the average velocity of a long-type run is hard to use to investigate this question because long-type runs can include pauses, and long pauses between periods of relatively fast motion would lead to a low average velocity, and we are interested in tug-of-wars that occur while the particle is moving. To address this question, we defined short-type runs that exclude pauses and are defined as any period of uninterrupted motion ended by a pause in motion, a reversal in travel direction, or cessation of motion along the microtubule (i.e., an apparent switch to actin-based motion). All run length data presented (Fig. 5 A, Fig. 6, A and C, and Table I) were derived from long-type run lengths. All velocity data (Fig. 5 B and Fig. 6, B and D) were derived from short-type runs.
Because the distribution of run lengths was very skewed, we could not simply average the data to estimate the mean. Instead, by taking the natural log of the lengths of runs, we were able to transform the data to a normal distribution. We then found the mean and standard error of the mean for this new dataset. Finally, we exponentiated the transformed mean to estimate the mean of the original dataset and exponentiated the mean plus (or minus) the standard error to estimate the confidence interval for the reported mean. The data shown in Fig. 5 A and Fig. 6, A and C, result from this modified averaging process.
A second method to deal with this issue involved fitting the distribution of run lengths to the sum of two decaying exponentials (in many cases, a single decaying exponential was not a good model for the distribution, as judged by reduced chi-squared values, but in all cases the double decaying exponential function was a good approximation, as judged by this criteria) (Table I). No changes were observed for the short decay constant; however, the long decay constant was altered in the different genetic backgrounds, as was the relative number of short versus long runs.
Microtubule imaging
For microtubule imaging, melanophores were microinjected with 2 mg/ml of purified bovine brain tubulin labeled with Alexa 488 fluorescent dye (Molecular Probes) according to the protocol described in Keating et al. (1997). Simultaneous imaging of melanosomes and microtubules was performed using a Nikon Diaphot 200 inverted microscope equipped with a Planapo 60× 1.4 NA objective. Images were collected with an Orca II cooled CCD camera (Hamamatsu Photonics) driven by Isee software. Images were projected onto the CCD chip with a magnification of 0.17 μm/pixel. Bright-field images of melanosomes were captured every 3 s and fluorescent images every 30 s.
Biochemical techniques
Melanosomes were purified according to Rogers et al. (1998). Cosedimentation of motor proteins with microtubules was performed as described by Reese and Haimo (2000). For Western blot analysis of microtubule pellets, samples were first run on SDS-polyacrylamide gels and stained with Coomassie blue R-250, and then the gels were scanned, and the load for Western blots was normalized according to the scan data. The same procedure was used to compare the amount of motor proteins on melanosomes purified from cells treated with MSH or melatonin. The tubulin band was used for normalization in the case of microtubule-binding experiments and the tyrosinase band in the case of melanosome experiments. For Western blots, we used mouse monoclonal antibodies against kinesin II k2.4 (Cole et al., 1993) and against dynein intermediate chain 70.1 (Sigma-Aldrich). Myosin V was visualized with an affinity purified rabbit polyclonal antibody DIL-2 against a recombinant fragment of the heavy chain produced according to Wu et al. (1997) and characterized previously (Rogers et al., 1999).
For quantitative analysis of myosin V on melanosomes, the number of organelles in samples was measured using a hemocytometer. Protein samples made from a known amount of melanosomes purified from cells dispersing and aggregating the pigment were serially diluted and loaded on a gel together with dilutions of a myosin V standard (recombinant DIL-2 fragment of the heavy chain [Wu et al., 1997]). Blots were probed with the antibody against myosin V, scanned, and samples within the linear range of the method (for both the standard and melanosomes) were used for calculations.
Acknowledgments
We would like to thank Kim Ly (University of Illinois) for help in data analysis and Alexander Minin (Russian Academy of Sciences, Moscow, Russia) for help with tubulin labeling.
S. Gross gratefully acknowledges support from University of California, Irvine start-up funds. V.I. Gelfand acknowledges support from the National Institutes of Health (GM-52111) and the National Science Foundation (95-13388).
S.P. Gross and M.C. Tuma contributed equally to this work.
M.C. Tuma's present address is Dept. of Molecular, Cellular, and Developmental Biology, Yale University, New Haven, CT 06520.
A.R. Reilein's present address is Dept. of Molecular and Cellular Physiology, Stanford University Medical Center, Palo Alto, CA 94304.
Footnotes
Abbreviations used in this paper: MSH, melanocyte-stimulating hormone; PTU, phenylthiourea.
References
- Bridgman, P.C. 1999. Myosin Va movements in normal and dilute-lethal axons provide support for a dual filament motor complex. J. Cell Biol. 146:1045–1060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown, S.S. 1999. Cooperation between microtubule- and actin-based motor proteins. Annu. Rev. Cell Dev. Biol. 15:63–80. [DOI] [PubMed] [Google Scholar]
- Cole, D.G., S.W. Chinn, K.P. Wedaman, K. Hall, T. Vuong, and J.M. Scholey. 1993. Novel heterotrimeric kinesin-related protein purified from sea urchin eggs. Nature. 366:268–270. [DOI] [PubMed] [Google Scholar]
- Daniolos, A., A.B. Lerner, and M.R. Lerner. 1990. Action of light on frog pigment cells in culture. Pigment Cell Res. 3:38–43. [DOI] [PubMed] [Google Scholar]
- Evan, G.I., G.K. Lewis, G. Ramsay, and J.M. Bishop. 1985. Isolation of monoclonal antibodies specific for human c-myc proto-oncogene product. Mol. Cell. Biol. 5:3610–3616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gross, S.P., M.A. Welte, S.M. Block, and E.F. Wieschaus. 2000. Dynein-mediated cargo transport in vivo. A switch controls travel distance. J. Cell Biol. 148:945–956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hancock, W.O., and J. Howard. 1998. Processivity of the motor protein kinesin requires two heads. J. Cell Biol. 140:1395–1405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hirschberg, K., C.M. Miller, J. Ellenberg, J.F. Presley, E.D. Siggia, R.D. Phair, and J. Lippincott-Schwartz. 1998. Kinetic analysis of secretory protein traffic and characterization of Golgi to plasma membrane transport intermediates in living cells. J. Cell Biol. 143:1485–1503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huang, J.D., S.T. Brady, B.W. Richards, D. Stenolen, J.H. Resau, N.G. Copeland, and N.A. Jenkins. 1999. Direct interaction of microtubule- and actin-based transport motors. Nature. 397:267–270. [DOI] [PubMed] [Google Scholar]
- Karcher, R.L., J.T. Roland, F. Zappacosta, M.J. Huddleston, R.S. Annan, S.A. Carr, and V.I. Gelfand. 2001. Cell cycle regulation of myosin-V by calcium/calmodulin-dependent protein kinase II. Science. 293:1317–1320. [DOI] [PubMed] [Google Scholar]
- Keating, T.J., J.G. Peloquin, V.I. Rodionov, D. Momcilovic, and G.G. Borisy. 1997. Microtubule release from the centrosome. Proc. Natl. Acad. Sci. USA. 94:5078–5083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Krendel, M., G. Sgourdas, and E.M. Bonder. 1998. Disassembly of actin filaments leads to increased rate and frequency of mitochondrial movement along microtubules. Cell Motil. Cytoskeleton. 40:368–378. [DOI] [PubMed] [Google Scholar]
- Kuznetsov, S.A., G.M. Langford, and D.G. Weiss. 1992. Actin-dependent organelle movement in squid axoplasm. Nature. 356:722–725. [DOI] [PubMed] [Google Scholar]
- Langford, G.M. 1995. Actin- and microtubule-dependent organelle motors: interrelationships between the two motility systems. Curr. Opin. Cell Biol. 7:82–88. [DOI] [PubMed] [Google Scholar]
- Maples, C.J., W.G. Ruiz, and G. Apodaca. 1997. Both microtubules and actin filaments are required for efficient postendocytotic traffic of the polymeric immunoglobulin receptor in polarized Madin-Darby canine kidney cells. J. Biol. Chem. 272:6741–6751. [DOI] [PubMed] [Google Scholar]
- Mermall, V., J.G. McNally, and K.G. Miller. 1994. Transport of cytoplasmic particles catalysed by an unconventional myosin in living Drosophila embryos. Nature. 369:560–562. [DOI] [PubMed] [Google Scholar]
- Morris, R.L., and P.J. Hollenbeck. 1995. Axonal transport of mitochondria along microtubules and F-actin in living vertebrate neurons. J. Cell Biol. 131:1315–1326. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reese, E.L., and L.T. Haimo. 2000. Dynein, dynactin, and kinesin II's interaction with microtubules is regulated during bidirectional organelle transport. J. Cell Biol. 151:155–166. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Rodionov, V.I., A.J. Hope, T.M. Svitkina, and G.G. Borisy. 1998. Functional coordination of microtubule-based and actin-based motility in melanophores. Curr. Biol. 8:165–168. [DOI] [PubMed] [Google Scholar]
- Rogers, S.L., and V.I. Gelfand. 1998. Myosin cooperates with microtubule motors during organelle transport in melanophores. Curr. Biol. 8:161–164. [DOI] [PubMed] [Google Scholar]
- Rogers, S.L., I.S. Tint, P.C. Fanapour, and V.I. Gelfand. 1997. Regulated bidirectional motility of melanophore pigment granules along microtubules in vitro. Proc. Natl. Acad. Sci. USA. 94:3720–3725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers, S.L., I.S. Tint, and V.I. Gelfand. 1998. In vitro motility assay for melanophore pigment organelles. Methods Enzymol. 298:361–372. [DOI] [PubMed] [Google Scholar]
- Rogers, S.L., R.L. Karcher, J.T. Roland, A.A. Minin, W. Steffen, and V.I. Gelfand. 1999. Regulation of melanosome movement in the cell cycle by reversible association with myosin V. J. Cell Biol. 146:1265–1276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tabb, J.S., B.J. Molyneaux, D.L. Cohen, S.A. Kuznetsov, and G.M. Langford. 1998. Transport of ER vesicles on actin filaments in neurons by myosin V. J. Cell Sci. 111:3221–3234. [DOI] [PubMed] [Google Scholar]
- Tuma, M.C., A. Zill, N. Le Bot, I. Vernos, and V. Gelfand. 1998. Heterotrimeric kinesin II is the microtubule motor protein responsible for pigment dispersion in Xenopus melanophores. J. Cell Biol. 143:1547–1558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Welte, M.A., S.P. Gross, M. Postner, S.M. Block, and E.F. Wieschaus. 1998. Developmental regulation of vesicle transport in Drosophila embryos: forces and kinetics. Cell. 92:547–557. [DOI] [PubMed] [Google Scholar]
- Wu, X., B. Bowers, Q. Wei, B. Kocher, and J.A. Hammer III. 1997. Myosin V associates with melanosomes in mouse melanocytes: evidence that myosin V is an organelle motor. J. Cell Sci. 110:847–859. [DOI] [PubMed] [Google Scholar]
- Wu, X., B. Bowers, K. Rao, Q. Wei, and J.A.R. Hammer. 1998. Visualization of melanosome dynamics within wild-type and dilute melanocytes suggests a paradigm for myosin V function in vivo. J. Cell Biol. 143:1899–1918. [DOI] [PMC free article] [PubMed] [Google Scholar]