

Abstract
Hippocalcin is a neuronal calcium sensor protein that possesses a Ca2+/myristoyl switch allowing it to translocate to membranes. Translocation of hippocalcin in response to increased cytosolic [Ca2+] was examined in HeLa cells expressing hippocalcin–enhanced yellow fluorescent protein (EYFP) to determine the dynamics and Ca2+ affinity of the Ca2+/myristoyl switch in living cells. Ca2+-free hippocalcin was freely diffusible, as shown by photobleaching and use of a photoactivable GFP construct. The translocation was dependent on binding of Ca2+ by EF-hands 2 and 3. Using photolysis of NP-EGTA, the maximal kinetics of translocation was determined (t 1/2 = 0.9 s), and this was consistent with a diffusion driven process. Low intensity photolysis of NP-EGTA produced a slow [Ca2+] ramp and revealed that translocation of hippocalcin–EYFP initiated at around 180 nM and was half maximal at 290 nM. Histamine induced a reversible translocation of hippocalcin–EYFP. The data show that hippocalcin is a sensitive Ca2+ sensor capable of responding to increases in intracellular Ca2+ concentration over the narrow dynamic range of 200–800 nM free Ca2+.
Keywords: calcium; neurons; hippocalcin; calcium sensors; EF-hand
Introduction
Calcium regulates numerous cellular processes and Ca2+ signals can vary from brief localized release events to a prolonged global rise (Bootman et al., 2001). These diverse Ca2+ signals are interpreted by Ca2+ sensor proteins (Carafoli, 2002). Little is known about how Ca2+ sensors react to differing Ca2+ signals, and calmodulin is the only EF-hand Ca2+-binding protein studied in living cells (Craske et al., 1999; Deisseroth et al., 1998; Milikan et al., 2002) and whose in vivo Ca2+ affinity has been determined (Persechini and Cronk, 1999). For other EF-hand Ca2+-binding proteins, only in vitro estimates of Ca2+ affinity are available. For an understanding of which Ca2+-binding proteins are functionally activated under particular Ca2+ signaling conditions, it will be essential to have direct information on their dynamics and Ca2+ affinities in living cells. The neuronal calcium sensor (NCS) proteins (Burgoyne and Weiss, 2001) are found mainly in neurons and neuroendocrine cells and are linked with modulation of neurotransmitter release (McFerran et al., 1998; Pongs et al., 1993), regulation of Ca2+ (Tsujimoto et al., 2002; Weiss et al., 2000; Weiss and Burgoyne, 2002) and K+ channels (An et al., 2000; Guo et al., 2002), regulation of phosphoinositide metabolism (Hendricks et al., 1999; Koizumi et al., 2002), phototransduction (Palczewski et al., 2000), and learning (Gomez et al., 2001). Most of the NCS proteins are NH2-terminally myristoylated. Studies on one member of the family, recoverin, showed that the myristoyl group is sequestered in the Ca2+-free form of the protein and following Ca2+-binding a substantial conformational change allows extrusion of the myristoyl group (Ames et al., 1997; Tanaka et al., 1995). This so-called Ca2+/myristoyl switch would allow recoverin to associate with membranes in a Ca2+-dependent manner. The Ca2+/myristoyl switch is used by some other NCS proteins (Ivings et al., 2002; O'Callaghan et al., 2002; Spilker et al., 2000, 2002), whereas others such as NCS-1 and guanylyl cyclase-activating protein (GCAP) 2 are membrane associated even at low [Ca2+] (O'Callaghan et al., 2002; Oleshevskaya et al., 1997).
Hippocalcin is an NCS protein most highly expressed in hippocampal neurons (Kobayashi et al., 1992). Its best characterised function is as an inhibitor of neuronal apoptosis through its interaction with neuronal apoptosis inhibitor protein (Lindholm et al., 2002). Hippocalcin has been shown to have a Ca2+/myristoyl switch mechanism within cells (O'Callaghan et al., 2002), allowing it to be cytosolic at resting [Ca2+] and translocate to intracellular membranes of the TGN and the plasma membrane in response to an increase in [Ca2+]. The Ca2+/myristoyl switch of recoverin has been examined using biochemical and structural approaches, but little work has examined the properties of the switch in NCS proteins in vivo. To address this issue, we have expressed hippocalcin–enhanced yellow fluorescent protein (EYFP) and observed its behavior in cells in response to controlled increases in intracellular [Ca2+] to characterize the dynamics and the Ca2+ sensitivity of a Ca2+/myristoyl switch protein in living cells. We demonstrate that hippocalcin is highly sensitive to cytosolic [Ca2+], but its translocation properties would require Ca2+ elevations prolonged for seconds to tens of seconds rather than very brief Ca2+ transients for a full response. The properties of hippocalcin would mean that it could integrate both temporal and spatial aspects of Ca2+ signals over a tight dynamic range.
Results and discussion
Translocation of hippocalcin in HeLa cells is dependent on Ca2+ binding
HeLa cells, which do not have endogenous hippocalcin, were transfected to express hippocalcin–EYFP, allowing the behavior of hippocalcin to be observed in isolation from neuronal-specific interacting proteins. In HeLa cells, NCS-1 is localized in resting cells to the plasma membrane and TGN—the same as the endogenous protein in neurons (Martone et al., 1999; O'Callaghan et al., 2002). Both hippocalcin–EYFP and untagged neurocalcin δ translocate to the same sites following elevation of [Ca2+], irrespective of time of transfection and expression levels (Ivings et al., 2002; O'Callaghan et al., 2002). The validity of using HeLa cells is supported by the finding that the endogenous closely related protein visinin-like protein (VILIP) 1 also translocates to the plasma membrane and TGN in hippocampal neurons (Spilker et al., 2002). Transfected HeLa cells were loaded with the Ca2+ indicator dye Fura red allowing cytosolic [Ca2+] to be measured in parallel. In an unstimulated HeLa cell, hippocalcin–EYFP was diffusely distributed throughout the cytosol and in the nucleus (Fig. 1). A robust and prolonged rise in cytosolic [Ca2+] induced by ionomycin gave a dramatic fall in fluorescence of Fura red and thus an increase in [Ca2+]. After 30 s hippocalcin–EYFP had translocated to a perinuclear compartment (TGN) and to patches on the plasma membrane (Fig. 1, upper panel; see also Figures S1 and S2, available at http://www.jcb.org/cgi/content/full/jcb.200306042/DC1). Nuclear hippocalcin–EYFP was retained in the nucleus. The translocation of hippocalcin–EYFP to membranes is dependent on its myristoylation (O'Callaghan et al., 2002) and was also dependent on the binding of Ca2+ using constructs with the glutamates at positions 85 and 121 mutated to glutamines to prevent Ca2+ binding by EF-hands 2 and 3. Hippocalcin (E85,121Q)-EYFP and hippocalcin(E121Q)-EYFP had a diffuse cytosolic distribution in cells at resting cytosolic [Ca2+] (Fig. 1). When Ca2+ was elevated, translocation of hippocalcin(E121Q)-EYFP occurred but was slower and not detectable until more than 90 s after Ca2+ elevation (Fig. 1, middle panel). In contrast, the distribution of hippocalcin (E85, E121Q)-EYFP was unaffected by the increase in cytosolic [Ca2+] (Fig. 1, lower panel), even after 220 s. These data are consistent with a Ca2+/myristoyl switch requiring Ca2+-binding to EF-hands 2 and 3 of hippocalcin. The slower kinetics with the EF-hand 3 mutant would be consistent with a reduction in Ca2+ affinity and a need for a higher threshold [Ca2+] to be reached. Hippocalcin binds 3 Ca2+ ions in vitro (Kobayashi et al., 1993) due to a nonfunctional EF-hand 1 and we do not rule out a requirement for EF-hand 4. Ca2+-binding to EF-hand 3 may initiate conformational changes in the protein allowing cooperative Ca2+ binding to the other EF-hands as in other NCS proteins (Cox et al., 1994; Permyakov et al., 2000; Senin et al., 2002).
Figure 1.
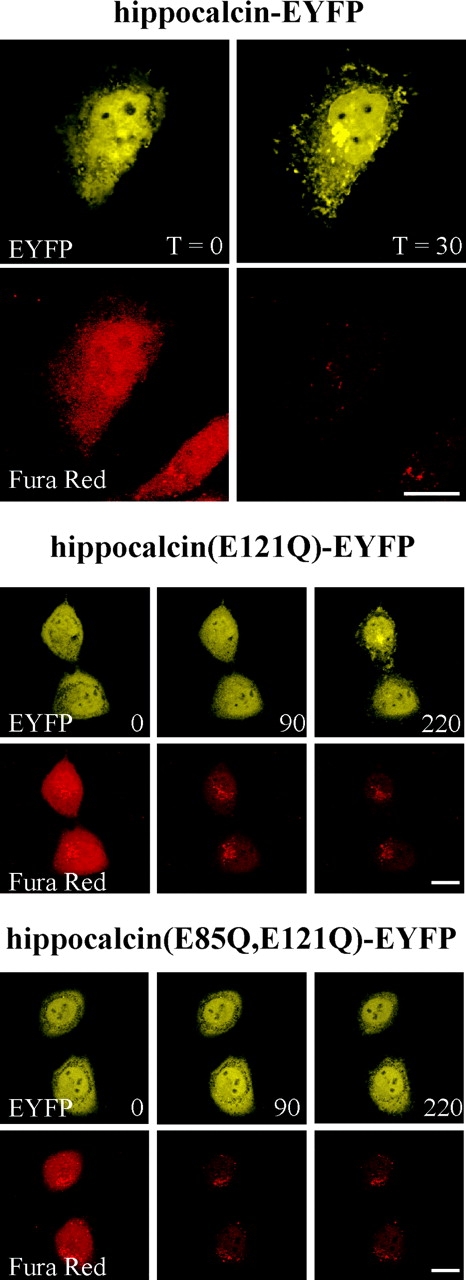
Requirement of EF-hands for translocation of hippocalcin–EYFP. HeLa cells were transfected to express hippocalcin–EYFP, hippocalcin (E121Q)-EYFP, or hippocalcin (E85Q, E121Q)-EYFP. After loading with Fura red, live cells were imaged by confocal microscopy and Ca2+ elevated by addition of 3 μM ionomycin. Images are shown immediately before addition (time 0) or at the indicated times after addition of ionomycin. In each case, images of Fura red are shown underneath the corresponding images of EYFP fusion proteins. Bars, 10 μM.
Hippocalcin is freely diffusible in the cytoplasm in resting cells
To determine that hippocalcin in its Ca2+ unbound form is freely diffusible, localized photobleaching was used to probe the movement of hippocalcin–EYFP in transfected HeLa cells (Fig. 2, A and B). A localized area of the cell (Fig. 2 B) was exposed to high-intensity 514-nm light for 30 s. During this period, the whole cytosol rapidly dimmed, indicating that hippocalcin–EYFP was capable of rapid diffusion. Fluorescence was retained in the nucleus, indicating that this is a diffusionally isolated region. The retention of nuclear fluorescence and that of surrounding cells indicates the regional specificity of the photobleaching. To examine the speed at which the diffusion of hippocalcin–EYFP occurs, we created a photoactivatable version termed hippocalcin–photoactivatable (PA)–GFP using photoactivatable GFP (Patterson and Lippincott-Schwartz, 2002). This protein was only dimly fluorescent (Fig. 2 C) until excited with 430-nm light. The illumination of half of a HeLa cell expressing hippocalcin-PA-GFP allowed the speed of diffusion of the photoactivated hippocalcin-PA-GFP to be measured. In the example shown, fluorescence intensity was monitored at two regions of interest that were 9.37-μm apart. The rate of diffusion of hippocalcin-PA-GFP from the delay between the two regions was 2.9 μm/s. This rate was comparable to that found for PA-GFP (3.2 μm/sec; data not shown). This demonstrates that at resting [Ca2+] cytosolic hippocalcin is not membrane-associated but is freely diffusible in the cytosol. The rate of diffusion would be sufficient to account for the translocation process.
Figure 2.

Demonstration that hippocalcin–EYFP is freely diffusible in resting cells. (A) HeLa cells were transfected to express hippocalcin–EYFP, which showed a diffuse cytosolic and nuclear localization. (B) One cell was locally photobleached within the indicated box for 30 s leading to a loss of cytosolic fluo-rescence. Nuclear fluorescence in the neighboring cells and in the nucleus of the photobleached cell was unaffected. (C) A cell transfected with pHippo-PA-GFP was illuminated with high-intensity 430-nm light on its right hand side to photoactivate the hippocalcin-PA-GFP. The images i–iv are shown before activation (i) and 2 (ii), 5 (iii), and 15 (iv) s after activation. (C) Time course of GFP fluorescence increases measured in the indicated regions of interest. Bar, 10 μm.
Maximal rate of translocation of hippocalcin–EYFP
Ionomycin produces a sustained but slow increase in cytosolic [Ca2+] in HeLa cells and so the dynamics of hippocalcin–EYFP translocation will be dependent on the rate of this Ca2+ rise. Indeed translocation of hippocalcin–EYFP after ionomycin addition occurs only after a lag period of around 2 s and then with a time constant of around 12 s. To assess the true maximum rate of Ca2+-dependent translocation and to rule out any effects on the kinetics of Ca2+-buffering by the expressed proteins, pHippo-EYFP transfected HeLa cells were loaded with the caged Ca2+ compound NP-EGTA. Photolysis of the NP-EGTA rapidly increased the cytosolic [Ca2+] to high levels indicated by the sharp drop in Fura red fluorescence and to an immediate initiation of translocation of hippocalcin (Fig. 3, A and B). This translocation could be fitted to first order kinetics with a half time of 0.9 s (mean value from 8 cells) and translocation was complete within <10 s. The initiation of translocation was concomitant with the elevation of [Ca2+], indicating that the conformational change induced by Ca2+ binding is not rate limiting for the translocation.
Figure 3.
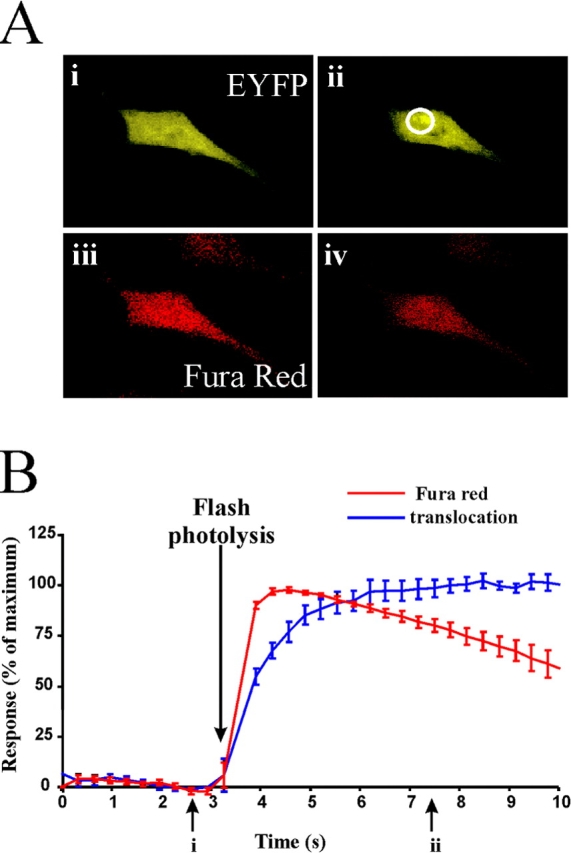
Rapid translocation of hippocalcin–EYFP in response to flash photolysis of NP-EGTA. (A) A HeLa cell expressing hippocalcin–EYFP was imaged for EYFP or Fura red before (i and iii) or after (ii and iv) photolysis of NP-EGTA (B). Fluorescence intensity was monitored in the region of interest and averaged to show the time course of changes in Fura red and hippocalcin–EYFP fluorescence near the TGN. Data shown as mean ± SEM (n = 8 cells).
The trigger [Ca2+] for translocation of hippocalcin–EYFP
To determine the threshold [Ca2+] for translocation, transfected HeLa cells loaded with Fura red and NP-EGTA were irradiated using low-intensity 360-nm light. This produced a slowly rising ramp in intracellular [Ca2+] seen as a decrease in Fura red fluorescence (Fig. 4 A). By calibrating the Fura red fluorescence, the intracellular [Ca2+] was calculated. The representative experimental trace (Fig. 4 B) shows the progressive rise in [Ca2+] starting at 100 nM. The initiation of translocation was seen when the [Ca2+] reached ∼180 nM and was half-maximal at around 320 nM. The data were replotted as the percentage of translocation versus [Ca2+] and could be fitted by a curve defined by the Hill equation with a K0.5 of 324 nM and a Hill coefficient of 3.3 (Fig. 4 C), showing high cooperativity in the response of hippocalcin to [Ca2+] elevation. The mean for the trigger concentration was 181 ± 9 nM and half maximal translocation was at 293 ± 20 nM free Ca2+ (11 cells). Translocation was complete as [Ca2+] reached ∼800 nM, indicating that hippocalcin has a dynamic range of 200–800 nM free Ca2+. This is more than tenfold as sensitive to Ca2+ than the Kd (5 μM) for recombinant hippocalcin (Kobayashi et al., 1993), demonstrating the problem with reliance on in vitro analyses. The Ca2+ affinity of hippocalcin is higher than that seen for calmodulin, which was around 1 μM free Ca2+ (Persechini and Cronk, 1999).
Figure 4.
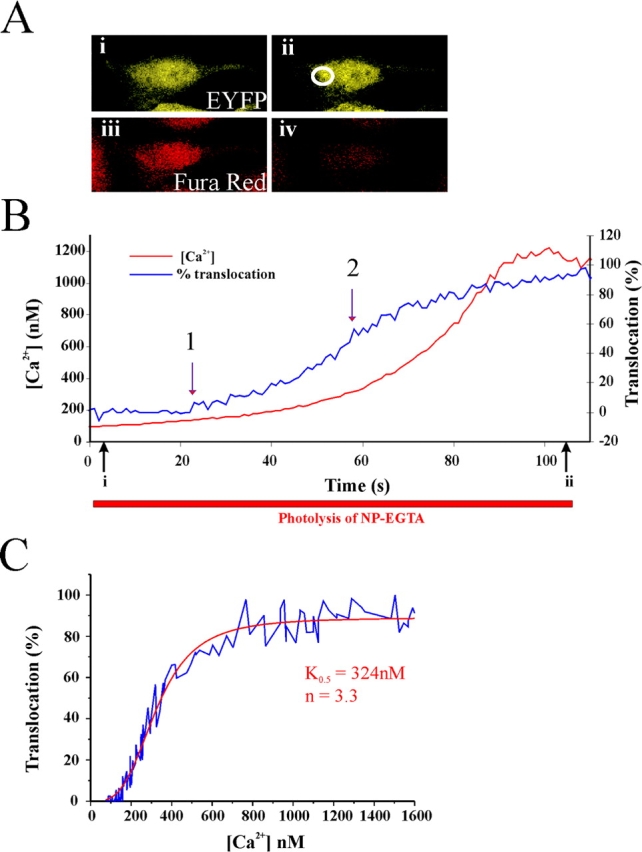
Assessment of the Ca2 + dependency of hippocalcin–EYFP translocation. (A) HeLa cells expressing hippocalcin–EYFP and loaded with Fura red were imaged before (i and iii) and after (ii and iv) release of Ca2+ by slow photolysis of NP-EGTA. (B) Changes in hippocalcin–EYFP fluorescence were monitored in the indicated region near the Golgi complex. Fura red fluorescence over the cell was converted to [Ca2+]. The initiation of translocation (1) and the time it was half-maximal (2) are indicated. (C) The data for percentage translocation were plotted against free Ca2+ concentration near the Golgi complex, and the curve derived by fitting the data to the Hill equation.
Hippocalcin responds to agonist-induced Ca2+ elevation and translocation is reversible
To examine the reversibility of the Ca2+/myristoyl switch, hippocalcin–EYFP expressing HeLa cells were exposed to NP-EGTA photolysis to induce translocation (Fig. 5 A, i and ii). This translocation was completely reversed after reduction of cytosolic [Ca2+] using 3 μM ionomycin in a Krebs-Ringer buffer bath solution with 3 mM EGTA present (Fig. 5 A, iii). Translocation was then induced again by the replacement of the bath buffer with Krebs-Ringer buffer containing 3 mM Ca2+ (Fig. 5 A, iv). To determine whether hippocalcin would respond to Ca2+ transients induced by physiological agonists we used histamine to stimulate transfected HeLa cells. Fig. 5 B shows the initial response in which rapid translocation occurred upon addition of histamine after a lag of around 2 s. Translocation was half-maximal at 278 nM free Ca2+, similar to that seen with NG-EGTA. Hippocalcin subsequently dissociated from the membranes (Fig. 5 C, iii), but hippocalcin could translocate again to the same sites when ionomycin was added (Fig. 5 C, iv). This indicates the hippocalcin can respond to agonist-induced elevation of [Ca2+] and also that it acts as a fully reversible switch.
Figure 5.
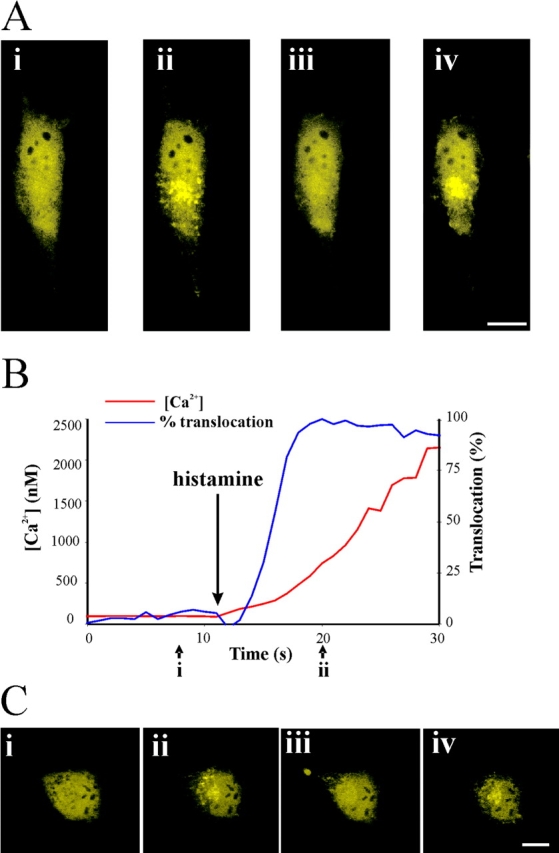
Hippocalcin–EYFP translocation is reversible and is elicited by agonist stimulation. (A) A HeLa cell expressing hippocalcin–EYFP was imaged before (i) or 10 s after (ii) Ca2+ release by rapid photolysis of NP-EGTA. External Ca2+ was reduced by addition of 3 μM ionomycin/3 mM EGTA (iii) and then elevated again by addition on 3 mM Ca2+ (iv). (B) Initial time course of changes in [Ca2+] and hippocalcin–EYFP fluorescence around the Golgi complex in response to histamine addition. (C) Images of hippocalcin–EYFP in a HeLa cell before (i) and 10 (ii) and 100 (iii) s after addition of 100 μM histamine, and a further 10 s after subsequent addition of 3 μM ionomycin. Bar, 10 μM.
Concluding remarks
These experiments have yielded new insights into the behavior of hippocalcin in living cells that indicate how it could integrate both temporal and spatial aspects of Ca2+ signals. The extent of translocation is not just a function of [Ca2+] but also the time that Ca2+ remains elevated. Hippocalcin translocation is more sensitive to increases in intracellular Ca2+ concentration than calmodulin but the full responses are on the time scale one seconds to tens of seconds. This is slower than the Ca2+ transients seen in neurons due to action potentials but longer-lasting Ca2+ elevations lasting up to hundreds of milliseconds are physiologically important in neurons for the induction of long-term potentiation or long-term depression (Sabatini et al., 2002; Sjostrom and Nelson, 2002). In a small neuronal process or dendritic spine, hippocalcin could diffuse sufficiently to translocate on such a time scale. One could imagine how the distinct properties of Ca2+ sensors such as calmodulin and hippocalcin could determine distinct cellular responses to differing Ca2+ signals.
Materials and methods
Plasmids and transfection of HeLa cells
EYFP-tagged hippocalcin (pHippo-EYFP) was made as described previously (O'Callaghan et al., 2002). pHippo(E121Q)-EYFP and pHippo (E85,121Q)-EYFP were made using the QuikChange™ site-directed mutagenesis kit (Stratagene). The photoactivatable hippocalcin construct pHippo-PA-GFP was made by cutting out the EYFP from pHippo-EYFP and replacing it with the PA-GFP from pPA-GFP-N1. pPA-GFP-N1 was generated with the mutations described previously (Patterson and Lippincott-Schwartz, 2002). HeLa cells were grown and transfected as described previously (O'Callaghan et al., 2002) and maintained for 8–96 h before use.
Confocal laser scanning microscopy on living cells
For confocal laser scanning microscopy, live transfected HeLa cells were examined with either a Zeiss LSM 150 confocal microscope or a Leica TCS-SP-MP microscope using a 63× water immersion objective with a 1.2 numerical aperture. Cells were loaded with Fura red (Molecular Probes) by incubation in 5 μM acetoxymethylester and loaded with NP-EGTA (Molecular Probes) by incubation in 10 μM acetoxymethylester in growth medium for 30 min. The cells were bathed in a Krebs-Ringers solution (145 mM NaCl, 5 mM KCl, 1.3 mM MgCl2, 1.2 mM NaH2PO4, 10 mM glucose, 20 mM HEPES; pH 7.4) with 3 mM CaCl2 and were excited at 488 nm and light collected at 625–725 nm for Fura red emission and at 525–590 for hippocalcin–EYFP emission. Ionomycin when used was added to the bath solution to a final concentration of 3 μM. With pHippo-PA-GFP, 488-nm illumination was used for excitation, and the emission was collected between 500–550 nm. Photoactivation of Hippo-PA-GFP was with 430-nm laser line illumination at full power of the laser on the Leica TCS-SP-MP system for 15 s. The photolysis of NP-EGTA was by illumination with 360-nm laser light at full power for rapid photolysis and at 6% power for generation of a [Ca2+] ramp. Histamine was added to the bath solution at a final concentration of 100 μM. For calibration of Fura red fluorescence as [Ca2+], the cells were treated with 3 μM ionomycin at the end of the experiment in the presence of 3 mM EGTA to determine Fmin. Due to problems with photobleaching leading to inaccurate direct determination of Fmax, a predicted Fmax value at resting [Ca2+] (FmaxP) was determined based on the use of a resting [Ca2+] of 100 nM that has been well established for HeLa cells (Thomas et al., 2000). [Ca2+] was calculated using a Kd of 140 nM. This calibration method would, if anything, overestimate rather than underestimate the [Ca2+] at which hippocalcin–EYFP translocation was observed. Data for percentage translocation versus [Ca2+] were fitted by nonlinear curve fitting using the Hill equation.
Online supplemental material
Figure S1 shows different confocal sections through the same HeLa cell after treatment with ionomycin. These were selected to show optimal images of the localization of hippocalcin–EYFP either at the TGN or at the plasma membrane. Figure S2 shows data averaged from four cells from imaging of hippocalcin–EYFP and Fura red fluorescence showing that translocation to the TGN or to the plasma membrane occurs with the same kinetics. Both figures are available at http://www.jcb.org/cgi/content/full/jcb.200306042/DC1.
Supplemental Material
Acknowledgments
We thank Dr. Mike Ashby and Nick Dolman for help in establishing the protocols for flash photolysis of NP-EGTA.
D.W. O'Callaghan was supported by a Wellcome Trust Prize studentship.
The online version of this article includes supplemental material.
Abbreviations used in this paper: EYFP, enhanced yellow fluorescent protein; GCAP, guanylyl cyclase-activating protein; NCS, neuronal calcium sensor; PA, photoactivatable; VILIP, visinin-like protein.
References
- Ames, J.B., R. Ishima, T. Tanaka, J.I. Gordon, L. Stryer, and M. Ikura. 1997. Molecular mechanics of calcium-myristoyl switches. Nature. 389:198–202. [DOI] [PubMed] [Google Scholar]
- An, W.F., M.R. Bowlby, M. Bett, J. Cao, H.P. Ling, G. Mendoza, J.W. Hinson, K.I. Mattsson, B.W. Strassle, J.S. Trimmer, and K.J. Rhodes. 2000. Modulation of A-type potassium channels by a family of calcium sensors. Nature. 403:553–556. [DOI] [PubMed] [Google Scholar]
- Bootman, M.D., P. Lipp, and M.J. Berridge. 2001. The organisation and functions of local Ca2+ signals. J. Cell Sci. 114:2213–2222. [DOI] [PubMed] [Google Scholar]
- Burgoyne, R.D., and J.L. Weiss. 2001. The neuronal calcium sensor family of Ca2+-binding proteins. Biochem. J. 353:1–12. [PMC free article] [PubMed] [Google Scholar]
- Carafoli, E. 2002. Calcium signalling: a tale for all seasons. Proc. Natl. Acad. Sci. USA. 99:1115–1122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox, J.A., I. Drussel, M. Comte, S. Nef, P. Nef, S.E. Lenz, and E.D. Gundelfinger. 1994. Cation binding and conformational changes in VILIP and NCS-1, two neuron-specific calcium-binding proteins. J. Biol. Chem. 269:32807–32814. [PubMed] [Google Scholar]
- Craske, H., T. Takeo, O. Gerasimenko, C. Vaillant, K. Torok, O.H. Petersen, and A.V. Tepikin. 1999. Hormone-induced secretory and nuclear translocation of calmodulin: Oscillations of calmodulin concentration with the nucleus as an integrator. Proc. Natl. Acad. Sci. USA. 96:4426–4431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deisseroth, K., E.K. Heist, and R.W. Tsien. 1998. Translocation of calmodulin to the nucleus supports CREB phosphorylation in hippocampal neurons. Nature. 392:198–202. [DOI] [PubMed] [Google Scholar]
- Gomez, M., E. De Castro, E. Guarin, H. Sasakura, A. Kuhara, I. Mori, T. Bartfai, C.I. Bargmann, and P. Nef. 2001. Ca2+ signalling via the neuronal calcium sensor-1 regulates associative learning and memory in C.elegans. Neuron. 30:241–248. [DOI] [PubMed] [Google Scholar]
- Guo, W., S.A. Malin, D.C. Johns, A. Jeromin, and J.M. Nerbonne. 2002. Modulation of Kv4-encoded K+ currents in the mammalian myocardium by neuronal calcium sensor-1. J. Biol. Chem. 277:26436–26443. [DOI] [PubMed] [Google Scholar]
- Hendricks, K.B., B.Q. Wang, E.A. Schnieders, and J. Thorner. 1999. Yeast homologue of neuronal frequenin is a regulator of phosphatidylinositol-4-OH kinase. Nat. Cell Biol. 1:234–241. [DOI] [PubMed] [Google Scholar]
- Ivings, L., S.R. Pennington, R. Jenkins, J.L. Weiss, and R.D. Burgoyne. 2002. Identification of calcium-dependent binding partners for the neuronal calcium sensor protein neurocalcin δ: interaction with actin, clathrin and tubulin. Biochem. J. 363:599–608. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kobayashi, M., K. Takamatsu, S. Saitoh, M. Miura, and T. Noguchi. 1992. Molecular cloning of hippocalcin, a novel calcium-binding protein of the recoverin family exclusively expressed in hippocampus. Biochem. Biophys. Res. Commun. 189:511–517. [DOI] [PubMed] [Google Scholar]
- Kobayashi, M., K. Takamatsu, S. Saitoh, and T. Noguchi. 1993. Myristoylation of hippocalcin is linked to its calcium-dependent membrane association properties. J. Biol. Chem. 268:18898–18904. [PubMed] [Google Scholar]
- Koizumi, S., P. Rosa, G.B. Willars, R.A.J. Challiss, E. Taverna, M. Francolini, M.D. Bootman, P. Lipp, K. Inoue, J. Roder, and A. Jeromin. 2002. Mechanisms underlying the neuronal calcium sensor-1 evoked enhancement of exocytosis in PC12 cells. J. Biol. Chem. 277:30315–30324. [DOI] [PubMed] [Google Scholar]
- Lindholm, D., E.A. Mercer, L.-Y. Yu, Y. Chen, J. Kukkonen, L. Korhonen, and U. Arumae. 2002. Neuronal apoptosis inhibitory protein: structural requirements for hippocalcin binding and effects on survival of NGF-dependent sympathetic neurons. Biochim. Biophys. Acta. 1600:138–147. [DOI] [PubMed] [Google Scholar]
- Martone, M.E., V.M. Edelmann, M.H. Ellisman, and P. Nef. 1999. Cellular and subcellular distribution of the calcium-binding protein NCS-1 in the central nervous system of the rat. Cell Tissue Res. 295:395–407. [DOI] [PubMed] [Google Scholar]
- McFerran, B.W., M.E. Graham, and R.D. Burgoyne. 1998. NCS-1, the mammalian homologue of frequenin is expressed in chromaffin and PC12 cells and regulates neurosecretion from dense-core granules. J. Biol. Chem. 273:22768–22772. [DOI] [PubMed] [Google Scholar]
- Milikan, J.M., T.D. Carter, J.H. Horne, A. Tzortzopoulos, K. Torok, and S.R. Bolsover. 2002. Integration of calcium signals by calmodulin in rat sensory neurons. Eur. J. Neurosci. 15:661–670. [DOI] [PubMed] [Google Scholar]
- O'Callaghan, D.W., L. Ivings, J.L. Weiss, M.C. Ashby, A.V. Tepikin, and R.D. Burgoyne. 2002. Differential use of myristoyl groups on neuronal calcium sensor proteins as a determinant of spatio-temporal aspects of Ca2+-signal transduction. J. Biol. Chem. 277:14227–14237. [DOI] [PubMed] [Google Scholar]
- Oleshevskaya, E.V., E.E. Hughes, J.B. Hurley, and A.M. Dizhoor. 1997. Calcium binding, but not calcium-myristoyl switch, controls the ability of guanyl cyclase-activating protein GCAP-2 to regulated photoreceptor guanyl cyclase. J. Biol. Chem. 272:14327–14333. [DOI] [PubMed] [Google Scholar]
- Palczewski, K., A. Polans, W. Baehr, and J.B. Ames. 2000. Ca2+-binding proteins in the retina: structure, function and the etiology of human visual diseases. Bioessays. 22:337–350. [DOI] [PubMed] [Google Scholar]
- Patterson, G.H., and J. Lippincott-Schwartz. 2002. A photoactivatable GFP for selective photolabeling of proteins and cells. Science. 297:1873–1877. [DOI] [PubMed] [Google Scholar]
- Permyakov, S.E., A.M. Cherskaya, I.I. Senin, A.A. Zargarov, S.V. Shulga-Morskoy, A.M. Alekseev, D.V. Zinchenko, V.M. Lipkin, P.P. Philippov, V.N. Uversky, and E.A. Permyakov. 2000. Effects of mutations in the calcium-binding sites of recoverin on its calcium affinity: evidence for successive filling of the calcium binding sites. Prot. Engin. 13:783–790. [DOI] [PubMed] [Google Scholar]
- Persechini, A., and B. Cronk. 1999. The relationship between the free concentrations of Ca2+ and Ca2+-calmodulin in intact cells. J. Biol. Chem. 274:6827–6830. [DOI] [PubMed] [Google Scholar]
- Pongs, O., J. Lindemeier, X.R. Zhu, T. Theil, D. Endelkamp, I. Krah-Jentgens, H.-G. Lambrecht, K.W. Koch, J. Schwemer, R. Rivosecchi, et al. 1993. Frequenin - A novel calcium-binding protein that modulates synaptic efficacy in the drosophila nervous system. Neuron. 11:15–28. [DOI] [PubMed] [Google Scholar]
- Sabatini, B.L., T.G. Oertner, and K. Svoboda. 2002. The life cycle of Ca2+ ions in dendritic spines. Neuron. 33:439–452. [DOI] [PubMed] [Google Scholar]
- Senin, I.I., T. Fischer, K.E. Komolov, D.V. Zinchenko, P.P. Philippov, and K.-W. Koch. 2002. Ca2+-myristoyl switch in the neuronal calcium sensor recoverin requires different functions of Ca2+ binding sites. J. Biol. Chem. 277:50365–50372. [DOI] [PubMed] [Google Scholar]
- Sjostrom, J.P., and S.B. Nelson. 2002. Spike timing, calcium signals and synaptic plasticity. Curr. Opin. Neurobiol. 12:305–314. [DOI] [PubMed] [Google Scholar]
- Spilker, C., K. Richter, K.H. Smalla, D. Manahan-Vaughan, E.D. Gundelfinger, and K.H. Braunewell. 2000. The neuronal EF-hand calcium-binding protein visinin-like protein-3 is expressed in cerebellar Purkinje cells and shows a calcium-dependent membrane association. Neuroscience. 96:121–129. [DOI] [PubMed] [Google Scholar]
- Spilker, C., T. Dresbach, and K.-H. Braunewell. 2002. Reversible translocation and activity-dependent localisation of the calcium-myristoyl switch protein VILIP-1 to different membrane compartments in living hippocampal neurons. J. Neurosci. 22:7331–7339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tanaka, T., J.B. Ames, T.S. Harvey, L. Stryer, and M. Ikura. 1995. Sequestration of the membrane targeting myristoyl group of recoverin in the calcium-free state. Nature. 376:444–447. [DOI] [PubMed] [Google Scholar]
- Thomas, D., P. Lipp, S.C. Tovey, M.J. Berridge, W. Li, R.W. Tsien, and M.D. Bootman. 2000. Microscopic properties of elementary Ca2+ release sites in non-excitable cells. Curr. Biol. 10:8–15. [DOI] [PubMed] [Google Scholar]
- Tsujimoto, T., A. Jeromin, N. Satoh, J.C. Roder, and T. Takahashi. 2002. Neuronal calcium sensor 1 and activity-dependent facilitation of P/Q-type calcium channel currents at presynaptic nerve terminals. Science. 295:2276–2279. [DOI] [PubMed] [Google Scholar]
- Weiss, J.L., D.A. Archer, and R.D. Burgoyne. 2000. NCS-1/frequenin functions in an autocrine pathway regulating Ca2+ channels in bovine adrenal chromaffin cells. J. Biol. Chem. 275:40082–40087. [DOI] [PubMed] [Google Scholar]
- Weiss, J.L., and R.D. Burgoyne. 2002. Sense and sensibility in the regulation of voltage-gated calcium channels. Trends Neurosci. 25:489–491. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}